
Submitted:
26 August 2024
Posted:
27 August 2024
You are already at the latest version
Abstract
Coilia nasus is an important fish resource in the Yangtze River, and the Yangtze River Estuary is a crucial migration pathway for it. In this study, we used otolith microchemistry to analyze the strontium/calcium (Sr/Ca) ratios and Sr contents in the sagitta otolith of C. nasus from the south branch (SB) and north branch (NB) of the Yangtze River Estuary, and obtained the diversity of migration patterns and spawning grounds distribution for C. nasus. The results indicate that C. nasus from both branches include two types of habitat history: freshwater (F)-brackish water (B) (Type I) and F-B-seawater (S) (Type II), with Type I being dominant at 62.50% in both branches. The C. nasus from SB comprises six migration patterns, while that from NB has seven migration patterns. The C. nasus from both branches hatch in F habitats. At the time of capture, the C. nasus from SB predominantly remain in F, accounting for 62.5%, while C. nasus from NB primarily stay in B, accounting for 87.5%. Throughout the migration process, C. nasus from both branches switch between different habitats, with C. nasus from NB exhibiting more frequent transitions between F and B, showing a greater reliance on the estuarine brackish habitat. The radius of the first blue region near the core (Lf) and freshwater coefficient (Fc) of otolith for C. nasus from both branches are divided into three groups: Long-distance freshwater dependence (LD), medium-distance freshwater-dependent (MD), and short-distance freshwater dependence (SD), with the LD only appearing in the SB, while the NB is primarily represented by the MD. There is a correlation between the differences in Lf among different groups of C. nasus and the differences in the distance from the spawning grounds of C. nasus in different sections of the Yangtze River to the Estuary (DYRE), reflecting the distribution pattern of C. nasus spawning grounds in different sections of the Yangtze River. This study provides theoretical guidance for the protection of migration pathway and maintenance of spawning grounds for C. nasus, which has significant practical value in the precise management of C. nasus resources in the Yangtze River Estuary.
Keywords:
Coilia nasus
; otolith microchemistry
; migration pattern
; spawning grounds distribution
; Yangtze River Estuary
1. Introduction
Coilia nasus belongs to the order Clupeiformes, family Engraulidae, genus Coilia. Widely distributed in the Northwest Pacific, including China, Korea and Japan [1]. In China, C. nasus is an important migratory economic fish in the middle and lower reaches of the Yangtze River. Every spring, the anadromous C. nasus migrate from the coast through estuaries to lakes along the middle and lower reaches of the Yangtze River to breed [2]. Historically, the C. nasus can be traced as far back as the lake of Dongtinghu (DTH), which is 1,400 km from the Yangtze River Estuary [3,4,5]. The highest production had reached 3,945 t (1973), with 390 t caught at the Yangtze River Estuary alone [6,7]. With the increase of fishing intensity, water pollution, and the construction of dams to block the upstream spawning channel, the production of C. nasus has been decreasing since the 1970s, and the population age and individual size have been miniaturized, the age of sexual maturity has been advanced, and there is an increase in the number of resident type C. nasus [8].
Previous studies generally categorized C. nasus into three different ecotypes: anadromous migratory type, freshwater resident type and landlocked type. However, this classification method can’t fully reflect different ecomorphotypes of C. nasus [9]. Habitat history and migration distance of C. nasus can be clearly determined by otolith microchemistry [10]. Otoliths are a type of calcareous stone carried by osteichthyes fish themselves, which are highly stable and not easily reabsorbed [11,12]. Otoliths are formed during the incubation period of fertilized eggs, after which they grow around a core deposit and record the life pattern and habitat processes experienced by the fish throughout its life [13]. The core corresponds to the early life stage of fish and records their spawning and natal habitats, while the edge corresponds to the capture and recent living environment [14]. There is a positive correlation between the trace element content of otoliths and the elemental content of the environment, especially strontium (Sr) and barium (Ba) [15]. The Sr is usually much higher in seawater (S) than in freshwater (F), which leads to an increase in the Sr/Ca ratio of otoliths when fishes enter seawater [16]. Using of otolith microchemistry provides a more objective and accurate reflection of fish habitat history and migratory patterns than relying on jaw-to-head ratios [9].
The current research on the habitat history of C. nasus in the Yangtze River and its connected lakes by otolith microchemistry is relatively extensive [2,5,8]. However, studies on the migration patterns and the spawning grounds distribution of C. nasus from the Yangtze River Estuary are limited [17]. This study conducts sampling in both the south branch (SB) and north branch (NB) of the Yangtze River estuary to analyze the habitat history and migration patterns of C. nasus from different sources. By analyzing the correlation between the radius of the first blue region near the core (Lf) of otolith and the distance from the spawning grounds of C. nasus in different sections of the Yangtze River to the Estuary (DYRE), this study aims to clarify the composition of migratory C. nasus in the Yangtze River Estuary and their upstream migration distribution areas. This research will provide theoretical guidance and technical support for the protection of C. nasus breeding populations, maintenance of key habitats, and precise management of C. nasus resources and their migration pathway.
2. Materials and Methods
2.1. Sampling Sites, Sample Collection, and Processing
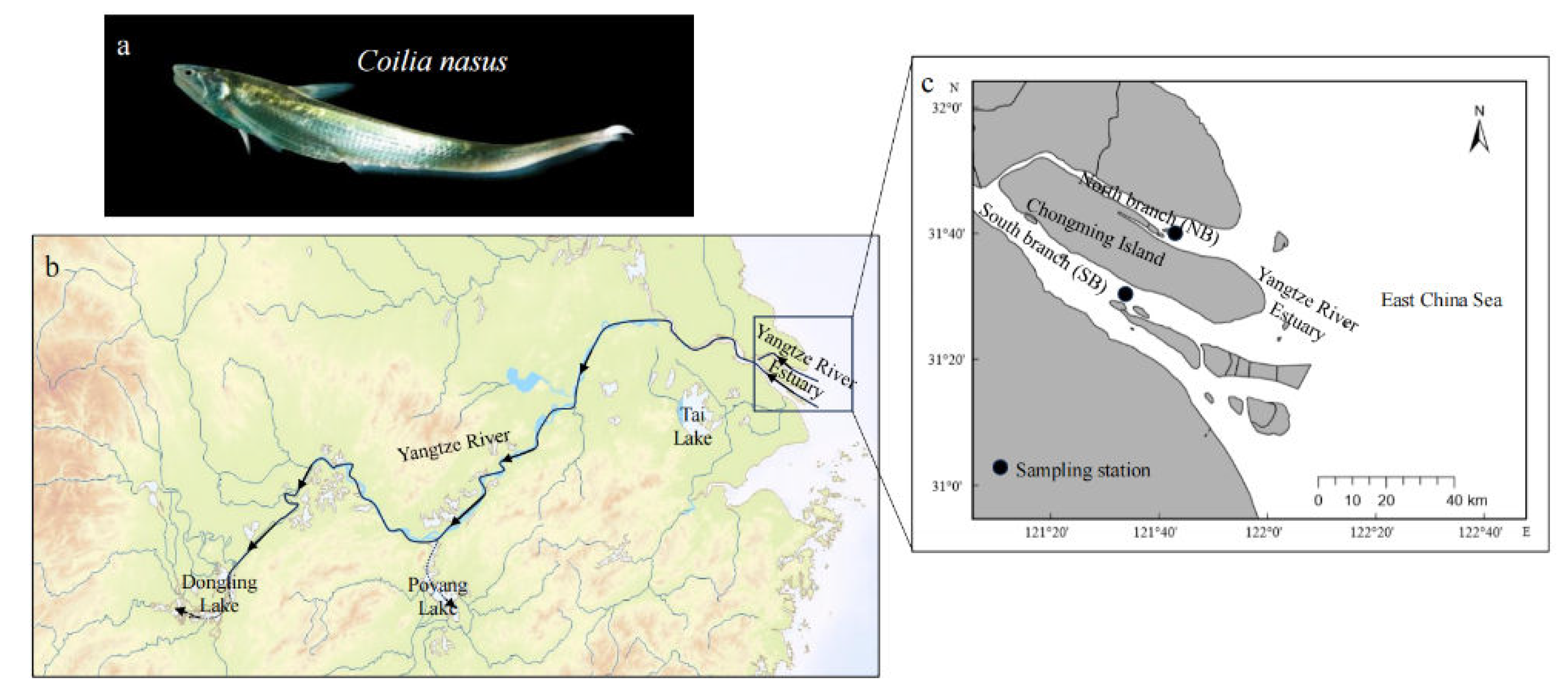
Sixteen fish samples were investigated, which were collected by trammel gill net from the south branch (SB: S1, S2, S3, S4, S5, S6, S7, S8) and north branch (NB: N1, N2, N3, N4, N5, N6, N7, N8) of the Yangtze River Estuary. Before otolith extraction, fish samples were routinely measured by electronic vernier caliper and electronic balance to obtain the total length, body length and wet weight. Eight individuals were collected from SB with the average total length, body length and wet weight are 332.25±28.43 mm, 306.00±26.57 mm and 110.97±28.42 g, another eight individuals were collected from NB with that of 245.00±12.39 mm, 221.13±13.17 mm and 33.37±7.44 g, respectively.
Figure 1.
Coilia nasus (a) sampling sites (b, c): North branch (NB) around 121.814°E, 31.662° N and south branch (SB) around 121.620°E, 31.505° N in the Yangtze River Estuary. The arrow in b indicates the direction of upstream migration of C. nasus.
Figure 1.
Coilia nasus (a) sampling sites (b, c): North branch (NB) around 121.814°E, 31.662° N and south branch (SB) around 121.620°E, 31.505° N in the Yangtze River Estuary. The arrow in b indicates the direction of upstream migration of C. nasus.
2.2. Otolith Treatment and Microchemical Analysis
Right sagittal otoliths were uniformly used for pretreatment and microchemical analysis. Otoliths were processed and microchemical analyzed according to previous methods [17], as follows: The otoliths were embedded in epoxy resin (Epofix, Struers, Copenhagen, Denmark), and then polished with an automated polishing wheel (Discoplan-TS mill, Struers Copenhagen, Denmark) to expose the core. All otoliths were then polished with a LaboPol-35 polishing machine (Struers, Copenhagen, Denmark) to remove scratches from the otolith surface. All otoliths were washed in a Milli-Q water for 5 min, followed by 6 rinses with Milli-Q water. Afterwards, all otoliths were dried in an oven at 40 °C and finally coated (36 A for 25 s) using a vacuum coater (EE420, JEOL Ltd., Tokyo, Japan). The quantitative line of Sr/Ca ratios and the X-ray intensity map of Sr content were carried out by EPMA (JXA-8100, JEOL Ltd., Tokyo, Japan). Calcite (CaCO3) and tausonite (SrTiO3) were used as standards for calibrating Ca and Sr measurements, respectively [18].
2.3. Data analysis
The line charts were drawn by Excel 365 MSO, where X-axis was the radius length of the otolith and Y-axis was Sr/Ca ratios which were customarily calculated and expressed as Sr/Ca×1000. To reflect the trend of Sr/Ca ratios of otoliths more directly, the results of quantitative analysis were trend-shifted by Regime detection 3.2 with a significance level of p = 0.05, a cut-off length of 5 and a Huber’s weight parameter of 1 [19]. To detect the significant differences in Sr/Ca ratios of otoliths, IBM SPSS Statistics v.27.0 was used for independent sample t-test and One-way analysis of variance (ANOVA). The Sr/Ca ratios of otolith could effectively correspond to the characteristics of salinity differences in different habitats as F, B and S: (1) In F, the Sr/Ca ratio was generally less than 3, which corresponds to the blue spectrum in Sr content; (2) in B, the Sr/Ca ratio was generally between 3 and 7, which corresponds to the green-yellow spectrum in Sr content; (3) in S, the Sr/Ca ratio was generally greater than 7, which corresponds to the red spectrum in Sr content [20]. Moreover, freshwater coefficient (Fc) refers to the ratio of the time spent in F to the whole life history in the early life history. The formula Fc=Lf/LT, was used to calculate freshwater coefficient, where LT was the analytical radius of the entire otolith [21].
3. Results
3.1. Sr/Ca Ratios Quantitative Line and Sr Content Analysis
Otoliths of C. nasus from the NB and SB of the Yangtze River Estuary showed various Sr/Ca ratios. As shown in Figure 2, the entire otolith quantification line was divided into three habitats based on the Sr/Ca ratios and their variations, including F, B and S. In terms of the starting point of the quantification line, C. nasus from both branches with the Sr/Ca ratios of less than 3 (below the green dotted line), and is distributed in F, all of which are freshwater origin and freshwater hatching type. The C. nasus from both branches showed different length of the quantitative line in the initial freshwater area (from the start of the quantitative line to the first intersection of the blue solid line with the green dotted line), indicating that their utilization of freshwater habitats varied during the early stages of their life history. From the spanning of the otolith quantification line between different habitats, the presence of individuals spanning 2 habitats (quantification line values are located below the red dotted line), switching between F and B, and the presence of individuals spanning 3 habitats (the quantification line intersects with both the green dotted line and the red dotted line), switching between F-B-S, it can be seen that both C. nasus from SB and NB comprise 2 types of habitat history types, F-B (Type I) and F-B-S (Type II).
Otoliths of C. nasus from SB and NB of the Yangtze River Estuary showed various Sr contents. As shown in Figure 3, otolith Sr content showed different colors variations from the core to the edge, and the colors variations in the facial distribution of Sr content corresponded to the different habitats showed in the quantitative line analysis (blue corresponds to F, yellow-green corresponds to B, and red corresponds to S) [22]. The core color of the otolith of C. nasus from both branches are blue, indicating that they are both of freshwater origin. The color distribution in the otolith of some individuals from SB and NB showed red areas, suggesting that the C. nasus from both branches have seawater habitats. The color change (the color alternated between blue, green and yellow) was observed in the otolith of C. nasus from both branches, suggesting that some individuals showed the reciprocal movement between F and B, and mainly using brackish water habitats in estuaries.
3.2. Habitat History Decomposition and Migration Pattern Analysis
Otoliths of C. nasus from SB and NB showed various Sr/Ca ratios and Sr contents. The otolith Sr/Ca ratios changes and the Sr contents distribution showed that there were two habitat history types for C. nasus from both branches, but they showed different migration patterns and paths in different types (Figure 2 and Figure 3). In order to explore the diversity of migratory patterns of C. nasus in both branches, the quantitative line was segmented different stages based on the trend of the otolith Sr/Ca ratios (Figure 2) and the color change of the Sr contents distribution (Figure 3). As shown in Table 1, there were significant differences between adjacent stages for the vast majority of individuals, except for N4, suggesting that there was a clear habitat transition between different stages, whereas the difference between stages 2 to 5 (B-F-B-F) for N4 was not significant, suggesting that frequent back and forth movements between F and B took place in the early stages of the migratory path for N4. From the changes Sr/Ca ratios and spectrum colors of different C. nasus in their migratory paths, we can see that there is a S stage (Sr/Ca ratios >7, red), of which are present in S1, S3 and S5 from the SB, and in N2, N3 and N4 from the NB. In terms of the number of transitions between different habitats, the frequency of transitions between F and B was higher for S5 (7 times) and S6 (6 times) from SB, and for N4 (10 times), N6 (8 times), N2 (6 times), and N7 (6 times) from NB, which indicated that some individuals from both branches were highly dependent on estuary brackish water habitat. The number of highly estuarine-dependent individuals (4) from NB and the transformation times in brackish water habitat of estuary (up to 10 times for N4) were higher than that from SB (the number of 2, and up to 7 times for S5), which shows that individuals from NB are more dependent on estuarine brackish habitats.
Based on the decomposition of the habitat history of C. nasus in Table 1 and the analysis of migration paths, it can be seen that between the two types of habitat history of C. nasus from both branches, Type I (F-B) was dominant, accounting for 62.5%. There were three migratory patterns in Type I for C. nasus from SB, with F-B-F-B-F was dominant (37.5%). There were four migratory patterns in type I for C. nasus from NB, with F-B was the most preferred (25.0%). In terms of the migratory path, the migratory starting point for C. nasus from both branches was F, and the end point of the habitat history included F and B, with F dominating for C. nasus from SB, accounting for 62.5%, and B dominating for C. nasus from NB, accounting for 87.5% (Table S1).
As shown in Figure 4, C. nasus from both branches have similar habitat histories, F-B and F-B-S. In terms of different migration patterns and paths, C. nasus had similar migration patterns in the same type of habitat history, but their migration paths were different. There is a difference in the starting point of migration path between SB and BN individuals. The starting point of the C. nasus from SB is far away from the Estuary, and its early life history has a longer time in freshwater. From the perspective of the transition between different habitats, the C. nasus from NB has a more frequent transition between F and B, and showed a greater dependence on brackish water in the estuary. In terms of the location and movement direction, the C. nasus form SB was mainly distributed in F when captured, with the movement direction shifting from B to F, showing an upward migratory movement trend, while that from NB was mainly distributed in B when captured, with the movement direction shifting from F to B, showing a reciprocal movement trend between B and F in the estuary.
3.3. Freshwater Dependence Analysis and Spawning Grounds Distribution Inference
The Lf and Fc can reflect the time of utilizing freshwater habitats for the C. nasus in its early life history, from hatching to entering the high salinity estuary, and reflect the distance between the spawning grounds in different sections of the Yangtze River to the Estuary [5,18]. As shown in Table S2, the Lf and Fc of different C. nasus were different, among which that of individuals from SB ranged from 300 to 1,210 μm, while that from NB ranged from 260 to 940 μm. The individuals with the longest Lf were distributed in SB, while that with the shortest in NB. The average Fc of C. nasus form SB is 0.36 and that from NB is 0.32, and the maximum Fc of the SB individuals is 0.63 and that of the NB individuals is 0.45. Analysis of the Lf and Fc of C. nasus from both branches showed that they can be clearly divided into different gradient groups of the long-distance freshwater dependence type (LD), the medium-distance freshwater-dependent type (MD) and the short-distance freshwater dependence type (SD) (Table S2), indicating that different individuals have different utilization times of F in the early stages of their life history. This not only reflects the different time from hatching to migration to the Estuary, but also reflects the different distances from the spawning ground in different sections of Yangtze to the Estuary [2,5,18]. As shown in Figure 5, there is a significant difference in Lf and Fc between different groups (P<0.05), while the difference between the same group is not significant (P>0.05), indicating that it is scientific and reliable to classify different freshwater dependence groups based on Lf and Fc. In terms of the composition of individuals with different freshwater-dependent types, there were three types of LD, MD and SD for C. nasus from SB, and two types of MD and SD for that from NB. Individuals from both branches had the MD, with three individuals from SB, accounting for 37.50%, and five individuals from NB, accounting for 62.50%. For MD individuals, the Lf and Fc of C. nasus from SB were 760.00±52.92 μm and 0.38±0.01, respectively, which were lower than those of C. nasus from NB with Lf and Fc were 786.00±105.97 μm and 0.42±0.04, respectively. Comparison is visible, the number, proportion, Lf and Fc of MD individuals from NB were higher than those from SB, and the MD individuals dominated in the NB. For the SD, the number of C. nasus individuals in both branches were three, and both accounted for 37.50%. In summary, the individuals of LD were distributed in SB, that of MD dominated in NB, and both branches contained the same proportion of SD.
Based on the analysis of the Lf of C. nasus otoliths from the Yangtze River Estuary, it was found that they could be clearly divided into three groups, with an average length of 1,135 µm in the longest group of LD, accounting for 12.5%, followed by MD, with an average length of 776 µm, accounting for 50.0%, and the shortest group of SD, with an average length of 295 µm, accounting for 37.5% (Figure 6A). In terms of the distribution of the different groups in SB and NB, LD were all distributed in SB, MD were mainly distributed in NB, and SD had the same proportion of distribution in both branches (Figure 6A). In terms of the composition of different groups, the individuals from SB included three types, with LD accounted for 25.00%, while that from NB included two types of MD and SD, and was dominated by MD with a predominance of 62.50% (Figure 6A). Since the Lf can visually reflect the distance of spawning grounds to the Estuary [5]. Corresponding analysis was conducted on the length of Lf and the distance of DYRE. It was found that there was an obvious correlation between Lf and DYRE (r=0.996). The Lf of LD group corresponds to DYRE of the spawning grounds near DTH (about 1,400 km away from the Estuary [5]), that MD group corresponds to that of the spawning grounds near PYH (about 800 km away from the Estuary [8]), and that the SD group corresponded to that of the spawning ground in the upper reaches of JY (> 220 km from the Estuary [1]) adjacent to the Yangtze River Estuary waters (Figure 6B).
4. Discussion
Earlier studies have focused on morphological features and geographical distribution to determine the ecological type of C. nasus [23]. Some studies have also used helminth marking, stable isotopes, molecular biology methods to distinguish the anadromous C. nasus from the non-anadromous C. nasus [24,25,26]. In recent years, the otolith microchemistry has been used to distinguish the freshwater sedentary C. nasus from the anadromous C. nasus more objectively, and the Sr content distribution can clearly distinguish the two ecologies [27,28]. In this study, all samples of C. nasus belonged to the long maxillary type (jaw length/head length ratio >1), and the Sr content diagrams showed that they all conformed to the physiological characteristics of the migratory type. There have been many studies on the migration habits of C. nasus in the Yangtze River. The C. nasus from Anqing section of the Yangtze River showed two types of migration, F-B and F-B-S [29], while the short-jawed C. nasus from Hexian section in Anhui Province showed two types of F and F-B, and the long-jawed C. nasus belonged to F-B-S [30]. The C. nasus from Nanjing section was anadromous and freshwater sedentary [31], while the C. nasus from Taizhou section in Jiangsu Province were all migratory [32]. The C. nasus from Jingjiang section were anadromous migratory long-jawed C. nasus as well as freshwater sedentary short-jawed C. nasus [33]. The results of the above study were highly compatible with the results of the present study, which showed that the habitat history of migratory C. nasus in the Yangtze River are classified into two types: F-B and F-B-S (Table S1 and Figure 4). In the aforementioned study, the C. nasus primarily originate from freshwater habitats in different sections of the Yangtze River, with habitat history types mainly including freshwater resident and upstream migratory types. However, for the C. nasus from the Yangtze River Estuary, their sources and distribution are relatively complex, involving various habitats such as F, B, and S. Currently, research on the habitat history and migratory patterns of C. nasus from the Yangtze River Estuary is relatively limited [17]. To better elucidate the migration patterns and upstream migration distances of C. nasus from different sources, this study conducted a detailed decomposition and quantitative analysis of the habitat history of C. nasus from SB and NB. This analysis identifies the composition of migration patterns and differences in migration paths of C. nasus from different branches of Yangtze River Estuary (Table S1, Figure 4).
In the life history of C. nasus, newly hatched fry leave freshwater to enter estuaries or even sea when they are less than one year old. After reaching maturity, most C. nasus anadromous migration in February-April, or as late as October, in search of suitable spawning grounds in the middle and lower reaches of the Yangtze River [1]. Previous studies have shown that most C. nasus have a freshwater origin, while a very few have a green core and originate from brackish waters [34]. Based on the starting values of Sr/Ca ratios, as well as the color of the otolith cores, all C. nasus in this research belonged to the freshwater origin and freshwater hatching type. Specifically, the early development of the C. nasus from the Yangtze River Estuary took place in freshwater, with spawning grounds potentially distributed in different freshwater areas along the Yangtze River. To further clarify the distribution pattern of spawning grounds for C. nasus from the Yangtze River Estuary, this study grouped C. nasus into three groups of LD, MD and SD based on the length of Lf to reflect the distance of DYRE [5]. A correspondence analysis was conducted between the length of Lf and the distance from traditional spawning grounds of C. nasus in various sections of the Yangtze River to the Estuary [3,4], revealing a clear correlation between these two (Figure 6). Analysis Lf of C. nasus from DTH showed an average length of 1,065 µm [5], consistent with that of LD group in this study (average of 1,135 µm). Therefore, it is speculated that the spawning grounds for LD group of C. nasus are distributed in DTH and nearby river sections (about 1,400 km from the Estuary in the middle reaches of the Yangtze River) [5]. For C. nasus from PYH and its connected river sections of Ganjiang, the shortest length of Lf is 650 µm [2], consistent with that in the MD group from this study. Hence, it is speculated that the spawning grounds for the MD group are located in PYH and adjacent river sections (about 800 km from the Estuary in the lower reaches of the Yangtze River) [2,8,35]. The average Lf of the SD group is 295 µm, suggesting that its spawning grounds are in the upstream of JY section (about 220 km from the Estuary), and near the Yangtze River estuary [1] (Figure 6).
This study confirms the existence of spawning grounds for C. nasus in different sections of the Yangtze River, highlighting the diversity of their spawning grounds distribution. C. nasus hatching from various spawning grounds may be at different developmental stages when they migrate to the Estuary, resulting in simultaneous presence of individuals with different life history stages such as larvae, juveniles, and adults in the estuarine waters. Each stage utilizes the habitat differently, and the Yangtze River Estuary can provide nursery, feeding, and migration pathways for C. nasus at different growth stages, reflecting the functional diversity of their habitat. The composition analysis of C. nasus from two branches show that the C. nasus from SB has LD type, while that from NB is primarily composed of MD type, with the same number of SD type individuals present in both branches. The differences in Lf among the various groups of C. nasus indicate that different early life history stages utilize freshwater habitats for varying durations, revealing different spatial distribution patterns of spawning grounds in different sections of the Yangtze River and highlighting the diversity of spawning ground distribution for C. nasus from the Yangtze River Estuary. By analyzing the composition of different freshwater-dependent types of C. nasus, it is found that the individuals migrated upstream to the middle reaches of the Yangtze River are distributed in SB, while individuals from both branches migrate to the lower reaches of the Yangtze River. The reproductive groups of C. nasus from the Yangtze River Estuary migrate upstream to spawning grounds in the middle and lower reaches of the Yangtze River, supporting the replenishment of the reproductive groups of C. nasus in various sections of the Yangtze River. Understanding the migration patterns and spawning ground distribution of C. nasus from the Yangtze River Estuary is of great theoretical and practical significance for the management of migration pathways, maintenance of spawning grounds, and precise resource control for C. nasus.
5. Conclusions
This study used otolith microchemistry to analyze the Sr/Ca ratios and Sr content distribution, obtaining habitat histories and migration patterns. Based on differences in Lf, it analyzed the upstream migration distances of C. nasus from the Yangtze River Estuary and the distribution patterns of their spawning grounds in different sections of the Yangtze River. The results indicate that C. nasus from both branches contain two types of habitat histories, both originating from freshwater. The C. nasus from SB exhibit six migration patterns, predominantly of F-B-F-B-F, while that from NB has seven migration patterns, mainly with F-B. In the migration paths of C. nasus from both branches, there are transitions between F and B, with the C. nasus from NB exhibiting more frequent back-and-forth transitions between these habitats. The C. nasus from SB shows a migration direction from B to F when captured, indicating an upstream migration trend, while that from NB show a migration direction from F to B, reflecting a back-and-forth migratory state within the estuarine brackish waters. The individuals from SB includes LD type, while that from NB is primarily composed of MD type, with the SD group equally distributed in both branches. The C. nasus from SB can migrate upstream to the spawning grounds near DTH, while the individuals from NB mainly migrate to the spawning grounds near PYH. This reflects the supporting role of C. nasus migrating upstream through the Yangtze River Estuary in replenishing the reproductive population in different sections of the Yangtze River, revealing the diversity of migration patterns and spawning ground distribution for C. nasus from different branches of the Yangtze River Estuary.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.Table S1: Habitat history and migration patterns of C. nasus from SB and NB of the Yangtze River Estuary; Table S2: Lf and Fc for C. nasus from SB and NB of the Yangtze River Estuary.
Author Contributions
Conceptualization and writing—original draft, C.S. and W.Y.; writing—review and editing, C.S., W.Y., F.Z. and F.L.; investigation and methodology, J.X. and R.L.; resources and funding acquisition, C.S., F.Z. and P.Z.; validation and supervision, F.L. and P. Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Key Research and Development Program of China (2022YFF0608203) and Central Public-interest Scientific Institution Basal Research Fund, CAFS (2023TD14).
Institutional Review Board Statement
The experiments comply with current laws in China. All the samples in this study were obtained from legal commercial fisheries, and the samples were dead when they were obtained.
Data Availability Statement
The data presented in this study are available in the article. Further information is available upon request from the corresponding author.
Acknowledgments
The authors acknowledge researcher Yang, J. and associate researcher Jiang, T. from the Key Laboratory of Fishery Ecological Environment Assessment and Resource Conservation in the Middle and Lower Reaches of the Yangtze River, Chinese Academy of Fishery Sciences for providing experimental instruments and processing the experimental materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhuang, P.; Zhang, T.; Li, S.F.; Ni, Y.; Wang, Y.H.; Deng, S.M.; Zhang, L.Z.; Ling, J.Z.; Hu, F.; Yang, G.; et al. Fishes of the Yangtze Estuary; China Agriculture Press: Beijing, China, 2018; pp. 124–127. [Google Scholar]
- Yang, Y.F.; Jiang, T.; Gao, X.P.; Xuan, Z.Y.; Chen, X.B.; Li, L.K.; Liu, H.B.; Yang, J. Discovery of anadromous Coilia nasus in the Ganjiang River, Lake Poyang Basin, China. J. Lake Sci. 2021, 33, 1595–1606. [Google Scholar]
- Yuan, C.M. Spawning migration of Coilia nasus. Bull. Biol. 1987, 12, 1–3. [Google Scholar]
- Zhu, D.L. Natural reproduction and observation of embryonic development of Coilia nasus from the Yangtze River. Fish. Sci. Technol. Inf. 1992, 19, 49–51. [Google Scholar]
- Xuan, Z.Y.; Jiang, T.; Liu, H.B.; Qiu, C.; Chen, X.B.; Yang, J. Are there still anadromous the estuarine tapertail anchovies Coilia nasus in Dongting lake? Acta Hydrob. Sin. 2020, 44, 838–843. [Google Scholar]
- Zhang, M.Y.; Xu, D.P.; Liu, K.; Shi, W.G. Studies on biological characteristics and change of resource of Coilia nasus schlegel in the lower research of the Yangtze River. Resour. Env. Yangtze Basin 2005, 14, 694–698. [Google Scholar]
- Li, Y.X.; He, W.P.; Liu, J.S.; Li, Z.J.; Xie, S.G. Annulus validation and age and growth estimation of anadromous Coilia ectenes in the Yangtze Estuary. Acta Hydrob. Sin. 2010, 34, 787–793. [Google Scholar] [CrossRef]
- Jiang, T.; Zhou, X.Q.; Liu, H.B.; Liu, H.Z.; Yang, J. Two microchemistry patterns in otoliths of Coilia nasus from Poyang Lake, China. J. Fish. China 2013, 37, 239–244. [Google Scholar] [CrossRef]
- Jiang, T.; Liu, H.B.; Xuan, Z.Y.; Chen, X.B.; Yang, J. Classification of ecomorphotypes of Coilia nasus from the middle and lower reaches of the Yangtze River Basin. J. Lake Sci. 2020, 32, 518–527. [Google Scholar]
- Hu, Y.H.; Jiang, T.; Liu, H.B.; Chen, X.B.; Yang, J. Habitat histories of different ecomorphotypes of Coilia nasus from the Yalu River in Dandong City of Liaoning Province based on otolith microchemical analysis. Mar. Fish. 2023, 45, 278–290. [Google Scholar]
- Xu, Q.; Ren, Q.Q.; Jiang, T.; Lin, B.A.; Jiang, X.B.; Yang, J.; Liu, M. Otolith microchemistry reveals diverse habitat uses and migratory patterns of two Coilia species (Engraulidae) in the Min River Estuary, southern China. Mar. Environ. Res. 2024, 193, 106296. [Google Scholar] [CrossRef]
- Zhao, Z.; Slypko, I.; Demianenko, K.; Zhu, G.P. Otolith chemistry reveals ontogenetic movement of the Antarctic toothfish (Dissostichus Mawsoni) in the Amundsen Sea Polynya, Antarctica. Fish. Res. 2024, 276, 107046. [Google Scholar] [CrossRef]
- Doerr, L.R.; Houghton, C.J.; Hansen, S.P.; Pangle, K.L.; Ransom, A.L.; Forsythe, P.S. Can otolith microchemistry identify the natal origin of larval lake whitefish Coregonus clupeaformis in the waters of Green Bay? J. Great Lakes Res. 2021, 47, 1771–1780. [Google Scholar] [CrossRef]
- Xu, Q.; Ren, Q.Q.; Jiang, T.; Jiang, C.R.; Fang, L.P.; Zhang, M.Z.; Yang, J.; Liu, M. Otolith microchemistry reveals various habitat uses and life histories of Chinese gizzard shad Clupanodon thrissa in the Min River and the estuary, Fujian Province, China. Fish. Res. 2023, 264, 106723. [Google Scholar] [CrossRef]
- Teichert, N.; Lizé, A.; Tabouret, H.; Roussel, J.M.; Bareille, G.; Trancart, T.; Acou, A.; Virag, L.S.; Pécheyran, C.; Carpentier, A.; et al. European flounder foraging movements in an estuarine nursery seascape inferred from otolith microchemistry and stable isotopes. Mar. Environ. Res. 2022, 182, 105797. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Guo, H.; Shen, H.; Li, X.; Tang, W.; Liu, J.; Jin, J.; Mi, Y. Preliminary results of Sr:Ca ratios of Coilia nasus in otoliths by Micro-PIXE. Nucl. Instrum. Meth. B. 2007, 260, 349–352. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.H.; Feng, G.P.; Yang, J.; Zhao, F.; Shen, C.C.; Song, C.; Jiang, T. Otolith microchemistry assessment: Evidence of migratory Coilia nasus of Yangtze River living in the Shengsi sea area. Fishes 2022, 7, 172. [Google Scholar] [CrossRef]
- Jiang, T.; Liu, H.B.; Shen, X.Q.; Shimasaki, Y.; Oshima, Y.; Yang, J. Life history variations among different population of Coilia nasus along the Chinese coast inferred from otolith microchemistry. J. Fac. Agr. Kyushu Univ. 2014, 59, 383–389. [Google Scholar] [CrossRef]
- Stirnimann, L.; Conversi, A.; Marini, S. Detection of regime shifts in the environment: testing “STARS” using synthetic and observed time series. ICES J. Mar. Sci. 2019, 76, 2286–2296. [Google Scholar] [CrossRef]
- Yang, J.; Arai, T.; Liu, H.; Miyazaki, N.; Tsukamoto, K. Reconstructing habitat use of Coilia mystus and Coilia ectenes of the Yangtze River estuary, and of Coilia ectenes of Taihu Lake, based on otolith strontium and calcium. J. Fish. Biol. 2006, 69, 1120–1135. [Google Scholar] [CrossRef]
- Kotake, A.; Arai, T.; Ozawa, T.; Nojima, S.; Miller, M.J.; Tsukamoto, K. Variation in migratory history of Japanese eels, Anguilla japonica, collected in coastal waters of the Amakusa Islands, Japan, inferred from otolith Sr/Ca ratios. Mar. Biol. 2003, 142, 849–854. [Google Scholar] [CrossRef]
- Sokta, L.; Jiang, T.; Liu, H.B.; Xuan, Z.Y.; Qiu, C.; Chen, X.B.; Yang, J. Loss of Coilia nasus habitats in Chinese freshwater lakes: An otolith microchemistry assessment. Heliyon 2020, 6, e04571. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.M.; Lin, J.B.; Qin, A.Z.; Liu, R.H. On the history and present situation of the taxonomy of Coilia in China——Some experiences on reforming the old taxonomy of fishes. J. Nanjing Univ. 1976, 2, 1–13. [Google Scholar]
- Li, W.X.; Wang, G.T. Helminth communities in Coilia nasus from anadromous freshwater and landlocked stocks. Chinese J. Zool. 2014, 49, 233–243. [Google Scholar]
- Wang, L.; Tang, W.Q.; Dong, W.X. The signatures of stable isotopes δ15N and δ13C in anadromous and non-anadromous Coilia nasus living in the Yangtze River, and the adjacent sea waters. J. Ocean Univ. China 2015, 14, 1053–1058. [Google Scholar] [CrossRef]
- Cheng, F.Y.; Wang, Q.; Delser, P.M.; Li, C.H. Multiple freshwater invasions of the tapertail anchovy (Clupeiformes: Engraulidae) of the Yangtze River. Ecol. Evol. 2019, 9, 12202–12215. [Google Scholar] [CrossRef]
- Xuan, Z.Y.; Jiang, T.; Liu, H.B.; Yang, J. Otolith microchemistry and microsatellite DNA provide evidence for divergence between estuarine tapertail anchovy (Coilia nasus) populations from the Poyang Lake and the Yangtze River Estuary of China. Reg. Stud. Mar. Sci. 2022, 56, 102649. [Google Scholar] [CrossRef]
- Xuan, Z.Y.; Jiang, T.; Liu, H.B.; Chen, X.B.; Yang, J. Otolith microchemical evidence revealing multiple spawning site origination of the anadromous tapertail anchovy (Coilia nasus) in the Changjiang (Yangtze) River Estuary. Acta Oceanol. Sin. 2023, 42, 120–130. [Google Scholar] [CrossRef]
- Li, M.M.; Jiang, T.; Chen, T.T.; Liu, H.B.; Yang, J. Otolith microchemistry of the estuarine tapertail anchovy Coilia nasus from the Anqing section of the Yangtze River and its significance for migration ecology. Acta Ecol. Sin. 2017, 37, 2788–2795. [Google Scholar]
- Li, M.M.; Jiang, T.; Khumbanyiwa, D.D.; Liu, H.B. , Yang, J. Reconstructing habitat history of Coilia nasus from the Hexian section of the Yangtze River in Anhui province by otolith microchemistry. Acta Hydrob. Sin. 2017, 41, 1054–1061. [Google Scholar]
- Chen, T.T.; Jiang, T.; Li, M.M.; Liu, H.B.; Yang, J. Inversion of habitat history for the long-jaw ecotype Coilia nasus collected from Nanjing section of the Yangtze River. J. Fish. China 2016, 40, 882–892. [Google Scholar]
- Hu, Y.H.; Jiang, T.; Liu, H.B.; Chen, X.B.; Yang, J. Otolith microchemical fingerprints of Coilia nasus from the Taizhou section of Changjiang River in Jiangsu Province. Chinese J. Ecol. 2024, 43, 967–974. [Google Scholar]
- Chen, T.T.; Jiang, T.; Lu, M.J.; Liu, H.B.; Yang, J. Microchemistry analysis of otoliths of Coilia nasus and Coilia brachygnathus from the Jingjiang section of the Yangtze River. J. Lake Sci. 2016, 28, 149–155. [Google Scholar]
- Liu, H.B.; Jiang, T.; Xuan, Z.Y.; Qiu, C.; Yang, J. Otolith microchemical analysis of Tapertail Anchovy Coilia nasus from Ariake sea and its adjacent tributaries in Japan. Fish. Sci. 2020, 39, 500–508. [Google Scholar]
- Jiang, T.; Yang, J.; Lu, M.J.; Liu, H.B.; Chen, T.T.; Gao, Y.W. Discovery of a spawning area for anadromous Coilia nasus Temminck et Schlegel, 1846 in Poyang Lake, China. J. Appl. Ichthyol. 2017, 33, 189–192. [Google Scholar] [CrossRef]
Figure 2.
Fluctuation in otolith Sr /Ca ratios along line transects from the core (0 µm) to the edge in the otoliths of C. nasus from SB and NB of the Yangtze River Estuary. Blue solid line represents the Sr/Ca ratios after trend conversion; light blue solid lines represent actual Sr/Ca ratios. Red dotted line represent border between seawater (S) and brackish water (B). Green dotted line represent border between B and freshwater (F).
Figure 2.
Fluctuation in otolith Sr /Ca ratios along line transects from the core (0 µm) to the edge in the otoliths of C. nasus from SB and NB of the Yangtze River Estuary. Blue solid line represents the Sr/Ca ratios after trend conversion; light blue solid lines represent actual Sr/Ca ratios. Red dotted line represent border between seawater (S) and brackish water (B). Green dotted line represent border between B and freshwater (F).
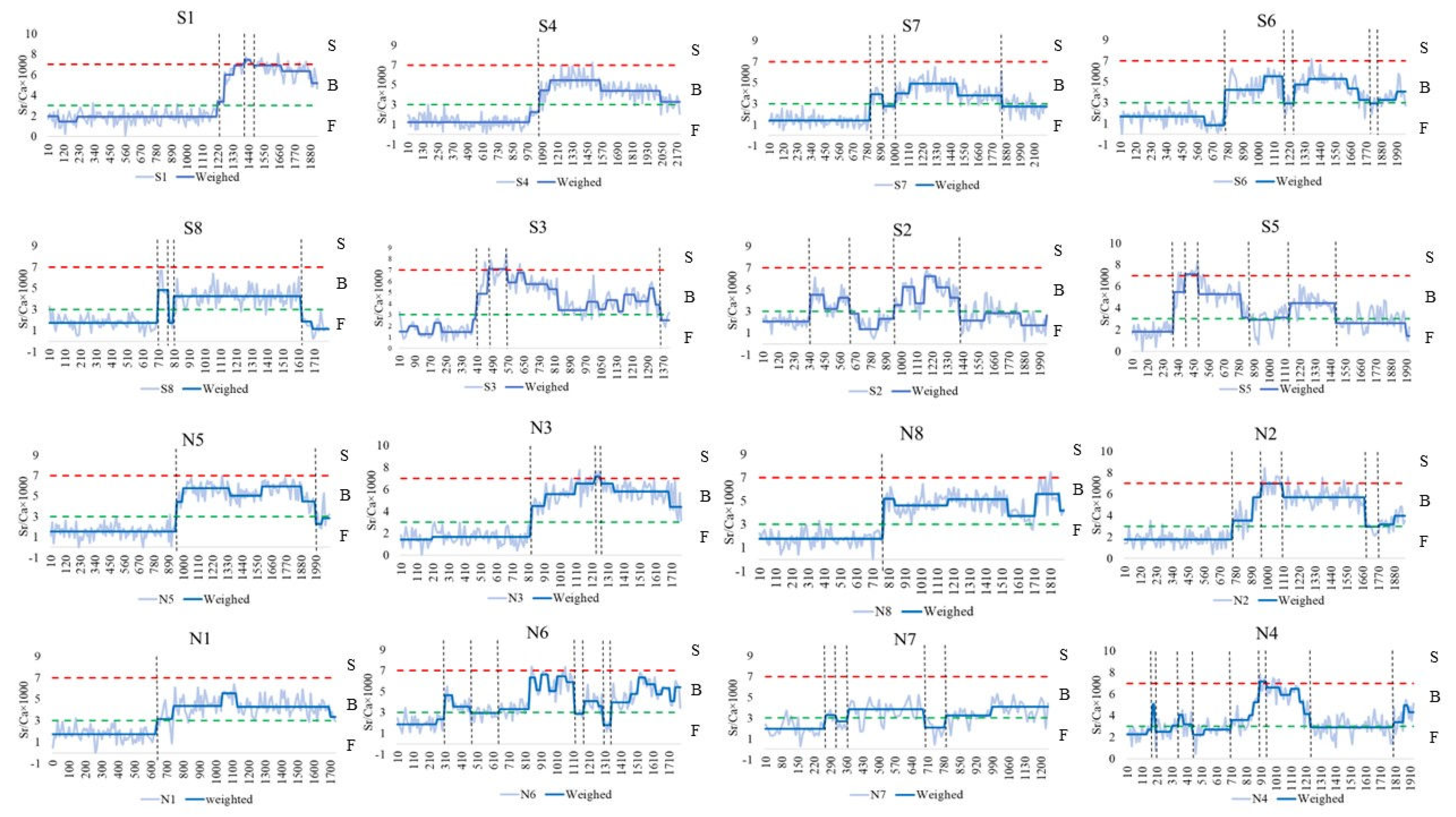
Figure 3.
Two-dimensional imaging using X-ray electron microprobe analysis of the Sr contents in otoliths of C. nasus from SB and NB of the Yangtze River Estuary. The values corresponding to Sr contents are represented by 17 colors from blue (lowest), through green and yellow, to red (highest).
Figure 3.
Two-dimensional imaging using X-ray electron microprobe analysis of the Sr contents in otoliths of C. nasus from SB and NB of the Yangtze River Estuary. The values corresponding to Sr contents are represented by 17 colors from blue (lowest), through green and yellow, to red (highest).
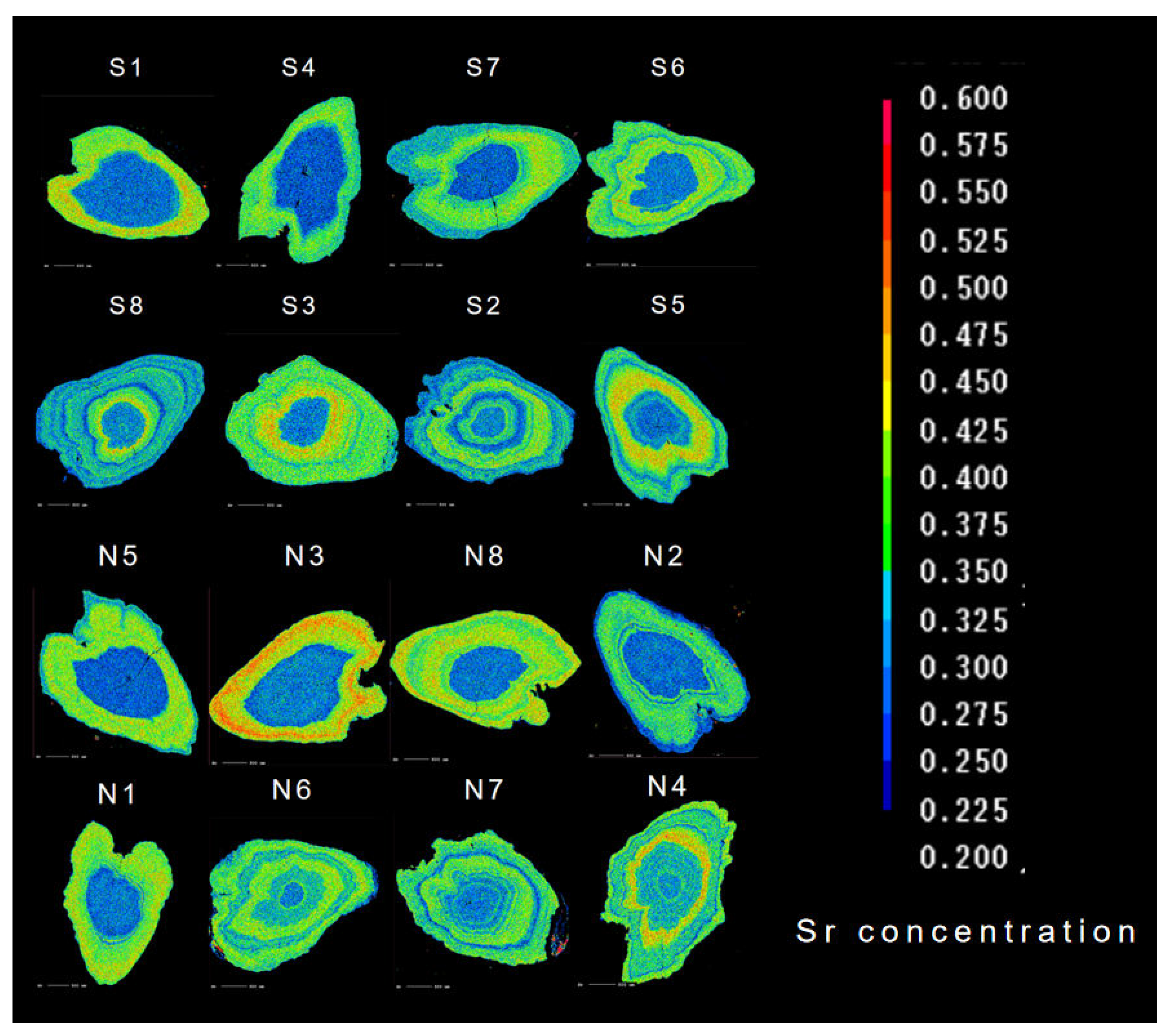
Figure 4.
Migration path simulation of C. nasus from SB and NB of the Yangtze River Estuary. The white closed loop line represents the migratory path of SB individuals, the black closed loop line represents the migratory path of NB individuals, the blue arrow indicates the direction of the start of the migratory path, and the red arrow indicates the direction of the end of the migratory path.
Figure 4.
Migration path simulation of C. nasus from SB and NB of the Yangtze River Estuary. The white closed loop line represents the migratory path of SB individuals, the black closed loop line represents the migratory path of NB individuals, the blue arrow indicates the direction of the start of the migratory path, and the red arrow indicates the direction of the end of the migratory path.
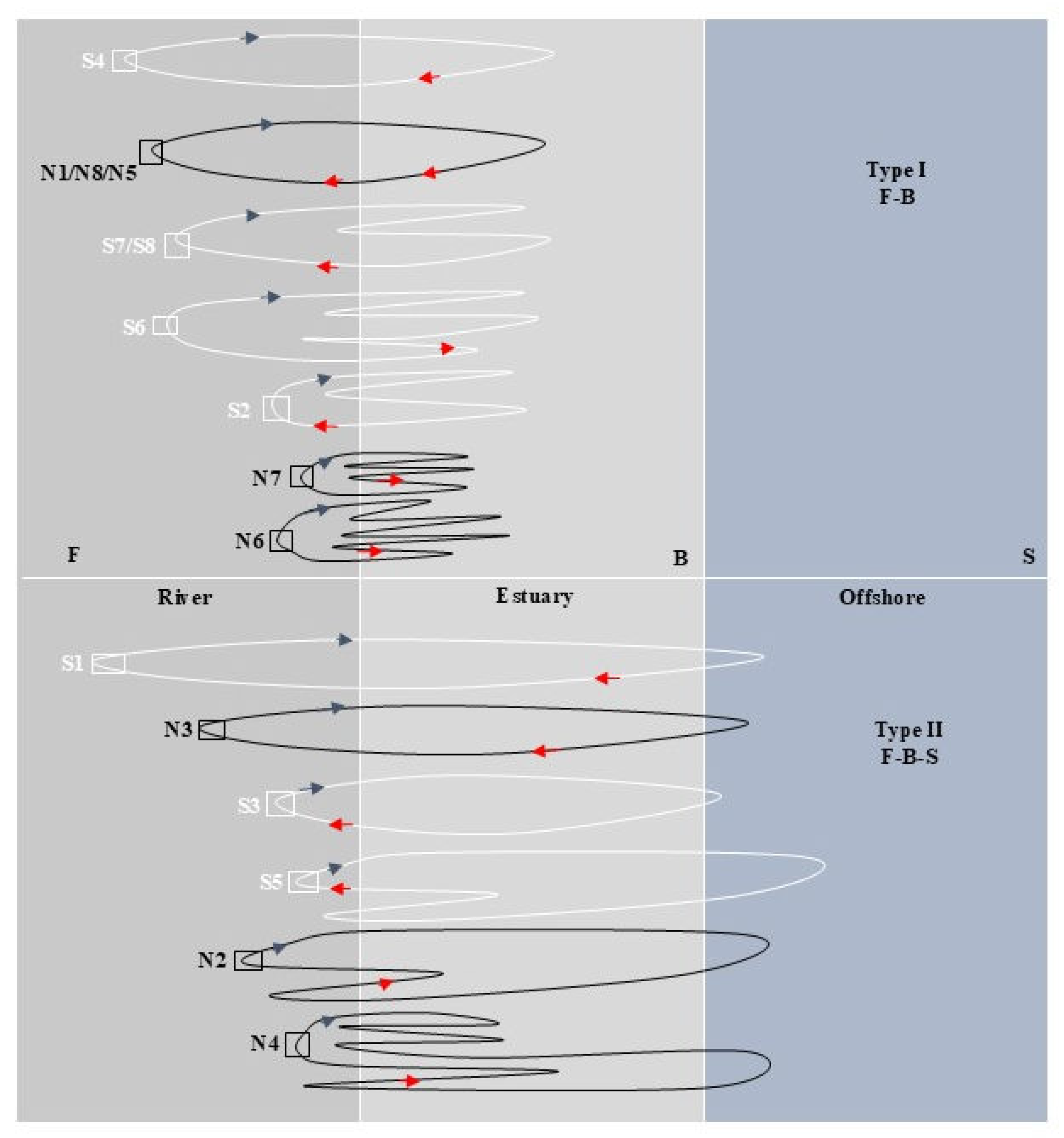
Figure 5.
Comparison of Lf and Fc of different freshwater dependent types of LD, MD and SD for C. nasus from SB and NB of the Yangtze River Estuary. Lf: the radius of the first blue region near the core, Fc: freshwater coefficient; LD: long-distance freshwater dependence type, MD: medium-distance freshwater-dependent type, SD: short-distance freshwater dependence type; Different large letters (A, B, C) indicate that there are significant differences among different groups of LD, MD and SD (P<0.05), while the same small letters (a) indicate that there are no significant differences for the same group between the individuals from SB and NB (P>0.05).
Figure 5.
Comparison of Lf and Fc of different freshwater dependent types of LD, MD and SD for C. nasus from SB and NB of the Yangtze River Estuary. Lf: the radius of the first blue region near the core, Fc: freshwater coefficient; LD: long-distance freshwater dependence type, MD: medium-distance freshwater-dependent type, SD: short-distance freshwater dependence type; Different large letters (A, B, C) indicate that there are significant differences among different groups of LD, MD and SD (P<0.05), while the same small letters (a) indicate that there are no significant differences for the same group between the individuals from SB and NB (P>0.05).
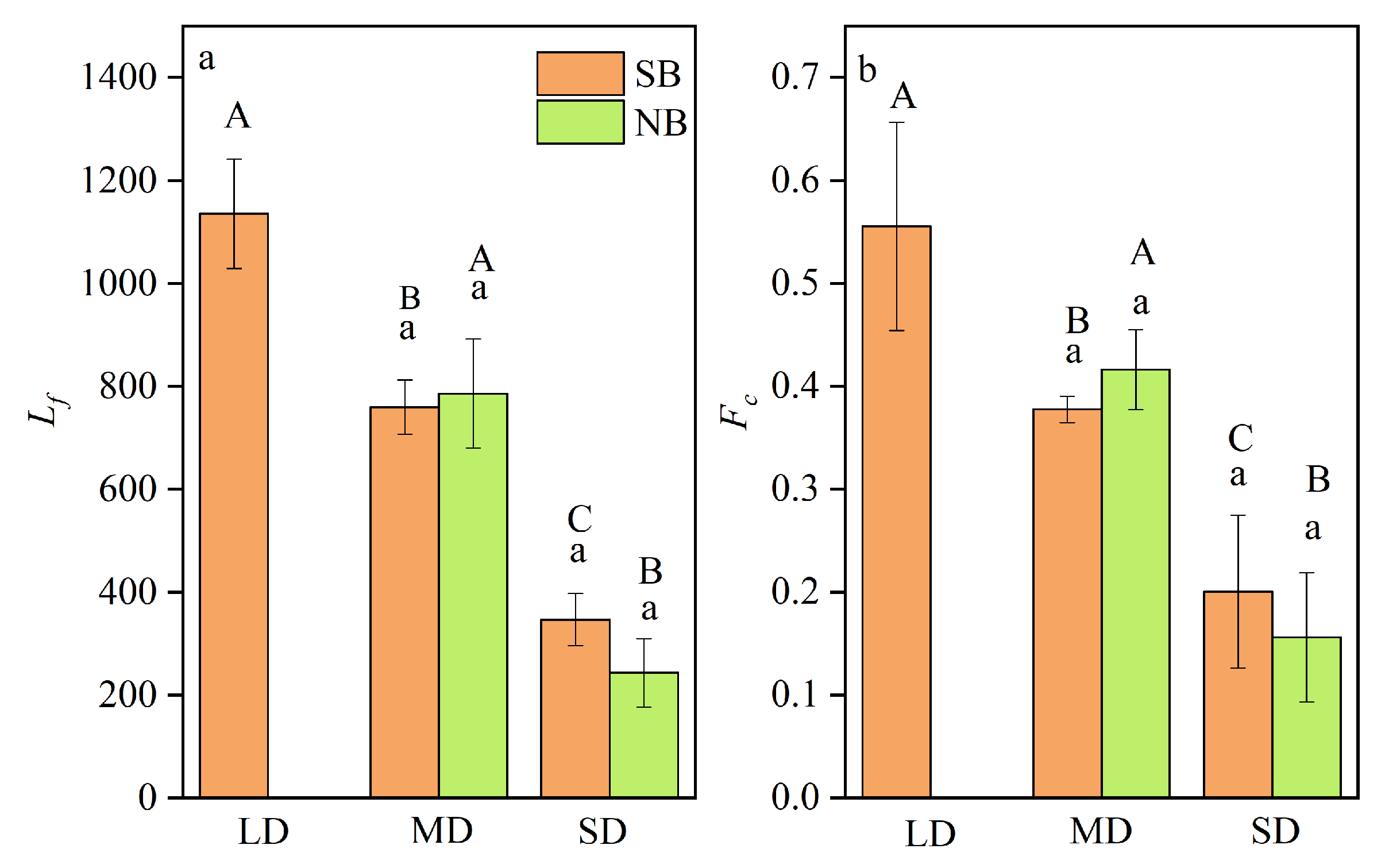
Figure 6.
Correspondence between Lf and DYRE. Lf, LD, MD, SD are the same with that in Figure 5. DYRE: the distance from the spawning grounds of C. nasus in different sections of the Yangtze River to the Estuary; YC, JJ, JY show the Yichang (YC), Jiujiang (JJ) and Jiangyin (JY) sections in the middle and lower reaches of the Yangtze River; JGJ shows the reef of Jigujiang (JGJ) in the Yangtze River Estuary (YRE). DTH, PYH show the lakes of Dongtinghu (DTH) and Poyanghu (PYH).
Figure 6.
Correspondence between Lf and DYRE. Lf, LD, MD, SD are the same with that in Figure 5. DYRE: the distance from the spawning grounds of C. nasus in different sections of the Yangtze River to the Estuary; YC, JJ, JY show the Yichang (YC), Jiujiang (JJ) and Jiangyin (JY) sections in the middle and lower reaches of the Yangtze River; JGJ shows the reef of Jigujiang (JGJ) in the Yangtze River Estuary (YRE). DTH, PYH show the lakes of Dongtinghu (DTH) and Poyanghu (PYH).
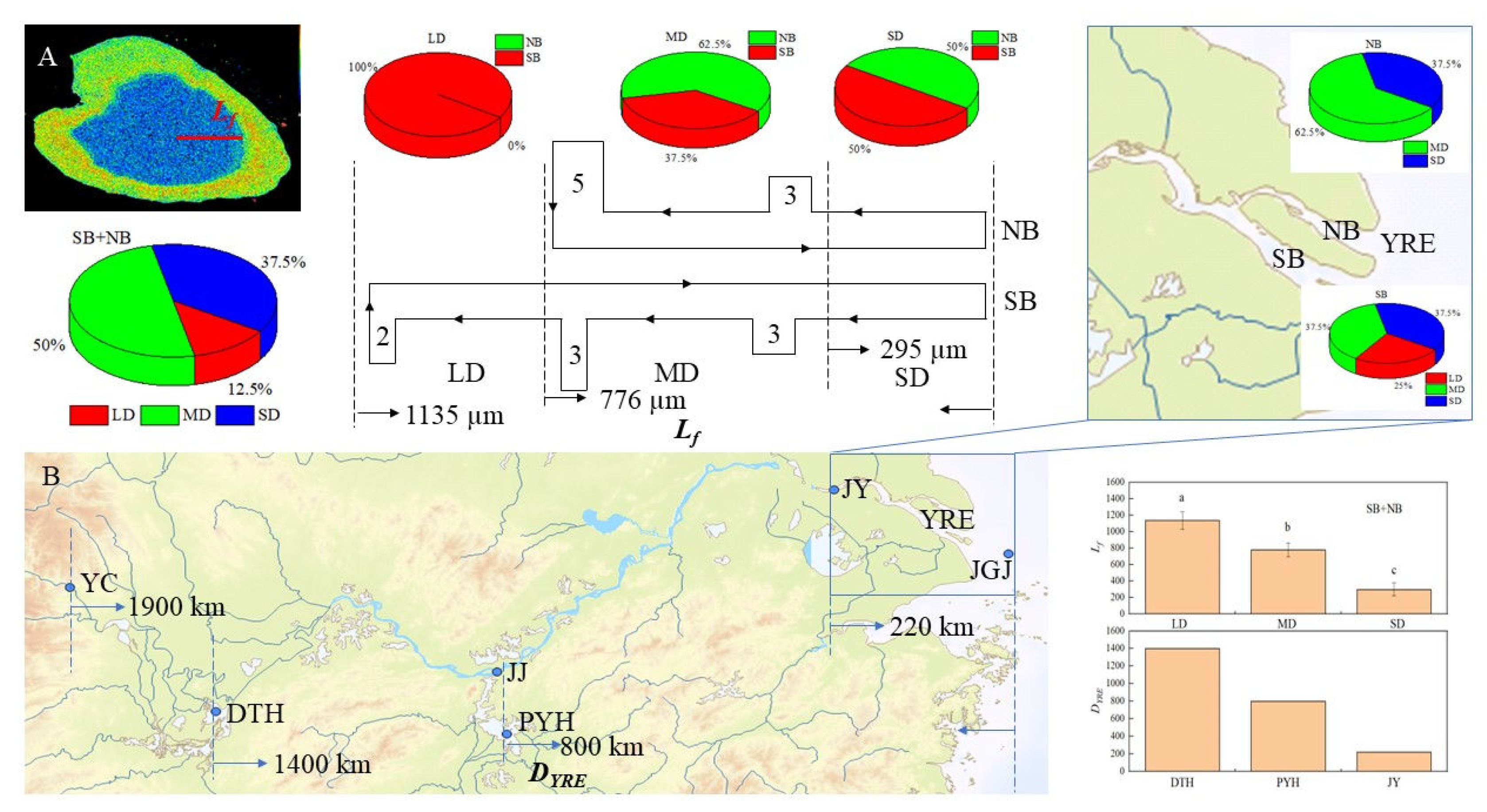

Table 1.
Changes in Sr/Ca ratios in otolith of C. nasus from SB and NB of Yangtze River Estuary.
| Water Area | Sample Number | Change Stages | Spectrum | Length from Otolith Core Radius/µm | Number of Element Points (N) | Sr/Ca Ratios(Mean ± SD) |
|---|---|---|---|---|---|---|
| SB | S4 | 1(F) | Blue | 0-1060 | 107 | 1.35±0.60a |
| 2(B) | Green-Yellow | 1070-2190 | 113 | 4.62±1.06b | ||
| S2 | 1(F) | Blue | 0-340 | 35 | 2.05±0.45a | |
| 2(B) | Green-Yellow | 350-650 | 31 | 3.92±0.89b | ||
| 3(F) | Blue | 660-940 | 29 | 1.89±0.73c | ||
| 4(B) | Green-Yellow | 950-1400 | 46 | 4.85±1.12d | ||
| 5(F) | Blue | 1410-2040 | 64 | 2.37±0.95a | ||
| S7 | 1(F) | Blue | 0-800 | 81 | 1.41±0.52a | |
| 2(B) | Green-Yellow | 810-900 | 10 | 3.88±0.53b | ||
| 3(F) | Blue | 910-990 | 9 | 2.73±0.35c | ||
| 4(B) | Green-Yellow | 1000-1840 | 85 | 4.31±0.89b | ||
| 5(F) | Blue | 1850-2200 | 36 | 2.70±0.67c | ||
| S8 | 1(F) | Blue | 0-700 | 71 | 1.72±0.60ac | |
| 2(B) | Green-Yellow | 710-770 | 7 | 5.11±1.09b | ||
| 3(F) | Blue | 780-820 | 5 | 2.32±0.95c | ||
| 4(B) | Green-Yellow | 830-1620 | 80 | 4.31±0.80d | ||
| 5(F) | Blue | 1630-1800 | 18 | 1.48±0.71a | ||
| S6 | 1(F) | Blue | 0-780 | 78 | 1.53±0.65a | |
| 2(B) | Green-Yellow | 790-1170 | 39 | 4.71±0.96b | ||
| 3(F) | Blue | 1180-1250 | 8 | 2.66±0.63c | ||
| 4(B) | Green-Yellow | 1260-1790 | 54 | 4.76±1.00b | ||
| 5(F) | Blue | 1800-1850 | 6 | 2.67±0.52c | ||
| 6(B) | Green-Yellow | 1860-2050 | 20 | 3.71±0.79d | ||
| S1 | 1(F) | Blue | 0-1210 | 122 | 1.85±0.59a | |
| 2(B) | Green-Yellow | 1220-1400 | 19 | 5.57±1.45b | ||
| 3(S) | Red | 1410-1470 | 7 | 7.38±0.21c | ||
| 4(B) | Green-Yellow | 1480-1930 | 46 | 6.43±0.75d | ||
| S3 | 1(F) | Blue | 0-400 | 41 | 1.63±0.62a | |
| 2(B) | Green-Yellow | 410-470 | 7 | 5.67±1.31b | ||
| 3(S) | Red | 480-560 | 9 | 7.03±0.80c | ||
| 4(B) | Green-Yellow | 570-1350 | 79 | 4.62±1.21d | ||
| 5(F) | Blue | 1360-1400 | 5 | 2.64±0.50e | ||
| S5 | 1(F) | Blue | 0-300 | 30 | 1.87±0.60a | |
| 2(B) | Green-Yellow | 310-410 | 11 | 5.93±1.32b | ||
| 3(S) | Red | 420-480 | 7 | 7.45±0.39c | ||
| 4(B) | Green-Yellow | 490-840 | 36 | 5.26±0.95d | ||
| 5(F) | Blue | 850-1030 | 19 | 2.53±0.95e | ||
| 6(B) | Green-Yellow | 1040-1490 | 46 | 4.18±0.84f | ||
| 7(F) | Blue | 1500-2010 | 52 | 2.88±0.86e | ||
| NB | N1 | 1(F) | Blue | 0-650 | 65 | 1.78±0.64a |
| 2(B) | Green-Yellow | 660-1730 | 108 | 4.34±0.93b | ||
| N8 | 1(F) | Blue | 0-770 | 78 | 1.77±0.61a | |
| 2(B) | Green-Yellow | 780-1890 | 112 | 4.80±0.94b | ||
| N5 | 1(F) | Blue | 0-940 | 93 | 1.57±0.57a | |
| 2(B) | Green-Yellow | 950-1980 | 104 | 5.43±0.80b | ||
| 3(F) | Blue | 1990-2090 | 11 | 2.99±1.05c | ||
| N7 | 1(F) | Blue | 0-260 | 27 | 2.02±0.75a | |
| 2(B) | Green-Yellow | 270-310 | 5 | 3.26±0.40b | ||
| 3(F) | Blue | 320-360 | 5 | 2.12±0.30a | ||
| 4(B) | Green-Yellow | 370-690 | 33 | 3.75±0.91b | ||
| 5(F) | Blue | 700-780 | 9 | 1.99±0.80a | ||
| 6(B) | Green-Yellow | 790-1230 | 45 | 3.63±0.86b | ||
| N6 | 1(F) | Blue | 0-300 | 31 | 1.99±0.62a | |
| 2(B) | Green-Yellow | 310-460 | 16 | 3.91±0.70b | ||
| 3(F) | Blue | 470-650 | 19 | 2.83±0.52a | ||
| 4(B) | Green-Yellow | 660-1110 | 46 | 4.94±1.50b | ||
| 5(F) | Blue | 1120-1170 | 6 | 2.88±0.24a | ||
| 6(B) | Green-Yellow | 1180-1270 | 10 | 4.08±0.72b | ||
| 7(F) | Blue | 1280-1340 | 7 | 2.36±1.21a | ||
| 8(B) | Green-Yellow | 1350-1780 | 44 | 4.92±1.01b | ||
| N3 | 1(F) | Blue | 0-820 | 82 | 1.66±0.62a | |
| 2(B) | Green-Yellow | 830-1220 | 40 | 5.55±1.04b | ||
| 3(S) | Red | 1230-1260 | 4 | 7.37±0.48c | ||
| 4(B) | Green-Yellow | 1270-1770 | 51 | 5.72±0.93b | ||
| N2 | 1(F) | Blue | 0-750 | 76 | 1.80±0.62a | |
| 2(B) | Green-Yellow | 760-970 | 22 | 4.41±1.44b | ||
| 3(S) | Red | 980-1080 | 11 | 7.05±0.81c | ||
| 4(B) | Green-Yellow | 1090-1680 | 60 | 5.75±0.79d | ||
| 5(F) | Blue | 1690-1770 | 9 | 2.62±0.37e | ||
| 6(B) | Green-Yellow | 1780-1950 | 18 | 3.55±0.70f | ||
| N4 | 1(F) | Blue | 0-170 | 18 | 2.32±0.53a | |
| 2(B) | Green-Yellow | 180-220 | 5 | 3.52±1.76bcf | ||
| 3(F) | Blue | 230-340 | 12 | 2.60±0.56ab | ||
| 4(B) | Green-Yellow | 350-440 | 10 | 3.46±0.86bcf | ||
| 5(F) | Blue | 450-700 | 26 | 2.66±0.63ab | ||
| 6(B) | Green-Yellow | 710-890 | 19 | 3.82±1.11cf | ||
| 7(S) | Red | 900-940 | 5 | 7.05±0.70d | ||
| 8(B) | Green-Yellow | 950-1260 | 32 | 5.68±1.31e | ||
| 9(F) | Blue | 1270-1820 | 56 | 2.92±0.82abc | ||
| 10(B) | Green-Yellow | 1830-1930 | 11 | 4.20±0.86f |
Note: Different superscript letters (a, b, c, d, e, f) indicated significant differences (P<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



