
Submitted:
07 August 2024
Posted:
27 August 2024
You are already at the latest version
Abstract
Diagenetic spheroids are rounded mineralised structures in sedimentary rocks that span several orders of size dimensions. These objects include rosettes spanning tens of micrometres, millimetre-sized granules, and centimetre- to metre-scale botryoids, nodules and concretions. Connections between diagenetic spheroids and chemically oscillating reactions (COR) have been established based on chemical and pattern similarities. Nevertheless, it is unknown if there are variations in their occurrences throughout geological time, especially during periods of global environmental change. This is because COR are pattern-forming reactions where carboxylic acids are decarboxylated by strong oxidants like sulphuric acid, bromate, or iodate. The hypothesis is that there should be a greater number of diagenetic spheroids at periods of increased atmospheric-oceanic oxygenation levels. Hence, this work presents the first compilation of occurrences of diagenetic spheroids between 0.541 to 1.8 billion years ago (Ga). The compiled patterned objects are those with concentric laminations and radially aligned crystals, which are patterns akin to equidistant circularly concentric waves that radially propagate in COR. Another characteristic of diagenetic spheroids, such as in botryoidal minerals, is the destructive interfere that forms circular twins; this is identical to the COR pattern formed when circular waves meet. The timeline of Proterozoic diagenetic spheroids produced in this work reveals a significant increase of occurrences across the Ediacaran, which is attributed to increased environmental oxidation state. During this interval, seawater is known to have become richer in sulphate and iodate, which create ideal conditions for the abiotic and diagenetic oxidation of biomass. Therefore, increased occurrences of diagenetic spheroids in the Neoproterozoic possibly represents sedimentological evidence for abiotic decarboxylation reactions and widespread COR in productive environments. The distribution of diagenetic spheroids observed during the Proterozoic may also be more broadly applicable to other epochs of Earth history.

Keywords:
concretion
; botryoid
; diagenesis
; carbon cycle
; fossilisation
; Belousov-Zhabotinsky
1. Introduction
Diagenesis describes the biological, chemical and physical processes that alter deposited sedimentary assemblages before complete lithification (Curtis, 1977; Burley et al., 1985). Concretions, nodules, botryoids, granules and rosettes are mineralised spheroidal structures that are prevalent in the sedimentological record and usually relate to diagenesis. These objects can display circularly concentric and radial mineral patterns, a limited dominant mineralogy, and common association with organic matter. They are also considered as abiotic in origin, hence they are here grouped as ‘diagenetic spheroids’. Diagenetic spheroids are distinguished according to size ranges. Mega-concretions can have a diameter as large as several metres, and rosettes are the smallest type with diameters <10-4 m (Gabriel et al., 2021). Both concretions and nodules have sizes from 10-2 m to 10-1 m. Whereas the mineralogy of concretions is akin to its host rock, the composition of nodules differs from their surrounding sediments (Mitsuchi, 1976). Granules are irregular spheroids that range between 10-2 m and 10-4 m in diameter. Botryoidal minerals include interconnected, or twinned, smooth spheroids, and their globular texture is grape-like, mammillary, or reniform in appearance (Papineau, 2020). Botryoids span a large diameter range from 10-1 m to 10-6 m.
Despite the array of descriptive terms, these objects commonly exhibit a sub-rounded morphology with circularly concentric and radially aligned geometric patterns (Papineau et al., 2017; 2021; Papineau, 2024). Occasionally, they exhibit a triaxial ellipsoidal or lenticular morphology. The term diagenetic spheroids groups those objects with identical range of patterns together, which may imply a similar sedimentary-diagenetic history (Papineau et al., 2017; 2021; 2023; Dodd et al., 2018; Papineau, 2020; 2024). Typical mineral compositions include microcrystalline quartz (chert, chalcedony) (Varkouhi et al., 2022; Varkouhi and Papineau, 2023), dolomite, pyrite, apatite and/or organic matter (Papineau et al., 2017; Gabriel et al., 2021). Fossils may also be encapsulated in diagenetic spheroids and usually have a comparable mineral composition. For instance, well-preserved Ediacaran microfossils, like Vendotaenia sp. and Oscillatoriopis obtusa, have been identified within chert concretions in the Shibantan Lagerstätte (Mason et al., 2017; Ding et al., 2019; Xiao et al., 2020).
The formation mechanism of diagenetic spheroids has yet to reach consensus. For instance, Dodd et al. (2018) disputed the wave-action model for granules in granular iron formation on the basis that the arrangement of fine equidistant, circularly concentric laminations and radially aligned acicular crystals are improbable under turbulence. Moreover, shallow marine wave agitation cannot explain the common association of granules with organic matter and well-preserved microfossils. Diagenetic spheroids are considered abiotic precipitates and are therefore possible indicators of physicochemical processes (Davies et al., 1978; Flannery et al., 2019). However, nucleation-controlled growth around a central mass (Lebron and Suarez, 1996; Fouke, 2011) is more likely to generate irregular clumps, rather than equidistant concentric laminations with density gradients. The Liesegang phenomenon is another abiotic process which is the purported origin of diagenetic spheroids (Liesegang, 1910), but it is unable to explain regular patterns of circular laminations (Papineau et al., 2021). Liesegang banding is produced by the precipitation and diffusion of metal cations within silica gels (Liesegang, 1910; Keller and Rubinow, 1981). Moreover, Liesegang rings have a gravity-driven linear arrangement (Sultan and Abdel-Rahman, 2013; Nabika et al., 2019), in contrast with concentric rings observed in diagenetic spheroids.
Alternatively, Bosak et al. (2010) proposed that diagenetic spheroids associated with microbialites and stromatolites may be mineralised gas bubbles generated by photosynthetic microorganisms. Internal features, such as mineral inclusions and organic matter, are absent within bubbles (Bosak et al., 2010). The gas-bubble theory therefore does not explain the occurrence of microfossils and concentric laminations in some diagenetic spheroids (Gabriel et al., 2021). Occurrences of microfossils, degraded organic matter, or highly variable carbon and sulphur isotope compositions in diagenetic spheroids have been interpreted to suggest a biologically mediated formation (Dahanayake et al., 1985; Dahanayake and Krumbein, 1986; Glasauer et al., 2013; Salama et al., 2012; Mason et al., 2017). For example, organic matter could act as nucleation sites, and mineral precipitation may be actively induced by microbial interaction with its niche (Diaz and Eberli, 2019).
COR have been proposed as an alternative formation model to explain the circularly concentric, radially aligned, and density gradient patterns often found in diagenetic spheroids (Papineau et al., 2016, 2017, 2021; Dodd et al., 2018; Gabriel et al., 2021; Varkouhi et al., 2022; Varkouhi and Papineau, 2023, Papineau, 2020; 2024). Zaikin and Zhabotinsky (1970) report that “radial circular waves of oxidation were propagated” when phenanthroline ferrous sulphate (ferroin) was added to an aqueous solution of bromate, bromide, malonic acid and sulphuric acid. Interference occurs when concentric waves from different initiation points intersect, producing ‘interconnected round arcs’ (or circular twins), known as Belousov–Zhabotinsky (B-Z) patterns (Zaikin and Zhabotinsky, 1970). B-Z reactions in Petri dishes display regular wave patterns and the effects of chemical turbulence. These circular waves span several size dimensions and are hence fractal patterns. In nature, abiotic carboxylic acids could be central to this reaction but alternatively, there are implications for biosignatures if the carboxylic acids are derived from biomass (Gabriel et al., 2021). The radially propagating circles have been frequently recreated and analysed in experimental settings (Briggs and Rauscher, 1973; Körös and Orbán, 1978; Epstein et al., 1983; Agladze et al., 1984; Field and Schneider, 1989; Zhabotinsky, 1991; Belmonte et al., 1997; Orbán et al., 2001; Chen et al., 2011; Papineau et al., 2017). Nevertheless, it was only recently proposed (Papineau, 2020; 2024; Papineau et al., 2021) that the patterns and products of COR are analogous with those of diagenetic spheroids. If COR occur naturally in the environment and indeed represent environmental proxies (Varkouhi and Papineau, 2023), then their occurrences might preserve evidence of global biogeochemical changes.
The Proterozoic spans from 2.5 to 0.541 Ga and includes a prolonged biogeochemical stasis between 1.8 and 0.8 Ga, dubbed the ‘Boring Billion’ (Brasier and Lindsay, 1998), or, perhaps less anthropocentrically, the “Balanced Billion” (Mitchell and Evans, 2024). This interval is characterised by tectonic and environmental stability, with steady isotopic compositions of carbon, sulphates and sulphides observed in marine sediments (Mukherjee et al., 2018). By contrast, the Neoproterozoic underwent large climatic variations due to major tectonic events like the Rodinian fragmentation and Gondwanan formation (Merdith et al., 2017), as well as major glaciations. Cap carbonates, which are sedimentary sequences that record deglaciations, have calcite and dolomite with strong depletions in 13C. These have been variably interpreted as evidence for the oxidation of dissolved or suspended organic matter in the water column (Fike et al., 2006), local methane seeps (Jiang et al., 2003), associated with anaerobic oxidation of methane (Bristow et al., 2011), or diagenetic oxidation of biomass in sediments (Ader et al., 2009). Therefore, a timeline of the abundances of diagenetic spheroids in geological strata between 1.8 Ga (start of the ‘Balanced Billion’) and 0.541 (Ediacaran-Cambrian boundary) may provide further insight into whether diagenetic spheroids are more likely to form under stable long-term geochemical conditions or dynamic ones, respectively. The emerging hypothesis is that an increased abundance of diagenetic spheroids near the end of the Neoproterozoic would be expected according to increased environmental oxidation because enhanced continental weathering provides an increased supply of dissolved ions in post glacial oceans. This work thus addresses two main scientific questions about diagenetic spheroids via a literature review:
- a)
- Are there secular variations in the abundance of diagenetic spheroids during the Proterozoic?
- b)
- Could Proterozoic diagenetic spheroids represent sedimentological signatures of abiotic carbon cycling favoured by environmental oxidation?
2. Approach for New Compilation
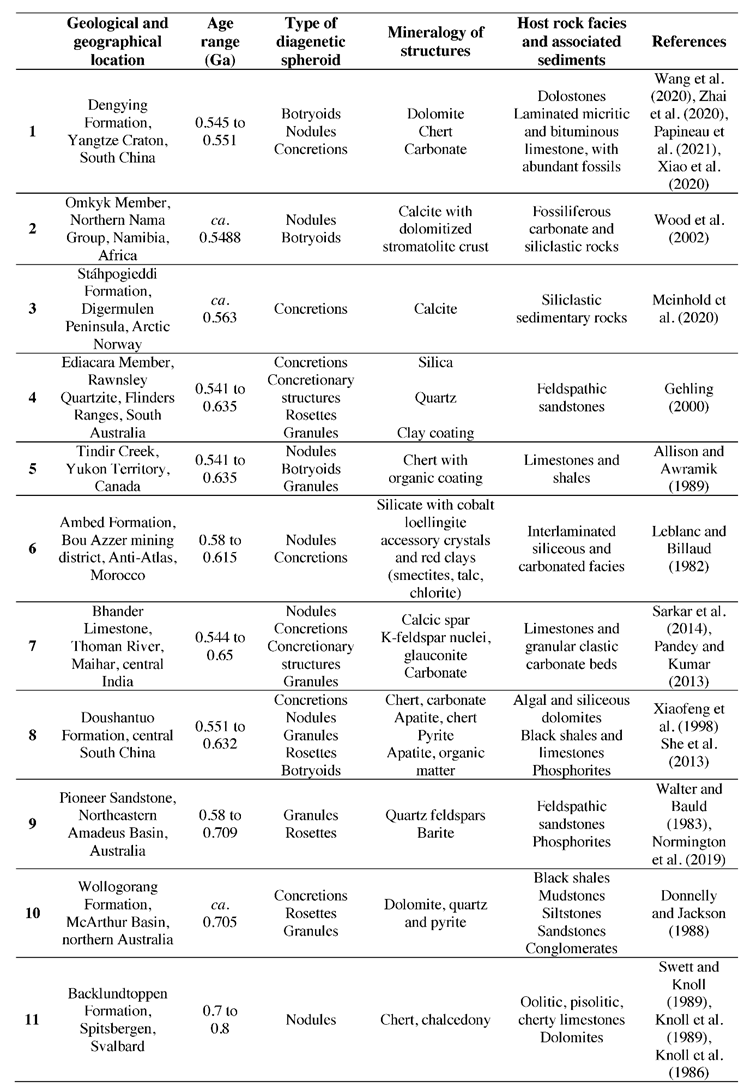
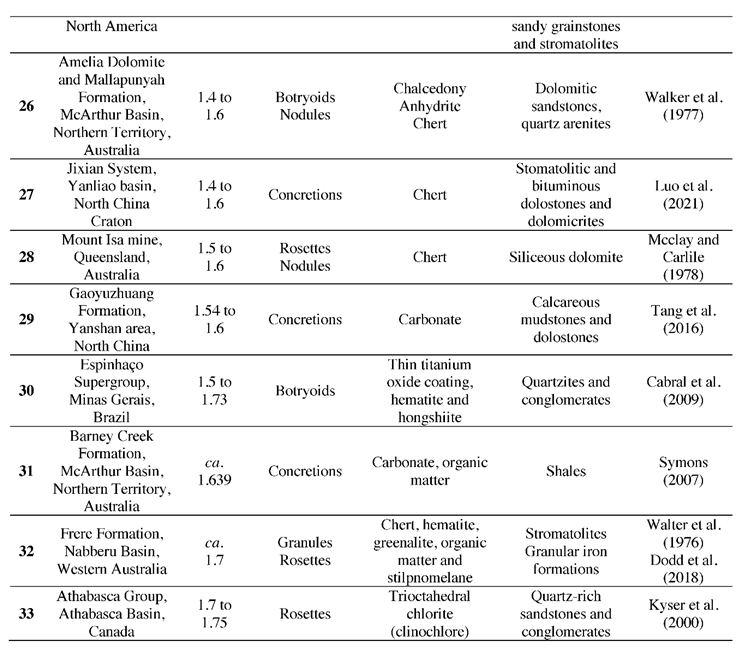
To determine the correlation between the occurrences of diagenetic spheroids during stable conditions and enhanced influxes of environmental oxygen, the abundance of concretions, nodules, botryoids, granules and rosettes in sedimentological beds between 0.541 to 1.8 Ga was collected (Table 1). Altogether, 72 occurrences of diagenetic spheroids from 33 formations were documented (Figure 1). Even though this is a comprehensive compilation of Proterozoic diagenetic spheroids, sample and formation bias is inherent within this collated data due to outcrop availability, the variable number of studies for each formation, the analytical approach used in publications, and whether authors reported occurrences of diagenetic spheroids of various sizes when they occurred. While these sampling biases are inherent to the geological and publication records, we argue that this compilation can still serve a useful purpose and show possible first-order trends. In the future, this compilation should be augmented, refined, and hopefully create a momentum that stimulates the report of occurrences of many more diagenetic spheroids, including at microscopic scales.
3. Observations from Compilation of Proterozoic Spheroids
3.1. Geometric Patterns of Diagenetic Spheroids
Concentric lamination patterns are displayed when multiple circles share the same centre. Within Proterozoic diagenetic spheroids, these are the most common type of geometric pattern, with 43% of the total collated diagenetic spheroids displaying circular laminations. However, this value is likely underestimated because the internal geometry of 36 compiled spheroids remains undetermined. Figure 2b shows an example of a carbonate concretion with circularly concentric layers and coloured mineral gradients. Botryoidal dolomites from the Dengying Formation (Figure 2c-e) demonstrate the formation of circular twins at the junctions where concentric laminations meet and destructively interfere. This arrangement can be considered as examples of crystal twinning within botryoidal minerals, which have several orders of magnitude sizes. Irregularly shaped cavity structures also form upon the destructive interference of circularly concentric laminations (Figure 2a). Variations in geometric patterns are dependent on the extent of the convergence of individual concentric circles. Therefore, fractal geometry occurs, since self-similar and circularly concentric patterns are observed at regular intervals over several orders of size magnitudes in Proterozoic diagenetic spheroids.
Centered-radial patterns are the next most common geometric pattern, with five recorded diagenetic spheroids during the Proterozoic (Figure 1). These are similar to circularly concentric patterns, but with the addition of radial fibrous crystals that have darker coloured banded zones emanating from a singular centre point. The laminations observed in Figure 2e have a rhythmically oscillating thin dark and thick brighter lamina with sharp boundaries. The dark and bright laminae are syntaxial, with the lighter laminae containing larger dolomite crystals (Zhai et al., 2020). Lenticular and triaxial ellipsoids are infrequent patterns and are typically demonstrated by nodules and concretions. Ellipsoidal shapes in diagenetic spheroids may be due to the original morphology of decomposing clumps of biomass.
3.2. Mineralogy and Palaeontology of Diagenetic Spheroids
Chert is the most common mineral, with 12 out of 72 occurrences of diagenetic spheroids exhibiting a chert mineralogy. Ten carbonate spheroids and one silicate spheroid are recorded, but both lack specificity since these terms represent mineral groups. For example, the typical anhydrous carbonate minerals are calcite, aragonite, and dolomite. Quartz and feldspars are common types of silicates, but further petrographic analyses of thin sections should be conducted more commonly on diagenetic spheroids to further determine their mineralogy. Quartz occurrences (8), dolomite occurrences (7) and calcite (6) are the next most common major minerals (see Table 1). Moreover, phosphorite nodules in siliceous dolomite and micritic limestone are abundant in the Doushantuo Formation in China (Xiaofeng et al., 1998). Botryoidal apatite occurs among granules in black organic-rich phosphorite of the Doushantuo Formation (She et al., 2013). Pyritic concretions also occur in the Mesoproterozoic Newland Formation in North America (Strauss and Schieber, 1990).
Shallow-marine depositional conditions nearby stromatolites or microbialites commonly apply to Proterozoic spheroids, with 7 diagenetic spheroids. In the Namibian Omkyk Member near the end of the Ediacaran, nodules are encrusted by a rim of dolomitised stromatolites and contain various microfossils (Figure 3a to 3c) (Wood et al., 2002). These purported metazoans are biomineralized and are oriented perpendicular to a fissure wall (Wood et al., 2002). Ediacaran microfossils can also be observed within diagenetic spheroids from China’s Shibantan Lagerstätte (Xiao et al., 2020). Chert nodules from Shibantan Member of the Dengying Formation also contain well preserved spheroidal microfossils such as Oscillatoriopsis obtusa, (Figure 3d) (Ding et al., 2019). Although the phylogenetic affinities of these microfossils are undetermined, due to their morphological similarity to photoautotrophs, they have been interpreted as benthic cyanobacteria (Xiao et al., 2020). These observations and inferences have also been made in the Doushantuo Formation where apatite granules commonly contain a range of microfossils (She et al., 2013). There are also well-preserved microfossils inside concretions of bitumen and chert from the Mesoproterozoic Xiamaling Formation in northern China (Wang et al., 2017). In chert granules from the Late Paleoproterozoic Frere Formation, subspherical and filamentous microfossils are clustered together and exhibit varying degrees of mineralisation (Figure 3e), whereas most are encrusted with a relatively thick layer of iron oxides, (Walter et al., 1976).
3.3. Distribution Trend of Diagenetic Spheroids
A greater number of occurrences of diagenetic spheroids during the Neoproterozoic, especially in the Ediacaran, can be observed compared to the Mesoproterozoic (Figure 1). The highest peak in the number of diagenetic spheroids thus coincides with positive excursions of δ13Ccarb values (Och and Shields, 2012; Shields et al., 2019), which is a major proxy for increased atmospheric oxygen. By contrast a limited number of spheroids occur in the 0.8-1.8 Ga interval, which coincides with the ‘Balanced Billion’. For instance, a significant decrease of diagenetic spheroids occurs during the early Tonian, because only two diagenetic spheroid occurrences from the Mackenzie Mountains in northwestern Canada have been recorded. There also appears to be no considerable changes in the number of diagenetic spheroids within the Mesoproterozoic, with a roughly similar amount across the Stenian, Ectasian and Calymmian (Figure 1).
4. Discussion
4.1. Geometric Patterns in Proterozoic Diagenetic Spheroids Compared to Those of COR
Under standard conditions, an unperturbed B-Z reaction produces random oxidation spots that propagate as a series of equidistant concentric rings (Zaikin and Zhabotinsky, 1970). Once a significant number of chemical waves have accumulated, destructive interference between waves occur (Figure 4b-c). At late reaction stages, linearly elongated cavities and twinned patterns occur at sites where chemical waves are mutually annihilated, causing the erasure of wave traces (Papineau et al., 2017, 2021).
The geometric patterns produced by COR are self-similar to the circularly concentric patterns found within diagenetic spheroids. Figure 4a shows an example of equidistant concentric laminations with no interference, perfectly recreating some of the COR patterns. Moreover, sub-parallel wavy layers caused by circular twins can be recreated by B-Z reactions due to destructive interference (Figure 4b). This effect appears related to surface tension in botryoids where the layers usually follow the cavity inside lining, whereas in a Petri dish the sub-parallel layers follow the glass wall. Circularly concentric waves can also have imperfect equidistant or non-equidistant laminations (Belmonte et al., 1997; Papineau, 2020), which is akin to the variable spacing distance displayed by laminations in some diagenetic spheroids. Mineral colour density gradients are also apparent in Figure 4b. Asymmetric irregular ellipses of concentric chemical waves are due to a higher concentration of nucleation spots and the resulting twins between chemical waves during radial diffusion (Figure 4c). Other diagenetic spheroids with these kinds of self-similar patterns that have previously been attributed an origin influenced by COR include rosettes of quartz ± apatite ± haematite ± carbonate ± organic matter (Papineau et al., 2016; 2022; Dodd et al., 2018), botryoids of malachite, quartz, and dolomite + organic matter (Papineau, 2020; Papineau et al., 2021; Gabriel and Papineau, 2021; Goodwin and Papineau, 2022; Varkouhi et al., 2022; Varkouhi and Papineau, 2023), granules of quartz ± apatite ± haematite ± magnetite ± organic matter ± dolomite ± pyrite (Papineau et al., 2017; Dodd et al., 2018), concretions or nodules of quartz ± haematite ± carbonate ± organic matter ± pyrite (Papineau et al., 2021; 2022; Papineau, 2024).
Oxidation spots in COR are chaotically distributed (Vidal and Pagola, 1989), which is akin to the spontaneous locations of diagenetic spheroids within sedimentary layers. Hastings et al. (2003) argued against random initiation by attributing microscopic fluctuations in the concentration of reactants to the initial nucleation of chemical activity. Despite the unpredictability of COR, these reactions can produce self-similar patterns such as circularly concentric equidistant laminations, twinned circles, cavity structures, radial diffusion, and colour gradients in laminations. All these self-similar patterns closely resemble those found in Proterozoic diagenetic spheroids. Oxidation spots with circularly concentric laminations in COR experiments have diameters that span sub-millimetre to decimetre sizes (Zaikin and Zhabotinsky, 1970; Chen et al., 2011). Therefore, COR display fractal patterns, akin to the fractal nature of patterns displayed by diagenetic spheroids.
Furthermore, strong colour gradients are produced during COR, which are analogous to alternating light and dark mineral laminations of organic matter within circularly concentric spheroids. The characteristic red and blue seen in COR experiments is produced via ferroin reduction and oxidation, respectively (Chen et al., 2011). The oscillatory patterns displayed in Figure 4 are initiated inside a ten centimetre diameter glass Petri dish and requires a film of COR solution (Papineau et al., 2021). Less distinguished chemical waves and cavity structures become apparent when some of the reactants are exhausted after around 30 minutes (Epstein et al., 1983). During diagenesis, COR must occur three-dimensionally and at much larger scales, which is a limitation not yet demonstrated by the COR model. Furthermore, COR experiments in Petri dishes can be restarted when the solution is shaken, which suggests that COR in nature would occur during the early stages of burial diagenesis and within relatively short time scales. Nevertheless, it likely took longer in environmental settings than in COR experiments due to the expected lower concentration of reactants. Despite these limitations, patterns of circularly concentric laminations, colour gradients, radial geometry, twins, and cavities are patterns found both in COR and in diagenetic spheroids (Figure 4), which constitutes a solid basis to argue for COR as a model for the formation of diagenetic spheroids.
4.2. Substances of COR Compared to Those of Proterozoic Diagenetic Spheroids
Experimental COR begin when a redox-sensitive metal-based catalyst is added to a solution with a strongly oxidising halogen with its complementary halide salt, malonic acid and sulphuric acid (Zaikin and Zhabotinsky, 1970, Belmonte et al., 1997). Variable concentrations can be utilised to initiate COR and a various reactants that can produce B-Z patterns (Agladze et al., 1984; Belmonte et al., 1997). Although many chemical reactions can occur during COR, two equations are particularly important:
In equation (1), iron in ferroin is oxidised from Fe2+ to Fe3+, which causes the observed colour change. In equation (2), the reaction involves decarboxylation of malonic acid (or other carboxylic acids), the production of carbon dioxide (seen as bubbles in COR experiments), and a change in oxidation state of bromine from +5 to -1. In COR, the chemical composition of products and intermediates remains undetermined, yet it can be predicted that the chemical waves are composed of variably oxidised ferroin and halogenated carboxylic acids (Papineau et al., 2021).
2Fe2+ + BrO3– + HBrO2 + 3H+ ↔ 2Fe3+ + 2HBrO2 + H2O + 2e-
3CH2 (COOH)2 + 4BrO3- → 4Br- + 9CO2 + 6H2 O
The concentration of bromide ions (Br−) has been argued to control the oscillating nature of the B-Z reaction (Orbán et al., 2001). When Br− exceeds the critical level, the oxidation of iron in ferroin is favoured, which generates a colour change in the diffusing circular waves from red (Fe2+) to blue (Fe3+). Other metals can also act as possible catalysts in COR (Belmonte et al., 1997). The reaction is thought to terminate when electron donor molecules like carboxylic acids or electron acceptors like bromate are exhausted or become too low in concentration (Papineau et al., 2020). Therefore, diagenetic environments that can concentrate reactants are ideal, such as evaporitic environments where halogens, sulphate, and biomass can accumulate.
Carboxylic acids and ketones are examples of other organic acids that can initiate COR (Belmonte et al., 1997; Orbán et at., 2001; Papineau et al., 2017). In nature, carboxylic acids are abundant because they are natural constituents of biomass, which decays in part through decarboxylation reactions (Bernard et al., 2007; Griffin et al., 2015; Sheik et al., 2020). Spheroidal microfossils such as Oscillatoriopsis obtusa and Namapoikia rietoogensis gen. et sp. nov. occur in Proterozoic diagenetic spheroids (Figure 3a-d) (Xiao et al., 2020; Wood et al., 2002), and other Proterozoic microfossils commonly occur in diagenetic spheroids (She et al., 2013; Wang et al., 2017). Subspherical and filamentous microfossils occur in granules from the late Palaeoproterozoic Frere Formation in the Nabberu Basin (Figure 3e) and they often exhibit iron oxide encrustation (Walter et al., 1976). The association of organic matter with iron oxides and pyrite could be because these minerals can be products of microbial sulphate reduction (Rueter et al., 1994; Muyzer and Stams, 2008). However, disseminations of iron oxides, pyrite, carbonate, and organic matter in diagenetic spheroids composed of quartz can also be interpreted to arise from abiotic COR because these can display many of the same self-similar patterns (Papineau et al., 2021; Varkouhi and Papineau, 2023; Papineau, 2024). Hence, other approaches are needed distinguish contributions from biological sulphate reduction and abiotic sulphate reduction in diagenetic spheroids, such as through large ranges of sulphur isotope ratios. However, COR have not yet been investigated for stable isotope fractionation on sulphur, carbon, iron, or halogens. Therefore, the abiotic putrefaction of microbial biomass in diagenetic environments is a likely process that uses organic acid reactants for COR (Papineau et al., 2021) and thus, the COR model can elegantly explain why Proterozoic diagenetic spheroids are often associated with fossils and organic matter.
For COR to occur naturally, productive, oxidising and evaporitic diagenetic environments are well-suited to decompose microbial biomass. Table 1 displays the variety of environments considered in this study and shows limestone, phosphorite, chert and quartzitic-sandstone most commonly contain diagenetic spheroids. The preservation of perfectly equidistant circularly concentric lamination patterns, radially aligned crystals and a limited range of chemical composition is not immediately compatible with the wave action model for the formation of diagenetic spheroids like botryoids and granules (Simonson, 2003; Pufahl and Fralick, 2004; Akin et al., 2013; Smith et al., 2017). Hence, while wave action cannot be excluded to play a role in rounding granules for instance (Dodd et al., 2018), it should be considered an insufficient model to explain the occurrence of specific geometric patterns within. The gas bubble model (Bosak et al., 2010) can also be ruled out based on its inability to explain why microfossils are commonly inside diagenetic spheroids. Considering their circular concentricity, mineralogy, and inclusions of organic matter and common microfossils, the COR model is the most convincing model for the formation of diagenetic spheroids (Papineau et al., 2016, 2017, 2021; Dodd et al., 2018; Gabriel et al., 2021; Varkouhi et al., 2022; Varkouhi and Papineau, 2023, Papineau, 2020; 2024).
During early diagenesis, the oxidation of organic acids through the decarboxylation of organic matter, was likely needed to initiate COR and produce spheroidal objects (Papineau et al., 2017, 2021). Originally, colloidal silica is present under alkaline conditions, but when carboxylic acids undergo decarboxylation, acid is produced and the pH is lowered (Papineau et al., 2021). If this occurs under alkaline conditions, it results in the precipitation of minerals such as quartz, apatite, carbonate and pyrite (Papineau et al., 2024). Microcrystalline quartz, cryptocrystalline apatite, micritic carbonate, nanoscopic pyrite and hematite, and finely disseminated organic matter are all commonly found within diagenetic spheroids (Table 1). Hence, these minerals are consistent with the COR model of diffusing reaction products and intermediates in colloidal gels with its consequent permineralisation under pH gradients. Therefore, in addition to the variety of geometric patterns, the mineralogy and chemical substances of diagenetic spheroids can also be explained by the COR model.
4.3. Influences of Atmospheric Oxygenation on Occurrences of Diagenetic Spheroids
A majority of diagenetic spheroids occur during the Neoproterozoic era (Figure 1). The increase in occurrences coincides with intense variations in δ13Ccarb, which is a well-known proxy for atmospheric oxygenation. Therefore, this is an indication that major perturbations in the carbon cycle are linked to increased occurrences of diagenetic spheroids and thus of COR in the environment. This observation also supports the COR model for the formation of circularly concentric laminations in diagenetic spheroids and for the preservation of microfossils in diagenetic spheroids. This statement is further supported by the smaller number of diagenetic spheroid occurrences during the ‘Balanced Billion’, when variations in δ13Ccarb were significantly smaller.
Under increased environmental oxygen, higher chemical weathering rates, and intense greenhouse conditions, more cations and anions are delivered into sea water (Hardisty et al. 2017). Hence during the Ediacaran, iodate, bromate and sulphate levels in seawater increased (Hardisty et al. 2017), thereby providing an increased availability of COR reactants. Post-glacial Neoproterozoic rocks of marine sedimentary origin also show increased variations in δ34Ssulphate and δ34Ssulphide, which imply increased availability of seawater sulphate (Ross et al., 1995; Hurtgen et al., 2005). Oxidised sulphur species can act as electron acceptors in the remineralization of organic carbon by sulphate-reducing bacteria, or abiotically as in the case of COR in the environment. Primary productivity of organic matter also increased during the Neoproterozoic and promoted a profound expansion of ecosystems, because sulphate and phosphate play central roles in biochemistry (Canfield and Raiswell, 1999). This biosphere expansion would enable an increased availability of biomass and carboxylic acids, which again are key reactants in COR. Lastly, environmental oxygenation during the Neoproterozoic would also have contributed to create redox gradients and thermodynamic disequilibria in sedimentary marine environments, thereby further stimulating COR.
A natural progression of this study includes the expansion of the considered age range to encompass the Phanerozoic eon (0.541 Ga until modern times) as well as into the Paleoproterozoic era and Archean eon. This study provides valuable new insights on the influence of intense variations in δ13Ccarb on the prevalence of diagenetic spheroids in sedimentary rocks and it is likely applicable to the Paleoproterozoic. An almost symmetrical distribution of the occurrences of diagenetic spheroids, with greater amounts at both ends of the Proterozoic, would therefore further support the newly proposed COR model. Since the connection between COR and diagenetic spheroids is still in its infancy, broader documentation of these spheroidal structures is needed to search for additional occurrences. This may eventually confirm that the COR model applies to all types of diagenetic spheroids, throughout Earth history. Diagenetic spheroids should then be regarded as sedimentological patterns produced abiotically from carbon cycling with carboxylic acids and CO2. While carboxylic acids are most likely of biological origin in most environments, but they may also be abiotic in origin, for instance in hydrothermal vent systems where Fischer-Tropsch Type synthesis takes place (McCollom et al., 1999). Hence, this study may even apply more broadly to oceanography and planetary sciences.
5. Conclusion
The classical COR is the B-Z reaction, which produces characteristic fractal patterns and CO2 bubbles from reactants such as malonic acid, bromate-bromide, sulphuric acid and ferroin. Self-similar fractal patterns, such as circularly concentric laminations, radial geometries, twins, and linearly elongated cavities occur in diagenetic spheroids and are identical to those produced by COR. These diagenetic spheroids include concretions, nodules, botryoids, granules and rosettes. Similarities between chemical reactants and products of COR and diagenetic spheroids have also been elucidated based on chemical substances. Hence diagenetic spheroids are interpreted to be formed by COR, based on their patterns and substances, and they can thus be considered as evidence of abiotic carbon cycling.
Because biomass is enriched in carboxylic acids, organic matter in sediments is affected by decarboxylation during the formation of diagenetic spheroids, hence the COR model explains why fossils and microfossils are common in these mineralised objects. In this first compilation of occurrences of diagenetic spheroids over geological history between 0.541 and 1.8 Ga, a secular peak in the number of their occurrences is documented during the late Neoproterozoic. This observation is consistent with the COR model because the Neoproterozoic oxygenation event resulted in high levels of biomass, increased oxidative weathering and greater amounts of oxidants, such as iodate and sulphate, all of which are linked to reactants in COR. These conditions during the Neoproterozoic were favourable to COR and resulted in increased occurrences of diagenetic spheroids in sedimentary rocks from this era. This study is a prelude to others which will cover the rest of Earth’s history.
References
- Ader, M., Macouin, M., Trindade, R.I.F., Hadrien, M., Yang, Z.Y., Sun, Z.M. and Besse, J. (2009). A multilayered water column in the Ediacaran Yangtze platform? Insights from carbonate and organic matter paired δ13C. Earth and Planetary Science Letters, 288(1-2), 213-227. DOI: 10.1016/j.epsl.2009.09.024.
- Agladze, K.I., Krinsky, V.I. and Pertsov, A.M. (1984). Chaos in the non-stirred Belousov–Zhabotinsky reaction is induced by interaction of waves and stationary dissipative structures. Nature, 308, 834-835. DOI: 10.1038/308834a0.
- Akin, S.J., Pufahl, P.K., Hiatt, E.E. and Pirajno, F. (2013). Oxygenation of shallow marine environments and chemical sedimentation in Palaeoproterozoic peritidal settings: Frere Formation, Western Australia. Sedimentology, 60(7), 1559-1582. DOI: 10.1111/sed.12038.
- Allison, C.W. and Awramik, S.M. (1989). Organic-walled microfossils from earliest Cambrian or latest Proterozoic Tindir Group rocks, northwest Canada. Precambrian Research, 43(4), 253-294. DOI: 10.1016/0301-9268(89)90060-0.
- Belmonte, A.L., Ouyang, Q. and Flesselles, J.M. (1997). Experimental survey of spiral dynamics in the Belousov-Zhabotinsky reaction. Journal de Physique II, 7(10), 1425-1468. DOI: 10.1051/jp2:1997195.
- Bernard, S., Benzerara, K., Beyssac, O., Menguy, N., Guyot, F., Brown Jr, G.E. and Goffé, B. (2007). Exceptional preservation of fossil plant spores in high-pressure metamorphic rocks. Earth and Planetary Science Letters, 262(1-2), 257-272. DOI: 10.1016/j.epsl.2007.07.041.
- Blake, T.S., Rothery, E., Muhling, J.R., Drake-Brockman, J.A.P., Sprigg, L.C., Ho, S.E., Rasmussen, B. and Fletcher, I.R. (2011). Two episodes of regional-scale Precambrian hydrothermal alteration in the eastern Pilbara, Western Australia. Precambrian Research, 188(1-4), 73-103. DOI: 10.1016/j.precamres.2011.03.010.
- Bosak, T., Bush, J.W.M., Flynn, M.R., Liang, B., Ono, S., Petroff, A.P. and Sim, M.S. (2010). Formation and stability of oxygen-rich bubbles that shape photosynthetic mats. Geobiology, 8(1), 45-55. DOI: 10.1111/j.1472-4669.2009.00227.x.
- Briggs, T.S. and Rauscher, W.C. (1973). An oscillating iodine clock. Journal of Chemical Education, 50(7), 496. DOI: 10.1021/ed050p496.
- Bristow, T.F., Bonifacie, M., Derkowski, A., Eiler, J.M. and Grotzinger, J.P. (2011). A hydrothermal origin for isotopically anomalous cap dolostone cements from south China. Nature, 474(7349), 68-71. DOI: 10.1038/nature10096.
- Bull, S., Selley, D., Broughton, D., Hitzman, M., Cailteux, J., Large, R. and McGoldrick, P. (2011). Sequence and carbon isotopic stratigraphy of the Neoproterozoic Roan Group strata of the Zambian copperbelt. Precambrian Research, 190(1-4), 70-89. DOI: 10.1016/j.precamres.2011.07.021.
- Cabral, A.R., Lehmann, B., Tupinambá, M., Schlosser, S., Kwitko-Ribeiro, R. and de Abreu, F.R. (2009). The platiniferous Au-Pd belt of Minas Gerais, Brazil, and genesis of its botryoidal Pt-Pd aggregates. Economic Geology, 104(8), 1265-1276. DOI: 10.2113/gsecongeo.104.8.1265.
- Burley, S.D., Kantorowicz, J.D. and Waugh, B. (1985). Clastic diagenesis. Geological Society, London, Special Publications, 18(1), 189-226. DOI: 10.1144/GSL.SP.1985.018.01.10.
- Cañadas, F., Papineau, D., Leng, M.J. and Li, C. (2022). Extensive primary production promoted the recovery of the Ediacaran Shuram excursion. Nature communications, 13, 148. DOI: 10.1038/s41467-021-27812-5.
- Canfield, D.E. and Raiswell, R. (1999). The evolution of the sulfur cycle. American Journal of Science, 299(7-9), 697-723. DOI: 10.2475/ajs.299.7-9.697.
- Chen, I.C., Kuksenok, O., Yashin, V.V., Moslin, R.M., Balazs, A.C. and Van Vliet, K.J. (2011). Shape-and size-dependent patterns in self-oscillating polymer gels. Soft Matter, 7(7), 3141-3146. DOI: 10.1039/C0SM01007C.
- Cohen, K.M., Finney, S.C., Gibbard, P.L. and Fan, J.X. (2013). The ICS International Chronostratigraphic Chart. Episodes, 36(3), 199-204.
- Collinson, J.D. (1983). Sedimentology of unconformities within a fluvio-lacustrine sequence; Middle Proterozoic of Eastern North Greenland. Sedimentary Geology, 34(2-3), 145-166. DOI: 10.1016/0037-0738(83)90084-2.
- Curtis, C.D. (1977). Geochemistry: sedimentary geochemistry: environments and processes dominated by involvement of an aqueous phase. Philosophical Transactions of the Royal Society of London. Series A, Mathematical and Physical Sciences, 286(1336), 353-372. DOI: 10.1098/rsta.1977.0123.
- Dahanayake, K., Gerdes, G. and Krumbein, W.E. (1985). Stromatolites, oncolites and oolites biogenically formed in situ. Naturwissenschaften, 72(10), 513-518. DOI: 10.1007/BF00367596.
- Dahanayake, K. and Krumbein, W.E. (1986). Microbial structures in oolitic iron formations. Mineralium Deposita, 21(2), 85-94. DOI: 10.1007/BF00204266.
- Davies, P.J., Bubela, B. and Ferguson, J. (1978). The formation of ooids. Sedimentology, 25(5), 703-730. DOI: 10.1111/j.1365-3091.1978.tb00326.x.
- Diaz, M.R. and Eberli, G.P. (2019). Decoding the mechanism of formation in marine ooids: A review. Earth-Science Reviews, 190, 536-556. DOI: 10.1016/j.earscirev.2018.12.016.
- Ding, W., Dong, L., Sun, Y., Ma, H., Xu, Y., Yang, R., Peng, Y., Zhou, C. and Shen, B. (2019). Early animal evolution and highly oxygenated seafloor niches hosted by microbial mats. Scientific Reports, 9(1), 13628. DOI: 10.1038/s41598-019-49993-2.
- Dodd, M.S., Papineau, D., She, Z., Fogel, M.L., Nederbragt, S. and Pirajno, F. (2018). Organic remains in late Palaeoproterozoic granular iron formations and implications for the origin of granules. Precambrian Research, 310, 133-152. DOI: 10.1016/j.precamres.2018.02.016.
- Donnelly, T.H. and Jackson, M.J. (1988). Sedimentology and geochemistry of a mid-Proterozoic lacustrine unit from northern Australia. Sedimentary geology, 58(2-4), 145-169. DOI: 10.1016/0037-0738(88)90067-X.
- Epstein, I.R., Kustin, K., De Kepper, P. and Orbán, M. (1983). Oscillating chemical reactions. Scientific American, 248(3), 112-123.
- Field, R.J. and Schneider, F.W. (1989). Oscillating chemical reactions and nonlinear dynamics. Journal of Chemical Education 66(3), 195. DOI: 10.1021/ed066p195.
- Fike, D.A., Grotzinger, J.P., Pratt, L.M. and Summons, R.E. (2006). Oxidation of the Ediacaran Ocean. Nature, 444, 744-747. DOI: 10.1038/nature05345.
- Flannery, D.T., Allwood, A.C., Hodyss, R., Summons, R.E., Tuite, M., Walter, M.R. and Williford, K.H. (2019). Microbially influenced formation of Neoarchean ooids. Geobiology, 17(2), 151-160. DOI: 10.1111/gbi.12321.
- Fouke, B.W. (2011). Hot-spring Systems Geobiology: abiotic and biotic influences on travertine formation at Mammoth Hot Springs, Yellowstone National Park, USA. Sedimentology, 58, 170-219. DOI: 10.1111/j.1365-3091.2010.01209.x.
- Gabriel, N.W., Papineau, D., She, Z., Leider, A. and Fogel, M.L. (2021). Organic diagenesis in stromatolitic dolomite and chert from the late Palaeoproterozoic McLeary Formation. Precambrian Research, 354, 106052. DOI: 10.1016/j.precamres.2020.106052.
- Gehling, J.G. (2000). Environmental interpretation and a sequence stratigraphic framework for the terminal Proterozoic Ediacara Member within the Rawnsley Quartzite, South Australia. Precambrian Research, 100(1-3), 65-95. DOI: 10.1016/S0301-9268(99)00069-8.
- Glasauer, S., Mattes, A. and Gehring, A. (2013). Constraints on the Preservation of Ferriferous Microfossils. Geomicrobiology Journal, 30(6), 479-489. DOI: 10.1080/01490451.2012.718408.
- Goddéris, Y., Donnadieu, Y., Dessert, C., Dupré, B., Fluteau, F., François, L.M., Meert, J., Nédélec, A. and Ramstein, G. (2007). Coupled modeling of global carbon cycle and climate in the Neoproterozoic: links between Rodinia breakup and major glaciations. Comptes Rendus Geoscience, 339(3-4), 212-222. DOI: 10.1016/j.crte.2005.12.002.
- Goode, A.D.T. and Hall, W.D.M. (1981). The middle Proterozoic Eastern Bangemall Basin, Western Australia. Precambrian Research, 16(1-2), 11–29. DOI: 10.1016/0301-9268(81)90003-6.
- Gray, C. (2002). An analysis of the Belousov-Zhabotinskii reaction. Rose-Hulman Undergraduate Mathematics Journal, 3(1), 1.
- Griffin, J.D., Zeller, M.A. and Nicewicz, D.A. (2015). Hydrodecarboxylation of carboxylic and malonic acid derivatives via organic photoredox catalysis: substrate scope and mechanistic insight. Journal of the American Chemical Society, 137(35), 11340-11348. DOI: 10.1021/jacs.5b07770.
- Hardisty, D.S., Lu, Z., Bekker, A., Diamond, C.W., Gill, B.C., Jiang, G., Kah, L.C., Knoll, A.H., Loyd, S.J., Osburn, M.R. and Planavsky, N.J. (2017). Perspectives on Proterozoic surface ocean redox from iodine contents in ancient and recent carbonate. Earth and Planetary Science Letters, 463, pp.159-170. DOI: 10.1016/j.epsl.2017.01.032.
- Hastings, H.M., Field, R.J. and Sobel, S.G. (2003). Microscopic fluctuations and pattern formation in a supercritical oscillatory chemical system. The Journal of Chemical Physics, 119(6), 3291-3296. DOI: 10.1063/1.1587700.
- Hofmann, H.J. and Altken, J.D. (1979). Precambrian biota from the Little Dal Group, Mackenzie Mountains, northwestern Canada. Canadian Journal of Earth Sciences, 16(1), 150-166. DOI: 10.1139/e79-014.
- Hurtgen, M.T., Arthur, M.A. and Halverson, G.P. (2005) Neoproterozoic sulfur isotopes, the evolution of microbial sulfur species, and the burial efficiency of sulfide as sedimentary pyrite. Geology, 33, 41-44; DOI: 10.1130/G20923.1.
- Jorgensen, B.B. (1979). A theoretical model of the stable sulfur isotope distribution in marine sediments. Geochimica et Cosmochimica Acta, 43(3), 363-374. DOI: 10.1016/0016-7037(79)90201-1.
- Karhu, J.A. and Holland, H.D. (1996). Carbon isotopes and the rise of atmospheric oxygen. Geology, 24(10), 867. DOI:10.1130/0091-7613(1996)024<0867:ciatro>2.3.co;2.
- Kaufman, A.J., Hayes, J.M., Knoll, A.H. and Germs, G.J. (1991). Isotopic compositions of carbonates and organic carbon from upper Proterozoic successions in Namibia: stratigraphic variation and the effects of diagenesis and metamorphism. Precambrian Research, 49(3-4), 301-327. DOI: 10.1016/0301-9268(91)90039-D.
- Keller, C.B., Husson, J.M., Mitchell, R.N., Bottke, W.F., Gernon, T.M., Boehnke, P., Bell, E.A., Swanson-Hysell, N.L. and Peters, S.E. (2018). Neoproterozoic glacial origin of the Great Unconformity. Proceedings of the National Academy of Sciences, 116(4), 1136-1145. DOI: 10.1073/pnas.1804350116.
- Keller, J.B. and Rubinow, S.I. (1981). Recurrent precipitation and Liesegang rings. The Journal of Chemical Physics, 74(9), 5000-5007. DOI: 10.1063/1.441752.
- Kinnaird, T.C., Prave, A.R., Kirkland, C., Horstwood, M., Parrish, R. and Batchelor, R.A. (2007). The late Mesoproterozoic–early Neoproterozoic tectonostratigraphic evolution of NW Scotland: the Torridonian revisited. Journal of the Geological Society, 164(3), 541-551. DOI: 10.1144/0016-76492005-096.
- Kirschvink, J.L. (1992). Late Proterozoic low-latitude global glaciation: the Snowball Earth. Cambridge University Press, Cambridge, 51-52.
- Knoll, A.H., Hayes, J.M., Kaufman, A.J., Swett, K. and Lambert, I.B. (1986). Secular variation in carbon isotope ratios from Upper Proterozoic successions of Svalbard and East Greenland. Nature, 321(6073), 832-838. DOI: 10.1038/321832a0.
- Knoll, A.H., Swett, K. and Burkhardt, E. (1989). Paleoenvironmental distribution of microfossils and stromatolites in the Upper Proterozoic Backlundtoppen Formation, Spitsbergen. Journal of Paleontology, 63(2), 129-145. DOI: 10.1017/S002233600001917X.
- Körös, E. and Orbán, M. (1978). Uncatalysed oscillatory chemical reactions. Nature, 273(5661), 371-372. DOI: 10.1038/273371b0.
- Kyser, T.K., Hiatt, E.E., Renac, C., Durocher, K., Holk, G.J. and Deckart, K. (2000). Diagenetic fluids in Paleo-and Meso-Proterozoic sedimentary basins and their implications for long protracted fluid histories. Mineralogical Association of Canada Short Course, 28, 225-262. DOI: 10.13140/2.1.1033.1847.
- Lascelles, D.F. (2007). Black smokers and density currents: A uniformitarian model for the genesis of banded iron-formations. Ore Geology Reviews, 32(1-2), 381-411. DOI: 10.1016/j.oregeorev.2006.11.005.
- Leblanc, M. and Billaud, P. (1982). Cobalt arsenide orebodies related to an upper Proterozoic ophiolite; Bou Azzer (Morocco). Economic Geology, 77(1), 162-175. DOI: 10.2113/gsecongeo.77.1.162.
- Lebron, I. and Suarez, D.L (1996). Calcite nucleation and precipitation kinetics as affected by dissolved organic matter at 25°C and pH > 7.5. Geochimica et Cosmochimica Acta, 60(15), 2765-2776. DOI: 10.1016/0016-7037(96)00137-8.
- Li, D., Luo, G., Yang, H., She, Z., Papineau, D. and Li, C. (2022). Characteristics of the carbon cycle in late Mesoproterozoic: Evidence from carbon isotope composition of paired carbonate and organic matter of the Shennongjia Group in South China. Precambrian Research, 377, 106726. DOI: 10.1016/j.precamres.2022.106726.
- Liesegang, R.E. (1910). Die Entstehung der Achate. Zentralblatt für Mineralogie, 11, 593-597.
- Liu, A.Q., Tang, D.J., Shi, X.Y., Zhou, L.M., Zhou, X.Q., Shang, M.H., Li, Y. and Song, H.Y. (2019). Growth mechanisms and environmental implications of carbonate concretions from the~ 1.4 Ga Xiamaling Formation, North China. Journal of Palaeogeography, 8(1), 20. DOI: 10.1186/s42501-019-0036-4.
- Liu, A., Tang, D., Shi, X., Zhou, X., Zhou, L., Shang, M., Li, Y. and Fang, H. (2020). Mesoproterozoic oxygenated deep seawater recorded by early diagenetic carbonate concretions from the Member IV of the Xiamaling Formation, North China. Precambrian Research, 341, 105667. DOI: 10.1016/j.precamres.2020.105667.
- Luo, J., Long, X., Bowyer, F.T., Mills, B.J.W., Li, J., Xiong, Y., Zhu, X., Zhang, K. and Poulton, S.W. (2021). Pulsed oxygenation events drove progressive oxygenation of the early Mesoproterozoic ocean. Earth and Planetary Science Letters, 559, 116754. DOI: 10.1016/j.epsl.2021.116754.
- Mason, R., Li, Y., Cao, K., Long, Y. and She, Z.B. (2017). Ediacaran macrofossils in Shunyang Valley, Sixi, Three Gorges district, Hubei Province, China. Journal of Earth Science, 28(4), 614-621. DOI: 10.1007/s12583-017-0773-1.
- McClay, K.R. and Carlile, D.G. (1978). Mid-Proterozoic sulphate evaporites at Mount Isa mine, Queensland, Australia. Nature, 274(5668), 240-241. DOI: 10.1038/274240a0.
- McCollom, T.M., Ritter, G. and Simoneit, B.R. (1999). Lipid synthesis under hydrothermal conditions by Fischer-Tropsch-type reactions. Origins of Life and Evolution of the Biosphere, 29(2), pp.153-166. DOI: 10.1023/A:1006592502746.
- Meinhold, G., Roberts, N.M.W., Arslan, A., Jensen, S., Ebbestad, J.O.R., Högström, A.E.S., Høyberget, M., Agić, H., Palacios, T. and Taylor, W.L. (2020). U–Pb dating of calcite in ancient carbonates for age estimates of syn- to post-depositional processes: a case study from the upper Ediacaran strata of Finnmark, Arctic Norway. Geological Magazine, 157(8), 1367-1372. DOI: 10.1017/S0016756820000564.
- Merdith, A.S., Collins, A.S., Williams, S.E., Pisarevsky, S., Foden, J.D., Archibald, D.B., Blades, M.L., Alessio, B.L., Armistead, S., Plavsa, D., Clark, C., and Muller, D. (2017) A full-plate global reconstruction of the Neoproterozoic. Gondwana Research 50, 84–134.
- Mitchell, R.N. and Evans, D.A.D. (2024) The balanced billion. GSA Today, February issue, 10-11.
- Mitsuchi, M. (1976). Characteristics and genesis of nodules and concretions occurring in soils of the R. Chinit area, Kompong Thom Province, Cambodia. Soil Science and Plant Nutrition, 22(4), 409-421. DOI: 10.1080/00380768.1976.10433003.
- Muyzer, G. and Stams, A.J. (2008). The ecology and biotechnology of sulphate-reducing bacteria. Nature Reviews Microbiology, 6(6), 441-454. DOI: 10.1038/nrmicro1892.
- Nabika, H., Itatani, M. and Lagzi, I. (2019). Pattern Formation in Precipitation Reactions: The Liesegang Phenomenon. Langmuir, 36(2), 481-497. DOI: 10.1021/acs.langmuir.9b03018.
- Nascimento, D.B., Schmitt, R.S., Ribeiro, A., Trouw, R.A., Passchier, C.W. and Basei, M.A. (2017). Depositional ages and provenance of the Neoproterozoic Damara Supergroup (northwest Namibia): Implications for the Angola-Congo and Kalahari cratons connection. Gondwana Research, 52, 153-171. DOI: 10.1016/j.gr.2017.09.006.
- Normington, V.J., Beyer, E.E., Whelan, J.A., Edgoose, C.J. and Woodhead, J.D. (2019). Summary of results—NTGS LA-ICP-MS Hf program: Amadeus Basin, July 2013–June 2015. Northern Territory Geological Survey Record, 5, 34.
- Och, L.M. and Shields-Zhou, G.A. (2012). The Neoproterozoic oxygenation event: Environmental perturbations and biogeochemical cycling. Earth-Science Reviews, 110(1-4), 26-57. DOI: 10.1016/j.earscirev.2011.09.004.
- Orbán, M., Kurin-Csörgei, K., Zhabotinsky, A.M. and Epstein, I.R. (2001). A new chemical system for studying pattern formation: Bromate–hypophosphite–acetone–dual catalyst. Faraday Discussions, 120, 11-19. DOI: 10.1039/B102885P.
- Ostwald, J. and Bolton, B.R. (1990). Diagenetic braunite in sedimentary rocks of the Proterozoic Manganese Group, Western Australia. Ore Geology Reviews, 5(4), 315-323. DOI: 10.1016/0169-1368(90)90036-M.
- Pandey, S.K. and Kumar, S. (2013). Organic walled microbiota from the silicified algal clasts, Bhander limestone, Satna area, Madhya Pradesh. Journal of the Geological Society of India, 82(5), 499-508. DOI: 10.1007/s12594-013-0181-9.
- Papineau, D. (2010). Global Biogeochemical Changes at Both Ends of the Proterozoic: Insights from Phosphorites. Astrobiology, 10(2), 165-181. DOI: 10.1089/ast.2009.0360.
- Papineau, D. (2020). Chemically oscillating reactions in the formation of botryoidal malachite. American Mineralogist: Journal of Earth and Planetary Materials, 105(4), 447-454. DOI: 10.2138/am-2020-7029.
- Papineau, D. (2024) Chemically oscillating reactions as a new model for the formation of mineral patterns in Agate geodes and concretions. Minerals, 14(2), 203. DOI:10.3390/min14020203.
- Papineau, D., Devine, K. and Nogueira, B.A. (2023). Self-similar patterns from abiotic decarboxylation metabolism through chemically oscillating reactions: A prebiotic model for the origin of life. Life, 13(2), 551. DOI: 10.3390/life13020551.
- Papineau, D., De Gregorio, B., Fearn, S., Kilcoyne, D., McMahon, G., Purohit, R. and Fogel, M. (2016). Nanoscale petrographic and geochemical insights on the origin of the Palaeoproterozoic stromatolitic phosphorites from Aravalli Supergroup, India. Geobiology, 14(1) 3-32. DOI: 10.1111/gbi.12164.
- Papineau, D., She, Z. and Dodd, M.S. (2017). Chemically-oscillating reactions during the diagenetic oxidation of organic matter and in the formation of granules in late Palaeoproterozoic chert from Lake Superior. Chemical Geology, 470, 33-54. DOI: 10.1016/j.chemgeo.2017.08.021.
- Papineau, D., Yin, J., Devine, K.G., Liu, D. and She, Z. (2021). Chemically Oscillating Reactions during the Diagenetic Formation of Ediacaran Siliceous and Carbonate Botryoids. Minerals, 11(10), 1060. DOI: 10.3390/min11101060.
- Postgate, J. (1959). Sulphate reduction by bacteria. Annual Reviews in Microbiology, 13, 505-520. DOI: 10.1146/annurev.mi.13.100159.002445.
- Pratt, B.R. (1998). Molar-tooth structure in Proterozoic carbonate rocks: Origin from synsedimentary earthquakes, and implications for the nature and evolution of basins and marine sediment. Geological Society of America Bulletin, 110(8), 1028-1045. DOI: 10.1130/0016-7606(1998)110<1028:MTSIPC>2.3.CO;2.
- Pufahl, P.K. and Fralick, P.W. (2004). Depositional controls on Palaeoproterozoic iron formation accumulation, Gogebic Range, Lake Superior region, USA. Sedimentology, 51(4), 791-808. DOI: 10.1111/j.1365-3091.2004.00651.x.
- Ross, G.M., Bloch, J.D. and Krouse, H.R. (1995). Neoproterozoic strata of the southern Canadian Cordillera and the isotopic evolution of seawater sulfate. Precambrian Research, 73(1-4), 71-99. DOI: 10.1016/0301-9268(94)00072-Y.
- Rowan, M.G., Hearon IV, T.E., Kernen, R.A., Giles, K.A., Gannaway-Dalton, C.E., Williams, N.J., Fiduk, J.C., Lawton, T.F., Hannah, P.T. and Fischer, M.P. (2019). A review of allochthonous salt tectonics in the Flinders and Willouran ranges, South Australia. Australian Journal of Earth Sciences, 67(6), 787-813. DOI: 10.1080/08120099.2018.1553063.
- Rowlands, N.J., Blight, P.G., Jarvis, D.M. and Von Der Borch, C.C. (1980). Sabkha and playa environments in late Proterozoic grabens, Willouran Ranges, South Australia. Journal of the Geological Society of Australia, 27(1-2), 55-68. DOI: 10.1080/00167618008729118.
- Rueter, P., Rabus, R., Wilkest, H., Aeckersberg, F., Rainey, F.A., Jannasch, H.W. and Widdel, F. (1994). Anaerobic oxidation of hydrocarbons in crude oil by new types of sulphate-reducing bacteria. Nature, 372, 455-458. DOI: 10.1038/372455a0.
- Salama, W., El Aref, M.M. and Gaupp, R. (2012). Mineral evolution and processes of ferruginous microbialite accretion - an example from the Middle Eocene stromatolitic and ooidal ironstones of the Bahariya Depression, Western Desert, Egypt. Geobiology, 11(1), 15-28. DOI: 10.1111/gbi.12011.
- Sarkar, S., Choudhuri, A., Banerjee, S., Van Loon, A.J. and Bose, P.K. (2014) Seismic and non-seismic soft-sediment deformation structures in the Proterozoic Bhander Limestone, central India. Geologos, 20(2), 89-103. DOI: 10.2478/logos-2014-0008.
- Schidlowski, M. (1989). Evolution of the sulphur cycle in the Precambrian. Evolution of the global biogeochemical sulphur cycle, 3-19.
- Schopf, J.W., Kudryavtsev, A.B., Sugitani, K. and Walter, M.R. (2010). Precambrian microbe-like pseudofossils: A promising solution to the problem. Precambrian Research, 179(1-4), 191-205. DOI: 10.1016/j.precamres.2010.03.003.
- She, Z., Strother, P., McMahon, G., Nittler, L.R., Wang, J., Zhang, J., Sang, L., Ma, C. and Papineau, D. (2013). Terminal Proterozoic cyanobacterial blooms and phosphogenesis documented by the Doushantuo granular phosphorites I: In situ micro-analysis of textures and composition. Precambrian Research, 235, 20-35. DOI: 10.1016/j.precamres.2013.05.011.
- Sheik, C.S., Cleaves, H.J., Johnson-Finn, K., Giovannelli, D., Kieft, T.L., Papineau, D., Schrenk, M.O. and Tumiati, S. (2020). Abiotic and biotic processes that drive carboxylation and decarboxylation reactions. American Mineralogist: Journal of Earth and Planetary Materials, 105(5), 609-615. DOI: 10.2138/am-2020-7166CCBYNCND.
- Shields, G.A., Mills, B.J.W., Zhu, M., Raub, T.D., Daines, S.J. and Lenton, T.M. (2019) Unique Neoproterozoic carbon isotope excursions sustained by coupled evaporite dissolution and pyrite burial. Nature Geoscience. 12, 823–827. DOI: 10.1038/s41561-019-0434-3.
- Simonson, B.M. (2003). Origin and evolution of large Precambrian iron formations. In: Chan, M.A. and Archer, A.W. (Eds.), Extreme depositional environments: Mega end members in geologic time. Geological Society of America, 231–244.
- Skotnicki, S.J. and Knauth, L.P. (2007). The Middle Proterozoic Mescal Paleokarst, Central Arizona, U.S.A.: Karst Development, Silicification, and Cave Deposits. Journal of Sedimentary Research, 77(12), 1046-1062. DOI: 10.2110/jsr.2007.094.
- Smith, A.J.B., Beukes, N.J., Gutzmer, J., Czaja, A.D., Johnson, C.M. and Nhleko, N. (2017). Oncoidal granular iron formation in the Mesoarchaean Pongola Supergroup, southern Africa: Textural and geochemical evidence for biological activity during iron deposition. Geobiology, 15(6), 731-749. DOI: 10.1111/gbi.12248.
- Smith, E.F., MacDonald, F.A., Crowley, J.L., Hodgin, E.B. and Schrag, D.P. (2016). Tectonostratigraphic evolution of the c. 780–730 Ma Beck Spring Dolomite: Basin Formation in the core of Rodinia. Geological Society, London, Special Publications, 424, 213-239. DOI: 10.1144/SP424.6.
- Southgate, P.N. (1986). Depositional environment and mechanism of preservation of microfossils, upper Proterozoic Bitter Springs Formation, Australia. Geology, 14(8), 683-686. DOI: 10.1130/0091-7613(1986)14<683:DEAMOP>2.0.CO;2.
- Stern, R.J., Avigad, D., Miller, N.R. and Beyth, M. (2006). Evidence for the Snowball Earth hypothesis in the Arabian-Nubian Shield and the East African Orogen. Journal of African Earth Sciences, 44(1), 1-20. DOI: 10.1016/j.jafrearsci.2005.10.003.
- Strauss, H. and Schieber, J. (1990). A sulfur isotope study of pyrite genesis: The mid-proterozoic Newland Formation, Belt Supergroup, Montana. Geochimica et Cosmochimica Acta, 54(1), 197-204. DOI: 10.1016/0016-7037(90)90207-2.
- Sultan, R.F. and Abdel-Rahman, M. (2013). On dynamic self-organization: examples from magmatic and other geochemical systems. Latin American Journal of Solids and Structures, 10(1), 59-73. DOI: 10.1590/S1679-78252013000100006.
- Symons, D.T.A. (2007). Paleomagnetism of the HYC Zn-Pb SEDEX deposit, Australia: evidence of an epigenetic origin. Economic Geology, 102(7), 1295-1310. DOI: 10.2113/gsecongeo.102.7.1295.
- Tang, D., Shi, X., Wang, X. and Jiang, G. (2016). Extremely low oxygen concentration in mid-Proterozoic shallow seawaters. Precambrian Research, 276, 145-157. DOI: 10.1016/j.precamres.2016.02.005.
- Timmons, J.M., Karlstrom, K.E., Heizler, M.T., Bowring, S.A., Gehrels, G.E. and Crossey, L.J. (2005). Tectonic inferences from the ca. 1255–1100 Ma Unkar Group and Nankoweap Formation, Grand Canyon: Intracratonic deformation and basin formation during protracted Grenville orogenesis. Geological Society of America Bulletin, 117(11-12), 1573-1595. DOI: 10.1130/B25538.1.
- Tucker, M.E. (1984). Calcitic, aragonitic and mixed calcitic-aragonitic ooids from the mid-Proterozoic Belt Supergroup, Montana. Sedimentology, 31(5), 627-644. DOI: 10.1111/j.1365-3091.1984.tb01227.x.
- Turner, E.C., Narbonne, G.M. and James, N.P. (1993). Neoproterozoic reef microstructures from the Little Dal Group, northwestern Canada. Geology, 21(3), 259-262. DOI: 10.1130/0091-7613(1993)021<0259:NRMFTL>2.3.CO;2.
- Valley, J.W., Eiler, J.M., Graham, C.M., Gibson, E.K., Romanek, C.S. and Stolper, E.M. (1997). Low-temperature carbonate concretions in the Martian meteorite ALH84001: Evidence from stable isotopes and mineralogy. Science, 275(5306), 1633-1638. DOI: 10.1126/science.275.5306.1633.
- Varkouhi, S. and Papineau, D. (2023). Silica botryoids from chemically oscillating reactions and as Precambrian environmental proxies. Geology, 51(7), 683–687. DOI: 10.1130/g50948.1.
- Varkouhi, S., Papineau, D., and Guo, Z. (2022) Botryoidal quartz as an abiotic signature in Palaeoarchean cherts of the Pilbara Supergroup, Western Australia. Precambrian Research. DOI: 10.1016/j.precamres.2022.106876.
- Vidal, C. and Pagola, A. (1989). Observed properties of trigger waves close to the center of the target patterns in an oscillating Belousov-Zhabotinskii reagent. The Journal of Physical Chemistry, 93(7), 2711-2716. DOI: 10.1021/j100344a004.
- Walker, R.N., Muir, M.D., Diver, W.L., Williams, N. and Wilkins, N. (1977). Evidence of major sulphate evaporite deposits in the Proterozoic McArthur Group, Northern Territory, Australia. Nature, 265, 526-529. DOI: 10.1038/265526a0.
- Walter, M.R. and Bauld, J. (1983). The association of sulphate evaporites, stromatolitic carbonates and glacial sediments: examples from the Proterozoic of Australia and the Cainozoic of Antarctica. Precambrian Research, 21(1-2), 129-148. DOI: 10.1016/0301-9268(83)90008-6.
- Walter, M.R., Goode, A.D.T. and Hall, W.D.M. (1976). Microfossils from a newly discovered Precambrian stromatolitic iron formation in Western Australia. Nature, 261, 221-223. DOI: 10.1038/261221a0.
- Wang, J., He, Z., Zhu, D., Liu, Q., Ding, Q., Li, S. and Zhang, D. (2020). Petrological and geochemical characteristics of the botryoidal dolomite of Dengying Formation in the Yangtze Craton, South China: Constraints on terminal Ediacaran “dolomite seas”. Sedimentary Geology, 406, 105722. DOI: 10.1016/j.sedgeo.2020.105722.
- Wang, X., Zhang, S., Wang, H., Canfield, D.E., Su, J., Hammarlund, E.U., and Bian, L. (2017) Remarkable preservation of microfossils and biofilms in Mesoproterozoic silicified bitumen concretions from northern China. Geofluids, DOI: 10.1155/2017/4818207.
- White, D. (1928). Algal Deposits of Unkar Proterozoic Age in the Grand Canyon, Arizona. Proceedings of the National Academy of Sciences, 14(7), 597-600. DOI: 10.1073/pnas.14.7.597.
- Wood, R.A., Grotzinger, J.P. and Dickson, J.A.D. (2002). Proterozoic modular biomineralized metazoan from the Nama Group, Namibia. Science, 296(5577), 2383-2386. DOI: 10.1126/science.1071599.
- Xiao, S., Chen, Z., Pang, K., Zhou, C. and Yuan, X. (2020). The Shibantan Lagerstätte: insights into the Proterozoic–Phanerozoic transition. Journal of the Geological Society, 178(1), 135. DOI: 10.1144/jgs2020-135.
- Xiaofeng, W., Erdtmann, B.D., Xiaohong, C. and Xiaodong, M. (1998). Integrated sequence-, bio- and chemostratigraphy of the terminal Proterozoic to Lowermost Cambrian "black rock series" from central South China. Episodes, 21(3), 178-189. DOI: 10.18814/epiiugs/1998/v21i3/007.
- Zaikin, A.N. and Zhabotinsky, A.M. (1970). Concentration Wave Propagation in Two-dimensional liquid-phase self-oscillating system. Nature, 225, 535-537. DOI: 10.1038/225535b0.
- Zhabotinsky, A.M. (1991). A history of chemical oscillations and waves. Chaos: An Interdisciplinary Journal of Nonlinear Science, 1(4), 379-386. DOI: 10.1063/1.165848.
- Zhai, X., Luo, P., Gu, Z., Jiang, H., Zhang, B., Wang, Z., Wang, T. and Wu, S. (2020). Microbial mineralization of botryoidal laminations in the Upper Ediacaran dolostones, Western Yangtze Platform, SW China. Journal of Asian Earth Sciences, 195(27), 104334. DOI: 10.1016/j.jseaes.2020.104334.
Figure 1.
Distribution of various beds of diagenetic spheroids in Proterozoic strata and specifically between 1.8 to 0.541 Ga. The centre of the diagenetic structures’ symbols is aligned to the median age of the formation, with age ranges and further mineralogical information simplified in Table 1. The dark grey outline indicates δ13Ccarb values across the Neoproterozoic and Mesoproterozoic interval obtained from Shields et al. (2019) and Li et al (2022), respectively. Numbers correspond to geological units: 1 = Dengying Formation; 2 = Omkyk Member, Northern Nama Group; 3 = Stáhpogieddi Formation; 4 = Ediacara Member; 5 = Tindir Creek; 6 = Ambed Formation; 7 = Bhander Limestone; 8 = Doushantuo Formation; 9 = Pioneer Sandstone; 10 = Wollogorang Formation; 11 = Backlundtoppen Formation; 12 = Beck Spring Dolomite; 13 = Bitter Springs Formation; 14 = Roan Group; 15 = Callanna Beds; 16 = Little Dal Group; 17 = Diabaig Formation; 18 = Nama Group; 19 = Apache Group; 20 = Bass limestone; 21 = Milbeena Bore; 22 = Independence Fjord Group; 23 = Newland Formation; 24 = Xiamaling Formation; 25 = Helena Formation; 26 = Amelia Dolomite and Mallapunyah Formation; 27 = Jixian System; 28 = Mount Isa mine; 29 = Gaoyuzhuang Formation; 30 = Espinhaço Supergroup; 31 = Barney Creek Formation; 32 = Frere Formation and 33 = Athabasca Group; see Table 1 for more information.
Figure 1.
Distribution of various beds of diagenetic spheroids in Proterozoic strata and specifically between 1.8 to 0.541 Ga. The centre of the diagenetic structures’ symbols is aligned to the median age of the formation, with age ranges and further mineralogical information simplified in Table 1. The dark grey outline indicates δ13Ccarb values across the Neoproterozoic and Mesoproterozoic interval obtained from Shields et al. (2019) and Li et al (2022), respectively. Numbers correspond to geological units: 1 = Dengying Formation; 2 = Omkyk Member, Northern Nama Group; 3 = Stáhpogieddi Formation; 4 = Ediacara Member; 5 = Tindir Creek; 6 = Ambed Formation; 7 = Bhander Limestone; 8 = Doushantuo Formation; 9 = Pioneer Sandstone; 10 = Wollogorang Formation; 11 = Backlundtoppen Formation; 12 = Beck Spring Dolomite; 13 = Bitter Springs Formation; 14 = Roan Group; 15 = Callanna Beds; 16 = Little Dal Group; 17 = Diabaig Formation; 18 = Nama Group; 19 = Apache Group; 20 = Bass limestone; 21 = Milbeena Bore; 22 = Independence Fjord Group; 23 = Newland Formation; 24 = Xiamaling Formation; 25 = Helena Formation; 26 = Amelia Dolomite and Mallapunyah Formation; 27 = Jixian System; 28 = Mount Isa mine; 29 = Gaoyuzhuang Formation; 30 = Espinhaço Supergroup; 31 = Barney Creek Formation; 32 = Frere Formation and 33 = Athabasca Group; see Table 1 for more information.
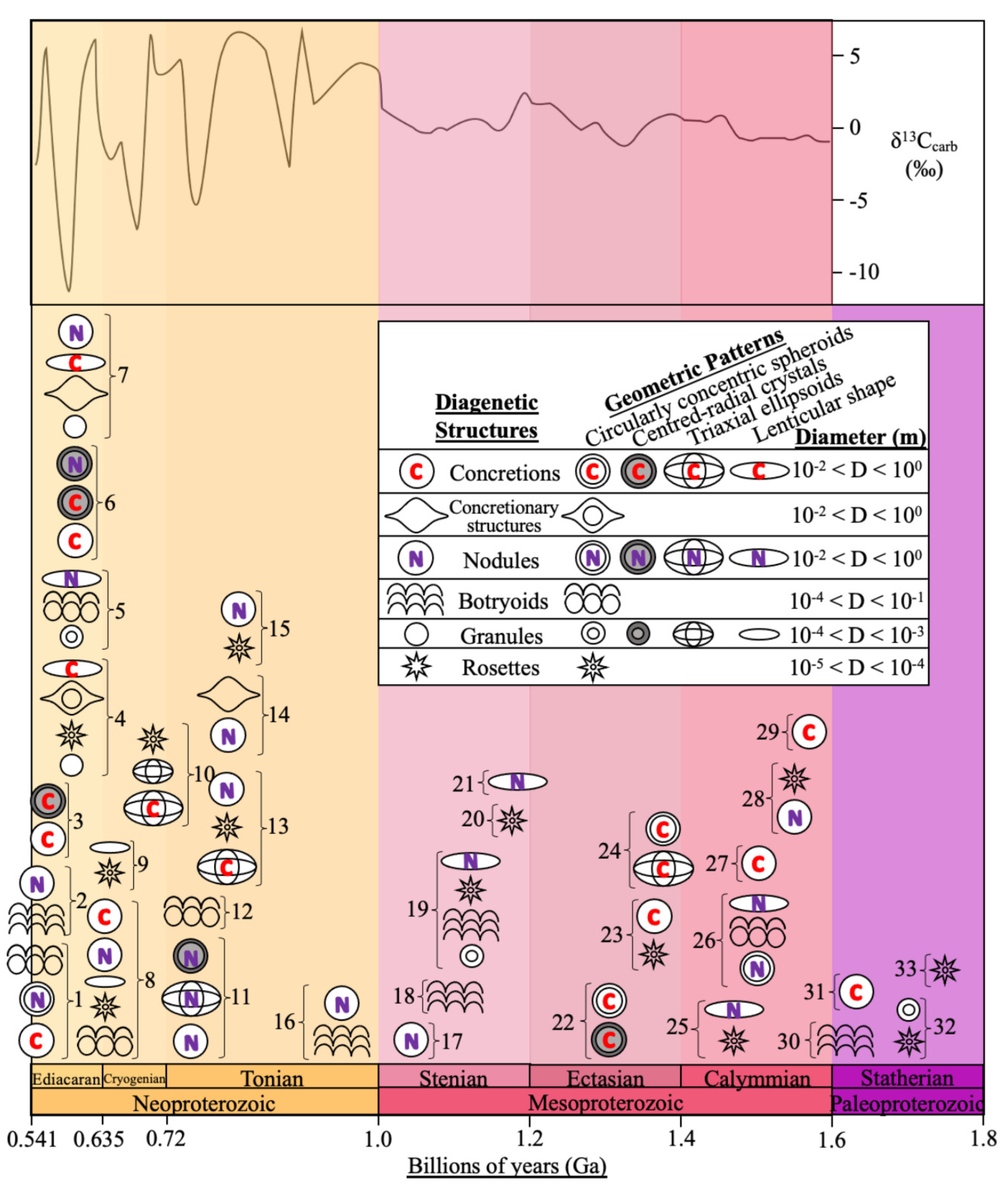
Figure 2.
A selection of common geometric patterns observed within diagenetic spheroids. (a) Botryoidal gypsum forms filled with quartz and acicular pseudomorphs from the 1.1 to 1.4 Ga basal Troy Quartzite in Arizona (Skotnicki and Knauth, 2007). (b) Carbonate concretion with concentric ring structures, from the Xiamaling Formation, 1.35 to 1.4 Ga (Liu et al., 2019). The “D” marks displacive growth whilst “R” displays the areas of replacive growth (Liu et al., 2019). (c) Botryoidal dolomite from the 0.545 to 0.551 Ga Dengying Formation in South China (Wang et al., 2020). (d) Cathodoluminescence photomicrograph displaying a cavity structure of botryoidal dolomite in the Dengying Formation (Wang et al., 2020). (e) Botryoidal laminations from the Dengying Formation (0.545 to 0.551 Ga) in the Gaojiashan outcrop (Zhai et al., 2020). Blue arrows showcase dark lamina (DL) whereas red arrows points towards brighter lamina (BL). All figures reproduced with permission.
Figure 2.
A selection of common geometric patterns observed within diagenetic spheroids. (a) Botryoidal gypsum forms filled with quartz and acicular pseudomorphs from the 1.1 to 1.4 Ga basal Troy Quartzite in Arizona (Skotnicki and Knauth, 2007). (b) Carbonate concretion with concentric ring structures, from the Xiamaling Formation, 1.35 to 1.4 Ga (Liu et al., 2019). The “D” marks displacive growth whilst “R” displays the areas of replacive growth (Liu et al., 2019). (c) Botryoidal dolomite from the 0.545 to 0.551 Ga Dengying Formation in South China (Wang et al., 2020). (d) Cathodoluminescence photomicrograph displaying a cavity structure of botryoidal dolomite in the Dengying Formation (Wang et al., 2020). (e) Botryoidal laminations from the Dengying Formation (0.545 to 0.551 Ga) in the Gaojiashan outcrop (Zhai et al., 2020). Blue arrows showcase dark lamina (DL) whereas red arrows points towards brighter lamina (BL). All figures reproduced with permission.

Figure 3.
Microfossil occurrences in Proterozoic diagenetic spheroids. (a) Outcrop image of nodules containing Namapoikia rietoogensis gen. et sp. nov. from the Omkyk Member, Nama Group, southern Namibia (Wood et al., 2002); (b) Longitudinal holotype and (c) Transverse section (Wood et al., 2002). (d) Oscillatoriopsis obtusa from the Shibantan Lagerstätte (Xiao et al., 2020). (e) A range of microfossils from granules in the Frere Formation, Nabberu Basin, Western Australia (Walter et al., 1976). Figures reproduced with permission.
Figure 3.
Microfossil occurrences in Proterozoic diagenetic spheroids. (a) Outcrop image of nodules containing Namapoikia rietoogensis gen. et sp. nov. from the Omkyk Member, Nama Group, southern Namibia (Wood et al., 2002); (b) Longitudinal holotype and (c) Transverse section (Wood et al., 2002). (d) Oscillatoriopsis obtusa from the Shibantan Lagerstätte (Xiao et al., 2020). (e) A range of microfossils from granules in the Frere Formation, Nabberu Basin, Western Australia (Walter et al., 1976). Figures reproduced with permission.

Figure 4.
Summative grid to compare the self-similar geometric patterns of diagenetic spheroids, shown in plane-polarised light, with those produced by COR. The COR experimental images are from Papineau et al. (2021) and Petri dishes are 10 cm in diameter. (a) Carbonate-apatite concretion with a circularly concentric lamination pattern in phosphorite, Doushantuo Formation in China (Papineau et al., in review). (b) Botryoids composed of fascicular slow dolomite (FSD) cements with chemical growth zonations and circular twins that grew from microbialite substrate, found in Dengying Formation, Yangtze Craton (Wang et al., 2020). The abbreviation of MD stands for micritic dolomite (Wang et al., 2020). (c) Apatite granule from the Doushantuo Formation that shows concentric gradients of brown coloured organic matter along with white arrows indicating linearly elongated cavity structures (She et al., 2013). Figures reproduced with permission.
Figure 4.
Summative grid to compare the self-similar geometric patterns of diagenetic spheroids, shown in plane-polarised light, with those produced by COR. The COR experimental images are from Papineau et al. (2021) and Petri dishes are 10 cm in diameter. (a) Carbonate-apatite concretion with a circularly concentric lamination pattern in phosphorite, Doushantuo Formation in China (Papineau et al., in review). (b) Botryoids composed of fascicular slow dolomite (FSD) cements with chemical growth zonations and circular twins that grew from microbialite substrate, found in Dengying Formation, Yangtze Craton (Wang et al., 2020). The abbreviation of MD stands for micritic dolomite (Wang et al., 2020). (c) Apatite granule from the Doushantuo Formation that shows concentric gradients of brown coloured organic matter along with white arrows indicating linearly elongated cavity structures (She et al., 2013). Figures reproduced with permission.
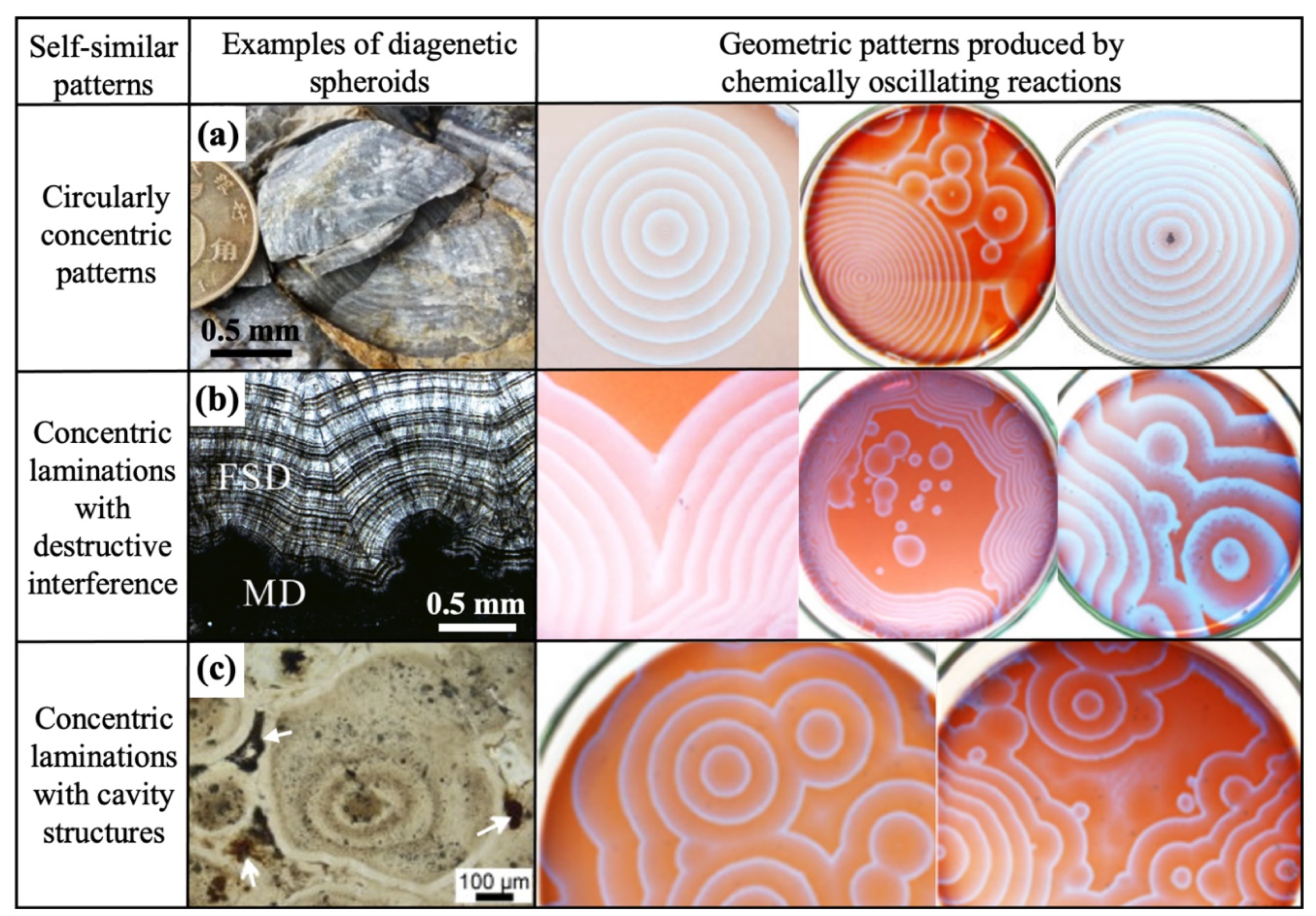

Table 1.
Proterozoic Diagenetic Spheroids.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



