
Submitted:
26 August 2024
Posted:
27 August 2024
You are already at the latest version
Abstract
Lung macrophage cells play a critical role in various lung disease, and their state can change depending on the progression of the disease by inducing either an inflammatory or anti-inflammatory state. In this review, we investigate the potential therapeutic effects of treatment with antioxidant nanoparticles in airway disease focusing on porcine reproductive and respiratory virus (PRRSV), considering Reactive Oxygen Species (ROS) as one of the factors that regulate M1 and M2 macrophages in the inflammatory and anti-inflammatory states, respectively. In addition, we examine the status of protein structure research on CD163 (one of the markers of anti-inflammatory M2 macrophages) in human and veterinary lung diseases and suggest directions for future applicable research areas.
Keywords:
ROS
; PRRSV
; respiratory disease
; M1/M2
; alveolar macrophages
; nanomaterials
1. Background
Lung disease has been one of the leading causes of death worldwide in humans for decades [1]. The spread of the COVID-19 pandemic a few years ago led to many casualties, beginning with coughing symptoms originating from the lungs [2]. This applies not only to human medicine but also to the veterinary field. Distemper in canines [3], Porcine Reproductive Respiratory Syndrome (PRRS) virus in porcines [4], and feline asthma [5] all show a high prevalence and morbidity. To effectively diagnose and treat lung disease, it is important to understand its underlying pathogenesis mechanisms. Specifically, many researchers have recently addressed the role of macrophages in lung diseases. Some studies have focused on the correlation between changes in the state of macrophages, such as general M1/M2, classically activated macrophages, and alternatively activated and lung disease immunity, such as chronic obstructive pulmonary disease (COPD) [6,7]. Another theory centers on the role and correlation of alveolar macrophages and interstitial macrophages in various lung diseases [8,9].
In this paper, we discuss the progression of these M1/M2 theories and suggest future directions for this area.
2. Lung Epithelial Cells and the Status of Common Lung Diseases Associated with M1/M2
There are a variety of lung diseases, including pneumothorax, pneumonia, flu, tuberculosis, and emphysema. Among them, asthma, COPD, and non-small-cell lung cancer are prevalent diseases that significantly harm public health, and much research is being conducted concerning both their pathogenesis and treatment. All chronic respiratory diseases have been reported to have prevalence rate of 7% worldwide in 2017 and COPD (3.9%), Asthma (3.5%) accounting for the majority[10].
Lung epithelial cells play an important role in those lung disease pathogenesis, and they include basal cells, secretory cells, type 1 pneumocytes, and type 2 pneumocytes. Additionally, alveolar macrophages and dendritic cells, along with epithelial cells, react to foreign antigens, and the former play an important role in the pathogenesis of asthma and COPD [11,12]. Harmful substances or cigarette smoking leads to macrophage stimulation, and along with lung epithelial cells, cytokines such as TNF (tumor necrosis factor)-a and IL (interleukin)-8 are secreted from epithelial cells, which induces fibrosis of the bronchial tubes and alveolar macrophage destruction, resulting in irreversible airway obstruction [13]. In addition to exposure to various environmental allergens, cell signals in allergic asthma are transmitted through the epigenetic regulation of histone protein modification, resulting in cytokines such as IL-4 and IL-13 that induce M2 polarization of macrophages [14]. Furthermore, cytokines and chemokines such as TNF-a, LPS, and Reactive Oxygen Species (ROS) also induce M1 polarization in acute lung injuries [15].
The M1 and M2 theories of macrophages were previously conceptualized by the Mantovani research team [16].
Macrophages are differentiated into M1 and M2 in inflammatory and anti-inflammatory states, respectively, depending on whether the immune response is Th1- or Th2-induced, and it has been revealed that ROS, among various cytokines, plays an important role in inducing an inflammatory state [17]. Tissue-stored macrophages can also receive cell signals, such as cytokines and chemokines, depending on the inflammatory situation, and flow into the blood vessels. When additional macrophages are needed, monocytes are differentiated into macrophages in the bone marrow. The states of these macrophages are also polarized from M0 to M1 or M2 to confront situations such as inflammation, cancer, and injury. They circulate within the blood vessels and are stored in the tissues to maintain balance and homeostasis between tissue-resident and circulating macrophages (Figure 1) [18].
Accordingly, researchers have used antioxidant treatments to alter macrophages to an anti-inflammatory state [19,20]. Furthermore, research has been conducted concerning the treatment of diseases through the anti-inflammatory state of macrophages [21]. Research is also being applied to cancers such as non-small-cell lung cancer [22].
Alveolar Macrophages and Interstitial Macrophages
Macrophages are classified into alveolar and interstitial depending on which part of the lung they are located, and research has continued to study the characteristics of these differentiated macrophages to reveal the pathogenesis of lung disease. Alveolar macrophages are known to adhere to and phagocytize antigens faster than interstitial macrophages. These two macrophages achieve homeostasis and change their state according to the inflammatory environment of the lung [8,23]. In this paper, we focus on the M1/M2 theories rather than alveolar/interstitial homeostasis.
3. Sequence–Structure–Function of Antigens and the Application of Macrophage Epitopes in Various Diseases, Including Lung Disease
Previous research demonstrating that the amino acid sequence determines both the protein structure and the function of the protein molecule has become widely known, and to date, research is being conducted to predict the structure of many proteins to diagnose or treat specific diseases [24]. For example, researchers recently discovered the structure of the coronavirus antigen’s spike protein, which was imitated to synthesize the most recent COVID-19 vaccine [25]. This sequence–structure–function theory has also been used for other rare diseases, such as pheochromocytoma, which is rare but fatal. Proteins such as chromogranin A have been used as biomarkers as a result of previous research to reveal their sequence and structure [26].
These efforts have also been applied to macrophages to reveal the structure of macrophage CD163 in inflammatory situations and the mechanism of disease [27] and it is described in Section 4 as GP2, GP4 protein structure. In particular, research is actively underway concerning the pathogenesis of PRRSV, which is one of the most widespread respiratory diseases, causing tremendous economic loss worldwide [28].
4. Lung Disease: Focusing on PRRSV
Porcine reproductive and respiratory syndrome virus (PRRSV) is agent of a mysterious pig disease that was first discovered in the United States and Canada in the late 1980s, causing respiratory diseases, miscarriages, and reproductive diseases such as infertility. In the 1990s, this disease was discovered in European countries such as the UK and France, and also in Korea, China, and Japan. In particular, the virus was isolated for the first time in Korea in 1994 [29]. Since then, PRRSV has caused serious economic damage all over the world and has existed through mutations such as North American type, European type, and highly pathogenic PRRSV.
PRRSV is small, single-stranded, positive-sense, enveloped RNA virus which belongs to the family Arteriviridae. Genomic RNA is 15kb long and consists of a part that translates non-structural proteins (ORF 1a, 1b) and structural proteins (ORFs 2, 3, 4, 5, 6, 7). ORF2, 3, 4, and 5 produce GP 2, 3, 4, and 5, ORF6 produces M protein, and ORF7 produces N protein. [30]. PRRSV is genetically divided into two types, which are European type1 and North American type2 virus. The most popular strains of each type are Lelystad, Netherlands [31] , European strain type1, and VR-2332 [32], North American strain type2. Both North American and European types are structured with a major envelope and minor envelope. The major envelope protein consists of M (174 residues in type2, 173 in type 1) and GP5 (200 residues in type2, 201 in type 1). Minor envelope protein GP2 (256 residues in type 2, 253 in type 1), GP3(254 residues in type 2, 265 in type 1)), GP4 (178 residues in type 2, 183 in type 1. M and GP5 form the domain structure, and GP2, 3, and 4 come together to form a functional structure. [33] (Figure 2)
PRRSV mainly infiltrates the respiratory system of pigs and causes reproductive and respiratory symptoms, and the most important host cells at this time are macrophages in the lung alveolar. Among macrophage viral receptors, CD163 and sialoadhesin have been studied the most. [33]. It is known that the macrophage CD163 marker interacts with viral GP2 and GP4, and the macrophage sialoadhesin marker interacts with GP5, but the specific interaction process has not been fully elucidated [33].
Interestingly, It has been revealed that porcine CD163 provide infection of both European PRRSV (type1) and North American PRRSV (type2) whereas human CD163 supports only type2 and whether type1 is involved is not revealed. Also, CD163 Knock-Out swine is resistant for both type 1,2 PRRSV whereas SRCR5-swap CD163 swine is only resistant for only type1, not type2 [34]. This fact implies the domain of SRCR5 CD163 macrophage receptor and PRRSV minor envelope GP2,4 are crucial keys to elucidate the pathogenesis of PRRSV and different virulence of type1 and type2 may be related to structural difference of interaction site due to residue difference between two types.
There are various routes of infection for PRRSV, including respiratory, oral [35], transdermal, and needle injection [36]. When the virus enters the body through various infection routes, primary proliferation occurs in bronchial mucus, lung parenchymal cell such as type1, 2 pneumocyte, and macrophages. Within 12 hours, secondary proliferation occurs in regional lymph nodes and enters the blood leading to viremia. Afterwards, the systemic infection progresses and becomes a subclinical infection or presents various clinical symptoms such as breathing difficulties, neurological symptoms, and abortion. After recovery, the virus is shed through blood, stool, saliva, semen through persistent infection [37].
When primary infection is caused by complex infection with pathogens such as PRRSV, mycoplasma, and Porcine circovirus (PCV), or when environmental stress such as contaminated gas and dust is combined, the pathogen phagocytosis ability of macrophages and the immunity of bronchial cilia decrease, making it easier for pathogens to infiltrate and proliferate in the body. In addition, complex infection with secondary pathogens such as Actinobacillus pleuropneumoniae, pasteurella multocida, and Haemophilus Parasuis can lead to Porcine respiratory disease complex (PRDC) [38] and further worsen clinical signs.
Clinical symptoms differ slightly depending on the age of the infected pig. For example, for sows, abortion, premature birth, delivery of weak piglets, and stillbirth are shown. [39]. Preweaning piglets have a high mortality rate of close to 100%, neurological symptoms, and respiratory difficulties. Poor growth and increased mortality in raised sows and piglets. In finishing pigs, loss of appetite and high fever are shown and in boars, sperm motility decreases, high fever are representative clinical signs [40].
In North America, Canada and the United States, and in Asia, China, Korea, Japan, and Asia, North American infections dominate, while in European countries such as Italy and Russia, European infections dominate. In particular, many European-type and North American-type complex infections are found in Asia, including Korea. [41]. The highly pathogenic PRRSV, a North American variant, is mainly spread in North America, and in Southeast Asia, including Vietnam, the highly pathogenic PRRSV variant is also widespread. [42]. Prevalence rate of PRRSV among continent is not well documented yet there have been many studies to elucidate the origin of the virus, both type1 [43] and type 2 [44] using genetic epidemiology and phylogenetic tree.
The virus can be excreted for a long time in the order of blood, urine, feces, nasal juice, saliva, and semen, and persistently infected pigs can excrete the virus through pharyngeal mucus for up to 5 months [45]. The spread of the virus includes intra-farm infection and inter-farm infection. Infections within the farm include placental infection, oral, nasal, genital infection, direct and indirect contact, fly, mosquito, air, and poor hygiene of worker infections. Farm-to-farm transmission includes infection through movement of infected pigs, semen, and contaminated vehicles and equipment [35].
Diagnosis of PRRSV can be divided into antigen detection and antibody detection, which can be performed on blood samples, tissues, saliva, semen, etc. There are many types of Antigen test including polymerase chain reaction (PCR), Immunohistochemistry (IHC), virus isolation and in-situ hybridization [46]. Through Antibody testing, Neutralizing Antibody is measured via enzyme-linked immunosorbent assay (ELISA), or indirect fluorescent antibody (IFA)
Additionally, enzyme-linked immune absorbent spot (ELISPOT) which measures PRRSV-specific interferon gamma (IFN-γ) in serum [47] is commonly used these days.
For preventing PRRSV, it is necessary to block horizontal transmission through all-in, all-out strategy and biosecurity, nursery depopulation, and improvement of breeding facilities are also crucial [48]. For monitoring the disease, conducting regular inspections, especially detailed inspections and serological tests for suspicious pigs are key to effectively protect the pig from PRRSV.
Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) is a significant pathogen in the swine industry, causing respiratory disease such as interstitial pneumonia combine with reproductive failure which evokes substantial economic losses worldwide. There are two main types of PRRSV. One is the European (type 1) virus and the other is North American (type 2) which were first isolated in the beginning of 1990s worldwide. Those two types show similar clinical signs except type 2 virus shows stronger virulence when coinfected [49].
PRRSV displays complex interactions with the host immune system, including modulating innate immune responses and employing various immune evasion strategies. One aspect of this dynamic interplay is the involvement of macrophages, which are critical players in the host’s antiviral defense. Particularly, alveolar macrophages, are among the first cells to encounter PRRSV upon respiratory infection among with typeⅡ pneumocyte [50]. When PRRSV encounter macrophage, sialoadhesin, the macrophage-specific receptor is responsible for internalization of the virus. Together with another macrophage marker, CD163, macrophage become more susceptible to PRRSV infection [51]. Not only PRRSV replicate in the host macrophage, they induce apoptosis [52]. Then the virus evades host humoral immunity and cell-mediated immunity which are proven by decrease in neutralizing antibody and PRRSV-specific interferon (IFN)-γ [53]. Interestingly, PRRSV also enters neuroendocrine cell which is shown by the biomarker chromogranin A [54] and it is prominent especially in Highly Pathogenic PRRSV (HP-PRRSV) which is originated from type 2 PPRSV. The higher concentration of neuroendocrine biomarkers in the highly pathogenic PRRSV of North American origin rather than the European type means that the North American virus may have a higher ability to penetrate neuroendocrine organs when it infiltrates the system of porcine. The specific mechanism for this requires additional research.
The virus has evolved various strategies to disrupt the host’s antiviral systems and promote its own survival. A crucial aspect of these strategies involves the interaction between PRRSV and the host’s immune cells, particularly macrophages. PRRSV has been shown to preferentially infect and replicate within M1 and M2 macrophage subsets, with distinct effects on their functions. For example, Type 2 PRRSV can induce M1 polarization of macrophages and Th1 response which are show by the host’s cell surface receptors, CD163 and proinflammatory genes, IFN-γ and IL-12 [55]. Exact mechanism of this process is not fully elucidated but follow up study is still on going.
Importantly, PRRSV infection can also modulate the polarization of macrophages, skewing them towards an M2-like phenotype. Since the M2 type induces a macrophage response related to anti-inflammation, the M2 state may be more advantageous for PRRSV in host invasion, replication, and survival than the inflammatory M1 macrophage state. As briefly described previously, PRRSV regulates the M1 and M2 differentiation of macrophages, thereby regulating the immune response and facilitating virus invasion [56]. If this control can be artificially controlled in the opposite direction to PRRSV through drug administration, a more efficient vaccine effect can be expected. Additionally, this macrophage differentiation strategy has the potential to be applied not only to PRRSV but also to other lung diseases. Many studies have shown that treatment with nanoparticles can provide an antioxidant effect, and it has been shown in vitro and in vivo experiments [57,58]. Through this antioxidant activity, Reactive Oxygen Species (ROS) can be removed which is essential chemokine for M1 polarization so that the state of macrophages can be changed from the M1 state to the M2 state eventually. Therefore, if an appropriate nanoparticle candidate is selected to scavenge ROS, it is possible to develop a lung disease medicine or adjuvant that is harmless in the body. Among the many candidates, selenium and magnesium, which are essential trace elements in the body, are harmless to the body and show antioxidant activity comparable to that of heavy metals such as silver or gold [59,60]. If such nanomaterial synthesis research is conducted along with macrophage differentiation, it will not only provide more effective PRRSV defense but also be applicable to various lung diseases. This could be promising strategy because in among various infectious disease such as human influenza, treating the disease with nanoparticle has been successful already decade ago [61] and it also has been applied to animal diseases in veterinary field these days [62]. Among with studies elucidate the structural interaction of CD 163 macrophage receptor, treating nanomaterial to control macrophage polarization can provide key factor to prevent lung diseases including PRRSV.
5. Therapeutic Effects of Changes in Macrophage Status through ROS Scavenging through Nanoparticles
Currently, various researchers, such as Korea’s Hyeon Taeghwan group and China’s Zhiyuan Zhong group, have conducted studies on the therapeutic effects of macrophage polarization in treating inflammatory diseases with nanoparticles. Successful research results have been achieved in the fields of Crohn’s disease IBD [63], sepsis [64], and rheumatism [65] using ROS scavenging.
Research has also been conducted on the role and differentiation of macrophages in various lung diseases, such as COPD [66], non-small-cell lung cancer [67], and acute/chronic inflammatory lung disease [68]. In particular, in non-small-cell lung cancer, the expression of CD68 and the M2 marker CD163 increased, whereas that of iNOS (the M1 marker) decreased [69].
Combining the concept of macrophage modulation through nanoparticles and the role of macrophages in lung diseases, the treatment of lung diseases is also progressing through macrophage modulation using nanoparticles [70]. However, this research has been conducted recently compared to other diseases. The goal of inducing macrophage differentiation differs depending on both the inflammatory and cancer situations. M2 are anti-inflammatory macrophages that promote tumor development and metastasis, whereas M1 are pro-inflammatory macrophages that suppress tumor development and metastasis [71].
When synthesizing nanoparticles using the method below (using selenium and manganese, which are essential trace elements, instead of ceria, which is a heavy metal), comparable ROS scavenging ability was found [72]. During the preliminary tests, selenium–albumin and manganese–albumin nanocomposites exhibited excellent antioxidative effects in vitro, as did ceria–albumin nanomaterials. Although more experiments and research should be conducted, including in vivo tests to apply this material, follow-up research can be conducted to investigate the therapeutic effects of lung disease inflammation (Figure 3). This strategy would not be suitable for lung cancer diseases, as inducing the M2 phenotype promotes tumor progression. It is also known that M2 macrophages cause a significant rise in asthma [73], COPD [74], and lung fibrosis [75]. However, the ROS scavenging strategy focuses on recovering M1/M2 homeostasis under bias towards the M1 population environment. Therefore, the mouse model incorporating this Se/Mn/albumin nanomaterial that is recommended for in vivo testing must consider macrophages in the M1 state that are dominant in severe and critical lung diseases that cause sepsis such as COVID-19.
Materials. All chemicals were purchased from Sigma-Aldrich and used as-received.
Ultrapure Millipore water (18.2 M Ω) was used. All glassware was washed with chromic acid lotion and then rinsed with ultrapure water. (Caution: chromic acid lotion is a very corrosive oxidizing agent, which should be handled with great care.)
Synthesis of CNLs. In a typical experiment, 0.25 mL 0.1 M Ce(NO3)3.6H2O aqueous solution was added to 4.5 mL 25 mg/mL BSA (or HSA) solution under vigorous stirring at 37 °C for 15 min. 0.3 mL 1 M KOH solution was quickly added into above mixture, and the reaction was kept stirring for 15 min. Then, the obtained CNLs were dialyzed in ultrapure water with dialysis bag (Thermo, 10 kd) for 24 h (notice: change the ultrapure water every 6 hours). For further purification, the CNLs were placed in sleeve tube (Millipore, 100 kd) with 4500 r/min by centrifugation and rinsed with ultrapure water repeatedly. Finally, the obtained CNLs were dispersed in water and stored at 4 °C before used.” [72] (direct quotation).
5.1. Effects of Nanoparticle Treatment on the Human Body
When treating the human body with nanoparticles, these could be deposited into the bone marrow, spleen, and CNS, which is a critical issue because the spleen is an organ where various immune cells are activated. We also have to consider accumulation of nanoparticle in human body when it is applied to PRRSV because of characteristic of economic, food animal. If heavy metal nanomaterials are deposited in the spleen, bone marrow, or CNS, there may be unexpected side-effects, which may cause more harm than good by treatment. According to a past study, a nanomaterial-bound anticancer drug showed higher bone marrow toxicity than a nanomaterial-free anticancer drug [76]. Therefore, researchers have tried to treat cancer or related diseases with nanomaterials while minimizing the impact on the spleen and bone marrow by directly delivering the nanoparticles to the target organ using other cells such as nanoparticle-attached red blood cells [77]. Nanoparticles must eventually be absorbed into the body or eliminated through the kidneys. The spleen is responsible for facilitating absorption into the body and removing blood cells through macrophages. Hepatic circulation, splenic uptake, and elimination of the nanoparticles are the critical parts of biodistribution [78,79]. Moreover, the spleen is an organ responsible for immunity in our body, and it contains macrophages in various states [80]. Depending on external stimulation, it is differentiated into the M1 or M2 state [81]. Spleen-derived macrophages play a protective role in lung inflammation [82]. Additionally, some studies have found decreased responsiveness in patients with interstitial lung disease due to splenic reuptake of the drug. [83]. Considering these points together, when treating lung disease using nanoparticles, it is imperative to consider side-effects due to nanoparticle absorption in the body and reuptake in the spleen, in addition to maintaining M1/M2 balance through the ROS scavenging strategy. This further supports the fact that nanoparticles should be synthesized not with heavy metals but with essential trace elements such as selenium and manganese, which can minimize side-effects, and albumin, which is a basic component of cells but has antioxidant properties. Various studies have also found selenium to be hepatoprotective against heavy metal toxicity [84,85]. Albumin treatment is beneficial as a natural lung surfactant [86], and it was shown that inhalable albumin-based nanoparticles exhibited excellent potency in lung cancer and was drug-resistant [87]. This suggests that, instead of commonly used heavy metal nanoparticles including silver, gold, and ceria, the former selenium–human albumin-based nanoparticle inhalant is an excellent candidate in treating lung diseases such as COVID-19 or sepsis-induced lung damage.
6. Macrophages and Lung Diseases in the Veterinary Field
Therefore, macrophages play an important role in the pathogenesis of lung diseases of various species, and in particular, research into M2 has been conducted by altering the state of macrophages from M1 to M2 through studies on the protein structure and function of CD163 and through treatment with nanoparticles to prevent disease.
Especially in PRRSV, Scavenging ROS through selenium-albumin nanomaterial treatment can lead to polarization of macrophage into M1 state to M2 state which express anti-inflammatory cytokines such as IL-10, IL-4, IL-13 result in upregulation of phagocytosis, PPAR signaling (Figure 3). The suggested treatments have caught the attention of many scientists in this field. Relevant research in the veterinary field is also in progress. For example, one study discovered that the CD163+ alveolar macrophages of sheep were latently infected with the OPP virus antigen. However, the direct role of macrophages in the infection mechanism has not yet been revealed. Furthermore, the role of porcine AM in the PRRS infection has not yet been fully elucidated. It was also structurally revealed that CD163, especially the SRCR5 receptor, plays a critical role [88]. Accordingly, our research team explored areas in which additional research could be conducted in the veterinary field and compared the amino acid sequences of CD163 macrophages from humans, dogs, cats, and pigs (Figure 4). As a result, except for cats, humans, dogs, and cats had almost identical amino acid sequences. Because the sequence of CD163 slightly differs in cats, there may be differences when applying the existing M1 and M2 theories to lung disease. Limited research has been conducted on cat CD163 and the infection mechanism of lung disease. If research on this progresses, new treatments may be developed for intractable lung diseases such as asthma, latent herpes infection, and chylothorax.
7. Conclusions and Perspectives
In this paper, we briefly reviewed the types of lung diseases in the human medicine and veterinary fields, the role and differentiation status of macrophages in lung diseases, the status of treatment of various diseases using macrophages, and nanomaterials that can be applied to lung diseases. Especially, macrophages play an important role in the pathogenesis of lung diseases of various species, and by treating nanomaterial which consist of trace essential elements, we can successfully scavenge ROS without damage or accumulation to the host body and lead M2 polarization from M1 state. It can be utilized into treatment of Covid-19 induced sepsis or other lung disease such as PRRSV. Furthermore, the role of porcine AM in the PRRS infection has not yet been fully elucidated. It was also structurally revealed that CD163, especially the SRCR5 receptor, plays a critical role in interaction with GP2, GP4 of epitope. Along with M1/M2 strategy, better understanding of functional domain could be the key to elucidate this interaction mechanism and will provide efficient protection of the disease. In human medicine, M1/M2 theories can be applied to lung inflammatory diseases by treating nanoparticles to polarize macrophages to M2 through ROS scavenging, especially selenium–manganese albumin nanomaterials, which are powerful antioxidants that are harmless to the human body. This study also addresses the importance of the impact of nanoparticles on human body organs such splenic reuptake and bone marrow deposition toxicity. In veterinary medicine, M1/M2 theories could be applied to porcine PRRS virus infection and other animal lung diseases, such as the sheep OPP virus. Finally, it suggests the need for follow-up research to explore the role of macrophages in various feline lung diseases through sequence–structure–function theory via structural research on feline CD163. We hope that many scientists, including our team, will proceed with the follow-up studies listed above.
Author Contributions
KC developed the idea, gathered the information and preliminary data, and wrote the manuscript. All authors reviewed the manuscript
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors
Conflicts of Interest
The authors claim no conflicts of interest.
Abbreviations
ROS (Reactive Oxygen Species), AM (alveolar macrophage), IM (interstitial macrophage), PRRSV (Porcine reproductive and respiratory syndrome virus)
References
- Schluger, N.W. and R. Koppaka, Lung disease in a global context. A call for public health action. Annals of the American Thoracic Society, 2014. 11(3): p. 407-416. [CrossRef]
- Shi, Y., et al., An overview of COVID-19. Journal of Zhejiang University. Science. B, 2020. 21(5): p. 343.
- Deem, S.L., et al., Canine distemper in terrestrial carnivores: a review. Journal of Zoo and Wildlife medicine, 2000. 31(4): p. 441-451.
- Chae, C., Commercial PRRS modified-live virus vaccines. Vaccines, 2021. 9(2): p. 185.
- Reinero, C.R., Advances in the understanding of pathogenesis, and diagnostics and therapeutics for feline allergic asthma. The Veterinary Journal, 2011. 190(1): p. 28-33. [CrossRef]
- Arora, S., et al., Macrophages: Their role, activation and polarization in pulmonary diseases. Immunobiology, 2018. 223(4-5): p. 383-396.
- Byrne, A.J., et al., Pulmonary macrophages: key players in the innate defence of the airways. Thorax, 2015. 70(12): p. 1189-1196.
- Shi, T., et al., Alveolar and lung interstitial macrophages: Definitions, functions, and roles in lung fibrosis. Journal of Leukocyte Biology, 2021. 110(1): p. 107-114.
- Liegeois, M., et al., The interstitial macrophage: A long-neglected piece in the puzzle of lung immunity. Cellular immunology, 2018. 330: p. 91-96.
- Soriano, J.B., et al., Prevalence and attributable health burden of chronic respiratory diseases, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. The Lancet Respiratory Medicine, 2020. 8(6): p. 585-596.
- Barnes, P.J., Alveolar macrophages as orchestrators of COPD. COPD: Journal of Chronic Obstructive Pulmonary Disease, 2004. 1(1): p. 59-70.
- Fricker, M.and P.G. Gibson, Macrophage dysfunction in the pathogenesis and treatment of asthma. European Respiratory Journal, 2017. 50(3). [CrossRef]
- Strzelak, A., et al., Tobacco Smoke Induces and Alters Immune Responses in the Lung Triggering Inflammation, Allergy, Asthma and Other Lung Diseases: A Mechanistic Review. Int J Environ Res Public Health, 2018. 15(5).
- Saradna, A., et al., Macrophage polarization and allergic asthma. Translational Research, 2018. 191: p. 1-14.
- Lu, H.-L., et al., Activation of M1 macrophages plays a critical role in the initiation of acute lung injury. Bioscience reports, 2018. 38(2): p. BSR20171555.
- Biswas, S.K.and A. Mantovani, Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nature immunology, 2010. 11(10): p. 889-896.
- Covarrubias, A. Byles, and T. Horng, ROS sets the stage for macrophage differentiation. Cell research, 2013. 23(8): p. 984-985.
- Gordon, S. and P.R. Taylor, Monocyte and macrophage heterogeneity. Nature reviews immunology, 2005. 5(12): p. 953-964.
- Tsai, C.-F., et al., Regulatory effects of quercetin on M1/M2 macrophage polarization and oxidative/antioxidative balance. Nutrients, 2021. 14(1): p. 67.
- Kwon, D.H., et al., Glutathione induced immune-stimulatory activity by promoting M1-like macrophages polarization via potential ROS scavenging capacity. Antioxidants, 2019. 8(9): p. 413.
- Griess, B., et al., Scavenging reactive oxygen species selectively inhibits M2 macrophage polarization and their pro-tumorigenic function in part, via Stat3 suppression. Free Radical Biology and Medicine, 2020. 147: p. 48-60.
- Hao, Y., et al., Critical Role of the Sulfiredoxin-Peroxiredoxin IV Axis in Urethane-Induced Non-Small Cell Lung Cancer. Antioxidants, 2023. 12(2): p. 367.
- Fathi, M., et al., Functional and morphological differences between human alveolar and interstitial macrophages. Experimental and molecular pathology, 2001. 70(2): p. 77-82.
- Whisstock, J.C. and A.M. Lesk, Prediction of protein function from protein sequence and structure. Quarterly reviews of biophysics, 2003. 36(3): p. 307-340.
- Wrapp, D., et al., Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science, 2020. 367(6483): p. 1260-1263.
- Choi, K. Han, and S.J. Kim, A systematic review of chromogranin A (CgA) and its biomedical applications, unveiling its structure-related functions. Journal of the Korean Physical Society, 2021. 78: p. 427-441.
- Etzerodt, A., et al., Structural basis for inflammation-driven shedding of CD163 ectodomain and tumor necrosis factor-α in macrophages. Journal of Biological Chemistry, 2014. 289(2): p. 778-788. [CrossRef]
- Ma, H., et al., The crystal structure of the fifth scavenger receptor cysteine-rich domain of porcine CD163 reveals an important residue involved in porcine reproductive and respiratory syndrome virus infection. Journal of Virology, 2017. 91(3): p. 10.1128/jvi. 01897-16.
- Kweon, C.-h., et al., Isolation of porcine reproductive and respiratory syndrome virus (PRRSV) in Korea. Korean Journal of Veterinary Research, 1994. 34(1): p. 77-83.
- Meulenberg, J., PRRSV, the virus. Veterinary research, 2000. 31(1): p. 11-21.
- Wensvoort, G., et al., Mystery swine disease in the Netherlands: the isolation of Lelystad virus. 1993.
- Collins, J.E., et al., Isolation of swine infertility and respiratory syndrome virus (isolate ATCC VR-2332) in North America and experimental reproduction of the disease in gnotobiotic pigs. Journal of Veterinary Diagnostic Investigation, 1992. 4(2): p. 117-126.
- Dokland, T., The structural biology of PRRSV. Virus research, 2010. 154(1-2): p. 86-97.
- Su, C.-M. R.R. Rowland, and D. Yoo, Recent advances in PRRS virus receptors and the targeting of receptor–ligand for control. Vaccines, 2021. 9(4): p. 354.
- Wills, R.W., et al., Porcine reproductive and respiratory syndrome virus: routes of excretion. Veterinary microbiology, 1997. 57(1): p. 69-81. [CrossRef]
- Prieto, C. and J.M. Castro, Porcine reproductive and respiratory syndrome virus infection in the boar: a review. Theriogenology, 2005. 63(1): p. 1-16.
- Beyer, J. , et al., Porcine reproductive and respiratory syndrome virus (PRRSV): kinetics of infection in lymphatic organs and lung. Journal of Veterinary Medicine, Series B, 2000. 47(1): p. 9-25.
- Bochev, I., Porcine respiratory disease complex (PRDC): A review. I. Etiology, epidemiology, clinical forms and pathoanatomical features. Bulgarian Journal of Veterinary Medicine, 2007. 10(3): p. 131-146.
- Pejsak, Z. Stadejek, and I. Markowska-Daniel, Clinical signs and economic losses caused by porcine reproductive and respiratory syndrome virus in a large breeding farm. Veterinary microbiology, 1997. 55(1-4): p. 317-322.
- Prieto, C., et al., Semen changes in boars after experimental infection with porcine reproductive and respiratory syndrome (PRRS) virus. Theriogenology, 1996. 45(2): p. 383-395.
- Ruedas-Torres, I., et al., The jigsaw of PRRSV virulence. Veterinary Microbiology, 2021. 260: p. 109168.
- Guo, B., et al., Chinese and Vietnamese strains of HP-PRRSV cause different pathogenic outcomes in United States high health swine. Virology, 2013. 446(1-2): p. 238-250.
- Balka, G., et al., Genetic diversity of PRRSV 1 in Central Eastern Europe in 1994–2014: origin and evolution of the virus in the region. Scientific Reports, 2018. 8(1): p. 7811.
- Jakab, S., et al., Genetic diversity of imported PRRSV-2 strains, 2005–2020, Hungary. Frontiers in Veterinary Science, 2022. 9: p. 986850.
- Pileri, E. and E. Mateu, Review on the transmission porcine reproductive and respiratory syndrome virus between pigs and farms and impact on vaccination. Veterinary research, 2016. 47(1): p. 108.
- Haynes, J., et al., Temporal and morphologic characterization of the distribution of porcine reproductive and respiratory syndrome virus (PRRSV) by in situ hybridization in pigs infected with isolates of PRRSV that differ in virulence. Veterinary pathology, 1997. 34(1): p. 39-43.
- Piras, F., et al., Porcine Reproductive and Respiratory Syndrome (PRRS) virus-specific interferon-γ+ T-cell responses after PRRS virus infection or vaccination with an inactivated PRRS vaccine. Viral immunology, 2005. 18(2): p. 381-389.
- Silva, G.S., et al., Development and validation of a scoring system to assess the relative vulnerability of swine breeding herds to the introduction of PRRS virus. Preventive veterinary medicine, 2018. 160: p. 116-122.
- Han, K., et al., Comparative pathogenesis of type 1 (European genotype) and type 2 (North American genotype) porcine reproductive and respiratory syndrome virus in infected boar. Virology journal, 2013. 10: p. 1-9.
- Beyer, J., et al., Arterivirus PRRSV: experimental studies on the pathogenesis of respiratory disease. Coronaviruses and Arteriviruses, 1998: p. 593-599.
- Van Gorp, H., et al., Sialoadhesin and CD163 join forces during entry of the porcine reproductive and respiratory syndrome virus. Journal of General Virology, 2008. 89(12): p. 2943-2953. [CrossRef]
- Choi, C. and C. Chae, Expression of tumour necrosis factor-α is associated with apoptosis in lungs of pigs experimentally infected with porcine reproductive and respiratory syndrome virus. Research in Veterinary Science, 2002. 72(1): p. 45-49.
- Mateu, E. and I. Díaz, The challenge of PRRS immunology. The Veterinary Journal, 2008. 177(3): p. 345-351.
- Tong, J. , et al., Hypothalamus-pituitary-adrenal axis involves in anti-viral ability through regulation of immune response in piglets infected by highly pathogenic porcine reproductive and respiratory syndrome virus. BMC veterinary research, 2018. 14: p. 1-7.
- Wahyuningtyas, R., et al., Recombinant antigen of type 2 porcine reproductive and respiratory syndrome virus (PRRSV-2) promotes M1 repolarization of porcine alveolar macrophages and Th1 type response. Vaccines, 2021. 9(9): p. 1009. [CrossRef]
- Gong, X., et al., Porcine reproductive and respiratory syndrome virus modulates the switch of macrophage polarization from M1 to M2 by upregulating moDC-released sCD83. Viruses, 2023. 15(3): p. 773.
- Sharpe, E. Andreescu, and S. Andreescu, Artificial nanoparticle antioxidants, in Oxidative stress: diagnostics, prevention, and therapy. 2011, ACS Publications. p. 235-253.
- Malhotra, S., et al., In vitro and in vivo antioxidant, cytotoxic, and anti--chronic inflammatory arthritic effect of selenium nanoparticles. Journal of Biomedical Materials Research Part B: Applied Biomaterials, 2016. 104(5): p. 993-1003.
- Battin, E.E. and J.L. Brumaghim, Antioxidant activity of sulfur and selenium: a review of reactive oxygen species scavenging, glutathione peroxidase, and metal-binding antioxidant mechanisms. Cell biochemistry and biophysics, 2009. 55(1): p. 1-23.
- Medhe, S. Bansal, and M.M. Srivastava, Enhanced antioxidant activity of gold nanoparticle embedded 3, 6-dihydroxyflavone: a combinational study. Applied Nanoscience, 2014. 4: p. 153-161.
- Okamoto, S., et al., Poly (γ-glutamic acid) nano-particles combined with mucosal influenza virus hemagglutinin vaccine protects against influenza virus infection in mice. Vaccine, 2009. 27(42): p. 5896-5905.
- Sun, M., et al., Toward innovative veterinary nanoparticle vaccines. Animal Diseases, 2024. 4(1): p. 14.
- Yang, J., et al., An oral nano-antioxidant for targeted treatment of inflammatory bowel disease by regulating macrophage polarization and inhibiting ferroptosis of intestinal cells. Chemical Engineering Journal, 2023. 465: p. 142940.
- Soh, M., et al., Ceria–Zirconia nanoparticles as an enhanced multi--antioxidant for sepsis treatment. Angewandte Chemie, 2017. 129(38): p. 11557-11561.
- Fang, H., et al., Macrophage-targeted hydroxychloroquine nanotherapeutics for rheumatoid arthritis therapy. ACS applied materials & interfaces, 2022. 14(7): p. 8824-8837.
- Eapen, M.S., et al., Abnormal M1/M2 macrophage phenotype profiles in the small airway wall and lumen in smokers and chronic obstructive pulmonary disease (COPD). Scientific reports, 2017. 7(1): p. 13392.
- Jackute, J., et al., Distribution of M1 and M2 macrophages in tumor islets and stroma in relation to prognosis of non-small cell lung cancer. BMC immunology, 2018. 19: p. 1-13.
- Lee, J.-W., et al., The role of macrophages in the development of acute and chronic inflammatory lung diseases. Cells, 2021. 10(4): p. 897.
- Almatroodi, S.A., et al., Characterization of M1/M2 tumour-associated macrophages (TAMs) and Th1/Th2 cytokine profiles in patients with NSCLC. Cancer Microenvironment, 2016. 9: p. 1-11.
- Wang, L., et al., Manipulation of macrophage polarization by peptide-coated gold nanoparticles and its protective effects on acute lung injury. Journal of nanobiotechnology, 2020. 18: p. 1-16.
- Schweer, D., et al., Tumor-associated macrophages and ovarian cancer: implications for therapy. Cancers, 2022. 14(9): p. 2220.
- Yang, Z., et al., Albumin-mediated biomineralization of shape-controllable and biocompatible ceria nanomaterials. ACS Applied Materials & Interfaces, 2017. 9(8): p. 6839-6848.
- Girodet, P.-O., et al., Alternative macrophage activation is increased in asthma. American journal of respiratory cell and molecular biology, 2016. 55(4): p. 467-475.
- Vlahos, R. and S. Bozinovski, Role of alveolar macrophages in chronic obstructive pulmonary disease. Frontiers in immunology, 2014. 5: p. 435.
- Braga, T.T. S.H. Agudelo, and N.O.S. Camara, Macrophages during the fibrotic process: M2 as friend and foe. Frontiers in immunology, 2015. 6: p. 602.
- Gibaud, S., et al., Increased bone marrow toxicity of doxorubicin bound to nanoparticles. European Journal of Cancer, 1994. 30(6): p. 820-826.
- Anselmo, A.C., et al., Delivering nanoparticles to lungs while avoiding liver and spleen through adsorption on red blood cells. ACS nano, 2013. 7(12): p. 11129-11137.
- Drozdov, A.S. I. Nikitin, and J.M. Rozenberg, Systematic review of cancer targeting by nanoparticles revealed a global association between accumulation in tumors and spleen. International journal of molecular sciences, 2021. 22(23): p. 13011.
- Demoy, M., et al., Spleen capture of nanoparticles: influence of animal species and surface characteristics. Pharmaceutical research, 1999. 16: p. 37-41.
- Den Haan, J.M. and G. Kraal, Innate immune functions of macrophage subpopulations in the spleen. Journal of innate immunity, 2012. 4(5-6): p. 437-445.
- Mulder, R. Banete, and S. Basta, Spleen-derived macrophages are readily polarized into classically activated (M1) or alternatively activated (M2) states. Immunobiology, 2014. 219(10): p. 737-745.
- Venosa, A., et al., Protective role of spleen-derived macrophages in lung inflammation, injury, and fibrosis induced by nitrogen mustard. American Journal of Physiology-Lung Cellular and Molecular Physiology, 2015. 309(12): p. L1487-L1498.
- Adams, H., et al., Altered splenic [89Zr] Zr-rituximab uptake in patients with interstitial lung disease not responding to rituximab: could this indicate a splenic immune-mediated mechanism? American Journal of Nuclear Medicine and Molecular Imaging, 2020. 10(4): p. 168.
- Newairy, A., et al., The hepatoprotective effects of selenium against cadmium toxicity in rats. Toxicology, 2007. 242(1-3): p. 23-30. [CrossRef]
- Messarah, M., et al., Hepatoprotective role and antioxidant capacity of selenium on arsenic-induced liver injury in rats. Experimental and toxicologic pathology, 2012. 64(3): p. 167-174.
- Holm, B. Notter, and J. Finkelstein, Surface property changes from interactions of albumin with natural lung surfactant and extracted lung lipids. Chemistry and physics of lipids, 1985. 38(3): p. 287-298.
- Choi, S.H., et al., Inhalable self-assembled albumin nanoparticles for treating drug-resistant lung cancer. Journal of controlled release, 2015. 197: p. 199-207.
- Ma, H., et al., Structural comparison of CD163 SRCR5 from different species sheds some light on its involvement in porcine reproductive and respiratory syndrome virus-2 infection in vitro. Veterinary Research, 2021. 52: p. 1-17.
- Barros, M.H.M., et al., Macrophage polarisation: an immunohistochemical approach for identifying M1 and M2 macrophages. PloS one, 2013. 8(11): p. e80908.
Figure 1.
Alveolar macrophages, interstitial macrophages, and monocytes (differentiated from bone marrow). Illustration of bones in a lung-inflammatory state. Monocytes are differentiated in the bone marrow, and alveolar macrophages are differentiated into the M2 state to phagocytose pollutants and fight inflammation [89].
Figure 1.
Alveolar macrophages, interstitial macrophages, and monocytes (differentiated from bone marrow). Illustration of bones in a lung-inflammatory state. Monocytes are differentiated in the bone marrow, and alveolar macrophages are differentiated into the M2 state to phagocytose pollutants and fight inflammation [89].
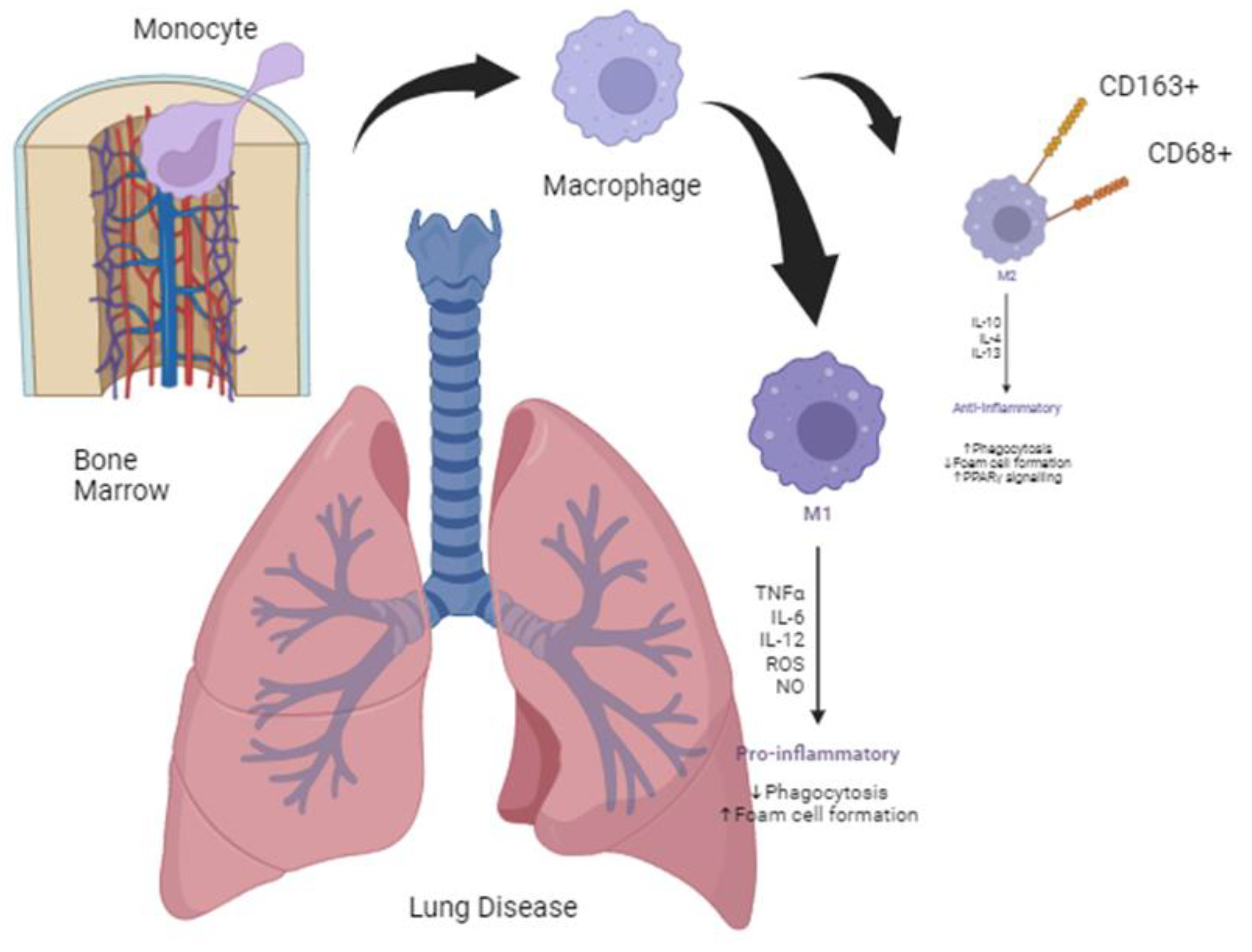
Figure 2.
Illustration of PRRSV structure made by SWISS-MODEL prediction program (https://swissmodel.expasy.org).
Figure 2.
Illustration of PRRSV structure made by SWISS-MODEL prediction program (https://swissmodel.expasy.org).
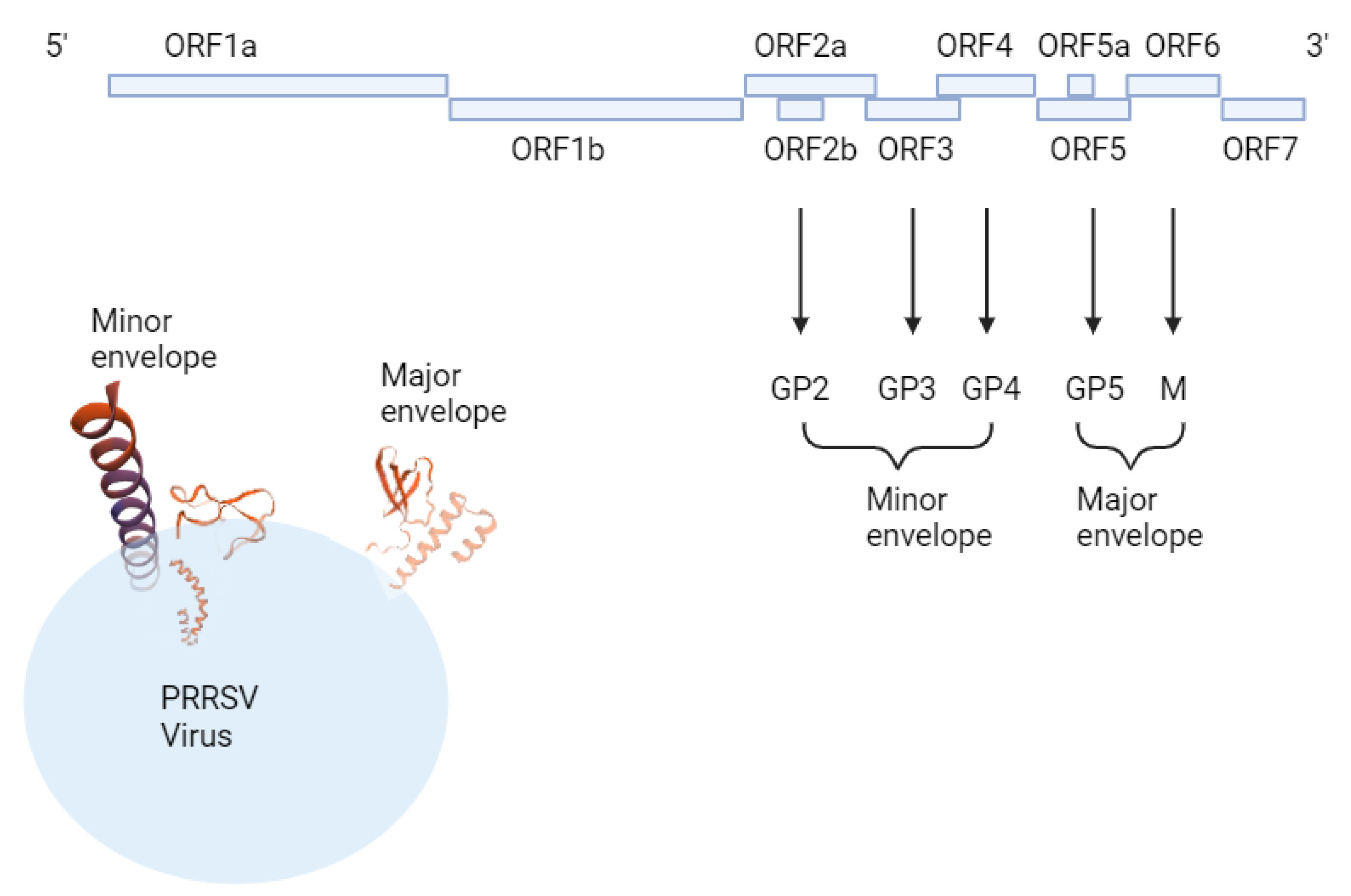
Figure 3.
The Mn-Se-HSA nanocomposite is used to treat inflammation by altering the state of macrophages from M1 and M0 to M2, which is anti-inflammatory by blocking NO and ROS that affect M1 polarization due to ROS scavenging.
Figure 3.
The Mn-Se-HSA nanocomposite is used to treat inflammation by altering the state of macrophages from M1 and M0 to M2, which is anti-inflammatory by blocking NO and ROS that affect M1 polarization due to ROS scavenging.
Figure 4.
Amino acid sequence comparison of human, canine, feline, and porcine macrophage CD163. Humans, canines, and porcines show almost similar sequences, except for felines. This suggests that, according to sequence–structure–function theory, the role of macrophages in lung disease may show similar tendencies in humans, canines, and porcines, but may differ in felines.
Figure 4.
Amino acid sequence comparison of human, canine, feline, and porcine macrophage CD163. Humans, canines, and porcines show almost similar sequences, except for felines. This suggests that, according to sequence–structure–function theory, the role of macrophages in lung disease may show similar tendencies in humans, canines, and porcines, but may differ in felines.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



