
Submitted:
26 August 2024
Posted:
27 August 2024
You are already at the latest version
Abstract
The utilisation of non-Saccharomyces yeasts in co-inoculation and non-thermal technologies for must sterilisation is becoming increasingly prevalent due to its notable utility and potential. This approach optimises the fermentation process, facilitates the production of wines with distinctive characteristics and enhances their stability. Two trials were conducted concurrently, designated as A and B. The same red must was utilized in both trials, with one undergoing UHPH treatment and the other serving as the control. The non-Saccharomyces yeasts were identical in both trials. In Trial A, different percentages of initial inoculum were employed with respect to the total volume of must to be fermented, with the objective of evaluating the influence and competitiveness between yeasts. Trial B was designed to investigate the impact of two nutrients that provide vitamins, energy and protection from oxidative stress on the development of these yeasts and their metabolic expression. A microbiological analysis was conducted, along with oenological parameter measurements, acidification assessments, volatile compound analyses, and spectrophotometric evaluations. It was observed in both trials that the prevalence of Lachancea thermotolerans (Lt) was longer than that of Metschnikowia pulcherrima (Mp), and that the use of quercetin + thiamine had a positive effect on yeast growth. Furthermore, the combination of Lt and Mp yeasts demonstrated remarkable synergy, resulting in the production of a substantial quantity of lactic acid (>5 g/L). With regard to aroma compounds, the UHPH must exhibited a nearly twofold increase in ethyl lactate. Additionally, the TPI was observed to be 8-10% higher in wines derived from UHPH musts, indicating that this technology may potentially safeguard against oxidation.
Keywords:
Wine
; fermentation
; non-Saccharomyces
; biocompatibility
; yeast nutrients
; non-thermal treatment
1. Introduction
Today, the use of non-Saccharomyces yeasts is increasingly widespread due to their high diversity and, therefore, their novel metabolic profiles that differentiate wines [1,2]. These distinctive attributes can facilitate the enhancement of relatively flat grape profiles, thereby yielding wines that are both expressive and well-received by consumers [3]. Furthermore, the potential for co-inoculation with non-Saccharomyces yeasts introduces the possibility of generating interesting synergies at the oenological level [4,5]. Currently, the most studied mixed and sequential inoculations are those of the genera: Lachancea, Metschnikowia, Hanseniaspora and Torulaspora [6,7]. The yeast species Lachancea thermotolerans is capable of producing up to 10-11% alcohol and generating up to 12 g/L of lactic acid through the metabolisation of sugars. Additionally, it increases ethyl lactate and lowers pH, rendering it an appealing yeast for use in quality wine production where wines tend to remain at high pH [8,9]. The yeast species Metschnikowia pulcherrima is capable of producing a high volatile acidity (>0.7g/L) [10]. Furthermore, the yeast produces pulcherriminic acid, which, depending on the amount synthesised, can have an antimicrobial effect [11]. However, it is also a yeast that is used as a starter due to its ability to generate highly distinctive esters, thiols and terpenes, including pear-scented ethyl octanoate and other aromas strongly associated with fermentation, such as 2,6-dimethoxyphenol, which evokes smoked foods [12,13].
The use of ternary fermentations with three or more yeasts is becoming increasingly prevalent in order to fully harness the expressive potential of wines. The aforementioned yeasts serve as illustrative examples of starter yeasts that can enhance the organoleptic characteristics of wines [14]. Non-Saccharomyces yeasts are not particularly competitive and demonstrate limited resistance to SO₂; therefore, the utilisation of non-thermal technologies, such as pulsed electric fields (PEF), pulsed light (PL), and ultra-high pressure homogenisation (UHPH), is a promising approach to reduce or eliminate the initial microbial load in the must, thereby facilitating the successful implantation of the desired yeasts [15,16,17]. In addition to the elimination of microorganisms, the UHPH technology inactivates oxidative enzymes by generating nanofragmentation due to the continuous pressurisation of the fluid to be used at a pressure exceeding 200 MPa, this process occurs without appreciable organoleptic changes [15,18]. The aim of this work was to analyse the biocompatibility of the following yeasts in ternary fermentations (co-inoculations), in all cases finishing the fermentation sequentially with Saccharomyces cerevisiae for the production of high quality wines in warm areas: L. thermotolerans and M. pulcherrima, with a S. cerevisiae strain as control and with two different conditions, trial A, with different volumes of initial inoculum and trial B, with the use of two nutrients, all with must untreated by UHPH and with must treated by UHPH.
2. Materials and Methods
2.1. Ultra-High Pressure Homogenisation of Musts
A pneumatic press was used to crush the Vitis vinifera L. Tempranillo grapes at a temperature of 15 °C. The resulting must was then separated into two parts, one part was treated by UHPH and the other part was used as a control. The tempranillo must was processed using a continuous UHPH system (60 L/h), at an inlet temperature of 23-25 °C and outlet temperature of 13-15 °C, valve temperature of 78-85 °C for only 0.02 s, and flow rate of 60 L/h at 300 ± 3 MPa. The total volume processed by the UHPH was 60 L [19], patented by the Universitat Autònoma de Barcelona and manufactured by Ypsicon Advanced Technologies in Barcelona, Spain (EP2409583B1).
2.2. Yeast Strains and Growing Media
The yeast strains used in this experimental set-up were all isolated at the Food Technology Laboratory of the Escuela Técnica Superior de Ingeniería Agronómica, Alimentaria y de Biosistemas (Universidad Politécnica de Madrid). The starter species used were two non-Saccharomyces yeast in co-inoculation (Lachancea thermotolerans (Lt) strain L3.1 and Metschnikowia pulcherrima (Mp) strain M29). All vinifications were completed with a sequential fermentation by inoculating the Saccharomyces cerevisiae (Sc) yeast strain 7VA on the seventh day after the start of fermentation to completely exhaust the sugars present in the must.
To achieve a high and similar population for the fermentation trials, all strains were grown for 24 h in liquid YEPD medium at a constant temperature of 24°C. The liquid medium for yeast growth was prepared by combining 1% yeast extract, 2% bacteriological peptone and 2% anhydrous D(+)-glucose from Panreac Química of Barcelona, Spain (Laboratorios Conda, Madrid, Spain). The culture medium was autoclaved at 120°C for 15 minutes.
2.3. Micro-Fermentation Trials
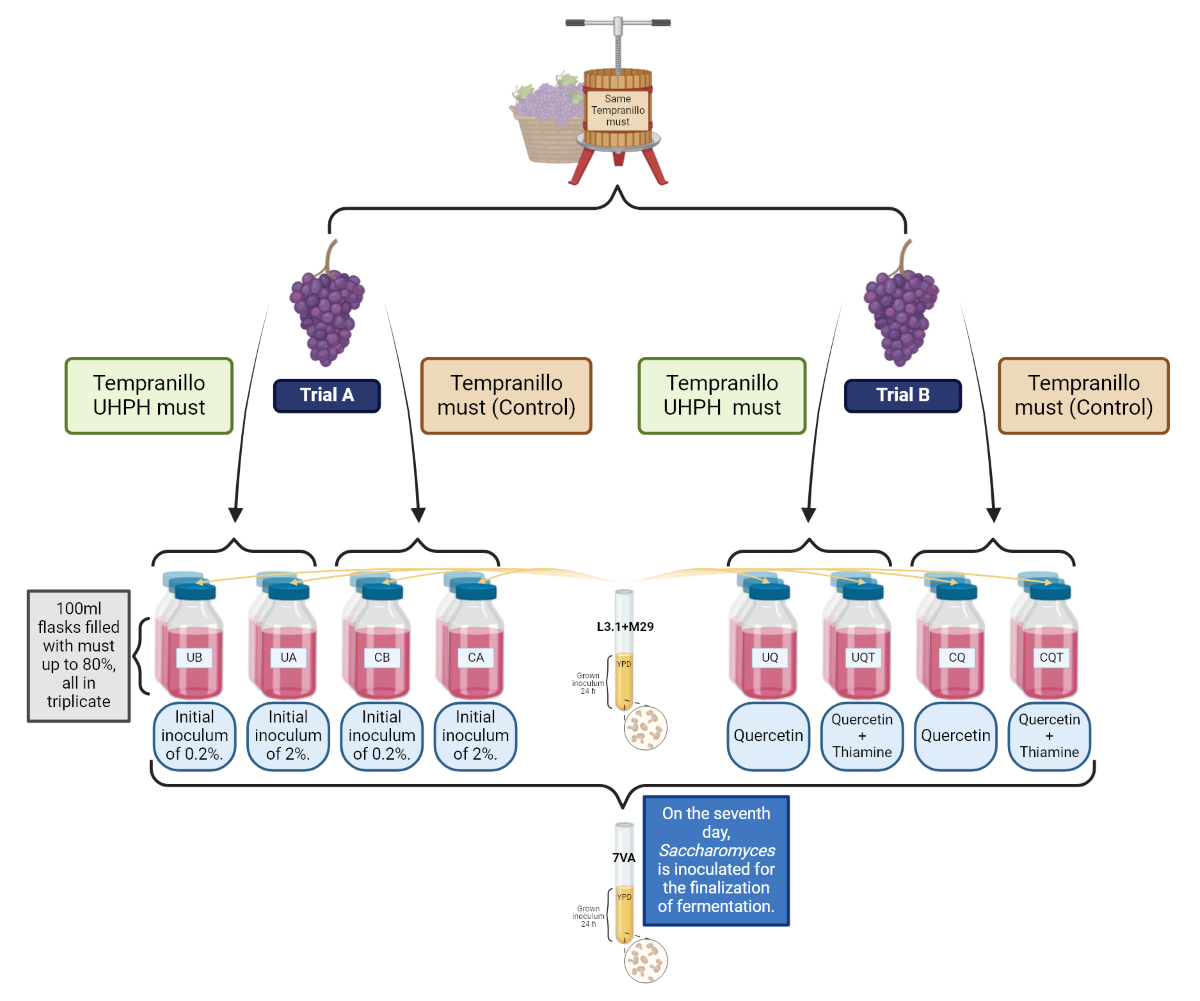
According to Figure 1, the microfermentation tests were performed in ISO flasks of 100 mL capacity, with 80 mL of must Vitis vinifera L. cv. Tempranillo with ~245 g/L of sugars and a pH of ~3.7. In total, two different musts were used, one of which was Tempranillo without any treatment and another must that was treated by UHPH according to the instructions in Section 2.1. Two trials were carried out at the same time with these yeasts and the same must but with different conditions. In the trial A, both non-Saccharomyces strains were inoculated in equal parts (50:50), but the volume of the initial inoculum was 2% (Ca and Ua) and 0.2% (Cb and Ub), to study the influence of UHPH on the implantation and biocompatibility of different inoculum volumes. In the trial B, both non-Saccharomyces strains were inoculated in equal parts (50:50) and at 2% inoculum volume, but adding quercetin (1g/L, “Cq” and “Uq”) to one batch and quercetin + thiamine (1g/L + 0.6mg/L, respectively, “Cqt” and “Uqt”) to the other batch, to evaluate the use of these fermentation activators on the metabolic performance of the co-inoculated non-Saccharomyces species. All fermentations were carried out in triplicate and continued for 25 days until sugar exhaustion (<2g/L). In all the vinifications, sequential fermentation with Sc took place on day 7 by adding 2% of the precultured strain 7VA. After completion of fermentations, five millilitres of each wine triplicate were filtered through 0.45 ηm MCE syringe membranes (Branchia, Dismadel, Madrid, Spain) for analytical characterisation.
2.4. Oenological Parameters
General oenological parameters were determined in grape juices and finished wines with the use of FTIR spectrometry (FOSS, Barcelona, Spain) and with the use of Y25 enzymatic analyser (Byosystems, Barcelona, Spain). The analysis comprised the determination of amino nitrogen, ammonia, total sugars, organic acids (tartaric acid and malic acid) in grape juices, while organic acids (lactic acid (by Y25), acetic acid), residual sugars (glucose and fructose), ethanol and pH were measured in finished wines. Additionally, pH values were measured with a GLP 21 Crison Instruments (Hach Lange Spain, S.L.U., Madrid, Spain). Five mL Samples were filtered with 0.45 ηm membrane and the CO2 trapped was removed with agitation.
2.5. Aroma Volatile Compounds
Volatile compounds were analysed in accordance with a previously described method [19]. The identification of volatile compounds was carried out with an Agilent Technologies™ 6850 chromatograph (Palo Alto, CA, USA) with a column DB-624 (60 m × 250 μm × 1.4 μm). The GC-FID parameters were as follows: injector’s temperature 250 °C; detector’s temperature 300 °C; and hydrogen was the carrying gas with a flow of 2.2 L/min and split ratio of 1:10. Finally, the temperature increased from 40 °C for 5 min to 250 °C with a gradient of 10 °C/min and was maintained for 5 min. The identification and the quantification of volatile organic compounds were performed on 1 mL of previously filtered samples and the use of individual calibration curves for the following: 2-phenylethyl acetate, 2-phenylethanol, ethyl acetate, isobutyl acetate, ethyl butyrate, isoamyl acetate, acetaldehyde, methanol, 1-propanol, diacetyl, 1-butanol, 2-butanol, isobutanol, acetoin, 2-methyl-1-butanol, 3-methyl-1-butanol, ethyl lactate, 2,3-butanediol, and 1-hexanol. Lastly, in accordance with the procedure previously described, 100 μL of 4-methyl-2-pentanol (500 mg/L) was used as internal standard.
2.6. Colour Parameters
The absorbance at 280, 420, 520 and 620 nm was determined at the end of fermentation using an Agilent 8453 spectrophotometer (Agilent Technologies S.L., Madrid, Spain) and a 1 mm pitch quartz cuvette, to comply with the standard method, the result is multiplied by 10 to emulate the optical passage through a 1 cm cuvette and parameters such as TPI, colour intensity and tonality were determined.
2.7. Statistics
ANOVA and the least significant difference (LSD) test were used to analyse differences and determine the means and standard deviations. PCA and all calculations were performed with the program PC Statgraphics v.5, (Graphics Software Systems, Rockville, MD, USA). The significance was set to p < 0.05.
3. Results and Discussion
Two studies have been carried out to look at the mixed implantation of non-Saccharomyces yeasts, on the one hand, with the use of different volumes of starter inoculum, and on the other hand, with the use of different nutrients. All this using the same must, but one part of the must had been treated with UHPH and the other not, in order to verify the effect of this sanitisation technology on the metabolic development of the yeasts used as starters. Non-thermal technologies such as the one used in these trials, in addition to pulsed light (PL), pulsed electric fields (PEFs) among others, are increasingly used in the field of oenology to partially or totally eliminate wild microflora from musts, without causing organoleptic changes, and thus be able to inoculate the desired yeasts with a higher success rate [16,17,20].
3.1. Evolution of the Inoculated Population
The yeast population and its evolution during fermentation (Figure 2) were monitored until day 6 through the CHROMagar™ Candida culture medium, just before the inoculation of S. cerevisiae on day 7. In Trial A (Figure 2A), the initial inocula exhibited a 10-fold difference, with one (6-log CFU/mL, 2% initial inoculum) being higher than the other (5-log CFU/mL, 0.2% initial inoculum), which was anticipated. On the third day, both yeasts demonstrated reduced growth in the UHPH-treated musts. In both cases, the UHPH-treated musts exhibited no indigenous yeasts at the outset or throughout fermentation. By the sixth day, the yeast populations had decreased overall by 1-log CFU/mL. The prevalence of L. thermotolerans was observed to persist for a longer period than that of M. pulcherrima. From day 3 onwards, the viability of the latter declined at a more rapid rate, which is likely attributable to its poor ethanol tolerance [21]. In general, the exponential growth of M. pulcherrima was observed to be lower, which is likely attributed to its relatively low competitiveness [22]. Furthermore, the use of UHPH-treated musts resulted in a reduction in yeast population growth, which is likely attributable to a decline in nutrient bioavailability and subsequent utilisation [23].
On the other hand, in trial B (Figure 2B) the yeast growth was similar, reaching more than 7-log in L. thermotolerans on the third day and almost 7-log on average in M. pulcherrima. Furthermore, it was observed that the combination of quercetin and thiamine resulted in an average increase of 0.5 log in population size compared to that observed in the presence of quercetin alone. For both yeast strains, a decline in population was observed from the sixth day of fermentation, similar to that observed in Trial A, only indigenous yeasts were present in the non-UHPH-treated controls. The combination of quercetin + thiamine appears to exert a beneficial effect on yeast growth in both control and UHPH must, potentially due to its capacity to safeguard yeast cells from oxidative stress [24,25]. The antimicrobial efficacy of UHPH was unmistakably demonstrated by the absence of indigenous yeast growth [15,19].
3.2. Oenological Parameters
The initial must had a sugar concentration of approximately 245 g/L. In Trial A, it was observed that the fermentation process was slower in the musts with UHPH, resulting in the accumulation of residual sugars at the end of fermentation (Table 1). This phenomenon can be attributed to the reduced availability of certain nitrogenous compounds that may be nano-encapsulated and less accessible [19]. Nevertheless, the greatest quantity of alcohol was achieved with the UHPH musts. With regard to total acidity, the highest levels were observed in the fermentation with control musts (without UHPH) and at high inoculum doses, with a value of 8.27 ± 0.26 g/L in tartaric acid equivalents. Volatile acidity was found to be below 0.5 g/L in all cases.
In contrast, the fermentation process in trial B (Table 2) was more homogeneous, although residual sugars below 0.5 g/L were present in the fermentations with UHPH must. In all cases, volatile acidity was below 0.5 g/L.
It was observed that fermentations with higher total acidity, both in trial A and especially in trial B, exhibited a reduction in ethanol production at the end of fermentation, reaching a level of approximately 1% v/v less than expected (~14.5% v/v). This phenomenon can be attributed to the high lactic acid production of L. thermotolerans, which utilizes a portion of the sugars present in the must to generate this elevated acidity [26]. Furthermore, although not the subject of investigation in these trials, numerous non-Saccharomyces yeasts have the capacity to produce pyruvic acid, glycerol and other compounds from sugars through secondary metabolic processes [27]. It should be noted that certain Mp strains have the capacity to reduce the final ethanol content by up to 1.6% v/v when co-inoculated with a Sc. [28]. In accordance with the characteristics of a mixed fermentation, the volatile acidity was found to be relatively low, with a concentration below 0.5 g/L in both trials, a finding that aligns with the observations of other researchers in this field [29].
3.3. Lactic Acidity and pH
It has been demonstrated that Lt is capable of producing lactic acid between days 3 and 6 of fermentation. However, this process is contingent upon a number of variables, including temperature, nutrients, and the competitive dynamics between yeast strains. Consequently, these factors must be considered in any analysis [5,30]. To check the effect of bioacidification with lactic acid on pH, we compared these results in trial A (Figure 3a) and saw a high correlation (> -0.9) where the Control sample with high initial inoculum (2%) produced a higher amount of lactic acid (4.30 ± 0.09 g/L) and, therefore, a lower pH, 3.20 ± 0.01. On the other hand, in trial B (Figure 3b) the production of lactic acid was more pronounced, especially in the must sample with UHPH and the nutrient quercetin, producing 6.18 ± 0.34 g/L lactic acid and a pH of 3.10 ± 0.03.
With these results it was observed that nutrients can generate a higher production of lactic acid compared to the average of Ca and Ua samples, producing 1.5 g/L more lactic acid in trial B Additionally, the observed synergy between Lt and Mp yeasts was found to be highly effective, without the occurrence of incompatibilities, as previously documented in other studies [5,31,32].
3.4. Colour Assessment
Their absorbances at different wavelengths were evaluated spectrophotometrically to determine, after fermentation, their colour intensity, hue and total polyphenol index (TPI). In trial A (Table 3), a considerably higher colour intensity than the rest was observed in the Control sample with the high initial inoculum (Ca), probably because the pH is lower and the anthocyanins are predominantly found in their protonated form and their colour is more reddish [33]. It is also noteworthy that both the hue and the IPT of the control samples were significantly lower than in the wines obtained from UHPH-treated musts.
The same evaluation was conducted with Trial B, and it was observed that the controls with both quercetin and quercetin + thiamine exhibited a higher colour intensity at the conclusion of fermentation than the UHPH. Furthermore, as observed in Trial A, the hue and IPT were significantly lower in the control wines compared to the UHPH.
Higher IPT in wines from UHPH musts means a higher concentration of these compounds and that this technology protects from oxidation [15]. It was also calculated that the samples that had produced more lactic acid in both trials A and B (Cq, Uq, Cqt, Uqt, Uqt and Ca) generated about 9% less yellow tints (420nm) [23,32].
3.5. Aromatic Profile
The volatile compounds that are produced during the fermentation process of wine play an important role in determining the wine's sensory profile, the inoculated yeasts are a significant factor in this process, influencing the types and quantities of aromas that are synthesised [34,35]. The volatile compounds of fermentation origin were subjected to analysis in both trials. In Trial A (Figure 4A), it was observed that fermentations with a high initial inoculum (Ca and Ua) produced lower amounts of higher alcohols and total esters. Furthermore, carbonyl compounds were generally low. Notably, for esters, excluding ethyl acetate, it was observed that UHPH wines contain almost twice as much ethyl lactate. Conversely, trial B revealed that the simultaneous addition of quercetin and thiamine resulted in the production of elevated levels of higher alcohols in the UHPH wines, while the Control wine exhibited enhanced formation of total esters (not including ethyl acetate) and carbonyl compounds.
In all cases, the higher alcohols were present in concentrations above 300 mg/L, which could impart subtle spicy and fusel oil aromas [36], it should be noted that the alcohol that exerted the most pronounced effect was 3-methyl-1-butanol (isoamyl alcohol), which produces complex, smoky, and roasted aromas when its concentration exceeds 30 mg/L [37,38]. The second most abundant alcohol was 1-propanol, with a mean of 67 mg/L. However, this was considerably below the perceived threshold of 830 mg/L [39]. This discrepancy can be attributed to the limited scale of the test, as at industrial levels, this alcohol does not typically emerge as a prominent factor [9,29]. The concentration of carbonyl compounds, which impart notes of candy and butter, was below the detection threshold for diacetyl (4–12 mg/L) and acetoin (150 mg/L) in both tests [40,41]. It can be stated that esters are aromas that are related to the amount of nitrogen (NFA) that the yeast is able to assimilate [39]. Ethyl acetate is a compound that, at concentrations below 60 mg/L, exhibits pleasant fruity aromas. In all fermentations, its concentration remained below this threshold, with an average of ~48 mg/L. Ethyl lactate, whose production is a direct consequence of lactic acid production by Lt [42], also remained below the perception threshold of 150 mg/L, with an average concentration of ~43 mg/L [42,43]. Both isoamyl acetate with sweet banana and fruit aroma with a hint of ripe essence and isobutyl acetate with tropical fruit aroma especially banana were detected in several samples but especially in the Control sample with quercetin + thiamine with ~4mg/L for both volatiles, which could be detectable as their perception threshold is below 2 mg/L for both volatiles [39,41]. It should be noted that, although not depicted in the figures, acetaldehyde was also analysed as a volatile compound, yielding values approximately 20 times higher in the control wines (approximately 300 mg/L). These elevated values may be attributed to a number of factors, including the composition of the medium, the nature of the insoluble materials employed to clarify the musts, the ageing/breeding procedure, the SO₂ content, the aeration, and most notably, the indigenous yeast population, which differs significantly between the control must and the UHPH must. It is highly probable that a population of S. cerevisiae beticus was present [44]. Additionally, the presence of non-Saccharomyces species, including C. stellata, Z. bailii, and S. pombe [45] as well as bacteria such as Lactobacillus plantarum, may contribute to the observed fermentation profile [46].
4. Conclusions
The use of UHPH eliminates the microflora naturally present in the grape must, facilitating the implantation of yeast ferments and not allowing spontaneous fermentations to take place. Moreover, the use of non-Saccharomyces yeasts can be strongly favoured in both mono- and co-inoculation. It should also be noted that both the nutrients (quercetin + thiamine) and the non-Saccharomyces yeasts studied produced some very interesting synergies that open up a very powerful field of study to be explored further.
Author Contributions
C.V. and C.E.; experimental work, methodology; C.V. Writing—original draft preparation review and editing; A.M.; project administration, data curation, conceptualisation and experimental design; J.M.D.F., I.L., C.L., C.G. and J.M.H.; critical reading. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Ministerio de Ciencia e Innovación (MICINN). Spain [grant number PID2021-124250OB-I00] and Programa Estatal de Fomento de la Investigación Científico-Técnica y su Transferencia del Ministerio de Ciencia e Innovación through Red de Investigación MALTA CONSOLIDER TEAM [grant number RED2022-134388-T].
References
- Civa V, Chinnici F, Picariello G, Tarabusi E, Bosaro M, Mannazzu I, et al. Non-Saccharomyces yeast derivatives: characterization of novel potential bio-adjuvants for the winemaking process. Curr Res Food Sci [Internet]. 2024 May 22 [cited 2024 May 27];100774. Available online: https://linkinghub.elsevier.com/retrieve/pii/S266592712400100X.
- Yang B, Liu S, Zang H, Dai Y, Zhang S, Lin X, et al. Flavor profile and quality of strawberry wine are improved through sequential fermentation with indigenous non-Saccharomyces yeasts and Saccharomyces cerevisiae. Food Biosci [Internet]. 2024 Jun 1 [cited 2024 May 27];59:104021. Available online: https://linkinghub.elsevier.com/retrieve/pii/S2212429224004516.
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl Microbiol Biotechnol [Internet]. 2016 Dec 27;100(23):9861–74. [CrossRef]
- Jolly NP, Augustyn OPH, Pretorius IS. The Effect of Non-Saccharomyces Yeasts on Fermentation and Wine Quality. South African J Enol Vitic [Internet]. 2003 May [cited 2022 May 28];24(2):55–62. Available online: https://www.journals.ac.za/index.php/sajev/article/view/2638.
- Vaquero C, Loira I, Heras JM, Carrau F, González C, Morata A. Biocompatibility in Ternary Fermentations With Lachancea thermotolerans, Other Non-Saccharomyces and Saccharomyces cerevisiae to Control pH and Improve the Sensory Profile of Wines From Warm Areas. Front Microbiol [Internet]. 2021 Apr 29 [cited 2021 Apr 29];12. Available online: https://www.frontiersin.org/articles/10.3389/fmicb.2021.656262/full.
- Sadineni V, Kondapalli N, Obulam VSR. Effect of co-fermentation with Saccharomyces cerevisiae and Torulaspora delbrueckii or Metschnikowia pulcherrima on the aroma and sensory properties of mango wine. Ann Microbiol [Internet]. 2012 Dec 22 [cited 2022 Jun 18];62(4):1353–6. Available online: https://annalsmicrobiology.biomedcentral.com/articles/10.1007/s13213-011-0383-6.
- Vaquero C, Escott C, Heras JM, Carrau F, Morata A. Co-inoculations of Lachancea thermotolerans with different Hanseniaspora spp.: Acidification, aroma, biocompatibility, and effects of nutrients in wine. Food Res Int [Internet]. 2022 Nov 1 [cited 2022 Sep 21];161:111891. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0963996922009498.
- Hranilovic A, Gambetta JM, Schmidtke L, Boss PK, Grbin PR, Masneuf-Pomarede I, et al. Oenological traits of Lachancea thermotolerans show signs of domestication and allopatric differentiation. Sci Rep [Internet]. 2018 Oct 4 [cited 2023 Jan 20];8(1):14812. Available online: https://www.nature.com/articles/s41598-018-33105-7.
- Vaquero C, Loira I, Bañuelos MA, Heras JM, Cuerda R, Morata A. Industrial Performance of Several Lachancea thermotolerans Strains for pH Control in White Wines from Warm Areas. Microorganisms [Internet]. 2020 Jun 1 [cited 2020 Jun 3];8(6):830. Available online: https://www.mdpi.com/2076-2607/8/6/830.
- Zhang M, Zhong T, Heygi F, Wang Z, Du M. Effects of inoculation protocols on aroma profiles and quality of plum wine in mixed culture fermentation of Metschnikowia pulcherrima with Saccharomyces cerevisiae. LWT [Internet]. 2022 May 1 [cited 2024 Aug 20];161:113338. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0023643822002730.
- Kántor A, Hutková J, Petrová J, Hleba L, Kačániová M. Antimicrobial activity of pulcherrimin pigment produced by Metschnikowia pulcherrima against various yeast species. J Microbiol Biotechnol food Sci [Internet]. 2015 Dec 1 [cited 2022 May 30];5(3):282–5. Available online: https://www.researchgate.net/publication/285310898.
- González-Royo E, Pascual O, Kontoudakis N, Esteruelas M, Esteve-Zarzoso B, Mas A, et al. Oenological consequences of sequential inoculation with non-Saccharomyces yeasts (Torulaspora delbrueckii or Metschnikowia pulcherrima) and Saccharomyces cerevisiae in base wine for sparkling wine production. Eur Food Res Technol [Internet]. 2015 May 7;240(5):999–1012. Available online: http://link.springer.com/10.1007/s00217-014-2404-8.
- Sadoudi M, Tourdot-Maréchal R, Rousseaux S, Steyer D, Gallardo-Chacón JJ, Ballester J, et al. Yeast-yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol [Internet]. 2012;32(2):243–53. [CrossRef]
- ŽENIŠOVÁ K, CABICAROVÁ T, SIDARI R, KOLEK E, PANGALLO D, SZEMES T, et al. Effects of co-fermentation with Lachancea thermotolerans or Metschnikowia pulcherrima on concentration of aroma compounds in Pinot Blanc wine. Food Nutr Res [Internet]. 2021 [cited 2024 May 30];60(1):87. Available online: https://openurl.ebsco.com/EPDB%3Agcd%3A2%3A21148786/detailv2?sid=ebsco%3Aplink%3Ascholar&id=ebsco%3Agcd%3A149520655&crl=c.
- Vaquero C, Escott C, Loira I, Guamis B, del Fresno JM, Quevedo JM, et al. Cabernet Sauvignon Red Must Processing by UHPH to Produce Wine Without SO2: the Colloidal Structure, Microbial and Oxidation Control, Colour Protection and Sensory Quality of the Wine. Food Bioprocess Technol [Internet]. 2022 Mar 4 [cited 2022 Nov 4];15(3):620–34. Available online: https://link.springer.com/article/10.1007/s11947-022-02766-8.
- Escott C, Vaquero C, del Fresno JM, Bañuelos MA, Loira I, Han S, et al. Pulsed Light Effect in Red Grape Quality and Fermentation. Food Bioprocess Technol [Internet]. 2017 Aug 5 [cited 2021 Sep 6];10(8):1540–7. Available online: https://link.springer.com/article/10.1007/s11947-017-1921-4.
- Vaquero C, Loira I, Raso J, Álvarez I, Delso C, Morata A. Pulsed Electric Fields to Improve the Use of Non-Saccharomyces Starters in Red Wines. Foods [Internet]. 2021 Jun 25 [cited 2022 Jan 11];10(7):1472. Available online: https://www.mdpi.com/2304-8158/10/7/1472/htm.
- Bañuelos MA, Loira I, Escott C, Del Fresno JM, Morata A, Sanz PD, et al. Grape processing by high hydrostatic pressure: Effect on use of non-Saccharomyces in must fermentation. Food Bioprocess Technol [Internet]. 2016;9(10):1769–78. [CrossRef]
- Bañuelos MA, Loira I, Guamis B, Escott C, Del Fresno JM, Codina-Torrella I, et al. White wine processing by UHPH without SO2. Elimination of microbial populations and effect in oxidative enzymes, colloidal stability and sensory quality. Food Chem. 2020 Dec 1;332:127417.
- Morata A, Loira I, Bañuelos MA, Puig-Pujol A, Guamis B, González C, et al. Use of Ultra High Pressure Homogenization to sterilize grape must. Roca P, editor. BIO Web Conf [Internet]. 2019 Oct 23;15:02035. Available online: https://www.bio-conferences.org/10.1051/bioconf/20191502035.
- Sadineni V, Kondapalli N, Obulam VSR. Effect of co-fermentation with Saccharomyces cerevisiae and Torulaspora delbrueckii or Metschnikowia pulcherrima on the aroma and sensory properties of mango wine. Ann Microbiol [Internet]. 2012 Dec 22 [cited 2024 Jun 12];62(4):1353–60. Available online: https://link.springer.com/articles/10.1007/s13213-011-0383-6.
- Puškaš VS, Miljić UD, Djuran JJ, Vučurović VM. The aptitude of commercial yeast strains for lowering the ethanol content of wine. Food Sci Nutr [Internet]. 2020 Mar 1 [cited 2024 Aug 16];8(3):1489–98. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/fsn3.1433.
- Escott C, Vaquero C, del Fresno JM, Topo A, Comuzzo P, Gonzalez C, et al. Effect of processing Verdejo grape must by UHPH using non- Saccharomyces yeasts in the absence of SO2. Sustain Food Technol [Internet]. 2024 Mar 21 [cited 2024 Aug 17];2(2):437–46. Available online: https://pubs.rsc.org/en/content/articlehtml/2024/fb/d3fb00226h.
- Labuschagne P, Divol B. Thiamine: a key nutrient for yeasts during wine alcoholic fermentation. Appl Microbiol Biotechnol [Internet]. 2021 Feb 6 [cited 2023 Jan 20];105(3):953–73. Available online: https://link.springer.com/article/10.1007/s00253-020-11080-2.
- Bayliak MM, Burdylyuk NI, Lushchak VI. Quercetin increases stress resistance in the yeast Saccharomyces cerevisiae not only as an antioxidant. Ann Microbiol [Internet]. 2016 Jun 14 [cited 2024 Aug 17];66(2):569–76. Available online: https://link.springer.com/articles/10.1007/s13213-015-1136-8.
- Ristic R, Hranilovic A, Li S, Longo R, Pham D-T, Qesja B, et al. Alcohol: Integrated strategies to moderate the alcohol content of wines. Wine Vitic J [Internet]. 2016;31(5):33–8. Available online: https://search.informit.org/doi/10.3316/informit.511380340406896.
- Ivit NN, Longo R, Kemp B. The Effect of Non-Saccharomyces and Saccharomyces Non-Cerevisiae Yeasts on Ethanol and Glycerol Levels in Wine. Fermentation [Internet]. 2020 Jul 30 [cited 2022 Jun 3];6(3):77. Available online: https://www.mdpi.com/2311-5637/6/3/77/htm.
- Gonzalez R, Quirós M, Morales P. Yeast respiration of sugars by non-Saccharomyces yeast species: A promising and barely explored approach to lowering alcohol content of wines. Trends Food Sci Technol. 2013;29(1):55–61.
- Gobbi M, Comitini F, Domizio P, Romani C, Lencioni L, Mannazzu I, et al. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol [Internet]. 2013;33(2):271–81. [CrossRef]
- Binati RL, Innocente G, Gatto V, Celebrin A, Polo M, Felis GE, et al. Exploring the diversity of a collection of native non-Saccharomyces yeasts to develop co-starter cultures for winemaking. Food Res Int [Internet]. 2019;122:432–42. [CrossRef]
- Vaquero C, Escott C, Heras JM, Carrau F, Morata A. Co-inoculations of Lachancea thermotolerans with different Hanseniaspora spp.: Acidification, aroma, biocompatibility, and effects of nutrients in wine. Food Res Int [Internet]. 2022 Nov 4 [cited 2022 Mar 1];161:111891. Available online: https://link.springer.com/article/10.1007/s11947-022-02766-8.
- Escott C, Vaquero C, Loira I, López C, González C, Morata A. Synergetic Effect of Metschnikowia pulcherrima and Lachancea thermotolerans in Acidification and Aroma Compounds in Airén Wines. Foods [Internet]. 2022 Nov 21 [cited 2022 Nov 22];11(22):3734. Available online: https://www.mdpi.com/2304-8158/11/22/3734/htm.
- Moreira, PF, Giestas L, Yihwa C, Vautier-Giongo C, Quina FH, Maçanita AL, et al. Ground- and Excited-State Proton Transfer in Anthocyanins: From Weak Acids to Superphotoacids. J Phys Chem A [Internet]. 2003 May 1 [cited 2024 Aug 22];107(21):4203–10. Available online: https://pubs.acs.org/doi/abs/10.1021/jp027260i.
- Jiang X, Lu Y, Liu SQ. Effects of different yeasts on physicochemical and oenological properties of red dragon fruit wine fermented with Saccharomyces cerevisiae, Torulaspora delbrueckii and Lachancea thermotolerans. Microorganisms. 2020;8(3):1–14.
- Lee SB, Banda C, Park HD. Effect of inoculation strategy of non-Saccharomyces yeasts on fermentation characteristics and volatile higher alcohols and esters in Campbell Early wines. Aust J Grape Wine Res. 2019;25(4):384–95.
- Saberi S, Cliff MA, van Vuuren HJJ. Impact of mixed S. cerevisiae strains on the production of volatiles and estimated sensory profiles of Chardonnay wines. Food Res Int [Internet]. 2012;48(2):725–35. [CrossRef]
- Antón-Díaz MJ, Suárez Valles B, Mangas-Alonso JJ, Fernández-García O, Picinelli-Lobo A. Impact of different techniques involving contact with lees on the volatile composition of cider. Food Chem [Internet]. 2016 Jan;190:1116–22. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0308814615008961.
- González Álvarez M, González-Barreiro C, Cancho-Grande B, Simal-Gándara J. Relationships between Godello white wine sensory properties and its aromatic fingerprinting obtained by GC–MS. Food Chem [Internet]. 2011 Dec;129(3):890–8. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0308814611007291.
- Carrau FM, Medina K, Farina L, Boido E, Henschke PA, Dellacassa E. Production of fermentation aroma compounds by Saccharomyces cerevisiae wine yeasts: effects of yeast assimilable nitrogen on two model strains. FEMS Yeast Res [Internet]. 2008 Nov;8(7):1196–207. Available online: https://academic.oup.com/femsyr/article-lookup/doi/10.1111/j.1567-1364.2008.00412.x.
- Rogerson FSS, Castro H, Fortunato N, Azevedo Z, Macedo A, De Freitas VAP. Chemicals with Sweet Aroma Descriptors Found in Portuguese Wines from the Douro Region: 2,6,6-Trimethylcyclohex-2-ene-1,4-dione and Diacetyl. J Agric Food Chem [Internet]. 2001 Jan 1;49(1):263–9. Available online: https://pubs.acs.org/doi/10.1021/jf000948c.
- Peinado RA, Moreno J, Medina M, Mauricio JC. Changes in volatile compounds and aromatic series in sherry wine with high gluconic acid levels subjected to aging by submerged flor yeast cultures. Biotechnol Lett [Internet]. 2004 May;26(9):757–62. Available online: http://link.springer.com/10.1023/B:BILE.0000024102.58987.de.
- Morata A, Bañuelos MA, Vaquero C, Loira I, Cuerda R, Palomero F, et al. Lachancea thermotolerans as a tool to improve pH in red wines from warm regions. Eur Food Res Technol [Internet]. 2019 Apr 16;245(4):885–94. Available online: http://link.springer.com/10.1007/s00217-019-03229-9.
- Perestrelo R, Silva C, Câmara JS. Madeira Wine Volatile Profile. A Platform to Establish Madeira Wine Aroma Descriptors. Molecules [Internet]. 2019 Aug 21;24(17):3028. Available online: https://www.mdpi.com/1420-3049/24/17/3028.
- Zea L, Serratosa MP, Mérida J, Moyano L. Acetaldehyde as Key Compound for the Authenticity of Sherry Wines: A Study Covering 5 Decades. Compr Rev Food Sci Food Saf [Internet]. 2015 Nov 1 [cited 2024 Jun 17];14(6):681–93. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/1541-4337.12159.
- Li E, Mira de Orduña R. Acetaldehyde kinetics of enological yeast during alcoholic fermentation in grape must. J Ind Microbiol Biotechnol [Internet]. 2017 Feb 1 [cited 2024 Jun 17];44(2):229–36. [CrossRef]
- Wang S, Li S, Zhao H, Gu P, Chen Y, Zhang B, et al. Acetaldehyde released by Lactobacillus plantarum enhances accumulation of pyranoanthocyanins in wine during malolactic fermentation. Food Res Int [Internet]. 2018 Jun 1 [cited 2024 Jun 17];108:254–63. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0963996918302084.
Figure 1.
Schematic representation of the experimental process for the evaluation of co-inoculation of non-Saccharomyces under different inoculation volumes and nutrient addition, and the use of UHPH-treated or untreated must.
Figure 1.
Schematic representation of the experimental process for the evaluation of co-inoculation of non-Saccharomyces under different inoculation volumes and nutrient addition, and the use of UHPH-treated or untreated must.
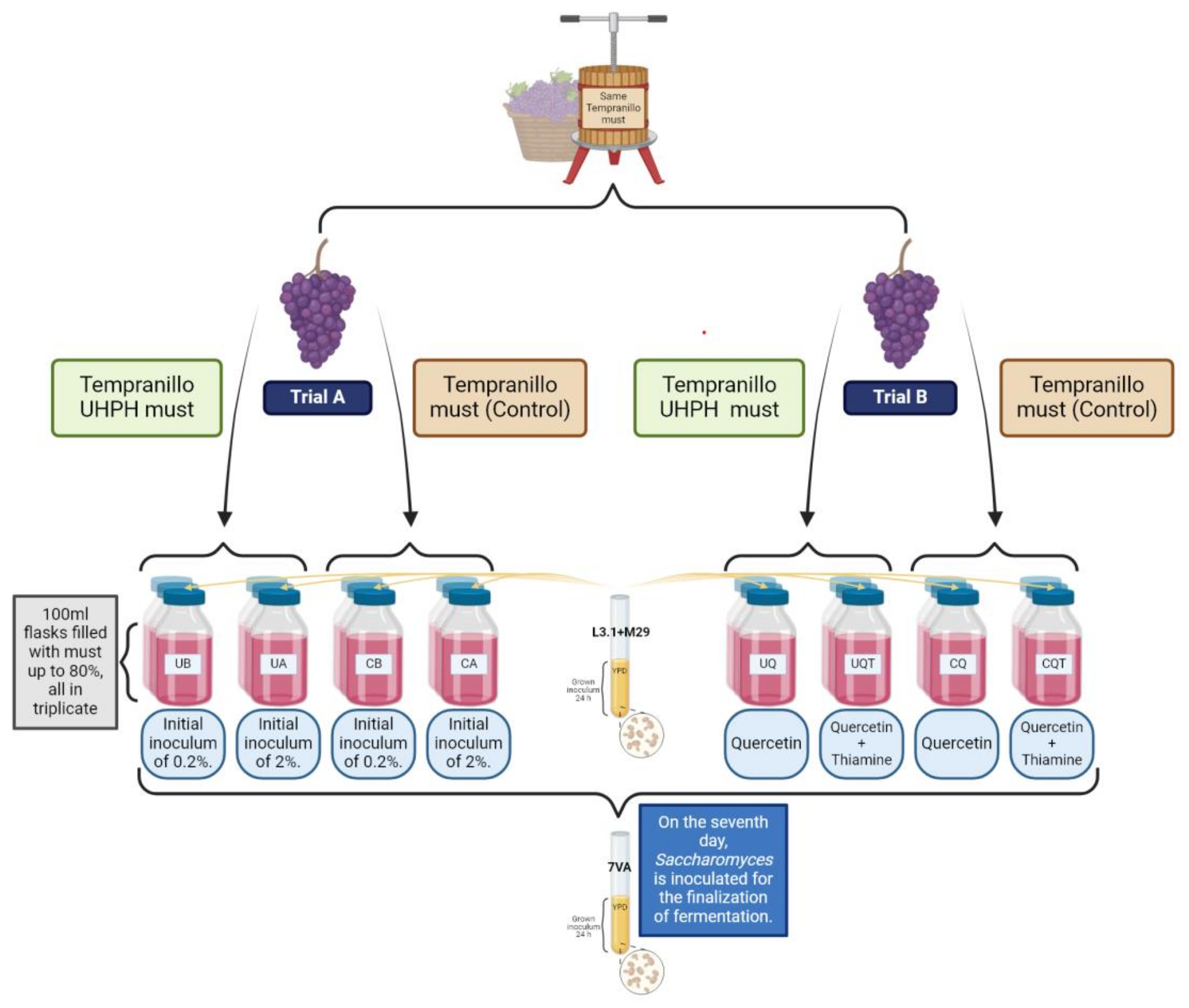
Figure 2.
(A). Evolution of yeast populations in the trial A with two different inoculum volumes (a = 2% inoculation volume, b = 0.2% inoculation volume, C = Control must and U = UHPH treated must). (B). Evolution of the yeast populations in the trial B with the addition of quercetin and quercetin + thiamine. Values are means ± SD (n = 3). A different letter for the same day means significant differences (p < 0.05).
Figure 2.
(A). Evolution of yeast populations in the trial A with two different inoculum volumes (a = 2% inoculation volume, b = 0.2% inoculation volume, C = Control must and U = UHPH treated must). (B). Evolution of the yeast populations in the trial B with the addition of quercetin and quercetin + thiamine. Values are means ± SD (n = 3). A different letter for the same day means significant differences (p < 0.05).
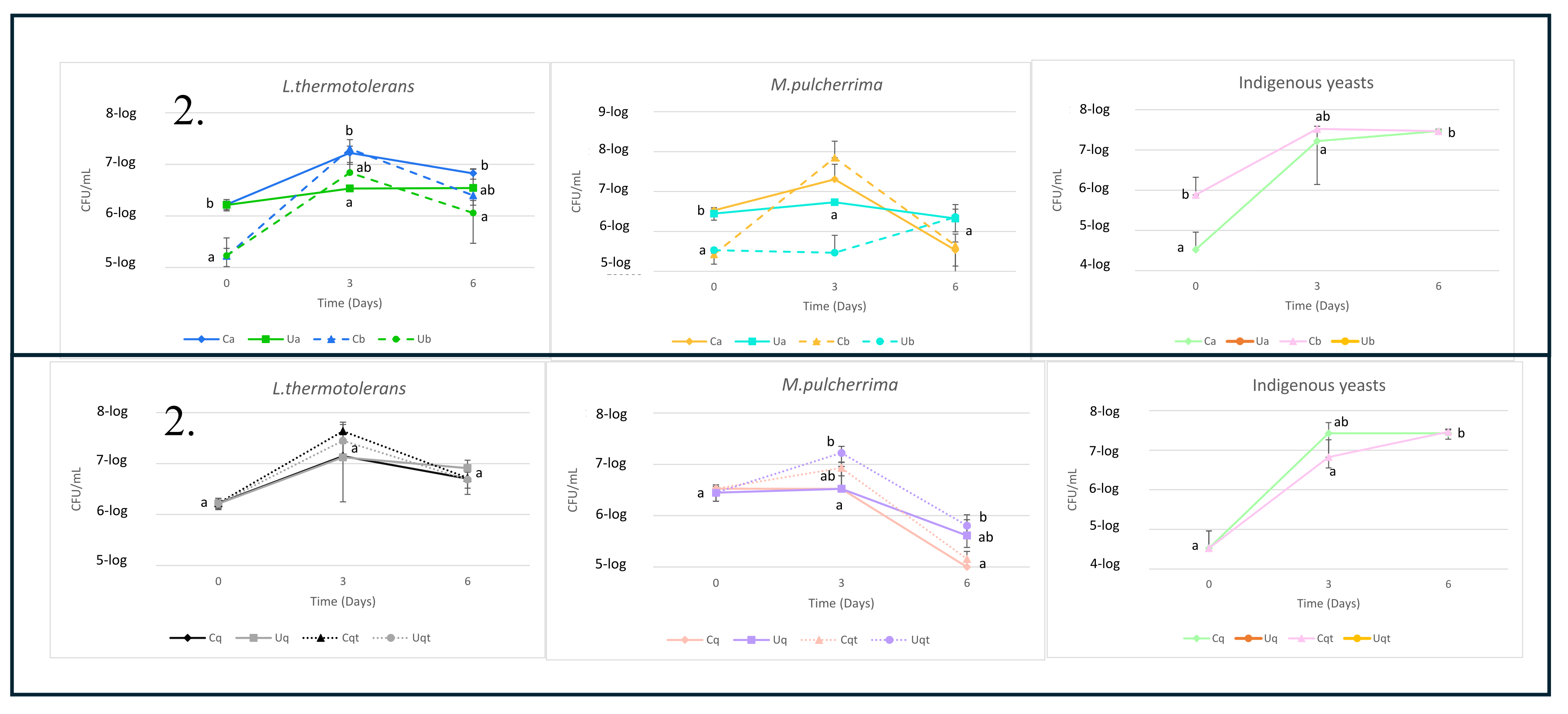
Figure 3.
(A). Lactic acid production (solid line) versus pH decrease (dashed line) in the trial A. (B). Lactic acid production (solid line) versus pH decrease (dashed line) in the trial B. a = 2% inoculation volume, b = 0.2% inoculation volume, C = Control must and U = UHPH treated must.
Figure 3.
(A). Lactic acid production (solid line) versus pH decrease (dashed line) in the trial A. (B). Lactic acid production (solid line) versus pH decrease (dashed line) in the trial B. a = 2% inoculation volume, b = 0.2% inoculation volume, C = Control must and U = UHPH treated must.
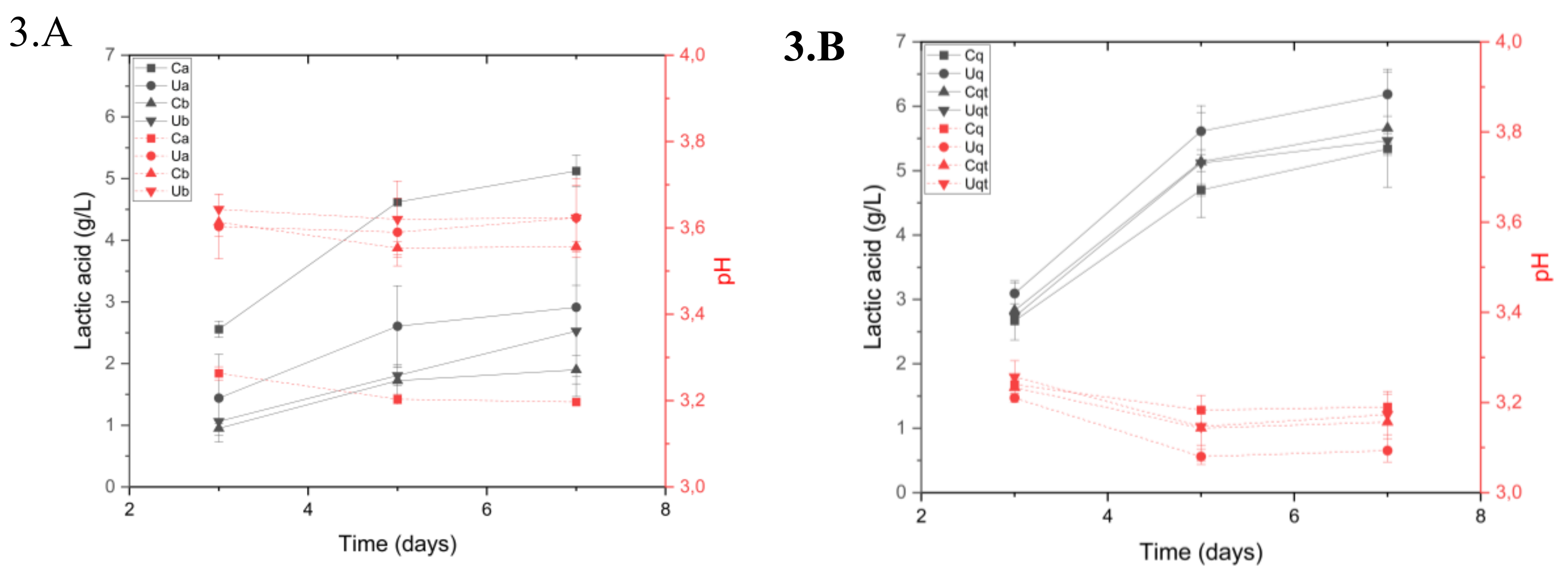
Figure 4.
(A) Aromatic compounds from the trial A. (B) Aromatic compounds from the trial B. Values are means ± SD (n = 3). A different letter for the same compound group means significant differences (p < 0.05).
Figure 4.
(A) Aromatic compounds from the trial A. (B) Aromatic compounds from the trial B. Values are means ± SD (n = 3). A different letter for the same compound group means significant differences (p < 0.05).
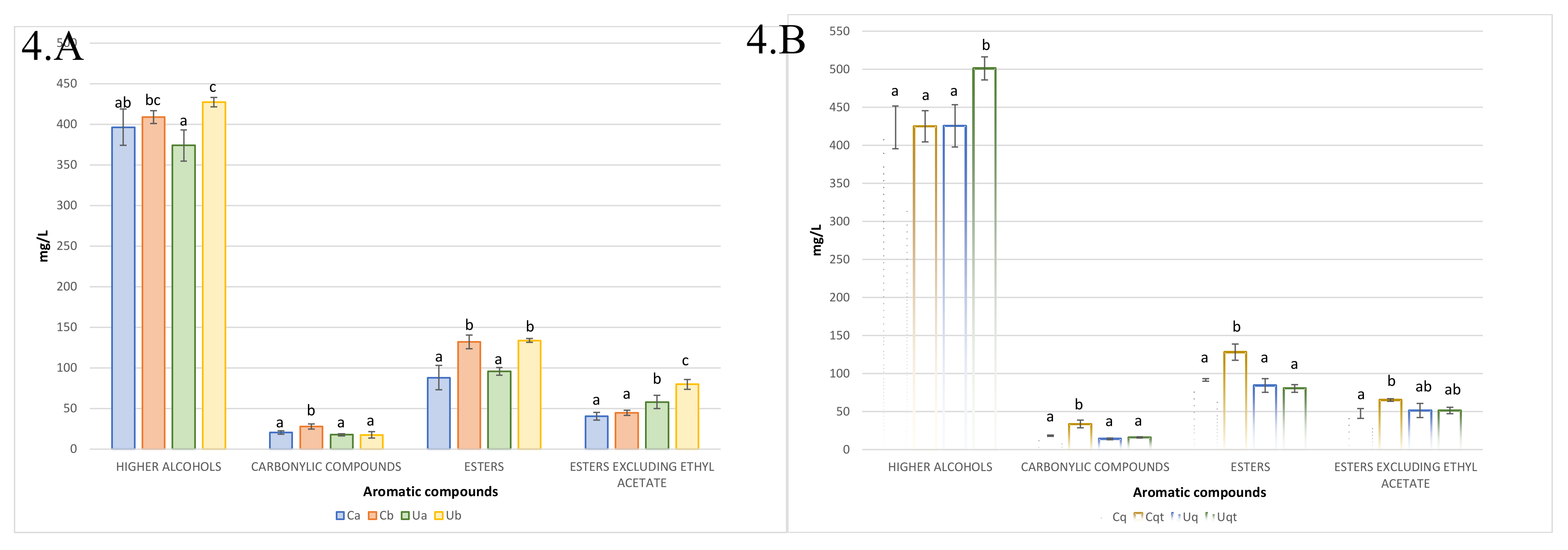

Table 1.
Oenological parameters of the first trial with different initial inoculum volumes. Different letters in each column indicate statistical differences (p < 0.05) between treatments in each fermentation scenario.
Table 1.
Oenological parameters of the first trial with different initial inoculum volumes. Different letters in each column indicate statistical differences (p < 0.05) between treatments in each fermentation scenario.
| Residual sugars (g/L) | Ethanol (% v/v) | Total acidity1 (g/L) | Volatile acidity2 (g/L) | |||||
|---|---|---|---|---|---|---|---|---|
| Trial/Days | 3 | 25 | 3 | 25 | 3 | 25 | 3 | 25 |
| Ca | 154.40 ± 2.23a | 0.00 ± 0.00a | 4.07 ± 0.15c | 12.25 ± 0.32a | 7.65 ± 0.03c | 8.27 ± 0.26c | 0.35 ± 0.01c | 0.44 ± 0.05c |
| Ua | 208.77 ± 6.43c | 2.07 ± 0.90b | 2.03 ± 0.15a | 14.09 ± 0.15c | 5.15 ± 0.46b | 6.41 ± 0.49b | 0.30 ± 0.02b | 0.29 ± 0.02b |
| Cb | 170.77 ± 3.10b | 0.00 ± 0.00a | 3.70 ± 0.26b | 13.52 ± 0.34b | 5.21 ± 0.09b | 5.30 ± 0.11a | 0.28 ± 0.02ab | 0.20 ± 0.02a |
| Ub | 216.30 ± 2.08d | 1.87 ± 0.49b | 1.93 ± 0.06a | 14.54 ± 0.15c | 4.30 ± 0.24a | 6.10 ± 0.52b | 0.26 ± 0.02a | 0.25 ± 0.06ab |
1 Expressed as tartaric acid. 2 Expressed as acetic acid.
Table 2.
Oenological parameters of the second trial with quercetin and quercetin + thiamine nutrients. Different letters in each column indicate statistical differences (p < 0.05) between treatments in each fermentation scenario.
Table 2.
Oenological parameters of the second trial with quercetin and quercetin + thiamine nutrients. Different letters in each column indicate statistical differences (p < 0.05) between treatments in each fermentation scenario.
| Residual sugar (g/L) | Ethanol (% v/v) | Total acidity1 (g/L) | Volatile acidity2 (g/L) | |||||
|---|---|---|---|---|---|---|---|---|
| Trial/Days | 3 | 25 | 3 | 25 | 3 | 25 | 3 | 25 |
| Cq | 154.43 ± 4.31a | 0.00 ± 0.00a | 4.17 ± 0.23a | 12.89 ± 0.17a | 7.71 ± 0.13c | 8.36 ± 0.24a | 0.35 ± 0.01a | 0.39 ± 0.03bc |
| Uq | 172.13 ± 4.88a | 0.30 ± 0.17b | 3.60 ± 0.26a | 13.56 ± 0.85a | 7.32 ± 0.13b | 9.20 ± 0.18b | 0.35 ± 0.02a | 0.30 ± 0.02a |
| Cqt | 163.93 ± 18.89a | 0.00 ± 0.00a | 3.67 ± 0.93a | 13.04 ± 0.26a | 7.71 ± 0.13c | 8.52 ± 0.31a | 0.34 ± 0.02a | 0.46 ± 0.07c |
| Uqt | 168.37 ± 2.47a | 0.47 ± 0.25b | 3.90 ± 0.10a | 13.56 ± 0.06a | 7.07 ± 0.07a | 8.74 ± 0.13a | 0.33 ± 0.02a | 0.32 ± 0.03ab |
1 Expressed as tartaric acid. 2 Expressed as acetic acid.
Table 3.
Trial A and trial B parameters of colour intensity, tonality and total polyphenols index (TPI) measured on the last day of fermentation (day 25). Values are means ± sd (n = 3). A different letter for the same parameter and assay means significant differences (p < 0.05).
Table 3.
Trial A and trial B parameters of colour intensity, tonality and total polyphenols index (TPI) measured on the last day of fermentation (day 25). Values are means ± sd (n = 3). A different letter for the same parameter and assay means significant differences (p < 0.05).
| COLOR INTENSITY (Absorbance units) | TONALITY (Adimensional) |
TPI (Absorbance unirts) |
||
|---|---|---|---|---|
| Trial A | Ca | 2.13 ± 0.01b | 6.46 ± 0.01a | 15.23 ± 0.25a |
| Ua | 1.78 ± 0.09a | 10.26 ± 0.10c | 17.19 ± 0.72b | |
| Cb | 1.64 ± 0.01a | 7.63 ± 0.01b | 15.76 ± 0.15a | |
| Ub | 1.74 ± 0.01a | 10.39 ± 0.01c | 17.06 ± 0.51b | |
| Trial B | Cq | 2.00 ± 0.05b | 6.53 ± 0.05a | 14.95 ± 0.15a |
| Uq | 1.48 ± 0.05a | 10.22 ± 0.06b | 16.03 ± 0.64ab | |
| Cqt | 2.10 ± 0.03b | 6.52 ± 0.04a | 15.10 ± 0.15a | |
| Uqt | 1.56 ± 0.10a | 10.79 ± 0.13b | 16.40 ± 1.00b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



