
Submitted:
27 August 2024
Posted:
28 August 2024
You are already at the latest version
Abstract
Zebrafish (Danio rerio) is an important animal model for a wide range of neurodegenerative diseases. However, obtaining the cellular resolution that is essential for studying the zebrafish brain remains challenging as it requires high-spatial resolution and signal-to-noise ratios (SNR). In the current study, we present the first MRI results of the zebrafish brain at the state-of-the-art magnetic field strength of 28.2 T. The performance of MRI at 28.2 T was compared to 17.6 T. A 20% improvement in SNR was observed at 28.2 T as compared to 17.6 T. Excellent contrast, resolution, and SNR allowed the identification of several brain structures. The normative T1 and T2 relaxation values were established over different zebrafish brain structures at 28.2 T. To zoom into the white matter structures, we applied diffusion tensor imaging (DTI) and obtained axial, radial, and mean diffusivity, as well as fractional anisotropy at a very high spatial resolution. Visualization of white matter structures was achieved by short-track track-density imaging by applying the constrained spherical deconvolution method (stTDI CSD). For the first time, an algorithm for stTDI with multi-shell multi-tissue (msmt) CSD was tested on zebrafish brain data. A significant reduction of false-positive tracks from grey matter signals was observed compared to stTDI with single-shell single-tissue (ssst) CSD. This allowed the non-invasive identification of white matter structures at high resolution and contrast. Our results show that ultra-high field DTI and tractography provide reproducible and quantitative maps of fibre organization from tiny zebrafish brains, which can be implemented in the future for a mechanistic understanding of disease-related microstructural changes in zebrafish models of various brain diseases.
Keywords:
Magnetic resonance imaging
; diffusion MRI
; White matter tractography
; Zebrafish
1. Introduction
Zebrafish (Danio rerio) has become an excellent animal model in the studies of diseases, biological pathways, genetics, and toxicology [1,2,3,4]. In the field of neurodegenerative conditions, zebrafish offer various models including those for Alzheimer’s, Parkinson’s, and Huntington’s disease [5,6,7]. Therefore, studying the zebrafish brain non-invasively might provide valuable information on the pathology and treatment of neurodegenerative disorders. Magnetic resonance imaging (MRI) is a well-established, non-invasive technique for neuroimaging in both human and animal models. In our previous reports, a successful examination of zebrafish was performed at high field (9.4 T) [8,9,10] and ultra-high field (17.6 T) MRI [11,12,13]. High-quality images gave access to anatomical details, allowing visualization of white matter (WM) lesions in zebrafish models of familial cystic leukoencephalopathy [9] and Lowe syndrome [10], as well as in vivo analysis of malignant melanoma tumors [13]. Additionally, in vivo high-resolution localized magnetic resonance spectroscopy (MRS) was successfully applied to obtain the neurochemical metabolite profile of adult zebrafish. However, obtaining the essential resolution for studying small structures with high signal-to-noise ratio (SNR) remains challenging.
Insight into the microstructural organization of the zebrafish brain could be obtained by diffusion-based MRI (dMRI), a powerful, non-invasive technique with high sensitivity for water movement [14]. Cellular structures hinder the microscopic random motion of water, making dMRI unique to study the microstructural organisation of tissue. Diffusion tensor imaging (DTI) is an extended dMRI method providing increased structural information by exploiting anisotropic diffusion effects. Diffusion tensors are calculated from directional differences in the MR signals and used to determine the axial diffusivity (D∥) from principal eigenvalue, the radial diffusivity (D⊥) from the average of two non-principal eigenvalues, the mean diffusivity (MD), and the fractional anisotropy (FA), the extent of directional preference. In the brain, anisotropic diffusion effects are most prominent in WM due to the ordered structures of its myelinated axon tracts [14]. Changes in the diffusion anisotropy of WM structures have been reported for many neurodegenerative diseases including Alzheimer’s disease, Parkinson’s disease, Amyotrophic Lateral Sclerosis (ALS), and Huntington’s Disease [15,16,17,18,19,20,21,22,23,24,25]. The directionality of WM structures is visualized by directional encoded colour (DEC) FA maps, combining FA with the directionality of the principal eigenvector. However, FA colour maps do not visualize the connectivity of WM tracks, nor does the technique account for crossing or closely passing fibres below the applied MRI resolution. In this regard, DTI offers the solution for visualizing WM tracks. DTI tractography is a distinct processing technique of DTI tensors and the only known non-invasive imaging technique for visualizing WM connectivity in the brain. DTI has been successfully used to probe the changes in brain connectivity during neurogenerative diseases in human subjects [26,27,28,29,30,31,32]. Combined with the high spatial resolution required for the neurological analysis of zebrafish, dMRI is very challenging for the zebrafish brain. Consequently, knowledge of diffusivity and connectivity in the zebrafish brain is limited. Freidlin et al. [33] obtained good contrast of the spinal cord in adult zebrafish by DTI, while Ullmann et al. [34], presented a DTI study in the zebrafish brain and obtained tractography maps using short-track track density imaging with single-shell single-tissue constrained spherical deconvolution (stTDI ssst-CSD). However, the analysis was performed on isolated brain tissue, rather than intact zebrafish. Additionally, Ullmann et al. performed DTI with a single non-zero b-value, eliminating the possibility of individually estimating WM, grey matter (GM), and cerebrospinal fluid (CSF) signals, therefore possibly resulting in errors and WM overestimation during tractography [35]. Recently, multi-shell multi-tissue (msmt) CSD algorithms were developed, deconvoluting WM, GM, and CSF responses [35]. By filtering GM- and CSF-like signals strongly present in ssst-CSD, false positive tracks are reduced [36,37]. We have recently utilized msmt CSD methods at 17.6 T to identify white matter structures in the zebrafish brain of a toll-like receptor 2 deficient zebrafish model, aiming to compare white matter integrity [38]. However, a comparison of ssst CSD and msmt-CSD has not been validated for the zebrafish brain. In the current work, ssst CSD and msmt CSD methods are compared, marking a critical step in adapting and validating these advanced imaging techniques for use in zebrafish neuroimaging.
Exploiting higher magnetic fields for imaging can potentially improve the neurological analysis of zebrafish brain as SNR increases with the applied magnetic field strength (B0). Consequently, increased spatial resolution can be obtained without the need for significant elongation of total acquisition time. In this study, the first MRI results at 28.2 T are presented that were obtained from the zebrafish brain. The performance of MRI at 28.2 T was compared to 17.6 T and a wide range of MR sequences were optimized, including anatomical imaging by rapid acquisition with relaxation enhancement (RARE). Moreover, diffusion weighted imaging (DWI) was applied to quantify apparent diffusion coefficient (ADC) values in several zebrafish brain regions. Furthermore, white matter tractography was conducted through DTI using stTDI CSD. Our findings not only include the initial results of stTDI through ssst-CSD on intact zebrafish, but also show the first outcomes of stTDI through msmt-CSD on the zebrafish brain.
2. Results and Discussion
This study presents the first MRI findings of the zebrafish brain using a state-of-the-art magnetic field strength of 28.2 T. The zebrafish has become a popular species for studying neurological disorders due to the growing understanding of its nervous system and the availability of numerous transgenic zebrafish models [5,6,9,10,13]. Non-invasive MRI methods have significant potential for investigating brain pathology in these models. However, obtaining the essential resolution for studying microstructures and diffusion processes with a high SNR remains challenging for zebrafish brain. In this work, we have optimized and successfully applied MRI methods at 28.2 T magnetic field strength to resolve microstructural details and white matter tracts in the young adult zebrafish brain.
2.1. Relaxation Times
The application of MRI at an ultra-high field of 28.2 T requires adjustment of image acquisition parameters, which are based on knowledge of the MR relaxation properties of the tissues being imaged. To establish regional T1 and T2 values of the zebrafish brain at 28.2 T, relaxation times were estimated in nine manually selected ROIs (Figure 1A); the vagal lobe (Xlo), granular layer of the cerebellar corpus (CeG), Telencephalon (Te), Olfactory bulb (OB), molecular layer of the cerebellar corpus (Cem), Cerebellar crest (CC), Optic tectum (OT), Caudal zone of the periventricular hypothalamus (Hc), and the ventral zone of the periventricular hypothalamus (Hv). Figure 1B shows relaxation times T1 and T2 for selected ROIs at 28.2 T and compared them with values obtained at 17.6 T. A clear increase of T1 and a decrease of T2 were observed in all selected ROIs at 28.2 T compared to 17.6 T. On average, T1 increased by a factor of 1.097 ± 0.015, while T2 decreased by a factor of 0.905 ± 0.011 at 28.2 T compared to 17.6 T. The observed shift in relaxation times at increasing B0 is consistent with previous reports [39,40,41]. Generally, T1 is proportional to B01/3. Here, all brain regions report a T1 increase below the theoretical expectations. This is consistent with T1 reported in mouse brain tissue at 9.4 T and 17.6 T, where an average T1 increase factor of 1.08 was reported, well below the theoretical increase factor of 1.23 [42]. For T2, the observed decrease at higher magnetic fields is consistent with previous reports as well. In mouse brain regions, increasing the magnetic field from 9.4 T to 17.6 T resulted in an average T2 decrease by a factor 0.72 [43], changing from 9.4 T to 11.7 T resulted in an average T2 decrease by a factor 0.83 [40]. Besides B0, the relaxation times of tissues depend on factors like age, gender, acquisition parameters, and pre-treatment (measuring in vivo or fixed) [44]. To minimize the effects of additional factors, measurements at 17.6 T and 28.2 T were performed for the same zebrafish and with identical acquisition parameters including TE, TR, RARE factor, number of averages, and acquisition bandwidth. Furthermore, both MR systems contained similar RF coils, gradient power supplies, and software. To summarize, we have established the regional T1 and T2 values of the healthy zebrafish brain at 28.2 T. Our findings indicate that T2 decreases while T1 continues to rise with increasing magnetic field strength up to the ultra-high 28.2 T level. In future studies, accurate knowledge of T1 and T2 values of various brain regions at 28.2 T can serve as a reference point for detecting regional relaxation changes related to diseases in zebrafish brain.
2.2. Anatomical Imaging
Figure 2A shows representative slices of images of the zebrafish brain acquired using the RARE sequence at 17.6 T and 28.2 T. Images were acquired using the same zebrafish samples and identical acquisition parameters at both magnetic field strengths, obtaining a resolution of 23 μm × 23 μm and a slice thickness of 200 μm. A clear improvement in SNR is observed at 28.2 T as compared to 17.6 T (Figure 2A). On average, an SNR improvement factor of 1.2 was found at 28.2 T as compared to 17.6 T in the zebrafish brain. Several brain structures could be clearly identified at 28.2 T (Figure 2B). Figure 2B shows anatomical images, obtained at 28.2 T, in the coronal and sagittal direction at a field of view (FOV) of 6 × 6 mm, an image size of 256 × 256 voxels, and a slice thickness of 100 μm, resulting in a spatial resolution of 23 × 23 × 100 μm. Excellent contrast, resolution, and SNR allowed for the identification of several brain structures that were verified by comparing them with detailed atlases of the zebrafish brain [45,46].
2.3. Diffusion Weighted Imaging (DWI)
To get further contrast for the identification of WM structures such as commissures, fibre tracts, nerves, as well as the CSF system, we applied diffusion-based MRI techniques. dMRI provides contrast based on the Brownian motion of water molecules [47]. MRI is made sensitive to diffusion by strong gradient pulses before and after a 180° refocusing pulse, of which the magnitude (G), duration (δ), and time interval (∆) are summarized in the b-value. In Figure 3A, the effect of the applied b-value in DWI experiments is shown for the zebrafish brain. At low b-values of 75 s/mm2, high contrast and SNR are obtained for all regions in the zebrafish head. At moderate b-values of 500 and 1000 s/mm2, the signal intensity of muscle and surrounding tissue is reduced, while a high contrast and SNR in the brain is maintained. In fact, DWI images obtained at 1000 s/mm2 show better contrast between various brain regions than those obtained at 75 and 500 s/mm2. This is clearer in the forebrain where the olfactory bulb gained significant contrast compared to anatomical imaging (Figure 2B & 3A). This allowed better contrast for the identification of various structures in the brain compared to anatomical images obtained by the RARE sequence (supplementary Figure S1). At a high b-values of 3500 s/mm2, the MR signal of all tissue drops, although the brain is still visible. Some signal from brain was even visible at very high b-values of 5000 and 7500 s/mm2. Figure 3B shows the ADC map. On average, the brain shows relatively low diffusivity (< 5 mm2/s) compared to most surrounding tissue (> 10 mm2/s), resulting in a clear high contrast between brain and the surrounding tissue. Figure 3C shows an ADC map of the brain region from the central imaging slice. In addition to excellent contrast between various brain regions, the CSF system is clearly visible, allowing for the identification of several ventricles. Brain structures including the diffusive nucleus of the inferior lobe, ventral zone of the periventricular hypothalamus, and medial zone of dorsal telencephalon, show relatively high diffusivity. On the other hand, structures, such as the olfactory bulb, cerebellar corpus, longitudinal torus, and medial longitudinal fascicle show relatively low diffusivity. Figure 3D shows estimated ADC values for selected ROIs obtained at 17.6 T and 28.2 T (see Figure 1A for ROIs). At both magnetic fields, the highest diffusivity is found in the telencephalon and ventral zone of the periventricular hypothalamus, while the lowest diffusivity is found in the olfactory bulb. Some differences in ADC values were seen at 28.2 T as compared to 17.6 T. Although ADC values are independent of the applied magnetic field strength, they are affected by SNR. Thus, reported differences at 28.2 T and 17.6 T likely originate from differences in SNR [48].
2.4. Diffusion Tensor Imaging (DTI)
Figure 4 shows representative 2D DTI results of adult zebrafish brain obtained at 28.2 T. All images show an identical sagittal slice, rostral (left) to caudal (right), acquired at a resolution of 25 μm x 25 μm, and a slice thickness of 200 μm. Figure 4A-C show D∥, D⊥, and MD maps of the zebrafish brain, respectively. Various structures could be identified by diffusivity-based contrast. The vascular system and specific brain regions, including the telencephalon, diffusive nucleus of the inferior lobe, and the ventral zone of the periventricular hypothalamus show a relatively high diffusivity. However, the olfactory bulb and the optic tectum show relatively low diffusivity compared to other brain structures. These results are consistent with observations made by DWI results shown in Figure 3. Furthermore, differences in the D∥ and D⊥ are observed, indicating anisotropic diffusivity effects. Figure 4D shows the fractional anisotropy map, visualizing the extent of anisotropic diffusivity in the zebrafish brain. High FA values (light colour) indicate anisotropic diffusion effects, while low FA values (dark colour) indicate more isotropic diffusion effects. It is well known that WM structures show highest FA values in the brain due to the ordered structures of its myelinated axon tracts [14]. Due to high FA values, WM brain structures show very high image contrast, allowing identification of various WM structures including the medial longitudinal fascicle, ansulate commissure, posterior commissure, and optic tract (indicated in Figure 4D). These structures could not be identified by anatomical imaging or DWI (Figure 2 & 3). Figure 4E shows the DEC FA colour map of the central sagittal slice of an adult zebrafish brain, obtained by combining directional information of the principal eigenvalues and FA maps. The FA colour map further confirms the identification of WM structures by FA, showing the rostral-caudal orientation of the medial longitudinal fascicle, and the medial-lateral orientation of the ansulate commissure, posterior commissure, and optic tract.
Figure 5A show representative super-resolution stTDI tractography maps of 2D DTI data. Basic tractography algorithms are deterministic, which allow fitting the diffusion tensors to the diffusion data by tracking paths through the principal eigenvector of the tensor [49]. However, deterministic algorithms do not allow for crossing or closely passing fibres, potentially leading to non-existing connections [50,51]. dMRI tractography with CSD processes crossing and closely passing fibres, based on the first-order integration over fibre orientation distribution functions (fODFs) [50]. These fODFs are estimated from response functions, in particular the estimated signal for single-fibre WM. In Figure 5A, CSD was performed by single-shell single-tissue (ssst) algorithms to estimate the response function. The stTDI maps by ssst-CSD show high-resolution fibre-tracts, providing directional information beyond the DTI spatial resolution. Similar stTDI maps have been generated from zebrafish brain in a previous DTI study performed at a lower magnetic field of 16.T [34]. However, the DTI measurement was performed on surgically isolated zebrafish brain treated with MRI contrast agent. In our work, we performed all the measurements of intact zebrafish at 28.2 T without any contrast agent. One of the shortcomings of performing CSD with ssst CSD algorithms is that it generates a track map without distinguishing individual anatomical structures (such as grey matter, CSF, and WM). Thus, potentially producing WM tracking errors and overestimation [35].
To produce tracks exclusively in the WM area, we applied multi-shell-multi-tissue algorithms to obtain stTDI maps (stTDI msmt-CSD). The msmt-CSD was originally developed and optimized for human brain data [37] and has also been applied to mouse brain DTI data [52]. However, it has not yet been applied to the zebrafish brain. In this study we implemented and successfully obtained the stTDI map by msmt-CSD in zebrafish brain (Figure 5B). The response function estimation for WM, GM, and CSF obtained with msmt-CSD derived from our DTI data set is shown in supplementary Figure S2. As can be seen from Figure 5B, stTDI by msmt-CSD produced tracks exclusively in WM areas. Furthermore, the stTDI map generated by msmt-CSD demonstrates a significant reduction in noise over those obtained by ssst-CSD (Figure 5), leading to improved identification of WM structures. Figure 5C&D show fODFs estimated by ssst and msmt CSD algorithms, respectively, from the optic tectum (see the highlighted area in Figure 5A&B). As shown in Figure 5D, the msmt-CSD fODFs have fewer lobes compared to the ssst-CSD shown in Figure 5C. This difference in lobe number indicates a reduced number of possible tracks obtained by the msmt-CSD method. Our results are consistent with earlier reports in human and mouse brain [37,52]. An increased number of tracks with ssst-CSD indicates tracking errors due to overestimation resulting in false positive WM tracks. Unlike msmt-CSD algorithms, ssst-CSD does not individually characterize WM, GM, and CSF signals, resulting in WM tracking errors and overestimation [35]. The development of the msmt-CSD algorithm revealed that WM, GM, and CSF have distinct dependencies on b-values [35], enabling the deconvolution of their individual signals. By filtering GM- and CSF signals, strongly present in ssst-CSD, the number of false positive WM tracks is reduced with the msmt-CSD algorithm [36,37].
Next, we acquire data using 3D DTI, as it allows for better visualization and quantification of white matter tracts, particularly in the areas with complex fibre orientation. The representative 3D DTI results of adult zebrafish brain, acquired at an isotropic resolution of 35 μm, are shown in supplementary Figure S3. From the 3D DTI data, FA maps and DEC FA colour maps were generated. Furthermore, 5 μm stTDI tractography maps were created from 3D DTI data (supplementary Figure S3). Both ssst-CSD and msmt-CSD algorithms were used to estimate the response function. Enhanced resolution for crossing and closely passing fibres was clearly obtained with msmt-CSD as compared to ssst-CSD. Similar to 2D DTI data, visualization of WM structures by stTDI with msmt-CSD is significantly improved compared to ssst-CSD.
Figure 6 demonstrates the capability to identify WM structures in the zebrafish brain using 3D super-resolution stTDI msmt-CSD. In the sagittal (Figure 6A), coronal (Figure 6B), and axial (Figure 6C) view, WM structures were identified at high contrast and resolution, that were verified by comparing them with detailed atlases of the zebrafish brain [45,46]. A selection of the commissures, fibre tracts, and nerves that could be assigned are shown. This study shows that DTI at ultra-high field in conjunction with stTDI msmt-CSD provide great resolution for visualization of white matter tracts in intact zebrafish without the need to isolate the brain.
3. Materials and Methods
3.1. Zebrafish Husbandry
The husbandry of adult zebrafish described in this study complied with guidelines from the local animal welfare committee of the university (License number: AVD1060020171767 and AVD10600202216175), following the international guidelines specified by the EU Animal Protection Directive 2010/63/EU, and was conducted according to standard protocols (www.zfin.org) as described previously [53]. In this study, adult zebrafish (wild-type, male, n = 6) were used. Adult zebrafish aged between 4 and 6 months were euthanized through immobilization by submersion in ice water (0-4°C) for at least 10 minutes following cessation of opercular movement and then fixed in 4% buffered paraformaldehyde (Zinc Formal-Fixx, ThermoShandon, UK) for 4 days and subsequently embedded in perfluoropolyether (Fomblin Y, Solvay Solexis S.P.A.) for MRI measurements.
3.2. Magnetic Resonance Imaging
MRI was performed in a 28.2 T (1.2 GHz) and a 17.6 T (750 MHz) vertical bore magnet (Bruker Biospin, Ettlingen, Germany). Both systems were equipped with a MICRO 5 gradient system (Gmax 3 T/m), a 5 mm birdcage RF coil, and a GREAT 60 gradient power supply. Data acquisition and processing were performed using Paravision 360 v3.3 (Bruker Biospin, Ettlingen, Germany). For all measurements, animals were transferred to 5 mm NMR tubes and embedded in Fomblin. To achieve maximal signal intensity, the position of the brain was aligned to the centre of the RF coil. Magnetic field homogeneity was achieved by shimming, up to the second order. A variety of scan protocols was used and optimized for the magnetic field strength of 28.2 T. Identical scan protocols were then used for both MR systems (17.6 T and 28.2 T). For anatomical imaging, a two-dimensional (2D) RARE sequence was used. Images were acquired with an echo time (TE) of 5.6 ms, a repetition time (TR) of 3000 ms, using 4 segmenting refocusing echoes (RARE factor), and a spatial resolution of 23 × 23 μm. RARE images were obtained at a slice thickness of 100 and 200 μm using 64 and 16 averages, respectively.
For the estimation of spin-spin relaxation times (T2) a 2D Multi-Slice Multi-Echo (MSME) protocol was used, based on the Carr-Purcell-Meiboom-Gill (CPMG) sequence [54]. MSME measurements were performed with 15 echo images per excitation at an echo spacing of 5 ms, a TR of 2500 ms, 4 averages, at a spatial resolution of 23 × 23 μm, and a slice thickness of 200 μm. For estimation of the spin-lattice relaxation times (T1), a 2D RARE protocol at variable Repetition times (VTR) was used. Signal intensities were measured at a TR of 300, 606, 967, 1408, 1974, 2767, 4100, and 10000 ms, with a TE of 3 ms, and 2 averages, at a spatial resolution of 47 × 47 μm and a slice thickness of 200 μm.
Diffusion weighted imaging (DWI) was performed with a 2D diffusion-weighted spin-echo sequence (DWI SE) with a TE of 20.2 ms, a TR of 1000 ms, 4 averages, at a spatial resolution of 23 × 23 μm and a slice thickness of 200 μm. MR signals were measured at b-values of 75, 500, 1000, 3500, 5000, and 7500 s/mm2. 2D DTI with echo planar imaging (EPI) was performed at an anisotropic resolution of 25 × 25 μm, at a slice thickness of 200 μm, with a TE of 12.4 ms, TR of 2000 ms, 32 averages, and an EPI factor of 8. Multi-shell experiments were performed with b-values of 4, 1000, 3500, and 6000 s/mm2 with 8, 12, 24, and 36 directions respectively. Three-dimensional (3D) DTI was performed at an isotropic spatial resolution of 35 μm, with a TE of 9.1 ms, TR of 2000 ms, 4 averages, and an EPI factor of 8. Multi-shell experiments were performed with b-values of 100, 1000, and 2500 s/mm2 with 4, 12, and 24 directions respectively. Automatic drift compensation was applied to compensate for possible B0 drift.
3.3. Data Processing
Identification of brain regions and WM structures was performed based on the Adult Zebrafish Brain Atlas [45], the topological atlas [46], and Ullmann et al.[34].
For the estimation of SNR, RARE intensity images were processed in Matlab (mathworks.com). Brain tissue was selected using the volume segmenter tool and the average signal intensity was calculated. For noise, 10 × 10 voxels were selected outside the fish. From these data, SNR was calculated using Eq.1.
where μS is the mean intensity of the brain signal, μN is the mean intensity of the noise and σN is the standard deviation of the noise. SNR maps were created by estimating SNR for each voxel inside the RARE intensity image. Signals outside the fish were set to zero by a signal intensity threshold.
[Eq.1] SNR = (μS – μN)/ σN
For the estimation of spin-lattice relaxation times (T1), spin-spin relaxation times (T2), and ADC values, nine regions of interest (ROIs) were manually selected in the zebrafish brain using Paravision 360 v3.3. T1 values were estimated by the image sequence analysis tool in Paravision using a nonlinear least square algorithm for the mono-exponential fit function [Eq.2] using RAREVTR data.
where It is the signal intensity at time t, A is the absolute bias, I0 is the signal intensity at time t0, t is the repetition time (ms), and T1 is longitudinal relaxation time (ms). From MSME data, T2 values were estimated using the image sequence analysis tool in Paravision with a nonlinear least square algorithm for the mono-exponential fit function [Eq.3].
where It is the signal intensity at time t, A is the absolute bias, I0 is the signal intensity at time t0, t is the echo time (ms), and T2 is the transverse relaxation time (ms). To reduce the effect of imperfect 180° pulses, uneven echoes were excluded from the data fitting [55]. ADC values were estimated from DWI data by the image sequence analysis tool in Paravision with a nonlinear least square algorithm for the mono-exponential fit function [Eq.4].
where Ib is the signal intensity at b, A is the absolute bias, I0 is the signal intensity at b = 0 mm2/s, b is the b-value (s/mm2), and ADC is the apparent diffusion coefficient (mm2/s).
[Eq.2] It = A + I0 ∙ (1- exp (-t/T1))
[Eq.3] It = A + I0 ∙ exp (-t/T2)
[Eq.4] Ib = A + I0 ∙ exp (-b*ADC)
Processing of 2D and 3D DTI data was performed by MRtrix3 [56]. Brain masks were manually created using the volume segmenter tool in Matlab. Using MRtrix3 functions, the denoising of DTI data was based on the random matrix theory [57]. Diffusion tensors were calculated by fitting the diffusion tensor to the log of the denoised DTI data by minimizing the weighted least-squares and iterated weighted least-squares [58,59]. From the tensors, the D∥, D⊥, MD, and FA were calculated. FA colour maps were generated by adding the directionality of the principal eigenvalue to FA maps. MRtrix3 was used to generate super-resolution (5 μm) stTDI maps from 2D and 3D DTI data. Constrained Spherical Deconvolution (CSD) was used to resolve crossing fibres by estimating fibre Orientation Distribution Functions (fODFs). Single-shell single-tissue (ssst) CSD response functions were estimated by the Tournier algorithm [60], using the highest b-value of the DTI dataset. fODFs were calculated with the ssst-CSD algorithm [61]. Multi-shell multi-tissue (msmt) CSD response functions were estimated by the Dhollander algorithm [36]. Crude segmentation of WM and GM/CSF voxels in 2D and 3D DTI data was performed at FA = 0.2 (default). WM, GM, and CSF voxel selection of 3D DTI data for response function estimation was performed with the top 0.5% of refined WM voxels, 2.0% of refined GM voxels, and 10% of refined GM voxels. For the 2D DTI data, 10% of the refined WM voxels were used in the final selection due to the lower total number of available voxels compared to the 3D DTI data set. Finally, msmt-CSD fODFs were calculated with the msmt-CSD algorithm [37]. Whole brain fibre tracking was performed with the iFOD1 algorithm [62]. 1 million (2D DTI data) and 10 million (3D DTI data) tracks were generated with a minimal track length of twice the voxel size and a maximal track length of ten times the voxel size. Fibre generation was automatically terminated once leaving the pre-defined brain mask. Additional parameters were used in their default mode [56]. Tractography results were transformed into super-resolution Track Density Imaging (TDI) [63], resulting in approximately 5μm isotropic resolution. The directionality of generated tracks is visualized by DEC.
4. Conclusions
In conclusion, here we have shown the potential of MRI techniques at ultra-high magnetic fields (28.2 T) to study the zebrafish brain non-invasively. Excellent contrast and SNR were obtained allowing for the identification of brain structures, as well as fine white matter structures in intact zebrafish brain. Furthermore, ultra-high field DTI was capable of generating reliable and quantitative representations of the fibre organization within the small brains of zebrafish. This opens up the possibility to study disease-related changes in white matter structure with super high resolution in a wide range of zebrafish models of human diseases.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Figure S1. Superior identification of brain structures based on diffusion weighted imaging (DWI) contrast as compared to RARE image contrast. Figure S2. Response function estimation from DTI dataset of adult zebrafish brain. Figure S3. 3D DTI results of adult zebrafish brain, showing coronal, sagittal, and axial slices, acquired at 28.2 T.
Author Contributions
R.S., A.A. H.P.S designed the experiments. R.S. and I.O. conducted MRI experiments. WH, YD zebrafish breading & sample preparations. R.S., I.O., A.P., & A.A. analysed the results. H.J.M.G, H.P.S, A.A. supervised the project. R.S., A.A wrote the manuscript. All authors reviewed the manuscript.
Funding
Experiments at the 1.2 GHz instrument were supported by uNMR-NL, an NWO-funded National Roadmap Large-Scale Facility of the Netherlands (project 184.032.207). Alia thanks the grant (SFB TRR-386 HYP*MOL) from Deutsche Forschungsgemeinschaft (DFG).
Data Availability Statement
The data presented in this study are available on request from the author (Y.K).
Acknowledgments
The authors thank Dr. Julia Krug & Dr. Jeanine Prompers for useful discussions and suggestions. We also acknowledge Dr. Andrei Gurinov for technical support at the uNMR-NL facility and Karthick Babu Sai Sankar Gupta and Fons Lefeber for help at 17.6 T. We would further like to thank Volker Lehmann and Thomas Oerther from Bruker for their help in troubleshooting.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bailone RL, Aguiar LKd, Roca RdO, Borra RC, Corrêa T, Janke H, Fukushima HCS (2019) “Zebrafish as an animal model for food safety research: trends in the animal research”. Food Biotechnology 33, 283-302.
- Cassar S, Adatto I, Freeman JL, Gamse JT, Iturria I, Lawrence C, Muriana A, Peterson RT, Van Cruchten S, Zon LI (2020) Use of Zebrafish in Drug Discovery Toxicology. Chem Res Toxicol 33, 95-118.
- Choi TY, Choi TI, Lee YR, Choe SK, Kim CH (2021) Zebrafish as an animal model for biomedical research. Exp Mol Med 53, 310-317.
- Ding Y, Haks MC, Forn-Cuni G, He J, Nowik N, Harms AC, Hankemeier T, Eeza MNH, Matysik J, Alia A, Spaink HP (2021) Metabolomic and transcriptomic profiling of adult mice and larval zebrafish leptin mutants reveal a common pattern of changes in metabolites and signaling pathways. Cell Biosci 11, 126.
- Bashirzade AA, Zabegalov KN, Volgin AD, Belova AS, Demin KA, de Abreu MS, Babchenko VY, Bashirzade KA, Yenkoyan KB, Tikhonova MA, Amstislavskaya TG, Kalueff AV (2022) Modeling neurodegenerative disorders in zebrafish. Neurosci Biobehav Rev 138, 104679.
- Wang X, Zhang JB, He KJ, Wang F, Liu CF (2021) Advances of Zebrafish in Neurodegenerative Disease: From Models to Drug Discovery. Front Pharmacol 12, 713963.
- Wang J, Cao H (2021) Zebrafish and Medaka: Important Animal Models for Human Neurodegenerative Diseases. Int J Mol Sci 22.
- Kabli S, Alia A, Spaink HP, Verbeek FJ, De Groot HJ (2006) Magnetic resonance microscopy of the adult zebrafish. Zebrafish 3, 431-439.
- Haud N, Kara F, Diekmann S, Henneke M, Willer JR, Hillwig MS, Gregg RG, Macintosh GC, Gartner J, Alia A, Hurlstone AF (2011) rnaset2 mutant zebrafish model familial cystic leukoencephalopathy and reveal a role for RNase T2 in degrading ribosomal RNA. Proc Natl Acad Sci U S A 108, 1099-1103.
- Ramirez IB, Pietka G, Jones DR, Divecha N, Alia A, Baraban SC, Hurlstone AF, Lowe M (2012) Impaired neural development in a zebrafish model for Lowe syndrome. Hum Mol Genet 21, 1744-1759.
- Kabli S, Spaink HP, De Groot HJ, Alia A (2009) In vivo metabolite profile of adult zebrafish brain obtained by high-resolution localized magnetic resonance spectroscopy. J Magn Reson Imaging 29, 275-281.
- Eeza MNH, Singer R, Ding Y, He J, Zuberi Z, Baelde HJ, de Groot HJM, Matysik J, Spaink HP, Alia A (2023) Probing microstructural changes in muscles of leptin-deficient zebrafish by non-invasive ex-vivo magnetic resonance microimaging. PLoS One 18, e0284215.
- Kabli S, He S, Spaink HP, Hurlstone A, Jagalska ES, De Groot HJ, Alia A (2010) In vivo magnetic resonance imaging to detect malignant melanoma in adult zebrafish. Zebrafish 7, 143-148.
- Alexander AL, Lee JE, Lazar M, Field AS (2007) Diffusion tensor imaging of the brain. Neurotherapeutics 4, 316-329.
- Auning E, Kjaervik VK, Selnes P, Aarsland D, Haram A, Bjornerud A, Hessen E, Esnaashari A, Fladby T (2014) White matter integrity and cognition in Parkinson’s disease: a cross-sectional study. BMJ Open 4, e003976.
- Bede P, Elamin M, Byrne S, McLaughlin RL, Kenna K, Vajda A, A. F, Bradley DG, Hardiman O (2014) Patterns of cerebral and cerebellar white matter degeneration in ALS. J Neurol Neurosurg Psychiatry 86.
- de Schipper LJ, Hafkemeijer A, Bouts M, van der Grond J, Marinus J, Henselmans JML, van Hilten JJ (2019) Age- and disease-related cerebral white matter changes in patients with Parkinson’s disease. Neurobiol Aging 80, 203-209.
- Gold BT, Johnson NF, Powell DK, Smith CD (2012) White matter integrity and vulnerability to Alzheimer’s disease: preliminary findings and future directions. Biochim Biophys Acta 1822, 416-422.
- Huang P, Xu X, Gu Q, Xuan M, Yu X, Luo W, Zhang M (2014) Disrupted white matter integrity in depressed versus non-depressed Parkinson’s disease patients: a tract-based spatial statistics study. J Neurol Sci 346, 145-148.
- Kantarci K, Murray ME, Schwarz CG, Reid RI, Przybelski SA, Lesnick T, Zuk SM, Raman MR, Senjem ML, Gunter JL, Boeve BF, Knopman DS, Parisi JE, Petersen RC, Jack CR, Jr., Dickson DW (2017) White-matter integrity on DTI and the pathologic staging of Alzheimer’s disease. Neurobiol Aging 56, 172-179.
- Novak MJ, Seunarine KK, Gibbard CR, Hobbs NZ, Scahill RI, Clark CA, Tabrizi SJ (2014) White matter integrity in premanifest and early Huntington’s disease is related to caudate loss and disease progression. Cortex 52, 98-112.
- Reading SA, Yassa MA, Bakker A, Dziorny AC, Gourley LM, Yallapragada V, Rosenblatt A, Margolis RL, Aylward EH, Brandt J, Mori S, van Zijl P, Bassett SS, Ross CA (2005) Regional white matter change in pre-symptomatic Huntington’s disease: a diffusion tensor imaging study. Psychiatry Res 140, 55-62.
- Stricker NH, Schweinsburg BC, Delano-Wood L, Wierenga CE, Bangen KJ, Haaland KY, Frank LR, Salmon DP, Bondi MW (2009) Decreased white matter integrity in late-myelinating fiber pathways in Alzheimer’s disease supports retrogenesis. Neuroimage 45, 10-16.
- van der Burgh HK, Westeneng HJ, Walhout R, van Veenhuijzen K, Tan HHG, Meier JM, Bakker LA, Hendrikse J, van Es MA, Veldink JH, van den Heuvel MP, van den Berg LH (2020) Multimodal longitudinal study of structural brain involvement in amyotrophic lateral sclerosis. Neurology 94, e2592-e2604.
- Zhang J, Yin X, Zhao L, Evans AC, Song L, Xie B, Li H, Luo C, Wang J (2014) Regional alterations in cortical thickness and white matter integrity in amyotrophic lateral sclerosis. J Neurol 261, 412-421.
- Abe O, Yamada H, Masutani Y, Aoki S, Kunimatsu A, Yamasue H, Fukuda R, Kasai K, Hayashi N, Masumoto T, Mori H, Soma T, Ohtomo K (2004) Amyotrophic lateral sclerosis: diffusion tensor tractography and voxel-based analysis. NMR Biomed 17, 411-416.
- Agosta F, Pagani E, Petrolini M, Caputo D, Perini M, Prelle A, Salvi F, Filippi M (2010) Assessment of white matter tract damage in patients with amyotrophic lateral sclerosis: a diffusion tensor MR imaging tractography study. AJNR Am J Neuroradiol 31, 1457-1461.
- Kitamura S, Kiuchi K, Taoka T, Hashimoto K, Ueda S, Yasuno F, Morikawa M, Kichikawa K, Kishimoto T (2013) Longitudinal white matter changes in Alzheimer’s disease: a tractography-based analysis study. Brain Res 1515, 12-18.
- Kok JG, Leemans A, Teune LK, Leenders KL, McKeown MJ, Appel-Cresswell S, Kremer HPH, de Jong BM (2020) Structural Network Analysis Using Diffusion MRI Tractography in Parkinson’s Disease and Correlations With Motor Impairment. Front Neurol 11, 841.
- Lo CY, Wang PN, Chou KH, Wang J, He Y, Lin CP (2010) Diffusion tensor tractography reveals abnormal topological organization in structural cortical networks in Alzheimer’s disease. J Neurosci 30, 16876-16885.
- Phillips O, Sanchez-Castaneda C, Elifani F, Maglione V, Di Pardo A, Caltagirone C, Squitieri F, Sabatini U, Di Paola M (2013) Tractography of the corpus callosum in Huntington’s disease. PLoS One 8, e73280.
- Zhang Y, Wu IW, Buckley S, Coffey CS, Foster E, Mendick S, Seibyl J, Schuff N (2015) Diffusion tensor imaging of the nigrostriatal fibers in Parkinson’s disease. Mov Disord 30, 1229-1236.
- Freidlin RZ, Morris HD, Horkay F, Pierpaoli C, Toyama R, Dawid IB, Basser PJ (2004) Diffusion Tensor MR Microscopy of Adult Zebrafish. Proc. Intl. Soc. Mag. Reson. Med. 11.
- Ullmann JF, Calamante F, Collin SP, Reutens DC, Kurniawan ND (2015) Enhanced characterization of the zebrafish brain as revealed by super-resolution track-density imaging. Brain Struct Funct 220, 457-468.
- Dhollander T, Raffelt D, Connelly A (2016) Unsupervised 3-tissue response function estimation from single-shell or multi-shell diffusion MR data without a co-registered T1 image.
- Dhollander T, Raffelt D, Mito R, Connelly A (2019) Improved white matter response function estimation for 3-tissue constrained spherical deconvolution. Conference Paper.
- Jeurissen B, Tournier JD, Dhollander T, Connelly A, Sijbers J (2014) Multi-tissue constrained spherical deconvolution for improved analysis of multi-shell diffusion MRI data. Neuroimage 103, 411-426.
- Singer, R, Oganezova, I, Hu W, Liu L, Ding Y, de Groot HJ, Spaink HP, Alia A (2024) Ultrahigh field diffusion magnetic resonance imaging uncovers intriguing microstructural changes in the adult zebrafish brain caused by Toll-like receptor 2 genomic deletion. NMR Biomed, p. e5170.
- Bottomley PA, Foster TH, Argersinger RE, Pfeifer LM (1984) A review of normal tissue hydrogen NMR relaxation times and relaxation mechanisms from 1-100 MHz: dependence on tissue type, NMR frequency, temperature, species, excision, and age. Med Phys 11, 425-448.
- de Graaf RA, Brown PB, McIntyre S, Nixon TW, Behar KL, Rothman DL (2006) High magnetic field water and metabolite proton T1 and T2 relaxation in rat brain in vivo. Magn Reson Med 56, 386-394.
- Korb JP, Bryant RG (2002) Magnetic field dependence of proton spin-lattice relaxation times. Magn Reson Med 48, 21-26.
- van de Ven RC, Hogers B, van den Maagdenberg AM, de Groot HJ, Ferrari MD, Frants RR, Poelmann RE, van der Weerd L, Kiihne SR (2007) T(1) relaxation in in vivo mouse brain at ultra-high field. Magn Reson Med 58, 390-395.
- Kara F, Chen F, Ronen I, de Groot HJ, Matysik J, Alia A (2013) In vivo measurement of transverse relaxation time in the mouse brain at 17.6 T. Magn Reson Med 70, 985-993.
- Hamilton N, Allen C, Reynolds S (2022) Longitudinal brain studies in adult zebrafish by MRI.
- Kenney JW, Steadman PE, Young O, Shi MT, Polanco M, Dubaishi S, Covert K, Mueller T, Frankland PW, A 3D adult zebrafish brain atlas (AZBA) for the digital age,eLife, http://azba.wayne.edu/, Accessed 09.
- Wullimann MF, Rupp B, Reichert H (1996) Neuroanatomy of the zebrafish brain A topological atlas, Basel Boston Berlin.
- Baliyan V, Das CJ, Sharma R, Gupta AK (2016) Diffusion weighted imaging: Technique and applications. World J Radiol 8, 785-798.
- Ogura A, Tamura T, Ozaki M, Doi T, Fujimoto K, Miyati T, Ito Y, Maeda F, Tarewaki H, Takahashi M (2015) Apparent Diffusion Coefficient Value Is Not Dependent on Magnetic Resonance Systems and Field Strength Under Fixed Imaging Parameters in Brain. J Comput Assist Tomogr 39, 760-765.
- Basser PJ, Pajevic S, Pierpaoli C, Duda J, Aldroubi A (2000) In vivo fiber tractography using DT-MRI data. Magnetic Resonance in Medicine 44, 625-632.
- Jeurissen B, Leemans A, Jones DK, Tournier JD, Sijbers J (2011) Probabilistic fiber tracking using the residual bootstrap with constrained spherical deconvolution. Hum Brain Mapp 32, 461-479.
- Jeurissen B, Leemans A, Tournier JD, Jones DK, Sijbers J (2013) Investigating the prevalence of complex fiber configurations in white matter tissue with diffusion magnetic resonance imaging. Hum Brain Mapp 34, 2747-2766.
- Jillings S, Morez J, Vidas-Guscic N, Van Audekerke J, Wuyts F, Verhoye M, Sijbers J, Jeurissen B (2020) Multi-tissue constrained spherical deconvolution in a murine brain. Proc. Intl. Soc. Mag. Reson. Med. 28.
- He J, Ding Y, Nowik N, Jager C, Eeza MNH, Alia A, Baelde HJ, Spaink HP (2021) Leptin deficiency affects glucose homeostasis and results in adiposity in zebrafish. J Endocrinol 249, 125-134.
- Carr HY, Purcell EM (1954) Effects of Diffusion on Free Precession in Nuclear Magnetic Resonance Experiments. Physical Review 94, 630-638.
- Milford D, Rosbach N, Bendszus M, Heiland S (2015) Mono-Exponential Fitting in T2-Relaxometry: Relevance of Offset and First Echo. PLoS One 10, e0145255.
- Tournier JD, Smith R, Raffelt D, Tabbara R, Dhollander T, Pietsch M, Christiaens D, Jeurissen B, Yeh CH, Connelly A (2019) MRtrix3: A fast, flexible and open software framework for medical image processing and visualisation. Neuroimage 202, 116137.
- Veraart J, Fieremans E, Novikov DS (2016) Diffusion MRI noise mapping using random matrix theory. Magn Reson Med 76, 1582-1593.
- Basser PJ, Mattiello J, Lebihan D (1994) Estimation of the Effective Self-Diffusion Tensor from the NMR Spin Echo. Journal of Magnetic Resonance B, 8.
- Veraart J, Sijbers J, Sunaert S, Leemans A, Jeurissen B (2013) Weighted linear least squares estimation of diffusion MRI parameters: strengths, limitations, and pitfalls. Neuroimage 81, 335-346.
- Tournier JD, Calamante F, Connelly A (2013) Determination of the appropriate b value and number of gradient directions for high-angular-resolution diffusion-weighted imaging. NMR Biomed 26, 1775-1786.
- Tournier JD, Calamante F, Connelly A (2007) Robust determination of the fibre orientation distribution in diffusion MRI: non-negativity constrained super-resolved spherical deconvolution. Neuroimage 35, 1459-1472.
- Tournier JD, Calamante F, Connelly A (2012) MRtrix: Diffusion tractography in crossing fiber regions. International Journal of Imaging Systems and Technology 22, 53-66.
- Calamante F, Tournier JD, Jackson GD, Connelly A (2010) Track-density imaging (TDI): super-resolution white matter imaging using whole-brain track-density mapping. Neuroimage 53, 1233-1243.
Figure 1.
T1 and T2 relaxation time measurements in various brain regions of adult zebrafish. [A] Anatomical RARE image of adult zebrafish indicating the location of ROIs selected for T1 and T2 quantification. [B] T1 and T2 relaxation time values in various zebrafish brain regions measured at 28.2 and 17.6 T. The difference between T1 and T2 values measured at two different magnetic field strengths is presented as fold change (Factor). Selected ROIs; Xlo – Vagal lobe; CC – Cerebellar crest; CeM – Cerebellar corpus, molecular layer; CeG – Cerebellar corpus, granular layer; OT – Optic Tectum; Te - Telencephalon; OB – Olfactory bulb; Hc – Caudal zone of periventricular hypothalamus; Hv – Ventral zone of periventricular hypothalamus. Data represent the mean T1 and T2 in ms ± standard error (SE) (Error bars); n=6.
Figure 1.
T1 and T2 relaxation time measurements in various brain regions of adult zebrafish. [A] Anatomical RARE image of adult zebrafish indicating the location of ROIs selected for T1 and T2 quantification. [B] T1 and T2 relaxation time values in various zebrafish brain regions measured at 28.2 and 17.6 T. The difference between T1 and T2 values measured at two different magnetic field strengths is presented as fold change (Factor). Selected ROIs; Xlo – Vagal lobe; CC – Cerebellar crest; CeM – Cerebellar corpus, molecular layer; CeG – Cerebellar corpus, granular layer; OT – Optic Tectum; Te - Telencephalon; OB – Olfactory bulb; Hc – Caudal zone of periventricular hypothalamus; Hv – Ventral zone of periventricular hypothalamus. Data represent the mean T1 and T2 in ms ± standard error (SE) (Error bars); n=6.
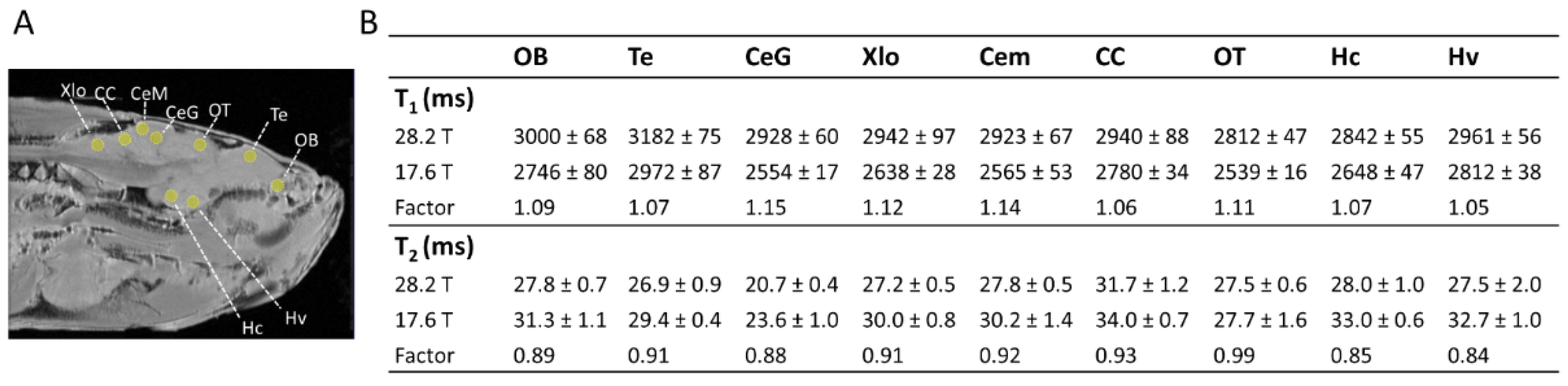
Figure 2.
Comparison of the anatomical images of the zebrafish brain measured at 17.6 T and 28.2 T. [A] Representative RARE images (left column) of the zebrafish brain acquired at 17.6 T and 28.2 T. RARE acquisition details: TR 3000 ms, TE 5.6 ms, 16 averages, resolution 23 μm × 23 μm × 200 μm, RARE factor 4. SNR maps generated from RARE images (right column); [B] RARE images of adult zebrafish brain region in coronal (left) and sagittal (right) view acquired at 28.2 T for structure identification. Acquisition details: TR 3000 s, TE 5.6 ms, 64 averages, resolution 23 μm × 23 μm × 100 μm, RARE factor 4. Identified structures are as follows: 1 – Central zone of dorsal telencephalon area; 2 – Lateral zone of the dorsal telencephalon; 3 – Medial zone of dorsal telencephalon; 4 – Posterior zone of dorsal telencephalon area; 5 – Dorsal habenular nucleus; 6 – Optic tectum; 7 – Tectal ventricle; 8 – Longitudinal torus; 9 – Periventricular gray zone of optic tectum; 10 – Ventrolateral nucleus of semicircular torus; 11 – Medial division of valvula cerebelli, molecular level; 12 - Medial division of valvula cerebelli, granular layer; 13 – Granular eminence; 14 – Cerebellar corpus, granular layer; 15 – Rhombencephalic ventricle; 16 – Caudal lobe of cerebellum; 17 – Medial octavolateralis nucleus; 18 – Facial lobe; 19 – Vagal lobe; 20 – Medial funicular nucleus; 21 – Cereberal corpus, molecular layer; 22 – Medial longitudinal fascicle; 23 – Dorsal sac, 24 – Cerebellar crest; 25 – Diffuse nucleus of the inferior lobe; 26 – Mammillary body; 27 – Caudal zone of periventricular hypothalamus; 28 – Ventral zone of periventricular hypothalamus; 29 – Parvocellular preoptic nucleus, anterior part; 30 – Ventral nucleus of ventral telencephalon area; 31 – Olfactory bulb; 32 – Interpeduncular nucleus.
Figure 2.
Comparison of the anatomical images of the zebrafish brain measured at 17.6 T and 28.2 T. [A] Representative RARE images (left column) of the zebrafish brain acquired at 17.6 T and 28.2 T. RARE acquisition details: TR 3000 ms, TE 5.6 ms, 16 averages, resolution 23 μm × 23 μm × 200 μm, RARE factor 4. SNR maps generated from RARE images (right column); [B] RARE images of adult zebrafish brain region in coronal (left) and sagittal (right) view acquired at 28.2 T for structure identification. Acquisition details: TR 3000 s, TE 5.6 ms, 64 averages, resolution 23 μm × 23 μm × 100 μm, RARE factor 4. Identified structures are as follows: 1 – Central zone of dorsal telencephalon area; 2 – Lateral zone of the dorsal telencephalon; 3 – Medial zone of dorsal telencephalon; 4 – Posterior zone of dorsal telencephalon area; 5 – Dorsal habenular nucleus; 6 – Optic tectum; 7 – Tectal ventricle; 8 – Longitudinal torus; 9 – Periventricular gray zone of optic tectum; 10 – Ventrolateral nucleus of semicircular torus; 11 – Medial division of valvula cerebelli, molecular level; 12 - Medial division of valvula cerebelli, granular layer; 13 – Granular eminence; 14 – Cerebellar corpus, granular layer; 15 – Rhombencephalic ventricle; 16 – Caudal lobe of cerebellum; 17 – Medial octavolateralis nucleus; 18 – Facial lobe; 19 – Vagal lobe; 20 – Medial funicular nucleus; 21 – Cereberal corpus, molecular layer; 22 – Medial longitudinal fascicle; 23 – Dorsal sac, 24 – Cerebellar crest; 25 – Diffuse nucleus of the inferior lobe; 26 – Mammillary body; 27 – Caudal zone of periventricular hypothalamus; 28 – Ventral zone of periventricular hypothalamus; 29 – Parvocellular preoptic nucleus, anterior part; 30 – Ventral nucleus of ventral telencephalon area; 31 – Olfactory bulb; 32 – Interpeduncular nucleus.
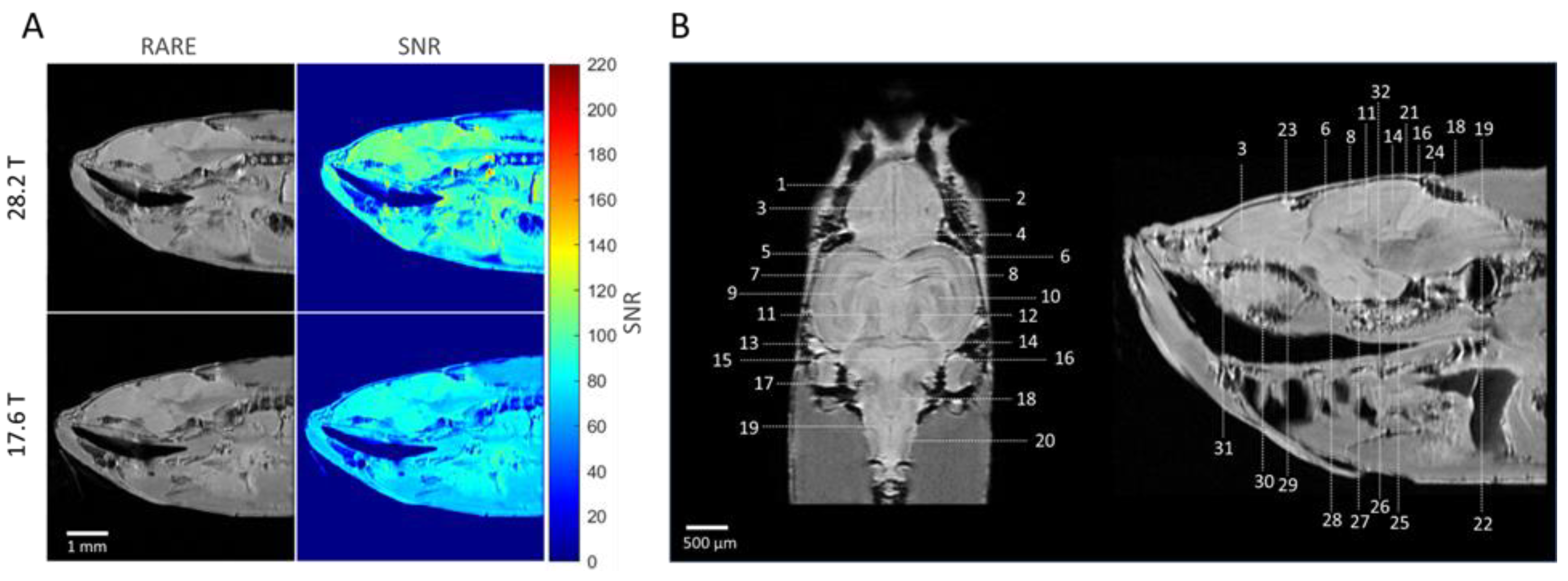
Figure 3.
Diffusion weighted imaging (DWI) of adult zebrafish brain acquired at 28.2 T. [A] A representative sagittal slice of DWI measurement taken in the head of a zebrafish at increasing b-values. Acquisition details: TR 1000 ms, TE 20.2 ms, 4 averages, resolution 23 μm × 23 μm × 200 μm, and effective b-value range 75, 500, 1000, 3500, 5000 and 7500 s/mm2; [B] Apparent Diffusion Coefficient (ADC) map estimated in brain area of zebrafish, showing high contrast between brain and surrounding tissue; [C] ADC map calculated in brain of zebrafish, demonstrating identification of several structures based on diffusion characteristics; 1 – Medial zone of dorsal telencephalon; 2 – Longitudinal torus; 3 – Cerebellar corpus; 4 – Rhombencephalic ventricle; 5 – Ventral rhombencephalic commisure; 6 – Medial longitudinal fascicle; 7 – Diffusive nucleus of the inferior lobe; 8 – Vasular lacuna of area postrema; 9 – ventral zone of periventricular hypothalamus; 10 – Diencephalic ventricle; 11 – Olfactory bulb; [D] ADC values in various brain regions acquired at 28.2 T and 17.6 T. The difference between ADC values measured at two different magnetic field strength is presented as fold change (Factor). Data represent the mean ADC (10-4 mm2 s-1) ± standard error (SE) (Error bars); n=6.
Figure 3.
Diffusion weighted imaging (DWI) of adult zebrafish brain acquired at 28.2 T. [A] A representative sagittal slice of DWI measurement taken in the head of a zebrafish at increasing b-values. Acquisition details: TR 1000 ms, TE 20.2 ms, 4 averages, resolution 23 μm × 23 μm × 200 μm, and effective b-value range 75, 500, 1000, 3500, 5000 and 7500 s/mm2; [B] Apparent Diffusion Coefficient (ADC) map estimated in brain area of zebrafish, showing high contrast between brain and surrounding tissue; [C] ADC map calculated in brain of zebrafish, demonstrating identification of several structures based on diffusion characteristics; 1 – Medial zone of dorsal telencephalon; 2 – Longitudinal torus; 3 – Cerebellar corpus; 4 – Rhombencephalic ventricle; 5 – Ventral rhombencephalic commisure; 6 – Medial longitudinal fascicle; 7 – Diffusive nucleus of the inferior lobe; 8 – Vasular lacuna of area postrema; 9 – ventral zone of periventricular hypothalamus; 10 – Diencephalic ventricle; 11 – Olfactory bulb; [D] ADC values in various brain regions acquired at 28.2 T and 17.6 T. The difference between ADC values measured at two different magnetic field strength is presented as fold change (Factor). Data represent the mean ADC (10-4 mm2 s-1) ± standard error (SE) (Error bars); n=6.
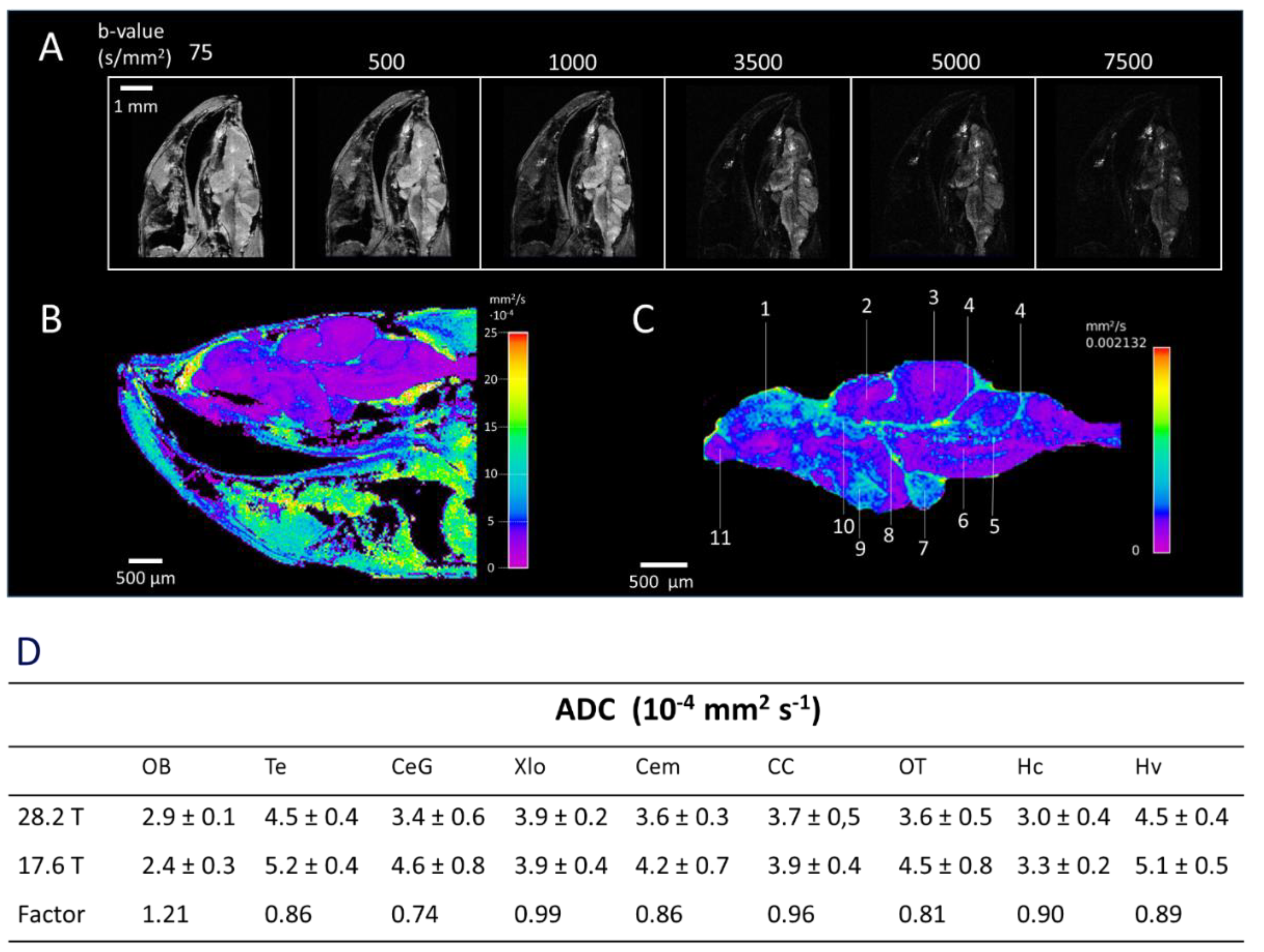
Figure 4.
White matter structure characterisation based on 2D diffusion tensor imaging (DTI) results of adult zebrafish brain acquired at 28.2 T.A representative central slice in sagittal view showing colour maps of [A] axial diffusivity; [B] radial diffusivity, and [C] mean diffusivity calculated from diffusion tensor data. [D] Fractional anisotropy; and [E] diffusion-encoded-colour (DEC) map of the fractional anisotropy (FA). DEC is used to indicate orientation; green – rostral/caudal, red – dorsal/ventral, and blue – medial/lateral. Abbreviation: MLF – the medial longitudinal fascicle; Cans – ansulate commissure; Cpost – posterior commissure, and OT – optic tract.
Figure 4.
White matter structure characterisation based on 2D diffusion tensor imaging (DTI) results of adult zebrafish brain acquired at 28.2 T.A representative central slice in sagittal view showing colour maps of [A] axial diffusivity; [B] radial diffusivity, and [C] mean diffusivity calculated from diffusion tensor data. [D] Fractional anisotropy; and [E] diffusion-encoded-colour (DEC) map of the fractional anisotropy (FA). DEC is used to indicate orientation; green – rostral/caudal, red – dorsal/ventral, and blue – medial/lateral. Abbreviation: MLF – the medial longitudinal fascicle; Cans – ansulate commissure; Cpost – posterior commissure, and OT – optic tract.
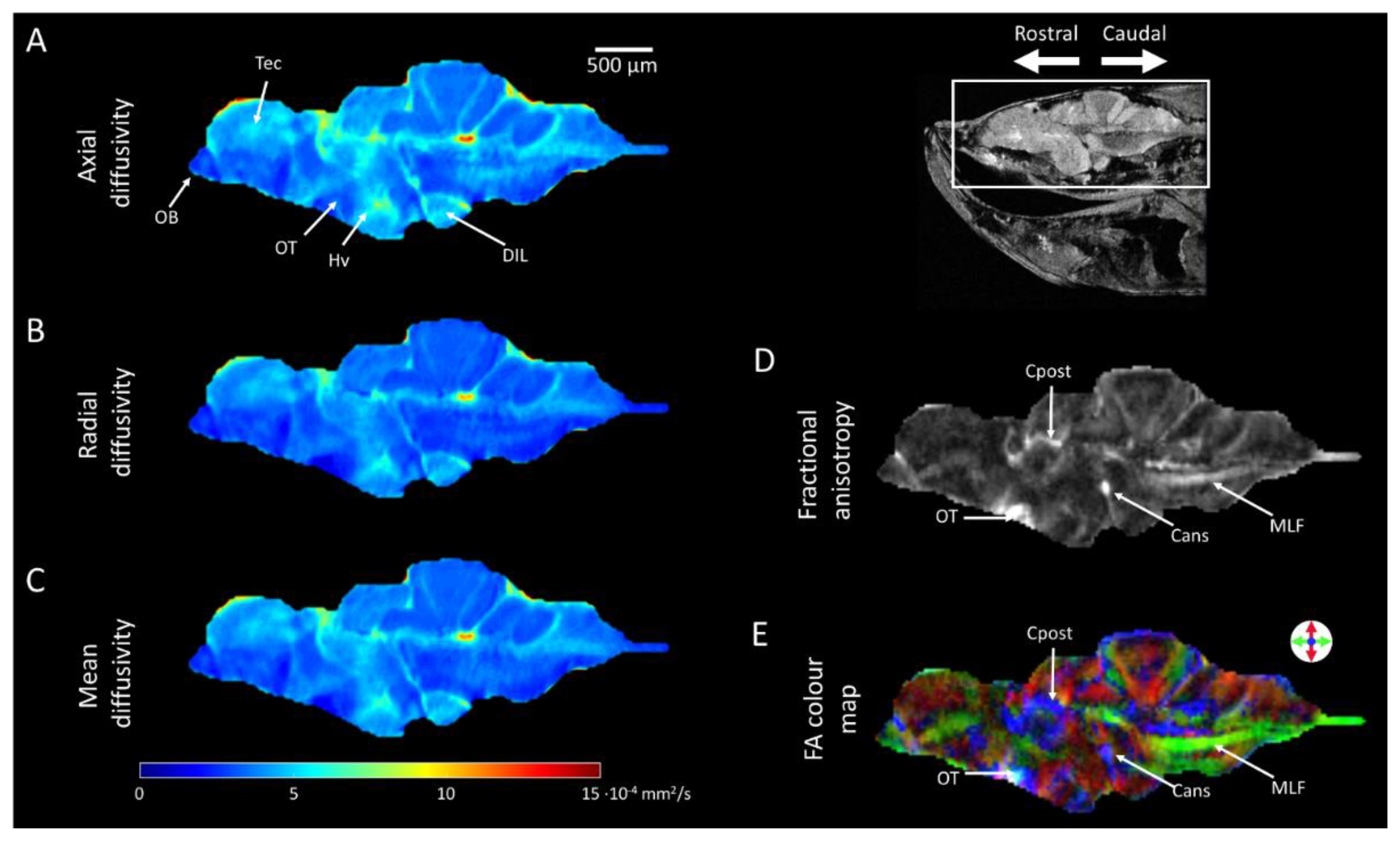
Figure 5.
Whole brain tractography of zebrafish brain. [A] Super-resolution DEC stTDI map calculated from single-shell single-tissue (ssst) CSD using the tournier algorithm [48] to estimate the response function. [B] Super-resolution DEC stTDI map calculated from white-matter response function estimated by multi-shell multi-tissue (msmt) CSD using the dHollander algorithm [35]. Tractography performed with one million streamlines, a minimal length of twice the voxel size (50 μm) and a maximum length of ten times the voxel size (250 μm); [C&D] fODFs located in the Optic Tectum isolated from the 2D stTDI ssst-CSD and msmt-CSD map, respectively. The location of these fODFs is highlighted in [A&B], respectively. DEC is used to indicate orientation; green – rostral/caudal, red – dorsal/ventral, and blue – medial/lateral.
Figure 5.
Whole brain tractography of zebrafish brain. [A] Super-resolution DEC stTDI map calculated from single-shell single-tissue (ssst) CSD using the tournier algorithm [48] to estimate the response function. [B] Super-resolution DEC stTDI map calculated from white-matter response function estimated by multi-shell multi-tissue (msmt) CSD using the dHollander algorithm [35]. Tractography performed with one million streamlines, a minimal length of twice the voxel size (50 μm) and a maximum length of ten times the voxel size (250 μm); [C&D] fODFs located in the Optic Tectum isolated from the 2D stTDI ssst-CSD and msmt-CSD map, respectively. The location of these fODFs is highlighted in [A&B], respectively. DEC is used to indicate orientation; green – rostral/caudal, red – dorsal/ventral, and blue – medial/lateral.
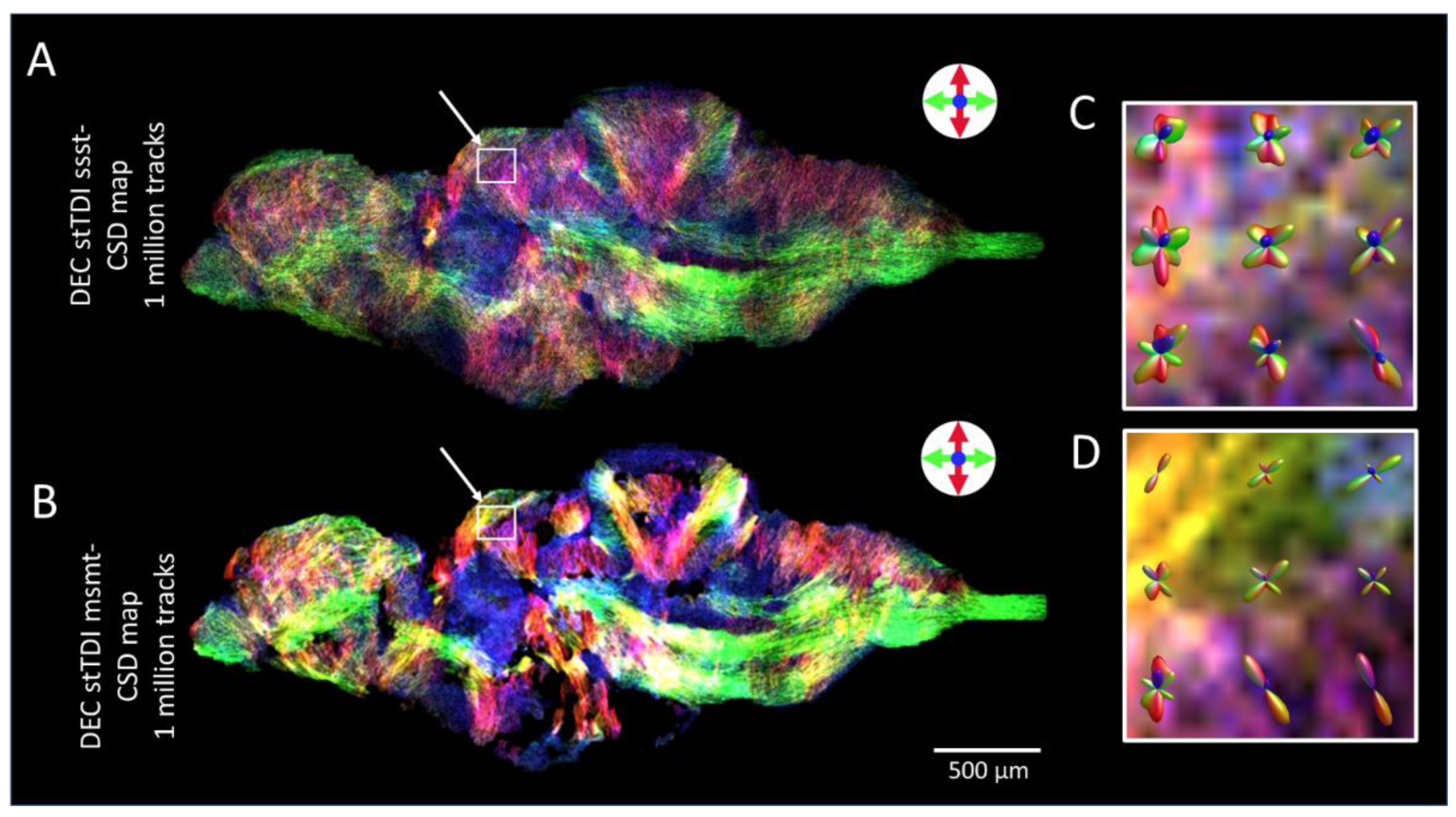
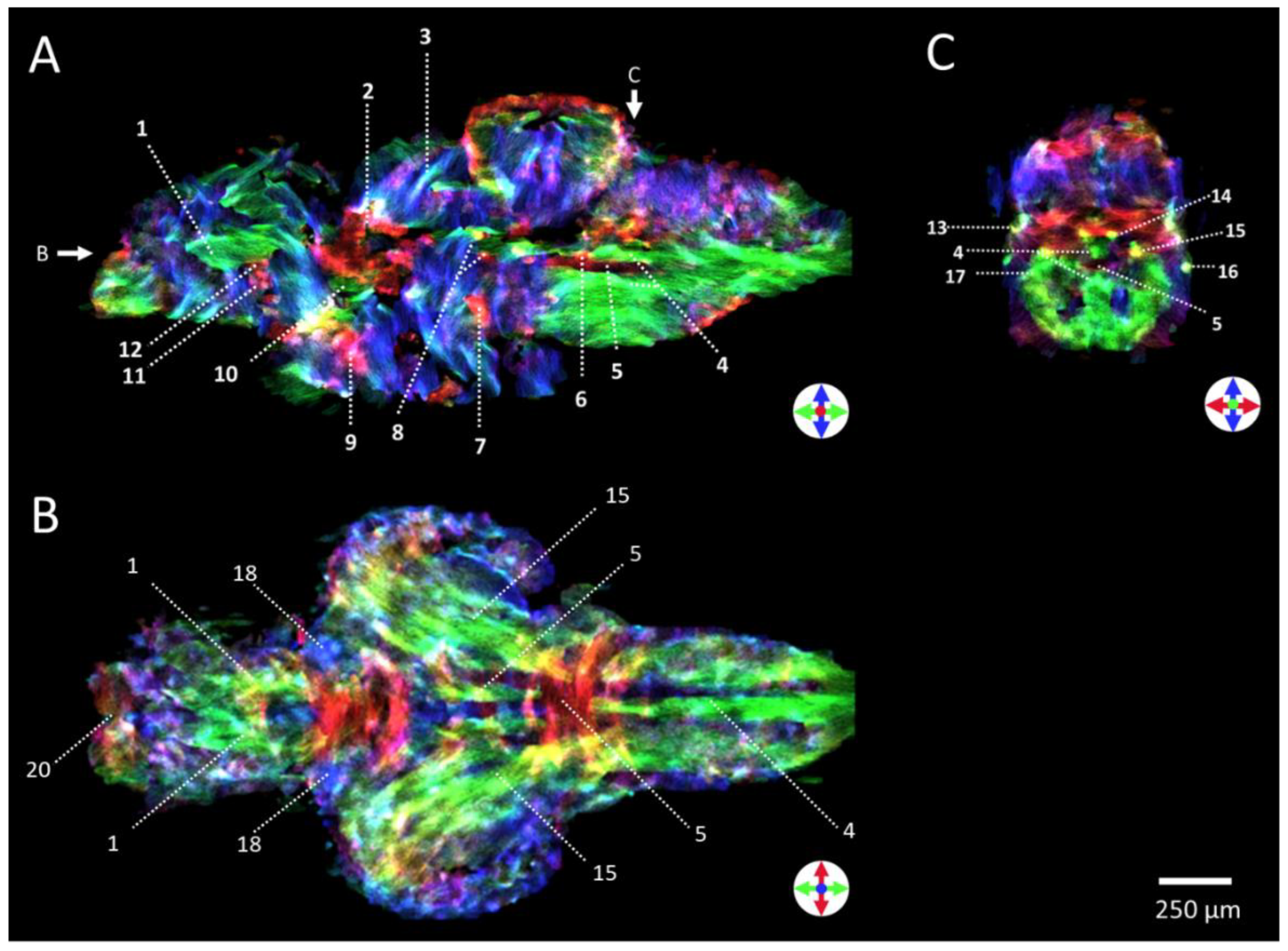
Figure 6.
3D super-resolution DEC stTDI msmt-CSD map of adult zebrafish brain. The DEC stTDI msmt-CSD maps were generated from 3D DTI data measured at 28.2 T. Shown are representative map in sagittal [A]; coronal [B]; and axial view [C]. 3D super-resolution DEC stTDI msmt-CSD map allowed identification of several WM structures in zebrafish brain: 1 (MOT) – Medial olfactory tract; 2 (Cpost) – Posterior commissure; 3 (Ctec) – tectal commissure; 4 (MLF) – Medial longitudinal fascicle; 5 (Cven) – Ventral rhombencephalic commissure; 6 (IAF) – Inner arcuate fibres; 7 (Cans) – Ansulate commissure; 8 (DiV) – Diencephalic ventricle; 9 (Cpop) – Postoptic commissure; 10 (MFB) – Medial forebrain bundle; 11 (CantV) – Anterior commissure; 12 (CantD) – Anterior commissure; 13 (PLLN) - Posterior lateral line nerve; 14 (VIIs) – Sensory root of the facial nerve; 15 (LLF) – Lateral longitudinal fascicle; 16 (ALLN) – Anterior lateral line nerves; 17 (SGT) – Secondary gustatory tract; 18 (DOT) – dorsomedial optic tract; 19 (Cgus) – Commissure of the secondary gustatory nuclei ; 20 (GL) – Glomerular layer of olfactory bulb. DEC is used to indicate orientation; green – rostral/caudal, blue – dorsal/ventral, and red – medial/lateral.
Figure 6.
3D super-resolution DEC stTDI msmt-CSD map of adult zebrafish brain. The DEC stTDI msmt-CSD maps were generated from 3D DTI data measured at 28.2 T. Shown are representative map in sagittal [A]; coronal [B]; and axial view [C]. 3D super-resolution DEC stTDI msmt-CSD map allowed identification of several WM structures in zebrafish brain: 1 (MOT) – Medial olfactory tract; 2 (Cpost) – Posterior commissure; 3 (Ctec) – tectal commissure; 4 (MLF) – Medial longitudinal fascicle; 5 (Cven) – Ventral rhombencephalic commissure; 6 (IAF) – Inner arcuate fibres; 7 (Cans) – Ansulate commissure; 8 (DiV) – Diencephalic ventricle; 9 (Cpop) – Postoptic commissure; 10 (MFB) – Medial forebrain bundle; 11 (CantV) – Anterior commissure; 12 (CantD) – Anterior commissure; 13 (PLLN) - Posterior lateral line nerve; 14 (VIIs) – Sensory root of the facial nerve; 15 (LLF) – Lateral longitudinal fascicle; 16 (ALLN) – Anterior lateral line nerves; 17 (SGT) – Secondary gustatory tract; 18 (DOT) – dorsomedial optic tract; 19 (Cgus) – Commissure of the secondary gustatory nuclei ; 20 (GL) – Glomerular layer of olfactory bulb. DEC is used to indicate orientation; green – rostral/caudal, blue – dorsal/ventral, and red – medial/lateral.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



