
Submitted:
26 August 2024
Posted:
28 August 2024
You are already at the latest version
Abstract
White oak mortality is a significant concern in forest ecosystems due to its impact on biodiversity and ecosystem functions. Understanding the factors influencing white oak mortality is crucial for effective forest management and conservation efforts. In this study, we aimed to investigate the spatial pattern of white oak mortality rates at different locations of the eastern US and explore the underlying processes involved with the spatial distribution patterns. Multicycle Forest Inventory and Analysis data were compiled to capture white oak plots across the eastern US. White oak mortality data were collected across plot systems that utilized Diameter at Breast Height between two periods. Ripley’s K function was used to study the spatial pattern of white oak mortality rate. Results found clustered patterns at both local and broad scales and random patterns at broad scales only across the southern and northern regions. However, the central region indicated both clustered and random patterns at local scale only. We found uniform patterns at a broad scale across all regions. Our findings demonstrated the importance of the spatial pattern of the white oak mortality rate and the underlying factors associated with it. This research can inform forest management practices for the conservation of white oak populations at varying scales. Future research is needed for a comprehensive assessment of biotic and abiotic factors for forest management strategies at various spatial scales aimed at mitigating white oak mortality.
Keywords:
white oak mortality
; spatial patterns
; clustered
; random
; uniform
1. Introduction
White oak (Quercus alba L.) is an important tree species both for ecological significance and economic importance across the eastern United States. White oaks are dominating canopy that serves a vital role in forming stand structures and composition. For a wide variety of creatures, including nesting birds and arboreal animals, its wide, spreading crown provides a friendly microhabitat [1,2]. Acorns and fallen leaves provide a wealth of food for wildlife, supporting complex food chains and promoting biodiversity [3,4]. Furthermore, white oaks' vast root systems create essential symbiotic connections with mycorrhizal fungi that aid in nutrient cycling and uptake. The tree's capacity to store large amounts of carbon helps to reduce greenhouse gas emissions and mitigate the effects of climate change [5,6]. The white oak's vital function in the forest ecosystems of the eastern United States is further demonstrated by its patterns of succession and regeneration. Because of its effective acorn dispersal, the tree offers a plentiful supply of seeds for spontaneous growth, guaranteeing the continuation of its lineage for future generations. However, white oaks population is in danger due to the increased declining situation associated with changes in forest stand dynamics.
Oak mortality is among the most observed phenomena in oak forests in the Eastern U.S. [7,8,9,10]. The mortality starts with the oak tree browning of leaf, turning black, curl-up, and finally falling to the ground [11,12,13,14,15]. Factors responsible for the oak decline are suspected long-term predisposing, short-term inciting, and contributing factors [12,13]. Predisposing factors are related to stand longevity and maturity, which are responsible for a tree's natural ability to growth inhibition and lead to injury-inducing agents. The inciting factors are related to physical or biological conditions, which are related to defoliating insects, hail, frost, and drought. Reports from inciting fac-tors showed typical crown dieback, browning, and new leaf emergence in dying trees, which eventually leads to deaths [16]. The contributing factors are related to pathogenic fungi and boring insects that ultimately kill the trees. This decline has grappled with notable drought outbreaks, late spring frost, the emergence of saprophytic fungus due to climate change [17], and oak borer attacks on the most vulnerable sites.
Typically, oak mortality has been targeted in red oak group species. More recently, white oak mortality (WOM) has become a prominent target across the eastern US. It seems that the spatial distribution pattern of WOM is not uniform. Poor resource sites [18] such as droughty, poor drainage, and soil nutrient deficiency are more prone to WOM. It is because low resources have led to declining and widespread regeneration failure. Scientists also reported WOM in higher-quality mesic sites where the forest has gone through a high stand density and maturity stage [19,20]. The mortality is more prevalent in the self-thinning stage, as the tree species under high stand density struggle to utilize maximum resources [21]. WOM has been observed in different topographies, from low-lying lands to valley floors [22]. North-facing slopes where sunlight is low are more prone to high oak mortality [23]. Besides, ecological stressors such as browsing, heavy shade, and disturbances have influenced white oaks in many parts of the eastern hardwood forests [22].
The impacts of white oak mortality are not only limited to individual tree species but also affect entire forest ecosystems and ecological processes. For instance, white oaks were dying in greater numbers, i.e., 30% of healthy crowns, which were less than 4 meter in width across the 516-ha area of Ozark Highlands [24]. Similarly, some 900-ha area of Baskett Wildlife Research and Education Center across the Ozark Border of Central Missouri depicted 10% of white oaks killed by drought-pathogen interactions [25]. White oak mortality across forest ecosystems can lead to changes in nutrient cycling, water regulation, and carbon sequestration [26,27,28]. White oak is a long-lived species and can sequester large amounts of carbon during its lifetime [29,30]. The decline of mature white oaks can disrupt carbon sinks and worsen the situation for climate change and carbon management [31,32]. White oaks vast canopy offers critical habitat and nutrition for a wide range of flora and fauna, promoting a complex web of biological interactions [33,34]. For instance, white oak is a dominant canopy species across the eastern US that shapes forest structure and composition influencing soil processes, species interactions, and wildlife habitat [35,36]. Its loss can lead to a change in forest dynamics such as impacting the recruitment of new individuals to alterations of the competitive relationship among plant species [37,38]. The decline in white oaks can impact numerous wildlife species and their microhabitats that rely upon shelter and foraging [39]. Moreover, the white oak decline can have adverse effects on species distribution and food chain interactions as well [40].
Several research concentrated on the spatial patterns of white oaks that utilized climate, topography, and several other factors combined as well as individually [41,42]. There is very little research done on white oak’s spatial patterns but those are focused on local level studies. The evidence did not find clear spatial patterns for white oaks across varying scales [43,44], which highlights the unique approach for our study. The aim of this study is (1) to identify the spatial distribution pattern of WOM across southern, central, and northern region of the eastern U.S. and (2) and explore the potential underlying processes behind the observed spatial patterns of WOM rate. We hypothesized that the observed spatial pattern of the WOM rate is randomly distributed at local scale due to localized environmental factors and stand-scale competition, while exhibiting clustering patterns at the broad scale, likely attributed to a larger-scale environmental gradients and climatic pattern.
2. Materials and Methods
2.1. Study Area
We selected the eastern United States as our study area, which predominantly covers oak forests [45]. It is comprised of ten states that cover a land area of 1,273,420,251.33 hectares (Figure 1). Our study area was further divided into southern region (low latitude: 30-to-36-degree N latitude), central region (medium latitude: 36-to-37.5-degree N latitude), and northern region (high latitude: 37.5-to-42.5-degree N latitude) WOM rate that contains 678, 747, and 797 plots, respectively. We did a subdivision to our study area due to regional variation as southern region differs from northern region and central region in terms of precipitation patterns and temperature. This divide enables us to capture variations in environmental factors that may affect tree mortality rates. Moreover, dividing the research area into southern, central, and northern regions allows us to investigate how ecological gradients affect WOM rates. For instance, latitudinal variations in soil types, topography, and disturbance regimes can have an impact on WOM mortality and forest health.
Our study area consists of varied tree species, and a wide range of climate, soil, and terrains [46,47,48]. The land structure in the eastern part of the study area is significantly Appalachian plateaus with low mountains, narrow valleys, and sharp edges. On the western portion, the area varies from highly flat central till plains to interior low plateau along with Ozark highlands. The area consists of a variety of ecological regions such as the Ouachita Mountains of Arkansas to the Northern Cumberland Mountains in West Virginia. In the south, central till plains Oak-Hickory of Illinois to Interior Low Plateau Highland Riff of Alabama.
The overall climate in these regions is long, hot summers and cool winters. The mean annual temperature ranges from 4 degrees to 18 degrees Celsius in the east-west gradient with warmer temperatures in the south. The precipitation ranges from 500 mm in the north-west to 1,650 mm in the southeast. Some Appalachian Mountains precipitation goes up to 2,000 mm during spring and fall. The soil types are mostly mollisols, inceptisols, alfisol, and ultisols. These areas consist of a xeric gradient characterized by thin, rocky soils, exposed south, southwest, and wet slopes [48]. Our study area was historically dominated by oaks, hickories, and pines. Today, most of the forest areas have been replaced by agriculture and rapid urbanization [47,49]. In our study area, there are deciduous types of forests such as oaks (Quercus spp.), hickory (Carya spp.), american beech (Fagus grandifolia), ash (Fraxinus spp.), and maple (Acer spp.).
Figure 1.
Study area showing forest covers across the eastern US.

2.2. Data Acquisition and Processing
2.2.1. Forest Inventory and Analysis Program
FIA program has been monitoring national forest resources across all ownerships in the US [50,51]. We used inventory data that were collected and processed on a relational data based on categorization into multiple phases. The relational database structure facilitates the integration of data from a variety of sources including historical records, satellite imaging, and remote sensing, which adds depth and diversity to the forest inventory dataset. This approach helped us for efficient data querying which is required for the development of reports, analysis, and summary statistics [50]. Our data has been tabulated based on relational database, which is organized, accessible, and consistent to support evidence-based decision-making and long-term forest management planning, and it can handle extensive and interdependent data link that are preferred for managing forest inventory data. For instance, stratified estimations were used to estimate population parameters for most of the variables depending on the scale and level of information [52]. We used inventoried data because it has an advantage over other databases as the plots are evenly distributed without any geospatial bias [53]. In our study area i.e., eastern US, FIA plots are recorded with at least one forested condition, which is remeasured again every five years. Each plot measures key attributes of all tree species including plots, surveyed years, and others [54]. Our plots were based on phase 2 and phase 3 ground plots in which every plot sample was measured with a fixed radius.
We used condition-level information from the sampled data of FIA. A condition was based on changes in land use or vegetation that fall inside a plot. Likewise, a condition has been determined by the ten percent crown or canopy cover of live tally trees in the forested condition [55]. We used condition-level data which has been efficiently designed in such a way that arbitrary numbers were assigned and defined using discrete variables such as forest type, stand size, species composition, stand structure, stand origin, ownership group, and disturbance history [55].
Our FIA periodic and annual inventory was recorded approximately every ten and five years, respectively. But both periodic and annual inventories contained differences in plot design, sampling, and measurement protocols. Therefore, it is not possible to combine both periodic and annual data with dissimilar samples. In this condition, we processed our data only from annualized survey years for efficient data collection relating to white oaks. For instance, our farthest annual data goes back to 1998 from Virginia, which is the oldest annual inventory among ten states. And most of the plot inventories of hardwood forests, especially white oaks, were fully recorded until 2019. Hence, a decision has been made to choose our timeframe from 1998 to 2019 to investigate WOM spatial patterns across the different locations of the eastern US.
2.2.2. Variables Selection for Live and Declining White Oaks
Our plot data were acquired from USDA Forest Service DataMart that had already been assigned to a stratum by forest inventory program across all states. A stratum refers to a set of plots that have similar classifications captured from remotely sensed imagery. Within the estimation unit of plot sampling, the weight of the stratum is based on the proportion of the stratum [50,55]. In the same set of our plot data, geographic coordinates (latitudes and longitudes) were recorded that captured a 0.40 ha (1 acre) sample area but not for all trees. We utilized plots from FIA that were located within one mile of the original location. It is because landowners prefer not to disclose their land or property publicly due to security reasons associated with it [55,56].
We processed our data from FIA that covered only white oak (Quercus alba L.). The reason behind this is the dearth of study on the regional pattern analysis of white oak in the eastern United States in particular. There were numerous oak species in the sample plot as well, but we limited our selection to plots with white oak trees, and we conducted our analysis from the oldest survey year 1998 up through 2019. The white oak tree data were extracted from USDA Forest Service in which we utilized attributes such as inventory year (INVYR) as surveyed year, county code (COUNTYCD) refers to a particular county number for a particular state, unique plot number (PLOT) assigned to each white oak, species code (SPCD) representing particular tree species, tree status code (STATUSCD) representing live tree (1) assigned by FIA, trees per acre unadjusted (TPA_UNADJ) refers to the number of trees per acre that the sample tree theoretically represents based on the sample design, cycles (CYCLE) refers to a number assigned to a set of plots, measured over a particular time, and current diameter (DIA) refers to diameter at breast height i.e., 1.37 meter (4.5 feet) above the ground line of the sample tree.
2.3. Data Processing and Analysis
2.3.1. Selection of Live and Declining White Oak Plots
Annualized data from 1998 to 2019 were processed to locate white oak plots (Table 1). After data processing and refinement of our variables i.e., plot, forest condition, and tree, we selected a plot to narrow our criteria i.e., locating white oaks' plots across accessible forest land. For instance, Missouri contained plot information as a unique plot number related to each county and cycle, and locations (latitudes and longitudes) with surveyed years. We utilized sampled plots that contained at least one accessible forest land condition on the plot. Most importantly, the condition table consisted of similar attributes except for locations that were useful to associated with plots. During this process, we narrowed our condition and tree data with similar criteria i.e., selecting accessible forest land only. We used other important attributes from tree data such as unique plot number, county code, tree species code, diameter, trees per acre, and cycle to identify all white oak trees.
We selected white oak trees identified with species code (e.g., 802) from the tree data. From the same tree data containing white oaks only, we determined the basal area i.e., Basal area of white oak = 0.005454 *(Current Diameter)2 * trees per acre unadjusted. Then, we obtained our information combining forest condition and tree level data based on unique plot numbers. Then, we extracted white oak basal area information from the same set of combined tree and forested condition data. Since we worked only with an annual inventory that clearly limited our timeframe for data processing to each state. For instance, we analyzed our Missouri white oak data in which cycle 5 consisted of survey years grouped from 1999 to 2003 (usually year-wise data). Similarly, other cycles had a set of survey years that went until 2019 which were grouped as cycle 6 (2004 to 2008), cycle 7 (2009 to 2013), and cycle 8 (2014 to 2019). We then grouped other states data with the same process and extracted white oaks from each cycle. To get white oaks plot within a given timeframe, we created an appropriate formula using county and plot number i.e., county code + (plot)/100,000 to connect one cycle to another. With all these processes, we identified all the declining plots of white oaks based on basal area change. We repeated the similar process while working with rest of the states from our study area connecting different cycles to get declining plots for white oaks. During this process, we only utilized the basal area from a single plot so that we could avoid plot repetition. Based on basal areas, we extracted 2,220 declining white oak plots from 7,405 live white oak plots across ten states. For all these data processing steps, we used ArcMap version 10.8.1.
After we finalized our WOM rate data, we further divided our study area into southern, central, and northern region WOM rates. By dividing the study area, we can unravel the complex interplay of ecological, climatic, and geographical factors influencing WOM, laying the groundwork for understanding and regulating spatial patterns at different scales and regions. Moreover, a regional division enables direct comparisons of our areas that identifies similarities and differences in WOM patterns, revealing whether certain spatial trends are consistent across regions or region-specific.

Table 2.
Ten states with inventory years, cycles, number of live white oak (WO) plots, and declining WO plots.
Table 2.
Ten states with inventory years, cycles, number of live white oak (WO) plots, and declining WO plots.
| State | Inventory Years | Cycles | Live WO plots | Declining WO plots |
|---|---|---|---|---|
| Alabama | 2001-2019 | 8,9, &10 | 716 | 166 |
| Arkansas | 2000-2019 | 8,9,10, &11 | 1,158 | 400 |
| Illinois | 2001-2019 | 5,6,7, & 8 | 244 | 63 |
| Indiana | 1999-2019 | 5,6,7, & 8 | 206 | 43 |
| Kentucky | 2000-2017 | 5,6,7, & 8 | 775 | 207 |
| Missouri | 1999-2019 | 5,6,7, & 8 | 1,544 | 527 |
| Ohio | 2001-2019 | 5,6,7, & 8 | 337 | 67 |
| Tennessee | 2000-2017 | 7,8,9, &10 | 909 | 305 |
| Virginia | 1998-2019 | 7,8,9,10, &11 | 855 | 314 |
| West Virginia | 2004-2019 | 6,7, & 8 | 661 | 128 |
| Total | 7,405 | 2,220 |
2.3.2. White Oak Mortality Rate Analysis
The mortality rate was calculated by using basal areas of surveyed cycles, which were grouped into five years. Some of the white oak plots were remeasured in the period from 1998 to 2019. Hence, we omitted those repeated plots to have accurate results. However, we summed up all those remeasured plots' basal areas to obtain the true basal area. We selected all the basal areas from declining plots of white oaks concerning surveyed years. The declining plots were chosen based on basal area change. Basal area change refers to the decrease in basal area from two consecutive cycles. For instance, we calculated basal area change using cycles as:
We did the same with all remaining cycles and chose decreased basal areas that directly referred to the declining white oak plots. We calculated the mortality rate in terms of the reduced basal area of white oaks divided by the differences in the surveyed year between two extreme cycles. All those declining plots representing mortality rates were extracted and compiled the results. We calculated the mortality rate as:
2.3.3. Spatial Distribution Analysis of WOM by Univariate Ripley’s K Function
Our data for white oak mortality was based on specific point events. Ripley’s K function was used for a comparison of observed spatial patterns against those expected under complete spatial randomness (CSR) that provides information for clustering, random, and uniform [60]. Other patterns do not justify the specific patterns such as CSR. Another reason is that edge effects are ignored and significantly impact other patterns that bring biased results [61,62]. However, the K function incorporates edge effects and produces valid results mitigating these issues. The use of Ripley’s K function highlights the visualization of spatial patterns at various scales i.e., small scales, larger scales, and their statistical significance whereas other patterns only highlight higher or lower concentrations without indicating particular patterns and statistical significance at varying scales.
We used Multi-Distance Spatial Cluster Analysis-Ripley’s K function tool [63], which uses nearest neighbor distance to process spatial distribution patterns. ArcMap version 10.8.1 was used for Ripley's K function which defines whether our spatial pattern is cluster, random, or uniform on each analysis scale [64,65]. All 2,220 WOM points from 1998 to 2019 were compiled that contained point locations i.e., longitudes and latitudes. In Ripley's K function tool, the initial distance was 50 km, and the increment distance was 10 km. Edge correction was done by using Ripley's edge correction formula [63], which automatically counts and measures points inside the study area. To test our null hypothesis of CSR, we formed upper and lower confidence envelopes of 99.9% generated from randomizations of 999 plot points using the default random generator in ArcGIS. We examined whether our theoretical curve (Ktheo) deviates from CSR or not. If the observed curve (Kobs) is above the theoretical curve and upper envelope, then there is spatial clustering. If it is below the theoretical curve and lower envelope, then WOM points follow spatial dispersion (uniform). There is also a greater chance that both observed and theoretical curves could line up denoting complete spatial randomness or random pattern. We used the transformed K function (L(d)) to represent the graphical interpretation of Ripley's K function.
3. Results
3.1. White Oak Mortality Spatial Distribution Patterns
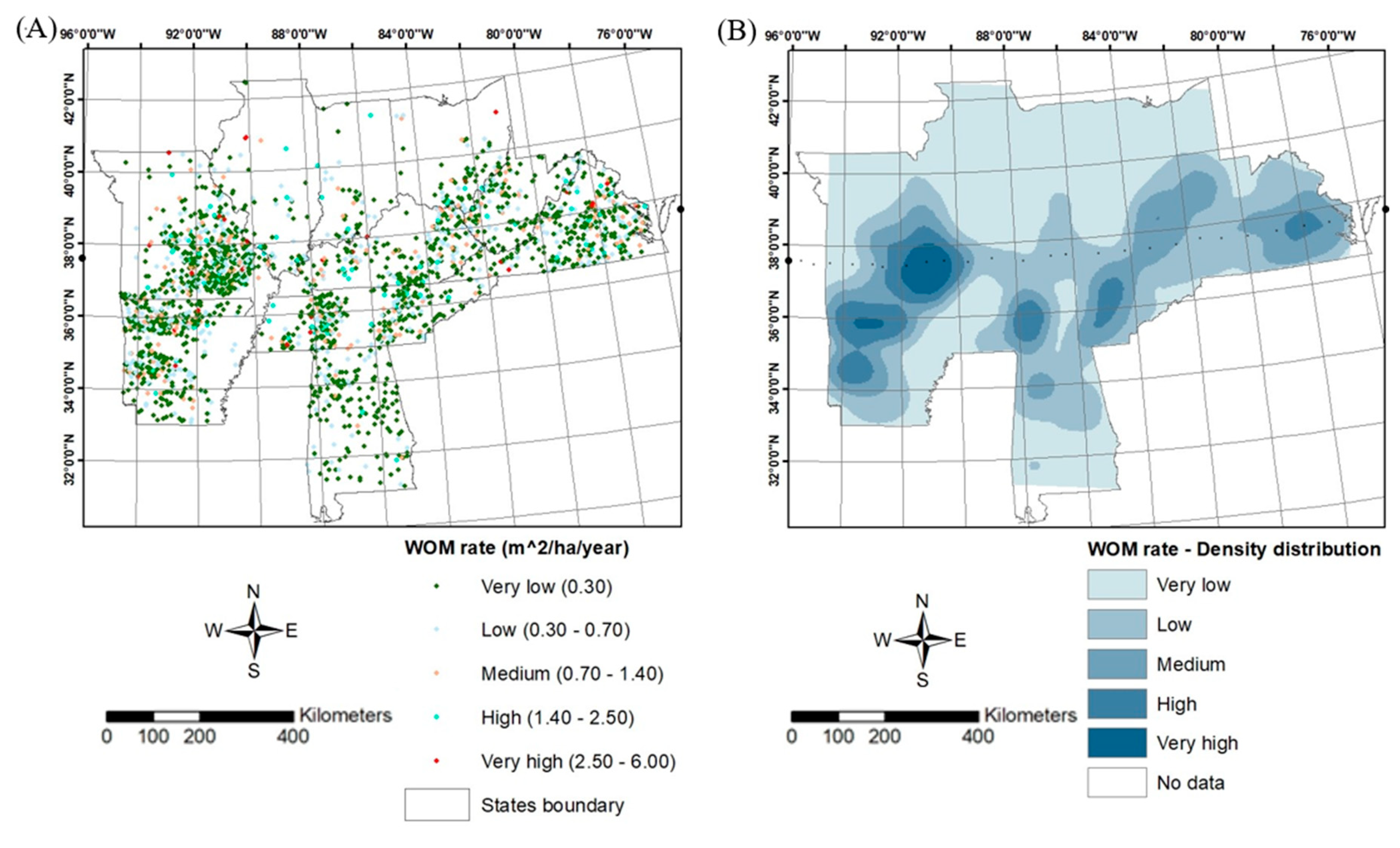
The white oak mortality (WOM) rate is spread throughout the eastern United States. However, there are differences in the magnitude of mortality rate and density distribution across different locations i.e., the southern, central, and northern regions of WOM rate. The southern region consists mostly of very low followed by low and medium WOM rates across different latitudes and longitudes (Figure 2A). However, there are also a few plots of high as well as very high mortality rates across this region. Despite the larger area of a low and very low density of WOM rate, some locations across the southern region depicted a high-density distribution of WOM rate (e.g., parts of Arkansas and Tennessee; Figure 2B).
Numerous plots of WOM rate were gathered that has been spread across most of the central region of the eastern US. Much of the central region showed very low and low WOM rates, however, there are also numerous plots related to medium, high, and very high WOM rates across this region (Figure 2A). This region has the most diversified forest landscape that consists of very high-density (e.g., parts of Missouri, Arkansas, Virginia etc.) and high-density distribution (e.g., parts of Missouri, Arkansas, Tennessee, Kentucky, and Virginia) of WOM rate as compared to southern and northern regions. Most of this region has been depicted with very low density as well as low and medium density distribution of WOM rate, which is greater than very high and high-density distribution (Figure 2B).
The majority of the northern region contains very few plots of WOM rate distributed across our study area (e.g., Illinois, Indiana, and Ohio). Though this region has very few plots of high to very high WOM rate (e.g., Missouri, west Virginia, and Virginia), the overall plot distribution to WOM rate is higher as compared to other regions (Figure 2A). The density distribution of WOM rate is mostly depicted very low and consists of a few areas of medium, high, and very high-density distribution to certain latitudes and longitudes. Despite the larger area of land mass, this region represented a very low plot distribution of WOM rate (Figure 2B).
The spatial distribution pattern of the WOM rate showed random patterns up to 3000 m, as the observed K value was inside the confidence envelopes, and we accept the null hypothesis (Figure 2). Beyond this point, we reject our null hypothesis, the spatial distribution of WOM rate showed clustered until 20,000 m is statistically significant. It is due to the spatial clustering of WOM rates, as indicated by the observed K value being significantly larger than the expected K function and falling outside the confidence envelopes. This clustering pattern of the WOM rate increases as the distance increases. Most of our pattern analysis for the WOM rate depicted clustered (non-random) patterns at a larger distance across the eastern US. Thus, our spatial distribution pattern analysis of WOM showed more clustering patterns than the random pattern as distance increases.
The kernel density maps for white oaks showed the cluster spatial distribution across the declining plots of white oaks (Figure 2). This density distribution indicated areas for higher or lower concentrations across the eastern US.
Figure 2.
Plots (A) and density distribution (B) showing WOM rate across different latitudes and longitudes.
Figure 2.
Plots (A) and density distribution (B) showing WOM rate across different latitudes and longitudes.
3.2. Spatial Distribution Patterns at Southern, Central, and Northern Regions WOM Rate
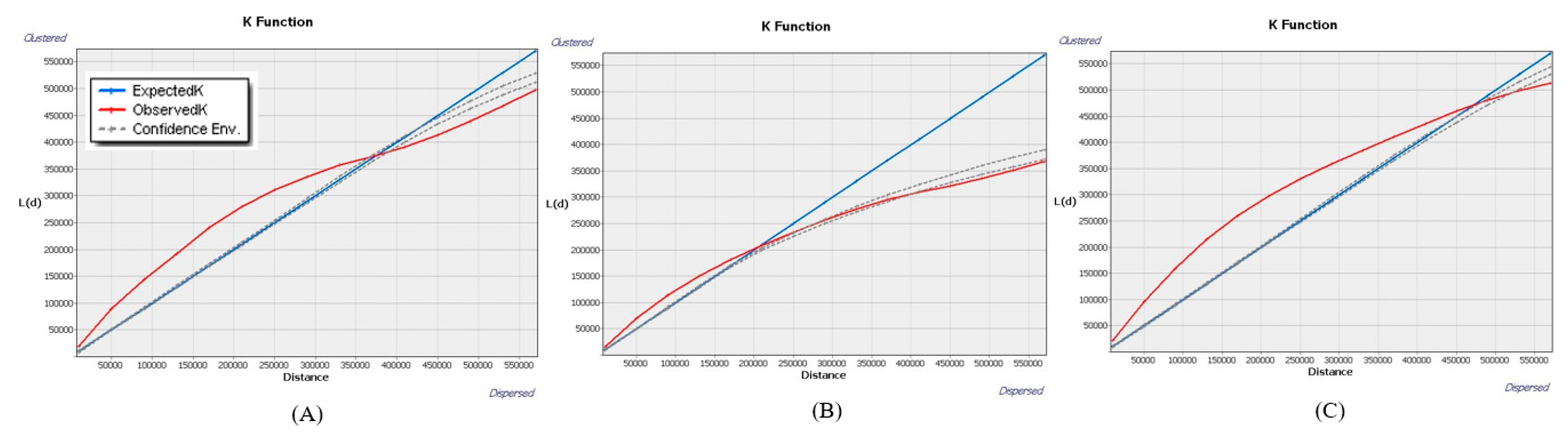
The spatial pattern of the WOM rate across the southern region depicted a clustering pattern until about 360 km, and we reject our null hypothesis. This is because the observed K function is above the expected K function. Also, the observed K function, statistically significant, is completely outside the envelope. This means points of WOM rate in the dataset are more closely spaced than would be expected under spatial randomness. However, the intensity of the clustering pattern, not constant, keeps increasing as the distance increases. Most of the southern region depicted aggregation to WOM rate across the local scale. We found the pattern is random at 360 km to 395 km as the K function, statistically not significant, is completely inside the envelopes. The random pattern of the WOM rate is across a broad scale but covers a low range of distance. This random pattern shows that the point of WOM rate has an equal chance of occurring anywhere in the southern region of our study area. The pattern is mostly uniform as the distance increases from 400km to 550km, and we reject our null hypothesis. It is because the observed K function across a broad scale is consistently below the envelope and is statistically significant as well. It shows that the points of the WOM rate are more regularly spaced than would be expected under spatial randomness. Also, there is a mostly uniform pattern across the larger scale which is more or less similar to the central and northern region WOM rate.
The central region of the WOM rate depicted a slightly lower intensity of observed K function at the local scale in comparison with patterns of southern and central region WOM rates. We found the clustering pattern until 200km across the central region where the intensity of clustering is much smaller, and we reject our null hypothesis. This clustering is because the observed K function, statistically significant, is outside the envelope. This indicates that there is a tendency for WOM rate points to be aggregated together in this region. Unlike other regions, the observed K function is much near to the random line at the local scale. Similarly, there is also a random pattern from 230km to 430 km as observed K function, statistically significant, is completely inside the envelope. It suggests that there is an equal chance of occurring points of WOM mostly at the medium scale than the local scale. However, broad scale is mostly facilitated with a uniform pattern starting from 430 km and beyond, and we reject our null hypothesis. And this uniform pattern is much away from the random line and contains a slightly lower intensity of observed K function in comparison to patterns of southern and northern region WOM rate. Overall, the pattern of WOM rate seems to be more random at the local scale, but at broad scale, it is less uniform, which is similar to southern region of the WOM rate as well.
The WOM rate across the higher latitude or northern region presented more clustering patterns (until 480km) than a random and uniform pattern. We found a complete clustering pattern across local, medium, and parts of broad scales in which the observed K function, statistically significant, is outside the envelope. The clustering pattern at the local scale starts with low intensity which is also much near the random line and slightly at the peak when it reaches around 20 km. This indicates that there is a tendency for WOM rate points to be aggregated or clustered together in this region. However, clustering slightly decreases as it reaches to medium scale and stops at 470km, which is at a broad scale. Unlike the southern and central WOM rates, patterns presented by the observed K function are varied across each scale. It is because random and uniform patterns are presented only across broad scales and across a few ranges of distance. Results show that the observed K function is below the expected K function at 480km to 520km, completely inside the envelope, denoting a random pattern at a broad scale. This tells us that the points are distributed in a manner consistent with complete spatial randomness (CSR). Beyond 520km, there is a uniform pattern at a broad scale but covers few distances in this region. The uniform pattern is since the observed K function is completely outside the envelope and is statistically significant as well, and we reject null hypothesis. This tells us that points are more evenly distributed across this region, and there is a tendency for points to avoid each other.
Figure 3.
Ripley’s K function showing various spatial patterns across (A) southern, (B) central, and (C) northern region WOM rates.
Figure 3.
Ripley’s K function showing various spatial patterns across (A) southern, (B) central, and (C) northern region WOM rates.
4. Discussion
Our study found that the southern region WOM rate showed a clustered pattern at the local scale, and the reason is that competition at stand level may play some dominant roles over the WOM rate. This confirms with other findings that aggregation factors were highly responsible for stand-level competition in most of the oak tree species [64,66]. Evidence supports that self-thinning, which is also called natural mortality, is mostly prevalent in the oak-dominated forest of eastern North America [67,68]. Self-thinning occurring mostly in the stem exclusion stage if deprived of sunlight, moisture, and space may lead to mortality. Also, the spatial pattern of the southern region WOM rate indicated a random pattern across a few ranges of broad scale. This random pattern is the indication of WOM occurring at regional sites including poor and good resource sites as well as various topographic and hydrological conditions [64,69,70]. This random pattern of WOM rate may be due to extended periods of drought. For instance, [44] reported that there was an extreme drought in which decline in white oaks increased from 6% (1999-2005) to 15% (2006-2010) across Ozark highlands of Arkansas and Missouri. Others found that the random pattern of WOM rate is also attributed to severe weather events such as storms, hurricanes, and droughts [71,72]. [11] and [73] documented that tropical winds followed by hurricanes killed randomly to many white oaks including other hardwood tree species and that may have resulted in a random pattern to WOM. Like the central and northern regions, our results reported that most of the southern region WOM rate had a significant uniform pattern on a broad scale. It is because the mortality in the mature stands of white oaks is not replacing themselves and would eventually generate a uniform pattern [74]. Other reasons leading to a uniform pattern might be due to uniformity in drought or flooding and homogenous soil conditions in the region [75,76].
The observed clustered pattern of WOM rate in the central region raises intriguing queries concerning the underlying mechanisms affecting the dynamics of white oak mortality. Although the observed patterns can be explained by local scale causes, our investigation of site scale processes, especially self-thinning, offers a plausible explanation for the observed patterns. It is because previous studies have found that self-thinning, a natural process characterized by intraspecific competition leading to mortality, may accumulate over time and produce a clustered distribution pattern [77]. For instance, oak forests across the central region of WOM rate (e.g., Illinois Ozark hills) had gone for massive competition with shade-tolerant species such as sugar maple, red maple, and American beech [47,79,80]. In this condition, the mortality events may be driven by competition for resources within stands [81]. The spatial clustering found in this region raises the possibility that these site-scale processes interact to affect WOM locally. However, our results also depicted a random pattern across the local scale of the central region. This random pattern may be due to environmental heterogeneity in the core area, which might result in a variety of microhabitats that have an impact on tree mortality rates [82,83]. For instance, disparities in soil types, moisture content, and other ecological parameters can lead to a distribution of WOM that appears random [84,85]. Others found that stochastic events such as disease outbreaks or drought can be the cause of random death patterns [86,87]. There is a greater chance for random mortality since drought followed by ice storms and other localized variables are impacting mortality in white oaks lacking a clear spatial pattern [44]. The observed randomness at the local scale could represent temporal dynamics, in which short-term variations in environmental variables impact mortality events [88,89]. Over time, these variations can result in inconsistent geographic clustering. Likewise, the randomness may be facilitated by the edge effects in which the edges of white oaks stand in this region and might have faced distinct ecological conditions compared to the interior. It is because researchers documented that adjacent forest canopies had demonstrated to restrict light availability and mitigated tree growth on the outer margins of even-aged regeneration methods and gaps may exacerbate random mortality patterns [90,91,92,93,94]. Our results also reported a uniform pattern in WOM rates at abroad scale that begs the question of what ecological factors are driving the dynamics of WOM. This unexpected uniformity could be caused by several factors. A uniform distribution of WOM rates may be facilitated by consistency in moisture content [95], and drought-pathogen interactions [96]. For instance, white oak stands throughout the large territory of the central region and has regular growth circumstances due to stable and consistent environmental conditions with an abundance of resources [97]. And this stability in the environment and resources supports a uniform pattern for tree species. Similarly, the central region has a diverse ecosystem caused by human-induced variables like land use patterns such as private landownership across most of the states (e.g., Kentucky, Tennessee, Virginia, etc.). And the uniform land fragmentation resulting from landownership has contributed to a uniform distribution of WOM [56,98].
Our results from spatial patterns at both local and broad scale clustering in the northern region WOM rate indicate that several ecological processes and spatial factors are playing vital roles. Specific areas with comparable environmental circumstances, such as soil compaction, moisture levels, and species interactions, are examples of local scale clustering in WOM rate [99,100]. There is a greater chance that a specific soil type promotes the growth of a specific disease impacting white oaks' health. For instance, white oaks are impacted by exotic insects and diseases, and affect the abundance and distribution across the local scale [101,102]. Furthermore, given that the majority of white oaks stands were restored following destruction and the ensuing agricultural abandonment about a century ago, it is possible that site scale processes like self-thinning in white oak stands may accumulate throughout the region, which may result in a region wide WOM [103,104,105]. Besides, elevation and distance from water have also an impact on the clustering observed at both local and broad scales. [74,106,107] reported that environmental variability, such as elevation changes or proximity to bodies of water, had led to both local and broad-scale clustering. This is because places with comparable topographical features demonstrate clustering tendencies as a result of consistent environmental variables. There is also a greater chance that broad-scale clustering could be attributed to regional variables impacting the dynamics of WOM rates. Regional variables including climate fluctuations, historical disruptions, or landscape features that influence death rates at larger spatial scales could all be considered [108,109,110]. We also found both random as well as uniform patterns across a broad scale. This could be due to various environmental stochastic processes. For instance, the northern region of the US is subjected to intermittent weather extremes, such as localized severe storms or extreme precipitation [111,112]. These stochastic processes result in random WOM patterns within specified broad scale ranges, reflecting the unpredictability of death events in such circumstances. A uniform WOM rate pattern at a broad scale might be the result of spatially homogeneous environmental factors. Factors include areas with constant soil types, climate, and land use methods such as private landownership, consistent logging, or land management techniques that may strongly exhibit uniform mortality rates [56,113,114]. This homogeneity may result in similar WOM rates across large spatial extents. There are also edge effects and landscape characteristics that can contribute to both random and uniform patterns at broad scales. The reason may include locations near forest boundaries or regions with distinct topographical features [115,116]. These localized impacts may provide randomness within specific broad-scale ranges while contributing to homogeneity in neighboring places.
5. Conclusions
Our study showed various types of spatial patterns of WOM rate across the eastern US. These patterns are clustered, random, and uniform across southern, central, and northern region WOM rates with varying scales. Our research highlights mainly a clustered pattern at the local scale across each region and offers important new insights into the spatial patterns of WOM rate in these regions. The observed clustered pattern at the local scale highlights the complex interplay between the white oak population and the specific environmental circumstances across the eastern US. As stand-level competition, topographical and edaphic factors are identified as determinants that contribute to localized clustering of WOM rates. The fact that self-thinning is associated with site-scale processes highlights how complicated the mechanisms affecting white oak stands are in these regions. Such intricate ecological interactions emphasize the need to take finer-scale environmental elements into account when studying mortality events. This localized clustering highlights the necessity for site-specific solutions that consider the variability of topographical and soil conditions. However, there is also a clustered pattern at a broad scale across southern and northern region WOM rates suggesting a significant role of climate, elevation, and proximity to water bodies in these regions.
Our examination of the regional patterns of WOM rate found a distinct random pattern at broad scales both in the southern and northern regions. This random pattern suggests factors at broader scales such as climate, geology, and stochastic events may have impacted white oak stands. Similarly, climatic stressors such as water scarcity and environmental strain, have been identified as major factors in the random pattern of WOM events both at local and broad scale [117].
Our study also unveiled a uniform pattern across all regions i.e., southern, central, and northern WOM rates. The reasons were associated with the presence of spatially homogeneous environmental conditions across a broad scale that may result in the uniform WOM pattern. Regions with consistent soil types, climate, and land use practices such as private landownerships and forest management techniques may suggest a uniform death rate across all regions. It is also evident that the land use practices, and stability of growth conditions emerge as significant factors of a uniform WOM distribution, giving valuable insights for forest management and conservation efforts in these regions exhibiting such patterns. However, further research is needed to investigate the specific mechanism underlying these relationships and to assess the long-term impacts on oak forest ecosystems. Future studies could explore the effects of multiple factors such as biotic and abiotic factors as well as land use practices, to obtain a more comprehensive understanding of the spatial pattern of WOM rate.
Author Contributions
Conceptualization, S.K.; methodology, S.K.; validation, S.K., and H.H.; writing—original draft preparation, S.K.; writing—review and editing, S.K., H.H., and S.B.; supervision, H.H.; project administration, H.H., and S.B.; funding acquisition, S.B. All authors have read and agreed to the published version of the manuscript.
Funding
United States Department of Agriculture/National Institute of Food and Agriculture 1890 Capacity Building Grant, Award number 2021-38821-34704.
Data Availability Statement
Not applicable.
Acknowledgments
We acknowledge the University of Missouri School of Natural Resources for its facilities and support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wohlleben The Hidden Life of Trees: What They Feel, How They Communicate. Wohlleben, Peter. Irish Forestry 2016, 74, 226–228.
- Newell, F.L.; Rodewald, A.D. Role of Topography, Canopy Structure, and Floristics in Nest-Site Selection and Nesting Success of Canopy Songbirds. For Ecol Manage 2011, 262, 739–749. [CrossRef]
- Parola, A.C.; Vesely, W.S.; Croasdaile, M.A. Geomorphic Characteristics of Streams in the Bluegrass Physiographic Region of Kentucky. Kentucky Division of 2007, 319.
- Tallamy, D.W. The Nature of Oaks: The Rich Ecology of Our Most Essential Native Trees; Timber Press, 2021;
- Creutzburg, M.K.; Scheller, R.M.; Lucash, M.S.; Evers, L.B.; Leduc, S.D.; Johnson, M.G. Bioenergy Harvest, Climate Change, and Forest Carbon in the Oregon Coast Range. GCB Bioenergy 2016, 8, 357–370. [CrossRef]
- Littlefield, C.E.; D’Amato, A.W. Identifying Trade-Offs and Opportunities for Forest Carbon and Wildlife Using a Climate Change Adaptation Lens. Conserv Sci Pract 2022, 4, 1–14. [CrossRef]
- Haavik, L.J.; Jones, J.S.; Galligan, L.D.; Guldin, J.M.; Stephen, F.M. Oak Decline and Red Oak Borer Outbreak: Impact in Upland Oak-Hickory Forests of Arkansas, USA. Forestry: An International Journal of Forest Research 2012, 85, 341–352. [CrossRef]
- Mccracken, K.E.; Witham, J.W.; Hunter, M.L. Relationships between Seed Fall of Three Tree Species and Peromyscus Leucopus and Clethrionomys Gapperi during 10 Years in an Oak-Pine Forest. J Mammal 1999, 80, 1288–1296. [CrossRef]
- Starkey, D.A.; Oliveria, F.; Mangini, A.; Mielke, M. NATURAL PHENOMENA, SEVERE OCCURRENCES. In Proceedings of the Upland Oak Ecology Symposium: History, Current Conditions, and Sustainability: Fayetteville, Arkansas, October 7-10, 2002; 2004; Vol. 73, p. 217.
- Tainter, F.H.; Retzlaff, W.A.; Starkey, D.A.; Oak, S.W. Decline of Radial Growth in Red Oaks Is Associated with Short-Term Changes in Climate. European Journal of Forest Pathology 1990, 20, 95–105.
- Greenberg, C.H. Long-Term Recovery Dynamics Following Hurricane-Related Wind Disturbance in a Southern Appalachian Forest. For Ecol Manage 2021, 502, 119704. [CrossRef]
- Reed, S.E.; English, J.T.; Muzika, R.M. Phytophthora Species Detected in Two Ozark Forests with Unusual Patterns of White Oak Mortality. Plant Dis 2019, 103, 102–109. [CrossRef]
- Shifley, S.R.; Fan, Z.; Kabrick, J.M.; Jensen, R.G. Oak Mortality Risk Factors and Mortality Estimation. For Ecol Manage 2006, 229, 16–26. [CrossRef]
- Heitzman, E.; Guldin, J.M. Impacts of Oak Decline on Forest Structure in Arkansas and Oklahoma : Preliminary Results. Methods 2004, 142–146.
- Spetich, M.A. Upland Oak Ecology Symposium: History, Current Conditions, and Sustainability. Uplannd Oak Ecology Symposium: History, Current Conditions, and Sustainability 2002, 318.
- Nagle, A.M.; Long, R.P.; Madden, L. V.; Bonello, P. Association of Phytophthora Cinnamomi with White Oak Decline in Southern Ohio. Plant Dis 2010, 94, 1026–1034. [CrossRef]
- Wang, C.; He, H.S.; Kabrick, J.M. A Remote Sensing-Assisted Risk Rating Study to Predict Oak Decline and Recovery in the Missouri Ozark Highlands, USA. GIsci Remote Sens 2008, 45, 406–425. [CrossRef]
- McConnell, M.E.; Balci, Y. Phytophthora Cinnamomi as a Contributor to White Oak Decline in Mid-Atlantic United States Forests. Plant Dis 2014, 98, 319–327. [CrossRef]
- Aldrich, P.R.; Parker, G.R.; Romero-severson, J.; Michler, C.H. Forest : 75 Years of Data. Forest Science 2005, 51.
- Dey, D. Dey DC 2002 Chapter 5 Oak Book for Oak Silviculture in Eastern North America. 2009.
- Greenberg, C.H.; Keyser, T.L.; Speer, J.H. Temporal Patterns of Oak Mortality in a Southern Appalachian Forest (1991-2006). Natural Areas Journal 2011, 31, 131–137. [CrossRef]
- Abrams, M.D. Where Has All the White Oak Gone? Bioscience 2003, 53, 927–939.
- Balci, Y.; Long, R.P.; Mansfield, M.; Balser, D.; MacDonald, W.L. Involvement of Phytophthora Species in White Oak (Quercus Alba) Decline in Southern Ohio. For Pathol 2010, 40, 430–442. [CrossRef]
- Fan, Z.; Kabrick, J.M.; Spetich, M.A.; Shifley, S.R.; Jensen, R.G. Oak Mortality Associated with Crown Dieback and Oak Borer Attack in the Ozark Highlands. For Ecol Manage 2008, 255, 2297–2305. [CrossRef]
- Wood, J.D.; Knapp, B.O.; Muzika, R.M.; Stambaugh, M.C.; Gu, L. The Importance of Drought-Pathogen Interactions in Driving Oak Mortality Events in the Ozark Border Region. Environmental Research Letters 2018, 13. [CrossRef]
- Chastain, R.A.; Currie, W.S.; Townsend, P.A. Carbon Sequestration and Nutrient Cycling Implications of the Evergreen Understory Layer in Appalachian Forests. For Ecol Manage 2006, 231, 63–77. [CrossRef]
- Peltzer, D.A.; Allen, R.B.; Lovett, G.M.; Whitehead, D.; Wardle, D.A. Effects of Biological Invasions on Forest Carbon Sequestration. Glob Chang Biol 2010, 16, 732–746. [CrossRef]
- Schlesinger, W.H.; Dietze, M.C.; Jackson, R.B.; Phillips, R.P.; Rhoades, C.C.; Rustad, L.E.; Vose, J.M. Forest Biogeochemistry in Response to Drought. Glob Chang Biol 2016, 22, 2318–2328. [CrossRef]
- Profft, I.; Mund, M.; Weber, G.E.; Weller, E.; Schulze, E.D. Forest Management and Carbon Sequestration in Wood Products. Eur J For Res 2009, 128, 399–413. [CrossRef]
- Wang, W.J.; He, H.S.; Fraser, J.S.; Thompson, F.R.; Shifley, S.R.; Spetich, M.A. LANDIS PRO: A Landscape Model That Predicts Forest Composition and Structure Changes at Regional Scales. Ecography 2014, 37, 225–229. [CrossRef]
- de Vries, W.; Du, E.; Butterbach-Bahl, K. Short and Long-Term Impacts of Nitrogen Deposition on Carbon Sequestration by Forest Ecosystems. Curr Opin Environ Sustain 2014, 9, 90–104. [CrossRef]
- Hui, D.; Deng, Q.; Tian, H.; Luo, Y. Handbook of Climate Change Mitigation and Adaptation; 2016; ISBN 9781461464310.
- Vesely, D.; Tucker, G. A Landowner’s Guide for Restoring and Managing Oregon White Oak Habitats. 2004.
- Keyser, P.D.; Fearer, T.; Harper, C.A. Managing Oak Forests in the Eastern United States; CRC Press, 2016;
- Allen, D.; Dick, C.W.; Strayer, E.; Perfecto, I.; Vandermeer, J. Scale and Strength of Oak–Mesophyte Interactions in a Transitional Oak–Hickory Forest. Canadian Journal of Forest Research 2018, 48, 1366–1372. [CrossRef]
- Martin, K.L.; Goebel, P.C. The Foundation Species Influence of Eastern Hemlock (Tsuga Canadensis) on Biodiversity and Ecosystem Function on the Unglaciated Allegheny Plateau. For Ecol Manage 2013, 289, 143–152. [CrossRef]
- Frelich, L.E.; Hale, C.M.; Scheu, S.; Holdsworth, A.R.; Heneghan, L.; Bohlen, P.J.; Reich, P.B. Earthworm Invasion into Previously Earthworm-Free Temperate and Boreal Forests. Biol Invasions 2006, 8, 1235–1245. [CrossRef]
- McDowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.; Aukema, B.H.; Bond-Lamberty, B.; Chini, L.; Clark, J.S.; Dietze, M.; Grossiord, C.; Hanbury-Brown, A.; et al. Pervasive Shifts in Forest Dynamics in a Changing World. Science (1979) 2020, 368, eaaz9463. [CrossRef]
- Hall, M. The Effects of Prescribed Burns on Oak ( Quercus Spp .) and Red Maple ( Acer Rubrum ) Stump Sprouts in Southeastern Ohio. 2011, 1–25.
- Muhly, T.B.; Hebblewhite, M.; Paton, D.; Pitt, J.A.; Boyce, M.S.; Musiani, M. Humans Strengthen Bottom-Up Effects and Weaken Trophic Cascades in a Terrestrial Food Web. PLoS One 2013, 8. [CrossRef]
- Fearer, T.M.; Norman, G.W.; Pack, J.C.; Bittner, S.; Healy, W.M. Influence of Physiographic and Climatic Factors on Spatial Patterns of Acorn Production in Maryland and Virginia, USA. J Biogeogr 2008, 35, 2012–2025. [CrossRef]
- LeBlanc, D.C.; Terrell, M.A. Comparison of Growth-Climate Relationships between Northern Red Oak and White Oak across Eastern North America. Canadian Journal of Forest Research 2011, 41, 1936–1947. [CrossRef]
- Dale, M.R.T. Spatial Pattern Analysis in Plant Ecology; Cambridge university press, 2000;
- Fan, Z.; Fan, X.; Crosby, M.K.; Moser, W.K.; He, H.; Spetich, M.A.; Shifley, S.R. Spatio-Temporal Trends of Oak Decline and Mortality under Periodic Regional Drought in the Ozark Highlands of Arkansas and Missouri. Forests 2012, 3, 614–631. [CrossRef]
- Clark, F.B.; Hutchinson, J.G. Central Hardwood Notes; US Department of Agriculture, Forest Service, North Central Forest~…, 1989;
- Cleland, D.T.; Freeouf, J.A.; Keys, J.E.; Nowacki, G.J.; Carpenter, C.A.; McNab, W.H. Ecological Subregions: Sections and Subsections for the Conterminous United States. Gen. Tech. Rep. WO-76D 2007, 242.
- Fralish, J.S. The Central Hardwood Forest: Its Boundaries and Physiographic Provinces. General Technical Report - North Central Research Station, USDA Forest Service 2003, 1–20.
- Wang, W.J.; He, H.S.; Thompson, F.R.; Fraser, J.S.; Hanberry, B.B.; Dijak, W.D. Importance of Succession, Harvest, and Climate Change in Determining Future Composition in U.S. Central Hardwood Forests. Ecosphere 2015, 6, 1–18. [CrossRef]
- Stambaugh, M.C.; Knapp, B.O.; Dey, D.C. Fire Ecology and Management of Forest Ecosystems in the Western Central Hardwoods and Prairie-Forest Border; 2021; ISBN 9783030732677.
- Bechtold, W.A.; Patterson, P.L. The Enhanced Forest Inventory and Analysis Program--National Sampling Design and Estimation Procedures; USDA Forest Service, Southern Research Station, 2005; Vol. 80.
- Tinkham, W.T.; Mahoney, P.R.; Hudak, A.T.; Domke, G.M.; Falkowski, M.J.; Woodall, C.W.; Smith, A.M.S. Applications of the United States Forest Inventory and Analysis Dataset: A Review and Future Directions. Canadian Journal of Forest Research 2018, 48, 1251–1268. [CrossRef]
- McRoberts, R.E.; Miles, P.D. United States of America. In National Forest Inventories: Assessment of Wood Availability and Use; Vidal, C., Alberdi, I.A., Hernández Mateo, L., Redmond, J.J., Eds.; Springer International Publishing: Cham, 2016; pp. 829–842 ISBN 978-3-319-44015-6.
- Smith, W.B. Forest Inventory and Analysis: A National Inventory and Monitoring Program. Environmental Pollution 2002, 116, 233–242. [CrossRef]
- Gray, A.; Brandeis, T.; Shaw, J.; McWilliams, W.; Miles, P. Forest Inventory and Analysis Database of the United States of America (FIA). Biodiversity & Ecology 2012, 4, 225–231. [CrossRef]
- Burrill, E.A.; Wilson, A.M.; Turner, J.A.; Pugh, S.A.; Menlove, J.; Christiansen, G.; Conkling, B.L.; David, W. The Forest Inventory and Analysis Database: Database Description and User Guide Version 8.0 for Phase 2. US Department of Agriculture, Forest Service 2018, 946.
- Khadka, S.; Gyawali, B.R.; Shrestha, T.B.; Cristan, R.; Banerjee, S. “Ban”; Antonious, G.; Poudel, H.P. Exploring Relationships among Landownership, Landscape Diversity, and Ecological Productivity in Kentucky. Land use policy 2021, 111, 105723. [CrossRef]
- Garnas, J.R.; Ayres, M.P.; Liebhold, A.M.; Evans, C. Subcontinental Impacts of an Invasive Tree Disease on Forest Structure and Dynamics. Journal of Ecology 2011, 99, 532–541. [CrossRef]
- Iverson, L.R.; Hutchinson, T.F.; Prasad, A.M.; Peters, M.P. Thinning, Fire, and Oak Regeneration across a Heterogeneous Landscape in the Eastern U.S.: 7-Year Results. For Ecol Manage 2008, 255, 3035–3050. [CrossRef]
- Yang, S.-I.; Brandeis, T.J. Estimating Maximum Stand Density for Mixed-Hardwood Forests among Various Physiographic Zones in the Eastern US. For Ecol Manage 2022, 521, 120420. [CrossRef]
- Robeson, S.M.; Li, A.; Huang, C. Point-Pattern Analysis on the Sphere. Spat Stat 2014, 10, 76–86. [CrossRef]
- Gavin, D.G. K1D: Multivariate Ripley’s K-Function for One-Dimensional Data. Oikos 2007, 1, 1–8.
- Haase, P. Spatial Pattern Analysis in Ecology Based on Ripley’s K-Function: Introduction and Methods of Edge Correction. Journal of Vegetation Science 1995, 6, 575–582. [CrossRef]
- Ripley, B.D. Spatial Statistics; John Wiley \& Sons, 2005;
- Miron, A.C.; Bezerra, T.G.; Nascimento, R.G.M.; Emmert, F.; Pereira, R.S.; Higuchi, N. Spatial Distribution of Six Managed Tree Species Is Influenced by Topography Conditions in the Central Amazon. J Environ Manage 2021, 281. [CrossRef]
- Wehenkel, C.; Brazão-Protázio, J.M.; Carrillo-Parra, A.; Martínez-Guerrero, J.H.; Crecente-Campo, F. Spatial Distribution Patterns in the Very Rare and Species-Rich Picea Chihuahuana Tree Community (Mexico). PLoS One 2015, 10, 1–19. [CrossRef]
- Condit, R. Spatial Patterns in the Distribution of Tropical Tree Species. Science (1979) 2000, 288, 1414–1418. [CrossRef]
- Bose, A.K.; Weiskittel, A.; Kuehne, C.; Wagner, R.G.; Turnblom, E.; Burkhart, H.E. Tree-Level Growth and Survival Following Commercial Thinning of Four Major Softwood Species in North America. For Ecol Manage 2018, 427, 355–364. [CrossRef]
- Dey, D.C. Sustaining Oak Forests in Eastern North America: Regeneration and Recruitment, the Pillars of Sustainability. Forest Science 2014, 60, 926–942. [CrossRef]
- Palik, B.; Mitchell, R.J.; Pecot, S.; Battaglia, M.; Mou, P. Spatial Distribution of Overstory Retention Influences Resources and Growth of Longleaf Pine Seedlings. Ecological Applications 2003, 13, 674–686. [CrossRef]
- Petroselli, A.; Vessella, F.; Cavagnuolo, L.; Piovesan, G.; Schirone, B. Ecological Behavior of Quercus Suber and Quercus Ilex Inferred by Topographic Wetness Index (TWI). Trees - Structure and Function 2013, 27, 1201–1215. [CrossRef]
- Muller, J.J.; Nagel, L.M.; Palik, B.J. Forest Adaptation Strategies Aimed at Climate Change: Assessing the Performance of Future Climate-Adapted Tree Species in a Northern Minnesota Pine Ecosystem. For Ecol Manage 2019, 451, 117539. [CrossRef]
- Lafon, C.W. Forest Disturbance by Ice Storms in Quercus Forests of the Southern Appalachian Mountains, USA. Ecoscience 2006, 13, 30–43. [CrossRef]
- Kim, D.; Millington, A.C.; Lafon, C.W. Biotic and Spatial Factors Potentially Explain the Susceptibility of Forests to Direct Hurricane Damage. J Ecol Environ 2019, 43, 1–12. [CrossRef]
- Aldrich, P.R.; Parker, G.R.; Ward, J.S.; Michler, C.H. Spatial Dispersion of Trees in an Old-Growth Temperate Hardwood Forest over 60 Years of Succession. For Ecol Manage 2003, 180, 475–491. [CrossRef]
- Luisa, B.G. The Ecology of Natural Disturbance and Patch Dynamics; Academic press, 2012;
- Pickett, S.T.; Wu, J.; Cadenasso., M.L. Patch Dynamics and the Ecology of Disturbed Ground: A Framework for Synthesis. Ecosystems of Disturbed Ground 1999, 707–722.
- Rozas, V. Structural Heterogeneity and Tree Spatial Patterns in an Old-Growth Deciduous Lowland Forest in Cantabria, Northern Spain. Plant Ecol 2006, 185, 57–72. [CrossRef]
- Fralish, J.S. The Keystone Role of Oak and Hickory in the Central Hardwood Forest. In Proceedings of the Upland Oak Ecology Symposium: History, Current Conditions, and Sustainability: Fayetteville, Arkansas, October 7-10, 2002; 2004; p. 78.
- Chapman, R.A.; Heitzman, E.; Shelton, M.G. Long-Term Changes in Forest Structure and Species Composition of an Upland Oak Forest in Arkansas. For Ecol Manage 2006, 236, 85–92. [CrossRef]
- Hoover, N.M. Succession of an Upland Oak/Hickory Forest in the Central Hardwood Region. 2018.
- Pretzsch, H.; Grote, R. Tree Mortality: Revisited under Changed Climatic and Silvicultural Conditions. 2023.
- Beckage, B.; Clark, J.S. Seedling Survival and Growth of Three Forest Tree Species: The Role of Spatial Heterogeneity. Ecology 2003, 84, 1849–1861. [CrossRef]
- Pérez-Ramos, I.M.; Urbieta, I.R.; Zavala, M.A.; Marañón, T. Ontogenetic Conflicts and Rank Reversals in Two Mediterranean Oak Species: Implications for Coexistence. Journal of Ecology 2012, 100, 467–477. [CrossRef]
- Pedersen, B.S. The Role of Stress in the Mortality of Midwestern Oaks As Indicated By Growth Prior To Death. Ecology 1998, 79, 79–93. [CrossRef]
- Casperson_et_al_1999.Pdf.Crdownload.
- Bréda, N.; Badeau, V. Forest Tree Responses to Extreme Drought and Some Biotic Events: Towards a Selection According to Hazard Tolerance? Comptes Rendus - Geoscience 2008, 340, 651–662. [CrossRef]
- Turner, M.G.; Calder, W.J.; Cumming, G.S.; Hughes, T.P.; Jentsch, A.; LaDeau, S.L.; Lenton, T.M.; Shuman, B.N.; Turetsky, M.R.; Ratajczak, Z.; et al. Climate Change, Ecosystems and Abrupt Change: Science Priorities. Philosophical Transactions of the Royal Society B: Biological Sciences 2020, 375. [CrossRef]
- Petritan, A.M.; Petritan, I.C.; Hevia, A.; Walentowski, H.; Bouriaud, O.; Sánchez-Salguero, R. Climate Warming Predispose Sessile Oak Forests to Drought-Induced Tree Mortality Regardless of Management Legacies. For Ecol Manage 2021, 491. [CrossRef]
- Kalyuzhny, M.; Kadmon, R.; Shnerb, N.M. A Neutral Theory with Environmental Stochasticity Explains Static and Dynamic Properties of Ecological Communities. Ecol Lett 2015, 18, 572–580. [CrossRef]
- Burton, P.J. Effects of Clearcut Edges on Trees in the Sub-Boreal Spruce Zone of Northwest-Central British Columbia. Silva Fennica 2002, 36, 329–352. [CrossRef]
- Gray, A.N.; Spies, T.A.; Easter, M.J. Microclimatic and Soil Moisture Responses to Gap Formation in Coastal Douglas-Fir Forests. Canadian Journal of Forest Research 2002, 32, 332–343. [CrossRef]
- Powers, M.D.; Pregitzer, K.S.; Palik, B.J. Physiological Performance of Three Pine Species Provides Evidence for Gap Partitioning. For Ecol Manage 2008, 256, 2127–2135. [CrossRef]
- Van Couwenberghe, R.; Collet, C.; Lacombe, E.; Pierrat, J.C.; Gégout, J.C. Gap Partitioning among Temperate Tree Species across a Regional Soil Gradient in Windstorm-Disturbed Forests. For Ecol Manage 2010, 260, 146–154. [CrossRef]
- Lhotka, J.M.; Stringer, J.W. Forest Edge Effects on Quercus Reproduction within Naturally Regenerated Mixed Broadleaf Stands. Canadian Journal of Forest Research 2013, 43, 911–918. [CrossRef]
- Hanson, P.J.; Todd, J.; Amthor, J.S. Erratum: A Six-Year Study of Sapling and Large-Tree Growth and Mortality Responses to Natural and Induced Variability in Precipitation and Throughfall (Tree Physiology 21 (345-358)). Tree Physiol 2001, 21, 1158.
- Wood, J.D.; Knapp, B.O.; Muzika, R.M.; Stambaugh, M.C.; Gu, L. The Importance of Drought-Pathogen Interactions in Driving Oak Mortality Events in the Ozark Border Region. Environmental Research Letters 2018, 13. [CrossRef]
- Hanberry, B.B.; Dey, D.C.; He, H.S. Regime Shifts and Weakened Environmental Gradients in Open Oak and Pine Ecosystems. PLoS One 2012, 7. [CrossRef]
- Yang, J.; He, H.S.; Shifley, S.R. Spatial Controls of Occurrence and Spread of Wildfires in the Missouri Ozark Highlands. Ecological Applications 2008, 18, 1212–1225. [CrossRef]
- Annighöfer, P.; Beckschäfer, P.; Vor, T.; Ammer, C. Regeneration Patterns of European Oak Species (Quercus Petraea (Matt.) Liebl., Quercus Robur L.) in Dependence of Environment and Neighborhood. PLoS One 2015, 10, 1–16. [CrossRef]
- Copes-Gerbitz, K.; Arabas, K.; Larson, E.; Gildehaus, S. A Multi-Proxy Environmental Narrative of Oregon White Oak (Quercus Garryana) Habitat in the Willamette Valley, Oregon. Northwest Science 2017, 91, 160–185. [CrossRef]
- Abrams, M.D. Where Has All the White Oak Gone? Bioscience 2003, 53, 927–939. [CrossRef]
- Lovett, G.M.; Weiss, M.; Liebhold, A.M.; Holmes, T.P.; Leung, B.; Lambert, K.F.; Orwig, D.A.; Campbell, F.T.; Rosenthal, J.; McCullough, D.G.; et al. Nonnative Forest Insects and Pathogens in the United States: Impacts and Policy Options. Ecological Applications 2016, 26, 1437–1455. [CrossRef]
- Southgate, E.W.B.R.; Thompson, J.E. Secondary Forest Succession in a Post-Agricultural Landscape in the Hudson Valley, New York. Northeast Nat (Steuben) 2014, 21. [CrossRef]
- Wang, J.; Maduako, I.N. Spatio-Temporal Urban Growth Dynamics of Lagos Metropolitan Region of Nigeria Based on Hybrid Methods for LULC Modeling and Prediction. Eur J Remote Sens 2018, 51, 251–265. [CrossRef]
- Wang, W.J.; He, H.S.; Thompson, F.R.; Fraser, J.S.; Dijak, W.D. Changes in Forest Biomass and Tree Species Distribution under Climate Change in the Northeastern United States. Landsc Ecol 2017, 32, 1399–1413. [CrossRef]
- Hosten, P.E.; Hickman, O.E.; Lake, F.K.; Lang, F.A.; Vesely, D. Oak Woodlands and Savannas. Restoring the Pacific Northwest: the art and science of ecological restoration in Cascadia 2006, 63–96.
- Hanberry, B.B.; Kabrick, J.M.; He, H.S. Densification and State Transition Across the Missouri Ozarks Landscape. Ecosystems 2014, 17, 66–81. [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate Change and Forest Disturbances: Climate Change Can Affect Forests by Altering the Frequency, Intensity, Duration, and Timing of Fire, Drought, Introduced Species, Insect and Pathogen Outbreaks, Hurricanes, Windstorms, Ice Storms, or Landslides. Bioscience 2001, 51, 723–734.
- Opdam, P.; Wascher, D. Climate Change Meets Habitat Fragmentation: Linking Landscape and Biogeographical Scale Levels in Research and Conservation. Biol Conserv 2004, 117, 285–297. [CrossRef]
- McEwan, R.W.; Dyer, J.M.; Pederson, N. Multiple Interacting Ecosystem Drivers: Toward an Encompassing Hypothesis of Oak Forest Dynamics across Eastern North America. Ecography 2011, 34, 244–256. [CrossRef]
- Barlow, M.; Gutowski, W.J.; Gyakum, J.R.; Katz, R.W.; Lim, Y.K.; Schumacher, R.S.; Wehner, M.F.; Agel, L.; Bosilovich, M.; Collow, A.; et al. North American Extreme Precipitation Events and Related Large-Scale Meteorological Patterns: A Review of Statistical Methods, Dynamics, Modeling, and Trends; Springer Berlin Heidelberg, 2019; Vol. 53; ISBN 0123456789.
- Walsh, C.R.; Patterson, R.T. Attribution of Observed Periodicity in Extreme Weather Events in Eastern North America. Earth and Space Science 2022, 9. [CrossRef]
- Keeley, J.E. Ecological Foundations for Fire Management in North American Forest and Shrubland Ecosystems; US Department of Agriculture, Forest Service, Pacific Northwest Research Station, 2009; Vol. 779.
- Foster, D.R.; Donahue, B.; Kittredge, D.; Motzkin, G.; Hall, B.; Turner, B.; Chilton, E. New England’s Forest Landscape. Agrarian Landscapes in Transition 2008, 44–88.
- Santos, B.A.; Arroyo-Rodríguez, V.; Moreno, C.E.; Tabarelli, M. Edge-Related Loss of Tree Phylogenetic Diversity in the Severely Fragmented Brazilian Atlantic Forest. PLoS One 2010, 5, 1–7. [CrossRef]
- Collins, R.J.; Carson, W.P. The Effects of Environment and Life Stage on Quercus Abundance in the Eastern Deciduous Forest, USA: Are Sapling Densities Most Responsive to Environmental Gradients? For Ecol Manage 2004, 201, 241–258. [CrossRef]
- Rogers, B.M.; Jantz, P.; Goetz, S.J. Vulnerability of Eastern US Tree Species to Climate Change. Glob Chang Biol 2017, 23, 3302–3320. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions, and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



