
Submitted:
30 August 2024
Posted:
30 August 2024
Read the latest preprint version here
Abstract
Developing new antibiotics poses a significant challenge in the fight against antimicrobial resistance (AMR), a critical global health threat responsible for approximately 5 million deaths annually. Finding new classes of antibiotics that are safe, have acceptable pharmacokinetic properties, and are appropriately active against pathogens is a lengthy and expensive process. Therefore, high-throughput platforms are needed to screen large libraries of synthetic and natural compounds. In this review, we present bacterial cytological profiling (BCP) as a rapid, scalable, and cost-effective method for identifying the mechanisms of action of antibiotics, offering a promising tool for combating AMR and drug discovery. We present the application of BCP for different bacterial organisms and different classes of antibiotics and discuss BCP's advantages, limitations, and potential improvements. Furthermore, we highlight the studies that have utilized BCP to investigate pathogens listed in the Bacterial Priority Pathogens List 2024 and we identify the pathogens whose cytological profiles are missing. Lastly, we explore the most recent artificial intelligence and deep learning techniques that could enhance the analysis of data generated by BCP, potentially advancing our understanding of antibiotic resistance mechanisms and the discovery of novel druggable pathways.
Keywords:
antibiotic resistance
; bacterial cytological profiling
; high-throughput screens
; antibiotic mechanism of action
; bacterial priority pathogen list
; cell segmentation
; machine learning
; deep learning
1. Introduction
Antimicrobial resistance (AMR) has been declared by the World Health Organization (WHO) as one of the top ten global public health threats facing humanity. It impacts both public health and global development, resulting in severe morbidity and mortality worldwide. AMR is defined as a condition characterized by the ability of microorganisms (bacteria, fungi, viruses, and parasites) to live and grow in the presence of antimicrobial agents that were previously reported to be effective against them. Resistance occurs through pathogen evolution, either naturally over time or acquired by the use of antimicrobial drugs, which make these drugs ineffective [1].
It has been estimated that in 2019 alone, antimicrobial resistance killed at least 1.27 million people globally, more deaths than HIV/AIDS or malaria, with 4.95 million deaths associated with AMR [2]. According to the Centers for Disease Control and Prevention’s Antibiotic Resistance Threats Report [3], in the United States, over 2.8 million antibiotic-resistant infections occur each year, leading to over 35,000 deaths. Furthermore, it has been estimated that, if the AMR trend continues, the cumulative loss to world economies might be as high as $100 trillion by 2050 [4]. These alarming statistics underscore the urgent need to develop effective therapeutics to combat antimicrobial resistance.
The efforts undertaken in the field of AMR until now have not been enough despite the research effort and inventive therapeutic approaches. Since 1940 antimicrobials have been used widely [5], [6] , in addition to treating infectious diseases, antibiotics made many modern medical procedures possible, including cancer treatment, organ transplants and open-heart surgery [7]. Even before 1940 and for about 60 years after, most antibiotics were discovered by screening soil samples for such natural products that kill bacteria, including known pathogens, first on culture plates and then in animal infection models [8], [7]. However, in the past 20 years, only two new antibiotic classes have been developed that are only effective for Gram-positive bacteria [9]. To overcome the burden of antibiotic resistance, new high- throughput screens based on bacterial cytological profiling have been developed.
This review emphasizes the use of bacterial cytological profiling (BCP) as a highly effective method for discovering novel antibiotics and rapidly identifying antibiotic targets in a cost-effective manner. BCP initially creates a library of bacterial cell shapes and sizes induced by antibiotics with known mechanisms of action. This library is then used to discover new antibiotics and classify antibiotics based on the components of bacterial cells they target. In this review, we highlight how BCP is used to improve our quantitative understanding of antibiotic pharmacodynamics and bacterial stress responses, as well as how BCP enhances the development of non-traditional antibacterial strategies such as phage therapies[10,11,12].
2. Antibiotic Mechanism of Action and Antibiotic Targets
AMR is a natural process that happens over time through genetic alterations or phenotypic changes [13] in pathogens. Its emergence and spread are accelerated by human activity, mainly the misuse and overuse of antimicrobials to treat, prevent or control infections in humans, animals, and plants. To effectively tackle antibiotic-resistant bacteria, it is essential to understand how antibiotics work, which is known as their mechanism of action (MOA), see Table 1. Understanding MOA involves studying how antibiotics affect bacterial physiology and molecular interaction with bacterial targets (Figure 1). However, determining MOA remains a serious challenge that limits both basic research and antibiotic discovery programs.
Traditionally, a variety of assays are performed to determine the MOA, by the evaluation of whether one of the five basic pathways is inhibited (Figure 1). These efforts typically begin with macromolecular synthesis (MMS) assays that use radioactively labeled precursors to determine whether a compound specifically inhibits peptidoglycan, lipid, protein, RNA or DNA synthesis or whether it blocks all simultaneously [14]. Despite being an important technique, MMS assays are time consuming and suffer from low resolution, low accuracy, and relatively low throughput [15].
To address the limitations associated with MMS assays, various alternative techniques for determining the mechanism of action (MOA) have been devised [32]. These include biochemical approaches, such as affinity chromatography that identify direct biophysical interactions between antimicrobials and their targets through an array where the antibiotic interacts with protein from whole-cell extracts[33,34,35]. Genetic approaches, such as selection for resistance and resistance screening [33], focus on the genetic comparison between non-resistant strains and strains that have evolved and mutated to become resistant to a specific antibiotic, this approach is often able to identify the molecular target of an antibiotic, the specific amino acid residues important for its interaction and the frequency with which resistance occurs.
While these methods offer various benefits, they also come with limitations that hinder their effectiveness. The main disadvantage of these methods is the required time to conduct the assay, moreover, these methods necessitate substantial quantities of purified compounds, which can be challenging as newly discovered lead compounds are frequently available in limited amounts.
Apart from all limitations in determining MOA, discovering new compounds that are active against Gram-positive and Gram-negative bacteria remains challenging. In the past 20 years, only two new antibiotic classes (lipopeptides and oxazolidinones) have been developed and approved by international drug agencies (US Food and Drug Administration and European Medicines Agency)—both of which provide coverage against Gram-positive bacteria [9]. The quinolones, discovered in 1962, were the last novel drug class identified to be active against Gram-negative bacteria [36].
From a technical perspective, finding new classes of antibiotics that are safe, have acceptable pharmacokinetic properties, and are appropriately active is a challenging issue [36]. Additionally, the profit margins for producing antibiotics are relatively low given the high cost for production and the lengthy timeline for research, testing, and approval [36]. Therefore, new, high-throughput screening platforms are needed for the fast and inexpensive screening of large libraries of synthetic and natural compounds that are highly effective against human pathogens[37,38]. The following section reviews the quick and scalable bacterial cytological profiling methods (BCPs) and discusses their availability for some of the most important human pathogens as outlined in the latest WHO 2024 report.
3. BCP to Identify the Mechanism of Action
In 2013, Poochit et al. designed Bacterial Cytological Profiling (BCP) analysis for E. coli cells using different classes of antibiotics [15] (Figure 2). BCP data are obtained using fluorescent microscopy of E. coli cells stained with fluorescent membrane and DNA dyes as well as fluorescent reporter for membrane permeability. Using image analysis software, various bacterial cell parameters, such as cell length, width, solidity, and DNA content, are extracted [15]. Subsequently, complex multidimensional data are analyzed using the Principal Component Analysis (PCA) technique, to cluster cells based on their cytological profile to identify the mechanism of action of known and unknown antibacterial compounds (Figure 2B, Box 1). Since the cytological profiling method produces data at a single-cell level [41], it identifies the metabolic pathways targeted by drugs or other toxins through their effects on bacterial chromosomal condensation, cell shape and overall cellular morphology [15] (Figure 2B). Furthermore, this approach can lead to the identification of antibiotics that are effective against multidrug-resistant bacteria [15].
Box 1. Principle Component Analysis (PCA)
PCA is a widely used statistical technique for analyzing data by identifying a set of principal components that capture the maximum variance in the data with the fewest components [44]. It involves transforming the original variables into a new set of uncorrelated variables, known as principal components, which are linear combinations of the original variables. These principal components are ordered so that the first component explains the maximum variance in the data, followed by the second component, and so on [44]. The process of PCA starts by calculating a matrix that shows how the original variables are related to each other. Then, it finds special vectors (eigenvectors) and values (eigenvalues) from this matrix. The eigenvectors show the new directions for the data, while the eigenvalues tell us how much of the data's variation is captured by each new direction. The principal components are then derived from these eigenvectors, and the data is projected onto these components to obtain the transformed dataset. PCA has diverse applications across almost all scientific fields, including biology, medicine, computer science, and geology. In the context of biomedical research, PCA has been utilized to analyze human cell atlas and prostate cancer risk prediction[45,46]. In computer science, PCA is employed for data processing, dimensionality reduction, and feature extraction[47,48,49]. In Bacterial Cytological Profiling, PCA is used to analyze microscopy data by grouping the phenotypic changes in bacteria after antibiotic exposure, thereby determining the target of the antibiotics (Figure 2B). Together, Principal Component Analysis is a powerful statistical method that enables the transformation of complex data into a new simplified coordinate system to reveal underlying patterns and structures. Its wide-ranging applications underscore its significance in various scientific disciplines.
The cytological profiles triggered by antibiotics reveal their damaging effects. For example, compounds that block translation led to chromosome compaction due to the interference with coordinated translation and insertion of proteins into the membrane, whereas compounds that block transcription lead to chromosome decondensation due to the absence of active RNA polymerase (Figure 2). BCP takes advantage of the limited presence of cell-cycle checkpoints in bacteria, combined with the precision of high-resolution imaging.
When stressed by antibiotics, bacteria show phenotypical changes that are characteristic of the antibiotic target. Treated bacteria transform into spheroplast, protoplast, ovoid or filamenting cells [50]. Spheroplast and protoplast are cells that have lost their peptidoglycan layer, spheroplasts are Gram-negative bacteria that keep their outer membrane, whereas protoplasts are formed from Gram-positive bacteria that lack both a peptidoglycan layer and an outer membrane [51]. Bacterial variants that lack a cell wall, encompassing both Gram-negative and Gram-positive bacteria, are known as L-forms[52,53,54,55].
Ovoid cells refer to bacterial rods (bacilli) that have decreased in length and become oval or round-shaped during antibacterial treatment[56,57]. Nomenclature in the literature varies, with some authors referring to them as ‘round forms’[58,59], ‘round cells’[60,61,62], ‘spherical forms’, ‘spherical cells’[63,64,65], or ‘coccoid forms’[66,67]. Filamentation, or cell elongation, occurs when rod-shaped bacteria (or sometimes cocci) produce peptidoglycan for their side walls but not for their division walls during growth, causing the cells to become unusually elongated [50]. Filamentation can occur following inhibition or disruption of peptidoglycan synthesis [56] but can also occur if DNA synthesis is inhibited[68,69] or DNA is damaged[70,71,72] by a process known as the SOS response that inhibits cell division [28](Figure 2).
Other phenotypic changes can be shown by bacteria under antibiotic exposure as altered cell size, localized swelling, bulge formation, blebbing, peptidoglycan thickening [50]. Phenotypical changes confer an increase in fitness to bacteria in the presence of antibiotics [73]. Resistance to antibiotics is commonly attained via reducing the intracellular concentration of the antibiotic or by reducing antibiotic binding affinities for their specific intracellular targets [74]. By using available BCP data, recent studies have shown that by reducing the surface-to-volume ratio (S/V), bacteria can effectively reduce the antibiotic concentration inside a cell, thereby promoting cell growth by decreasing antibiotic influx [74]. Similarly, an increase in S/V can also provide adaptive benefits by increasing the rate of nutrient uptake or by increasing the rate of antibiotic efflux[74,75,76]. These studies explain how cell shape transformations promote bacterial survival under antibiotic treatments – pointing towards potential new druggable targets that control cell shape and size under stress.
BCP has been successfully employed to study the MOA of various antibacterial agents, including azithromycin [77], diphenylureas [78] and thailandamide [79]. It has also been used to identify the cellular pathways targeted by anticancer metal complexes [80], to study the response of bacteria to antibiotics in different growth conditions [81]. Additionally, BCP has been used to identify the cellular pathways targeted by antibacterial molecules affecting different cellular pathways [82], [83], making it a valuable tool not only for determining antibacterial targets but also to potentially identify novel MOA i.e., ones that target new proteins or new pathways (Figure 3).
BCP offers unprecedented insights not only into antibiotic therapies but also into phage therapies, revealing how phages exploit or disrupt bacterial cellular processes[10,11,12,84]. BCP enables the visualization of distinct cytological changes within bacterial cells during phage infection. These cytological signatures not only uncover the pathways and cellular targets that phages manipulate to propagate their lifecycle but also highlight the role of bacterial defense mechanisms in combating phage infection [11]. BCP has demonstrated how the overexpression of phage-related proteins can induce specific cytological changes that inhibit phage replication, showcasing the potential for targeting bacterial metabolic pathways to modulate phage activity [10]. Additionally, BCP has been instrumental in assessing the impact of various antibiotics on phage replication, revealing that certain antibiotics can synergize with phages to enhance bacterial cell lysis, while others inhibit phage propagation by disrupting essential bacterial processes [11]. This dual capability of BCP to monitor both the direct effects of phage infection and the influence of external agents such as antibiotics makes it an invaluable tool in studying phage-bacteria dynamics.
4. BCP of Important Human Pathogens
Most importantly, BCP has been successfully used to study some of the most important human pathogens from the WHO Bacterial Priority Pathogens List (Table 2). In 2017, using a multicriteria decision analysis technique, WHO experts together with researchers from the Division of Infectious Diseases at the University of Tübingen, Germany, developed the first Bacterial Priority Pathogen List (BPPL) to guide investment into the R&D of new antibacterials for guiding AMR surveillance, prevention and control [85]. During the past 7 years, the antibiotic development pipeline brought to the market nine new antibiotics with in-vitro or in-vivo activity against the 2017 BPPL “critical” priority pathogens, although resistant strains have since been described for almost all of them[86,87]. In 2024, WHO published an updated list to address current challenges and provide essential guidance for policymakers, national health authorities and others involved in decisions about R&D and investment. The 2024 BPPL includes 15 families of antibiotic-resistant pathogens, grouped into critical, high and medium categories of priority for R&D and public health measures [88] (Table 2). Out of 15 families, bacterial cytological profiling is not available for 30 % of them: Non-typhoidal Salmonella, Neisseria gonorrhoeae, Group A and B Streptococci, Haemophilus influenzae. Therefore, urgent BCPs regarding these severe pathogenic organisms are needed.
5. BCP to Identify New Druggable Cell Pathways
BCP is used to scan new antibacterial components to identify their specific targets (Figure 3). As demonstrated, BCP effectively differentiates between various morphological changes induced by different antibiotics, thereby providing insights into the antibiotic’s mechanism of action. If a novel antibiotic places bacteria in a distinct region of the PCA plot compared to known antibiotic targets, it could indicate a new pathway target or MOA previously uncharacterized (Figure 3). For example, if the PCA analysis shows that the morphology of bacteria treated with a new antibiotic clusters in a region associated with membrane or RNA targets (Arrows 2 and 3 in Figure 3), it directly indicates the antibiotic's mode of action. Conversely, if the antibiotic's effect causes a morphology change that places bacteria in a novel zone, as illustrated with Arrow 4, it may suggest the discovery of a new antibacterial pathway or target.
Together, BCP significantly enhances drug development processes by offering a precise, fast and systematic method for characterizing the effects of new antibacterial agents. Its ability to identify target-specific morphological changes provides a comprehensive tool for uncovering novel antibiotic targets and advancing our understanding of bacterial physiology.
6. BCP Limitations
Even with all the advantages we mentioned about BCP, it has certain limitations. BCP can identify the general target of an antibiotic, but it cannot provide precise information about the exact site within the target that is affected. For instance, while BCP can indicate that an antibiotic targets the ribosome, it cannot specify which part of the ribosome is involved.
BCP requires staining chemicals to evaluate DNA content and cell size and shape, with fluorescent dye intensity being a crucial variable (Table 3). These factors are essential for determining the mechanism of action of antibiotics. A plethora of dyes, protein fusions, and reporter strains are available and have been used in BCP, enabling both rapid mode of action categorization and in-depth analysis of antibiotic mechanisms. However, the plethora of possible dyes, strains, and assays can make it difficult to choose which specific phenotypic experiments are the most suited for a given purpose (Table 3). Furthermore, bacterial cell physiology is complex, and many cellular processes are intimately interconnected, e.g., by co-dependent regulation or metabolic flux [103], [104].
7. BCP Potential Improvements
Despite the cell walls being one of the primary antibiotic targets, direct visualization methods of cell wall synthesis and remodeling during antibiotic exposure have been lacking in BCP approaches developed so far (Table 3). In 2012, Kuru et al. discovered a groundbreaking method for bacterial cell wall staining using fluorescent amino acids [121]. Cell wall provides the shape and structural integrity to the cell. It is made of peptidoglycan (PG), which consists of glycan strands cross-linked by d-amino acid (DAA) [122]. The team introduced HADA and NADA, two fluorescent d-amino acids (FDAAs) attached to a d-amino acid backbone (3-amino-d-alanine). This chemical biology approach aims to detect and visualize the exact location and amount of new peptidoglycan layer synthesis in bacteria. By using HADA or NADA as fluorescently labeled peptidoglycan components during cell wall synthesis, the technique allows researchers to observe morphological changes in bacteria over time. This provides a chronological account of shifts in peptidoglycan synthesis within individual cells across various bacterial species. This is relevant as HADA and NADA can be implemented in the methodology of BCP as dyes to unravel the growth modes of bacteria under antibiotic exposure as they exhibit a diverse array of growth patterns that confer selective advantages in their environments [123], [74].
Other fluorescent dyes are available to quantitatively probe bacterial physiological states: ThT and DiBAC4 for bacterial membrane potential[124,125,126,127], carboxy-H2DCFDA for reactive oxygen species (ROS) [127], and DAF-FM for reactive nitrogen species (RNS) [127]. By integrating membrane potential, ROS, and RNS into cytological profiles could provide additional information regarding bacterial physiology and bacterial stress response during antibiotic treatment.
8. Image Analysis Tools for BCP and Data Availability
Although fluorescent microscopy is a valuable tool for characterizing cellular and subcellular structures and dynamics, quantitative analysis of microscopy images remains a persistent challenge[119,128]. Once the fluorescence microscopy is done, analyzing the images obtained is a critical step for accurately determining cell morphologies. This is especially evident in the study of bacterial shapes since their cell body is composed of a small number of pixels (for example, ~ 100–300 px [2] for E. coli in typical experiments) [129]. At this scale, accurate subcellular localization requires defining the cell boundary with single-pixel precision or more desirable sub-pixel resolution [130].
In addition to their small size, bacteria exhibit a diverse array of shapes. While many frequently studied bacteria can be adequately represented as rods or spheres, there is growing interest in those with more complex forms or the atypical shapes that emerge after exposure to antibiotics, such as seen in Caulobacter crescentus [73]. Cell segmentation allows us to analyze microscopy images more precisely to identify these different shapes. Segmentation is a method of partitioning an image into distinct regions with similar attributes, such as texture, intensity, or gray level, with the goal of distinguishing the cells from the background to enable quantitative analysis such as cell feature extraction [130].
Cell segmentation is a complex problem that extends far beyond microbiological research where advances can be lagging as expertise is less common; thus, many solutions are currently available within ImageJ [128,131,132] which broaden the number of potential users, but many more also use stand-alone image-analysis programs which can at times be less supported and less accessible [133,134]. Many traditional solutions use image-processing techniques such as the Otsu thresholding method to segment isolated cells [130]; however, this approach performs poorly on cells in close contact and it requires image-by-image tuning to optimize parameters. Furthermore, whilst implementing image-by-image thresholds instead of one static threshold can potentially improve segmentation, it also introduces bias [135].
Classical image segmentation techniques have been in use since the 1960s[136,137], laying the groundwork for the more advanced artificial intelligence methods used nowadays. In 2016 SuperSegger was developed to address thresholding issues in classical segmentation specifically in bacterial phase-contrast images and is an interesting example of classical segmentation combined with Deep learning [138].This program utilizes traditional image filtering techniques now with a shallow neural network to correct for errors that thresholding and watershed segmentation tend to produce [139]. This would soon be overshadowed by other Deep neural networks (DNNs) which have now become the backbone of most Deep learning segmentation methods and are widely recognized as superior tools for cell segmentation [140]. This has marked a successful trend in using classical techniques to supplement or in conjunction with DNN methods for cell segmentation. However, as showcased in Table 3 there is currently a significant gap in Deep learning being used in BCP. Unlike traditional image processing, machine-learning approaches such as DNNs require training on a ground-truth dataset of cells and corresponding labels. Trained DNNs are thus limited in applicability to images that are representative of those in the training dataset and require greater knowledge to set up than popular classical segmentation methods [141]. Early DNN approaches were based on the Mask R-CNN architecture, whereas more recent algorithms such as StarDist, Cellpose and MiSiC are based on the U-net architecture[131,132,142,143] but new Transformer-based object detection frameworks may begin to surpass them. We have already seen remarkable results from team Osi lab in the multimodality cell segmentation challenge, with their algorithm outcompeting all other entries and some established and popular algorithms[128,133].
Among easily available Deep learning segmentation algorithms, Pachitariu et al. showed that Cellpose outperforms the popular programs Mask R-CNN and StarDist on a variety of cell types and cell-like objects, distinguishing it as a relatively powerful general solution for cell segmentation [131]. Cutler et al. evaluated the performance of (at the time) state-of-the-art cell segmentation algorithms on a diverse collection of bacterial cells. Their findings motivated the design of a new algorithm, Omnipose, which substantially outperforms all segmentation algorithms tested across a wide range of bacterial cell sizes, morphologies, and optical characteristics and as such has been used extensively in literature [129]. Omnipose, whilst well supported and widely used in bioimage analysis as a generalist segmentation algorithm is now demonstrably worse than more recent generalist algorithms for many cellular morphologies. The winner of the Multimodality cell segmentation challenge published in 2024, a competition for the best generalist bioimage segmentation algorithms was a team led by Gihun Lee presenting their algorithm, MEDIAR[128,133]. The competition used 1000 images of multiple organisms from over 20 labs; 50 different experiments; using brightfield, fluorescent, PC and DIC microscopy techniques. Importantly, there was also a direct comparison between models submitted by the contestants, and models from the widely acclaimed Cellpose and Omnipose showcasing the higher accuracy of the entries.. Beyond segmentation, Deep learning has been used to predict morphologies of unlabeled cells, allowing researchers to simulate more tags than are currently possible to image at once and potentially extending the power of cytological profiling[144,145,146].
Segmented data availability (Table 3) is invaluable for scientific communities and accelerates new findings. By using published BCP data and mathematical modelling, the researchers uncovered the robustness of scaling behavior between cell surface area and volume in E. coli [75] and B. subtilis [76], inferred cell physiological alterations upon antibiotic treatments [147], and proposed a new antibiotic resistance pathway mediated by cell surface-to-volume ratio (S/V) transformations [74]. Therefore, the availability of BCP data is good practice and should be considered as a benchmark for all future BCP platforms, especially for pathogenic bacteria (Table 2).
9. Conclusions
Despite significant advances in research and the development of new tools, combating antimicrobial resistance (AMR) requires a multifaceted approach. Continued investment in research and development, global collaboration, and the effective implementation of surveillance and prevention strategies are crucial. Bacterial Cytological Profiling (BCP) stands out as a rapid and cost-effective technique that facilitates drug discovery by revealing the mechanism of action of novel antibacterial agents through detailed physiological and morphological analysis. Furthermore, BCP could be used to identify phenotypic changes when multiple antibiotics are used, revealing unique or overlapping cell morphologies induced by these combinations [41]. However, systematic explorations of cytological profiles for drug combinations are still missing.
Apart from bacteria, cytological profiling methods are also widely used for other organisms such as yeast[148,149], fungi [150], and human cells[151,152,153]. However, deep learning techniques employed for yeast and human cells, unfortunately, have not yet been applied to bacteria. Therefore, wider availability, applications and integration of machine learning tools across different scientific fields are needed.
Besides BCP being used to discover new antibiotics, BCP has been used to investigate complex interactions between bacteria and their predators – bacteriophages[10,11,12]. By leveraging high-resolution imaging, BCP enables the identification of metabolic pathways and cellular processes targeted by phages and antibiotics, both individually and in combination, offering valuable insights into the molecular mechanisms governing the phage-bacteria interaction, ultimately paving the way for more effective phage-based antibacterial therapies.
Acknowledgments
We gratefully acknowledge funding from BBSRC (BB/Y009002/1).
References
- Kim, C., Holm, M., Frost, I., Hasso-Agopsowicz, M. & Abbas, K. Global and regional burden of attributable and associated bacterial antimicrobial resistance avertable by vaccination: modelling study. BMJ Glob. Health 8, e011341 (2023). [CrossRef]
- Murray, C. J. L. et al. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. The Lancet 399, 629–655 (2022). [CrossRef]
- Centers for Disease Control and Prevention (U.S.). Antibiotic Resistance Threats in the United States, 2019. https://stacks.cdc.gov/view/cdc/82532 (2019) . [CrossRef]
- 160518_Final paper_with cover.pdf.
- GARDNER, A. D. Morphological Effects of Penicillin on Bacteria. Nature 146, 837–838 (1940). [CrossRef]
- Gardner, A. & others. Microscopical Effect of Penicillin on Spores and Vegetative Cells of Bacilli. Lancet 658–9 (1945). [CrossRef]
- Hutchings, M. I., Truman, A. W. & Wilkinson, B. Antibiotics: past, present and future. Curr. Opin. Microbiol. 51, 72–80 (2019).
- Walsh, C. Molecular mechanisms that confer antibacterial drug resistance. Nature 406, 775–781 (2000). [CrossRef]
- Luepke, K. H. et al. Past, Present, and Future of Antibacterial Economics: Increasing Bacterial Resistance, Limited Antibiotic Pipeline, and Societal Implications. Pharmacother. J. Hum. Pharmacol. Drug Ther. 37, 71–84 (2017). [CrossRef]
- Deep, A., Liang, Q., Enustun, E., Pogliano, J. & Corbett, K. D. Architecture and activation mechanism of the bacterial PARIS defence system. Nature 1–8 (2024) . [CrossRef]
- Tsunemoto, H., Sugie, J., Enustun, E., Pogliano, K. & Pogliano, J. Bacterial cytological profiling reveals interactions between jumbo phage φKZ infection and cell wall active antibiotics in Pseudomonas aeruginosa. PloS One 18, e0280070 (2023). [CrossRef]
- Birkholz, E. A. et al. An intron endonuclease facilitates interference competition between coinfecting viruses. Science 385, 105–112 (2024). [CrossRef]
- Corona, F. & Martinez, J. L. Phenotypic Resistance to Antibiotics. Antibiotics 2, 237–255 (2013). [CrossRef]
- Cotsonas King, A. & Wu, L. Macromolecular Synthesis and Membrane Perturbation Assays for Mechanisms of Action Studies of Antimicrobial Agents. Curr. Protoc. Pharmacol. 47, (2009). [CrossRef]
- Nonejuie, P., Burkart, M., Pogliano, K. & Pogliano, J. Bacterial cytological profiling rapidly identifies the cellular pathways targeted by antibacterial molecules. Proc. Natl. Acad. Sci. 110, 16169–16174 (2013). [CrossRef]
- Lima, L. M., Silva, B. N. M. da, Barbosa, G. & Barreiro, E. J. β-lactam antibiotics: An overview from a medicinal chemistry perspective. Eur. J. Med. Chem. 208, 112829 (2020). [CrossRef]
- Baquero, F. & Levin, B. R. Proximate and ultimate causes of the bactericidal action of antibiotics. Nat. Rev. Microbiol. 19, 123–132 (2021). [CrossRef]
- Reynolds, P. E. Structure, biochemistry and mechanism of action of glycopeptide antibiotics. Eur. J. Clin. Microbiol. Infect. Dis. 8, 943–950 (1989). [CrossRef]
- Jerala, R. Synthetic lipopeptides: a novel class of anti-infectives. Expert Opin. Investig. Drugs 16, 1159–1169 (2007). [CrossRef]
- O’Rourke, A. et al. Mechanism-of-Action Classification of Antibiotics by Global Transcriptome Profiling. Antimicrob. Agents Chemother. 64, e01207-19 (2020). [CrossRef]
- PubChem. Cerulenin. https://pubchem.ncbi.nlm.nih.gov/compound/5282054.
- Davis, B. D., Chen, L. L. & Tai, P. C. Misread protein creates membrane channels: an essential step in the bactericidal action of aminoglycosides. Proc. Natl. Acad. Sci. U. S. A. 83, 6164–6168 (1986). [CrossRef]
- Chopra, I. & Roberts, M. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance. Microbiol. Mol. Biol. Rev. 65, 232–260 (2001). [CrossRef]
- Vázquez-Laslop, N. & Mankin, A. S. How Macrolide Antibiotics Work. Trends Biochem. Sci. 43, 668–684 (2018). [CrossRef]
- Tenson, T., Lovmar, M. & Ehrenberg, M. The Mechanism of Action of Macrolides, Lincosamides and Streptogramin B Reveals the Nascent Peptide Exit Path in the Ribosome. J. Mol. Biol. 330, 1005–1014 (2003). [CrossRef]
- Swaney, S. M., Aoki, H., Ganoza, M. C. & Shinabarger, D. L. The oxazolidinone linezolid inhibits initiation of protein synthesis in bacteria. Antimicrob. Agents Chemother. 42, 3251–3255 (1998). [CrossRef]
- Correia, S., Poeta, P., Hébraud, M., Capelo, J. L. & Igrejas, G. Mechanisms of quinolone action and resistance: where do we stand? J. Med. Microbiol. 66, 551–559 (2017).
- Ojkic, N. et al. A Roadblock-and-Kill Mechanism of Action Model for the DNA-Targeting Antibiotic Ciprofloxacin. Antimicrob. Agents Chemother. 64, e02487-19 (2020). [CrossRef]
- Wong, W. R., Oliver, A. G. & Linington, R. G. Development of Antibiotic Activity Profile Screening for the Classification and Discovery of Natural Product Antibiotics. Chem. Biol. 19, 1483–1495 (2012). [CrossRef]
- Kohanski, M. A., Dwyer, D. J. & Collins, J. J. How antibiotics kill bacteria: from targets to networks. Nat. Rev. Microbiol. 8, 423–435 (2010). [CrossRef]
- WHO. AWaRe classification of antibiotics for evaluation and monitoring of use, 2023. (2023).
- Silver, L. L. Challenges of Antibacterial Discovery. Clin. Microbiol. Rev. 24, 71–109 (2011). [CrossRef]
- Hudson, M. A. & Lockless, S. W. Elucidating the Mechanisms of Action of Antimicrobial Agents. mBio 13, e02240-21. [CrossRef]
- Hage, D. S. et al. Pharmaceutical and biomedical applications of affinity chromatography: Recent trends and developments. J. Pharm. Biomed. Anal. 69, 93–105 (2012). [CrossRef]
- Franken, H. et al. Thermal proteome profiling for unbiased identification of direct and indirect drug targets using multiplexed quantitative mass spectrometry. Nat. Protoc. 10, 1567–1593 (2015). [CrossRef]
- Tacconelli, E. et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 18, 318–327 (2018). [CrossRef]
- Lewis, K. Platforms for antibiotic discovery. Nat. Rev. Drug Discov. 12, 371–387 (2013). [CrossRef]
- Lewis, K. et al. Sophisticated natural products as antibiotics. Nature 632, 39–49 (2024). [CrossRef]
- Gould, K. Antibiotics: from prehistory to the present day. J. Antimicrob. Chemother. 71, 572–575 (2016). [CrossRef]
- WHO. 2021 Antibacterial Agents in Clinical and Preclinical Development: An Overview and Analysis. (World Health Organization, 2022).
- Samernate, T. et al. High-Resolution Bacterial Cytological Profiling Reveals Intrapopulation Morphological Variations upon Antibiotic Exposure. Antimicrob. Agents Chemother. 67, e01307-22 (2023). [CrossRef]
- Balaban, N. Q. et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 17, 441–448 (2019). [CrossRef]
- Kussell, E., Kishony, R., Balaban, N. Q. & Leibler, S. Bacterial Persistence. Genetics 169, 1807–1814 (2005).
- Bailey, S. Principal Component Analysis with Noisy and/or Missing Data. Publ. Astron. Soc. Pac. 124, 1015–1023 (2012). [CrossRef]
- Van Den Broeck, T. et al. The Role of Single Nucleotide Polymorphisms in Predicting Prostate Cancer Risk and Therapeutic Decision Making. BioMed Res. Int. 2014, 1–16 (2014). [CrossRef]
- Regev, A. et al. The Human Cell Atlas. eLife 6, e27041 (2017).
- Liu, Z., Liu, J., Ghosh, S., Han, J. & Scarlett, J. Generative Principal Component Analysis. (2022).
- Ghadban, N., Honeine, P., Mourad-Chehade, F., Francis, C. & Farah, J. Diffusion strategies for in-network principal component analysis. in 2014 IEEE International Workshop on Machine Learning for Signal Processing (MLSP) 1–6 (IEEE, Reims, France, 2014). [CrossRef]
- Siirtola, H., Saily, T. & Nevalainen, T. Interactive Principal Component Analysis. in 2017 21st International Conference Information Visualisation (IV) 416–421 (IEEE, London, 2017). [CrossRef]
- Cushnie, T. P. T., O’Driscoll, N. H. & Lamb, A. J. Morphological and ultrastructural changes in bacterial cells as an indicator of antibacterial mechanism of action. Cell. Mol. Life Sci. 73, 4471–4492 (2016). [CrossRef]
- Gebicki, J. M. & James, A. M. The Preparation and Properties of Spheroplasts of Aerobacter aerogenes. J. Gen. Microbiol. 23, 9–18 (1960). [CrossRef]
- Errington, J. L-form bacteria, cell walls and the origins of life. Open Biol. 3, 120143 (2013). [CrossRef]
- Allan, E. J., Hoischen, C. & Gumpert, J. Bacterial L-forms. Adv. Appl. Microbiol. 68, 1–39 (2009).
- Mercier, R., Kawai, Y. & Errington, J. General principles for the formation and proliferation of a wall-free (L-form) state in bacteria. eLife 3, e04629 (2014). [CrossRef]
- Errington, J. Cell wall-deficient, L-form bacteria in the 21st century: a personal perspective. Biochem. Soc. Trans. 45, 287–295 (2017). [CrossRef]
- Spratt, B. G. Distinct penicillin binding proteins involved in the division, elongation, and shape of Escherichia coli K12. Proc. Natl. Acad. Sci. U. S. A. 72, 2999–3003 (1975). [CrossRef]
- Spratt, B. G. & Pardee, A. B. Penicillin-binding proteins and cell shape in E. coli. Nature 254, 516–517 (1975). [CrossRef]
- Curtis, N. A., Orr, D., Ross, G. W. & Boulton, M. G. Affinities of penicillins and cephalosporins for the penicillin-binding proteins of Escherichia coli K-12 and their antibacterial activity. Antimicrob. Agents Chemother. 16, 533–539 (1979). [CrossRef]
- Di Modugno, E. et al. In vitro activity of the tribactam GV104326 against gram-positive, gram-negative, and anaerobic bacteria. Antimicrob. Agents Chemother. 38, 2362–2368 (1994). [CrossRef]
- Bernabeu-Wittel, M. et al. Morphological changes induced by imipenem and meropenem at sub-inhibitory concentrations in Acinetobacter baumannii. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 10, 931–934 (2004). [CrossRef]
- Differences in mode of action of (β-lactam antibiotics influence morphology, LPS release and in vivo antibiotic efficacy. CoLab https://colab.ws/articles/10.1177%2F096805199600300306.
- de Pedro, M. A., Donachie, W. D., Höltje, J.-V. & Schwarz, H. Constitutive Septal Murein Synthesis in Escherichia coli with Impaired Activity of the Morphogenetic Proteins RodA and Penicillin-Binding Protein 2. J. Bacteriol. 183, 4115–4126 (2001). [CrossRef]
- Sumita, Y., Fukasawa, M. & Okuda, T. Comparison of two carbapenems, SM-7338 and imipenem: affinities for penicillin-binding proteins and morphological changes. J. Antibiot. (Tokyo) 43, 314–320 (1990).
- Dalhoff, A., Nasu, T. & Okamoto, K. Target affinities of faropenem to and its impact on the morphology of gram-positive and gram-negative bacteria. Chemotherapy 49, 172–183 (2003). [CrossRef]
- Horii, T., Kobayashi, M., Sato, K., Ichiyama, S. & Ohta, M. An in-vitro study of carbapenem-induced morphological changes and endotoxin release in clinical isolates of gram-negative bacilli. J. Antimicrob. Chemother. 41, 435–442 (1998). [CrossRef]
- Perumalsamy, H., Jung, M. Y., Hong, S. M. & Ahn, Y.-J. Growth-Inhibiting and morphostructural effects of constituents identified in Asarum heterotropoides root on human intestinal bacteria. BMC Complement. Altern. Med. 13, 245 (2013). [CrossRef]
- Nickerson, W. J. & Webb, M. Effect of folic acid analogues on growth and cell division of nonexacting microorganisms. J. Bacteriol. 71, 129–139 (1956). [CrossRef]
- Elliott, T. S. J., Shelton, A. & Greenwood, D. The response of Escherichia coli to ciprofloxacin and norfloxacin. J. Med. Microbiol. 23, 83–88 (1987). [CrossRef]
- Chen, K., Sun, G. W., Chua, K. L. & Gan, Y.-H. Modified Virulence of Antibiotic-Induced Burkholderia pseudomallei Filaments. Antimicrob. Agents Chemother. 49, 1002–1009 (2005). [CrossRef]
- Uphoff, S., Reyes-Lamothe, R., Garza de Leon, F., Sherratt, D. J. & Kapanidis, A. N. Single-molecule DNA repair in live bacteria. Proc. Natl. Acad. Sci. 110, 8063–8068 (2013). [CrossRef]
- Jones, E. C. & Uphoff, S. Single-molecule imaging of LexA degradation in Escherichia coli elucidates regulatory mechanisms and heterogeneity of the SOS response. Nat. Microbiol. 6, 981–990 (2021). [CrossRef]
- Jaramillo-Riveri, S. et al. Growth-dependent heterogeneity in the DNA damage response in Escherichia coli. Mol. Syst. Biol. 18, e10441 (2022).
- Banerjee, S. et al. Mechanical feedback promotes bacterial adaptation to antibiotics. Nat. Phys. 17, 403–409 (2021). [CrossRef]
- Ojkic, N., Serbanescu, D. & Banerjee, S. Antibiotic Resistance via Bacterial Cell Shape-Shifting. mBio 13, e00659-22 (2022). [CrossRef]
- Ojkic, N., Serbanescu, D. & Banerjee, S. Surface-to-volume scaling and aspect ratio preservation in rod-shaped bacteria. eLife 8, e47033 (2019). [CrossRef]
- Ojkic, N. & Banerjee, S. Bacterial cell shape control by nutrient-dependent synthesis of cell division inhibitors. Biophys. J. 120, 2079–2084 (2021). [CrossRef]
- Lin, L. et al. Azithromycin Synergizes with Cationic Antimicrobial Peptides to Exert Bactericidal and Therapeutic Activity Against Highly Multidrug-Resistant Gram-Negative Bacterial Pathogens. EBioMedicine 2, 690–698 (2015). [CrossRef]
- Mohammad, H. et al. Bacteriological profiling of diphenylureas as a novel class of antibiotics against methicillin-resistant Staphylococcus aureus. PLOS ONE 12, e0182821 (2017). [CrossRef]
- Wu, Y. & Seyedsayamdost, M. R. The Polyene Natural Product Thailandamide A Inhibits Fatty Acid Biosynthesis in Gram-Positive and Gram-Negative Bacteria. Biochemistry 57, 4247–4251 (2018). [CrossRef]
- Sun, Y., Heidary, D. K., Zhang, Z., Richards, C. I. & Glazer, E. C. Bacterial Cytological Profiling Reveals the Mechanism of Action of Anticancer Metal Complexes. Mol. Pharm. 15, 3404–3416 (2018). [CrossRef]
- Dillon, N. A. et al. Characterizing the response of Acinetobacter baumannii ATCC 17978 to azithromycin in multiple in vitro growth conditions. Preprint a (2020). [CrossRef]
- Araújo-Bazán, L., Ruiz-Avila, L. B., Andreu, D., Huecas, S. & Andreu, J. M. Cytological Profile of Antibacterial FtsZ Inhibitors and Synthetic Peptide MciZ. Front. Microbiol. 7, (2016). [CrossRef]
- Andreu, J. M., Huecas, S., Araújo-Bazán, L., Vázquez-Villa, H. & Martín-Fontecha, M. The Search for Antibacterial Inhibitors Targeting Cell Division Protein FtsZ at Its Nucleotide and Allosteric Binding Sites. Biomedicines 10, 1825 (2022). [CrossRef]
- Soonthonsrima, T. et al. Phage-induced bacterial morphological changes reveal a phage-derived antimicrobial affecting cell wall integrity. Antimicrob. Agents Chemother. 67, e00764-23 (2023). [CrossRef]
- World Healt Organization. WHO Bacterial Priority Pathogens List, 2017. (2017).
- Di Bella, S. et al. Resistance to ceftazidime/avibactam in infections and colonisations by KPC-producing Enterobacterales: a systematic review of observational clinical studies. J. Glob. Antimicrob. Resist. 25, 268–281 (2021).
- Butler, M. S. et al. Analysis of the Clinical Pipeline of Treatments for Drug-Resistant Bacterial Infections: Despite Progress, More Action Is Needed. Antimicrob. Agents Chemother. 66, e01991-21. [CrossRef]
- Geneva: World Health Organization. WHO Bacterial Priority Pathogens List, 2024: bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance. (2024).
- Htoo, H. H. et al. Bacterial Cytological Profiling as a Tool To Study Mechanisms of Action of Antibiotics That Are Active against Acinetobacter baumannii. Antimicrob. Agents Chemother. 63, 10.1128/aac.02310-18 (2019). [CrossRef]
- Coram, M. A. et al. Morphological Characterization of Antibiotic Combinations. ACS Infect. Dis. 8, 66–77 (2022). [CrossRef]
- Montaño, E. T. et al. Bacterial Cytological Profiling Identifies Rhodanine-Containing PAINS Analogs as Specific Inhibitors of Escherichia coli Thymidylate Kinase In Vivo. J. Bacteriol. 203, 10.1128/jb.00105-21 (2021). [CrossRef]
- Nonejuie, P. et al. Application of bacterial cytological profiling to crude natural product extracts reveals the antibacterial arsenal of Bacillus subtilis. J. Antibiot. (Tokyo) 69, 353–361 (2016). [CrossRef]
- Sun, D. et al. Intrinsic Antibacterial Activity of Xeruborbactam In Vitro: Assessing Spectrum and Mode of Action. Antimicrob. Agents Chemother. 66, e00879-22. [CrossRef]
- Sridhar, S. et al. High-Content Imaging to Phenotype Antimicrobial Effects on Individual Bacteria at Scale. mSystems 6, e00028-21. [CrossRef]
- Smith, T. C. et al. Morphological profiling of tubercle bacilli identifies drug pathways of action. Proc. Natl. Acad. Sci. U. S. A. 117, 18744–18753 (2020). [CrossRef]
- Allen, R. et al. An arylsulphonamide that targets cell wall biosynthesis in Mycobacterium tuberculosis. 2024.07.22.604653 Preprint a (2024). [CrossRef]
- López-Jiménez, A. T. et al. High-content high-resolution microscopy and deep learning assisted analysis reveals host and bacterial heterogeneity during Shigella infection. eLife 13, (2024).
- Werth, B. J. et al. Defining Daptomycin Resistance Prevention Exposures in Vancomycin-Resistant Enterococcus faecium and E. faecalis. Antimicrob. Agents Chemother. 58, 5253–5261 (2014). [CrossRef]
- Quach, D. T., Sakoulas, G., Nizet, V., Pogliano, J. & Pogliano, K. Bacterial Cytological Profiling (BCP) as a Rapid and Accurate Antimicrobial Susceptibility Testing Method for Staphylococcus aureus. EBioMedicine 4, 95–103 (2016). [CrossRef]
- Kalla, G. Using Bacterial Cytological Profiling to Study the Interactions of Bacteria and the Defense Systems of Multicellular Eukaryotes. (University of California San Diego, USA, 2020).
- Blaskovich, M. A. T. et al. The antimicrobial potential of cannabidiol. Commun. Biol. 4, 1–18 (2021). [CrossRef]
- Sakoulas, G. et al. Examining the Use of Ceftaroline in the Treatment of Streptococcus pneumoniae Meningitis with Reference to Human Cathelicidin LL-37. Antimicrob. Agents Chemother. 59, 2428–2431 (2015). [CrossRef]
- Schäfer, A.-B. et al. Dissecting antibiotic effects on the cell envelope using bacterial cytological profiling: a phenotypic analysis starter kit. Microbiol. Spectr. 12, e03275-23 (2024). [CrossRef]
- Serbanescu, D., Ojkic, N. & Banerjee, S. Cellular resource allocation strategies for cell size and shape control in bacteria. FEBS J. 289, 7891–7906 (2022). [CrossRef]
- Htoo, H. H. et al. Bacterial Cytological Profiling as a Tool To Study Mechanisms of Action of Antibiotics That Are Active against Acinetobacter baumannii. Antimicrob. Agents Chemother. 63, 10.1128/aac.02310-18 (2019). [CrossRef]
- Samernate, T. et al. High-Resolution Bacterial Cytological Profiling Reveals Intrapopulation Morphological Variations upon Antibiotic Exposure. Antimicrob. Agents Chemother. 67, e01307-22 (2023). [CrossRef]
- Tsunemoto, H., Sugie, J., Enustun, E., Pogliano, K. & Pogliano, J. Bacterial cytological profiling reveals interactions between jumbo phage φKZ infection and cell wall active antibiotics in Pseudomonas aeruginosa. PLOS ONE 18, e0280070 (2023). [CrossRef]
- Lamsa, A. et al. Rapid Inhibition Profiling in Bacillus subtilis to Identify the Mechanism of Action of New Antimicrobials. ACS Chem. Biol. 11, 2222–2231 (2016). [CrossRef]
- Herschede, S. R., Salam, R., Gneid, H. & Busschaert, N. Bacterial cytological profiling identifies transmembrane anion transport as the mechanism of action for a urea-based antibiotic. Supramol. Chem. 34, 26–33 (2022). [CrossRef]
- Schäfer, A.-B. et al. Dissecting antibiotic effects on the cell envelope using bacterial cytological profiling: a phenotypic analysis starter kit. Microbiol. Spectr. 12, e03275-23 (2024). [CrossRef]
- Araújo-Bazán, L., Ruiz-Avila, L. B., Andreu, D., Huecas, S. & Andreu, J. M. Cytological Profile of Antibacterial FtsZ Inhibitors and Synthetic Peptide MciZ. Front. Microbiol. 7, (2016). [CrossRef]
- Santos, T. M. A. et al. Small Molecule Chelators Reveal That Iron Starvation Inhibits Late Stages of Bacterial Cytokinesis. ACS Chem. Biol. 13, 235–246 (2018). [CrossRef]
- Montaño, E. T. et al. Bacterial Cytological Profiling Identifies Rhodanine-Containing PAINS Analogs as Specific Inhibitors of Escherichia coli Thymidylate Kinase In Vivo. J. Bacteriol. 203, 10.1128/jb.00105-21 (2021). [CrossRef]
- Ulloa, E. R. et al. Azithromycin Exerts Bactericidal Activity and Enhances Innate Immune Mediated Killing of MDR Achromobacter xylosoxidans. Infect. Microbes Dis. 2, 10 (2020). [CrossRef]
- de Wet, T. J., Winkler, K. R., Mhlanga, M., Mizrahi, V. & Warner, D. F. Arrayed CRISPRi and quantitative imaging describe the morphotypic landscape of essential mycobacterial genes. eLife 9, e60083 (2020). [CrossRef]
- Smith, T. C. et al. Morphological profiling of tubercle bacilli identifies drug pathways of action. Proc. Natl. Acad. Sci. 117, 18744–18753 (2020). [CrossRef]
- Mayer, B., Schwan, M., Thormann, K. M. & Graumann, P. L. Antibiotic Drug screening and Image Characterization Toolbox (A.D.I.C.T.): a robust imaging workflow to monitor antibiotic stress response in bacterial cells in vivo. F1000Research 10, 277 (2022).
- Ouyang, X. et al. Classification of antimicrobial mechanism of action using dynamic bacterial morphology imaging. Sci. Rep. 12, 11162 (2022). [CrossRef]
- Mistretta, M. et al. Dynamic microfluidic single-cell screening identifies pheno-tuning compounds to potentiate tuberculosis therapy. Nat. Commun. 15, 4175 (2024). [CrossRef]
- El-sagheir, A. M. K. et al. Rational design, synthesis, molecular modeling, biological activity, and mechanism of action of polypharmacological norfloxacin hydroxamic acid derivatives. RSC Med. Chem. 14, 2593–2610 (2023).
- Kuru, E. et al. In Situ Probing of Newly Synthesized Peptidoglycan in Live Bacteria with Fluorescent D -Amino Acids. Angew. Chem. Int. Ed. 51, 12519–12523 (2012). [CrossRef]
- Vollmer, W., Blanot, D. & de Pedro, M. A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 32, 149–167 (2008). [CrossRef]
- Young, K. D. The Selective Value of Bacterial Shape. Microbiol. Mol. Biol. Rev. 70, 660–703 (2006). [CrossRef]
- Stratford, J. P. et al. Electrically induced bacterial membrane-potential dynamics correspond to cellular proliferation capacity. Proc. Natl. Acad. Sci. 116, 9552–9557 (2019). [CrossRef]
- Prindle, A. et al. Ion channels enable electrical communication in bacterial communities. Nature 527, 59–63 (2015). [CrossRef]
- De Souza-Guerreiro, T. C. et al. Membrane potential dynamics unveil the promise of bioelectrical antimicrobial susceptibility Testing (BeAST) for anti-fungal screening. mBio 15, e01302-24 (2024).
- Wong, F. et al. Cytoplasmic condensation induced by membrane damage is associated with antibiotic lethality. Nat. Commun. 12, 2321 (2021). [CrossRef]
- Ma, J. et al. The multimodality cell segmentation challenge: toward universal solutions. Nat. Methods 21, 1103–1113 (2024). [CrossRef]
- Cutler, K. J., Stringer, C., Wiggins, P. A. & Mougous, J. D. Omnipose: a high-precision morphology-independent solution for bacterial cell segmentation. [CrossRef]
- Bali, A. & Singh, S. N. A Review on the Strategies and Techniques of Image Segmentation. in 2015 Fifth International Conference on Advanced Computing & Communication Technologies 113–120 (2015). [CrossRef]
- Stringer, C., Wang, T., Michaelos, M. & Pachitariu, M. Cellpose: a generalist algorithm for cellular segmentation. Nat. Methods 2020 181 18, 100–106 (2020). [CrossRef]
- StarDist: Application of the deep-learning tool for phase-contrast cell images - 2020 - Wiley Analytical Science. Analytical Science Article DO Series https://analyticalscience.wiley.com/do/10.1002/was.000400090/.
- Lou, W. et al. Multi-stream Cell Segmentation with Low-level Cues for Multi-modality Images. Preprint at http://arxiv.org/abs/2310.14226 (2023).
- Paintdakhi, A. et al. Oufti: An integrated software package for high-accuracy, high-throughput quantitative microscopy analysis. Mol. Microbiol. 99, 767–777 (2016).
- Danielsen, J. & Nordenfelt, P. Computer Vision-Based Image Analysis of Bacteria. in Bacterial Pathogenesis: Methods and Protocols (eds. Nordenfelt, P. & Collin, M.) 161–172 (Springer, New York, NY, 2017). [CrossRef]
- THE ANALYSIS OF CELL IMAGES* - Prewitt - 1966 - Annals of the New York Academy of Sciences - Wiley Online Library. https://nyaspubs.onlinelibrary.wiley.com/doi/abs/10.1111/j.1749-6632.1965.tb11715.x.
- Mendelsohn, M. L., Kolman, W. A., Perry, B. & Prewitt, J. M. S. Computer Analysis of Cell Images. Postgrad. Med. 38, 567–573 (1965). [CrossRef]
- Stylianidou, S., Brennan, C., Nissen, S. B., Kuwada, N. J. & Wiggins, P. A. SuperSegger: robust image segmentation, analysis and lineage tracking of bacterial cells. Mol. Microbiol. 102, 690–700 (2016). [CrossRef]
- Chai, B., Efstathiou, C., Yue, H. & Draviam, V. M. Opportunities and challenges for deep learning in cell dynamics research. Trends Cell Biol. 0, (2023). [CrossRef]
- Jan, M., Spangaro, A., Lenartowicz, M. & Mattiazzi Usaj, M. From pixels to insights: Machine learning and deep learning for bioimage analysis. BioEssays 46, 2300114 (2024). [CrossRef]
- O’Mahony, N. et al. Deep Learning vs. Traditional Computer Vision. in Advances in Computer Vision (eds. Arai, K. & Kapoor, S.) 128–144 (Springer International Publishing, Cham, 2020). [CrossRef]
- Falk, T. et al. U-Net: deep learning for cell counting, detection, and morphometry. Nat. Methods 16, 67–70 (2019). [CrossRef]
- Panigrahi, S. et al. Misic, a general deep learning-based method for the high-throughput cell segmentation of complex bacterial communities. eLife https://elifesciences.org/articles/65151 (2021). [CrossRef]
- Osokin, A., Chessel, A., Salas, R. E. C. & Vaggi, F. GANs for Biological Image Synthesis. in 2017 IEEE International Conference on Computer Vision (ICCV) 2252–2261 (2017). doi:10.1109/ICCV.2017.245.
- Chan, C. K., Hadjitheodorou, A., Tsai, T. Y.-C. & Theriot, J. A. Quantitative comparison of principal component analysis and unsupervised deep learning using variational autoencoders for shape analysis of motile cells. 2020.06.26.174474 Preprint a (2020). [CrossRef]
- Goldsborough, P., Pawlowski, N., Caicedo, J. C., Singh, S. & Carpenter, A. E. CytoGAN: Generative Modeling of Cell Images. 227645 Preprint a (2017). [CrossRef]
- Cylke, C., Si, F. & Banerjee, S. Effects of antibiotics on bacterial cell morphology and their physiological origins. Biochem. Soc. Trans. 50, 1269–1279 (2022). [CrossRef]
- Chessel, A. & Carazo Salas, R. E. From observing to predicting single-cell structure and function with high-throughput/high-content microscopy. Essays Biochem. 63, 197–208 (2019). [CrossRef]
- Chong, Y. T. et al. Yeast Proteome Dynamics from Single Cell Imaging and Automated Analysis. Cell 161, 1413–1424 (2015). [CrossRef]
- McMahon, C. L. et al. Development of an Imaging Flow Cytometry Method for Fungal Cytological Profiling and Its Potential Application in Antifungal Drug Development. J. Fungi 9, 722 (2023). [CrossRef]
- McDiarmid, A. H. et al. Morphological profiling in human neural progenitor cells classifies hits in a pilot drug screen for Alzheimer’s disease. Brain Commun. 6, fcae101 (2024). [CrossRef]
- Ren, E. et al. Deep Learning-Enhanced Morphological Profiling Predicts Cell Fate Dynamics in Real-Time in hPSCs. (2021). [CrossRef]
- Perlman, Z. E. et al. Multidimensional Drug Profiling By Automated Microscopy. Science 306, 1194–1198 (2004). [CrossRef]
Figure 1.
Antibiotic targets in bacteria. Antibiotics typically kill bacteria by targeting at least one of the five cell components: cell wall, cell membrane, ribosome, DNA and RNA. Antibiotics interfere with the synthesis of or directly damage cell structures to inhibit bacterial growth or irreversible damage bacterial integrity. Antibiotics can inhibit the synthesis of some essential components, such as folate synthesis, a precursor for DNA synthesis. Antibiotics are transported into cells across the cell membranes or through membrane porins that facilitate antibiotic transport. Bacterial cells can also eliminate antibiotics by using efflux pumps, which are transport proteins that actively expel antibiotics out of the cell.
Figure 1.
Antibiotic targets in bacteria. Antibiotics typically kill bacteria by targeting at least one of the five cell components: cell wall, cell membrane, ribosome, DNA and RNA. Antibiotics interfere with the synthesis of or directly damage cell structures to inhibit bacterial growth or irreversible damage bacterial integrity. Antibiotics can inhibit the synthesis of some essential components, such as folate synthesis, a precursor for DNA synthesis. Antibiotics are transported into cells across the cell membranes or through membrane porins that facilitate antibiotic transport. Bacterial cells can also eliminate antibiotics by using efflux pumps, which are transport proteins that actively expel antibiotics out of the cell.
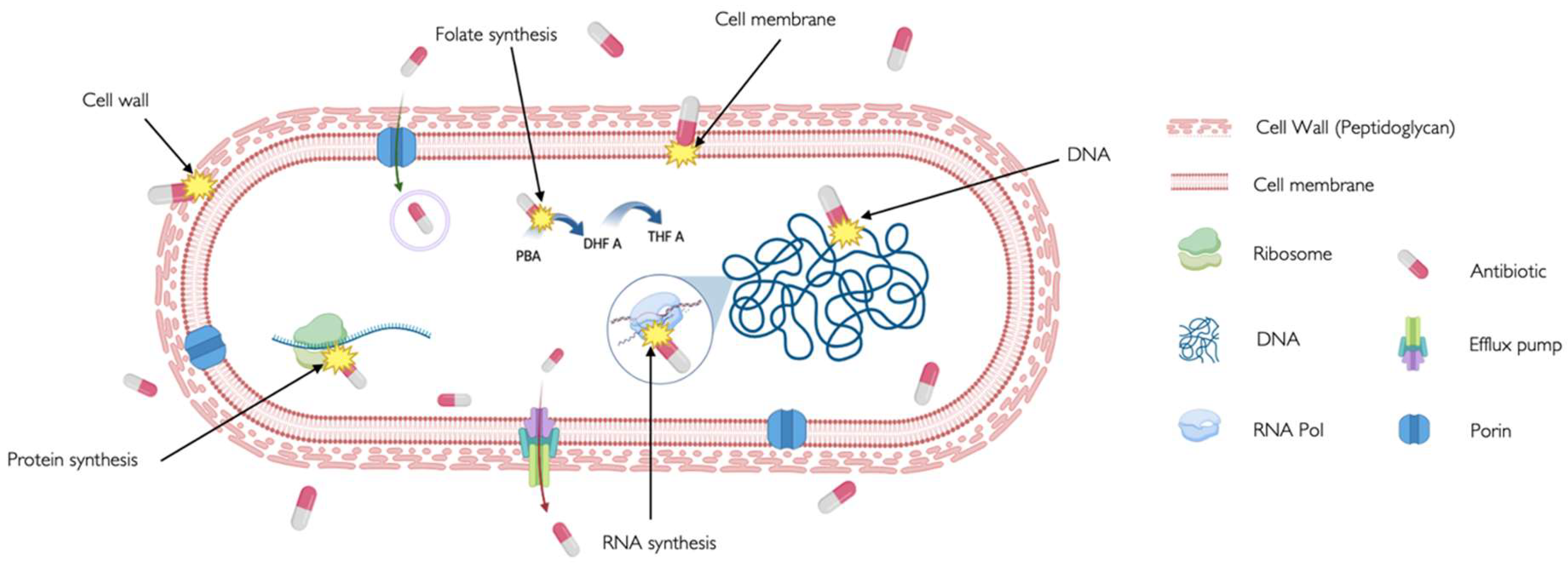
Figure 2.
Bacterial Cytological Profiling. (A) Different morphologies adopted by bacteria under the stress of antibiotics targeting different pathways. The cartoon drawings are based on the microscopy images from Poochit et al [15]., where bacterial cells were treated with antibiotics targeting five major biosynthetic pathways (DNA, Ribosome, RNA, Cell Wall, Membrane), using fluorescent dyes FM4-64 (red) and DAPI (blue) to stain bacterial membranes and DNA respectively. Scale bar, 1 μm. (B) Principal Component Analysis (PCA) is used to cluster different bacterial cell shapes based on the antibiotic mechanism of action. Each point on the graph represents a single cell. The graph also illustrates that when green dots, representing a characteristic morphology of a DNA-targeting antibiotic, cluster with orange dots, representing untreated bacteria, it indicates no morphological change and suggests possible antibiotic resistance or persistence[42,43].
Figure 2.
Bacterial Cytological Profiling. (A) Different morphologies adopted by bacteria under the stress of antibiotics targeting different pathways. The cartoon drawings are based on the microscopy images from Poochit et al [15]., where bacterial cells were treated with antibiotics targeting five major biosynthetic pathways (DNA, Ribosome, RNA, Cell Wall, Membrane), using fluorescent dyes FM4-64 (red) and DAPI (blue) to stain bacterial membranes and DNA respectively. Scale bar, 1 μm. (B) Principal Component Analysis (PCA) is used to cluster different bacterial cell shapes based on the antibiotic mechanism of action. Each point on the graph represents a single cell. The graph also illustrates that when green dots, representing a characteristic morphology of a DNA-targeting antibiotic, cluster with orange dots, representing untreated bacteria, it indicates no morphological change and suggests possible antibiotic resistance or persistence[42,43].
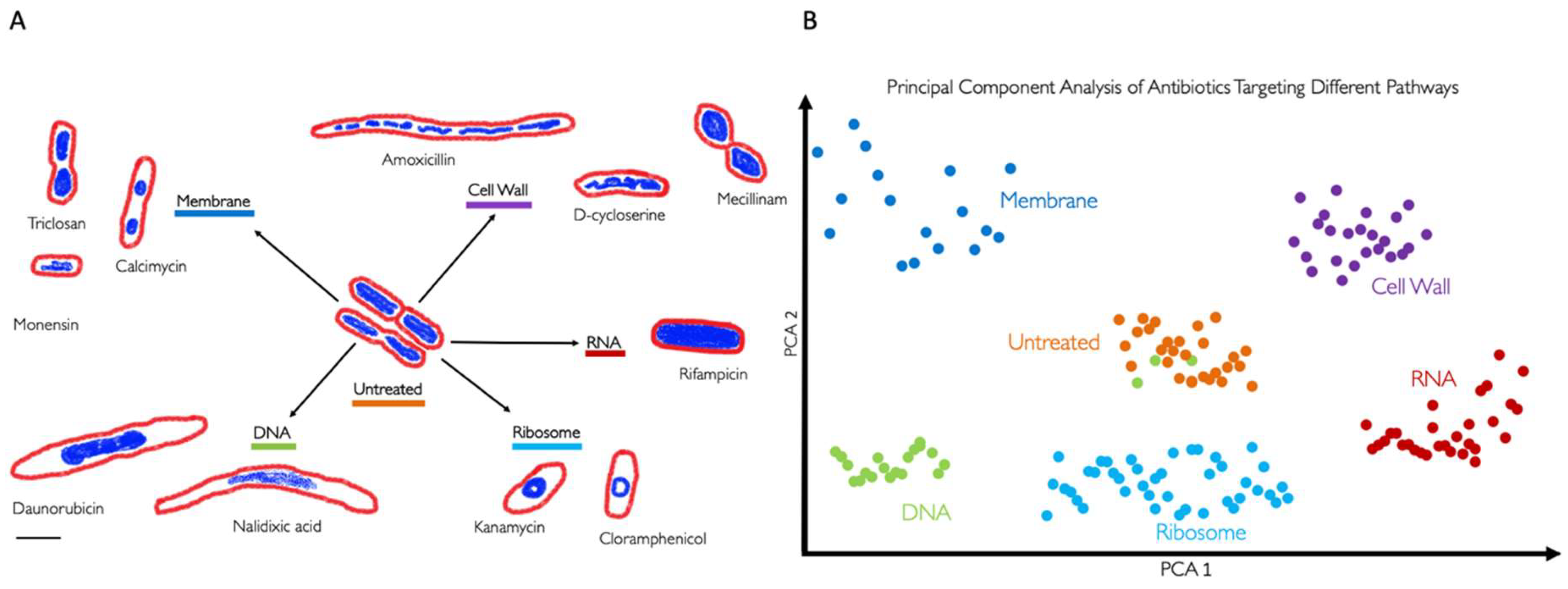
Figure 3.
Representation of Principal Component Analysis (PCA) using bacterial morphologies to determine the mechanism of action (MOA) of a novel antibiotic. Arrow 1 indicates the antibiotic used against certain bacteria, which can change their shape depending on the antibiotic's MOA. If the bacteria exhibit a morphology as indicated by arrow 2, the antibiotic targets the membrane. Conversely, if the bacteria display a morphology as indicated by arrow 3, the antibiotic targets RNA. However, if the morphology is completely different from the known and clustered morphologies, as shown by arrow 4, it suggests that the antibiotic targets a novel pathway and if the bacteria do not show any change, it suggests that they are not susceptible to this antibiotic or, in the worst-case scenario, that they are resistant to the antibiotic.
Figure 3.
Representation of Principal Component Analysis (PCA) using bacterial morphologies to determine the mechanism of action (MOA) of a novel antibiotic. Arrow 1 indicates the antibiotic used against certain bacteria, which can change their shape depending on the antibiotic's MOA. If the bacteria exhibit a morphology as indicated by arrow 2, the antibiotic targets the membrane. Conversely, if the bacteria display a morphology as indicated by arrow 3, the antibiotic targets RNA. However, if the morphology is completely different from the known and clustered morphologies, as shown by arrow 4, it suggests that the antibiotic targets a novel pathway and if the bacteria do not show any change, it suggests that they are not susceptible to this antibiotic or, in the worst-case scenario, that they are resistant to the antibiotic.
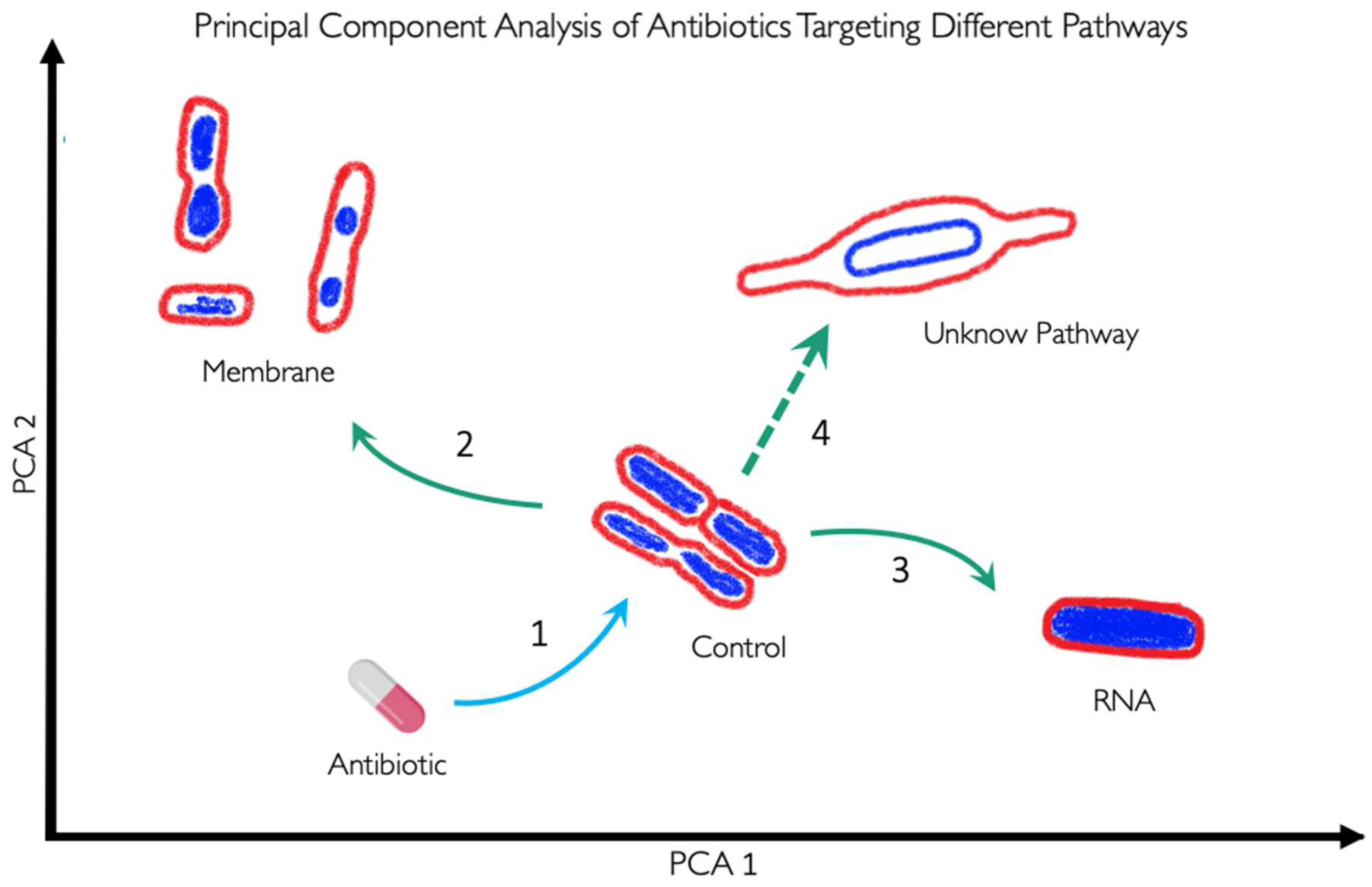

Table 1.
General classification of antibiotics based on their target and chemical structure, including their mechanism of action (MOA), examples of each antibiotic type, and their typical clinical uses.
Table 1.
General classification of antibiotics based on their target and chemical structure, including their mechanism of action (MOA), examples of each antibiotic type, and their typical clinical uses.
| TARGET | CHEMICAL STRUCTURE | MECHANISM OF ACTION (MOA) | GENERIC NAME EXAMPLES | USE |
|---|---|---|---|---|
| CELL WALL | β-Lactams | Inhibit penicillin-binding proteins (PBPs) that crosslink peptidoglycan chains in the bacterial cell wall [16], disrupting cell wall integrity and causing cell lysis [17]. | Penicillins, cephalosporins, cephamycins, carbapenems, and others. | To treat a variety of infections, including skin infections, chest infections, urinary tract infections sepsis and meningitis. |
| Glycopeptides | Target gram-positive bacteria by binding to the acyl-D-Ala-D-Ala terminus to the growing peptidoglycan and then cross-linking peptides within and between peptidoglycan [18]. | Vancomicyn | Last resort medication for the treatment of sepsis and lower respiratory tract, skin, and bone infections caused by Gram-positive bacteria. | |
| MEBRANE | Lipopeptides | Insert in the cell membrane and cause depolarization, reducing the ability to create ATP and cell death [19]. | Daptomycin, Colistin | For treatment of complicated skin and skin-structure infections associated to Gram-positive bacteria. |
| FATTY ACID SYNTHESIS | Chlorophenol | Block the reduction step of the fatty acid synthesis pathway by inhibiting an enoyl-ACP reductase (fabI) [20]. | Triclosan | Added to many consumer products as soaps, body washes and toothpastes, intended to reduce or prevent bacterial contamination. |
| Oxirane carboxylic acids | Irreversibly binds to fatty acid synthase, specifically b-ketoacyl-acyl carrier protein synthase. In sterol synthesis, inhibits HMG-CoA synthetase activity [21]. |
Cerulenin | Antifungal agent whose activity interferes with or otherwise acts to prevent the formation of fatty acids and sterols. With selective cytotoxicity to cancer cells | |
| PROTEIN SYNTHESIS | Aminoglycosides | Interact with the 30s ribosomal subunit of 16S RNA causing misreading and/or truncated proteins and cell death [17], [22]. | Gentamicin, tobramicin, kanamycin | To treat mainly very serious illnesses and infections such as sepsis, as they can cause very serious side effects. |
| Tetracyclines | Inhibit translation by binding to 16S rRNA of the 30S ribosomal subunit, preventing tRNA binding to 30S [23]. | Tetracycline, doxycycline and lymecycline | To treat a wide range of infections, acne and skin conditions as rosacea. | |
| Macrolides | It binds to the 23S rRNA of the 50S ribosomal subunit, leading to the production of incomplete peptide chains [24]. | Azithromicin, erythromycin and clarithromicyn | Particularly useful to treat lung and chest infections, as an alternative for people with a penicillin allergy or penicillin-resistant strains. | |
| Lincosamide | It binds to the 50S ribosome subunit to stimulate dissociation of the peptidyl- tRNA molecule from the ribosomes during elongation [25] | Clindamicyn | Primarily used to treat gram-positive bacterial infections in which there is resistance or intolerance to penicillin. | |
| Oxazolidinones | Limit translation by binding to 23S rRNA of the 50S subunit and preventing the formation of a functional 70S subunit [26]. | Linezolid | Active against multidrug-resistant staphylococci, streptococci, and enterococci. | |
| DNA SYNTHESIS | Fluoroquinoles | Inhibit DNA replication by targeting DNA gyrase and topoisomerase IV[27,28]. | Ciprofloxacin and levofloxacin | Broad-spectrum antibiotics that are used to treat a wide range of infections, especially respiratory and urinary tract infections. Not commonly used due to their risk of serious side effects. |
| Sulfonamides | Competitive inhibitor of Dihydropteroate synthase (DHPS) involved in folate synthesis [29] | Sulfamethazine, sulfapyridine | Utilized in the treatment of tonsillitis, septicemia, meningococcal meningitis, bacillary dysentery, and number of infections of urinary tract | |
| RNA SYNTHESIS | Rifamycins | It binds to the RNA polymerase and blocks the RNA synthesis [30] | Rifapentine, Rifampin | Effective against mycobacteria, and are therefore used to treat tuberculosis, leprosy, and mycobacterium avium complex (MAC) infections. |
Table 2.
BCP Analysis for the WHO Bacterial Priority Pathogens List (2024). In third column, this table indicates whether bacterial have been studied using Bacterial Cytological Profiling (BCP) or not. We consider any BCP done in the wild-type strain rather than in the resistant strain.
Table 2.
BCP Analysis for the WHO Bacterial Priority Pathogens List (2024). In third column, this table indicates whether bacterial have been studied using Bacterial Cytological Profiling (BCP) or not. We consider any BCP done in the wild-type strain rather than in the resistant strain.
| Bacteria | Resistant to | Bacterial Cytological Profilling (BCP) |
|---|---|---|
| Priority 1. Critical group | ||
| Acinetobacter baumannii | Carbapenems | Yes[41,77,81,89] |
| Enterobacteriaceae* | Third generation cephalosporine | Yes[15,80,90,91,92] |
| Enterobacteriaceae** | Carbapenems, ESBL-producing | Yes[77,93,94] |
| Rifampicin-Resistant Tuberculosis (RR-TB)*** | Rifampicin | Yes[95,96] |
| Priority 2. High group | ||
| Salmonella Thypi | Fluoroquinolones | Yes [94] |
| Shigella spp. | Fluoroquinolones | Yes [97] |
| Enterococcus faecium | Vancomycin | Yes [98] |
| Pseudomonas aeruginosa | Carbapenems | Yes[11,77,93,94] |
| Non-typhoidal Salmonella | Fluoroquinoles | No |
| Neisseria gonorrhoeae | Cephalosporin, Fluoroquinolonas | No |
| Staphylococcus aureus | Methicillin and vancomycin | Yes[78,99,100,101] |
| Priority 3. Medium group | ||
| Group A Streptococci | Macrolide | No |
| Streptococcus pneumoniae | Macrolide/No sensitivity to penicillin | Yes [102] |
| Haemophilus influenzae | Ampicillin | No |
| Group B Streptococci | Penicillin | No |
* The BCP column uses Escherichia coli as a reference for this group ** The BCP column uses Klebsiella pneumoniae as a reference for this group ***RR-TB was evaluated independently in a tailored approach so it was technically “not” included in the list but after the evaluation by specialists, it was determined as a critically dangerous bacteria therefore. RR-TB stands apart from the list due to the distinct nature of its evaluation process.
Table 3.
Organisms used in BCP and how images were analysed.
| Organism | Dyes/Fluorophores | Processed Data available | Segmentation | Feature extraction | Source |
|---|---|---|---|---|---|
| Acinetobacter baumannii and E. coli | FM4-64 DAPI SYTOX-Green |
Yes | CellProfiler | CellProfiler | [105] |
| Acinetobacter baumannii | FM4-64 DAPI SYTOX-Green |
No | Ilastic | CellProfiler | [106]* |
| Pseudomonas aeruginosa | FM4-64 DAPI |
Yes | Manually (FIJI/ImageJ ) |
Manually (FIJI/ImageJ) |
[107]* |
| S. aureus | FM4-64 DAPI SYTOX-Green |
Yes | Semi-Manual (FIJI/ImageJ) | Semi-Manual (FIJI/ImageJ) | [15]* |
| S. aureus | FM4-64 DAPI SYTOX-Green WGA-647 |
Yes | CellProfiler | CellProfiler | [99]* |
|
S. aureus, S. Typhimurium, and K. pneumoniae |
FM4-64 DAPI SYTOX-Green |
Yes | Harmony | Harmony | [94] |
| B. subtilis | FM 4-64 DAPI SYTOX Green |
Yes | CellProfiler | CellProfiler FIJI |
[108]* |
| B. subtilis | FM4-64 DAPI SYTOX-Green |
No | CellProfiler | CellProfiler | [109]* |
| B. subtilis | Nile red DAPI |
No | MicrobeJ | MicrobeJ | [110]* |
|
Bacillus subtilis, E. coli |
FM4-64 DAPI GFP |
No | Wasabi software (Hamamatsu) | Wasabi software (Hamamatsu) | [111] |
| E. coli | FM4-64 Hoechst-33342 Dendra2 protein |
No | FIJI/ImageJ | FIJI/ImageJ | [80] |
|
E. coli Caulobacter crescentus |
FM4-64 DAPI |
No | Oufti | Oufti | [112]* |
| E. coli | FM4-64 DAPI SYTOX-Green |
No | Semi-Manual (FIJI/ImageJ) | Semi-Manual (FIJI/ImageJ) | [113]* |
| Achromobacter xylosoxidans | FM4-64 DAPI SYTOX-Green NBD Azithromycin |
No | FIJI/ImageJ and CellProfiler | FIJI/ImageJ and CellProfiler | [114] |
| M. smegmatis | ParB-mCherry | Yes | MicrobeJ | MicrobeJ | [115]* |
| M. tuberculosis Erdman | FM4-64FX SYTO 24 |
Yes | MorphEUS | MorphEUS | [116]* |
| Shewanella putrefaciens | Ffh-mVenus FtsY-mVenus uL1-mVenus |
Yes | FIJI/ImageJ | FIJI/ImageJ | [117]* |
| V. parahaemolyticus | FM4-64 DAPI |
No | FIJI/ImageJ | FIJI/ImageJ | [84] |
| Bacillus subtilis | FM4-64 DAPI SYTOX-Green SYTO-9 |
No | - | Manual w/ FIJI | [118]* |
|
M. smegmatis M. tuberculosis |
FM4-64 GFP CellROX |
Yes | Omnipose | Manual w/ FIJI or with Cell Counter installation in FIJI Custom python script |
[119] |
|
E. coli B. subtilis |
FM4-64 DAPI GFP DiSC |
No | FIJI/ImageJ and MicrobeJ | FIJI/ImageJ and MicrobeJ | [120] |
*Pipelines, scripts, or instructions are detailed and/or included in the paper. Programs are also widely accessible.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



