
Submitted:
29 August 2024
Posted:
30 August 2024
You are already at the latest version
Abstract
Mosquitoes are responsible for the transmission of numerous pathogens, including Plasmodium parasites, arboviruses and filarial worms. They pose a significant risk to public health with over 200 million cases of malaria per annum and approximately four billion people at risk of arthropod-borne viruses (arboviruses). Mosquito populations are geographically expanding into temperate regions and their distribution is predicted to continue increasing. Mosquito symbionts, including fungi, bacteria and viruses, have desirable traits for mosquito disease control including spreading horizontally and vertically through mosquito populations and potentially colonising multiple important vector species. Paratransgenesis, genetic modification of mosquito symbionts with effectors to target the pathogen rather than the vector, is a promising strategy to prevent the spread of mosquito-borne diseases. A variety of effectors can be expressed but venom toxins are excellent effector candidates because they are target specific, potent, and, stable. However, the only toxins to be explored in mosquito paratransgenesis to date are Scorpine and mutated phospholipase A2. To enhance the scope, effectiveness, and durability of paratransgenesis, an expanded arsenal of effectors is required. This review discusses other potential toxins effectors for future paratransgenesis studies based on prior in vitro and in vivo antiparasitic and antiviral studies and highlights the need for further research and investment in this area. In terms of mosquito-borne diseases, paratransgenesis strategies have been developed to target Plasmodium. We postulate the potential to apply this principle to target arboviruses using antiviral toxin effectors.
Keywords:
Plasmodium
; rift valley fever virus
; yellow fever virus
; Japanese encephalitis virus
; West Nile virus
; chikungunya virus
; dengue virus
; zika virus
; snake
; scorpion
; spider
; Aedes
; Anopheles
Overall Introduction and Aims
Mosquito-borne pathogens such as parasites and arthropod-borne viruses (arboviruses) pose a significant risk to public health [1]. Malaria is one of the most common parasitic diseases globally with an estimated 249 million cases and 608,000 deaths reported in 2022, mostly in children under five in sub-Saharan Africa [2]. Over half the world’s population are at risk of infection by arboviruses, including Rift Valley fever virus (RVFV), yellow fever virus (YFV), Zika virus (ZIKV), Japanese encephalitis virus (JEV), West Nile virus (WNV), Dengue virus (DENV), and Chikungunya virus (CHIKV) [3].
Due to climate change and other anthropogenic factors, the burden of mosquito-borne diseases is intensifying [4,5,6]. There is no single solution for the control of mosquito-borne disease and multiple strategies are required. This multi-pronged approach will require location-specific strategies influenced by environmental and economic factors, governing bodies and disease prevalence [7]. In this regard, novel strategies and tools are urgently required to develop an integrated control strategy.
Paratransgenesis, the genetic engineering of symbionts with anti-pathogenic effectors to control disease transmission, represents a potentially promising strategy. The technique was originally developed by Beard et al. to control Rhodnius prolixus (triatomine/kissing bug) from spreading the causal parasite of Chagas disease (Trypanosoma cruzi). A gram-positive bacteria, Rhodococcus rhodnii, that occurs at high concentrations within the hindgut of R. prolixus was genetically engineered to express a trypanocidal immune peptide, Cecropin A. This resulted in a decreased T. cruzi infection rate in R. prolixus and was approved as an integrated pest management program in South and Central America [8].
Paratransgenesis offers several advantages. It is scalable because transgenic microorganisms can be grown to large quantities at low cost. The technique is not limited to single mosquito species because symbiotics can potentially colonise multiple important vector species. Moreover, the symbiont can be maintained within the ecosystem by vertical, horizontal, and trans-stadial transmission, mitigating the need for re-introduction [9,10]. Finally, and perhaps most importantly, it is a manipulable system that can be altered to target different pathogens or keep pace with resistance by exploiting different effectors. As such, the discovery and development of novel anti-pathogen molecules is critical for paratransgenesis implementation.
Venom toxins are excellent candidates for effectors in paratransgenesis. Venoms are complex mixtures of toxic proteins, peptides, and small molecules that are delivered through the infliction of a wound [11,12]. Venoms of hymenopteran insects such as bees and wasps are diverse, consisting of peptides, enzymes, and neurotransmitters [13], whilst scorpion and spider venoms largely consist of neurotoxins, which modulate a variety of channels including voltage-gated potassium, sodium, and calcium ion channels, acid sensing ion channels, calcium-activated potassium channels, glutamate receptors, and glutamate transporters [14,15]. Snake venom consists of haemotoxins, cytotoxins, and neurotoxins that can be grouped into superfamilies by structure, with phospholipase A2s (PLA2s), snake venom metalloproteinases, snake venom serine proteinases, and three-finger toxins being the most abundant [16]. Venom toxins have high specificity, potency, and stability [11], and are less susceptible to bioaccumulation than chemical insecticides [17]. The venoms of many hymenopteran insects, scorpions, spiders, and snakes have been studied for their potential antiparasitic [18,19,20] and antiviral [21,22,23,24] properties. This review highlights the untapped potential of venom toxins as effectors in paratransgenesis. We discuss successful paratransgenesis studies that have been undertaken with antimalarial venom toxins as proof of principle and the need for specific screening of venom toxins to identify effectors is highlighted. Regarding mosquito-borne diseases, paratransgenesis strategies have focused on targeting Plasmodium, the causal agent of malaria. However, we suggest that paratransgenesis could be applicable to target arboviruses through the use of antiviral venom toxins.
This review highlights the untapped potential of venom toxins as effectors in paratransgenesis. We discuss successful paratransgenesis studies that have been undertaken with antimalarial venom toxins as proof of principle and the need for specific screening of venom toxins to identify effectors is highlighted. Regarding mosquito-borne diseases, paratransgenesis strategies have focused on targeting Plasmodium, the causal agent of malaria. However, we suggest that paratransgenesis could be applicable to target arboviruses through the use of antiviral venom toxins.
Antiparasitic Venom Toxins as Effectors to Target Plasmodium
Previous mosquito paratransgenesis strategies have focussed on targeting Plasmodium, the casual parasite of malaria. The species of Plasmodium responsible for causing malaria in humans are P. falciparum, P. vivax, P. ovale, P. knowlesi, and P. malariae, with the former being responsible for more than 90% of the malaria deaths. Mosquitoes from the Anopheles genus are responsible for the transmission of malaria. Paratransgenesis targeting Plasmodium must use effectors that inhibit the parasite stages within the mosquito: gametes, ookinetes, oocysts or sporozoites (Figure 1) [25]. Two venom toxins have effectively been utilised as effectors (Table 1): Scorpine, an excitatory neurotoxin from Pandinus imperator with antibacterial and antiparasitic properties, and mPLA2, a PLA2 from bee venom with a point mutation (H67N) to prevent enzyme activity and toxicity to bacteria. mPLA2 expressed in Escherichia coli induced a moderate reduction of oocyst numbers from P. berghei, a rodent malaria model, when fed to Anopheles stephensi, however, the bacterium survived poorly in the mosquito [26]. mPLA2 and Scorpine expressed in P. agglomerans [27] and Serratia [28] were able to effectively colonise the midgut of An. gambiae and decreased the number of P. falciparum oocytes in infected mosquitoes. Scorpine has also been expressed in Asaia [29], a bacteria found in Anopheles sp., Aedes aegypti, and Aedes albopictus [30,31,32,33,34,35,36] that is transmitted vertically, horizontally and transstadially [31]. Transgenic Asaia expressing Scorpine significantly reduced the number of P. berghei oocytes in the mosquito midgut, however, constitutive expression of the toxin compromised bacterial fitness. To improve bacterial fitness, blood meal inducible promoters within the mosquito microbiome were identified and used to conditionally express Scorpine. This enabled Asaia to maintain fitness and compete with wild type Asaia, whilst oocyst midgut number in A. stephensi decreased by approximately 90% and prevalence decreased by up to 20%, indicating a decrease in infection potential [37]. Future work should assess transgenic bacteria in semi-field trials to assess suitability for practical use within the field.
Despite promising preliminary mosquito paratransgenesis data, only a limited number of effector molecules have been assessed with mPLA2 and Scorpine being the only venom toxins effectors that have been experimentally tested in mosquito paratransgenesis. An expanded arsenal of molecules is required to allow a multi-faceted and adaptable approach to paratransgenesis. Importantly, expression of multiple effectors has been shown to enhance efficacy [27,28] and can enable several stages of the pathogen life cycle to be targeted, increasing robustness. The risk of resistance development can be reduced through identification and use of multiple effectors with different mechanisms of actions and/or broad-spectrum actions. There is also a potential to target multiple pathogens through co-expression of effectors or use of effectors with multiple mechanism of actions. Finally, it is important to have a diverse effector library available to mitigate resistance and enable new paratransgenesis replacement strategies.
Venom toxins have the potential to act as effectors due to their antiparasitic activity (Supplementary Table S1, Figure 1). However, the majority of these studies have focussed on the intraerythrocytic asexual stages of Plasmodium within the mammalian host [19], in line with research more applicable to the identification of antimalaria therapeutics. Few studies have screened toxins to identify effectors for paratransgenesis, but for effectors to be useful they must target the Plasmodium stages occurring in the mosquito [25].
Several α-helical linear peptides such as Anoplin and Mastoparan X from wasp venom [38], Melittin from European honeybee venom [38], and MeuTXKβ [39] from Mesobuthus scorpion venom inhibit ookinete development. Whilst, another linear helical peptide, specifically scorpion toxin VmCT1 from Vaejovis mexicanus is effective in vitro against P. gallinaceum sporozoites, a poultry model of the last stage of Plasmodium development within the mosquito [40]. Antimicrobial peptides from scorpions including Scorpine and synthetic peptides based on Vejovine and Hadrurin also inhibit ookinete development in vitro [41,42,43]. In vivo studies have found PLA2 derived from the venom of the rattlesnake Crotalus adamanteus reduced the number of oocysts by 99% when mixed with cultured P. falciparum gametocytes and fed to An. gambiae or An. stephensi mosquitos [44]. A similar reduction in P. gallinaceum oocyst number in Ae. aegypti was achieved. Interestingly, the PLA2 toxin did not affect ookinete viability, but acted on the midgut surface, preventing ookinete maturation to oocytes. A similar effect was observed for a PLA2 from bee venom in Ae. fluviatilis [45] and Melittin [38].
Ookinete to oocyst development in the midgut, the bottleneck of malaria transmission [46], is the best target for effector screening. The emphasis should be on P. falciparum, the species primarily infecting humans and causing most malaria deaths. Although experiments with P. falciparum are challenging and limited to in vitro culture of the intraerythrocytic stages of the parasite, ookinetes can be generated in vitro by gametocyte differentiation in specialised medium enabling toxins to be rapidly screened [47]. However, studies have mainly been performed with P. berghei and P. gallineceum, species that infect rodents and poultry, respectively, as described above. This is because they can be maintained as intraerythrocytic stages in mice or chickens to generate a high density of gametocytes, the sexual stage that is required for oocyst development within the mosquito. An ideal high-throughput pipeline would be the screening of toxins on ookinete development in vitro with successful candidates taken forward to in vivo development of Plasmodium in the mosquito, as described by Carter et al., 2013 [38].
In addition, downstream assays must be undertaken to ensure the toxin is specific for the pathogen. This should involve assessing any potential effect of mosquito fitness, as performed in a previous study to identify effectors [38]. Assays included basic in vitro cell viability assays on mosquito cell lines and feeding toxins to mosquitoes. However, the compatibility of the toxins with the mosquito symbiont must also be assessed by performing minimum inhibitory concentration assays. From an ecological perspective it is important to assess effects on other insects, especially pollinator species, using similar assays. From a safety perspective, mammalian toxicity should also be reviewed using in vivo and in vitro assays. These types of studies have largely been neglected to date.
Could Paratransgenesis Be Used to Target Arboviruses?
Paratransgenesis to target arboviruses has not been attempted thus far but the antiviral properties of venom toxins are encouraging for this strategy. Antiviral compounds can target various stages of virus infection including pre-entry and/or post-entry stages [48]. Compounds can inactivate the virus pre-entry by inactivating the virus before it attaches to the cell, a process known as neutralisation; inhibiting surface proteins required for attachment; inhibiting virus endocytosis or inhibiting fusion of the viral envelope and host cell membrane. Alternatively, compounds can act at the post-entry stages, by inhibiting viral uncoating, replication, transcription, translation, virus assembly, and virus release. Antivirals can also induce the host immune response, through stimulating the production of interferons, other cytokines, and chemokines, affecting both pre- and post-entry stages. Targeting any of these stages within the mosquito midgut, as the location of arbovirus infection after the mosquito takes a blood meal from an infected host, has the potential to prevent viral dissemination into salivary glands. Blocking this step, as with Plasmodium, would prevent the mosquito becoming infectious and transmitting the arbovirus and has been suggested previously as a strategy to control arbovirus transmission [49].
Venom toxins have shown antiviral activity against ZIKV, DENV, YFV, JEV, and CHIKV. However, there is limited research on the antiviral properties of venom toxins against RVFV. Many antiviral venom toxins have been shown to target the pre-entry stages, the most studied of which being group I and II snake venom PLA2 toxins (Figure 2, Supplementary Table S2). Group I PLA2, consisting of PLA2 produced by Elapidae (cobras, mambas, coral snakes) and Hydrophidae (sea snakes) whilst group II PLA2 are produced by Viperidae (rattlesnakes) [12]. Group II PLA2s derived from Bothrops alteratus [50], B. leucurus [51], and B. asper venom [52] can neutralise several strains of DENV, whilst group II PLA2s from B. jararacussu [53,54] and Crotalus durissus terrificus venom have shown inhibition activity against YFV, CHIKV, DENV, and ZIKV [55,56,57,58,59]. LaPLA2-1, a group III PLA2 from the scorpion Liocheles australasiae, can neutralise DENV and JEV [60]. Interestingly, DENV propagated in mosquito cell lines was more sensitive to Mt-I, a catalytically inactive PLA2 from B. asper venom, than viruses propagated in mammalian cells [52]. Neutralisation by group I, II, and III PLA2 likely occurs by hydrolysis of the virus lipid bilayer [55,59,60]. Viral neutralisation has also been shown with to occur with ZY13, a peptide analogue of cathelicidin from Bungarus fasciatus venom [61] and the Scorpine-like peptide Smp76 from Scorpio maurus palmatus venom [62,63].
Venom toxins can also inhibit virus post-entry stages (Figure 2). Ev37, a Scorpine-like peptide from scorpion Euscorpiops validus venom that selectively inhibit Kv1.3 potassium channel, prevents viral genome release into the cytoplasm by acidifying viral genome-containing vesicles preventing membrane fusion [64]. The host defense peptide Av-LCTX-An1a from Alopecosa nagpag spider venom can inhibit viral protease activity preventing virus maturation [62]. Studies assessing the host immune response have found that Scorpine-like peptide rSmp76 from scorpion Scorpio maurus palmatus venom and ZY13 have antiviral effects by activating interferon signaling [61,62]. However, it is important to stress that most of these studies have been undertaken with mammalian cell lines, and their translatability into mosquito cells is unknown. Promisingly, recombinant Scorpine generated in Anopheles gambie cells can inhibit DENV serotype 2 replication in mosquito cells [42] showing the potential of venom toxins to have antiviral activity within mosquitoes. Similar studies with the aforementioned venom toxins are necessary to determine if the antiviral activity seen within mammalian cells is transferable to mosquitoes. In vivo studies assessing viral load, for example by RT-qPCR and plaque assays, in mosquitoes fed with toxins and virus must also be conducted to confirm in vitro findings.
Screening to identify new potential antiviral effectors should involve assessing the ability of toxins to inhibit virus at all stages of infections. Viral neutralization should be assessed by incubating the test compound with the virus then assessing virus titre. Effect of the toxins at pre-entry stages should be evaluated by simultaneously adding the toxin and virus to mosquito cells at 4 °C (to prevent virus internalisation) and quantifying the levels of bound virus, as well as simultaneously adding the compounds and virus at 37 °C to determine effects on virus internalization and entry. Toxins should also be added after viral infection to evaluate post-entry antiviral activity. Finally, the ability of the toxin to induce a cellular antiviral response can be determined by addition of the toxin to the host cells pre-viral infection. Similarly, with antiparasitic effectors, any potential candidates should be further tested to ensure the toxin does not affect the fitness of the mosquito and symbiont. This is a vital step before moving forward with genetically engineering the symbiont.
Conclusions
Venoms contain a highly diverse library of bioactive and stable peptides with antiparasitic and antiviral properties. Studies have shown that using venom toxins as transgenes in paratransgenesis can be useful for the control of mosquito-borne pathogens, specifically Plasmodium. However, few studies have screened toxins with the goal of identifying effector molecules and therefore the choice of potential effectors is limited. Here, we have reviewed the toxin literature and have highlighted potential effector candidates for future paratransgenesis studies. However, we stress that additional screening with the aim of identifying effectors is vital. These studies should involve in vitro and in vivo studies to select antiviral and antiparasitic toxins that target appropriate stages of the pathogen life cycle, and that do not affect mosquito or symbiont fitness. We also argue the paratransgenesis strategy should be expanded to attempt to target arboviruses. The studies discussed here provide a strong foundation for further research in this area to identify toxin effector candidates.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
List of Abbreviations
Arbovirus, arthropod-borne viruses; CHIKV, chikungunya virus; DENV, dengue virus; JEV, Japanese encephalitis virus; mPLA2, inactive mutant (H67N) phospholipase A2; PLA2, phospholipase A2; RVFV, Rift Valley fever phlebovirus; WNV, West Nile virus; YFV, yellow fever virus; ZIKV, Zika virus.
References
- Lee H, Halverson S, Ezinwa N. Mosquito-Borne Diseases. Primary Care - Clinics in Office Practice. 2018, 45, 393–407. [Google Scholar] [CrossRef] [PubMed]
- World malaria report 2023. World Health Organisation. Geneva: Licence: CC BY-NC-SA 3.0 IGO.; 2023. p. 1–356.
- Global Arbovirus Initiative: preparing for the next pandemic tackling mosquito-borne viruses with epidemic and pandemic potential. World Health Organization. Geneva: Licence: CC BY-NCSA 3.0 IGO; 2024. p. 1–24.
- Wu X, Lu Y, Zhou S, Chen L, Xu B. Impact of climate change on human infectious diseases: Empirical evidence and human adaptation. Environ Int. 2016, 86, 14–23. [Google Scholar] [CrossRef]
- Semenza JC, Rocklöv J, Ebi KL. Climate change and cascading risks from infectious disease. Infect Dis Ther. 2022, 11, 1371–90. [Google Scholar] [CrossRef] [PubMed]
- Fouque F, Reeder JC. Impact of past and on-going changes on climate and weather on vector-borne diseases transmission: a look at the evidence. Infect Dis Poverty. 2019, 8, 1–9. [Google Scholar]
- Baitharu I, Shroff S, Naik PP, Sahu JK. Environmental Management and Sustainable Control of Mosquito Vector: Challenges and Opportunities. Molecular Identification of Mosquito Vectors and Their Management. Singapore: Springer Singapore; 2020. p. 129–47.
- Durvasula R, V.; , Gumbs A, Panackal A, Kruglov O, Aksoy S, Merrifield RB, et al. Prevention of insect-borne disease: An approach using transgenic symbiotic bacteria. Proc Natl Acad Sci U S A. 1997, 94, 3274–8. [Google Scholar] [CrossRef]
- Ratcliffe NA, Furtado Pacheco JP, Dyson P, Castro HC, Gonzalez MS, Azambuja P, et al. Overview of paratransgenesis as a strategy to control pathogen transmission by insect vectors. Parasit Vectors. 2022, 15, 1–31. [Google Scholar]
- Wilke ABB, Marrelli MT. Paratransgenesis: A promising new strategy for mosquito vector control. Parasit Vectors. 2015, 8, 1–9. [Google Scholar]
- Casewell NR, Wüster W, Vonk FJ, Harrison RA, Fry BG. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol Evol. 2013, 28, 219–29. [Google Scholar] [CrossRef]
- Mackessy, SP. Venom production and secretion in reptiles. Journal of Experimental Biology. 2022, 225, 227348. [Google Scholar] [CrossRef]
- Guido-Patiño JC, Plisson F. Profiling hymenopteran venom toxins: Protein families, structural landscape, biological activities, and pharmacological benefits. Toxicon X. 2022, 14, 100119. [Google Scholar]
- Escoubas P, Diochot S, Corzo G. Structure and pharmacology of spider venom neurotoxins. Biochimie. 2000, 82, 893–907. [Google Scholar] [CrossRef] [PubMed]
- Xia Z, He D, Wu Y, Kwok HF, Cao Z. Scorpion venom peptides: Molecular diversity, structural characteristics, and therapeutic use from channelopathies to viral infections and cancers. Pharmacol Res. 2023, 197, 106978. [Google Scholar] [CrossRef]
- Gutiérrez JM, Calvete JJ, Habib AG, Harrison RA, Williams DJ, Warrell DA. Snakebite envenoming. Nat Rev Dis Primers. 2017, 3, 1–21. [Google Scholar]
- King, GF. Tying pest insects in knots: the deployment of spider-venom-derived knottins as bioinsecticides. Pest Manag Sci. 2019, 75, 2437–45. [Google Scholar] [CrossRef]
- Almeida JR, Gomes A, Mendes B, Aguiar L, Ferreira M, Brioschi MBC, et al. Unlocking the potential of snake venom-based molecules against the malaria, Chagas disease, and leishmaniasis triad. Int J Biol Macromol. 2023, 242 Pt. 2, 124745. [Google Scholar]
- Salimo ZM, Barros AL, Adrião AAX, Rodrigues AM, Sartim MA, de Oliveira IS, et al. Toxins from animal venoms as a potential source of antimalarials: A comprehensive review. Toxins (Basel). 2023, 15, 375. [Google Scholar] [CrossRef] [PubMed]
- Samy RP, Foo SL, Franco OL, Stiles BG, Kumar AP, Sethi G, et al. Identification of natural peptides as a new class of antimalarial drugs by in silico approaches. Frontiers in Bioscience. 2017, 9, 88–110. [Google Scholar] [CrossRef]
- Utkin Y, Siniavin A, Kasheverov I, Tsetlin V. Antiviral effects of animal toxins: is there a way to drugs? Int J Mol Sci. 2022, 23, 3634. [Google Scholar] [CrossRef]
- da Mata ÉCG, Mourão CBF, Rangel M, Schwartz EF. Antiviral activity of animal venom peptides and related compounds. Journal of Venomous Animals and Toxins Including Tropical Diseases 2017, 23, 1–12. [Google Scholar]
- Lima WG, Maia CQ, de Carvalho TS, Leite GO, Brito JCM, Godói IPD, et al. Animal venoms as a source of antiviral peptides active against arboviruses: a systematic review. Arch Virol. 2022, 167, 1763–72. [Google Scholar] [CrossRef] [PubMed]
- Teixeira SC, Borges BC, Oliveira VQ, Carregosa LS, Bastos LA, Santos IA, et al. Insights into the antiviral activity of phospholipases A2 (PLA2s) from snake venoms. Int J Biol Macromol. 2020, 164, 616–25. [Google Scholar] [CrossRef] [PubMed]
- Whitten MMA, Shiao SH, Levashina EA. Mosquito midguts and malaria: cell biology, compartmentalization and immunology. Parasite Immunol. 2006, 28, 121–30. [Google Scholar] [CrossRef]
- Riehle MA, Moreira CK, Lampe D, Lauzon C, Jacobs-Lorena M. Using bacteria to express and display anti-Plasmodium molecules in the mosquito midgut. Int J Parasitol. 2007, 37, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Wang S, Ghosh AK, Bongio N, Stebbings KA, Lampe DJ, Jacobs-Lorena M. Fighting malaria with engineered symbiotic bacteria from vector mosquitoes. Proc Natl Acad Sci U S A. 2012, 109, 12734–9. [Google Scholar] [CrossRef]
- Wang S, Dos-Santos ALA, Huang W, Liu KC, Oshaghi MA, Wei G, et al. Driving mosquito refractoriness to Plasmodium falciparum with engineered symbiotic bacteria. Science (1979). 2017, 357, 1399–402. [Google Scholar]
- Bongio NJ, Lampe DJ. Inhibition of Plasmodium berghei development in mosquitoes by effector proteins secreted from Asaia sp. bacteria using a novel native secretion signal. PLoS One. 2015, 10, e0143541. [Google Scholar]
- De Freece C, Damiani C, Valzano M, D’Amelio S, Cappelli A, Ricci I, et al. Detection and isolation of the α-proteobacterium Asaia in Culex mosquitoes. Med Vet Entomol. 2014, 28, 438–42. [Google Scholar] [CrossRef]
- Favia G, Ricci I, Damiani C, Raddadi N, Crotti E, Marzorati M, et al. Bacteria of the genus Asaia stably associate with Anopheles stephensi , an Asian malarial mosquito vector. Proc Natl Acad Sci U S A. 2007, 104, 9047–51. [Google Scholar] [CrossRef]
- Maffo CGT, Sandeu MM, Fadel AN, Tchouakui M, Nguete DN, Menze B, et al. Molecular detection and maternal transmission of a bacterial symbiont Asaia species in field-caught Anopheles mosquitoes from Cameroon. Parasit Vectors. 2021, 14, 1–11. [Google Scholar]
- Rami A, Raz A, Zakeri S, Dinparast Djadid N. Isolation and identification of Asaia sp. in Anopheles spp. mosquitoes collected from Iranian malaria settings: Steps toward applying paratransgenic tools against malaria. Parasit Vectors. 2018, 11, 1–8. [Google Scholar]
- Zouache K, Raharimalala FN, Raquin V, Tran-Van V, Raveloson LHR, Ravelonandro P, et al. Bacterial diversity of field-caught mosquitoes, Aedes albopictus and Aedes aegypti , from different geographic regions of Madagascar. FEMS Microbiol Ecol. 2011, 75, 377–89. [Google Scholar] [CrossRef] [PubMed]
- Chouaia B, Rossi P, Montagna M, Ricci I, Crotti E, Damiani C, et al. Molecular evidence for multiple infections as revealed by typing of Asaia bacterial symbionts of four mosquito species. Appl Environ Microbiol. 2010, 76, 7444–50. [Google Scholar] [CrossRef] [PubMed]
- Boissière A, Tchioffo MT, Bachar D, Abate L, Marie A, Nsango SE, et al. Midgut microbiota of the Malaria mosquito vector Anopheles gambiae and interactions with Plasmodium falciparum infection. PLoS Pathog. 2012, 8, e1002742. [Google Scholar]
- Shane JL, Grogan CL, Cwalina C, Lampe DJ. Blood meal-induced inhibition of vector-borne disease by transgenic microbiota. Nat Commun. 2018, 9, 1–10. [Google Scholar]
- Carter V, Underhill A, Baber I, Sylla L, Baby M, Larget-Thiery I, et al. Killer bee molecules: Antimicrobial peptides as effector molecules to target sporogonic stages of Plasmodium. PLoS Pathog. 2013, 9, e1003790. [Google Scholar]
- Zhu S, Gao B, Aumelas A, del Carmen Rodríguez M, Lanz-Mendoza H, Peigneur S, et al. MeuTXKβ1, a scorpion venom-derived two-domain potassium channel toxin-like peptide with cytolytic activity. Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 2010, 1804, 872–83. [Google Scholar] [CrossRef] [PubMed]
- Pedron C, Silva A, Torres M, de Oliveira C, Andrade G, Cerchiaro G, et al. Net charge tuning modulates the antiplasmodial and anticancer properties of peptides derived from scorpion venom. J Pept Sci. 2021, 27, e3296. [Google Scholar] [CrossRef]
- Sánchez-Vásquez L, Silva-Sanchez J, Jiménez-Vargas JM, Rodríguez-Romero A, Muñoz-Garay C, Rodríguez MC, et al. Enhanced antimicrobial activity of novel synthetic peptides derived from vejovine and hadrurin. Biochim Biophys Acta Gen Subj. 2013, 1830, 3427–36. [Google Scholar] [CrossRef]
- Carballar-Lejarazú R, Rodríguez MH, De La Cruz Hernández-Hernández F, Ramos-Castañeda J, Possani LD, Zurita-Ortega M, et al. Recombinant scorpine: A multifunctional antimicrobial peptide with activity against different pathogens. Cellular and Molecular Life Sciences. 2008, 65, 3081–92. [Google Scholar] [CrossRef]
- Conde R, Zamudio FZ, Rodríguez MH, Possani LD. Scorpine, an anti-malaria and anti-bacterial agent purified from scorpion venom. FEBS Lett. 2000, 471, 165–8. [Google Scholar] [CrossRef]
- Zieler H, Keister DB, Dvorak JA. A snake venom phospholipase A2 blocks malaria parasite development in the mosquito midgut by inhibiting ookinete association with the midgut surface. J Exp Biol. 2001, 204, 4157–67. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues FG, Santos MN, De Carvalho TXT, Rocha BC, Riehle MA, Pimenta PFP, et al. Expression of a mutated phospholipase A2 in transgenic Aedes fluviatilis mosquitoes impacts Plasmodium gallinaceum development. Insect Mol Biol. 2008, 17, 175–83. [Google Scholar] [CrossRef] [PubMed]
- Smith RC, Vega-Rodríguez J, Jacobs-Lorena M. The Plasmodium bottleneck: Malaria parasite losses in the mosquito vector. Mem Inst Oswaldo Cruz. 2014, 109, 644–61. [Google Scholar] [CrossRef]
- Delves M, Lafuente-Monasterio MJ, Upton L, Ruecker A, Leroy D, Gamo FJ, et al. Fueling open innovation for malaria transmission-blocking drugs: hundreds of molecules targeting early parasite mosquito stages. Front Microbiol. 2019, 10, 1–10. [Google Scholar]
- Lee MF, Wu YS, Poh CL. Molecular mechanisms of antiviral agents against Dengue virus. Viruses. 2023, 15, 1–37. [Google Scholar]
- Beard CB, O’Neill SL, Tesh RB, Richards FF, Aksoy S. Modification of arthropod vector competence via symbiotic bacteria. Parasitology Today. 1993, 9, 179–83. [Google Scholar] [CrossRef]
- Dias EHV, de Sousa Simamoto BB, da Cunha Pereira DF, Ribeiro MSM, Santiago FM, de Oliveira F, et al. Effect of BaltPLA2, a phospholipase A2 from Bothrops alternatus snake venom, on the viability of cells infected with dengue virus. Toxicology in Vitro. 2023, 88, 1–6. [Google Scholar]
- Cecilio AB, Caldas S, De Oliveira RA, Santos ASB, Richardson M, Naumann GB, et al. Molecular characterization of Lys49 and Asp49 phospholipases A2 from snake venom and their antiviral activities against Dengue virus. Toxins (Basel). 2013, 5, 1780–98. [Google Scholar] [CrossRef]
- Brenes H, Loría GD, Lomonte B. Potent virucidal activity against Flaviviridae of a group IIA phospholipase A2 isolated from the venom of Bothrops asper. Biologicals. 2020, 63, 48–52. [Google Scholar] [CrossRef]
- Cassani NM, Santos IA, Grosche VR, Ferreira GM, Guevara-Vega M, Rosa RB, et al. Roles of Bothrops jararacussu toxins I and II: Antiviral findings against Zika virus. Int J Biol Macromol. 2023, 227, 630–40. [Google Scholar] [CrossRef]
- Ayusso GM, Lima MLD, da Silva Sanches PR, Santos IA, Martins DOS, da Conceição PJP, et al. The dimeric peptide (KKYRYHLKPF)2K shows broad-spectrum antiviral activity by inhibiting different steps of chikungunya and zika virus infection. Viruses. 2023, 15, 1–17. [Google Scholar]
- Muller VD, Soares RO, Santos-Junior NN Dos, Trabuco AC, Cintra AC, Figueiredo LT, et al. Phospholipase A2 isolated from the venom of Crotalus durissus terrificus inactivates dengue virus and other enveloped viruses by disrupting the viral envelope. PLoS One. 2014, 9, e112351. [Google Scholar]
- Muller VDM, Russo RR, Oliveira Cintra AC, Sartim MA, De Melo Alves-Paiva R, Figueiredo LTM, et al. Crotoxin and phospholipases A 2 from Crotalus durissus terrificus showed antiviral activity against dengue and yellow fever viruses. Toxicon. 2012, 59, 507–15. [Google Scholar] [CrossRef] [PubMed]
- Russo RR, dos Santos Júnior NN, Cintra ACO, Figueiredo LTM, Sampaio SV, Aquino VH. Expression, purification and virucidal activity of two recombinant isoforms of phospholipase A 2 from Crotalus durissus terrificus venom. Arch Virol. 2019, 164, 1159–71. [Google Scholar] [CrossRef] [PubMed]
- Santos IA, Shimizu JF, de Oliveira DM, Martins DOS, Cardoso-Sousa L, Cintra ACO, et al. Chikungunya virus entry is strongly inhibited by phospholipase A2 isolated from the venom of Crotalus durissus terrificus. Sci Rep. 2021, 11, 1–12. [Google Scholar]
- Chen M, Aoki-Utsubo C, Kameoka M, Deng L, Terada Y, Kamitani W, et al. Broad-spectrum antiviral agents: secreted phospholipase A2 targets viral envelope lipid bilayers derived from the endoplasmic reticulum membrane. Sci Rep. 2017, 7, 1–8. [Google Scholar]
- Miyashita M, Mitani N, Kitanaka A, Yakio M, Chen M, Nishimoto S, et al. Identification of an antiviral component from the venom of the scorpion Liocheles australasiae using transcriptomic and mass spectrometric analyses. Toxicon. 2021, 191, 25–37. [Google Scholar] [CrossRef]
- Xing M, Ji M, Hu J, Zhu T, Chen Y, Bai X, et al. Snake cathelicidin derived peptide inhibits zika virus infection. Front Microbiol. 2020, 11, 1–11. [Google Scholar]
- Ji Z, Li F, Xia Z, Guo X, Gao M, Sun F, et al. The scorpion venom peptide Smp76 inhibits viral infection by regulating Type-I interferon response. Virol Sin. 2018, 33, 545–56. [Google Scholar] [CrossRef]
- El-Bitar AMH, Sarhan M, Abdel-Rahman MA, Quintero-Hernandez V, Aoki-Utsubo C, Moustafa MA, et al. Smp76, a scorpine-Like peptide isolated from the venom of the scorpion Scorpio maurus palmatus , with a potent antiviral activity against Hepatitis C virus and Dengue virus. Int J Pept Res Ther. 2020, 26, 811–21. [Google Scholar] [CrossRef]
- Li F, Lang Y, Ji Z, Xia Z, Han Y, Cheng Y, et al. A scorpion venom peptide Ev37 restricts viral late entry by alkalizing acidic organelles. Journal of Biological Chemistry. 2019, 294, 182–94. [Google Scholar] [CrossRef] [PubMed]
- Flipse J, Wilschut J, Smit JM. Molecular mechanisms involved in antibody-dependent enhancement of dengue virus infection in humans. Traffic. 2013, 14, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Fox JM, Pierson TC. Chikungunya virus assembly and egress. Nat Microbiol. 2022, 7, 1112–3. [Google Scholar] [CrossRef] [PubMed]
- Ji M, Zhu T, Xing M, Luan N, Mwangi J, Yan X, et al. An antiviral peptide from Alopecosa nagpag spider targets NS2B-NS3 protease of flaviviruses. Toxins (Basel). 2019, 11, 584. [Google Scholar] [CrossRef]
Figure 1.
Overview of in vitro and in vivo studies showing activity of toxins against the mosquito stages of Plasmodium sp. Venom toxins that have shown anti-plasmodial activity within mosquitos (in vivo, pink panel), and anti-plasmodial activity when added directly to isolated Plasmodium stages (in vitro, orange panels). The cycle shows the developmental stages of Plasmodium sp. within the mosquito vector and inhibitory arrows indicate the stage targeted by the venom toxin. Plasmodium male and female gametocytes, the sexual stages that differentiate into gametes, are taken up by the mosquito in a bloodmeal and merge to form a zygote which develops into a motile ookinete. The ookinetes invade the epithelial lining of the mosquito midgut and differentiate into oocysts that generate sporozoites which are released into the hemolymph, invade the salivary gland and are injected in the skin of the next host during a blood meal.
Figure 1.
Overview of in vitro and in vivo studies showing activity of toxins against the mosquito stages of Plasmodium sp. Venom toxins that have shown anti-plasmodial activity within mosquitos (in vivo, pink panel), and anti-plasmodial activity when added directly to isolated Plasmodium stages (in vitro, orange panels). The cycle shows the developmental stages of Plasmodium sp. within the mosquito vector and inhibitory arrows indicate the stage targeted by the venom toxin. Plasmodium male and female gametocytes, the sexual stages that differentiate into gametes, are taken up by the mosquito in a bloodmeal and merge to form a zygote which develops into a motile ookinete. The ookinetes invade the epithelial lining of the mosquito midgut and differentiate into oocysts that generate sporozoites which are released into the hemolymph, invade the salivary gland and are injected in the skin of the next host during a blood meal.
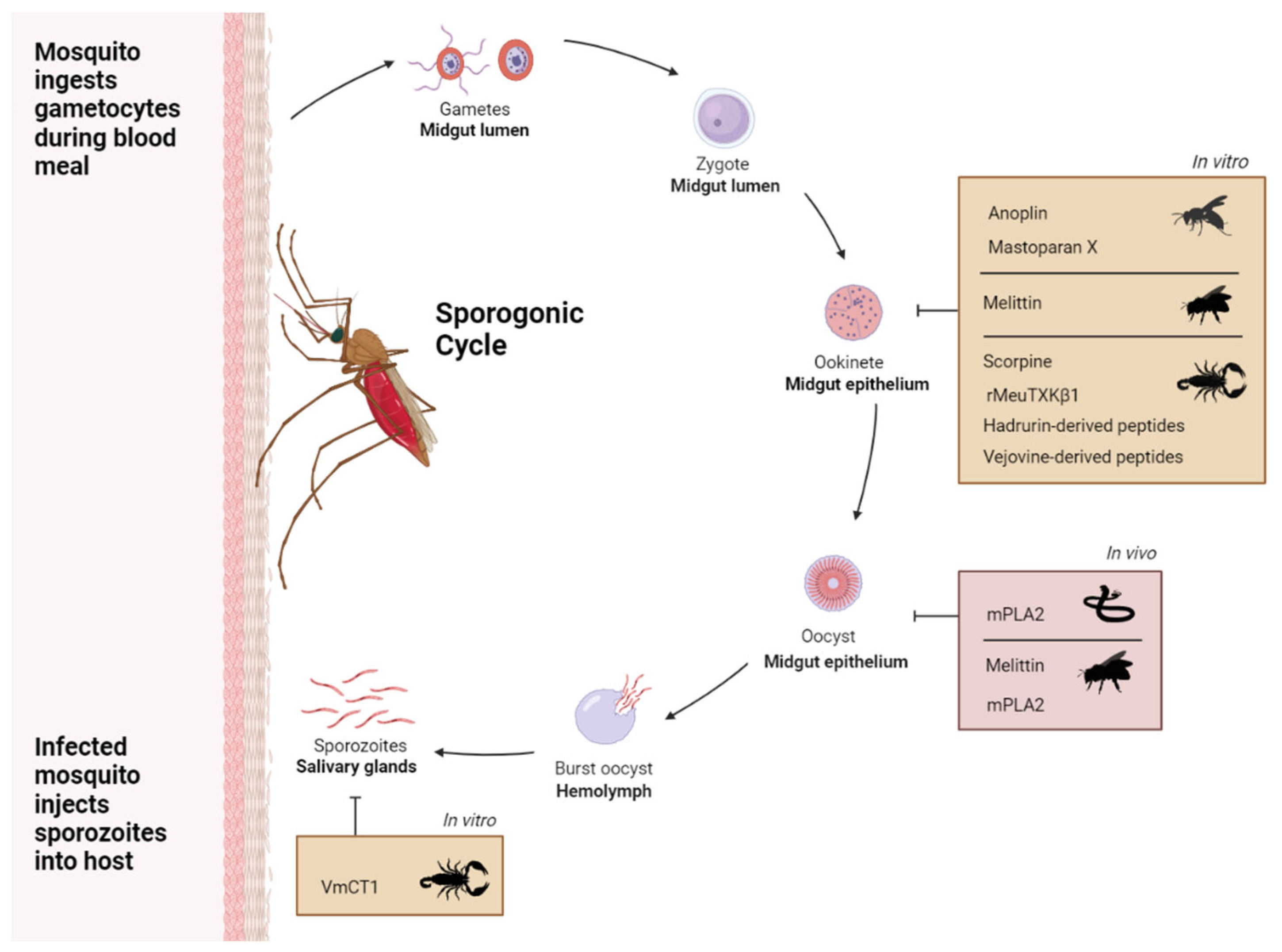
Figure 2.
Orthoflavivirus (ZIKV, DENV, YFV, JEV) and Alphavirus chikungunya (CHIKV) stages of infection and venom toxin targets. Venom toxins that have shown antiviral activity against (A) Orthoflavivirus and (B) Alphavirus chikungunya. The illustration shows the infection stages of the two classes of virus and inhibitory arrows indicate the stage targeted by the venom toxin. Viral particles attach and are internalized through clathrin-mediated endocytosis. Acidification of the endosome facilitates membrane fusion. (A) Once the Orthoflavivirus capsid protein is released, the capsid is disassembled, and virus genomic RNA transported to the ER for translation and replication. Immature virions bud from the ER and undergo maturation in the trans-golgi network. Virus exits by exocytosis [65]. (B) After the CHIKV genome release, viral RNA is directly translated into non-structural proteins (nsPs) and forms replication spherules, where viral genome replication occurs. Viral RNA is then translated to produce structural proteins. Capsid proteins and genomic RNA are assembled in the cytoplasm to form icosahedral nucleocapsid, and other structural polyproteins are translocated into ER for post-translational modification and delivered to the cell surface through the secretory pathway. The virus budding occurs when the nucleocapsid assembles with the modified structural proteins [66].
Figure 2.
Orthoflavivirus (ZIKV, DENV, YFV, JEV) and Alphavirus chikungunya (CHIKV) stages of infection and venom toxin targets. Venom toxins that have shown antiviral activity against (A) Orthoflavivirus and (B) Alphavirus chikungunya. The illustration shows the infection stages of the two classes of virus and inhibitory arrows indicate the stage targeted by the venom toxin. Viral particles attach and are internalized through clathrin-mediated endocytosis. Acidification of the endosome facilitates membrane fusion. (A) Once the Orthoflavivirus capsid protein is released, the capsid is disassembled, and virus genomic RNA transported to the ER for translation and replication. Immature virions bud from the ER and undergo maturation in the trans-golgi network. Virus exits by exocytosis [65]. (B) After the CHIKV genome release, viral RNA is directly translated into non-structural proteins (nsPs) and forms replication spherules, where viral genome replication occurs. Viral RNA is then translated to produce structural proteins. Capsid proteins and genomic RNA are assembled in the cytoplasm to form icosahedral nucleocapsid, and other structural polyproteins are translocated into ER for post-translational modification and delivered to the cell surface through the secretory pathway. The virus budding occurs when the nucleocapsid assembles with the modified structural proteins [66].


Table 1.
Summary of the toxin effectors used in paratransgenesis studies that target Plasmodium. Abbreviation: mPLA2, inactive mutant (H67N) PLA2.
Table 1.
Summary of the toxin effectors used in paratransgenesis studies that target Plasmodium. Abbreviation: mPLA2, inactive mutant (H67N) PLA2.
| Venom Effector | Mosquito Species | Vector Symbiont | Effect | Ref |
|---|---|---|---|---|
| Scorpine | An. stephensi | Asaia bogorensis | 63% reduction in oocyst number of Plasmodium berghei | [29] |
| Scorpine | An. stephensi | Asaia bogorensis | 90% reduction in oocyst number of Plasmodium berghei | [37] |
| mPLA2 | An. stephensi | Escherichia coli | 23% reduction in oocyst number of Plasmodium berghei | [26] |
| Scorpine & mPLA2 | An.gambiea An. stephensi | Pantoea agglomerans | 97.8% reduction (scorpine) and 85.3% reduction (mPLA2) in oocyst number of Plasmodium falciparum | [27] |
| Scorpine & mPLA2 | An. gambiae | Serratia marcescens | 93% reduction (scorpine) and 86% reduction (mPLA2) in oocyst number of Plasmodium falciparum | [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



