
Submitted:
01 September 2024
Posted:
03 September 2024
You are already at the latest version
Abstract
Leucine is an essential amino acid that cannot be produced endogenously in the human body and therefore needs to be obtained from dietary sources. Leucine plays a pivotal role in stimulating muscle protein synthesis along with isoleucine and valine as the group of branched-chain amino acids making them as one of the most popular dietary supplements for athletes and gym-goers. Individual effects of leucine, however, are not fully clarified, as most of the studies focused on the grouped effects of branched-chain amino acids. In recent years, leucine and its metabolites have been shown to stimulate muscle protein synthesis mainly via mammalian target of rapamycin complex 1 signaling pathway, thereby improving muscle atrophy in cancer cachexia. Interestingly, cancer research suggests that leucine may have either anti-cancer or carcinogenic effects. In the current manuscript, we aim to review leucine's roles in muscle protein synthesis, tumor suppression, and tumor progression, specifically summarizing the molecular mechanisms of leucine's action. The role of leucine is controversial in hepatocellular carcinoma, whereas its carcinogenic effects have been demonstrated in breast and pancreatic cancers. In summary, leucine being used as nutritional supplement for athletes needs more attention, as its oncogenic effects may have been identified by recent studies. Anti-cancer or carcinogenic effects of leucine in various cancers should be further investigated to have clear conclusions.
Keywords:
amino acid
; leucine
; muscle protein
; cancer
; signaling pathway
Introduction
From the 20 amino acids assembling human proteins 11 are classified as nonessential and 9 as essential. Nonessential amino acids are synthesized in the body, while the essential ones must be supplemented via diet to maintain human health [1]. Of the 9 essential amino acids, leucine (Leu), isoleucine (Ile), and valine (Val) are grouped as branched-chain amino acids (BCAAs) due to their branched aliphatic side chains [2]. BCAAs play pivotal role in various metabolic reactions and act as biochemical regulators of protein turnover [3,4,5]. BCAAs supplementation has increased in sports nutrition and became a vital part of the daily diet of bodybuilders and gym-goers, as BCAAs promote protein synthesis preventing its breakdown induced by intensive exercise [6,7,8]. They also play fundamental role in muscle's post-exercise recovery [9,10], and are implicated in the delay of fatigue by reducing brain's uptake of tryptophan and synthesis of 5-hydroxytryptamine [11,12]. These functions make BCAA's a “panacea” for professional and amateur athletes. As a result of such attention, the world sales of BCAA's have been stably increasing from year to year [13,14]. Interestingly, the potential benefits of BCAAs on performance, strength gains, and muscle mass have been a subject of ongoing debate [15,16,17]. Adding to this controversy, recent studies have shown that BCAA metabolism may be also associated with the progression of various types of tumors [18,19].
Even though the three BCAAs exhibit significant differences in their biological effects, they are often studied together as a single group, leading to potentially erroneous assumptions about their individual impacts [2]. Therefore, in this review, we primarily focus on Leu, as it is considered the most influential regulator and signaling molecule among the BCAAs [17,20].
1. Leucine and Muscle Protein Synthesis: Mechanisms and Impacts
The muscle protein mass is maintained by a dynamic equilibrium between muscle protein synthesis (MPS) and muscle protein breakdown (MPB) determining the net protein balance [21]. Leu has been shown to play a particularly central role in MPS [22] stimulating a robust MPS response in humans at a relatively low dose of about 3g, even in the absence of other amino acids [23]. Unlike other essential amino acids, Leu is mainly metabolized within the skeletal muscle where with the help of the mitochondrial branched-chain amino transferase 2 (BCAT2) Leu is converted to its keto-acid, α-ketoisocaproate (KIC), which can be further metabolized either to isovaleryl-CoA by branched-chain α-keto acid dehydrogenase complex (BCKDC) or to β-hydroxy-β-methylbutyrate (HMB) by KIC dioxygenase ultimately serving as energetic substrate for the citric acid cycle to produce energy (Figure 1) [23,24]. Both Leu itself and its metabolites, KIC and HMB, have been shown to increase MPS (Figure 1) [23,25,26]. Due to Leu's crucial role in regulating protein metabolism, it is used to combat protein loss in patients with various pathological conditions. This is supported by recent trials demonstrating that Leu supplementation improved sarcopenia in older adults [27,28,29,30]. The mechanism by which Leu and its metabolites enhance protein synthesis in muscle involves the activation of the mechanistic target of rapamycin complex 1 (mTORC1) signaling pathway [31,32]. This pathway is a key regulator that integrates various cellular processes, including protein synthesis [33]. mTORC1 senses signals from both intra- and extracellular cues including the availability of amino acids. As a result, nutrient sensing and responding to their availability are considered the primary functions of mTORC1. Specific transports are involved in bringing amino acids into the cells. For Leu, the solute carrier family 7 member 5 (SLC7A5)/SLC3A2, a heterodimeric bidirectional transporter, is responsible for transporting extracellular leucine into the cells in exchange for intracellular L-glutamine [34]. Within the cell, the lysosome is considered a key site for amino acid sensing, where specific proteins, known as amino acid sensors, help mTORC1 detect amino acids. For Leu, the major detectors in the cytosol are leucine-tRNA ligase (LARS1) and sestrin 2 (SESN2) [35,36,37,38]. These sensors closely interact with Ras-related GTP-binding proteins (RagA, B, C, and D), a small GTPase family, as well as GTPase-activating proteins, such as the multiprotein complex GTPase-activating proteins toward Rags (GATOR) such as GATOR1 and GATOR2, to communicate information about amino acid availability to mTORC1 [36]. Upon Leu availability, LARS1 specifically interacts with the RagA-RagC or RagB-RagD heterodimers, promoting the proper nucleotide loading state, which is essential for mTORC1 activation. LARS1 functions as a GTPase-activating protein towards RagD-GTP, which further facilitates the activation of the mTORC1 pathway [36]. Additionally, Leu binds to SESN2, causing its dissociation from GATOR2, which removes the inhibitory effect of GATOR1 on mTORC1, ultimately activating the pathway (Figure 1) [39,40]. Once activated, mTORC1 phosphorylates p70 ribosomal protein S6 kinase 1 (RPS6KB1) and eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1). The phosphorylation of RPS6KB1 enhances its kinase activity, further promoting protein translation, while the phosphorylation of 4E-BP1 prevents its association with eukaryotic translation initiation factor 4E (eIF4E), allowing translation initiation to occur (Figure 1) [41]. Through these mechanisms, Leu increases MPS via the activation of the mTORC1 pathway. However, Leu metabolites appear to be sensed differently from the Leu-sensing pathway described above. For example, KIC has been shown to increase protein synthesis by promoting the phosphorylation of mTORC1 substrates, RPS6KB1 and 4E-BP1 [26,42], although the upstream mechanism by which mTORC1 senses KIC remains unclear. More research has been conducted on HMB, another Leu metabolite. Earlier studies demonstrated that HMB increases the phosphorylation of mTORC1 downstream targets, RPS6KB1 and 4E-BP1, thereby enhancing skeletal MPS [43,44]. An independent study explored how HMB activates nutrient sensing upstream of mTORC1, suggesting that the effects of HMB on MPS may be mediated by the phosphatidylinositol 3-kinase (PI3K)-RAC-α serine/threonine-protein kinase (AKT1)-mTORC1 signaling axis [45]. Although the PI3K-AKT1 signaling pathway primarily transmits signals from growth factors and cytokines to mTORC1, acting as a major upstream regulator of the mTORC1 pathway, this study indicates that HMB might also influence mTORC1 through this pathway (Figure 1) [46,47]. PI3K, when activated in response to growth factors and cytokines, phosphorylates and activates AKT1. Active AKT1 then phosphorylates and inactivates TSC2, a component of the tuberous sclerosis complex (TSC), which includes TSC1, TSC2, and TBC1D7. The TSC complex is a negative regulator of mTORC1, so its inactivation leads to the activation of the mTORC1 pathway [48,49]. Girón et al. [45] demonstrated that HMB phosphorylates AKT1, mTOR, and its downstream targets, RPS6KB1 and 4E-BP1, thereby increasing protein synthesis. However, this study did not investigate whether AKT1 activates mTORC1 through the TSC complex, making it difficult to conclude whether HMB exerts its effects via the PI3K-AKT1-mTORC1 signaling axis. A more recent study provided evidence that HMB stimulates protein synthesis in skeletal muscle by inducing the autophosphorylation of mTOR, leading to the activation of the mTORC1 pathway without involving the SESN2-GATOR2 or Rag GTPase family proteins [32] (Figure 1). These findings suggest that Leu metabolites, such as KIC and HMB, may be sensed independently of the traditional Leu-sensing pathway. However, further research is needed to reach a consensus on the mechanisms by which Leu metabolites influence protein synthesis.
2. Leucine and Cancer Cachexia
As mentioned above, Leu has been shown to increase MPS and MPB [50,51]. In recent years, its potential to improve muscle wasting in cancer cachexia (CC) has been extensively studied [52,53,54]. CC is a complex syndrome characterized by glucose intolerance, loss of body fat, significant body weight loss due to muscle mass depletion, and malnutrition. As a result, nutritional supplementation including Leu, has been explored as a novel and promising therapeutic approach to protect cancer-associated cachexia [55]. However, concerns have arisen that Leu supplementation might also promote cancer growth and aggressiveness [56,57,58], suggesting that Leu could be a double-edged sword in cancer treatment.
2.1. Anti-Tumor Effects of Leucine
Recent investigations have explored the use of Leu as a potential adjunct therapy in cancer treatment, focusing on its effects in tumor-bearing models and various cancer cell lines.
Viana et al. observed that a diet rich in Leu led to a metabolic shift of Walker 256 rat in tumor towards a less glycolytic profile [54]. This shift resulted in reduced tumor glucose uptake, decreased tumor aggressiveness and metastatic sites in rats, but with no significant change in tumor size. Both Walker 256 rat tumor cells and tumor biopsies from rats fed the Leu-rich diet showed an increase in oxygen consumption, which was accompanied by an upregulation of mitochondrial genes, including proliferator-activated receptor γ coactivator-1 α (PGC-1α), nuclear respiratory factor -1 (NRF-1), cyclooxygenase (COX) 5a, citrate synthetase (CS), and cytochrome C, indicating enhanced mitochondrial biogenesis and oxidative phosphorylation (OXPHOS) (Figure 2). These findings suggest that Leu induces a metabolic shift in Walker-256 tumors, favoring OXPHOS over glycolysis, both in vitro and in vivo [54]. Cytotoxic effects of L-Leu supplementation on cancer have been demonstrated in hepatocellular carcinoma (HCC) cell lines [59]. In these studies, Leu supplementation exerted a dose-dependent cytotoxic effect and induced apoptosis, which was attributed to a decrease in insulin-like growth factor 1 (IGF-1) levels and an increase in p53 levels, leading to the inhibition of the PI3K/AKT1/mTORC1 signaling pathway (Figure 2). The combination of Leu supplementation with indoleamine 2,3-dioxygenase 1 (IDO1) and tryptophan 2,3-dioxygenase (TDO) inhibitors has been proposed as a promising novel therapeutic approach. IDO1 and TDO are enzymes involved in the catabolism of tryptophan into kynurenine, a substrate in the kynurenine pathway (KP), which is known for its potent immunomodulatory effects that cancer cells exploit to evade immune system destruction (Figure 2) [60,61,62,63]. Leu, as a substrate of the system L transporter, competes with kynurenine. Consequently, Leu limits the powerful immunosuppressive effects of the KP and restores antitumor immunity when used in conjunction with IDO1 and TDO inhibitors [64]. Additionally, kynurenine interacts with transcription factors such as the aryl hydrocarbon receptor, which promotes the differentiation and activation of immunosuppressive T-regulatory cells, while inhibiting the proliferation of T cells and natural killer cells by increasing the expression of programmed cell death protein 1 (PD-1), a regulatory checkpoint molecule for T cells [65,66,67,68]. A recent study further supported the antitumor effects of Leu, particularly when combined with PD-1 inhibition. This combination was shown to reverse immune regulation and enhance the antitumor activity of CD8+ tumor-infiltrating lymphocytes (TILs), mediated by the activation of the mTORC1 signaling pathway (Figure 2) [69]. The inhibitory effect of Leu, in combination with IDO1 and TDO inhibitors on kynurenine, represents a promising therapeutic approach for the future.
2.2. Tumorigenic Effects of Leucine
Although the studies mentioned above in Section 2.1 highlighted the anticancer effects of Leu, an increasing body of research suggests that Leu may also have oncogenic effects. Like other amino acids, Leu can serve as an energy source, potentially fueling cancer development [70]. The oncogenic effects of Leu have been studied in various cancers, including hepatocellular carcinoma (HCC), breast cancer, and pancreatic cancer. In contrast to the findings of Hassan et al. [59], Chen et al. [71] demonstrated that the growth of HCC cell lines depends on the presence of Leu, with deprivation significantly reducing the proliferation, migration, and invasion of cancer cells. This oncogenic effect was linked to the regulation of Leu metabolism by enzymes such as the mitochondrial methylcrotonoyl-CoA carboxylase (MCC) enzyme, which is highly expressed and associated with poor prognosis in HCC patients [71] (Figure 3). The oncogenic role of Leu in HCC has also been supported by recent metabolomics studies, which identified Leu as one of the amino acids significantly upregulated in both HCC tumor tissues and the serum of HCC patients [72]. As a result, dietary amino acid supplementation has become an increasingly researched area as a potential cancer treatment strategy (Figure 3). In vitro studies on breast cancer have shown that Leu deprivation inhibits cancer cell proliferation and induces apoptosis, while in vivo studies demonstrated inhibited tumor growth [73]. Another recent study on breast cancer found that a high-fat diet, leading to the abundant release of Leu, promoted cancer progression in tumor-bearing mice. This effect was mediated by the activation of the mTOR signaling pathway, which led to the differentiation and infiltration of polymorphonuclear myeloid-derived suppressor cells, a factor associated with poor clinical outcomes in breast cancer patients [74]. Complementing these observations, the oncogenic effects of Leu have also been demonstrated in bladder cancer and pancreatic cancer. Long-term supplementation with excessive amounts of Leu promoted bladder carcinogenesis in rats [58,75], suggesting that both the duration and amount of supplementation play a role in Leu's oncogenic potential. In pancreatic cancer, Leu supplementation increased tumor growth in both lean and overweight mice, but through different mechanisms. In lean mice, Leu promotes tumor growth by activating the mTOR signaling pathway, while in overweight mice, Leu supplementation increases the amount of glucose available to tumor cells, further accelerating tumor growth (Figure 3) [56].
In a separate study, Leu was found to promote the growth and proliferation of pancreatic cancer cells by stimulating the expression of SESN2 and increasing phosphorylated mTOR (p-mTOR), indicating the involvement of the mTOR signaling pathway in pancreatic cancer development. The use of mTOR inhibitors and SESN2 expression vectors in this study clearly demonstrated that SESN2 promotes glycolysis in pancreatic cancer through the mTOR signaling pathway (Figure 3) [73].
Conclusions
Leu, as part of BCAAs has become one of the most popular sport supplements due to its role in promoting MPS [17]. In the light of this, Leu and its metabolites have been found to improve muscle wasting in CC [76]. However, the role of Leu remains controversial in certain cancers [59,71]. Through this review of the literature, Leu has been identified as a 'dichotomous' amino acid, exhibiting both anticancer and carcinogenic effects. In conclusion, Leu has been found to exhibits significant anticancer activity [54,59,64,69], leading to cell death by regulating tumor cell metabolism, apoptosis, and immune signaling pathways. On the other hand, Leu also demonstrates oncogenic activity, promoting tumor cell proliferation through various mechanisms depending on the cancer type. For example, Leu can regulate its own metabolism in HCC [71,72], increase glucose availability to tumor cells, or activate the mTOR signaling pathway in pancreatic, breast, and bladder cancers [56,73,74,75,77]. The carcinogenic effects of Leu may be achieved in several ways such as by supporting the metabolic reprogramming of cancer cells, similar to other amino acids [78], or by stimulating tumorigenesis through the modulation of oncogenic signaling pathways [79]. Additionally, the duration and amount of Leu supplementation appear to influence its carcinogenic activity [58,75]. The dual nature of Leu in cancer biology reveals its complexity and emphasizes the need for careful evaluation. The future use of Leu, either as a nutritional supplement or a potential cancer treatment, will depend on further research clarifying its effects. Moreover, suggestions on targeting enzymes involved in Leu metabolism and Leu dietary interventions as treatment strategies may present challenges, which may arise due to the complex nature of Leu metabolism, individual variability in response, potential side effects, and limited research on long-term effects and optimal dosages.
Author Contributions
Conceptualization: T.T, B.A. Resources: B.A., Z.O. Writing – original draft preparation B.A., Z.O. Writing – review and editing: T.T, F.M, O.K, T.A, B.S. B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Nazarbayev University under Collaborative Research Grants OPCRP2023015.
Data Availability Statement
All relevant data are available within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wu, G.; Wu, Z.; Dai, Z.; Yang, Y.; Wang, W.; Liu, C.; Wang, B.; Wang, J.; Yin, Y. Dietary Requirements of “Nutritionally Non-Essential Amino Acids” by Animals and Humans. Amino Acids 2013, 44, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Neinast, M.; Murashige, D.; Arany, Z. Branched Chain Amino Acids. Annu. Rev. Physiol. 2019, 81, 139–164. [Google Scholar] [CrossRef] [PubMed]
- Lal, H.; Chugh, K. Metabolic and Regulatory Effects of Branched Chain Amino Acid Supplementation. Nutrition Research 1995, 15, 1717–1733. [Google Scholar] [CrossRef]
- May, M.E.; Buse, M.G. Effects of Branched-Chain Amino Acids on Protein Turnover. Diabetes Metab. Rev. 1989, 5, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.R.; Phillips, S.M.; Stellingwerff, T.; Rezzi, S.; Bruce, S.J.; Breton, I.; Thorimbert, A.; Guy, P.A.; Clarke, J.; Broadbent, S.; et al. A Protein–Leucine Supplement Increases Branched-Chain Amino Acid and Nitrogen Turnover But Not Performance. Medicine & Science in Sports & Exercise 2012, 44, 57–68. [Google Scholar] [CrossRef]
- Churchward-Venne, T.A.; Burd, N.A.; Phillips, S.M. Nutritional Regulation of Muscle Protein Synthesis with Resistance Exercise: Strategies to Enhance Anabolism. Nutr Metab (Lond) 2012, 9, 40. [Google Scholar] [CrossRef]
- Jackman, S.R.; Witard, O.C.; Philp, A.; Wallis, G.A.; Baar, K.; Tipton, K.D. Branched-Chain Amino Acid Ingestion Stimulates Muscle Myofibrillar Protein Synthesis Following Resistance Exercise in Humans. Front. Physiol. 2017, 8, 390. [Google Scholar] [CrossRef]
- Wang, C.; Guo, F. Branched Chain Amino Acids and Metabolic Regulation. Chin. Sci. Bull. 2013, 58, 1228–1235. [Google Scholar] [CrossRef]
- Lynch, C.J.; Adams, S.H. Branched-Chain Amino Acids in Metabolic Signalling and Insulin Resistance. Nat Rev Endocrinol 2014, 10, 723–736. [Google Scholar] [CrossRef]
- Rahimi, M.H.; Shab-Bidar, S.; Mollahosseini, M.; Djafarian, K. Branched-Chain Amino Acid Supplementation and Exercise-Induced Muscle Damage in Exercise Recovery: A Meta-Analysis of Randomized Clinical Trials. Nutrition 2017, 42, 30–36. [Google Scholar] [CrossRef]
- Blomstrand, E. A Role for Branched-Chain Amino Acids in Reducing Central Fatigue. The Journal of Nutrition 2006, 136, 544S–547S. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, M.; Sisti, D.; Amatori, S.; Donati Zeppa, S.; Annibalini, G.; Piccoli, G.; Vallorani, L.; Benelli, P.; Rocchi, M.B.L.; Barbieri, E.; et al. Effects of a Commercially Available Branched-Chain Amino Acid-Alanine-Carbohydrate-Based Sports Supplement on Perceived Exertion and Performance in High Intensity Endurance Cycling Tests. Journal of the International Society of Sports Nutrition 2020, 17, 6. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, R.R. Branched-Chain Amino Acids and Muscle Protein Synthesis in Humans: Myth or Reality? Journal of the International Society of Sports Nutrition 2017, 14, 30. [Google Scholar] [CrossRef]
- Global BCAA Market [2024-2032] | Advanced Research Report.
- Gleeson, M. Interrelationship between Physical Activity and Branched-Chain Amino Acids. The Journal of Nutrition 2005, 135, 1591S–1595S. [Google Scholar] [CrossRef]
- Marcon, M.; Zanella, P.B. The Effect of Branched-Chain Amino Acids Supplementation in Physical Exercise: A Systematic Review of Human Randomized Controlled Trials. Science & Sports 2022, S0765159721002008. [Google Scholar] [CrossRef]
- Plotkin, D.L.; Delcastillo, K.; Van Every, D.W.; Tipton, K.D.; Aragon, A.A.; Schoenfeld, B.J. Isolated Leucine and Branched-Chain Amino Acid Supplementation for Enhancing Muscular Strength and Hypertrophy: A Narrative Review. International Journal of Sport Nutrition and Exercise Metabolism 2021, 31, 292–301. [Google Scholar] [CrossRef]
- Han, L.; Dong, L.; Leung, K.; Zhao, Z.; Li, Y.; Gao, L.; Chen, Z.; Xue, J.; Qing, Y.; Li, W.; et al. METTL16 Drives Leukemogenesis and Leukemia Stem Cell Self-Renewal by Reprogramming BCAA Metabolism. Cell Stem Cell 2023, 30, 52–68.e13. [Google Scholar] [CrossRef]
- Lee, J.H.; Cho, Y.; Kim, J.H.; Kim, J.; Nam, H.Y.; Kim, S.W.; Son, J. Branched-Chain Amino Acids Sustain Pancreatic Cancer Growth by Regulating Lipid Metabolism. Exp Mol Med 2019, 51, 1–11. [Google Scholar] [CrossRef]
- Columbus, D.A.; Fiorotto, M.L.; Davis, T.A. Leucine Is a Major Regulator of Muscle Protein Synthesis in Neonates. Amino Acids 2015, 47, 259–270. [Google Scholar] [CrossRef]
- Kang, M.C. Muscle Protein Metabolism in Critically Illness. Surg Metab Nutr 2020, 11, 35–39. [Google Scholar] [CrossRef]
- Zaromskyte, G.; Prokopidis, K.; Ioannidis, T.; Tipton, K.D.; Witard, O.C. Evaluating the Leucine Trigger Hypothesis to Explain the Post-Prandial Regulation of Muscle Protein Synthesis in Young and Older Adults: A Systematic Review. Front. Nutr. 2021, 8, 685165. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.J.; Hossain, T.; Hill, D.S.; Phillips, B.E.; Crossland, H.; Williams, J.; Loughna, P.; Churchward-Venne, T.A.; Breen, L.; Phillips, S.M.; et al. Effects of Leucine and Its Metabolite Β-hydroxy-β-methylbutyrate on Human Skeletal Muscle Protein Metabolism. The Journal of Physiology 2013, 591, 2911–2923. [Google Scholar] [CrossRef] [PubMed]
- Ananieva, E.A.; Powell, J.D.; Hutson, S.M. Leucine Metabolism in T Cell Activation: mTOR Signaling and Beyond. Advances in Nutrition 2016, 7, 798S–805S. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Li, F.; Song, B.; Zheng, C.; Zhong, Y.; Xu, K.; Kong, X.; Yin, Y.; Wang, W.; Shu, G. β-Hydroxy-β-Methyl Butyrate, but Not α-Ketoisocaproate and Excess Leucine, Stimulates Skeletal Muscle Protein Metabolism in Growing Pigs Fed Low-Protein Diets. Journal of Functional Foods 2019, 52, 34–42. [Google Scholar] [CrossRef]
- Escobar, J.; Frank, J.W.; Suryawan, A.; Nguyen, H.V.; Van Horn, C.G.; Hutson, S.M.; Davis, T.A. Leucine and α-Ketoisocaproic Acid, but Not Norleucine, Stimulate Skeletal Muscle Protein Synthesis in Neonatal Pigs. The Journal of Nutrition 2010, 140, 1418–1424. [Google Scholar] [CrossRef] [PubMed]
- Cereda, E.; Pisati, R.; Rondanelli, M.; Caccialanza, R. Whey Protein, Leucine- and Vitamin-D-Enriched Oral Nutritional Supplementation for the Treatment of Sarcopenia. Nutrients 2022, 14, 1524. [Google Scholar] [CrossRef]
- Gielen, E.; Beckwée, D.; Delaere, A.; De Breucker, S.; Vandewoude, M.; Bautmans, I.; the Sarcopenia Guidelines Development Group of the Belgian Society of Gerontology and Geriatrics (BSGG); Bautmans, I. ; Beaudart, C.; Beckwée, D.; et al. Nutritional Interventions to Improve Muscle Mass, Muscle Strength, and Physical Performance in Older People: An Umbrella Review of Systematic Reviews and Meta-Analyses. Nutrition Reviews 2021, 79, 121–147. [Google Scholar] [CrossRef]
- Martínez-Arnau, F.M.; Fonfría-Vivas, R.; Buigues, C.; Castillo, Y.; Molina, P.; Hoogland, A.J.; van Doesburg, F.; Pruimboom, L.; Fernández-Garrido, J.; Cauli, O. Effects of Leucine Administration in Sarcopenia: A Randomized and Placebo-Controlled Clinical Trial. Nutrients 2020, 12, 932. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Bise, T.; Shimazu, S.; Tanoue, M.; Tomioka, Y.; Araki, M.; Nishino, T.; Kuzuhara, A.; Takatsuki, F. Effects of a Leucine-Enriched Amino Acid Supplement on Muscle Mass, Muscle Strength, and Physical Function in Post-Stroke Patients with Sarcopenia: A Randomized Controlled Trial. Nutrition 2019, 58, 1–6. [Google Scholar] [CrossRef]
- Bodine, S.C. The Role of mTORC1 in the Regulation of Skeletal Muscle Mass. Fac Rev 2022, 11. [Google Scholar] [CrossRef]
- Suryawan, A.; Rudar, M.; Fiorotto, M.L.; Davis, T.A. Differential Regulation of mTORC1 Activation by Leucine and β-Hydroxy-β-Methylbutyrate in Skeletal Muscle of Neonatal Pigs. Journal of Applied Physiology 2020, 128, 286–295. [Google Scholar] [CrossRef]
- Akbay, B.; Shmakova, A.; Vassetzky, Y.; Dokudovskaya, S. Modulation of mTORC1 Signaling Pathway by HIV-1. Cells 2020, 9, 1090–1090. [Google Scholar] [CrossRef]
- Nicklin, P.; Bergman, P.; Zhang, B.; Triantafellow, E.; Wang, H.; Nyfeler, B.; Yang, H.; Hild, M.; Kung, C.; Wilson, C.; et al. Bidirectional Transport of Amino Acids Regulates mTOR and Autophagy. Cell 2009, 136, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Chantranupong, L.; Wolfson, R.L.; Orozco, J.M.; Saxton, R.A.; Scaria, S.M.; Bar-Peled, L.; Spooner, E.; Isasa, M.; Gygi, S.P.; Sabatini, D.M. The Sestrins Interact with GATOR2 to Negatively Regulate the Amino-Acid-Sensing Pathway Upstream of mTORC1. Cell Reports 2014, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Jeong, S.J.; Park, M.C.; Kim, G.; Kwon, N.H.; Kim, H.K.; Ha, S.H.; Ryu, S.H.; Kim, S. Leucyl-tRNA Synthetase Is an Intracellular Leucine Sensor for the mTORC1-Signaling Pathway. Cell 2012, 149, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Ro, S.-H.; Kim, M.; Park, H.-W.; Semple, I.A.; Park, H.; Cho, U.-S.; Wang, W.; Guan, K.-L.; Karin, M.; et al. Sestrin2 Inhibits mTORC1 through Modulation of GATOR Complexes. Scientific Reports 2015, 5, 9502–9502. [Google Scholar] [CrossRef]
- Parmigiani, A.; Nourbakhsh, A.; Ding, B.; Wang, W.; Kim, Y.C.; Akopiants, K.; Guan, K.-L.; Karin, M.; Budanov, A.V. Sestrins Inhibit mTORC1 Kinase Activation through the GATOR Complex. Cell Reports 2014, 9, 1281–1291. [Google Scholar] [CrossRef]
- Saxton, R.A.; Knockenhauer, K.E.; Wolfson, R.L.; Chantranupong, L.; Pacold, M.E.; Wang, T.; Schwartz, T.U.; Sabatini, D.M. Structural Basis for Leucine Sensing by the Sestrin2-mTORC1 Pathway. Science 2016, 351, 53–58. [Google Scholar] [CrossRef]
- Wolfson, R.L.; Chantranupong, L.; Saxton, R.A.; Shen, K.; Scaria, S.M.; Cantor, J.R.; Sabatini, D.M. Sestrin2 Is a Leucine Sensor for the mTORC1 Pathway. Science 2016, 351, 43–48. [Google Scholar] [CrossRef]
- Yang, M.; Lu, Y.; Piao, W.; Jin, H. The Translational Regulation in mTOR Pathway. Biomolecules 2022, 12, 802. [Google Scholar] [CrossRef]
- Moghei, M.; Tavajohi-Fini, P.; Beatty, B.; Adegoke, O.A.J. Ketoisocaproic Acid, a Metabolite of Leucine, Suppresses Insulin-Stimulated Glucose Transport in Skeletal Muscle Cells in a BCAT2-Dependent Manner. American Journal of Physiology-Cell Physiology 2016, 311, C518–C527. [Google Scholar] [CrossRef]
- Kao, M.; Columbus, D.A.; Suryawan, A.; Steinhoff-Wagner, J.; Hernandez-Garcia, A.; Nguyen, H.V.; Fiorotto, M.L.; Davis, T.A. Enteral β-Hydroxy-β-Methylbutyrate Supplementation Increases Protein Synthesis in Skeletal Muscle of Neonatal Pigs. American Journal of Physiology-Endocrinology and Metabolism 2016, 310, E1072–E1084. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.J.; Hossain, T.; Limb, M.C.; Phillips, B.E.; Lund, J.; Williams, J.P.; Brook, M.S.; Cegielski, J.; Philp, A.; Ashcroft, S.; et al. Impact of the Calcium Form of β-Hydroxy-β-Methylbutyrate upon Human Skeletal Muscle Protein Metabolism. Clinical Nutrition 2018, 37, 2068–2075. [Google Scholar] [CrossRef] [PubMed]
- Girón, M.D.; Vílchez, J.D.; Salto, R.; Manzano, M.; Sevillano, N.; Campos, N.; Argilés, J.M.; Rueda, R.; López-Pedrosa, J.M. Conversion of Leucine to β-Hydroxy-β-Methylbutyrate by α-Keto Isocaproate Dioxygenase Is Required for a Potent Stimulation of Protein Synthesis in L6 Rat Myotubes: HMB Is a Potent Stimulator of Protein Synthesis. Journal of Cachexia, Sarcopenia and Muscle 2016, 7, 68–78. [Google Scholar] [CrossRef]
- Huang, H.; Long, L.; Zhou, P.; Chapman, N.M.; Chi, H. mTOR Signaling at the Crossroads of Environmental Signals and T-cell Fate Decisions. Immunological Reviews 2020, 295, 15–38. [Google Scholar] [CrossRef]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef]
- Cai, S.-L.; Tee, A.R.; Short, J.D.; Bergeron, J.M.; Kim, J.; Shen, J.; Guo, R.; Johnson, C.L.; Kiguchi, K.; Walker, C.L. Activity of TSC2 Is Inhibited by AKT-Mediated Phosphorylation and Membrane Partitioning. Journal of Cell Biology 2006, 173, 279–289. [Google Scholar] [CrossRef]
- Dibble, C.C.; Elis, W.; Menon, S.; Qin, W.; Klekota, J.; Asara, J.M.; Finan, P.M.; Kwiatkowski, D.J.; Murphy, L.O.; Manning, B.D. TBC1D7 Is a Third Subunit of the TSC1-TSC2 Complex Upstream of mTORC1. Molecular Cell 2012, 47, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Garlick, P.J. The Role of Leucine in the Regulation of Protein Metabolism. The Journal of Nutrition 2005, 135, 1553S–1556S. [Google Scholar] [CrossRef]
- Ham, D.J.; Caldow, M.K.; Lynch, G.S.; Koopman, R. Leucine as a Treatment for Muscle Wasting: A Critical Review. Clinical Nutrition 2014, 33, 937–945. [Google Scholar] [CrossRef]
- Beaudry, A.G.; Law, M.L. Leucine Supplementation in Cancer Cachexia: Mechanisms and a Review of the Pre-Clinical Literature. Nutrients 2022, 14, 2824. [Google Scholar] [CrossRef] [PubMed]
- Cruz, B.; Oliveira, A.; Gomes-Marcondes, M.C.C. L-Leucine Dietary Supplementation Modulates Muscle Protein Degradation and Increases pro-Inflammatory Cytokines in Tumour-Bearing Rats. Cytokine 2017, 96, 253–260. [Google Scholar] [CrossRef]
- Viana, L.R.; Tobar, N.; Busanello, E.N.B.; Marques, A.C.; De Oliveira, A.G.; Lima, T.I.; Machado, G.; Castelucci, B.G.; Ramos, C.D.; Brunetto, S.Q.; et al. Leucine-Rich Diet Induces a Shift in Tumour Metabolism from Glycolytic towards Oxidative Phosphorylation, Reducing Glucose Consumption and Metastasis in Walker-256 Tumour-Bearing Rats. Sci Rep 2019, 9, 15529. [Google Scholar] [CrossRef] [PubMed]
- Maschke, J.; Kruk, U.; Kastrati, K.; Kleeberg, J.; Buchholz, D.; Erickson, N.; Huebner, J. Nutritional Care of Cancer Patients: A Survey on Patients’ Needs and Medical Care in Reality. Int J Clin Oncol 2017, 22, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.A.; Lashinger, L.M.; Rasmussen, A.J.; Hursting, S.D. Leucine Supplementation Differentially Enhances Pancreatic Cancer Growth in Lean and Overweight Mice. Cancer Metab 2014, 2, 6. [Google Scholar] [CrossRef]
- Schrems, E.R.; Haynie, W.S.; Perry, R.A.; Morena, F.; Cabrera, A.R.; Rosa-Caldwell, M.E.; Greene, N.P.; Washington, T.A. Leucine Supplementation Exacerbates Morbidity in Male but Not Female Mice with Colorectal Cancer-Induced Cachexia. Nutrients 2023, 15, 4570. [Google Scholar] [CrossRef]
- Xie, X.-L.; Wei, M.; Yunoki, T.; Kakehashi, A.; Yamano, S.; Kato, M.; Wanibuchi, H. Long-Term Treatment with l-Isoleucine or l-Leucine in AIN-93G Diet Has Promoting Effects on Rat Bladder Carcinogenesis. Food and Chemical Toxicology 2012, 50, 3934–3940. [Google Scholar] [CrossRef]
- Hassan, Y.A.; Helmy, M.W.; Ghoneim, A.I. Combinatorial Antitumor Effects of Amino Acids and Epigenetic Modulations in Hepatocellular Carcinoma Cell Lines. Naunyn-Schmiedeberg’s Arch Pharmacol 2021, 394, 2245–2257. [Google Scholar] [CrossRef]
- Ball, H.J.; Fedelis, F.F.; Bakmiwewa, S.M.; Hunt, N.H.; Yuasa, H.J. Tryptophan-Catabolizing Enzymes €“ Party of Three. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef]
- Hoffmann, D.; Dvorakova, T.; Stroobant, V.; Bouzin, C.; Daumerie, A.; Solvay, M.; Klaessens, S.; Letellier, M.-C.; Renauld, J.-C.; Van Baren, N.; et al. Tryptophan 2,3-Dioxygenase Expression Identified in Human Hepatocellular Carcinoma Cells and in Intratumoral Pericytes of Most Cancers. Cancer Immunology Research 2020, 8, 19–31. [Google Scholar] [CrossRef]
- Pilotte, L.; Larrieu, P.; Stroobant, V.; Colau, D.; Dolušić, E.; Frédérick, R.; De Plaen, E.; Uyttenhove, C.; Wouters, J.; Masereel, B.; et al. Reversal of Tumoral Immune Resistance by Inhibition of Tryptophan 2,3-Dioxygenase. Proc. Natl. Acad. Sci. U.S.A. 2012, 109, 2497–2502. [Google Scholar] [CrossRef] [PubMed]
- Uyttenhove, C.; Pilotte, L.; Théate, I.; Stroobant, V.; Colau, D.; Parmentier, N.; Boon, T.; Van Den Eynde, B.J. Evidence for a Tumoral Immune Resistance Mechanism Based on Tryptophan Degradation by Indoleamine 2,3-Dioxygenase. Nat Med 2003, 9, 1269–1274. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Tomek, P. Tryptophan: A Rheostat of Cancer Immune Escape Mediated by Immunosuppressive Enzymes IDO1 and TDO. Front. Immunol. 2021, 12, 636081. [Google Scholar] [CrossRef] [PubMed]
- Chiesa, M.D.; Carlomagno, S.; Frumento, G.; Balsamo, M.; Cantoni, C.; Conte, R.; Moretta, L.; Moretta, A.; Vitale, M. The Tryptophan Catabolite L-Kynurenine Inhibits the Surface Expression of NKp46- and NKG2D-Activating Receptors and Regulates NK-Cell Function. Blood 2006, 108, 4118–4125. [Google Scholar] [CrossRef]
- Liu, Y.; Liang, X.; Dong, W.; Fang, Y.; Lv, J.; Zhang, T.; Fiskesund, R.; Xie, J.; Liu, J.; Yin, X.; et al. Tumor-Repopulating Cells Induce PD-1 Expression in CD8+ T Cells by Transferring Kynurenine and AhR Activation. Cancer Cell 2018, 33, 480–494.e7. [Google Scholar] [CrossRef] [PubMed]
- Mezrich, J.D.; Fechner, J.H.; Zhang, X.; Johnson, B.P.; Burlingham, W.J.; Bradfield, C.A. An Interaction between Kynurenine and the Aryl Hydrocarbon Receptor Can Generate Regulatory T Cells. The Journal of Immunology 2010, 185, 3190–3198. [Google Scholar] [CrossRef]
- Rad Pour, S.; Morikawa, H.; Kiani, N.A.; Yang, M.; Azimi, A.; Shafi, G.; Shang, M.; Baumgartner, R.; Ketelhuth, D.F.J.; Kamleh, M.A.; et al. Exhaustion of CD4+ T-Cells Mediated by the Kynurenine Pathway in Melanoma. Sci Rep 2019, 9, 12150. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, H.; Liu, W.; Yan, S.-M.; Li, Y.; Tan, L.; Chen, Y.; Liu, J.; Peng, Z.; Yuan, Y.; et al. Amino Acids and RagD Potentiate mTORC1 Activation in CD8 + T Cells to Confer Antitumor Immunity. J Immunother Cancer 2021, 9, e002137. [Google Scholar] [CrossRef]
- Keenan, M.M.; Chi, J.-T. Alternative Fuels for Cancer Cells. The Cancer Journal 2015, 21, 49–55. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Zhang, X.-N.; Xu, C.-Z.; Zhou, D.-H.; Chen, J.; Liu, Z.-X.; Sun, Y.; Huang, W.; Qu, L.-S. MCCC2 Promotes HCC Development by Supporting Leucine Oncogenic Function. Cancer Cell Int 2021, 21, 22. [Google Scholar] [CrossRef]
- Morine, Y.; Utsunomiya, T.; Yamanaka-Okumura, H.; Saito, Y.; Yamada, S.; Ikemoto, T.; Imura, S.; Kinoshita, S.; Hirayama, A.; Tanaka, Y.; et al. Essential Amino Acids as Diagnostic Biomarkers of Hepatocellular Carcinoma Based on Metabolic Analysis. Oncotarget 2022, 13, 1286–1298. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Wang, C.; Yin, H.; Yu, J.; Chen, S.; Fang, J.; Guo, F. Leucine Deprivation Inhibits Proliferation and Induces Apoptosis of Human Breast Cancer Cells via Fatty Acid Synthase. Oncotarget 2016, 7, 63679–63689. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, X.; Zou, Y.; Gong, J.; Ge, Z.; Lin, X.; Zhang, W.; Huang, H.; Zhao, J.; Saw, P.E.; et al. A High-Fat Diet Promotes Cancer Progression by Inducing Gut Microbiota–Mediated Leucine Production and PMN-MDSC Differentiation. Proc. Natl. Acad. Sci. U.S.A. 2024, 121, e2306776121. [Google Scholar] [CrossRef]
- Gi, M.; Wanibuchi, H. Roles of Leucine and Isoleucine in Experimental Models of Bladder Carcinogenesis. Food Safety 2015, 3, 136–142. [Google Scholar] [CrossRef]
- Aversa, Z.; Costelli, P.; Muscaritoli, M. Cancer-Induced Muscle Wasting: Latest Findings in Prevention and Treatment. Ther Adv Med Oncol 2017, 9, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhu, H.; Weng, M.; Zhang, H.; Wang, C.; Sun, L. CC-223, NSC781406, and BGT226 Exerts a Cytotoxic Effect Against Pancreatic Cancer Cells via mTOR Signaling. Front. Pharmacol. 2020, 11, 580407. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, X.; Cheng, C.; Yu, W.; Yi, P. Metabolism of Amino Acids in Cancer. Front. Cell Dev. Biol. 2021, 8, 603837. [Google Scholar] [CrossRef]
- Chen, J.; Cui, L.; Lu, S.; Xu, S. Amino Acid Metabolism in Tumor Biology and Therapy. Cell Death Dis 2024, 15, 42. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the action of Leu and its metabolites. Solute carrier family 7 member 5 (SLC7A5) transports extracellular Leu into the cells. Amino acid sensors, leucyl-tRNA ligase (LARS1) and sestrin 2 (SESN2), detect cytosolic Leu. SESN2 alleviates the inhibitory function of GTPase-activating protein toward Rags 1 (GATOR1) and GTPase-activating protein toward Rags 2 (GATOR2), leading to the activation of the mechanistic target of rapamycin complex 1 (mTORC1) signaling pathway, which further promotes muscle protein synthesis (MPS). Leu metabolites also promote MPS via mTORC1. Branched-chain amino transferase 2 (BCAT2) converts Leu to α-ketoisocaproate (KIC), which phosphorylates p70 ribosomal protein S6 kinase 1 (RPS6KB1) and eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1), both markers of mTORC1 activation, thereby increasing MPS. KIC can be further metabolized to β-hydroxy-β-methylbutyrate (HMB) by KIC dioxygenase and to isovaleryl-CoA by branched-chain α-keto acid dehydrogenase complex (BCKDC). Rag GTPases are crucial in this pathway. RagA/B typically bind GTP when Leu is abundant, while RagC/D are in the GDP-bound state. The RagA-RagC and RagB-RagD heterodimers are key pairs interacting with mTORC1 on the lysosomal membrane, activating it in response to Leu and promoting MPS. Both HMB and isovaleryl-CoA are involved in energy production. HMB increases MPS by activating the AKT1-mTORC1 signaling pathway. However, whether HMB promotes MPS via the AKT1-mTORC1 axis is not fully characterized, as the effect of HMB on the tuberous sclerosis complex 1-tuberous sclerosis complex 2-TBC1 domain family member 7 (TSC1-TSC2-TBC1D7) complex, the intermediate between AKT1 and mTORC1.
Figure 1.
Schematic representation of the action of Leu and its metabolites. Solute carrier family 7 member 5 (SLC7A5) transports extracellular Leu into the cells. Amino acid sensors, leucyl-tRNA ligase (LARS1) and sestrin 2 (SESN2), detect cytosolic Leu. SESN2 alleviates the inhibitory function of GTPase-activating protein toward Rags 1 (GATOR1) and GTPase-activating protein toward Rags 2 (GATOR2), leading to the activation of the mechanistic target of rapamycin complex 1 (mTORC1) signaling pathway, which further promotes muscle protein synthesis (MPS). Leu metabolites also promote MPS via mTORC1. Branched-chain amino transferase 2 (BCAT2) converts Leu to α-ketoisocaproate (KIC), which phosphorylates p70 ribosomal protein S6 kinase 1 (RPS6KB1) and eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1), both markers of mTORC1 activation, thereby increasing MPS. KIC can be further metabolized to β-hydroxy-β-methylbutyrate (HMB) by KIC dioxygenase and to isovaleryl-CoA by branched-chain α-keto acid dehydrogenase complex (BCKDC). Rag GTPases are crucial in this pathway. RagA/B typically bind GTP when Leu is abundant, while RagC/D are in the GDP-bound state. The RagA-RagC and RagB-RagD heterodimers are key pairs interacting with mTORC1 on the lysosomal membrane, activating it in response to Leu and promoting MPS. Both HMB and isovaleryl-CoA are involved in energy production. HMB increases MPS by activating the AKT1-mTORC1 signaling pathway. However, whether HMB promotes MPS via the AKT1-mTORC1 axis is not fully characterized, as the effect of HMB on the tuberous sclerosis complex 1-tuberous sclerosis complex 2-TBC1 domain family member 7 (TSC1-TSC2-TBC1D7) complex, the intermediate between AKT1 and mTORC1.
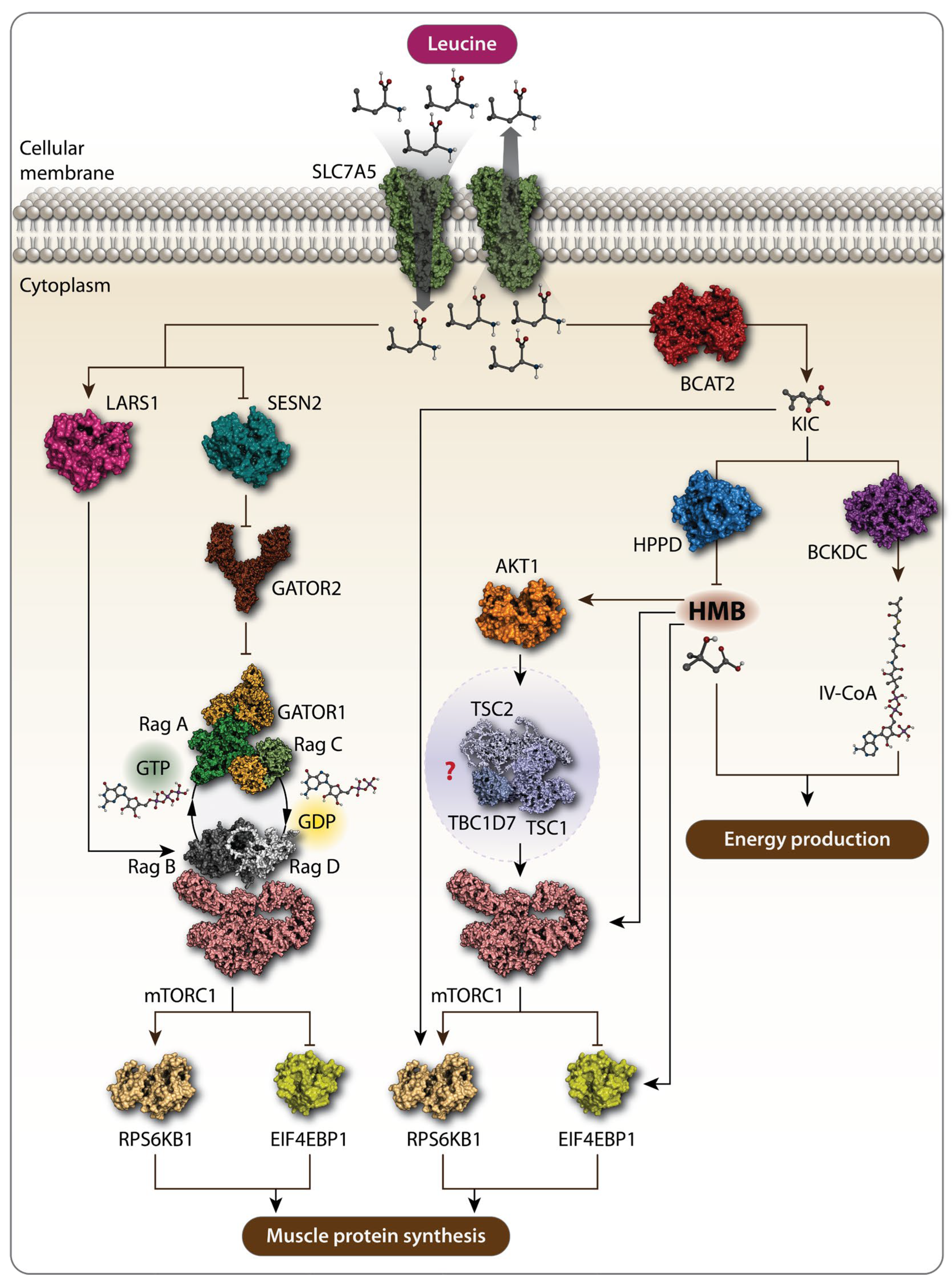
Figure 2.
Leu supplementation leads to tumor cell death by inducing apoptosis and oxidative stress. Leu supplementation can lead to tumor cells death via three mechanisms: (1) Modulation of tumor cell metabolism. Leu supplementation favors oxidative phosphorylation (OXPHOS) over glycolysis by increasing oxygen consumption and mitochondrial biogenesis, while reducing glucose consumption. This metabolic shift promotes apoptosis and oxidative stress in tumor cells; (2) Regulation of the mTORC1 signaling through kinase pathway. Leu supplementation inhibits the expression of insulin-like growth factor 1 (IGF-1) and increases p53 levels. This results in the inhibition of the PI3K/AKT1/mTORC1 signaling axis in tumor cells, leading to apoptosis and tumor cell death. (3) Modulation of the Immune Response. Leu supplementation enhances anti-tumor immunity by inhibiting immunosuppressive regulatory T cells. This effect is mediated by mTORC1 activation, which leads to the inhibition of programmed cell death protein 1 (PD-1). Additionally, Leu competes with kynurenine in the kynurenine pathway (KP), thereby restoring the antitumor effects of T cells and natural killer (NK) cells. Therefore, the inhibitory effect of Leu, in combination with indoleamine 2,3-dioxygenase 1 (IDO1) and tryptophan 2,3-dioxygenase (TDO) inhibitors on kynurenine.
Figure 2.
Leu supplementation leads to tumor cell death by inducing apoptosis and oxidative stress. Leu supplementation can lead to tumor cells death via three mechanisms: (1) Modulation of tumor cell metabolism. Leu supplementation favors oxidative phosphorylation (OXPHOS) over glycolysis by increasing oxygen consumption and mitochondrial biogenesis, while reducing glucose consumption. This metabolic shift promotes apoptosis and oxidative stress in tumor cells; (2) Regulation of the mTORC1 signaling through kinase pathway. Leu supplementation inhibits the expression of insulin-like growth factor 1 (IGF-1) and increases p53 levels. This results in the inhibition of the PI3K/AKT1/mTORC1 signaling axis in tumor cells, leading to apoptosis and tumor cell death. (3) Modulation of the Immune Response. Leu supplementation enhances anti-tumor immunity by inhibiting immunosuppressive regulatory T cells. This effect is mediated by mTORC1 activation, which leads to the inhibition of programmed cell death protein 1 (PD-1). Additionally, Leu competes with kynurenine in the kynurenine pathway (KP), thereby restoring the antitumor effects of T cells and natural killer (NK) cells. Therefore, the inhibitory effect of Leu, in combination with indoleamine 2,3-dioxygenase 1 (IDO1) and tryptophan 2,3-dioxygenase (TDO) inhibitors on kynurenine.
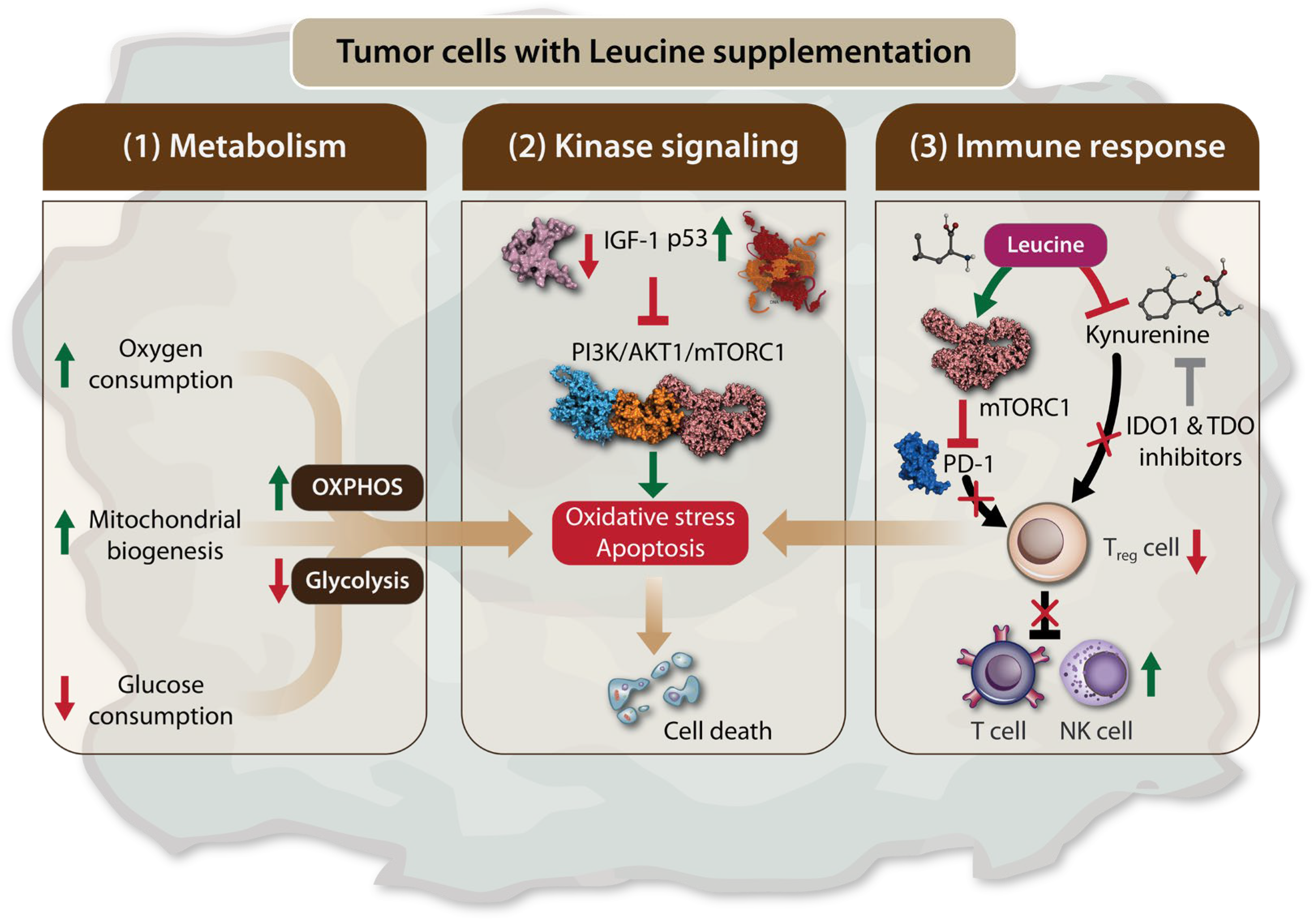
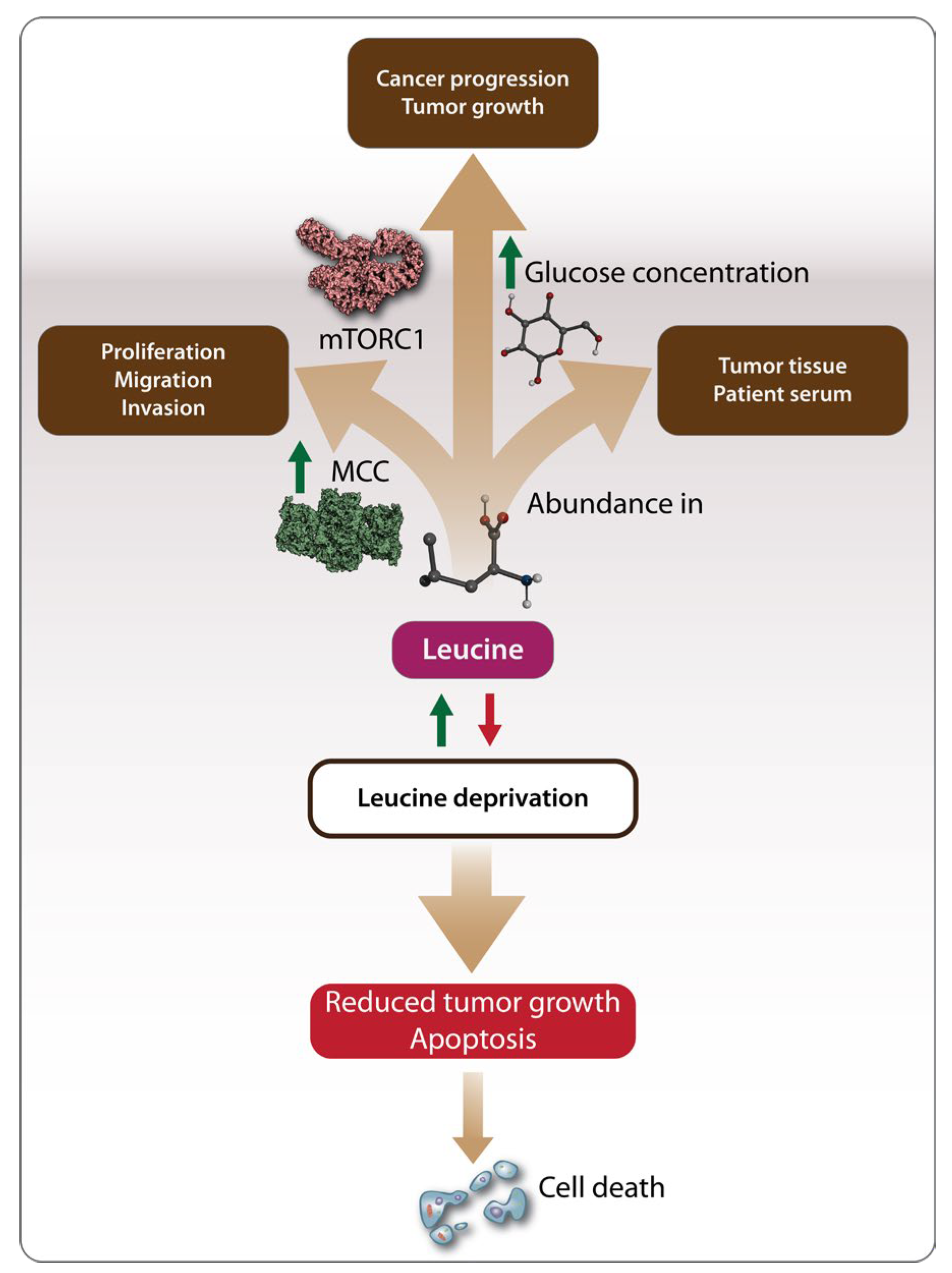
Figure 3.
Mechanisms driving the oncogenic effects of Leu. Methylcrotonoyl-CoA carboxylase (MCC) regulates Leu metabolism promoting tumor cell proliferation, migration, and invasion. Additionally, Leu provides energy to tumor cells by activating the mTORC1 pathway and facilitating glucose uptake, which serves as a necessary energy source for tumor cells. Leu has been found to be abundant in both tumor tissues and the serum of patients with hepatocellular carcinoma (HCC). Consequently, Leu deprivation has been proposed as a potential cancer treatment strategy leading to reduced tumor growth and apoptosis.
Figure 3.
Mechanisms driving the oncogenic effects of Leu. Methylcrotonoyl-CoA carboxylase (MCC) regulates Leu metabolism promoting tumor cell proliferation, migration, and invasion. Additionally, Leu provides energy to tumor cells by activating the mTORC1 pathway and facilitating glucose uptake, which serves as a necessary energy source for tumor cells. Leu has been found to be abundant in both tumor tissues and the serum of patients with hepatocellular carcinoma (HCC). Consequently, Leu deprivation has been proposed as a potential cancer treatment strategy leading to reduced tumor growth and apoptosis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



