
Submitted:
02 September 2024
Posted:
03 September 2024
You are already at the latest version
Abstract
Extracellular vesicles (EVs) are natural carriers of biomolecules that play a crucial role in cell-to-cell communication and tissue homeostasis under normal and pathological conditions, including inflammatory diseases and cancer. Since the discovery of the pro-regenerative and immune-modulating properties of EVs, EV-based therapeutics have entered clinical trials for conditions such as myocardial infarction and autoimmune diseases, among others. Due to their unique advantages—such as superior bioavailability, substantial packaging capacity, and the ability to traverse biological barriers—EVs are regarded as a promising platform for targeted drug delivery. However, achieving sufficient accumulation of therapeutic agents at the target site necessitates a larger quantity of EVs per dose compared to using EVs as standalone drugs. This challenge can be addressed by administering larger doses of EVs, increasing the drug dosage per administration, or enhancing the selective accumulation of EVs at target cells. In this review, we will discuss methods to improve the isolation and purification of EVs, approaches to enhance cargo packaging—including proteins, RNAs, and small-molecule drugs—and technologies for displaying targeting ligands on the surface of EVs to facilitate improved targeting. Ultimately, this guide can be applied to the development of novel classes of EV-based therapeutics and to overcoming existing technological challenges.
Keywords:
targeted drug delivery
; nanotherapeutics
; surface display
1. Introduction
Extracellular vesicles (EVs) are particles constituted by a lipid bilayer that are naturally released from almost all cell types into the extracellular space. Although EVs were initially envisioned as carriers of cellular waste, they are now recognized as key players in cell-to-cell communication [1]. EVs have emerged as a superior delivery system, offering biological compatibility, biodegradability, low toxicity, and immunogenicity [2]. Furthermore, EVs can cross many biological barriers (e.g., blood-brain barrier [3]) and deliver nucleic acids, proteins, small-molecule compounds, and other cargo [4].
Extracellular vesicles (EVs) are considered a more “natural” delivery system, as they are derived from cells and possess specific surface markers such as CD47, which functions as a “don’t eat me” signal that inhibits phagocytosis by macrophages [5,6]. This signaling mechanism leads to enhanced retention of EVs in circulation compared to synthetic nanocarriers like liposomes. Furthermore, EVs produced from mesenchymal stem cells exhibit additional properties, including improved targeting to inflamed tissues and immunoregulatory capabilities [7]. Several EV-based therapeutics have entered phases III-IV of clinical trials and may receive approval for medical applications in the near future [8]. However, the manufacturing of EVs in accordance with good manufacturing practice (GMP) principles is hindered by factors such as batch-to-batch variability and a lack of standardization in EV research [9,10]. Additionally, the isolation and purification processes for EVs are costly, complex, and yield low quantities, making it challenging to scale up EV production. The application of EVs as drug delivery vehicles also necessitates effective strategies for encapsulating various cargo types (such as small-molecule drugs, RNA, proteins, and others) and ensuring targeted delivery to specific tissues through the incorporation of targeting molecules. Some of these issues have been addressed over the past decade, resulting in effective protocols for the cost-efficient production of EVs and the incorporation of desired cargo for delivery, including industrial-scale manufacturing [11]. In this review, we will summarize methods for the production and purification of EVs, as well as techniques for cargo loading. Additionally, we will discuss the primary approaches for the presentation of targeting ligands in EV-based drug delivery systems. Overall, this manuscript aims to serve as a basic guide for developing novel EV-based therapeutics and solving the key fundamental and technological challenges at different stages of drug development.
2. EV Preparation
EVs represent a group of heterogeneous vesicles that are generally divided into three subcategories based on their biogenesis: exosomes (30–120 nm), ectosomes (30 nm–1 µm), and apoptotic bodies (100 nm–5 µm) [12]. At the same type, classification of EVs is evolving, and many distinct types of EVs were characterized. Current landscape of EVs includes small high-density lipoprotein (HLD), low density lipoprotein (LDL), supermeres, intermediate-density lipoprotein (IDL), exomeres, VLDL and vaults with a size of 5-72 nm, exosomes, arrestin domain-containing protein 1-mediated microvesicle (ARMM), small ectosomes and supramolecular attack particle (SMAP) which size ranges from 30 to 150 nm. Larger EVs with a size from 70 nm to several µm include chylomicron remnants, chylomicrons, microvesicles, apoptotic bodies or vesicles, migrasomes, large oncosomes and exophers. The types have distinct sizes, biochemical composition, and morphology [12]. Exosomes are formed by the inward budding of multivesicular bodies that later fuse with the cell surface, resulting in vesicle release. Ectosomes bud directly from the plasma membrane, and apoptotic EVs form during apoptosis by cell fragmentation. Recently, more subtypes have been discovered, such as migrasomes [13] and exophers [14]; their capacity to specifically deliver molecules for therapeutic purposes is yet to be discovered (Figure 1). The EVs mentioned below mostly represent particles <1000 nm in diameter, like exosomes and the classic type of ectosomes a.k.a microvesicles.
EVs are naturally released from recipient cells into the extracellular space. However, after production, EVs are contaminated with cells, host cell DNA, host proteins, and cell culture media supplements (e.g., fetal bovine serum [FBS]), and thus must be isolated and purified from cell culture media for further use. As EVs are usually manufactured using adherent cells (e.g., stem cells), they can be easily isolated and purified in one step by processing the conditioned media [15].
3. Purification Methods
3.1. Size-Based Approaches
Plenty of protocols exist for EV isolation, including several combined methods [16,17]. The gold standard for EV separation is differential ultracentrifugation (dUC, or UC) that combines serial centrifugation steps (Figure 2A). A standard protocol comprises centrifugation at low speed (300 × g) to remove cells; at medium speed (2000 × g) to remove larger cell debris; centrifugation at high speed (10000 × g) or filtration through a 0.22 μm filter to remove large non-exosomal EVs; and two long, high-speed centrifugation (100 000 × g; 60–70 min) steps to pellet and wash EVs [18]. dUC allows high concentration of EVs, but its application for industrial-scale EV manufacturing is complicated [19]. Currently, dUC is the best-characterized method that is used in roughly half the studies related to EVs [20]. However, separation of EVs with dUC has many disadvantages, including vesicle aggregation [21], disruption [22] and loss [23], contamination with free proteins and their aggregates [24], and low isolation yield (~25–30%) [25]. Moreover, dUC is time-consuming and has high equipment requirements. EV pelleting efficiency is dependent on parameters such as sample viscosity and centrifuge acceleration, rotor type and characteristics [19]. Intra- and interlaboratory variability, as well as poor scalability, make following GMP principles difficult with this method [26].
Density gradient ultracentrifugation (DGU) is a variation of dUC that uses a medium (usually a solution of sucrose or iodixanol [27]) of gradated density (Figure 2B). During the centrifugation process, EVs migrate to their equilibrium fractions according to their size, shape, and density. DGU offers the highest EV purity with almost no protein contamination [28], as well as reduces vesicle damage and aggregation [11], but the method is tedious, expensive, low yield, and requires post-purification from gradient media, thus often resulting in considerable EV loss [19].
During ultrafiltration (UF), a sample is passed through a semi-permeable membrane, so small contaminants (below membrane pore size cut-off) are removed into the filtrate whereas the desired particles are collected in the retentate (Figure 3A). To remove proteins and small molecules, cut-offs of ~300–750 kDa (~30–75 nm) are a common choice, whereas filters with a higher molecular weight cut-off (i.e., 1000 kDa) can be useful to remove some smaller EVs [29]. UF is preferred by many researchers as it is fast, simple, and does not require expensive equipment [30]. UF is considered time-saving and cost-effective, it allows for concentrating EVs, offering a greater isolation yield than dUC [31]. Also, UF method is established in the industry for protein production and is therefore easily scalable [32]. However, larger particles that cannot migrate through the membrane can accumulate on the filter upstream surface and block filtration of smaller molecules, resulting in a “cake” formation effect that decreases sample purity [11,33]. Other disadvantages of UF are possible EV deformation [19], non-specific adsorption of EVs on the filter surface, and filter pore clogging [34].
To reduce the “cake” formation effect, asymmetric depth filtration (ADF) has been proposed for EV isolation [35]. Depth filters use a porous filtration medium with larger diameter pores to retain particles throughout the medium rather than on the filter’s surface. The filter pores have a tortuous geometry, which allows them to capture larger particles (Figure 3B). Thanks to this setting, soluble components and small molecules are eluted first, while larger components elute later or get trapped within the filter. DF is a simple, fast, and inexpensive method that provides less filter clogging since the entire filter medium participates in fractionation rather than just its surface.
Tangential flow filtration (TFF), also known as cross-flow filtration, is a more sophisticated version of UF that is based on the same principle. However, in the TFF system, the stream flows tangentially across the UF membrane and a second flow is directed through the membrane (Figure 3C). Smaller particles are filtered through the membrane, and larger particles are flushed from the filter by tangential flow. TFF is a cyclic process, and filtration can be repeated several times to increase sample purity or perform buffer exchange. TFF is time-saving, reduces material deposition, provides high EV yields, and allows for vesicle concentration [36,37,38]. TFF can be used to isolate EVs from large volumes (thousands of liters) and is established as a scalable industrial approach [37]. However, TFF can result in vesicle contamination with large proteins and DNA [15,39], and therefore often requires a second process (e.g., size exclusion chromatography) to obtain optimal EV purity [38].
Size exclusion chromatography (SEC) is another prominent method that separates particles by size as they pass through a column filled with porous beads [40]. Components larger than the pore size are unable to enter the pores of the column and thus elute first, while smaller molecules or particles become trapped inside beads and elute later. This method efficiently eliminates smaller molecules, proteins, and small particles in combination with mild processing conditions that preserve vesicle structure [41]. SEC provides high purity of isolated EVs, but it is a laborious process that may require specialized equipment [42,43]. Although SEC may be used for scaled preparations, it requires a pre-concentration step for processing of large amounts of EV-enriched media [44] and has relatively low yield [45].
3.2. Precipitation-Based Methods
Precipitation-based methods (Figure 4A) commonly utilize hydrophilic (e.g., polyethylene glycol [PEG]) or cationic polymers (e.g., protamine) to induce reversible EV aggregation, allowing their separation by centrifugal forces or filtration [44]. Many commercial isolation kits are based on precipitation (ExoQuick, Exo-Prep, Total Exosome Isolation Kit) due to its simplicity and low EV damage [46]. Precipitation-based methods do not require costly or specialized equipment and can be applied to large sample volumes required to obtain preparative EV amounts [47]. However, precipitation-based methods are costly, result in high amounts of contaminants, including proteins, that co-precipitate with the vesicles, reducing sample purity. Polymer residues must also be removed, which can lead to EV loss [15]. These disadvantages make precipitation-based methods unsuitable for use in clinical therapy without additional purification steps [48].
3.3. Affinity-Based Approaches
EVs can be isolated via immunoaffinity methods based on EV-specific membrane protein molecules, such as CD9, CD63, and CD81. [49]. In immunoaffinity-based isolation, antibodies are usually attached to magnetic beads to form antibody-conjugated magnetic beads that can pull EV populations from crude material (Figure 4B) [15]. Immunoaffinity methods offer high purity [50] and reduce contamination with soluble proteins [33]. However, EV markers are not present on every EV subtype [51]; for instance, only a small number of EVs are positive for both CD63 and CD81 [52]. Moreover, the interaction between the antibody and the EV marker is not easily disrupted. Harsh elution conditions may destroy EV structure [53].
Recently, a protocol named EV-Elute has been proposed, which allows recovery of over 70% of bound EVs from antibody-conjugated magnetic beads without compromising EV integrity [54]. This is achieved by optimizing components of the elution buffer and conditions used for the elution process. Other disadvantages of immunoaffinity methods include high antibody costs, relatively low EV yield [33], and method difficulties for isolation from large quantities [55]. On the other hand, aptamers can be used instead of antibodies to lower the cost [56]; aptamer-marker interactions are also easily reversible in mild conditions that allow to particle isolation that preserves their integrity.
3.4. Chromatography Approaches
To obtain high-purity EVs, multimodal flowthrough chromatography (MFC; also known as bind-elute chromatography) has been introduced [57]. MFC combines size-exclusion and bind-elute chromatography. MFC resins have an inert shell permeated with size-selective pores that surround an absorptive core. When small molecules and proteins enter the pores, they bind to the absorptive core via hydrophobic and charge interactions. Other molecules larger than the size of the selective pores, including EVs, are excluded from the shell and can be collected in the flowthrough. MFC allows isolating EVs with higher purity (comparable to 2D culture and SEC) even from highly contaminated bioreactor preparations and with negligible sample dilution compared to SEC [57].
Anion-exchange chromatography (AIEX) is another chromatography-based method that is applied for EV isolation. EVs can be separated by binding to the positively charged column due to negatively charged phospholipids found in EV membranes [58]. AIEX can be scaled to an industrial level and shows higher sample yield, purity, and size distribution than dUC [59]. Despite its advantages, AIEX can provide only limited specificity due to co-isolation of different EV subtypes and sample contamination with other negatively charged molecules (nucleic acids, proteins, and other) [60,61], in addition to potential damage to the vesicles due to harsh conditions (e.g., acidic buffers) [41].
A combination of methods is also used to isolate highly purified EVs: indeed, ~60% of current studies combine several methods for EV isolation [62]. Combined EV isolation assures better purity and specificity than single isolation methods [32,63], and the shortcomings of one approach can be addressed by another method. At the moment, the most optimal methods are SEC in combination with either TFF or UF. As aforementioned, both TFF and UF are able to concentrate the samples but yield less pure EV fractions, whereas SEC can result in relatively high EV purity but dilutes the sample. By combining SEC with TFF or UF, highly concentrated EV fractions with even higher purity can be obtained [64]. Despite the advantages, every additional purification step increases processing time and reduces overall EV yield [65].

Table 1.
Comparison of different EV isolation approaches.
| Method | Principle | Scalability | Yield | EV Damage | Purity | Equipment requirement | Cost | Additional Pre/Post-steps | Time | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|
| Differential ultracentrifugation (dUC) | Serial UC steps | + | + | ↑↑↑ | ++ | +++ | + | No | ↑↑↑ | [19,21,22,23,24,25,26] |
| Density gradient ultracentrifugation (DGU) | Separation of EVs by density using gradient medium | + | + | ↑ | ++++ | +++ | ++ | Yes (media removal) | ↑↑↑ | [11,19,28] |
| Ultrafiltration (UF) | Filtration through semi-permeable membranes | +++ | +++ | ↑↑ | +++ | ++ | ++ | No | ↑/↑↑ | [11,19,30,31,32,33,34] |
| Asymmetric depth filtration (DF) | Filtration through porous medium | ++ | ++ | ↑ | +++ | ++ | ++ | No | ↑↑ | [35] |
| Tangential flow filtration (TFF) | Cross-flow filtration through membranes | ++++ | +++ | ↑ | +++ | ++ | ++ | No | ↑↑ | [11,15,36,37,38,39] |
| Precipitation approaches | EV sedimentation using polymers | +++ | ++++ | ↑↑/↑↑↑ | + | + | ++ | Yes (polymer removal) | ↑↑↑ | [11,15,46,47,48] |
| Affinity-based isolation | EV capture via specific interactions with EV markers | +/++ | ++ | ↑/↑↑ | ++++ | ++ | +++ | No | ↑↑ | [33,50,51,52,53,55] |
| Size exclusion chromatography (SEC) | Separation by size through a bead-filled column | +++ (combined with UF/TFF) | ++ | ↑ | +++ | ++ | + | Yes (Pre-Concentration) | ↑↑ | [41,42,43,44,45] |
| Multimodal flowthrough chromatography (MFC) | Combination of size-exclusion and bind-elute chromatography | +++ (combined with pre-concentration) | ++ | ↑ | ++++ | ++ | ++ | Yes (Pre-Concentration) | ↑↑ | [57] |
| Anion-exchange chromatography (AIEX) | Binding of EVs to positively charged column | +++ | +++ | ↑↑ | ++ | ++ | ++ | Yes (Buffer exchange) | ↑↑↑ | [41,59,60,61] |
3.5. Challenges of EV Preparation
Low EV yield is a major problem for EV research. Poor EV yields occur for two reasons: 1) small amounts of EVs produced by the cells and 2) EV loss during isolation. One liter of conditioned culture media yields approximately 109–1011 EVs using dUC, which is usually enough for only one experiment in one mouse in preclinical studies [66]. The expected number of EVs needed for therapeutic applications is ~1013 EVs per dose per patient [44]. There are many methods to increase EV production by cells, such as mechanical stimulation, serum deprivation, hypoxia, and supplementation with small-molecule drugs and additives [11], but these approaches can be associated with cell damage and reduced EV quality. At the same time, using dynamic and static 3-dimensional cell cultures can substantially boost EV production in a scalable manner [67]. However, low yields after isolation are still an issue.
These issues have led researchers to explore alternative EV sources, such as EVs derived from bovine milk or plants. While these sources allow robust production of EVs with high yields, they cannot substitute human EVs for all applications and can cause adverse and unpredictable effects [64]. Non-human EVs can be immunogenic or allergenic depending on the administration route, dosage, and number/frequency of doses [68]. Safety of these materials must be validated for each individual case.
There are currently no methods for simultaneously achieving high EV yield and high purity. Highly purifying techniques always lower EV yields [69]. Sufficient separation from co-purifying components is a challenge because EVs are contaminated with host nucleic acids, and proteins and their aggregates. Carbohydrates (e.g., hyaluronic acid) can be overlooked contaminants as well [70]. Another possible contaminant is FBS, a common supplement for cell culturing, which is high in endogenous EVs [71]. However, serum-free conditions can cause stress-induced phenotypic changes in cultured cells, promoting release of EVs containing reactive oxygen species (ROS) and stress-related proteins [72]. Human platelet lysate is often used as a substitute for FBS, as it is a safe and effective supplement that creates a xeno-free environment suitable for culturing various cell types while reducing the immunologic risks associated with FBS [73]. On the other hand, the variability of platelet concentrations and the presence of additional bioactive factors in human platelet lysate may affect cell behavior and EV characteristics, compromising the reproducibility and standardization of prepared EVs. Nevertheless, some contaminants can supply EVs with different traits; for example, hyaluronic acid has immunomodulatory properties that may be associated with successful EV applications in cancer therapy and immunotherapy [74]. Viral contamination must also be monitored, especially for clinical treatment, because viral particles and EVs are of similar size [15].
Separating specific EV subpopulations from cell culture for further analysis is still a problem. The heterogeneity of EVs poses a challenge to their application as therapeutic carriers in clinical treatments. For instance, in one study, three EV subpopulations with distinct sizes, protein marker content, morphology, and different functional properties were separated using SEC, demonstrating similar results across different cell types [75].
Additionally, appropriate conditions for storing, handling, and transporting EVs must be determined. One comprehensive study showed that freezing EVs in phosphate-buffered saline (PBS) greatly reduced EV recovery, and proposed PBS supplemented with human serum albumin and trehalose as the optimal EV storage method [76]. Other recent studies have utilized EV lyophilization to facilitate handling and transportation and to prolong the shelf-life of the final product [77,78]. However, further exploration of effects of lyophilization on the integrity of EV membranes is necessary [79]. Finally, different storage conditions may be optimal for EVs from different cell sources or for EV subpopulations [80].
Scalable manufacturing and batch-to-batch variability of EVs are other crucial issues facing clinical therapy. To address these, isolation methods suitable for large-scale applications, such as TFF, SEC, or MFC, can be applied. Bioreactors can be used to produce therapeutically relevant concentrations of EVs while maintaining consistent particle size and phenotype [81,82].
Another problem is the biomolecular (protein) corona and effects of different isolation methods on its composition. The surface of most of nanoparticles (NPs) is well known to play a pivotal role in their behavior and functionality. The biomolecular corona can be characterized as spontaneous adsorption of proteins, lipids, sugar moieties, nucleic acids, and metabolites onto NP surfaces [83] due to their interaction with NP surface molecules in biological fluids. Notably, the corona is formed rapidly (<30 seconds) [84]. After corona formation, NPs’ physicochemical properties, targeting abilities, and biological responses are dictated by the adsorbed molecules [83,85], posing difficulties for predicting how the biomolecular corona may affect NP activity in vivo due to its dynamic nature. Coronas form on the surface of different kinds of NPs, including lipid-based NPs [86,87], polymeric NPs [88], and inorganic NPs [89]. Recently, biomolecular coronas were shown to be generated around EVs as well [90]. Additionally, it was demonstrated that the isolation method can deform the biomolecular corona: UC and TFF may change its composition and abrogate EV function [91].
4. EV Cargo Loading Methods
Application of EVs as delivery platform requires efficient methods for packaging therapeutic cargo into the vesicles. Payload loading can be endogenous (based on the natural ability of cells to form EVs) or exogenous (loading molecules into EVs after the EVs have been isolated). Whether endogenous or exogenous loading methods are used, the cargo can be loaded either passively or actively.
During passive loading, cargo is simply incubated with donor cells or directly with EVs. This technique can be used to load small molecules, particularly hydrophobic small-molecule drugs. However, the incubation method is rarely used due to its low loading efficiency [92] and poor reproducibility [93]. Other methods are categorized as active loading techniques, and include sonication, electroporation, freeze-thawing, transfection, and extrusion.
4.1. Endogenous Loading
Endogenous loading, or pre-loading, is an approach based on the ability of donor cells to incorporate molecules into EVs during EV biogenesis (Figure 5). Several methods of endogenous loading are used for RNA, protein and small molecules packaging, including simple incubation, application of EV-associated motifs, interaction of target molecules with plasma membrane of membrane proteins, viral protein-assisted loading and others. The main advantage of endogenous loading is the preservation of EV membrane integrity [94].
4.1.1. Cells Transfection/Transduction for RNA Loading
This approach often utilizes transfecting or transducing producer cells with vectors encoding therapeutic molecules (or with therapeutic molecules directly, for example, with siRNA), but cells can also be simply co-incubated with putative cargo for this purpose. One drawback of using transfection for EV loading is the potential alteration of the structure and properties of EVs by the transfection reagents [95]. Another concern is cytotoxicity and difficulty to completely remove transfection reagent [96,97]. Furthermore, transfection reagents can form lipid micelles with loaded molecules that are similar in size to and imitate cargo-loaded EVs, leading to possible sample contamination [98]. Additionally, transfection reagents can affect EV cargo delivery to recipient cells [99,100]. Although endogenous loading appears to be less effective in terms of loading small molecules, it seems to be more suitable for loading larger molecules [101].
4.1.2. EV-Associated Motifs for RNA Loading
EV-associated motifs (EXOmotifs or “zipcodes”) are special motifs usually found in 3′-untranslated regions (3′-UTRs) of RNA. They interact with EV biogenesis machinery and help with RNA loading into EVs. EXOmotifs either recruit RNA binding proteins (RBPs) that mediate the packaging of therapeutic RNA into EVs, or possibly modulate direct RNA interaction with lipid membranes during EV biogenesis [102]. EV-associated motifs have been found in both mRNAs [103] and microRNAs (miRNAs) [104]. hnRNPA2B1 ribonucleoprotein (RNP) can directly interact with GGAG motifs and control miRNA sorting [105], whereas SYNCRIP protein is able to bind to GGCU motifs of miRNA [106]. EXOmotifs can direct RNA of interest to be packaged inside EVs. For instance, miRNA targeting the ATXN3 gene was engineered using ExoMotif GGAG, resulting in its 3-fold enrichment in EVs [107]. However, the loading process tends to be tissue-specific and may be influenced by secondary and tertiary RNA structures [102,107].
The Psi (Ψ+) signal is an alternative to EV-associated motifs. It can be found in retrovirus genomes (e.g., of human immunodeficiency virus [HIV]) and act as a packaging signal for the retroviral RNA genome [108]. This signal specifically interacts with Gag protein and allows the transport of RNA into virus-like EV-mimetic particles (VLPs) that are structurally similar to EVs. Inserting a Ψ+ signal into the RNA sequence can allow RNA loading into VLPs. One study used the nanomembrane-derived extracellular vesicles for the delivery of macromolecular cargo (NanoMEDIC) system for single guide RNA (sgRNA) packaging by incorporating Ψ+, along with the Tat activation response element (TAR) to increase expression of CRISPR-Cas sgRNA [109]. To liberate sgRNA from the long mRNA transcript, self-cleaving ribozymes from hammerhead and hepatitis delta virus flanked the sgRNA sequence. The Ψ+-mediated packaging loaded sgRNA 3 times more efficiently than stochastic loading. The main drawback of this system is the use of Gag and Tat proteins, which are associated with oxidative stress and can induce cellular toxicity and immunogenicity when used in vivo [110].
4.1.3. Interaction of RNA with Proteins Enriched within EV Membranes
Another strategy for packaging RNA into EVs is through indirect interactions of the desired molecules with EV membrane proteins. For this purpose, RNA-binding proteins (RBPs) can be fused to proteins that are abundant in EVs, like tetraspanins CD9, CD63, and CD81. Then, RBPs can specifically bind to sequence and/or structural motifs of RNA via one or several RNA-binding domains (RBDs), thereby packaging RNA into EVs [111]. For instance, CD9 was fused with human antigen R (HuR) RBP that mediates interaction between AU-rich elements (AREs) in the target RNA and HuR [112]. Loaded RNA was localized inside EVs, increasing loading of Cas9 mRNA and miR-155 by up to 10-fold and 7-fold, respectively.
Several studies have utilized CD63-mediated packaging. The targeted and modular EV loading (TAMEL) approach was proposed as a platform for RNA loading [113]. Here, RNA is engineered with MS2 stem loops that can specifically interact with MS2 bacteriophage coat protein (MCP) fused to CD63 protein. This technique improves RNA packaging into EVs up to 6-fold, but low endosomal escape of loaded RNAs was observed. An alternative approach uses a specific Com/com interaction between the inserted RNA aptamer com and Com protein fused to CD63 [114]. By additionally overexpressing vesicular stomatitis virus protein G (VSV-G) in the donor cells, EVs were loaded with CRISPR-Cas RNP complexes and demonstrated efficient endosomal escape and gene editing, up to 10-fold higher than EVs without an RNP enriching mechanism. Viral VSV-G protein increase EVs secretion and provide broad tissue tropism of particles, as many types of cells express receptor for VSV-G. The main disadvantage of packaging using 3′-UTR-mediated mRNA modifications is activation of nonsense-mediated RNA decay (NMD) that causes destabilization of mRNA and its degradation in both packaging cells and acceptor cells [115,116].
To overcome the obstacles of low mRNA stability and insufficient loading, a novel TAMEL-based platform has been developed [117]. The authors used an engineered RBD derived from Pumilio and FBF homology domain (PUFe) fused to CD63, while the target RNA was engineered with 3′-UTR repeats to specifically interact with the RBD. CD63-PUFe showed 1.7-fold to 4.3-fold better loading efficiency than CD63-MCP in three independent experiments, suggesting that RBD choice and design influence the efficiency of RNA loading. Additionally, co-expressed VSV-G and cytoplasmic poly(A)-binding protein (PABPc) improve EV cargo delivery and ensure better stability of mRNA, respectively. PABPc binds to the poly(A)-tail of mRNA and protects it from degradation through the NMD pathway. Indeed, addition of PABPc improved mRNA enrichment in EVs: the PABPc/mock ratio varied from 3- to 14-fold [117]. Nevertheless, the absolute number of loaded RNA per EV may still be insufficient to provide clinically relevant effects.
Another RNA-RBP interaction was used as the basis for a two-component system called exosomal transfer into cells (EXOtic). CD63 was fused with L7Ae archaeal ribosomal protein that can specifically interact with a cognate C/D box of the engineered cargo RNA [118]. Connexin 43 protein, a cytosolic delivery helper enriched in EVs, or the more effective mutant S368A protein, was utilized to facilitate the fusion of EVs with the endosomal membrane or the formation of channels that allow the escape of EV content from endosomal compartment. By using the EXOtic system, catalase mRNA was successfully delivered into cells, reducing the production of ROS and cell death induced by neurotoxin in vivo.
Moreover, another approach can be applied for packaging of sgRNA, which is a component of a genome editing system called CRISPR-Cas. sgRNA can be packaged passively due to the formation of sgRNA-Cas9 complexes. For instance, coupled fusion of GFP with CD63 and fusion of GFP-specific nanobodies with Cas9 leads to the formation of RNP complexes and their subsequent loading into EVs, but with unsatisfactory efficiency [119]. Notably, Cas9 mRNA is undetectable in EVs when mRNA is used instead of sgRNA, suggesting that sgRNA packaging into EVs is a result of its co-recruitment as a passenger in a complex with Cas9 [120].
4.1.4. RNA Enrichment on the Plasma Membrane
An alternative method for loading endogenous RNA uses a light-inducible dimerization system based on CRY2-CIB1 and MS2–MCP interaction modules [121]. CIBN (a truncated version of CIB1) protein is fused to a palmitoylation sequence (palm) that induces protein enrichment in the cell membrane, while CRY2 is conjugated to MCP protein. Application of blue light induces reversible binding of membrane-enriched CIBN with CRY2-MCP and subsequent accumulation of MS2-RNA on the plasma membrane via MCP-MS2 interaction. The amount of thus loaded miRNA is 14-fold higher than in EVs obtained from cells cultured without light illumination. However, the system relies on RNA loading during EV biogenesis [121], and blue light illumination may affect cell viability through ROS generation [122].
4.2. Endogenous Protein Loading
A number of papers demonstrate CRISPR-Cas9 delivery approaches, and thus, in certain protocols, protein loading can accompany RNA loading. CRISPR-Cas possesses great potential in therapy of cancer, hereditary and infectious diseases [123,124,125,126,127], and EVs are considered as an important carrier for CRISPR-Cas delivery. To preserve the functional activity of the packaged protein, it must be released from the vesicle membrane into recipient cells. Here, we focus on some examples of protein loading that have not been described in the previous section. Along with transfection of donor cells, a plethora of methods has been developed to endogenously load cargo proteins into EVs (Figure 6). These methods can be divided into three distinct strategies:
- 1)
- Fusing cargo to proteins enriched on EV membranes;
- 2)
- Using post-translational modifications of the cargo proteins;
- 3)
- Viral protein-assisted loading.
4.2.1. Fusion with Proteins Enriched within EV Membranes
Similar to RNA packaging, protein of interest can be fused to proteins that are enriched in EVs. For this purpose, a light-inducible loading system called exosomes for protein loading via optically reversible protein-protein interactions (EXPLOR) has been proposed [128]. Two recombinant proteins are overexpressed in this system: CIBN is conjugated to the EV-associated CD9, while the cargo protein is fused to CRY2. Similarly to the RNA loading method discussed above, blue light illumination induced dimerization of CRY2-CIBN, with subsequent enrichment of the protein of interest in EVs. In the absence of blue light, engineered cargo detaches and releases into EV intraluminal space. This method demonstrated efficient delivery of Bax protein, super-repressor IκB protein, and Cre enzyme. However, the EXPLOR system shows limited loading efficiency with an average of 1–2 cargo molecules per particle, and although blue light did not cause cell toxicity in the study, it could still potentially be toxic to cells.
Another method is based on the use of CD63 for packaging and subsequent cleavage of the cargo into a soluble active form, releasing it into the EV lumen. A recent study employed engineered self-cleaving pH-sensitive intein to develop systems for cytosolic protein delivery, termed VSV-G plus EV-sorting Domain-Intein-Cargo (VEDIC) and VSV-G-Foldon-Intein-Cargo (VFIC) [129]. Treating reporter cells with collected particles derived from CD63-Intein-Cre and VSV-G-expressing cells leads to significant gene editing activity. To further improve the VEDIC system, the VFIC system expressed through a single vector was developed. Also, VSV-G itself has been shown to act as an efficient EV sorting domain without requiring the co-expression of an EV sorting protein such as CD63. Furthermore, to improve VSV-G trimer formation and function, T4 fibritin trimerization motif (foldon) was added to the construct. The efficiency of cargo delivery using the VFIC system turned out to be significantly higher (about 2–3 times) than using the VEDIC system.
Gesicles are particles generated with the help of viral proteins (VSV-G). Overexpressing VSV-G induces generation of many VLPs with the encapsulated cargo of interest [130]. Due to the high prevalence of low-density lipoprotein (LDL) receptors, which serve as cellular receptors for vesicular stomatitis virus, gesicles have broad tropism [131]. This method was subsequently used as the basis for a trigger-dependent system based on the interaction of FK506-binding protein (FKBP12) and FKBP12-rapamycin-binding protein (FRB) [132]. In this system, gesicles are produced for CRISPR/Cas9 RNP delivery using a system composed of 4 components: membrane-associated protein produced by the interaction between two domains (FKBP12 and FRB), VSV-G, Cas9, and the chosen gRNA. In the presence of rapamycin (A/C heterodimerizer), the two domains of the protein are brought together to form a complete protein that is enriched in EV membranes. Gesicle-mediated CRISPR/Cas9 RNP delivery targeting the HIV long terminal repeat (LTR) reduced copy numbers of HIV provirus and the viral protein Nef. However, VSV-G may have immunogenic properties that need to be addressed for clinical applications [133].
Another approach can be used with the specific EV population called arrestin domain-containing protein 1- (ARRDC1) mediated microvesicles (ARMMs) [134]. ARRDC1 plays a crucial role in ARMM biogenesis at the plasma membrane. Its overexpression triggers outward membrane budding, leading to a rise in ARMM production [80]. Proteins of interest can be directly fused with ARRDC1 and incorporated into ARMMs. To deliver additionally deliver mRNA, some authors utilized the viral TAT-TAR system [134], in which ARRDC1 is fused with TAT protein; after transfection, ARMMs are collected. RNA cargo is loaded into EVs due to the specific interaction between TAR RNA loop and TAT protein. In this study, ARMMs efficiently delivered functional p53 and p53 mRNA, which was translated into functional protein. However, for packaging of larger proteins (Cas9), researchers decided to use the interaction of ARRDC1 with WW-domain-containing proteins instead of direct protein cargo fusion with ARRDC1. ARRDC1 acts as an adapter recruiting ubiquitin-protein ligases to proteins with WW-domain-containing proteins. Because of this, WW-Cas9, along with its associated sgRNA, is successfully incorporated into ARMMs [134].
In addition, some other EV-sorting proteins, like tetraspanin 14 (TSPAN14) [135], TSPAN2, and TSPAN3 [136] can be employed to load proteins into EVs. For instance, in one study, TSPAN2 and TSPAN3 proteins outperformed the well-studied CD63 scaffold in EV-sorting ability and were utilized for robust luminal loading [136]. Furthermore, brain acid soluble protein 1 (BASP1) can be used to incorporate various proteins into the lumen of EVs, since BASP1 can bind to the inner leaflet of the plasma membrane and promote cargo sorting into EVs [137].
4.2.2. Post-Translational Modifications of the Protein
Post-translational modifications (PTM) can change the properties of a newly translated protein by proteolytic cleavage or addition of functional groups. PTM can guide proteins to be packaged inside EVs by distinct mechanisms. For instance, polyubiquitinated cargoes activate the ESCRT-dependent sorting mechanism [138], whereas myristoylated cargoes can associate with the cell membrane, which increases their encapsulation into EVs [139].
Ubiquitination [140], myristoylation [139], and palmitoylation [109] are commonly used tags for favoring the loading of proteins into EVs. Furthermore, as mentioned, N-glycosylation sites and WW tags can act as PTM sites. All these PTM promote cargo protein incorporation, while some PTM can also contribute to EV uptake by recipient cells and signal transduction [141]. PTM of a protein of interest can promote its relatively efficient loading into EVs. For example, myristoylation-palmitoylation-palmitoylation motif (MysPalm) was demonstrated to be the best method for protein loading, allowing the highest amount of Cas9 protein per EV, more effective than fusing cargo protein with CD9, Rab5c, or CD81 [120].
Another study implemented a similar approach (TOP-EVs) to the above-mentioned four-component gesicle system, but added myristoylation tags to the protein [142]. Rapamycin-inducible heterodimers T82L mutant FRB (DmrC) with FKBP12 (DmrA) were used for drug-inducible dimerization. DmrC was fused to the target protein, whereas DmrA contained an N-myristoylation sequence for enrichment of DmrA and, thus, the final protein, in the cellular membrane. VSV-G was also co-transfected along with a protein of interest as an endosomal escape booster. This study shows successful loading of GFP into TOP-EVs in the presence of rapamycin, with more than 60% of TOP-EVs containing GFP and an average of over 90 GFP molecules per vesicle. Furthermore, efficient intraluminal loading of Cre recombinase and CRISPR/Cas9 RNP into TOP-EVs and their delivery to the cells are demonstrated.
Another possible protein-loading mechanism relies on the use of protein motifs. A recent study showed that lysosome-associated membrane protein 2, isoform A (Lamp2A) can participate in the loading of cytosolic proteins into a subpopulation of exosomes [143]. It is a novel mechanism independent of the ESCRT machinery. Proteins with amino acid sequences biochemically related to the KFERQ motif can be loaded into a specific exosome subpopulation. When a KFERQ-like motif is added to the sequence of mCherry fluorescent protein, an approximately 7-fold increase in the presence of mCherry in sEVs is observed. However, it is unclear whether the addition of a KFERQ-like motif results in protein incorporation by a distinct mechanism or if these motifs also act as potential sites for PTM. Further studies are needed to fully elucidate the loading mechanism of proteins with KFERQ-like motifs and further applicability of this method for protein packaging into EVs.
Problematically, PTM are most likely to be cell-specific, complicating their application in protein loading. As another drawback, PTM can be temporary and, thus, lead to protein loss before EV sorting. For instance, ubiquitinated proteins can be deubiquitinated en route to EVs [141]. Moreover, some EV isolation methods, like dUC, can alter PTM and thus alter functionality of the cargo proteins.
4.2.3. Viral Protein-Assisted Loading
Several previously mentioned systems have used VSV-G to enhance endosomal escape and cargo delivery. In this section, we will focus on some examples of viral protein-assisted loading.
Nanoblades are biological EV-mimetic nanoparticles that were originally developed from murine leukemia VLPs for the delivery of Cas9-sgRNA RNPs [144]. The approach utilizes the ability of retroviral Gag proteins to form artificial VLPs in the presence of fusogenic VSV-G and deliver them into the cytoplasm by fusion with target cells. Gag proteins (either from murine leukemia virus [MLV]) or HIV are fused with the protein of interest, with a proteolytic cleavage site designed between the desired protein and Gag. Co-expression of wild-type Gag-Pol protein is required to release the protein of interest from Gag into the intraluminal space via protease-mediated cleavage. Treatment of cells with isolated nanoparticles (Nanoblades) containing incorporated CRISPR-Cas9 system demonstrate more efficient and rapid induction of double-stranded DNA breaks than CRISPR-Cas loaded via transfection or electroporation. However, this method requires co-expression of Gag-Pol protein, which competes for space with the protein of interest, resulting in less protein loaded per VLP [109]. Moreover, immunogenicity of viral proteins presents a challenge for clinical use [145]. Another disadvantage is that HIV protease produced by self-cleavage of the Gag-Pol polyprotein can mediate the degradation of packaged cargo proteins.
VSV-G protein can be exchanged to another viral protein for nanoblades preparation. One group used baboon retroviral envelope glycoprotein (BaEV) to transduce hematopoietic stem cells (HSCs) using nanoblades due to the lack of LDL receptor on HSCs [144]. The obtained VLPs result in 50% genome editing when CRISPR-Cas9 system is packaged. This makes nanoblades complicated to use, however, because cells must be transfected with plasmids that encode Gag fused with the required protein, Gag-Pol protein, VSV-G, and BaEV.
Since the discovery of nanoblades, a few studies have been conducted using this method. One study confirmed the ability of HIV-derived nanoblades to incorporate and deliver cargo, along with MLV-derived nanoblades [146]. By additionally co-transfecting BaEV, CRISPR-Cas9 system is delivered to human T cells, B cells, and hematopoietic stem and progenitor cells with no significant toxicity. In a recent study, nanoblades are used for gene knockout in organoids from murine prostate and both murine and human rectal organoids [147]. Similarly, no toxicity is seen in the organoids and no obvious off-target effects are observed after CRISPR-Cas9 knockout.
To improve the shortcomings of nanoblades, the previously mentioned ligand-induced EV packaging system termed NanoMEDIC was developed [109]. It is designed to deliver RNPs without the need for a proteolytic cleavage site, and is based on the association of FKBP12 and FRB in the presence of a rapamycin analog. Cells stably expressing sgRNA are transfected with plasmids encoding VSV-G, Cas9-FRB, and FKBP12-Gag containing a myristoylation motif for enrichment on the plasma membrane. Effective incorporation of RNPs into VLPs is only possible in the presence of a rapamycin analog. The NanoMEDIC system shows over 90% exon skipping efficiency in skeletal muscle cells with no observed cellular toxicity. Confirming the shortcomings of the previously mentioned nanoblades, removing the fused Pol protein containing an HIV protease cleavage site results in higher editing efficiency. As expected, the same editing efficiency is demonstrated when HIV protease inhibitor Darunavir is coupled with FKBP12-Gag-Pol, compared to transfection with FKBP12-Gag. Nevertheless, Gag and HIV-1 Tat (required to drive the expression of sgRNA in producer cells) proteins can be immunogenic when used in vivo.
To sum up, although VLPs resemble EVs and can provide efficient protein loading, viral components can trigger inflammatory responses in vivo, and VLPs may have some properties that are more inherent to viral vectors than natural EVs.
4.3. Exogenous Loading
Exogenous loading, also known as direct loading or post-loading, is a method for packaging therapeutics into purified EVs. Exogenous loading allows a wider range of therapeutics to be loaded into EVs, as cells are not influenced by therapeutics that may change cell viability or alter their phenotype. For instance, cell genomes do not change when a gene editing system is exogenously loaded [148].
Exogenous loading is performed using physical methods (electroporation, sonication, heat shock, freeze-thawing, transfection, or extrusion) or chemical methods (surfactant permeabilization, pH gradient modification, hypotonic dialysis, or lipid conjugation) (Figure 7).
4.3.1. Physical Methods
Electroporation is commonly used due to its cost efficiency and simplicity of operation [96]. A pulsed electrical current is applied to the sample to form temporary pores in the EV membrane. Besides small molecules and nucleic acids, this method can be utilized to load protein cargo [149]. Electroporation is an efficient and fast technique but can induce precipitation of cargo RNA [150] and change morphology of EV membranes with further EV aggregation [151]. Moreover, parameters like condenser capacity, voltage, number of pulses, pulse length, and interval duration should be optimized for each application [152]. To minimize EV aggregation after electroporation, the use of media containing 50 mM trehalose in PBS has been proposed [153]. Alternatively, buffers containing 400 mM sucrose in PBS can be utilized to improve loading efficiency while maintaining similar EV recovery [154].
Sonication is another physical method for exogenous cargo loading. With this method, extra mechanical shear force (sound energy) is applied to the sample and temporary pores are generated. Sonication allows high loading efficiency but can alter EV zeta potential and cause membrane damage and aggregation of EVs [152,155,156].
Heat shock is based on a sudden and significant increase in temperature to generate transient pores in the EV membrane. This method is relatively simple and easy to operate, does not require expensive equipment, and provides approximately the same loading efficiency as electroporation [157]. Possible drawbacks of the heat shock method include destabilization of the loaded nucleic acids, changes in the fluidity of EV membranes, and denaturation of membrane proteins [158,159].
Freeze-thawing is also based on temporary disruption of EV membrane integrity that may be associated with damage caused by the formation of ice crystals [160]. Freeze-thawing is a simple method that does not require any costly equipment. Despite its advantages, this method has relatively low drug loading efficiency [152]. During sudden temperature change, EV aggregation or destruction can occur, as well as induced protein damage [160,161].
Extrusion is another approach for exogenous cargo loading. A mixture of EVs and drugs is loaded into a syringe-based lipid extruder with 100–400 nm pores, and the EV membrane continuity decreases, resulting in cargo encapsulation within EVs [162]. Extrusion provides relatively high loading rates, improved EV size uniformity and homogeneity, does not require chemical additives or reagents, and is relatively cost-effective [96]. However, since extrusion can cause mechanical stress, it can lead to EV membrane reorganization and zeta potential changes, which can modulate immune responses and increase EV toxicity [96,152,163].
4.3.2. Chemical Methods
Chemical methods are based on application of chemical substances to increase permeability of EV membranes for cargo loading. Effectiveness of chemical methods are highly dependent of chemical structure of desired cargo.
EVs can be loaded with nucleic acids in a similar manner to cell transfection. Transfection with Exo-Fect (a cationic transfection reagent) is the most efficient method for miRNA loading, compared to electroporation, heat shock, saponin-based loading, and modification of miRNA with cholesterol [164]. This efficiency is observed for EVs isolated from various sources. Polyvalent cationic transfection reagent forms a complex with nucleic acids that facilitates loading via charge-charge interactions between the complex and EVs. EV transfection does not depend on cargo concentration and, therefore, improved loading is anticipated [165]. Nevertheless, transfection can affect EV mean size and zeta potential [156] and cause problems such as cytotoxicity. Moreover, difficulties in eliminating transfection reagent residue still remain; as aforementioned, lipid micelles from transfection reagents can mimic cargo-loaded EVs [166].
Chemical permeabilization mainly utilizes saponin, a surfactant molecule that can complex with cholesterol in cell membranes, generating pores [167]. Permeabilization with saponin is a relatively cheap and simple technique that provides high loading efficiency [168]. Nowadays, chemical permeabilization is rarely used due to saponin’s toxicity and hemolytic activity in vivo [155]. Saponin can damage EV membranes and change EV zeta potential, and not all cargo molecules can be mixed with surfactants [96,156].
pH gradient modification was originally developed to encapsulate weakly basic drugs (e.g., doxorubicin [DOX]) within liposomes [169]. When the drug is incubated at neutral pH with particles that have an interior acidic core, it diffuses into the lumen of the particles. While a pH gradient is more effective for entrapping basic drugs, a high transmembrane pH gradient (pH ~2) can still be applied for negatively charged molecules. Thus, a pH gradient across EV membranes can be used to load nucleic acids into EVs. To achieve this, one group dehydrated EVs in 70% ethanol and then rehydrated in an acidic citrate buffer (pH = 2.5) to establish the pH gradient [170]. This approach allowed enhanced loading of miRNA, siRNA, and single-stranded DNA into EVs with about the same loading efficiency as achieved by sonication, electroporation, or chemical permeabilization. Simultaneously, size and zeta potential were unaffected by this method [152]. However, addition of ethanol causes EV protein degradation and changes EV stability [148]; for example, milk-derived EVs are unstable during the ethanol dehydration step [171]. Moreover, low pH may induce acid hydrolysis of lipids like phosphatidylcholine, enhancing drug leakage and particle instability [172].
Hypotonic dialysis is a method for loading cargo during EV swelling. A hypotonic solution is added to form temporary pores in EVs, allowing water-soluble molecules to move into the vesicles with the water flow. Then EVs are dispersed in an isotonic solution, which favors EVs transition to their normal state. Hypotonic dialysis is a simple technique that provides high efficiency and does not require any specialized equipment but can alter the size and charge of EVs and cause degradation of protein and peptide cargo [152,173]. Furthermore, one study showed relatively low EV uptake by the cells after cargo loading with this method [168].
Lipid conjugation may also be used to improve loading of therapeutic RNAs and other hydrophilic ligands (e.g., peptides) into EVs. Covalent conjugation of RNA with lipids is mostly utilized by cholesterol conjugation, but fatty acids, sterols, and vitamins can also be anchored to the RNA of interest. For instance, conjugation with docosanoic acid and α-tocopheryl succinate shows the same or better siRNA loading into EVs than conjugation with cholesterol [174]. Vitamin E-conjugated siRNA supports the best loading into EVs, outperforming cholesterol-conjugated RNA. Hydrophobicity drives integration of lipid-conjugated hydrophilic molecules into the EV membrane during simple co-incubation. Synthesized hydrophobic RNAs package into EVs more efficiently [174]. Despite the advantages of lipid-conjugated RNA, most RNA attaches to the EV surface instead of being incorporated [175]: Reduced zeta potential and increased EV size are noted after loading, indicating deposition of RNA on EV surface [176]. Furthermore, hydrophilic cholesterol-anchored ligands are shed from EVs in presence of serum in less than two hours [177], potentially explaining the generally low efficiency of silencing activity of cholesterol-conjugated siRNA transferred by EVs in vivo.
5. Challenges of Loading Cargo into EVs
Although numerous loading techniques for therapeutic molecules have been developed, they still suffer from inadequate efficiency inferior to that of lipid-based NPs [178]. Large hydrophilic molecules are particularly difficult to package due to the presence of EV lipid bilayer membranes that protect naturally incorporated material.
Endogenous loading provides continuous production of drug-loaded EVs without damaging EV membranes, but most pre-loading methods are time-consuming and laborious. Compared to exogenous loading, endogenous packaging results in low loading efficiency for smaller molecules [93,173]. Moreover, it is hard to control the amount of packaged cargo into EVs with exogenous loading, and this amount depends on transfection efficiency and viability of the producing cells. As another drawback, loaded molecules may influence EV-producing cells, impacting their physiology and, therefore, changing the constitution of released EVs and/or limiting EV production [179].
Exogenous loading has its own disadvantages. EVs have plenty of proteins and nucleic acids inside, and it is unclear how much more therapeutics can be loaded efficiently [180]. As mentioned, most exogenous packaging methods can disrupt EV integrity, thereby altering the payload and delivery vehicle [142]. Furthermore, exogenous loading shows lower loading efficiency for larger molecules, such as proteins and nucleic acids larger than 1000 bp [181].
6. Surface Display of Functional Moieties on EVs
Fusion with EV-specific proteins is the most commonly used approach to display functional moieties on the surface of EVs. Displayed proteins or peptides can be used as targeting molecules, receptor agonists or antagonists, etc. Proteins or peptides can be included in extracellular loops of tetraspanins (CD63, CD9, and CD81) or fused to the extracellular domains of proteins highly enriched in EVs, such as lysosome-associated membrane protein 2, isoform B (Lamp2B) or lactadherin. Moreover, functional moieties can be displayed on the surface of EVs by employing transmembrane proteins enriched in EVs, such as prostaglandin F2 receptor inhibitor (PTGFRN), TSPAN14, TSPAN2, and TSPAN3.
Lamp2b is the most frequently used protein for displaying targeting moieties through cell engineering. For instance, Lamp2 fused to the neuron-specific rabies viral glycoprotein (RVG) peptide delivered EVs containing siRNA targeting GAPDH specifically to neurons, microglia, and oligodendrocytes in a mouse model [182]. Upon intravenous administration of these decorated RVG-modified EVs, strong neuron-specific knockdown of GAPDH gene expression was observed.
Lactadherin is a secretory protein that specifically binds to the EV surface through phosphatidylserine and becomes a membrane-associated protein [183]. Fusing anti-HER2 single-chain variable fragments with C1C2 domain of lactadherin protein demonstrates their specific delivery to HER2-positive human breast tumor xenografts [184]. In another study of in vivo tracing of EVs, a fusion protein was designed consisting of Gaussia luciferase (gLuc), C1C2 domains of lactadherin, and the N-terminal secretion signal of lactadherin [185]. This protein resulted in strong luciferase activity in the EV-containing fraction.
Functional moieties also can be included in the large extracellular loop (2nd loop) of CD63 for surface display. For example, the large extracellular loop of CD63 was genetically engineered for surface display of the albumin-binding domain to increase EV circulation time [186]. However, surface display of large molecules can compromise stability and EV-sorting ability of the fusion protein [187]. Alternatively, targeting molecules can be fused with specific CP05 peptide that are binds specifically to CD63 molecules at EV surface and expose targeting molecule. For example, functionalizing of EVs with muscle-targeting peptide and EV loading with splice–correcting oligomer for dystrophin exon skipping induced functional rescue and formation of dystrophin-positive muscle fibers in mdx mice [188].
Alternalive technology for ligand display is an anchoring via lipid molecules. In this technology, targeting molecule is fused with lipid (for example, cholesterol or phospholipid). Incubation of construct with EV result in incorporation of lipid molecule into EV lipid membrane and surface display of targeting ligand [189,190,191]. For example, Pi et al., used cholesterol-conjugated arrow-shaped RNA modified with PSMA-specific or EGFR-specific RNA aptamers or folate molecule for targeting in prostate cancer, breast cancer and colorectal cancer cells in mice models, respectively [190].
Another approach was demonstrated using glycophosphatidylinositol (GPI) anchor signaling peptide derived from decay-accelerating factor (DAF) fused to EG1a nanobody with high affinity to EGFR. As a result, targeting nanobodies were enriched in EVs and exposed at EV surface leading to increased uptake by EGFR-overexpressing cells compared to unfunctionalized vesicles [192].
While most studies for EV surface display have used protein and peptide ligands, glycan ligands can also be used for cell-specific targeting. This is accomplished by co-expressing a glycosylation domain inserted into the large extracellular loop of CD63 and fucosyltransferase VII (FUT7) or IX (FUT9) responsible for surface display of the glycan of interest [187]. FUT7 and FUT9 catalyze fucosylation of one of the residues of a short glycosylation domain. These structures act as glycan ligands and mediate adhesion to activated endothelial cells or dendritic cells, respectively. Various available glycosyltransferases can be used for surface display of glycans with different biological functions, making this approach attractive.
Exposure of targeting molecules independently of surface proteins can be performed by click-chemistry approach. In this way, exact chemical groups of all surface protein are interact with reactogenic chemical compound in mild conditions forming activated chemical group. During second step, targeting molecule react with activated group by forming covalent links, resulting in exposure of targeting molecule. Reactions can be performer in neutral physiological buffers without catalysts. MSC-derived EVs modified with c(RGDyK) peptide and loaded with anti-inflammatory drug curcumin were tested in cerebral artery occlusion mice model. Functionalized particles efficiently binded to target integrin αvβ3 overexpressed in ischemic brain sites and reduced inflammation and cells apoptosis at the site of ischemia [193].
Displayed molecules on the EV surface may be unstable in vivo and can be attacked by membrane proteases [194]. Another challenge is the efficient display of functional moieties on the surface of EVs. For instance, some peptides fused to Lamp2b were not displayed efficiently on the EV surface [195]. Moreover, EV markers are not found on all EVs, or are not present at densities optimal for engineering [196]. Overall, efficient surface exposure of desired molecules is an area of further research.
7. Conclusions
There are currently no FDA-approved EV products, and many studies are still in the early stages, indicating that EV-based therapeutics have a long way to go before they can be widely used in clinical therapy. Moreover, issues like short half-life of EVs and cell type-specific targeting remain and hinder EV use [197]. Despite the promising applications of EVs, major limitations linger in our current understanding of EV biogenesis and cargo encapsulation. Understanding of this processes will allow to extremely improve packaging of therapeutic cargo to improve efficiency of delivery by extracellular vesicles for most perspective drugs, for example, CRISPR-Cas gene editing systems. For example, efficiency of CRISPR-Cas packaging in EV-mimetic Nanoblades system could achieve tens of Cas-protein copies per vesicle that result in superior gene editing efficiency [144]. Improvement in surface display of targeting ligands will open a way to highly specific biodistribution of vesicles into target tissue, resulting in further increase of drug delivery by EVs.
Author Contributions
Sergey Brezgin: conceptualization, writing original draft, review and editing; Oleg Danilik – conceptualization, writing original draft; Alexandra Yudaeva – writing-review and editing, visualization; Artyom Kachanov – writing-review and editing, visualization; Anastasiya Kostyusheva – writing-original draft, writing-review and editing, visualization; Ivan Karandashov – writing-review and editing, visualization; Natalia Ponomareva – writing-review and editing; Andrey A. Zamyatnin Jr. – writing-review and editing, resources, supervision; Alessandro Parodi – writing-original draft, writing-review and editing, supervision; Vladimir Chulanov – writing-review and editing, supervision, funding acquisition; Dmitry Kostyushev – conceptualization, resources, writing-original draft, writing-review and editing, supervision, project administration.
Funding
This study was supported by RSF grant no. 20-15-00373-P. Sections 3 and 5 were funded by Academic Leadership Program Priority 2030 (Sechenov University).
Data Availability Statement
No data was generated in this manuscript.
Conflicts of Interest
Nothing to declare.
References
- Ginini, L.; Billan, S.; Fridman, E.; Gil, Z. Insight into Extracellular Vesicle-Cell Communication: From Cell Recognition to Intracellular Fate. Cells 2022, 11, 1375. [Google Scholar] [CrossRef] [PubMed]
- Cecchin, R.; Troyer, Z.; Witwer, K.; Morris, K.V. Extracellular vesicles: The next generation in gene therapy delivery. Mol. Ther. 2023, 31, 1225–1230. [Google Scholar] [CrossRef]
- Banks, W.A.; Sharma, P.; Bullock, K.M.; Hansen, K.M.; Ludwig, N.; Whiteside, T.L. Transport of Extracellular Vesicles across the Blood-Brain Barrier: Brain Pharmacokinetics and Effects of Inflammation. Int. J. Mol. Sci. 2020, 21, 4407. [Google Scholar] [CrossRef]
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef]
- Wei, Z.; Chen, Z.; Zhao, Y.; Fan, F.; Xiong, W.; Song, S.; Yin, Y.; Hu, J.; Yang, K.; Yang, L.; et al. Mononuclear phagocyte system blockade using extracellular vesicles modified with CD47 on membrane surface for myocardial infarction reperfusion injury treatment. Biomaterials 2021, 275, 121000. [Google Scholar] [CrossRef]
- Parodi, A.; Molinaro, R.; Sushnitha, M.; Evangelopoulos, M.; Martinez, J.O.; Arrighetti, N.; Corbo, C.; Tasciotti, E. Bio-inspired engineering of cell- and virus-like nanoparticles for drug delivery. Biomaterials 2017, 147, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.T.; Hosseini-Beheshti, E.; Afrose, D.; Ding, X.; Xia, B.; Grau, G.; Little, C.; McClements, L.; Li, J. Extracellular Vesicles from Mesenchymal Stromal Cells for the Treatment of Inflammation-Related Conditions. Int. J. Mol. Sci. 2021, 22, 3023. [Google Scholar] [CrossRef] [PubMed]
- Duong, A.; Parmar, G.; Kirkham, A.M.; Burger, D.; Allan, D.S. Registered clinical trials investigating treatment with cell-derived extracellular vesicles: a scoping review. Cytotherapy 2023, 25, 939–945. [Google Scholar] [CrossRef]
- Herrmann, I.K.; Wood, M.J.A.; Fuhrmann, G. Extracellular vesicles as a next-generation drug delivery platform. Nat. Nanotechnol. 2021, 16, 748–759. [Google Scholar] [CrossRef]
- Nieuwland, R.; Falcón-Pérez, J.M.; Théry, C.; Witwer, K.W. Rigor and standardization of extracellular vesicle research: Paving the road towards robustness. J. Extracell. Vesicles 2020, 10. [Google Scholar] [CrossRef]
- Brezgin, S.; Parodi, A.; Kostyusheva, A.; Ponomareva, N.; Lukashev, A.; Sokolova, D.; Pokrovsky, V.S.; Slatinskaya, O.; Maksimov, G.; Zamyatnin, A.A.; et al. Technological aspects of manufacturing and analytical control of biological nanoparticles. Biotechnol. Adv. 2023, 64, 108122. [Google Scholar] [CrossRef]
- Jeppesen, D.K.; Zhang, Q.; Franklin, J.L.; Coffey, R.J. Extracellular vesicles and nanoparticles: emerging complexities. Trends Cell Biol. 2023, 33, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, Y.; Peng, J.; Wu, D.; Zhao, X.; Cui, Y.; Chen, L.; Yan, X.; Du, Y.; Yu, L. Discovery of the migrasome, an organelle mediating release of cytoplasmic contents during cell migration. Cell Res. 2014, 25, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Melentijevic, I.; Toth, M.L.; Arnold, M.L.; Guasp, R.J.; Harinath, G.; Nguyen, K.C.; Taub, D.; Parker, J.A.; Neri, C.; Gabel, C.V.; et al. elegans neurons jettison protein aggregates and mitochondria under neurotoxic stress. Nature 2017, 542, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Colao, I.L.; Corteling, R.; Bracewell, D.; Wall, I. Manufacturing Exosomes: A Promising Therapeutic Platform. Trends Mol. Med. 2018, 24, 242–256. [Google Scholar] [CrossRef]
- EV-TRACK Consortium; Van Deun, J.; Mestdagh, P.; Agostinis, P.; Akay, Ö.; Anand, S.; Anckaert, J.; Martinez, Z.A.; Baetens, T.; Beghein, E.; et al. EV-TRACK: Transparent reporting and centralizing knowledge in extracellular vesicle research. Nat. Methods 2017, 14, 228–232. [Google Scholar] [CrossRef]
- Gardiner, C.; Di Vizio, D.; Sahoo, S.; Théry, C.; Witwer, K.W.; Wauben, M.; Hill, A.F. Techniques used for the isolation and characterization of extracellular vesicles: Results of a worldwide survey. J. Extracell. Vesicles 2016, 5, 32945. [Google Scholar] [CrossRef]
- Thery, C.; Clayton, A.; Amigorena, S.; Raposo, G. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 2006, 3. [Google Scholar] [CrossRef]
- Konoshenko, M.Y.; Lekchnov, E.A.; Vlassov, A.V.; Laktionov, P.P. Isolation of Extracellular Vesicles: General Methodologies and Latest Trends. BioMed Res. Int. 2018, 2018, 8545347. [Google Scholar] [CrossRef]
- Zarovni, N.; Corrado, A.; Guazzi, P.; Zocco, D.; Lari, E.; Radano, G.; Muhhina, J.; Fondelli, C.; Gavrilova, J.; Chiesi, A. Integrated isolation and quantitative analysis of exosome shuttled proteins and nucleic acids using immunocapture approaches. Methods 2015, 87, 46–58. [Google Scholar] [CrossRef]
- Witwer, K.W.; Buzás, E.I.; Bemis, L.T.; Bora, A.; Lässer, C.; Lötvall, J.; Nolte-’t Hoen, E.N.; Piper, M.G.; Sivaraman, S.; Skog, J.; et al. Standardization of sample collection, isolation and analysis methods in extracellular vesicle research. J. Extracell. Vesicles 2013, 2, 20360. [Google Scholar] [CrossRef] [PubMed]
- Linares, R.; Tan, S.; Gounou, C.; Arraud, N.; Brisson, A.R. High-speed centrifugation induces aggregation of extracellular vesicles. J. Extracell. Vesicles 2015, 4, 29509. [Google Scholar] [CrossRef] [PubMed]
- Lobb, R.j.; Becker, M.; Wen, S.W.; Wong, C.S.F.; Wiegmans, A.P.; Leimgruber, A.; Möller, A. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef]
- Benayas, B.; Morales, J.; Egea, C.; Armisén, P.; Yáñez-Mó, M. Optimization of extracellular vesicle isolation and their separation from lipoproteins by size exclusion chromatography. J. Extracell. Biol. 2023, 2, e100. [Google Scholar] [CrossRef]
- Livshits, M.A.; Khomyakova, E.; Evtushenko, E.G.; Lazarev, V.N.; Kulemin, N.A.; Semina, S.E.; Generozov, E.V.; Govorun, V.M. Isolation of exosomes by differential centrifugation: Theoretical analysis of a commonly used protocol. Sci. Rep. 2015, 5, 17319–17319. [Google Scholar] [CrossRef] [PubMed]
- Lamparski, H.G.; Metha-Damani, A.; Yao, J.-Y.; Patel, S.; Hsu, D.-H.; Ruegg, C.; Le Pecq, J.-B. Production and characterization of clinical grade exosomes derived from dendritic cells. J. Immunol. Methods 2002, 270, 211–226. [Google Scholar] [CrossRef]
- Tauro, B.J.; Greening, D.W.; Mathias, R.A.; Ji, H.; Mathivanan, S.; Scott, A.M.; Simpson, R.J. Comparison of ultracentrifugation, density gradient separation, and immunoaffinity capture methods for isolating human colon cancer cell line LIM1863-derived exosomes. Methods 2012, 56, 293–304. [Google Scholar] [CrossRef]
- Onódi, Z.; Pelyhe, C.; Nagy, C.T.; Brenner, G.B.; Almási, L.; Kittel, M.; Manček-Keber, M.; Ferdinandy, P.; Buzás, E.I.; Giricz, Z. Isolation of High-Purity Extracellular Vesicles by the Combination of Iodixanol Density Gradient Ultracentrifugation and Bind-Elute Chromatography From Blood Plasma. Front. Physiol. 2018, 9, 1479. [Google Scholar] [CrossRef]
- Welsh, J.A.; Goberdhan, D.C.I.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.P.; Erdbrügger, U.; et al. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J. Extracell. Vesicles 2024, 13, e12404. [Google Scholar] [CrossRef]
- Sanz-Ros, J.; Mas-Bargues, C.; Romero-García, N.; Huete-Acevedo, J.; Dromant, M.; Borrás, C. Extracellular Vesicles as Therapeutic Resources in the Clinical Environment. Int. J. Mol. Sci. 2023, 24, 2344. [Google Scholar] [CrossRef]
- El Baradie, K.B.Y.; Nouh, M.; Iii, F.O.; Liu, Y.; Fulzele, S.; Eroglu, A.; Hamrick, M.W. Freeze-Dried Extracellular Vesicles From Adipose-Derived Stem Cells Prevent Hypoxia-Induced Muscle Cell Injury. Front. Cell Dev. Biol. 2020, 8, 181. [Google Scholar] [CrossRef]
- Benedikter, B.J.; Bouwman, F.G.; Vajen, T.; Heinzmann, A.C.A.; Grauls, G.; Mariman, E.C.; Wouters, E.F.M.; Savelkoul, P.H.; Lopez-Iglesias, C.; Koenen, R.R.; et al. Ultrafiltration combined with size exclusion chromatography efficiently isolates extracellular vesicles from cell culture media for compositional and functional studies. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Visan, K.S.; Wu, L.-Y.; Voss, S.; Wuethrich, A.; Möller, A. Status quo of Extracellular Vesicle isolation and detection methods for clinical utility. Semin. Cancer Biol. 2023, 88, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Polyakov, Y.S.; Zydney, A.L. Ultrafiltration membrane performance: Effects of pore blockage/constriction. J. Membr. Sci. 2013, 434, 106–120. [Google Scholar] [CrossRef]
- Chernyshev, V.S.; Chuprov-Netochin, R.N.; Tsydenzhapova, E.; Svirshchevskaya, E.V.; Poltavtseva, R.A.; Merdalimova, A.; Yashchenok, A.; Keshelava, A.; Sorokin, K.; Keshelava, V.; et al. Asymmetric depth-filtration: A versatile and scalable method for high-yield isolation of extracellular vesicles with low contamination. J. Extracell. Vesicles 2022, 11, e12256. [Google Scholar] [CrossRef] [PubMed]
- Liangsupree, T.; Multia, E.; Riekkola, M.-L. Modern isolation and separation techniques for extracellular vesicles. J. Chromatogr. A 2020, 1636, 461773. [Google Scholar] [CrossRef] [PubMed]
- Busatto, S.; Vilanilam, G.; Ticer, T.; Lin, W.-L.; Dickson, D.W.; Shapiro, S.; Bergese, P.; Wolfram, J. Tangential Flow Filtration for Highly Efficient Concentration of Extracellular Vesicles from Large Volumes of Fluid. Cells 2018, 7, 273. [Google Scholar] [CrossRef]
- Visan, K.S.; Lobb, R.J.; Ham, S.; Lima, L.G.; Palma, C.; Edna, C.P.Z.; Wu, L.; Gowda, H.; Datta, K.K.; Hartel, G.; et al. Comparative analysis of tangential flow filtration and ultracentrifugation, both combined with subsequent size exclusion chromatography, for the isolation of small extracellular vesicles. J. Extracell. Vesicles 2022, 11, e12266. [Google Scholar] [CrossRef]
- Heath, N.; Grant, L.; De Oliveira, T.M.; Rowlinson, R.; Osteikoetxea, X.; Dekker, N.; Overman, R. Rapid isolation and enrichment of extracellular vesicle preparations using anion exchange chromatography. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Böing, A.N.; van der Pol, E.; Grootemaat, A.E.; Coumans, F.A.W.; Sturk, A.; Nieuwland, R. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J. Extracell. Vesicles 2014, 3. [Google Scholar] [CrossRef]
- Staubach, S.; Bauer, F.N.; Tertel, T.; Börger, V.; Stambouli, O.; Salzig, D.; Giebel, B. Scaled preparation of extracellular vesicles from conditioned media. Adv. Drug Deliv. Rev. 2021, 177, 113940. [Google Scholar] [CrossRef]
- Muller, L.; Hong, C.-S.; Stolz, D.B.; Watkins, S.C.; Whiteside, T.L. Isolation of biologically-active exosomes from human plasma. J. Immunol. Methods 2014, 411, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.D.; Shah, S. Methods of isolating extracellular vesicles impact down-stream analyses of their cargoes. Methods 2015, 87, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Lins, P.M.P.; Pirlet, E.; Szymonik, M.; Bronckaers, A.; Nelissen, I. Manufacture of extracellular vesicles derived from mesenchymal stromal cells. Trends Biotechnol. 2023, 41, 965–981. [Google Scholar] [CrossRef]
- Sidhom, K.; Obi, P.O.; Saleem, A. A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef]
- Wang, L.; Wang, D.; Ye, Z.; Xu, J. Engineering Extracellular Vesicles as Delivery Systems in Therapeutic Applications. Adv. Sci. 2023, 10, e2300552. [Google Scholar] [CrossRef]
- Ludwig, A.; De Miroschedji, K.; Doeppner, T.R.; Börger, V.; Ruesing, J.; Rebmann, V.; Durst, S.; Jansen, S.; Bremer, M.; Behrmann, E.; et al. Precipitation with polyethylene glycol followed by washing and pelleting by ultracentrifugation enriches extracellular vesicles from tissue culture supernatants in small and large scales. J. Extracell. Vesicles 2018, 7, 1528109. [Google Scholar] [CrossRef]
- Rekker, K.; Saare, M.; Roost, A.M.; Kubo, A.-L.; Zarovni, N.; Chiesi, A.; Salumets, A.; Peters, M. Comparison of serum exosome isolation methods for microRNA profiling. Clin. Biochem. 2014, 47, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Névo, N.; Jouve, M.; Valenzuela, J.I.; Maurin, M.; Verweij, F.J.; Palmulli, R.; Lankar, D.; Dingli, F.; Loew, D.; et al. Specificities of exosome versus small ectosome secretion revealed by live intracellular tracking of CD63 and CD9. Nat. Commun. 2021, 12, 1–18. [Google Scholar] [CrossRef]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in Exosome Isolation Techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef]
- Doyle, L.; Wang, M. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- Karimi, N.; Dalirfardouei, R.; Dias, T.; Lötvall, J.; Lässer, C. Tetraspanins distinguish separate extracellular vesicle subpopulations in human serum and plasma – Contributions of platelet extracellular vesicles in plasma samples. J. Extracell. Vesicles 2022, 11, e12213. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, D.; Ho, E.A. Challenges in the development and establishment of exosome-based drug delivery systems. J. Control. Release 2020, 329, 894–906. [Google Scholar] [CrossRef] [PubMed]
- De Voogt, W.S.; Frunt, R.; Leandro, R.M.; Triesscheijn, C.S.; Monica, B.; Paspali, I.; Tielemans, M.; Francois, J.J.J.M.; Seinen, C.W.; De Jong, O.G.; et al. EV-Elute: A Universal Platform for Enrichment of Functional Surface Marker-Defined Extracellular Vesicle Subpopulations (Preprint). BioRxiv 2023. [Google Scholar]
- Matsuzaka, Y.; Yashiro, R. Advances in Purification, Modification, and Application of Extracellular Vesicles for Novel Clinical Treatments. Membranes 2022, 12, 1244. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Mao, J.; Barrero, R.A.; Wang, P.; Zhang, F.; Wang, T. Development of a CD63 Aptamer for Efficient Cancer Immunochemistry and Immunoaffinity-Based Exosome Isolation. Molecules 2020, 25, 5585. [Google Scholar] [CrossRef]
- Bonner, S.E.; van de Wakker, S.I.; Phillips, W.; Willms, E.; Sluijter, J.P.G.; Hill, A.F.; Wood, M.J.A.; Vader, P. Scalable purification of extracellular vesicles with high yield and purity using multimodal flowthrough chromatography. J. Extracell. Biol. 2024, 3, e138. [Google Scholar] [CrossRef]
- Hallal, S.; Tűzesi, G.E.; Grau, G.E.; Buckland, M.E.; Alexander, K.L. Understanding the extracellular vesicle surface for clinical molecular biology. J. Extracell. Vesicles 2022, 11, e12260. [Google Scholar] [CrossRef]
- Seo, N.; Nakamura, J.; Kaneda, T.; Tateno, H.; Shimoda, A.; Ichiki, T.; Furukawa, K.; Hirabayashi, J.; Akiyoshi, K.; Shiku, H. Distinguishing functional exosomes and other extracellular vesicles as a nucleic acid cargo by the anion-exchange method. J. Extracell. Vesicles 2022, 11, e12205. [Google Scholar] [CrossRef]
- Notarangelo, M.; Zucal, C.; Modelska, A.; Pesce, I.; Scarduelli, G.; Potrich, C.; Lunelli, L.; Pederzolli, C.; Pavan, P.; la Marca, G.; et al. Ultrasensitive detection of cancer biomarkers by nickel-based isolation of polydisperse extracellular vesicles from blood. EBioMedicine 2019, 43, 114–126. [Google Scholar] [CrossRef]
- Koch, L.F.; Best, T.; Wüstenhagen, E.; Adrian, K.; Rammo, O.; Saul, M.J. Novel insights into the isolation of extracellular vesicles by anion exchange chromatography. Front. Bioeng. Biotechnol. 2024, 11, 1298892. [Google Scholar] [CrossRef] [PubMed]
- Stam, J.; Bartel, S.; Bischoff, R.; Wolters, J.C. Isolation of extracellular vesicles with combined enrichment methods. J. Chromatogr. B Analyt Technol. Biomed. Life Sci. 2021, 1169, 122604. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Nguyen, L.T.H.; Hickey, R.; Walters, N.; Wang, X.; Kwak, K.J.; Lee, L.J.; Palmer, A.F.; Reátegui, E. Immunomagnetic sequential ultrafiltration (iSUF) platform for enrichment and purification of extracellular vesicles from biofluids. Sci. Rep. 2021, 11, 1–17. [Google Scholar] [CrossRef]
- Paganini, C.; Palmiero, U.C.; Pocsfalvi, G.; Touzet, N.; Bongiovanni, A.; Arosio, P. Scalable Production and Isolation of Extracellular Vesicles: Available Sources and Lessons from Current Industrial Bioprocesses. Biotechnol. J. 2019, 14, e1800528. [Google Scholar] [CrossRef]
- Vo, N.; Tran, C.; Tran, N.H.B.; Nguyen, N.T.; Nguyen, T.; Ho, D.T.K.; Nguyen, D.D.N.; Pham, T.; Nguyen, T.A.; Phan, H.T.N.; et al. A novel multi-stage enrichment workflow and comprehensive characterization for HEK293F-derived extracellular vesicles. J. Extracell. Vesicles 2024, 13, e12454. [Google Scholar] [CrossRef]
- Wang, S.; Li, C.; Yuan, Y.; Xiong, Y.; Xu, H.; Pan, W.; Pan, H.; Zhu, Z. Microvesicles as drug delivery systems: A new frontier for bionic therapeutics in cancer. J. Drug Deliv. Sci. Technol. 2023, 79. [Google Scholar] [CrossRef]
- Johnston, J.; Stone, T.; Wang, Y. Biomaterial-enabled 3D cell culture technologies for extracellular vesicle manufacturing. Biomater. Sci. 2023, 11, 4055–4072. [Google Scholar] [CrossRef]
- Paolini, L.; Monguió-Tortajada, M.; Costa, M.; Antenucci, F.; Barilani, M.; Clos-Sansalvador, M.; Andrade, A.C.; Driedonks, T.A.P.; Giancaterino, S.; Kronstadt, S.M.; et al. Large-scale production of extracellular vesicles: Report on the “massivEVs” ISEV workshop. J. Extracell. Biol. 2022, 1. [Google Scholar] [CrossRef]
- Ng, C.Y.; Kee, L.T.; Al-Masawa, M.E.; Lee, Q.H.; Subramaniam, T.; Kok, D.; Ng, M.H.; Law, J.X. Scalable Production of Extracellular Vesicles and Its Therapeutic Values: A Review. Int. J. Mol. Sci. 2022, 23, 7986. [Google Scholar] [CrossRef]
- Goncalves, J.P.; Ghebosu, R.E.; Tan, X.N.S.; Iannotta, D.; Koifman, N.; Wolfram, J. Hyaluronic acid: An overlooked extracellular vesicle contaminant. J. Extracell. Vesicles 2023, 12, e12362. [Google Scholar] [CrossRef]
- Shelke, G.V.; Lässer, C.; Gho, Y.S.; Lötvall, J. Importance of exosome depletion protocols to eliminate functional and RNA-containing extracellular vesicles from fetal bovine serum. J. Extracell. Vesicles 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lee, Y.; Johansson, H.J.; Mäger, I.; Vader, P.; Nordin, J.Z.; Wiklander, O.P.B.; Lehtiö, J.; Wood, M.J.A.; EL Andaloussi, S. Serum-free culture alters the quantity and protein composition of neuroblastoma-derived extracellular vesicles. J. Extracell. Vesicles 2015, 4, 26883. [Google Scholar] [CrossRef] [PubMed]
- Castiglia, S.; Mareschi, K.; Labanca, L.; Lucania, G.; Leone, M.; Sanavio, F.; Castello, L.; Rustichelli, D.; Signorino, E.; Gunetti, M.; et al. Inactivated human platelet lysate with psoralen: a new perspective for mesenchymal stromal cell production in Good Manufacturing Practice conditions. Cytotherapy 2014, 16, 750–763. [Google Scholar] [CrossRef] [PubMed]
- Marinho, A.; Nunes, C.; Reis, S. Hyaluronic Acid: A Key Ingredient in the Therapy of Inflammation. Biomolecules 2021, 11, 1518. [Google Scholar] [CrossRef]
- van de Wakker, S.I.; Bauzá-Martinez, J.; Arceo, C.R.; Manjikian, H.; Blok, C.J.B.S.; Roefs, M.T.; Willms, E.; Maas, R.G.C.; Pronker, M.F.; de Jong, O.G.; et al. Size matters: Functional differences of small extracellular vesicle subpopulations in cardiac repair responses. J. Extracell. Vesicles 2024, 13, e12396. [Google Scholar] [CrossRef]
- Görgens, A.; Corso, G.; Hagey, D.W.; Wiklander, R.J.; Gustafsson, M.O.; Felldin, U.; Lee, Y.; Bostancioglu, R.B.; Sork, H.; Liang, X.; et al. Identification of storage conditions stabilizing extracellular vesicles preparations. J. Extracell. Vesicles 2022, 11, e12238. [Google Scholar] [CrossRef]
- Trenkenschuh, E.; Richter, M.; Heinrich, E.; Koch, M.; Fuhrmann, G.; Friess, W. Enhancing the Stabilization Potential of Lyophilization for Extracellular Vesicles. Adv. Heal. Mater. 2021, 11, 2100538. [Google Scholar] [CrossRef]
- Susa, F.; Limongi, T.; Borgione, F.; Peiretti, S.; Vallino, M.; Cauda, V.; Pisano, R. Comparative Studies of Different Preservation Methods and Relative Freeze-Drying Formulations for Extracellular Vesicle Pharmaceutical Applications. ACS Biomater. Sci. Eng. 2023, 9, 5871–5885. [Google Scholar] [CrossRef]
- Ghodasara, A.; Raza, A.; Wolfram, J.; Salomon, C.; Popat, A. Clinical Translation of Extracellular Vesicles. Adv. Heal. Mater. 2023, 12, e2301010. [Google Scholar] [CrossRef]
- Elsharkasy, O.M.; Nordin, J.Z.; Hagey, D.W.; De Jong, O.G.; Schiffelers, R.M.; Andaloussi, S.E.; Vader, P. Extracellular vesicles as drug delivery systems: Why and how? Adv. Drug Deliv. Rev. 2020, 159, 332–343. [Google Scholar] [CrossRef]
- Jeske, R.; Liu, C.; Duke, L.; Castro, M.L.C.; Muok, L.; Arthur, P.; Singh, M.; Jung, S.; Sun, L.; Li, Y. Upscaling human mesenchymal stromal cell production in a novel vertical-wheel bioreactor enhances extracellular vesicle secretion and cargo profile. Bioact. Mater. 2023, 25, 732–747. [Google Scholar] [CrossRef] [PubMed]
- Mendt, M.; Kamerkar, S.; Sugimoto, H.; McAndrews, K.M.; Wu, C.-C.; Gagea, M.; Yang, S.; Blanko, E.V.R.; Peng, Q.; Ma, X.; et al. Generation and testing of clinical-grade exosomes for pancreatic cancer. J. Clin. Investig. 2018, 3. [Google Scholar] [CrossRef]
- Mahmoudi, M.; Landry, M.P.; Moore, A.; Coreas, R. The protein corona from nanomedicine to environmental science. Nat. Rev. Mater. 2023, 8, 422–438. [Google Scholar] [CrossRef] [PubMed]
- Tenzer, S.; Docter, D.; Kuharev, J.; Musyanovych, A.; Fetz, V.; Hecht, R.; Schlenk, F.; Fischer, D.; Kiouptsi, K.; Reinhardt, C.; et al. Rapid formation of plasma protein corona critically affects nanoparticle pathophysiology. Nat. Nanotechnol. 2013, 8, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Corbo, C.; Molinaro, R.; Parodi, A.; Furman, N.E.T.; Salvatore, F.; Tasciotti, E. The Impact of Nanoparticle Protein Corona on Cytotoxicity, Immunotoxicity and Target Drug Delivery. Nanomedicine 2015, 11, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Francia, V.; Schiffelers, R.M.; Cullis, P.R.; Witzigmann, D. The Biomolecular Corona of Lipid Nanoparticles for Gene Therapy. Bioconjugate Chem. 2020, 31, 2046–2059. [Google Scholar] [CrossRef]
- Corbo, C.; Molinaro, R.; Taraballi, F.; Furman, N.E.T.; Sherman, M.B.; Parodi, A.; Salvatore, F.; Tasciotti, E. Effects of the protein corona on liposome–liposome and liposome–cell interactions. Int. J. Nanomed. 2016, 3049–3063. [Google Scholar]
- Lima, T.; Bernfur, K.; Vilanova, M.; Cedervall, T. Understanding the Lipid and Protein Corona Formation on Different Sized Polymeric Nanoparticles. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Durán, N.; Silveira, C.P.; Durán, M.; Martinez, D.S.T. Silver nanoparticle protein corona and toxicity: a mini-review. J. Nanobiotechnology 2015, 13, 1–17. [Google Scholar] [CrossRef]
- Tóth, E.; Turiák, L.; Visnovitz, T.; Cserép, C.; Mázló, A.; Sódar, B.W.; Försönits, A.I.; Petővári, G.; Sebestyén, A.; Komlósi, Z.; et al. Formation of a protein corona on the surface of extracellular vesicles in blood plasma. J. Extracell. Vesicles 2021, 10, e12140. [CrossRef]
- Wolf, M.; Poupardin, R.W.; Ebner-Peking, P.; Andrade, A.C.; Blöchl, C.; Obermayer, A.; Gomes, F.G.; Vari, B.; Maeding, N.; Eminger, E.; et al. A functional corona around extracellular vesicles enhances angiogenesis, skin regeneration and immunomodulation. J. Extracell. Vesicles 2022, 11, e12207. [Google Scholar] [CrossRef] [PubMed]
- Haney, M.J.; Klyachko, N.L.; Zhao, Y.; Gupta, R.; Plotnikova, E.G.; He, Z.; Patel, T.; Piroyan, A.; Sokolsky, M.; Kabanov, A.V.; et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J. Control. Release 2015, 207, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Godbout, K.; Lamothe, G.; Tremblay, J.P. CRISPR-Cas9 delivery strategies with engineered extracellular vesicles. Mol. Ther. - Nucleic Acids 2023, 34, 102040. [Google Scholar] [CrossRef]
- Walker, S.; Busatto, S.; Pham, A.; Tian, M.; Suh, A.; Carson, K.; Quintero, A.; Lafrence, M.; Malik, H.; Santana, M.X.; et al. Extracellular vesicle-based drug delivery systems for cancer treatment. Theranostics 2019, 9, 8001–8017. [Google Scholar] [CrossRef]
- Han, Y.; Jones, T.W.; Dutta, S.; Zhu, Y.; Wang, X.; Narayanan, S.P.; Fagan, S.C.; Zhang, D. Overview and Update on Methods for Cargo Loading into Extracellular Vesicles. Processes 2021, 9, 356. [Google Scholar] [CrossRef]
- Dimik, M.; Abeysinghe, P.; Logan, J.; Mitchell, M. The exosome: a review of current therapeutic roles and capabilities in human reproduction. Drug Deliv. Transl. Res. 2022, 13, 473–502. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Yang, Q.; Sun, X.; Wang, Y. Recent Advancements in the Loading and Modification of Therapeutic Exosomes. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef]
- Kenari, A.N.; Cheng, L.; Hill, A.F. Methods for loading therapeutics into extracellular vesicles and generating extracellular vesicles mimetic-nanovesicles. Methods 2020, 177, 103–113. [Google Scholar] [CrossRef]
- McCann, J.; Sosa-Miranda, C.D.; Guo, H.; Reshke, R.; Savard, A.; Buzatto, A.Z.; Taylor, J.A.; Li, L.; Gibbings, D.J. Contaminating transfection complexes can masquerade as small extracellular vesicles and impair their delivery of RNA. J. Extracell. Vesicles 2022, 11. [Google Scholar] [CrossRef]
- McConnell, R.E.; Youniss, M.; Gnanasambandam, B.; Shah, P.; Zhang, W.; Finn, J.D. Transfection reagent artefact likely accounts for some reports of extracellular vesicle function. J. Extracell. Vesicles 2022, 11. [Google Scholar] [CrossRef]
- Rädler, J.; Gupta, D.; Zickler, A.; EL Andaloussi, S. Exploiting the biogenesis of extracellular vesicles for bioengineering and therapeutic cargo loading. Mol. Ther. 2023, 31, 1231–1250. [Google Scholar] [CrossRef]
- Dellar, E.R.; Hill, C.; Melling, G.E.; Carter, D.R.; Baena-Lopez, L.A. Unpacking extracellular vesicles: RNA cargo loading and function. J. Extracell. Biol. 2022, 1. [Google Scholar] [CrossRef] [PubMed]
- Bolukbasi, M.F.; Mizrak, A.; Ozdener, G.B.; Madlener, S.; Ströbel, T.; Erkan, E.P.; Fan, J.-B.; O Breakefield, X.; Saydam, O. miR-1289 and “Zipcode”-like Sequence Enrich mRNAs in Microvesicles. Mol. Ther. - Nucleic Acids 2012, 1, e10. [Google Scholar] [CrossRef]
- Temoche-Diaz, M.M.; Shurtleff, M.J.; Nottingham, R.M.; Yao, J.; Fadadu, R.P.; Lambowitz, A.M.; Schekman, R.; States, U. Distinct mechanisms of microRNA sorting into cancer cell-derived extracellular vesicle subtypes. eLife 2019, 8. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Gutiérrez-Vázquez, C.; Sánchez-Cabo, F.; Pérez-Hernández, D.; Vázquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sánchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef]
- Santangelo, L.; Giurato, G.; Cicchini, C.; Montaldo, C.; Mancone, C.; Tarallo, R.; Battistelli, C.; Alonzi, T.; Weisz, A.; Tripodi, M. The RNA-Binding Protein SYNCRIP Is a Component of the Hepatocyte Exosomal Machinery Controlling MicroRNA Sorting. Cell Rep. 2016, 17, 799–808. [Google Scholar] [CrossRef]
- Rufino-Ramos, D.; Albuquerque, P.R.; Leandro, K.; Carmona, V.; Martins, I.M.; Fernandes, R.; Henriques, C.; Lobo, D.; Faro, R.; Perfeito, R.; et al. Extracellular vesicle-based delivery of silencing sequences for the treatment of Machado-Joseph disease/spinocerebellar ataxia type 3. Mol. Ther. 2023, 31, 1275–1292. [Google Scholar] [CrossRef] [PubMed]
- Kuzembayeva, M.; Dilley, K.; Sardo, L.; Hu, W.-S. Life of psi: How full-length HIV-1 RNAs become packaged genomes in the viral particles. Virology 2014, 454-455, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Gee, P.; Lung, M.S.Y.; Okuzaki, Y.; Sasakawa, N.; Iguchi, T.; Makita, Y.; Hozumi, H.; Miura, Y.; Yang, L.F.; Iwasaki, M.; et al. Extracellular nanovesicles for packaging of CRISPR-Cas9 protein and sgRNA to induce therapeutic exon skipping. Nat. Commun. 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Valuev-Elliston, V.T.; Ivanova, O.N.; Kochetkov, S.N.; Starodubova, E.S.; Bartosch, B.; Isaguliants, M.G. Oxidative Stress during HIV Infection: Mechanisms and Consequences. Oxid. Med. Cell Longev. 2016, 2016, 8910396. [Google Scholar] [CrossRef]
- Hentze, M.W.; Castello, A.; Schwarzl, T.; Preiss, T. A brave new world of RNA-binding proteins. Nat. Rev. Mol. Cell Biol. 2018, 19, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, X.; Wei, M.; Gao, X.; Zhao, L.; Shi, R.; Sun, W.; Duan, Y.; Yang, G.; Yuan, L. In Vitro and in Vivo RNA Inhibition by CD9-HuR Functionalized Exosomes Encapsulated with miRNA or CRISPR/dCas. Nano Lett. 2018, 19, 19–28. [Google Scholar] [CrossRef]
- Hung, M.E.; Leonard, J.N. A platform for actively loading cargo RNA to elucidate limiting steps in EV-mediated delivery. J. Extracell. Vesicles 2016, 5, 31027. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Lyu, P.; Yoo, K.; Yadav, M.K.; Singh, R.; Atala, A.; Lu, B. Engineered extracellular vesicles as versatile ribonucleoprotein delivery vehicles for efficient and safe CRISPR genome editing. J. Extracell. Vesicles 2021, 10, e12076. [Google Scholar] [CrossRef]
- Kebaara, B.W.; Atkin, A.L. Long 3′-UTRs target wild-type mRNAs for nonsense-mediated mRNA decay in Saccharomyces cerevisiae. Nucleic Acids Res. 2009, 37, 2771–2778. [Google Scholar] [CrossRef]
- Li, W.; Maekiniemi, A.; Sato, H.; Osman, C.; Singer, R.H. An improved imaging system that corrects MS2-induced RNA destabilization. Nat. Methods 2022, 19, 1558–1562. [Google Scholar] [CrossRef]
- Zickler, A.M.; Liang, X.; De Luca, M.; Gupta, D.; Corso, G.; Errichelli, L.; Hean, J.; Kamei, N.; Niu, Z.; Zhou, G.; Roudi, S.; Nordin, J.; Castilla-Llorente, V.; Andaloussi, S.E.L. Novel endogenous engineering platform for robust loading and delivery of functional mRNA by extracellular vesicles. BioRxiv 5330, 2023.03.17.533081. Available online: https://www.biorxiv.org/content/10.1101/2023.03.17.533081v1%0Ahttps://www.biorxiv.org/content/10.1101/2023.03.17.533081v1.abstract%0Ahttps://www.biorxiv.org/content/10.1101/2023.03.17.533081v1%0Ahttps://www.biorxiv.org/content/10.1101/2023.03.17.533081v1.
- Kojima, R.; Bojar, D.; Rizzi, G.; Hamri, G.C.-E.; El-Baba, M.D.; Saxena, P.; Ausländer, S.; Tan, K.R.; Fussenegger, M. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson’s disease treatment. Nat. Commun. 2018, 9, 1305. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Zhang, X.; Xie, F.; Xu, B.; Xie, P.; Yang, T.; Shi, Q.; Zhang, C.-Y.; Zhang, Y.; Chen, J.; et al. An engineered exosome for delivering sgRNA:Cas9 ribonucleoprotein complex and genome editing in recipient cells. Biomater. Sci. 2020, 8, 2966–2976. [Google Scholar] [CrossRef]
- Osteikoetxea, X.; Silva, A.; Lázaro-Ibáñez, E.; Salmond, N.; Shatnyeva, O.; Stein, J.; Schick, J.; Wren, S.; Lindgren, J.; Firth, M.; et al. Engineered Cas9 extracellular vesicles as a novel gene editing tool. J. Extracell. Vesicles 2022, 11, e12225. [Google Scholar] [CrossRef]
- Huang, L.; Gu, N.; Zhang, X.; Wang, D. Light-Inducible Exosome-Based Vehicle for Endogenous RNA Loading and Delivery to Leukemia Cells. Adv. Funct. Mater. 2019, 29. [Google Scholar] [CrossRef]
- King, A.; Gottlieb, E.; Brooks, D.G.; Murphy, M.P.; Dunaief, J.L. Mitochondria-derived Reactive Oxygen Species Mediate Blue Light–induced Death of Retinal Pigment Epithelial Cells¶. Photochem. Photobiol. 2004, 79, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Kostyushev, D.; Brezgin, S.; Kostyusheva, A.; Zarifyan, D.; Goptar, I.; Chulanov, V. Orthologous CRISPR/Cas9 systems for specific and efficient degradation of covalently closed circular DNA of hepatitis B virus. Cell. Mol. Life Sci. 2019, 76, 1779–1794. [Google Scholar] [CrossRef]
- Kostyushev, D.; Brezgin, S.; Kostyusheva, A.; Ponomareva, N.; Bayurova, E.; Zakirova, N.; Kondrashova, A.; Goptar, I.; Nikiforova, A.; Sudina, A.; et al. Transient and tunable CRISPRa regulation of APOBEC/AID genes for targeting hepatitis B virus. Mol. Ther. - Nucleic Acids 2023, 32, 478–493. [Google Scholar] [CrossRef] [PubMed]
- Kostyushev, D.; Kostyusheva, A.; Brezgin, S.; Ponomareva, N.; Zakirova, N.F.; Egorshina, A.; Yanvarev, D.V.; Bayurova, E.; Sudina, A.; Goptar, I.; et al. Depleting hepatitis B virus relaxed circular DNA is necessary for resolution of infection by CRISPR-Cas9. Mol. Ther. - Nucleic Acids 2023, 31, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-S.; Li, Q.-C.; Yin, C.-Q.; Xue, W.; Song, C.-Q. Advances in CRISPR/Cas-based Gene Therapy in Human Genetic Diseases. Theranostics 2020, 10, 4374–4382. [Google Scholar] [CrossRef]
- Song, X.; Liu, C.; Wang, N.; Huang, H.; He, S.; Gong, C.; Wei, Y. Delivery of CRISPR/Cas systems for cancer gene therapy and immunotherapy. Adv. Drug Deliv. Rev. 2021, 168, 158–180. [Google Scholar] [CrossRef]
- Yim, N.; Ryu, S.-W.; Choi, K.; Lee, K.R.; Lee, S.; Choi, H.; Kim, J.; Shaker, M.R.; Sun, W.; Park, J.-H.; et al. Exosome engineering for efficient intracellular delivery of soluble proteins using optically reversible protein–protein interaction module. Nat. Commun. 2016, 7, 12277. [Google Scholar] [CrossRef]
- Liang, X.; Gupta, D.; Xie, J.; Van Wonterghem, E.; Van Hoecke, L.; Hean, J.; Niu, Z.; Wiklander, O.; Zheng, W.; Wiklander, R.J.; et al. Multimodal engineering of extracellular vesicles for efficient intracellular protein delivery. bioRxiv 2023. [Google Scholar] [CrossRef]
- Mangeot, P.-E.; Dollet, S.; Girard, M.; Ciancia, C.; Joly, S.; Peschanski, M.; Lotteau, V. Protein Transfer Into Human Cells by VSV-G-induced Nanovesicles. Mol. Ther. 2011, 19, 1656–1666. [Google Scholar] [CrossRef]
- Finkelshtein, D.; Werman, A.; Novick, D.; Barak, S.; Rubinstein, M. LDL receptor and its family members serve as the cellular receptors for vesicular stomatitis virus. Proc. Natl. Acad. Sci. 2013, 110, 7306–7311. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.A.; Coke, L.M.; Richie, C.T.; Fortuno, L.V.; Park, A.Y.; Harvey, B.K. Gesicle-Mediated Delivery of CRISPR/Cas9 Ribonucleoprotein Complex for Inactivating the HIV Provirus. Mol. Ther. 2019, 27, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Kuate, S.; Stahl-Hennig, C.; Stoiber, H.; Nchinda, G.; Floto, A.; Franz, M.; Sauermann, U.; Bredl, S.; Deml, L.; Ignatius, R.; et al. Immunogenicity and efficacy of immunodeficiency virus-like particles pseudotyped with the G protein of vesicular stomatitis virus. Virology 2006, 351, 133–144. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, J.; Kadungure, T.; Beyene, J.; Zhang, H.; Lu, Q. ARMMs as a versatile platform for intracellular delivery of macromolecules. Nat. Commun. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Silva, A.M.; Lázaro-Ibáñez, E.; Gunnarsson, A.; Dhande, A.; Daaboul, G.; Peacock, B.; Osteikoetxea, X.; Salmond, N.; Friis, K.P.; Shatnyeva, O.; et al. Quantification of protein cargo loading into engineered extracellular vesicles at single-vesicle and single-molecule resolution. J. Extracell. Vesicles 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Rädler, J.; Sork, H.; Niu, Z.; Roudi, S.; Bost, J.P.; Görgens, A.; Zhao, Y.; Mamand, D.R.; Liang, X.; et al. Identification of scaffold proteins for improved endogenous engineering of extracellular vesicles. Nat. Commun. 2023, 14, 1–14. [Google Scholar] [CrossRef]
- Dooley, K.; McConnell, R.E.; Xu, K.; Lewis, N.D.; Haupt, S.; Youniss, M.R.; Martin, S.; Sia, C.L.; McCoy, C.; Moniz, R.J.; et al. A versatile platform for generating engineered extracellular vesicles with defined therapeutic properties. Mol. Ther. 2021, 29, 1729–1743. [Google Scholar] [CrossRef]
- Wei, H.; Chen, Q.; Lin, L.; Sha, C.; Li, T.; Liu, Y.; Yin, X.; Xu, Y.; Chen, L.; Gao, W.; et al. Regulation of exosome production and cargo sorting. Int. J. Biol. Sci. 2021, 17, 163–177. [Google Scholar] [CrossRef]
- Whitley, J.A.; Kim, S.; Lou, L.; Ye, C.; Alsaidan, O.A.; Sulejmani, E.; Cai, J.; Desrochers, E.G.; Beharry, Z.; Rickman, C.B.; et al. Encapsulating Cas9 into extracellular vesicles by protein myristoylation. J. Extracell. Vesicles 2022, 11, e12196. [Google Scholar] [CrossRef]
- Cheng, Y.; Schorey, J.S. Targeting soluble proteins to exosomes using a ubiquitin tag. Biotechnol. Bioeng. 2015, 113, 1315–1324. [Google Scholar] [CrossRef]
- Carnino, J.M.; Ni, K.; Jin, Y. Post-translational Modification Regulates Formation and Cargo-Loading of Extracellular Vesicles. Front. Immunol. 2020, 11, 948. [Google Scholar] [CrossRef] [PubMed]
- Ilahibaks, N.F.; Ardisasmita, A.I.; Xie, S.; Gunnarsson, A.; Brealey, J.; Vader, P.; de Jong, O.G.; de Jager, S.; Dekker, N.; Peacock, B.; et al. TOP-EVs: Technology of Protein delivery through Extracellular Vesicles is a versatile platform for intracellular protein delivery. J. Control. Release 2023, 355, 579–592. [Google Scholar] [CrossRef]
- Ferreira, J.V.; Soares, A.d.R.; Ramalho, J.; Carvalho, C.M.; Cardoso, M.H.; Pintado, P.; Carvalho, A.S.; Beck, H.C.; Matthiesen, R.; Zuzarte, M.; et al. LAMP2A regulates the loading of proteins into exosomes. Sci. Adv. 2022, 8, eabm1140. [Google Scholar] [CrossRef] [PubMed]
- Mangeot, P.E.; Risson, V.; Fusil, F.; Marnef, A.; Laurent, E.; Blin, J.; Mournetas, V.; Massouridès, E.; Sohier, T.J.M.; Corbin, A.; et al. Genome editing in primary cells and in vivo using viral-derived Nanoblades loaded with Cas9-sgRNA ribonucleoproteins. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Megede, J.Z.; Chen, M.-C.; Doe, B.; Schaefer, M.; Greer, C.E.; Selby, M.; Otten, G.R.; Barnett, S.W. Increased Expression and Immunogenicity of Sequence-Modified Human Immunodeficiency Virus Type 1 gag Gene. J. Virol. 2000, 74, 2628–2635. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Guerrero, A.; Recalde, M.J.A.; Mangeot, P.E.; Costa, C.; Bernadin, O.; Périan, S.; Fusil, F.; Froment, G.; Martinez-Turtos, A.; Krug, A.; et al. Baboon Envelope Pseudotyped “Nanoblades” Carrying Cas9/gRNA Complexes Allow Efficient Genome Editing in Human T, B, and CD34+ Cells and Knock-in of AAV6-Encoded Donor DNA in CD34+ Cells. Front. Genome Ed. 2021, 3. [Google Scholar] [CrossRef]
- Tiroille, V.; Krug, A.; Bokobza, E.; Kahi, M.; Bulcaen, M.; Ensinck, M.M.; Geurts, M.H.; Hendriks, D.; Vermeulen, F.; Larbret, F.; et al. Nanoblades allow high-level genome editing in murine and human organoids. Mol. Ther. - Nucleic Acids 2023, 33, 57–74. [Google Scholar] [CrossRef]
- Kostyushev, D.; Kostyusheva, A.; Brezgin, S.; Smirnov, V.; Volchkova, E.; Lukashev, A.; Chulanov, V. Gene Editing by Extracellular Vesicles. Int. J. Mol. Sci. 2020, 21, 7362. [Google Scholar] [CrossRef] [PubMed]
- Wan, T.; Zhong, J.; Pan, Q.; Zhou, T.; Ping, Y.; Liu, X. Exosome-mediated delivery of Cas9 ribonucleoprotein complexes for tissue-specific gene therapy of liver diseases. Sci. Adv. 2022, 8, eabp9435. [Google Scholar] [CrossRef]
- Kooijmans, S.A.A.; Stremersch, S.; Braeckmans, K.; De Smedt, S.C.; Hendrix, A.; Wood, M.J.A.; Schiffelers, R.M.; Raemdonck, K.; Vader, P. Electroporation-induced siRNA precipitation obscures the efficiency of siRNA loading into extracellular vesicles. J. Control. Release 2013, 172, 229–238. [Google Scholar] [CrossRef]
- Johnsen, K.B.; Gudbergsson, J.M.; Skov, M.N.; Christiansen, G.; Gurevich, L.; Moos, T.; Duroux, M. Evaluation of electroporation-induced adverse effects on adipose-derived stem cell exosomes. Cytotechnology 2016, 68, 2125–2138. [Google Scholar] [CrossRef]
- Kimiz-Gebologlu, I.; Oncel, S.S. Exosomes: Large-scale production, isolation, drug loading efficiency, and biodistribution and uptake. J. Control. Release 2022, 347, 533–543. [Google Scholar] [CrossRef]
- Hood, J.L.; Scott, M.J.; Wickline, S.A. Maximizing exosome colloidal stability following electroporation. Anal. Biochem. 2014, 448, 41–49. [Google Scholar] [CrossRef]
- Lennaárd, A.J.; Mamand, D.R.; Wiklander, R.J.; EL Andaloussi, S.; Wiklander, O.P.B. Optimised Electroporation for Loading of Extracellular Vesicles with Doxorubicin. Pharmaceutics 2021, 14, 38. [Google Scholar] [CrossRef]
- Peng, H.; Ji, W.; Zhao, R.; Yang, J.; Lu, Z.; Li, Y.; Zhang, X. Exosome: a significant nano-scale drug delivery carrier. J. Mater. Chem. B 2020, 8, 7591–7608. [Google Scholar] [CrossRef]
- Hettich, B.F.; Bader, J.J.; Leroux, J. Encapsulation of Hydrophilic Compounds in Small Extracellular Vesicles: Loading Capacity and Impact on Vesicle Functions. Adv. Heal. Mater. 2021, 11, 2100047. [Google Scholar] [CrossRef]
- Joshi, B.; Ortiz, D.; Zuhorn, I. Converting extracellular vesicles into nanomedicine: loading and unloading of cargo. Mater. Today Nano 2021, 16, 100148. [Google Scholar] [CrossRef]
- Zhang, D.; Lee, H.; Zhu, Z.; Minhas, J.K.; Jin, Y. Enrichment of selective miRNAs in exosomes and delivery of exosomal miRNAs in vitro and in vivo. Am. J. Physiol. Cell. Mol. Physiol. 2017, 312, L110–L121. [Google Scholar] [CrossRef]
- Lepock, J.R.; Frey, H.E.; Ritchie, K.P. Protein denaturation in intact hepatocytes and isolated cellular organelles during heat shock. J. Cell Biol. 1993, 122, 1267–1276. [Google Scholar] [CrossRef]
- Oshchepkova, A.; Zenkova, M.; Vlassov, V. Extracellular Vesicles for Therapeutic Nucleic Acid Delivery: Loading Strategies and Challenges. Int. J. Mol. Sci. 2023, 24, 7287. [Google Scholar] [CrossRef]
- Cao, E.; Chen, Y.; Cui, Z.; Foster, P.R. Effect of freezing and thawing rates on denaturation of proteins in aqueous solutions. Biotechnol. Bioeng. 2003, 82, 684–690. [Google Scholar] [CrossRef]
- Xi, X.-M.; Chen, M.; Xia, S.-J.; Lu, R. Drug loading techniques for exosome-based drug delivery systems. An International Journal of Pharmaceutical Sciences 2021, 76, 61–67. [Google Scholar] [CrossRef]
- Danilushkina, A.A.; Emene, C.C.; Barlev, N.A.; Gomzikova, M.O. Strategies for Engineering of Extracellular Vesicles. Int. J. Mol. Sci. 2023, 24, 13247. [Google Scholar] [CrossRef]
- de Abreu, R.C.; Ramos, C.V.; Becher, C.; Lino, M.; Jesus, C.; Martins, P.A.d.C.; Martins, P.A.T.; Moreno, M.J.; Fernandes, H.; Ferreira, L. Exogenous loading of miRNAs into small extracellular vesicles. J. Extracell. Vesicles 2021, 10, e12111. [Google Scholar] [CrossRef]
- Roerig, J.; Schulz-Siegmund, M. Standardization Approaches for Extracellular Vesicle Loading with Oligonucleotides and Biologics. Small 2023, 19, e2301763. [Google Scholar] [CrossRef]
- Nordin, J.Z. Transfection reagents affect Extracellular Vesicle cargo transfer to recipient cells: The importance of appropriate controls in EV research. J. Extracell. Vesicles 2022, 11. [Google Scholar] [CrossRef]
- Komuro, H.; Aminova, S.; Lauro, K.; Harada, M. Advances of engineered extracellular vesicles-based therapeutics strategy. Sci. Technol. Adv. Mater. 2022, 23, 655–681. [Google Scholar] [CrossRef]
- Fuhrmann, G.; Serio, A.; Mazo, M.; Nair, R.; Stevens, M.M. Active loading into extracellular vesicles significantly improves the cellular uptake and photodynamic effect of porphyrins. J. Control. Release 2015, 205, 35–44. [Google Scholar] [CrossRef]
- Mayer, L.; Bally, M.; Cullis, P. Uptake of adriamycin into large unilamellar vesicles in response to a pH gradient. Biochim. et Biophys. Acta (BBA) - Biomembr. 1986, 857, 123–126. [Google Scholar] [CrossRef]
- Jeyaram, A.; Lamichhane, T.N.; Wang, S.; Zou, L.; Dahal, E.; Kronstadt, S.M.; Levy, D.; Parajuli, B.; Knudsen, D.R.; Chao, W.; et al. Enhanced Loading of Functional miRNA Cargo via pH Gradient Modification of Extracellular Vesicles. Mol. Ther. 2019, 28, 975–985. [Google Scholar] [CrossRef]
- Roerig, J.; Mitrach, F.; Schmid, M.; Hause, G.; Hacker, M.C.; Wölk, C.; Schulz-Siegmund, M. Synergistic siRNA Loading of Extracellular Vesicles Enables Functional Delivery into Cells. Small Methods 2022, 6, e2201001. [Google Scholar] [CrossRef]
- Liu, P.; Chen, G.; Zhang, J. A Review of Liposomes as a Drug Delivery System: Current Status of Approved Products, Regulatory Environments, and Future Perspectives. Molecules 2022, 27, 1372. [Google Scholar] [CrossRef]
- Sutaria, D.S.; Badawi, M.; Phelps, M.A.; Schmittgen, T.D. Achieving the Promise of Therapeutic Extracellular Vesicles: The Devil is in Details of Therapeutic Loading. Pharm. Res. 2017, 34, 1053–1066. [Google Scholar] [CrossRef]
- Biscans, A.; Haraszti, R.A.; Echeverria, D.; Miller, R.; Didiot, M.-C.; Nikan, M.; Roux, L.; Aronin, N.; Khvorova, A. Hydrophobicity of Lipid-Conjugated siRNAs Predicts Productive Loading to Small Extracellular Vesicles. Mol. Ther. 2018, 26, 1520–1528. [Google Scholar] [CrossRef]
- Haraszti, R.A.; Miller, R.; Didiot, M.-C.; Biscans, A.; Alterman, J.F.; Hassler, M.R.; Roux, L.; Echeverria, D.; Sapp, E.; DiFiglia, M.; et al. Optimized Cholesterol-siRNA Chemistry Improves Productive Loading onto Extracellular Vesicles. Mol. Ther. 2018, 26, 1973–1982. [Google Scholar] [CrossRef]
- Didiot, M.-C.; Hall, L.M.; Coles, A.H.; A Haraszti, R.; Godinho, B.M.; Chase, K.; Sapp, E.; Ly, S.; Alterman, J.F.; Hassler, M.R.; et al. Exosome-mediated Delivery of Hydrophobically Modified siRNA for Huntingtin mRNA Silencing. Mol. Ther. 2016, 24, 1836–1847. [Google Scholar] [CrossRef]
- Tréton, G.; Sayer, C.; Schürz, M.; Jaritsch, M.; Müller, A.; Matea, C.-T.; Stanojlovic, V.; Melo-Benirschke, H.; Be, C.; Krembel, C.; et al. Quantitative and functional characterisation of extracellular vesicles after passive loading with hydrophobic or cholesterol-tagged small molecules. J. Control. Release 2023, 361, 694–716. [Google Scholar] [CrossRef]
- Meng, W.; He, C.; Hao, Y.; Wang, L.; Li, L.; Zhu, G. Prospects and challenges of extracellular vesicle-based drug delivery system: considering cell source. Drug Deliv. 2020, 27, 585–598. [Google Scholar] [CrossRef]
- Piffoux, M.; Volatron, J.; Cherukula, K.; Aubertin, K.; Wilhelm, C.; Silva, A.K.; Gazeau, F. Engineering and loading therapeutic extracellular vesicles for clinical translation: A data reporting frame for comparability. Adv. Drug Deliv. Rev. 2021, 178, 113972. [Google Scholar] [CrossRef]
- O’brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Lamichhane, T.N.; Raiker, R.S.; Jay, S.M. Exogenous DNA Loading into Extracellular Vesicles via Electroporation is Size-Dependent and Enables Limited Gene Delivery. Mol. Pharm. 2015, 12, 3650–3657. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Liu, Q.; Li, D.; Pan, X.; Liang, Y. Targeted therapy using engineered extracellular vesicles: principles and strategies for membrane modification. J. Nanobiotechnology 2023, 21, 1–28. [Google Scholar] [CrossRef]
- Wang, J.-H.; Forterre, A.V.; Zhao, J.; Frimannsson, D.O.; Delcayre, A.; Antes, T.J.; Efron, B.; Jeffrey, S.S.; Pegram, M.D.; Matin, A.C. Anti-HER2 scFv-Directed Extracellular Vesicle-Mediated mRNA-Based Gene Delivery Inhibits Growth of HER2-Positive Human Breast Tumor Xenografts by Prodrug Activation. Mol. Cancer Ther. 2018, 17, 1133–1142. [Google Scholar] [CrossRef]
- Takahashi, Y.; Nishikawa, M.; Shinotsuka, H.; Matsui, Y.; Ohara, S.; Imai, T.; Takakura, Y. Visualization and in vivo tracking of the exosomes of murine melanoma B16-BL6 cells in mice after intravenous injection. J. Biotechnol. 2013, 165, 77–84. [Google Scholar] [CrossRef]
- Liang, X.; Niu, Z.; Galli, V.; Howe, N.; Zhao, Y.; Wiklander, O.P.B.; Zheng, W.; Wiklander, R.J.; Corso, G.; Davies, C.; et al. Extracellular vesicles engineered to bind albumin demonstrate extended circulation time and lymph node accumulation in mouse models. J. Extracell. Vesicles 2022, 11, e12248. [Google Scholar] [CrossRef]
- Zheng, W.; He, R.; Liang, X.; Roudi, S.; Bost, J.; Coly, P.; van Niel, G.; Andaloussi, S.E.L. Cell-specific targeting of extracellular vesicles through engineering the glycocalyx. J. Extracell. Vesicles 2022, 11, e12290. [Google Scholar] [CrossRef]
- Gao, X.; Ran, N.; Dong, X.; Zuo, B.; Yang, R.; Zhou, Q.; Moulton, H.M.; Seow, Y.; Yin, H. Anchor peptide captures, targets, and loads exosomes of diverse origins for diagnostics and therapy. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Tian, B.; Liu, J.; Yang, L.; Zeng, L.; Chen, T.; Hong, A.; Wang, X. Nucleolin-targeted Extracellular Vesicles as a Versatile Platform for Biologics Delivery to Breast Cancer. Theranostics 2017, 7, 1360–1372. [Google Scholar] [CrossRef]
- Pi, F.; Binzel, D.W.; Lee, T.J.; Li, Z.; Sun, M.; Rychahou, P.; Li, H.; Haque, F.; Wang, S.; Croce, C.M.; et al. Nanoparticle orientation to control RNA loading and ligand display on extracellular vesicles for cancer regression. Nat. Nanotechnol. 2017, 13, 82–89. [Google Scholar] [CrossRef]
- Zou, J.; Shi, M.; Liu, X.; Jin, C.; Xing, X.-J.; Qiu, L.; Tan, W. Aptamer-Functionalized Exosomes: Elucidating the Cellular Uptake Mechanism and the Potential for Cancer-Targeted Chemotherapy. Anal. Chem. 2019, 91, 2425–2430. [Google Scholar] [CrossRef]
- Kooijmans, S.A.A.; Aleza, C.G.; Roffler, S.R.; van Solinge, W.W.; Vader, P.; Schiffelers, R.M. Display of GPI-anchored anti-EGFR nanobodies on extracellular vesicles promotes tumour cell targeting. J. Extracell. Vesicles 2016, 5, 31053. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Zhang, H.-X.; He, C.-P.; Fan, S.; Zhu, Y.-L.; Qi, C.; Huang, N.-P.; Xiao, Z.-D.; Lu, Z.-H.; Tannous, B.A.; et al. Surface functionalized exosomes as targeted drug delivery vehicles for cerebral ischemia therapy. Biomaterials 2017, 150, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Zhang, B.; Ocansey, D.K.W.; Xu, W.; Qian, H. Extracellular vesicles: A bright star of nanomedicine. Biomaterials 2020, 269, 120467. [Google Scholar] [CrossRef] [PubMed]
- Hung, M.E.; Leonard, J.N. Stabilization of Exosome-targeting Peptides via Engineered Glycosylation. J. Biol. Chem. 2015, 290, 8166–8172. [Google Scholar] [CrossRef]
- Witwer, K.W. On your MARCKS, get set, deliver: Engineering extracellular vesicles. Mol. Ther. 2021, 29, 1664–1665. [Google Scholar] [CrossRef]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular vesicles for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156. [Google Scholar] [CrossRef]
Figure 1.
Biogenesis of extracellular vesicles. Exosome biogenesis: 1.1 Early endosome forms from cell membrane invagination. 1.2 During the maturation process, membrane invaginations form on endosomal membrane, which subsequently bud into endosomal lumen, becoming intraluminal vesicles. 1.3 Multivesicular body fuses with cellular membrane, releasing exosomes into the extracellular space. Ectosome biogenesis: 2.1 Ectosome biogenesis happens through direct membrane budding and fission. Migrasome biogenesis: 3.1 Migrasomes form on retraction fibers during cellular migration due to interactions with extracellular matrix. Apoptotic bodies biogenesis: 4.1 Apoptotic bodies from fragmented cellular membrane during apoptosis. Exopher biogenesis: Exopher forms by cell membrane budding. For a period of time it remains connected to the cell with a nanotube, but is subsequently disconnected from the cell. 1.1 Early endosomes are formed through the invagination of the cell membrane. 1.2 During the maturation process, membrane invaginations occur on the endosomal membrane, which subsequently bud into the endosomal lumen, resulting in the formation of intraluminal vesicles. 1.3 Multivesicular bodies fuse with the cellular membrane, releasing exosomes into the extracellular space. Ectosome Biogenesis: 2.1 Ectosomes are generated through direct membrane budding and fission. Migrasome Biogenesis: 3.1 Migrasomes form on retraction fibers during cellular migration as a result of interactions with the extracellular matrix. Apoptotic Body Biogenesis: 4.1 Apoptotic bodies arise from the fragmentation of the cellular membrane during apoptosis. Exopher Biogenesis: Exophers are formed by the budding of the cell membrane. For a period, they remain connected to the cell via a nanotube but are subsequently detached from the cell.
Figure 1.
Biogenesis of extracellular vesicles. Exosome biogenesis: 1.1 Early endosome forms from cell membrane invagination. 1.2 During the maturation process, membrane invaginations form on endosomal membrane, which subsequently bud into endosomal lumen, becoming intraluminal vesicles. 1.3 Multivesicular body fuses with cellular membrane, releasing exosomes into the extracellular space. Ectosome biogenesis: 2.1 Ectosome biogenesis happens through direct membrane budding and fission. Migrasome biogenesis: 3.1 Migrasomes form on retraction fibers during cellular migration due to interactions with extracellular matrix. Apoptotic bodies biogenesis: 4.1 Apoptotic bodies from fragmented cellular membrane during apoptosis. Exopher biogenesis: Exopher forms by cell membrane budding. For a period of time it remains connected to the cell with a nanotube, but is subsequently disconnected from the cell. 1.1 Early endosomes are formed through the invagination of the cell membrane. 1.2 During the maturation process, membrane invaginations occur on the endosomal membrane, which subsequently bud into the endosomal lumen, resulting in the formation of intraluminal vesicles. 1.3 Multivesicular bodies fuse with the cellular membrane, releasing exosomes into the extracellular space. Ectosome Biogenesis: 2.1 Ectosomes are generated through direct membrane budding and fission. Migrasome Biogenesis: 3.1 Migrasomes form on retraction fibers during cellular migration as a result of interactions with the extracellular matrix. Apoptotic Body Biogenesis: 4.1 Apoptotic bodies arise from the fragmentation of the cellular membrane during apoptosis. Exopher Biogenesis: Exophers are formed by the budding of the cell membrane. For a period, they remain connected to the cell via a nanotube but are subsequently detached from the cell.
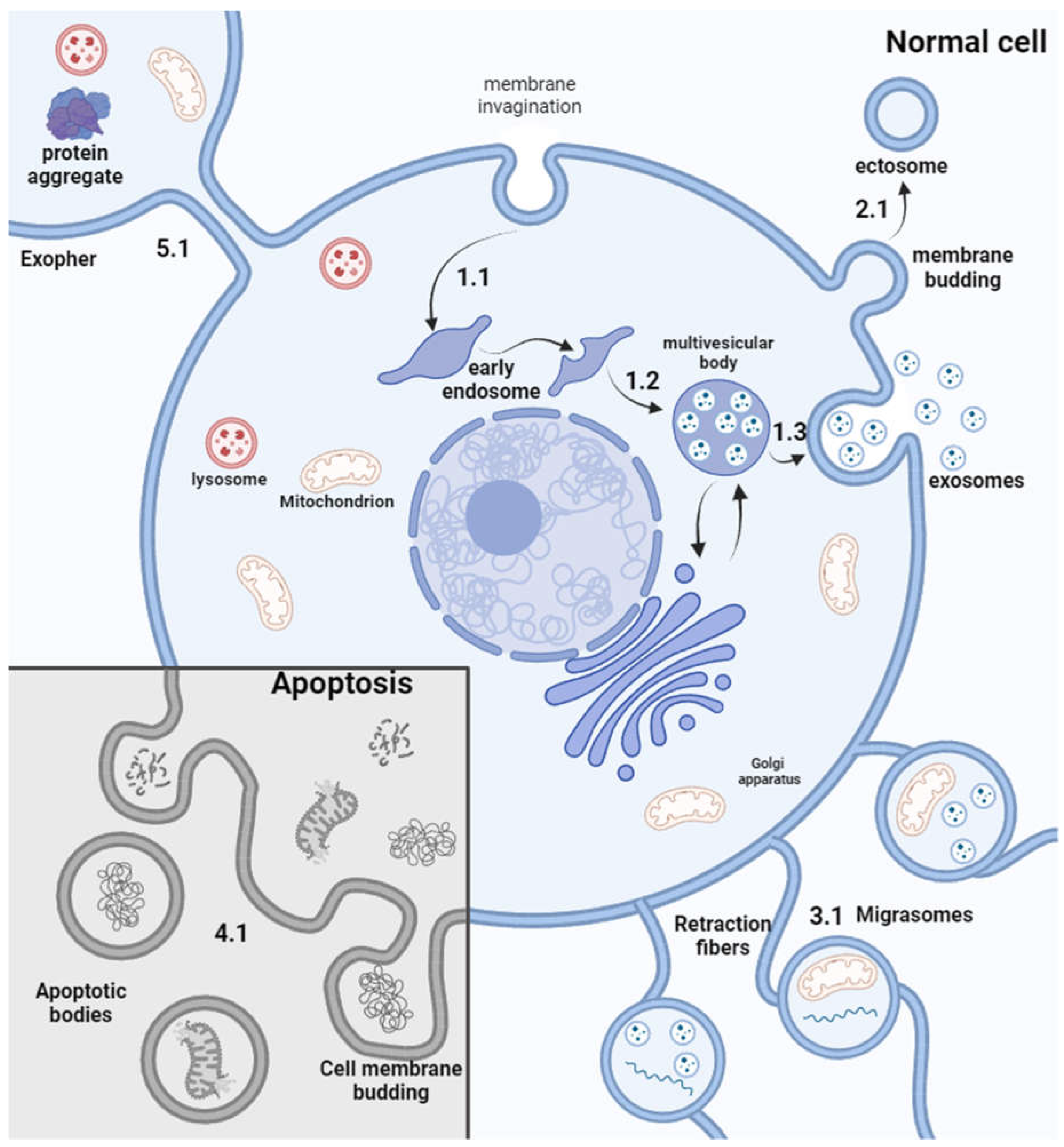
Figure 2.
Ultracentrifugation-based EV purificaiton methods. A. Differential ultracentrifugation begins with several pre-centrifugation steps to remove cells, cell debris, and large non-exosomal vesicles. Subsequently, extracellular vesicles (EVs) are pelleted using an ultracentrifuge. The supernatant, which contains contaminants, is discarded, and the EVs are resuspended in an appropriate buffer. B. Density gradient ultracentrifugation also involves multiple pre-centrifugation steps to eliminate large contaminants. Following pre-centrifugation, the sample is applied to the density gradient, and ultracentrifugation is conducted. The fraction containing the desired EV population is then collected.
Figure 2.
Ultracentrifugation-based EV purificaiton methods. A. Differential ultracentrifugation begins with several pre-centrifugation steps to remove cells, cell debris, and large non-exosomal vesicles. Subsequently, extracellular vesicles (EVs) are pelleted using an ultracentrifuge. The supernatant, which contains contaminants, is discarded, and the EVs are resuspended in an appropriate buffer. B. Density gradient ultracentrifugation also involves multiple pre-centrifugation steps to eliminate large contaminants. Following pre-centrifugation, the sample is applied to the density gradient, and ultracentrifugation is conducted. The fraction containing the desired EV population is then collected.
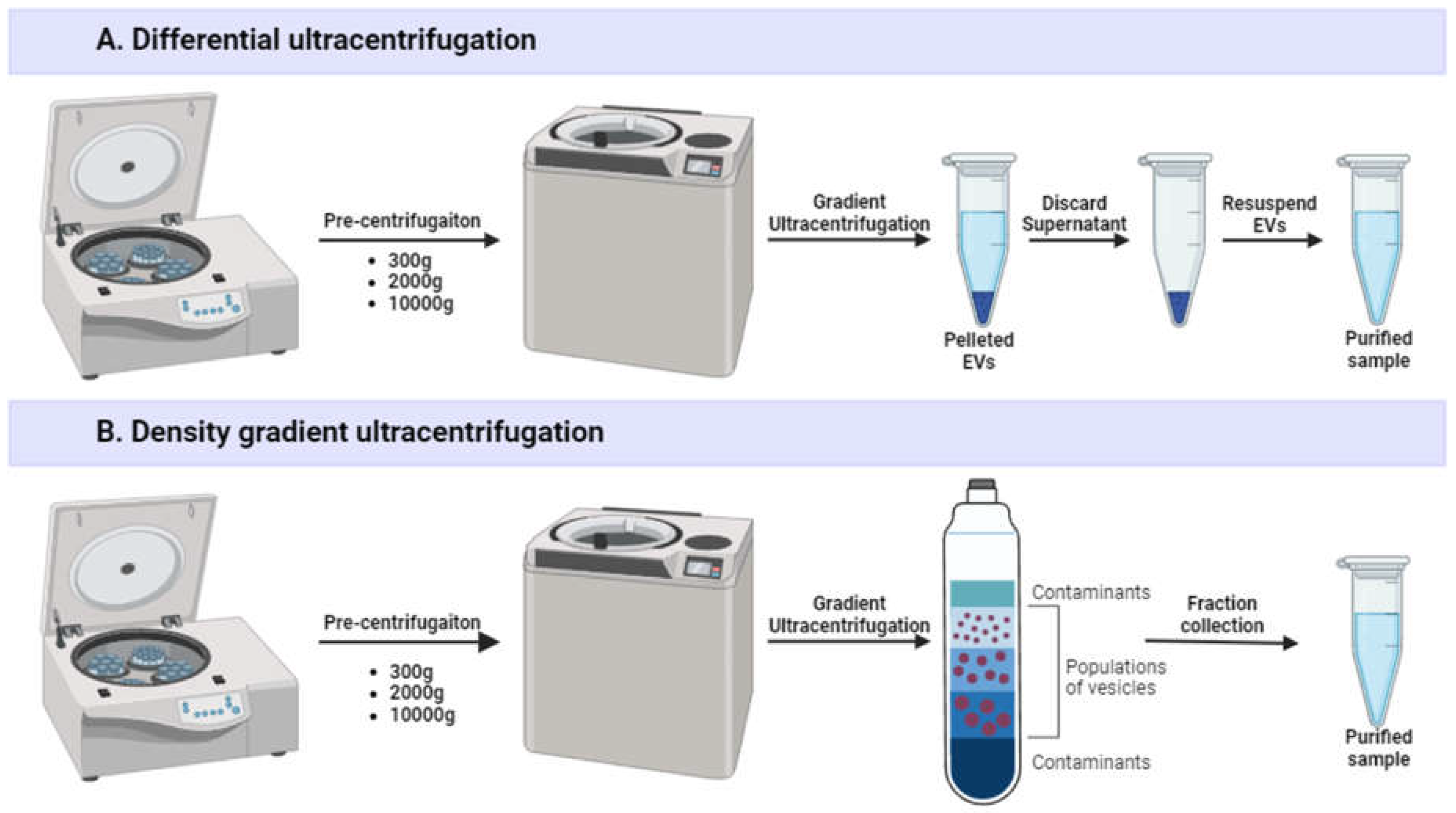
Figure 3.
Ultrafiltration-based EV purification methods. A. During ultrafiltration, pressure forces particles through the filter membrane. Smaller particles permeate the membrane and are discarded, while EVs cannot permeate the membrane and remain in the retentate. However, the inability of larger particles to permeate the membrane leads to the formation of a cake, which hinders the passage of residual smaller molecules through the membrane, resulting in reduced purification efficiency. B. During asymmetric depth filtration, pressure forces the sample through a filter membrane characterized by asymmetric irregular pores. These irregular pores allow particles to enter the membrane, but their movement is impeded based on size. Smaller particles can move further or even pass through the membrane, while larger particles become entrapped and can be eluted later. C. Tangential flow filtration is based on the continuous cyclic flow of particles through a closed system that incorporates a filter membrane into the wall of a filter cartridge. The pressure in the system drives the sample through the membrane, while the feed flow prevents cake formation.
Figure 3.
Ultrafiltration-based EV purification methods. A. During ultrafiltration, pressure forces particles through the filter membrane. Smaller particles permeate the membrane and are discarded, while EVs cannot permeate the membrane and remain in the retentate. However, the inability of larger particles to permeate the membrane leads to the formation of a cake, which hinders the passage of residual smaller molecules through the membrane, resulting in reduced purification efficiency. B. During asymmetric depth filtration, pressure forces the sample through a filter membrane characterized by asymmetric irregular pores. These irregular pores allow particles to enter the membrane, but their movement is impeded based on size. Smaller particles can move further or even pass through the membrane, while larger particles become entrapped and can be eluted later. C. Tangential flow filtration is based on the continuous cyclic flow of particles through a closed system that incorporates a filter membrane into the wall of a filter cartridge. The pressure in the system drives the sample through the membrane, while the feed flow prevents cake formation.
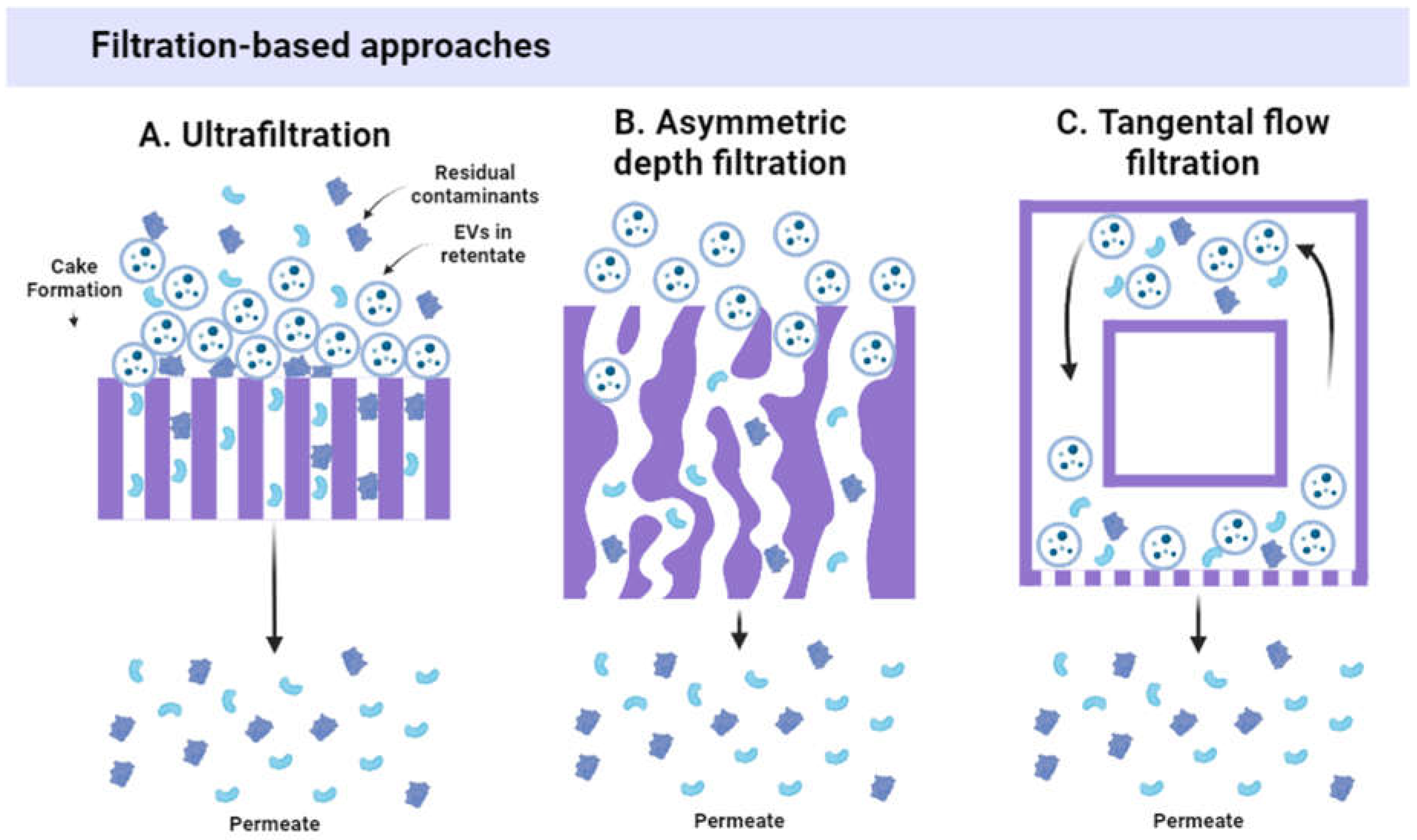
Figure 4.
Precipitation and affinity-based EV purification methods. A. Addition of cationic polymer induces EV aggregation. The sample is centrifuged, which results in EV pelleting and some of the contaminants residing in supernatant. Supernatant is subsequently removed and EVs are resuspended in suitable buffer. B. During affinity purificaiton, antibodies against EV markers conjugated to the magnetic beads are added to the sample. After the binding of antibodies, the beads are pulled down by magnetic force. The supernatant is replaced by the elution buffer, resulting in the release of the EVs. Afterward, the sample is collected.
Figure 4.
Precipitation and affinity-based EV purification methods. A. Addition of cationic polymer induces EV aggregation. The sample is centrifuged, which results in EV pelleting and some of the contaminants residing in supernatant. Supernatant is subsequently removed and EVs are resuspended in suitable buffer. B. During affinity purificaiton, antibodies against EV markers conjugated to the magnetic beads are added to the sample. After the binding of antibodies, the beads are pulled down by magnetic force. The supernatant is replaced by the elution buffer, resulting in the release of the EVs. Afterward, the sample is collected.
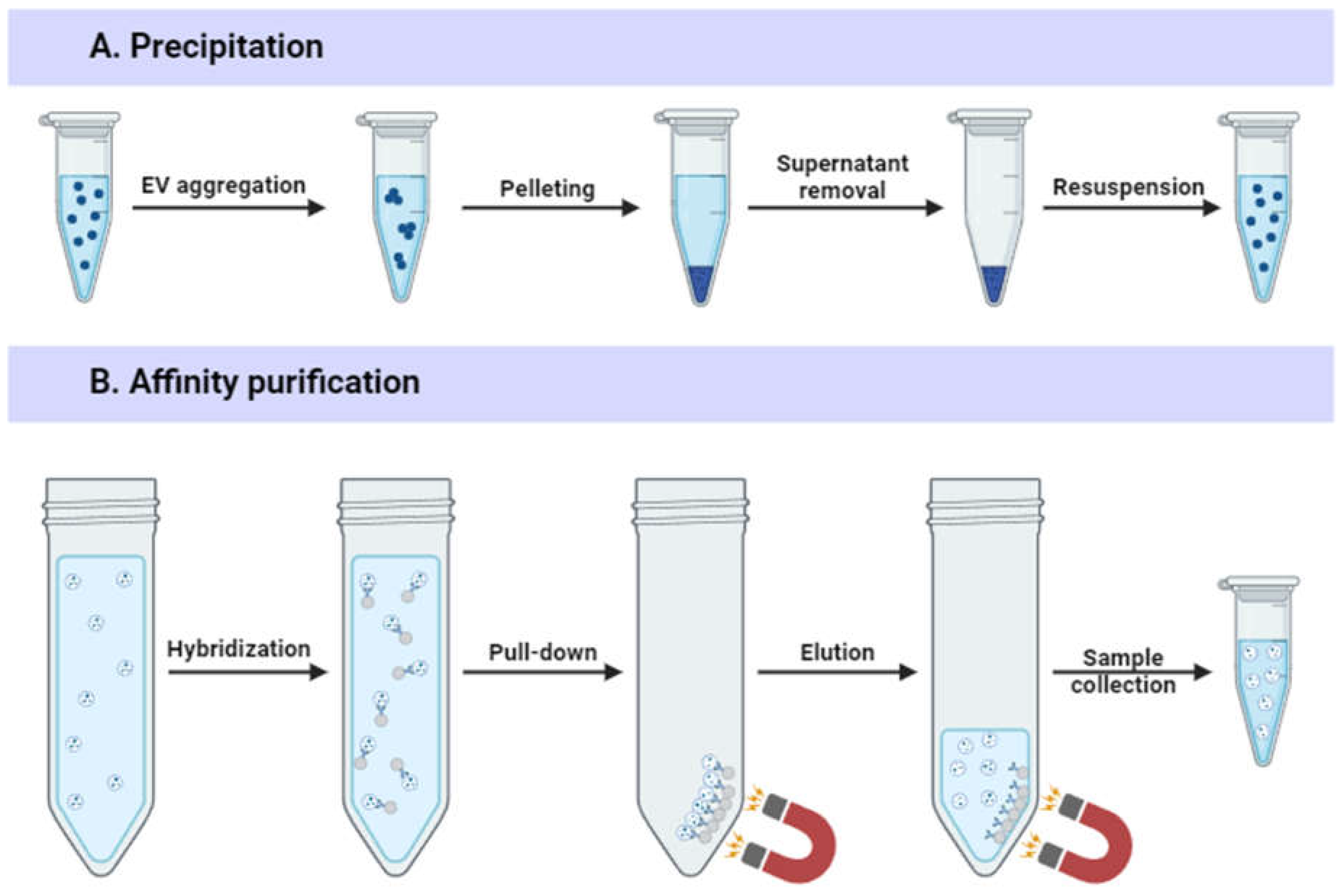
Figure 5.
Endogenous RNA loading methods. A) RNA is delivered into the cell via transfection with RNA or a plasmid encoding the RNA. B) The EV-associated motif in RNA facilitates the binding of EV-associated RNA-binding proteins (RBPs). C) EV-associated RBPs assist in the sorting of RNA into extracellular vesicles (EVs). D) A chimeric construct consisting of an RBP and an EV membrane protein (MP) binds to the RBP-binding site in RNA. E) The RBP-MP complex bound to RNA is sorted to the EV membrane. F) A chimeric protein consisting of myristoylated FKBP12 and GAG binds to the Psi signal in RNA. G) Due to myristoylation, the RNA-bound construct is sorted into EVs. H) A ribozyme encoded in the RNA induces self-cleavage and release of the RNA. I) MCP fused to CRY2 binds to the MSB site in the RNA. J) Blue light induces dimerization of CRY2 with palmitoylated CIBN, which is enriched on the cell membrane. K) The dimer bound to RNA is sorted into the EV. L) In the absence of blue light, the dimer disassembles.
Figure 5.
Endogenous RNA loading methods. A) RNA is delivered into the cell via transfection with RNA or a plasmid encoding the RNA. B) The EV-associated motif in RNA facilitates the binding of EV-associated RNA-binding proteins (RBPs). C) EV-associated RBPs assist in the sorting of RNA into extracellular vesicles (EVs). D) A chimeric construct consisting of an RBP and an EV membrane protein (MP) binds to the RBP-binding site in RNA. E) The RBP-MP complex bound to RNA is sorted to the EV membrane. F) A chimeric protein consisting of myristoylated FKBP12 and GAG binds to the Psi signal in RNA. G) Due to myristoylation, the RNA-bound construct is sorted into EVs. H) A ribozyme encoded in the RNA induces self-cleavage and release of the RNA. I) MCP fused to CRY2 binds to the MSB site in the RNA. J) Blue light induces dimerization of CRY2 with palmitoylated CIBN, which is enriched on the cell membrane. K) The dimer bound to RNA is sorted into the EV. L) In the absence of blue light, the dimer disassembles.

Figure 6.
Endogenous protein loading methods. A) The protein of interest (POI) is fused with CRY2. Upon exposure to blue light, the POI-CRY2 complex dimerizes with CIBN, which is fused to an EV membrane protein (MP). B) In the absence of blue light, the dimer disassembles, releasing the protein into the EV lumen. C) The chimeric protein, consisting of the POI, a self-cleaving intein, and the MP, is sorted into the EV. D) The intein cleaves the construct, releasing the POI into the EV lumen. E) Post-translational modifications (PTMs), such as myristoylation or palmitoylation, facilitate the sorting of the protein into EVs. F) Rapamycin induces dimerization of SpCas9-FRB with FKBP-VSV-g. Due to VSV-g, the construct is sorted into the EV. G) The POI, fused to a WW signal, binds to ARRDC1, leading to their sorting into ARRDC1-mediated microvesicles.
Figure 6.
Endogenous protein loading methods. A) The protein of interest (POI) is fused with CRY2. Upon exposure to blue light, the POI-CRY2 complex dimerizes with CIBN, which is fused to an EV membrane protein (MP). B) In the absence of blue light, the dimer disassembles, releasing the protein into the EV lumen. C) The chimeric protein, consisting of the POI, a self-cleaving intein, and the MP, is sorted into the EV. D) The intein cleaves the construct, releasing the POI into the EV lumen. E) Post-translational modifications (PTMs), such as myristoylation or palmitoylation, facilitate the sorting of the protein into EVs. F) Rapamycin induces dimerization of SpCas9-FRB with FKBP-VSV-g. Due to VSV-g, the construct is sorted into the EV. G) The POI, fused to a WW signal, binds to ARRDC1, leading to their sorting into ARRDC1-mediated microvesicles.
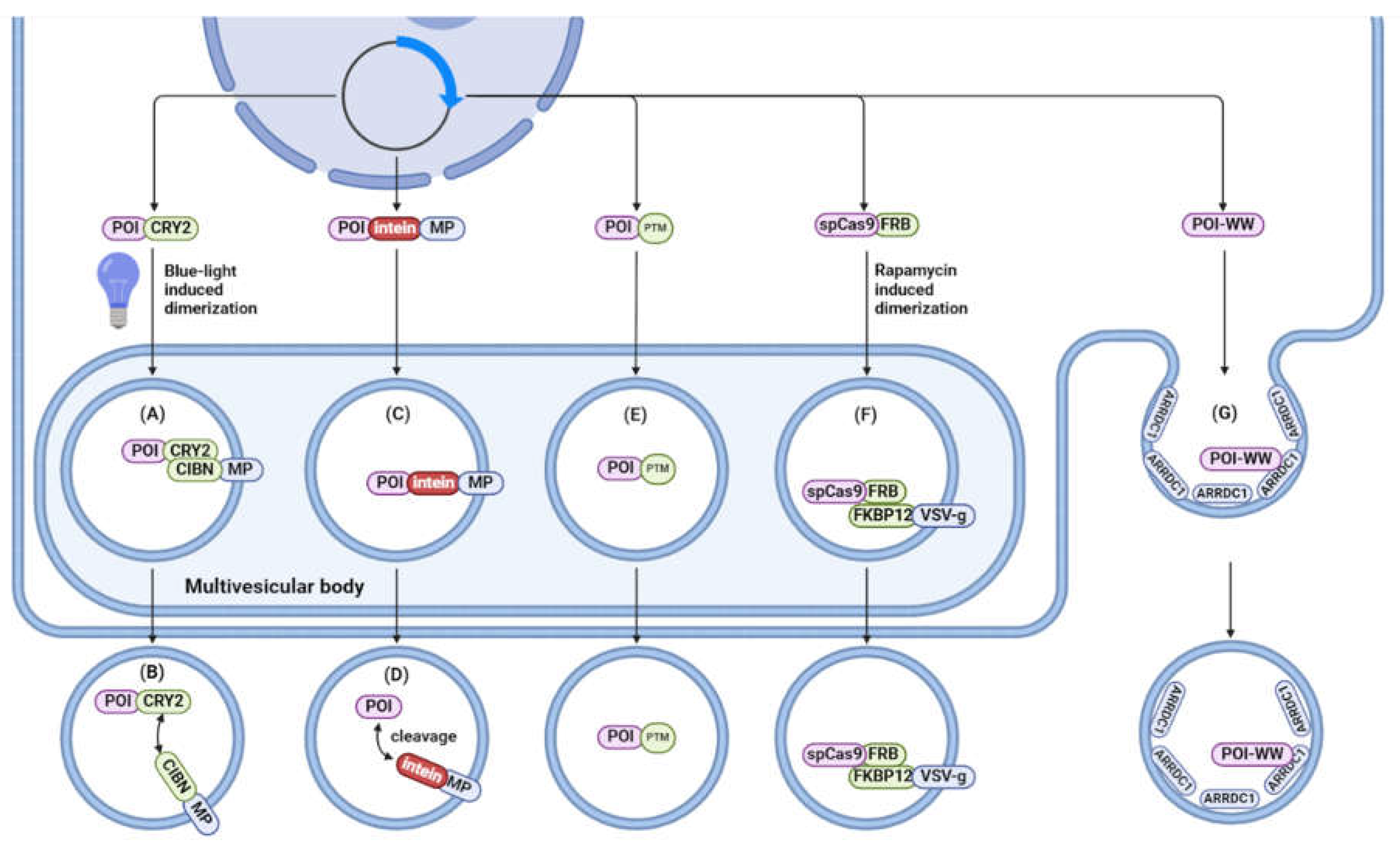
Figure 7.
Exogenous cargo loading methods. A) Several methods, including sonication, electroporation, freeze-thaw cycles, heat shock, chemical permeabilization, and hypotonic dialysis, induce transient pore formation in membranes. Through these pores, cargo from the extravesicular space can enter the vesicle. Subsequently, the membrane reseals, trapping the cargo inside. B) Co-extrusion leads to a transient decrease in membrane continuity, facilitating the entry of cargo into EVs. C) Similar to cells, EVs can be transfected with nucleic acids using cationic transfection reagents. D) Cargo can be conjugated to lipids and subsequently incorporated into the EV membrane; however, the cargo may exist both within the vesicle and on its surface.
Figure 7.
Exogenous cargo loading methods. A) Several methods, including sonication, electroporation, freeze-thaw cycles, heat shock, chemical permeabilization, and hypotonic dialysis, induce transient pore formation in membranes. Through these pores, cargo from the extravesicular space can enter the vesicle. Subsequently, the membrane reseals, trapping the cargo inside. B) Co-extrusion leads to a transient decrease in membrane continuity, facilitating the entry of cargo into EVs. C) Similar to cells, EVs can be transfected with nucleic acids using cationic transfection reagents. D) Cargo can be conjugated to lipids and subsequently incorporated into the EV membrane; however, the cargo may exist both within the vesicle and on its surface.
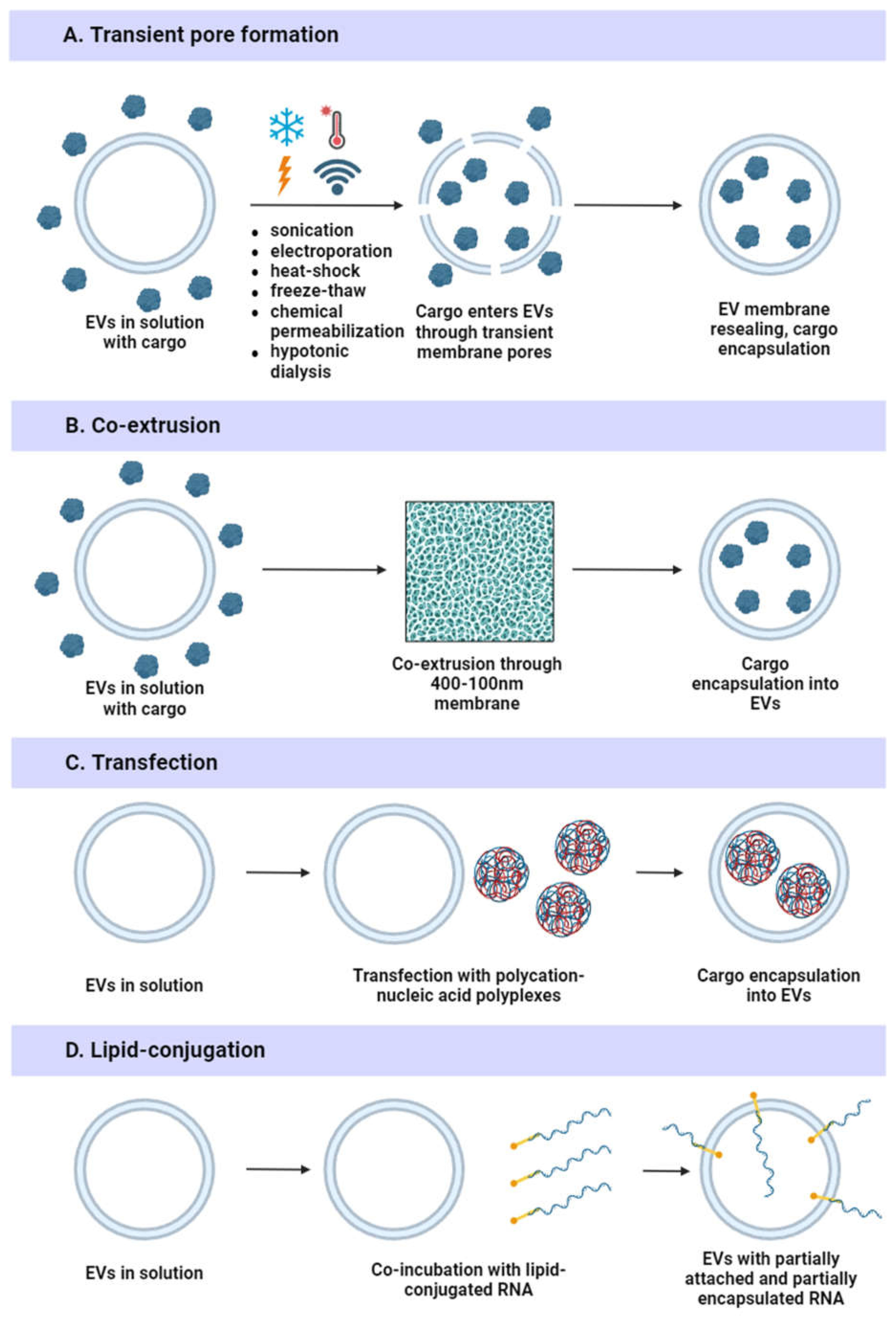
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



