
Submitted:
04 September 2024
Posted:
09 September 2024
You are already at the latest version
Abstract
Background and Objectives: Most of the research of the BRCA1 inactivation is focused on monoallelic germline alterations and loss of heterozygosity in tumour. The aim of this study was to identify characteristic transcriptomic pattern of monoallelic somatic BRCA1 inactivation and estimate its impact to the event-free breast cancer survival prognosis. Materials and Methods: We conducted global transcriptome sequencing of breast cancer tissue samples to identify differentially expressed genes and signalling pathways associated with monoallelic somatic BRCA1 inactivation. The study group involved 36 patient samples categorized based on BRCA1 inactivation status. Subsequently, the differential gene expression and Kaplan-Meier analyses in the groups with and without monoallelic somatic BRCA1 inactivation were performed. Results: Kaplan-Meier analysis showed tendency for longer event-free survival in patients with monoallelic somatic BRCA1 inactivation, suggesting somatic BRCA1 inactivation is favourable prognostic. Differential gene expression analysis followed by the STRING tool enrichment analysis showed significant enrichment of proteins in the extracellular region and extracellular space. Conclusion: In this study we identified transcriptomic profile of differentially expressed genes TPSD1, FABP4, CARTPT and MMP9 as indicative for homologous recombination impaired tumours with tendency for better therapy results.
Keywords:
breast cancer
; transcriptome
; BRCA1 somatic inactivation
; differential gene expression
1. Introduction
Breast cancer is the most common cancer among women worldwide and possesses a growing burden on global public health. It is a diverse disease, both biologically and molecularly, and is associated with environmental and genetic risk factors such as pathogenic variants in the BRCA1 and BRCA2 genes, which contribute to the development of malignancy [1].
The BRCA1 and BRCA2 genes have vital roles in preserving the integrity of the genome and suppressing tumour formation. Both genes facilitate DNA repair through homologous recombination and reactivating replication processes. They are essential for ensuring accurate and efficient restoration of damaged DNA, thereby preventing the accumulation of genetic changes that can lead to cancer development [2].
Germline alterations in the BRCA1 and BRCA2 genes are responsible for an increased susceptibility and higher likelihood of developing breast and ovarian cancers. These genetic variations, which result in the Hereditary Breast and Ovarian Cancer (HBOC) syndrome, account for around 16% from all breast cancer cases [3].
Somatic inactivation of BRCA1/2 genes plays a significant role in the development and progression of breast cancer. While germline BRCA1/2 mutations are well-known risk factors, somatic mutations occurring specifically within the tumour cells which can also lead to the loss of BRCA1/2 function [3]. Somatic BRCA1/2 inactivation results in the disruption of crucial DNA repair mechanisms, including homologous recombination (HR), leading to genomic instability and an increased propensity for the accumulation of additional genetic alterations. These somatic mutations often manifest as large-scale genomic rearrangements, including deletions, insertions, and copy number alterations.
The inactivation of the BRCA1 gene can occur in hereditary cases which is characterized by pathogenic variants in the gene sequence, and sporadic cases. The majority of breast cancer cases are considered sporadic, indicating they are not inherited or linked to genetic syndromes. While the sequence of the BRCA1 gene remains mostly unchanged, there is a specific methylation pattern on the CpG Island region in the BRCA1 promoter. Focal hypermethylation at the tip regions of this CpG island prevents the gene from being transcribed. Alterations in DNA methylation profiles can contribute to the development of sporadic breast cancer without changes in the underlying DNA sequence of the BRCA1 gene [4,5].
The loss of BRCA1/2 function contributes to the impairment of DNA repair and renders cancer cells highly sensitive to DNA-damaging agents, such as platinum-based chemotherapy and poly(ADP-ribose) polymerase (PARP) inhibitors [6,7]. Beside the mutational epigenetic changes in the BRCA1 gene, the breast cancer is also characterized by certain types of somatic mutational signatures, including ”BRCAness” - an HR deficient tumour without pathogenic variants in BRCA1 or BRCA2. Such tumours also are associated with better response to the platinum or PARP inhibitor therapy (Bodily et al., 2020; Polak et al., 2017; Nik-Zainal et al., 2016). The classification of cancer mutational signatures includes HR deficient cancers in base substitution signature 3 and 8, which are characterised with absence of BRCA1 and BRCA2 functions [8]. Combinations of base substitution, indel and rearrangement mutational signatures is proposed as predictive biomarkers of HR deficiency for responsiveness to cisplatin and PARP inhibitors treatment than BRCA1/2 mutations or promoter methylation alone [8].
Drawing on the parallels with ovarian cancer, where BRCA (germline and somatic) mutations confer sensitivity to platinum-based chemotherapy and PARP inhibitors [9,10,11], incorporating BRCA status into clinical decision-making could improve outcomes and personalize therapy for breast cancer patients. Previous research by Maksimenko et al. [12] showed improved breast cancer-specific survival rates in triple-negative breast cancer patients with BRCA1 founder mutations compared to non-carriers. Identifying somatic BRCA1/2 inactivation in breast cancer patients can therefore have significant implications for treatment decisions and prognostic assessments. This enables the selection of targeted therapies that exploit the vulnerabilities of BRCA1/2-deficient tumours. Additionally, studying the patterns and consequences of somatic BRCA1/2 inactivation can provide insights into the mechanisms driving tumour progression and aid in developing novel therapeutic strategies and biomarkers for breast cancer patients with these specific alterations.
2. Materials and Methods
Study Group
36 fresh frozen tissue samples isolated from surgery material were selected from the repository of Riga Stradins University, Institute of Oncology (RSU IO). All patient samples had confirmed breast cancer diagnosis and were not treated with systemic treatment before the surgery. Patients underwent surgery and subsequent systemic therapy at P. Stradins Clinical University Hospital from 2016 to 2018. Study group consisted of TNBC, luminal, HER2-positive samples. Patient’s median age at diagnosis was 59 (ranging from 31 to 81). Only patients without germline BRCA1/BRCA2 variants were included. Clinical information (Supplement 1) was collected from medical records, including information on cancer diagnosis, recurrence and survival status which was used for Kaplan-Mayer analysis and log-rank test to assess event-free survival, defined as the time from cancer diagnosis to recurrence with average follow-up 71 months (19-91).
This study was approved by the Central Medical Ethics Committee (Nr.1/18-09-19 (19.09.2018)). The informed consent was signed by all study participants.
MLPA
All tumour samples underwent MLPA testing for BRCA1 gene. DNA was isolated using QIAamp DNA mini kit (Qiagen) according to manufacturer’s protocol. Isolated DNA underwent MLPA analysis following manufacturer’s protocols, ME001 Tumour suppressor probemix 1 (MRC Holland). Samples with BRCA1 gene promoter deletion or methylation were defined as monoallelic somatic inactivation.
RNA-Sequencing
The total RNA was isolated using TRIzol (Life Technologies) reagent followed by Direct-zolTM RNA MiniPrep (Zymo Research) RNA purification according to the manufacturer’s protocol. RNA concentration was measured with Qubit and NanoDrop according to manufacturer’s protocols.
Library preparation: cDNA libraries were constructed using MGIEasy RNA directional library prep set (MGI) according to manufacturer’s protocol followed by NGS sequencing with MGISEQ-200RS High-throughput Sequencing Set (PE100) (MGI).
Bioinformatics and Statistical Analysis
Obtained raw sequenced reads were analysed with CLC genomic workbench to filter raw reads, check sequencing error rate (Q20 and Q30) and CG content check. CLC Genomics Workbench (Qiagen) software was used for high-quality reads alignment to the reference genome; RPKMs (reads per kilobase of exon model per million mapped reads) were calculated for each annotated gene; differentially expressed genes (or transcripts) between two groups were determined.
The Negative binomial generalised Linear model embedded in CLC Genomic Workbench was used to capture differential gene expression (DEGs) between two study groups. Genes after Bonferroni correction (p<0.05) and with max group mean >10 of average expression were included in further analysis.
The STRING (Search Tool for the Retrieval of Interacting Genes/Proteins) analysis is a bioinformatics tool that evaluates and visualizes protein-protein interaction networks. It was used to predict and display interactions between proteins, analysing insights into cellular processes and functional relationships [13].
Statistical analysis was performed using R software, utilizing the ‘survival’ package for Kaplan-Meier analysis and employing the Peto & Peto method for log-rank testing.
3. Results
Based on MLPA results the group of 36 samples was divided in two study groups based on BRCA1 status – with monoallelic somatic BRCA1 inactivation and without inactivation. 16 samples had monoallelic promoter region deletion and one sample showed hypermethylation of BRCA1 promoter region. 19 samples did not reveal deletion or methylation of the BRCA1 promoter region. Based on these results two study groups were formed - “BRCA1-” with monoallelic somatic inactivation and “BRCA1+” with two active BRCA1 gene alleles. The analysis of clinical information (including grade, stage and molecular type of cancer, as well as Ki67 index) did not indicate statistically significant differences between the groups (Supplement 1).
The Kaplan-Mayer analysis indicated tendency for longer event-free survival (p<0.09; HR 5.17, 95% CI 0.60 and 44.3) in the group with the BRCA1 inactivation (Figure 1).
The RNA sequencing data analysis revealed 39 DEGs between the study groups. Among these, 23 genes exhibited upregulation (Table 1) while 16 genes were downregulated (Table 2) in the BRCA1- group. All genes, except 4 are protein coding genes (the rest 4 are lncRNAs or rRNAs, not shown in the tables).
To explore functional association between proteins coded by the identified DEGs, the STRING analysis tool [13] was used. The acquired results provided insights into the molecular processes and pathways potentially associated with the monoallelic somatic BRCA1 gene inactivation in breast cancer (Figure 2).
The STRING tool enrichment analysis shows significant enrichment of proteins in the extracellular region (GO:0005576) and extracellular space (GO:0005615). Key genes implicated in these categories include TPSD1, FABP4, ORM1, ALPI, CARTPT, TRH, CSN3, MMP9 among others.
The STRING tool enrichment analysis results also implies a potential connection or shared molecular pathways between breast cancer and thyroid dysfunction (not shown in the Table 3). This association is important as thyroid dysfunction has been implicated in various physiological and pathological conditions. Genes identified in both pathways are TRH, IRS4, CHGB, and CGA.
4. Discussion
Our study was based on global transcriptome sequencing of breast cancer tissue samples to identify differentially expressed genes and signalling pathways associated with monoallelic somatic BRCA1 inactivation. The Kaplan-Meier analysis to assess the differences in event-free survival between two distinct groups was performed. Notably, the findings revealed tendency for positive effect on event-free survival in the group with monoallelic BRCA1 inactivation. This observation, indicated by a p<0.09, suggests that breast cancer patients with BRCA1 promoter inactivation may experience improved outcomes in terms of event-free survival. Further investigation into these finding could offer valuable insights into the underlying mechanisms driving the disease and would help to develop more targeted and effective therapeutic strategies for breast cancer patients.
The detailed analysis using the STRING database discovered functional associations of DEGs changes in the molecules pathways related to breast cancer with somatic monoallelic BRCA1 inactivation. The enrichment analysis, focused on the extracellular region and extracellular space, has unveiled proteins with potential implications for the tumour microenvironment and intercellular communication in the context of cancer. Among the key genes found in these enriched categories, TPSD1, FABP4, CARTPT, TRH, CSN3, MMP9 and others stand out, suggesting their critical roles in cancer progression.
The extracellular region and extracellular space are one of main components in the tumour microenvironment, contributing significantly to cancer progression and metastasis [14]. Proteins identified in these categories often participate in intricate signalling networks, modulating cell behaviour, angiogenesis, and immune responses within the tumour microenvironment. Furthermore, our previous study on the transcriptome of TNBC tumours revealed that differentially expressed genes (DEGs) were associated with processes such as extracellular matrix organization, collagen fibril organization, and the composition of collagen-containing extracellular matrix [15].
TPSD1 appears to be upregulated in study group with somatic monoallelic BRCA1 inactivation. TPSD1 gene codes for tryptase delta, which is secreted by mast cells. Mast cells (MCs) play a role in extracellular matrix degradation, angiogenesis, and immune responses through the release of various bioactive substances, including tryptases. Kankkunen et al observed a substantial increase in the presence of tryptase-containing MCs in malignant breast carcinomas compared to benign lesions [16]. The density of MCs, along with their release of tryptases, has been correlated with cancer growth, particularly in facilitating angiogenesis [16]. Mice deficient in mast cells, and subsequently, tryptase secretion, exhibit reduced susceptibility to carcinogenic agents [17,18]. Although TPSD1 shows increased expression in group with better event-free survival, its role in the context of BRCA1-deficient tumours requires further exploration.
Fatty Acid Binding Protein 4 (FABP4), also known as adipocyte protein 2 (aP2), is a member of the FABP family, playing a crucial role in lipid metabolism and cellular signalling. FABP4 is primarily expressed in adipocytes and macrophages, where it facilitates the transportation of fatty acids and other lipophilic molecules within cells [19]. While its role in obesity-related metabolic disorders has been extensively studied, emerging evidence suggests its involvement in various cancers, including breast cancer. Recent studies demonstrate that adipose FABP4 promotes obesity-associated breast cancer development, thus suggesting FABP4 as a novel player linking obesity and breast cancer risk [20,21].
In breast cancer, FABP4 expression and function is linked to the tumour microenvironment and cancer progression. Research indicates that FABP4 is often upregulated in breast cancer tissues, promoting aggressive phenotypes. The higher expression of FABP4 has been associated with increased cell proliferation, migration, and invasion, contributing to tumour growth and metastasis. Moreover, FABP4 has been implicated in promoting angiogenesis, a critical process for the establishment and progression of solid tumours, including breast cancer [20,22]
Interestingly, our study group’s observation of lower FABP4 expression in breast cancer patients with monoallelic somatic BRCA1 inactivation suggesting a potential regulatory role of BRCA1 and FABP4 expression. The crosstalk between BRCA1 and FABP4 may involve complex signalling pathways that influence lipid metabolism and tumour progression. This unique molecular profile, characterized by lower FABP4 expression in the context of somatic BRCA1 inactivation, could be associated with a less aggressive tumour phenotype and, consequently, better event-free survival outcomes.
Lower FABP4 expression levels potentially influence response to therapy, including hormone-based treatments. While the exact mechanisms linking FABP4 expression, BRCA1 inactivation, and clinical outcomes require further exploration, these findings open avenues for personalized therapeutic strategies and highlight the importance of understanding the molecular complexity of breast cancer. Recent study by Kast et al, found association of higher BMI and weight gain in adult life were risk factors for postmenopausal breast cancer in BRCA1 variant carriers [23]. The link between BMI, weight gain, and breast cancer risk in BRCA1 carriers emphasizes the importance of lifestyle factors in modifying cancer susceptibility.
CARTPT, known for its role in cocaine- and amphetamine-regulated transcript signalling, has been implicated in breast cancer cell survival and tamoxifen resistance, underscoring its relevance in therapeutic response and tumour behaviour [24].
Research into cocaine- and amphetamine-regulated transcript (CART) unveils its expression in both primary and metastatic breast cancer, appearing as an independent predictor of poor prognosis in oestrogen receptor-positive, lymph node-negative tumours [24]. CART plays a multifaceted role by amplifying the transcriptional activity of oestrogen receptor alpha (ERα) through the mitogen-activated protein kinase (MAPK) pathway in a ligand-independent manner. In various cancer cell lines, CARTPT acts as an oncogene, promoting cellular survival through the activation of the ERK pathway, stimulation of pro-survival molecules, inhibition of apoptosis, and an increase in cyclin D1 levels. Particularly in breast cancer, CART emerges as a safeguard, protecting tumour cells from tamoxifen-induced cell death and underscoring its pivotal role in cancer pathogenesis [24,25].
Study group with somatically inactivated BRCA1 gene showed decreased CARTPT expression, it is conceivable that the compromised DNA repair mechanisms resulting from BRCA1 inactivation may contribute to altered gene expression patterns, including downregulation of CARTPT.
This complexity may offer explanation into Kaplan-Meier plot (Figure 1), revealing that patient group with somatic monoallelic BRCA1 inactivation experience prolonged event-free survival. Notably, half of these patients underwent hormone therapy, either tamoxifen or anastrozole. This observation suggests sensitivity to hormone therapy, potentially contributing to the extended event-free survival observed in this patient subgroup.
Matrix metalloproteinase 9 (MMP9) is a member of the matrix metalloproteinase family, a group of enzymes that play a crucial role in the degradation and remodelling of the extracellular matrix (ECM). The extracellular matrix is a complex network of proteins and carbohydrates that provides structural support to cells and regulates various cellular processes, including cell adhesion, migration, and signalling [26,27,28]. It’s interesting that in our results increased matrix metalloproteinase 9 (MMP9) expression shows tendency for better event-free survival, although MMP9 is typically associated with promoting cancer progression, invasion, and metastasis. However, it is important to recognize that the role of MMP9 in cancer is complex. Several factors may contribute to this observation. MMP9 has both pro-tumorigenic and anti-tumorigenic functions.
While MMP9 is often linked with promoting invasion and metastasis, it can also have beneficial effects, such as influencing the immune response, modulating the tumour microenvironment, and facilitating tissue repair [27,29,30]. There are no studies investigating how somatic inactivation of BRCA1 would be related to increased MMP9 expression, however hypothetical explanation would be that genomic instability resulting from BRCA1 inactivation may induce an inflammatory response within the tumour microenvironment. Inflammatory signals are known to influence MMP9 expression, and this could contribute to increased MMP9 levels.
In the context of BRCA1-related functions, these genes may contribute to the complex regulatory network associated with the BRCA1 pathway, influencing cellular responses, immune modulation, and therapeutic resistance in breast cancer.
The sample size and heterogeneity of molecular subtypes between the study groups may set some limitations to the current study. Further studies are needed for clinical validation of identified transcriptomic profile
5. Conclusions
Kaplan-Meier analysis indicates a favourable impact on event-free survival in patients with somatic monoallelic BRCA1 inactivation, highlighting the potential clinical significance of this subtype for tailored patient care. STRING database analysis identifies alterations in key genes associated with cancer progression within the extracellular microenvironment, with genes like TPSD1 and FABP4 showing differential expression in patients with monoallelic somatic BRCA1 inactivation, potentially contributing to improved outcomes. Additionally, the involvement of genes like CARTPT and MMP9 hints at complex interactions between DNA repair mechanisms and endocrine therapies.
The transcriptomic profile of differentially expressed genes TPSD1, FABP4, CARTPT and MMP9 identified in the group with monoallelic somatic inactivation can be indicative for homologous recombination impaired tumours with tendency for better therapy results.
Author Contributions
Design of the Experiment: Z.D., E.M., E.K. and M.N.M; Data Examination: E.K., M.N.M., N.K. and P.L.; Conducting the Experiment: E.K and N.K.; Data Analysis and Manuscript Writing: E.K., M.N.M., Z.D., E.S., J.G. and A.I.; Guidance and Consultation on the Experimental Design: Z.D., E.M., J.G., E.S., A.I. and P.L.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and study was approved by the Central Medical Ethics Committee (Nr.1/18-09-19 (19.09.2018)).
Informed Consent Statement
The informed consent was signed by all study participants.
Data Availability Statement
The datasets generated and analyzed during the present study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Clinical data of patients involved in the study
| Characteristic | Study population (n=36) | BRCA1 - group (n=17) | BRCA1 + group (n=19) | p-value |
| Age, median (range) | 59.5 (37-81) | 58 (38-81) | 68 (37-76) | |
| T stage, n (%) | 0.16 | |||
| Tis | 1 (2.8) | 0 (0) | 1 (5) | |
| T1 | 10 (27.8) | 4 (24) | 6 (32) | |
| T2 | 21 (58.3) | 13 76) | 8 (42) | |
| T3 | 3 (8.3) | 0 (0) | 3 (16) | |
| T4 | 1 (2.8) | 0 (0) | 1 (5) | |
| N stage, n(%) | 0.24 | |||
| N0 | 26 (72.2) | 13 (76.5) | 13 (68.4) | |
| Nmic | 2 (5.6) | 2 (11.8) | 0 (0) | |
| N1 | 7 (19.4) | 2 (11.8) | 5 (26.3) | |
| N3 | 1 (2.8) | 0 (0) | 1 (5.3) | |
| Clinical stage | n (%) | 0.25 | ||
| 0 | 1 (2.8) | 0 (0) | 1 (5.2) | |
| IA | 9 (25) | 3 (17.6) | 6 (31.8) | |
| IB | 1 (2.8) | 1 (5.9) | 0 (0) | |
| IIA | 14 (38.9) | 10 (58.9) | 4 (21.1) | |
| IIB | 8 (22.1) | 3 (17.6) | 5 (26.3) | |
| IIIA | 1 (2.8) | 0 (0) | 1 (5.2) | |
| IIIB | 1 (2.8) | 0 (0) | 1 (5.2) | |
| IIIC | 1 (2.8) | 0 (0) | 1 (5.2) | |
| Grade | n (%) | 0.74 | ||
| G1 | 5 (13.9) | 3 (17.6) | 2 (10.5) | |
| G2 | 22 (61.1) | 10 (58.8) | 12 (63.2) | |
| G3 | 6 (16.7) | 2 (11.8) | 4 (21.1) | |
| not known | 3 (8.3) | 2 (11.8) | 1 (5.3) | |
| Ki67 index: | n (%) | 0.34 | ||
| <20% | 15 (41.7) | 5 (29.4) | 10 (52.6) | |
| >20% | 15 (41.7) | 9 (52.9) | 6 (31.6) | |
| Unknown | 6 (16.7) | 3 (17.6) | 3 (15.8) | |
| Molecular type: | n (%) | 0.28 | ||
| Luminal A | 10 (27.8) | 3 (17.6) | 7 (36.8) | |
| Luminal B | 13 (36.2) | 6 (35.3) | 7 (36.8) | |
| TNBC | 7 (19.4) | 4 (23.5) | 3 (15.8) | |
| HER2 positive | 3 (8.3) | 3 (17.6) | 0 (0) | |
| Unknown | 3 (8.3) | 1 (5.9) | 2 (10.5) | |
| Adjuvant treatment: | n | |||
| Chemotherapy | 14 | 7 | 7 | |
| Endocrine therapy (Tamoxifen or AI) | 13 | 6 | 7 | |
| Radiotherapy | 26 | 11 | 15 | |
| Trastuzumab | 2 | 2 | 0 | |
References
- Y. Feng et al., “Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis,” Genes Dis, vol. 5, no. 2, pp. 77–106, Jun. 2018. [CrossRef]
- Q. Jiang and R. A. Greenberg, “Deciphering the BRCA1 Tumor Suppressor Network,” Journal of Biological Chemistry, vol. 290, no. 29, pp. 17724–17732, Jul. 2015. [CrossRef]
- B. W. X. Lim et al., “Somatic inactivation of breast cancer predisposition genes in tumors associated with pathogenic germline variants,” JNCI: Journal of the National Cancer Institute, vol. 115, no. 2, pp. 181–189, Feb. 2023. [CrossRef]
- R. D. Nindrea, W. A. Harahap, T. Aryandono, and L. Lazuardi, “Association of BRCA1 Promoter Methylation with Breast Cancer in Asia: A Meta- Analysis.,” Asian Pac J Cancer Prev, vol. 19, no. 4, pp. 885–889, Apr. 2018. [CrossRef]
- G. Arpino et al., “Tumor characteristics and prognosis in familial breast cancer,” BMC Cancer, vol. 16, no. 1, p. 924, Dec. 2016. [CrossRef]
- P. C. Fong et al., “Inhibition of Poly(ADP-Ribose) Polymerase in Tumors from BRCA Mutation Carriers,” New England Journal of Medicine, vol. 361, no. 2, pp. 123–134, Jul. 2009. [CrossRef]
- N. Turner, A. Tutt, and A. Ashworth, “Hallmarks of ‘BRCAness’ in sporadic cancers,” Nat Rev Cancer, vol. 4, no. 10, pp. 814–819, Oct. 2004. [CrossRef]
- S. Nik-Zainal et al., “Landscape of somatic mutations in 560 breast cancer whole-genome sequences,” Nature, vol. 534, no. 7605, pp. 47–54, Jun. 2016. [CrossRef]
- E. M. Walsh et al., “Olaparib Use in Patients With Metastatic Breast Cancer Harboring Somatic BRCA1/2 Mutations or Mutations in Non-BRCA1/2, DNA Damage Repair Genes,” Clin Breast Cancer, vol. 22, no. 4, pp. 319–325, Jun. 2022. [CrossRef]
- J. Ledermann et al., “Olaparib maintenance therapy in patients with platinum-sensitive relapsed serous ovarian cancer: a preplanned retrospective analysis of outcomes by BRCA status in a randomised phase 2 trial,” Lancet Oncol, vol. 15, no. 8, pp. 852–861, Jul. 2014. [CrossRef]
- K. P. Pennington et al., “Germline and Somatic Mutations in Homologous Recombination Genes Predict Platinum Response and Survival in Ovarian, Fallopian Tube, and Peritoneal Carcinomas,” Clinical Cancer Research, vol. 20, no. 3, pp. 764–775, Feb. 2014. [CrossRef]
- J. MAKSIMENKO et al., “Prognostic role of BRCA1 mutation in patients with triple-negative breast cancer,” Oncol Lett, vol. 7, no. 1, pp. 278–284, Jan. 2014. [CrossRef]
- D. Szklarczyk et al., “STRING v11: protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets,” Nucleic Acids Res, vol. 47, no. D1, pp. D607–D613, Jan. 2019. [CrossRef]
- D. F. Quail and J. A. Joyce, “Microenvironmental regulation of tumor progression and metastasis,” Nat Med, vol. 19, no. 11, pp. 1423–1437, Nov. 2013. [CrossRef]
- E. Kuzņecova et al., “Identification of Altered Transcripts and Pathways in Triple Negative Breast Cancer,” Proceedings of the Latvian Academy of Sciences. Section B. Natural, Exact, and Applied Sciences., vol. 77, no. 1, pp. 33–40, Feb. 2023. [CrossRef]
- J.-P. Kankkunen, I. T. Harvima, and A. Naukkarinen, “Quantitative analysis of tryptase and chymase containing mast cells in benign and malignant breast lesions,” Int J Cancer, vol. 72, no. 3, pp. 385–388, Jul. 1997. [CrossRef]
- A. Iamaroon, S. Pongsiriwet, S. Jittidecharaks, K. Pattanaporn, S. Prapayasatok, and S. Wanachantararak, “Increase of mast cells and tumor angiogenesis in oral squamous cell carcinoma,” Journal of Oral Pathology & Medicine, vol. 32, no. 4, pp. 195–199, Apr. 2003. [CrossRef]
- J. Wedemeyer and S. J. Galli, “Decreased susceptibility of mast cell-deficient Kit/Kit mice to the development of 1, 2-dimethylhydrazine-induced intestinal tumors,” Laboratory Investigation, vol. 85, no. 3, pp. 388–396, Mar. 2005. [CrossRef]
- M. Furuhashi, S. Saitoh, K. Shimamoto, and T. Miura, “Fatty Acid-Binding Protein 4 (FABP4): Pathophysiological Insights and Potent Clinical Biomarker of Metabolic and Cardiovascular Diseases,” Clin Med Insights Cardiol, vol. 8s3, p. CMC.S17067, Jan. 2014. [CrossRef]
- S. Liu et al., “FABP4 in obesity-associated carcinogenesis: Novel insights into mechanisms and therapeutic implications,” Front Mol Biosci, vol. 9, Aug. 2022. [CrossRef]
- J. Zeng, E. R. Sauter, and B. Li, “FABP4: A New Player in Obesity-Associated Breast Cancer,” Trends Mol Med, vol. 26, no. 5, pp. 437–440, May 2020. [CrossRef]
- J. Hao et al., “Expression of Adipocyte/Macrophage Fatty Acid–Binding Protein in Tumor-Associated Macrophages Promotes Breast Cancer Progression,” Cancer Res, vol. 78, no. 9, pp. 2343–2355, May 2018. [CrossRef]
- K. Kast et al., “Associations of height, body mass index, and weight gain with breast cancer risk in carriers of a pathogenic variant in BRCA1 or BRCA2: the BRCA1 and BRCA2 Cohort Consortium,” Breast Cancer Research, vol. 25, no. 1, p. 72, Jun. 2023. [CrossRef]
- D. J. Brennan et al., “The cocaine- and amphetamine-regulated transcript mediates ligand-independent activation of ERα, and is an independent prognostic factor in node-negative breast cancer,” Oncogene, vol. 31, no. 30, pp. 3483–3494, Jul. 2012. [CrossRef]
- H. J. Kang, Y. W. Yi, H. J. Kim, Y. B. Hong, Y. S. Seong, and I. Bae, “BRCA1 negatively regulates IGF-1 expression through an estrogen-responsive element-like site,” Cell Death Dis, vol. 3, no. 6, pp. e336–e336, Jun. 2012. [CrossRef]
- R. Roy, J. Yang, and M. A. Moses, “Matrix Metalloproteinases As Novel Biomarker s and Potential Therapeutic Targets in Human Cancer,” Journal of Clinical Oncology, vol. 27, no. 31, pp. 5287–5297, Nov. 2009. [CrossRef]
- M. K. Jena and J. Janjanam, “Role of extracellular matrix in breast cancer development: a brief update,” F1000Res, vol. 7, p. 274, Jun. 2018. [CrossRef]
- K. Dzobo and C. Dandara, “The Extracellular Matrix: Its Composition, Function, Remodeling, and Role in Tumorigenesis,” Biomimetics, vol. 8, no. 2, p. 146, Apr. 2023. [CrossRef]
- M. D. Martin and L. M. Matrisian, “The other side of MMPs: Protective roles in tumor progression,” Cancer and Metastasis Reviews, vol. 26, no. 3–4, p. 717, Dec. 2007. [CrossRef]
- H. Huang, “Matrix Metalloproteinase-9 (MMP-9) as a Cancer Biomarker and MMP-9 Biosensors: Recent Advances,” Sensors, vol. 18, no. 10, p. 3249, Sep. 2018. [CrossRef]
Figure 1.
Kaplan-Meier plot of event-free survival based on BRCA1 monoallelic inactivation status.
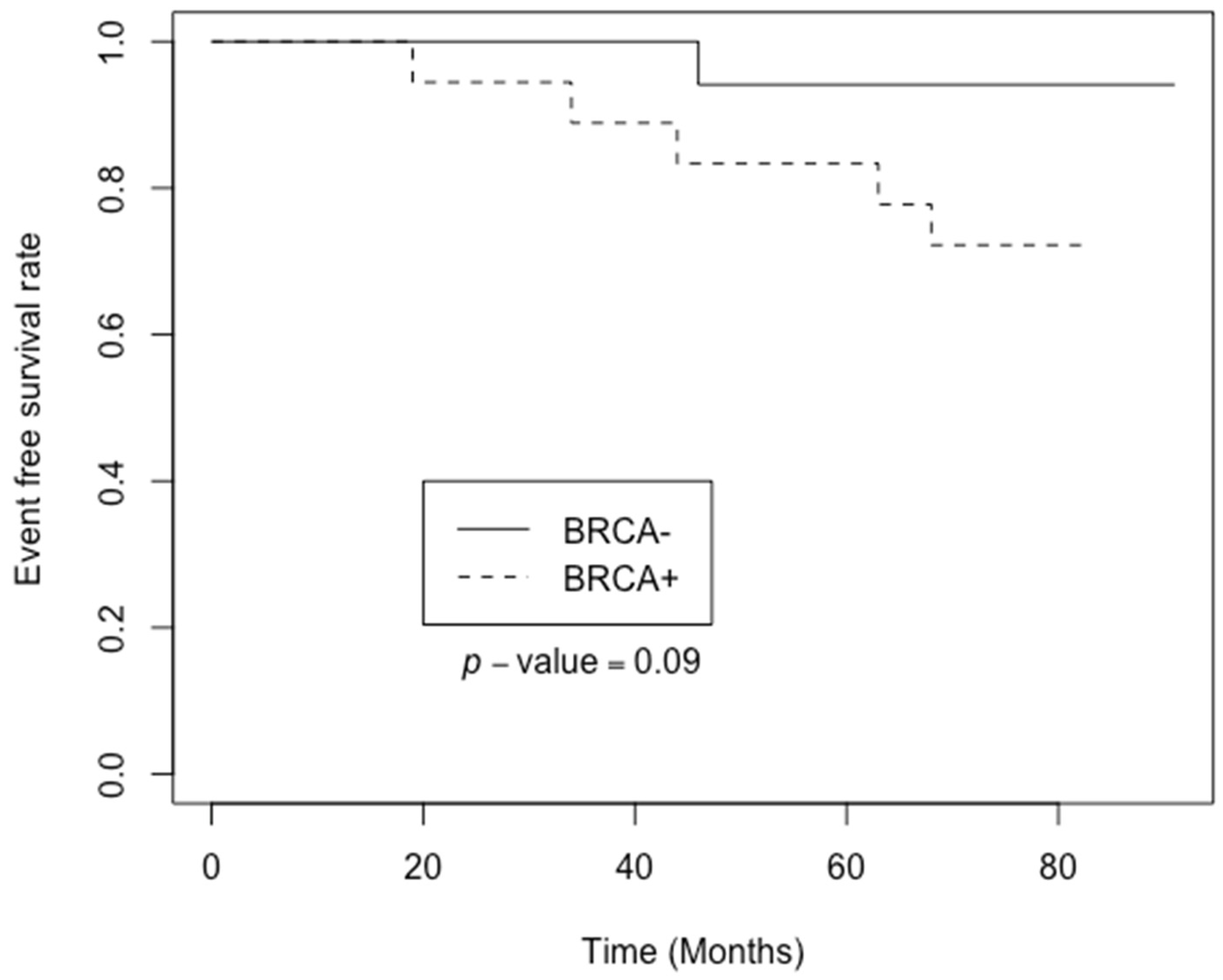
Figure 2.
The DEGs coded protein interaction in BRCA1- group.
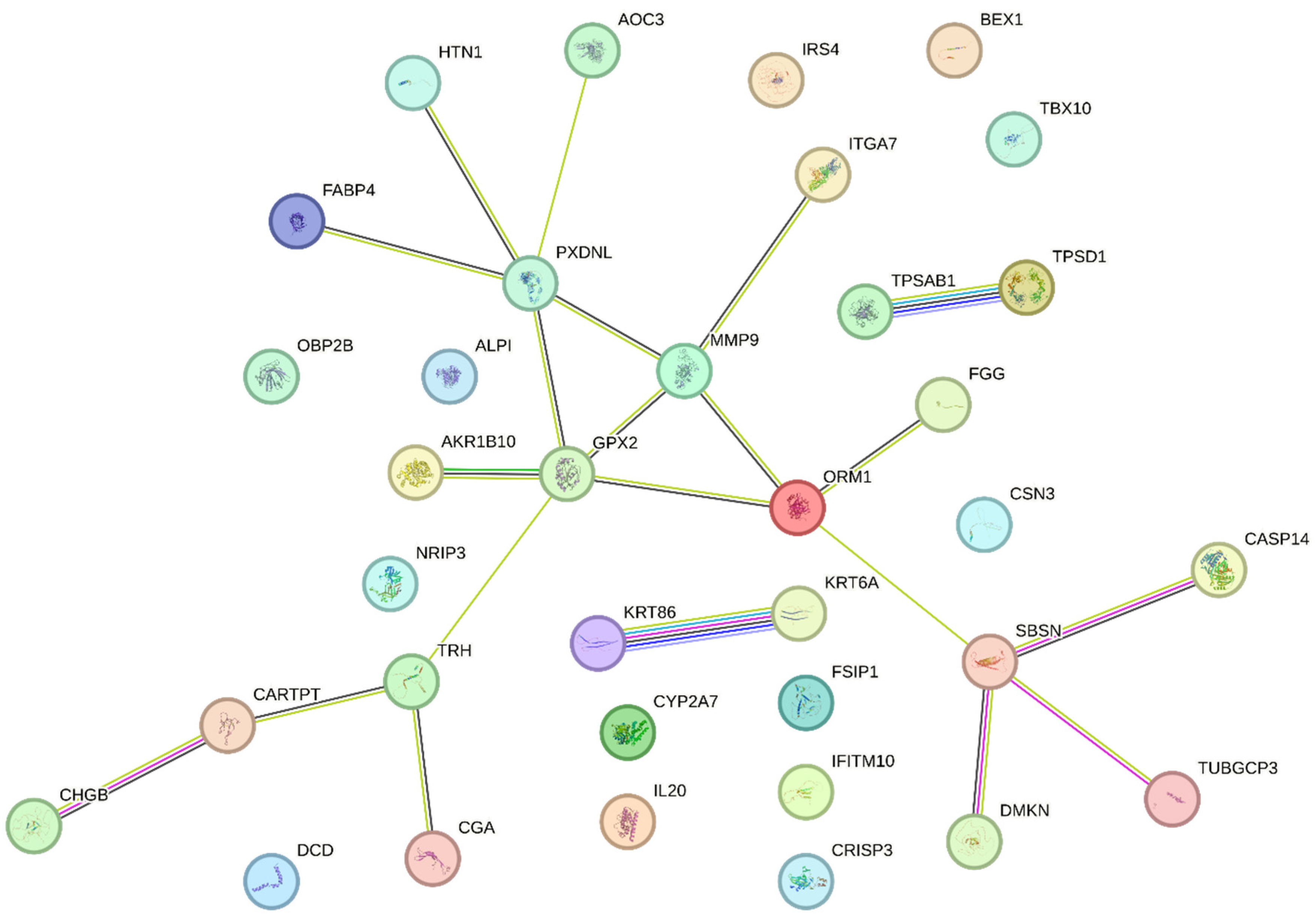

Table 1.
The list of upregulated DEGs in BRCA1-group. Bonferroni adjustment p <0.05; Max group mean threshold of >10.
Table 1.
The list of upregulated DEGs in BRCA1-group. Bonferroni adjustment p <0.05; Max group mean threshold of >10.
| Gene | Log₂ fold change | Fold change | p-value | HGNC |
|---|---|---|---|---|
| NRIP3 | 2.37 | 5.17 | 1.81E-08 | HGNC:1167 |
| TUBGCP3 | 2.42 | 5.37 | 7.66E-08 | HGNC:18598 |
| GPX2 | 2.72 | 6.60 | 6.25E-08 | HGNC:4554 |
| PXDNL | 2.84 | 7.15 | 3.21E-07 | HGNC:26359 |
| FSIP1 | 3.06 | 8.32 | 5.6E-09 | HGNC:21674 |
| IL20 | 3.35 | 10.22 | 1.31E-07 | HGNC:6002 |
| MMP9 | 3.75 | 13.46 | 6.45E-08 | HGNC:7176 |
| TPSD1 | 3.81 | 14.06 | 3.6E-07 | HGNC:14118 |
| TPSAB1 | 3.85 | 14.43 | 1.45E-08 | HGNC:12019 |
| TRH | 4.36 | 20.56 | 2.56E-10 | HGNC:12298 |
| AKR1B10 | 4.37 | 20.75 | 5.45E-07 | HGNC:382 |
| ORM1 | 4.52 | 22.86 | 4.8E-07 | HGNC:8498 |
| CGA | 4.75 | 26.90 | 1.18E-06 | HGNC:1885 |
| BEX1 | 4.87 | 29.27 | 5.66E-10 | HGNC:1036 |
| TBX10 | 5.72 | 52.82 | 9.21E-10 | HGNC:11593 |
| FGG | 5.81 | 56.16 | 5.42E-08 | HGNC:3694 |
| CASP14 | 6.27 | 77.34 | 4.16E-10 | HGNC:1502 |
| CRISP3 | 7.06 | 133.61 | 1.41E-16 | HGNC:16904 |
| CSN3 | 7.10 | 137.58 | 1.92E-10 | HGNC:2446 |
| HTN1 | 9.80 | 891.01 | 1.74E-12 | HGNC:5283 |
| ALPI | 10.61 | 1566.05 | 4.67E-16 | HGNC:437 |
Table 2.
The list of downregulated DEGs in the BRCA1-group. Bonferroni adjustment p<0.05; Max group mean threshold of >10.
Table 2.
The list of downregulated DEGs in the BRCA1-group. Bonferroni adjustment p<0.05; Max group mean threshold of >10.
| Gene | Log₂ fold change | Fold change | p-value | HGNC |
| CARTPT | -9.88 | -944.90 | 4.29E-20 | HGNC:24323 |
| SBSN | -8.98 | -504.29 | 2.71E-19 | HGNC:24950 |
| IRS4 | -8.00 | -256.83 | 1.06E-16 | HGNC:6128 |
| CHGB | -7.93 | -243.21 | 2.68E-17 | HGNC:1930 |
| CYP2A7 | -7.44 | -173.43 | 4.19E-22 | HGNC:2611 |
| KRT6A | -5.37 | -41.46 | 4.43E-10 | HGNC:6443 |
| DCD | -5.27 | -38.53 | 4.59E-09 | HGNC:14669 |
| OBP2B | -4.04 | -16.48 | 1.14E-10 | HGNC:23381 |
| KRT86 | -3.62 | -12.30 | 7.03E-09 | HGNC:6463 |
| DMKN | -3.30 | -9.83 | 1.16E-08 | HGNC:25063 |
| FABP4 | -3.09 | -8.49 | 2.55E-07 | HGNC:3559 |
| IFITM10 | -2.23 | -4.68 | 3.22E-08 | HGNC:40022 |
| AOC3 | -2.03 | -4.08 | 3.7E-07 | HGNC:550 |
| ITGA7 | -1.85 | -3.61 | 1.25E-06 | HGNC:6143 |
Table 3.
Results of gene enrichment analysis.
| #Category | Term ID | Term description | Genes from input | Strength | False discovery rate | Matching proteins in the network (labels) |
|---|---|---|---|---|---|---|
| GO Component | GO:0005576 | Extracellular region | 23/4175 | 0.49 | 3.74e-05 | TPSD1, FABP4, ORM1, ALPI, CARTPT, TRH, CSN3, FGG, DMKN, TPSAB1, PXDNL, AKR1B10, IL20, MMP9, CHGB, KRT6A, CRISP3, DCD, HTN1, SBSN, KRT86, CGA, OBP2B |
| GO Component | GO:0005615 | Extracellular space | 20/3247 | 0.54 | 5.90e-05 | TPSD1, FABP4, ORM1, CARTPT, CSN3, FGG, DMKN, TPSAB1, PXDNL, IL20, MMP9, CHGB, KRT6A, CRISP3, DCD, HTN1, SBSN, KRT86, CGA, OBP2B |
| COMPARTMENTS | GOCC:0005576 | Extracellular region | 17/2079 | 0.66 | 4.18e-05 | FABP4, ORM1, ALPI, CARTPT, TRH, CSN3, FGG, TPSAB1, PXDNL, AKR1B10, IL20, MMP9, CHGB, CRISP3, DCD, HTN1, CGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



