
Submitted:
18 October 2024
Posted:
24 October 2024
You are already at the latest version
Abstract
Candida albicans, a significant human pathogen due to its biofilm-forming ability, poses a serious health risk. This study investigates the antifungal activity and biofilm inhibition potential of Tea Tree Oil (TTO) from Melaleuca alternifolia cultivated in Vietnam. Renowned for its bioactive compounds, including terpinen-4-ol, 1,8-cineole, and terpinolene, the essential oil of M. alternifolia, commonly referred to as TTO, is scrutinized for its impact on C. albicans. The study meticulously determines the Minimum Inhibitory Concentration (MIC) and Minimum Fungicidal Concentration (MFC) of TTO against C. albicans in both planktonic and biofilm states. Additionally, it assesses the biofilm-forming capacity of C. albicans and establishes the Minimum Biofilm Inhibitory Concentration (MBIC) and Minimum Biofilm Eradication Concentration (MBEC) of TTO. Employing a liquid culture dilution method within a defined concentration range of 0.1 μL/mL to 10 μL/mL, the experimental approach adheres to established protocols. The outcomes reveal a noteworthy MIC and MIC80 of 0.1 μL/mL and 2 μL/mL, respectively, and an MFC of 10 μL/mL for TTO against C. albicans. Furthermore, TTO exhibits an MBIC of 2 μL/mL, showcasing its efficacy in inhibiting biofilm formation. However, the MBEC of TTO remains uncertain, possibly due to insufficient concentration tested, which may not effectively disrupt over 50% of the pre-formed biofilm mass within 24 hours. These findings highlight TTO’s significant potential as both an antifungal agent and a biofilm inhibitor against C. albicans, providing essential insights that could guide the development of new therapeutic approaches for treating fungal infections.
Keywords:
Candida albicans
; Tea tree oil
; biofilm
; antifungal
; biofilm inhibition
; modified tea tree oil formulation
; Minimum Fungicidal Concentration (MFC)
; Minimum Biofilm Inhibitory Concentration (MBIC)
; Minimum Biofilm Eradication Concentration (MBEC)
1. Introduction
Candida albicans is widely prevalent in the human microbiota and possesses opportunistic pathogenic traits, contributing to the development of serious, sometimes fatal, bloodstream infections. [1] The crucial virulence factor in its pathogenesis lies in the initiation of adhesion and the subsequent formation of biofilms on both abiotic and biotic surfaces. [2] The growing apprehension about drug resistance emphasizes the urgency of discovering new agents, leading to an increased focus on exploring novel antifungal compounds effective against biofilms. [3] Traditional medicinal plants and alternative therapies have emerged as promising sources for identifying innovative antimicrobial agents.
Natural products play a crucial role in the exploration of novel antifungal therapies. Numerous studies have underscored the antimicrobial effectiveness of these products against oral pathogens, positioning herbal agents as potential tools for preventing and managing infectious oral diseases. [4,5] In Vietnam, Melaleuca alternifolia, a member of the Myrtaceae family native to Australia commonly known as tea tree, produces an essential oil known as tea tree oil (TTO) through the steam distillation of its leaves. Containing around 100 compounds, among which terpinen-4-ol constitutes a minimum of 30% of the oil, and with notable presence of 1,8-cineole and terpinolene, TTO displays a diverse array of biological activities. These actions aid in efficiently handling diverse microorganisms, improving approaches to tackle persistent infections such as candidiasis, pharyngitis, vaginitis, and respiratory tract diseases. TTO influences biological membranes by compromising their integrity and impeding enzyme function, leading to heightened membrane fluidity and the release of intracellular components. [6] As highlighted by Cox and Markham (2007), the hydrophobic nature of its chemical constituents enables them to accumulate in lipid-rich membrane environments, resulting in structural and functional damage.
Carson et al. (2006) conducted a study [7] that determined the concentration ranges for key terpenes, alcohols, and ethers in TTO. The study also provided guidelines stating that the terpinen-4-ol content should be a minimum of 30% and the 1,8-cineole content should not exceed 15% of the oil volume. Due to its antimicrobial, antifungal, and anti-inflammatory properties, terpinen-4-ol, which is the main active component in TTO, has attracted significant attention. [8] Notably, terpinen-4-ol induces membrane disruption, compromising the integrity and physiology of microbial cells. [7,9,10,11] By virtue of its low concentrations, it demonstrates non-toxicity towards fibroblasts and epithelial cells, making it suitable for topical use with minimal adverse effects
In this study, the chemical composition of Tea Tree Oil (TTO) was analyzed using gas chromatography-mass spectrometry (GC-MS), identifying a range of bioactive compounds. The major components include eucalyptol (18-cineole), which constitutes 29.41% of the oil. This compound plays a significant role in enhancing the permeability of microbial cell membranes, facilitating the penetration of other compounds and disrupting internal cellular processes [12]. Terpinolene, accounting for 8.63%, is another major compound known for its strong antimicrobial and antifungal properties, disrupting the cell membranes of C. albicans and inhibiting fungal growth [7]. Additionally, γ-terpinene (7.92%) has antioxidant and antimicrobial properties that further contribute to the disruption of microbial cell structures [9], while α-terpineol (5.67%) has been shown to effectively inhibit the growth of C. albicans and other bacteria [13]. Furthermore, the most bioactive component, terpinen-4-ol, is known for its membrane-disrupting effects on C. albicans, altering membrane permeability and causing ion imbalance, which leads to microbial cell death [11]. These findings highlight the complex chemical profile of TTO and its potent antifungal and biofilm-inhibitory properties, supporting its use as an effective agent against C. albicans.
This study aims to evaluate the antifungal and biofilm-inhibitory potential of a modified formulation of TTO derived from Melaleuca alternifolia cultivated in Vietnam. We investigate its MIC, MFC, MBIC, and MBEC against C. albicans. By exploring both planktonic and biofilm states of C. albicans, this study provides a comprehensive evaluation of TTO’s potential as a novel therapeutic agent for managing fungal infections, particularly those complicated by biofilm formation. Additionally, we highlight the novelty of using a modified TTO formulation with enhanced concentrations of its key bioactive components to assess its efficacy compared to previously studied formulations.
2. Material and Methods
Microorganisms, cultivation environment, and conditions. A laboratory intervention experiment was conducted at the Faculty of Chemical Engineering, Ho Chi Minh City University of Technology (HCMUT), Vietnam National University-Ho Chi Minh City (VNU-HCMC). The microbial strain under investigation was the C. albicans strain (ATCC 24433) purchased from KwikStik Co., Ltd, Vietnam. They were activated and cultured in a self-prepared liquid Hansen medium (comprising 50 g H2SO4, 3 g KH2PO4, 3 g MgSO4.7H2O, 10 g peptone, 20 g agar, 1000 mL distilled water, adjusted to pH 6), at room temperature, aerobic conditions, and shaken at 150 rpm for 24 hours. The Hansen agar medium used for C. albicans growth had the following composition: 50 g/L H₂SO₄, 3 g/L KH₂PO₄, 3 g/L MgSO₄·7H₂O, 10 g/L peptone, 20 g/L agar, and 1 L distilled water. The pH was adjusted to 6.0 before autoclaving at 121°C for 15 minutes, as per standard protocols [14,15,16].
Bioactive compounds. This study utilized pure tea tree oil (Melaleuca alternifolia) obtained from Thuy Moc Viet Co., Ltd, Vietnam. The chemical components of the tea tree oil were analyzed by the Institute of Materials Science - Vietnam Academy of Science and Technology. Additionally, the antifungal agent Amphotericin B (AMB) was purchased from Maxxcare VP Co., Ltd, India, and used as a positive control in antifungal activity assays and biofilm inhibition studies against C. albicans.
Preparation of Emulsions. Due to the hydrophobic nature of tea tree oil (TTO), it was emulsified with sterile distilled water containing 0.5% Tween 80 before being mixed into the culture medium, following the method described by Van Nguyen, H., et al (2017) [17]. However, some adjustments were made to ensure compatibility with experimental conditions, with final concentrations achieved after dilution being 0.1 μL, 1 μL, 2 μL, 5 μL, 10 μL/mL. The entire experiment was conducted at these five concentration thresholds. The AMB antifungal agent used as a positive control was also dissolved in sterile distilled water containing 0.5% Tween 80 and volumes were calculated to achieve final concentrations of 0.1 μg, 1 μg, 2 μg, 5 μg, 10 μg/mL. The overall experimental sequence is illustrated in Figure 1A.
Selection of Culture Medium. The Mueller-Hinton medium, supplemented with 0.5% Tween 80, was chosen over RPMI-1640 due to its enhanced compatibility with hydrophobic substances like TTO. This combination ensures better solubility and diffusion of the essential oil in the aqueous medium, which is critical for accurate determination of the Minimum Inhibitory Concentration (MIC) and Minimum Fungicidal Concentration (MFC). While RPMI-1640 is commonly used in antifungal testing, the choice of Mueller-Hinton was based on its success in studies with essential oils, ensuring consistent and reproducible results under the specific conditions of this research [18,19,20].
Determining the Minimum Inhibitory Concentration (MIC) and Minimum Fungicidal Concentration (MFC). For MIC determination, C. albicans was first cultured in Hansen medium, incubated at room temperature with shaking at 150 rpm for 24 hours. The cells were centrifuged at 4000 rpm for 20 minutes, washed with PBS, and resuspended in Mueller-Hinton medium. Cell density was adjusted to an OD range of 0.119-0.14 at 530 nm (approximately 1×10⁶-6.2×10⁶ CFU/mL). In the test tubes, 4 mL of Mueller-Hinton medium containing the cell suspension was mixed with 1 mL of emulsified TTO, yielding final concentrations of 0.1, 1, 2, 5, and 10 μL/mL. Controls included AMB (0.1-10 μg/mL) and untreated Mueller-Hinton medium. After a 24-hour incubation at room temperature, the optical density was measured at 610 nm using a UV-VIS spectrophotometer to determine the MIC, defined as the lowest concentration that inhibited fungal growth compared to controls [21,22,23,24] (Figure 1B).
To establish MFC, 50 μL samples from tubes exhibiting MIC were spread onto Hansen agar and incubated at room temperature for 48 hours. The MFC was identified as the lowest concentration showing no visible fungal growth.
MIC represents the lowest concentration inhibiting visible fungal growth, while MIC80 is the concentration causing an 80% reduction in optical density compared to the control. MFC is defined as the lowest concentration showing no colony formation on agar after 48 hours [22,24]
Following that, 4 mL of Mueller-Hinton medium containing fungal cell suspension was added to each test tube. Then, 1 mL of TTO, emulsified with sterile distilled water containing 0.5% Tween 80, was added to achieve final concentrations of 0.1 μL, 1 μL, 2 μL, 5 μL, 10 μL/mL in the medium. The positive control (AMB) consisted of 4 mL of Mueller-Hinton medium containing fungal cell suspension and 1 mL of AMB mixed in sterile distilled water containing 0.5% Tween 80, resulting in final concentrations of 0.1 μg, 1 μg, 2 μg, 5 μg, 10 μg/mL. The negative control contained 4 mL of Mueller-Hinton medium with fungal cell suspension and 1 mL of sterile distilled water containing 0.5% Tween 80. Blank samples were prepared similarly but without fungal cell suspension. The tubes were then incubated at room temperature for 24 hours. After incubation, the optical density was measured at a wavelength of 610 nm using a UV-VIS spectrophotometer to determine the MIC of TTO in inhibiting fungal growth.
From the tubes identified as MICs, 50 μL of these samples were spread onto the surface of Hansen medium and incubated at room temperature for 48 hours to determine the MFC of TTO. The MIC and MFC experiments were conducted independently three times, and the averages were calculated.
MIC is defined as the lowest concentration of TTO or antifungal agent that inhibits the fungal growth, as indicated by a reduction in optical density compared to the control sample [16,25]. MIC80 is the lowest concentration of tea tree oil or antifungal agent causing an 80% reduction in optical density compared to the control sample [16,25]. MFC is the lowest concentration of TTO or antifungal agent demonstrating no growth of fungal colonies on the agar surface after 48 hours of incubation [26,27].
Investigation of the biofilm-forming ability of C. albicans. The experimental procedure is illustrated in Figure 1C. C. albicans strains were cultured in Hansen medium with additional glucose concentrations of 2.5%, 5%, and 10% (w/v) in 2 mL round-bottom Eppendorf tubes.
Materials preparation followed Pereira et al. (2015) [28] with minor adjustments. C. albicans was cultured on agar for 48 hours, then inoculated into Hansen liquid medium at room temperature for 24 hours with shaking at 150 rpm. After centrifugation at 4000 rpm for 20 minutes and two PBS washes, the pellet was resuspended in Hansen medium with glucose concentrations of 2.5%, 5%, and 10% (w/v). The cell density was adjusted to approximately 107 CFU/mL [OD530 nm values between 0.39 and 0.43, equivalent to 1.26×107 - 6.49×107 CFU/mL] [29]. Subsequently, 1 mL of the standard inoculum [yeast cell suspension in Hansen medium with 2.5%, 5%, and 10% glucose (w/v)] was aspirated into 2 mL Eppendorf tubes and incubated undisturbed at room temperature for 24 hours.
Biofilm formation capability was assessed by quantifying total biomass using the Crystal Violet (CV) staining method, adapted from Silva et al. (2009) [30]. After a 24-hour incubation, wells were washed with sterile PBS (pH 7.2) three times to remove residual medium and non-adherent fungal cells. Methanol (1 mL) was added and removed after 15 minutes. Eppendorf tubes were air-dried, followed by the addition of 1 mL CV 1% (v/v) for 5 minutes. After washing with PBS, 33% (v/v) acetic acid (1 mL) was added, and absorbance was read at 570 nm using a UV-VIS spectrophotometer. The experiment was conducted in triplicate, with three parallel samples for each repetition.
In this study, we opted to use 2 mL round-bottom Eppendorf microtubes instead of 96-well microplates to grow C. albicans biofilms. Microtubes provide a more enclosed environment, which helps prevent medium evaporation and ensures stable conditions for biofilm formation during long-term experiments, especially under static conditions [31]. Additionally, microtubes allow for more accurate control of aeration and mixing when placed on a shaker, facilitating better biofilm formation by reducing the risk of desiccation or inconsistent nutrient diffusion. Microtubes also simplify handling during the washing and staining steps required for biofilm quantification assays, enhancing reproducibility due to the consistent geometry of the tubes [32].
Biofilm-forming ability, assessed by total biomass quantification, followed Alves et al.‘s method (2023) [33]:
ODc = ODnc + 3SD
Where:
ODc: The cut-off optical density
ODnc: The mean OD of the negative control
SD: The standard deviations of OD values of negative control samples (ODs)
Positive samples (ODs > ODc) were considered biofilm producers, categorized as:
ODs ≤ ODc: no biofilm producer
ODc < ODs ≤ 2ODc: week biofilm producer
2ODc < ODs ≤ 4ODc: moderate biofilm producer
4ODc < ODs: strong biofilm producer
Determining the Minimum Biofilm Inhibitory Concentration (MBIC) and Minimum Biofilm Eradication Concentration (MBEC). The experimental sequence is depicted in Figure 1D and Figure 1E. The cell density in the C. albicans yeast suspension for MBIC and MBEC determination should be around 107 CFU/mL, [OD530 nm values between 0.39 and 0.43, approximately 1.26×107 - 6.49×107 CFU/mL] [14] and must be re-suspended in Hansen medium (the Hansen medium composition was selected from the C. albicans biofilm formation capability assessment experiment).
MBIC and MBEC are defined as the lowest concentration of essential oil that inhibits over 50% of biofilm formation and the lowest concentration of essential oil that eradicates over 50% of pre-formed biofilm, respectively [34].
MBIC and MBEC were determined following Purwasena et al.‘s method (2020) [34], with adjustments. C. albicans yeast cell suspension (900 μL) in 2 mL Eppendorf tubes was prepared. Different concentrations of essential oil (0.1 μL, 1 μL, 2 μL, 5 μL, 10 μL/mL) were added in 100 μL increments. Positive controls (using AMB) were also prepared at concentrations of 0.1 μg, 1 μg, 2 μg, 5 μg, 10 μg/mL. The tubes were statically incubated for 24 hours at room temperature. After incubation, the medium was removed, and tubes were washed with sterile PBS buffer (pH 7.2) three times to eliminate free cells. Biofilms were stained with 0.1% crystal violet for 30 minutes. After staining, crystal violet was removed, and tubes were rinsed with PBS until runoff was colorless. Finally, biofilms were dissolved in 95% ethanol for 15 minutes, and absorbance at 595 nm was measured using a UV-VIS spectrophotometer to determine MBIC and MBEC. The untreated sample contained only the growth medium, sterile distilled water with 0.5% Tween 80, and yeast cell suspension. The negative control contained only the growth medium. The experiment was repeated four times, and average values were obtained.
Statistical analysis. We used Dixon’s Q test to identify and remove outliers when analyzing microbial growth or biofilm assay data. For each concentration tested in the assays mentioned above, we calculated the arithmetic mean and standard deviation (SD), which were presented as a relative percentage compared to the control sample (set at 100%). Dixon’s Q test was utilized to identify outliers in the data obtained from the biofilm assays [35]. We performed a one-way analysis of variance (ANOVA) with a significance level of p < 0.05 to assess the statistical significance of the disparities observed between the control and adjuvant effects [36]. Statistical analysis was conducted using standard protocols for microbial diversity and biofilm formation studies [37]. The approval of an ethics committee is not necessary for this study.
3. Results
Morphology of C. albicans. In terms of morphological characteristics, C. albicans fungal cells were uniformly observed in a yeast form (Figure 2a). C. albicans cells were streaked on the surface of the Hansen agar medium and incubated at room temperature (Figure 2b). Regarding the macroscopic morphology of C. albicans, after 24 hours of incubation, C. albicans colonies grew in circular shapes, appearing opaque white, with an approximate diameter of about 0.5 mm (Figure 2b-A). After 48 hours of incubation, C. albicans colonies exhibited an increase in diameter (Figure 2b-B).
Investigation of the antifungal activity of TTO against C. albicans. The results determined by two criteria, MIC and MFC, are presented in Figure 3 and Table 1.
Figure 3A illustrates that both the antifungal agent AMB and TTO at all tested concentrations exhibit a decrease in optical density compared to the negative control (-), indicating MIC values of 0.1 μg/mL for AMB and 0.1 μL/mL for TTO. All concentrations of the positive control (treated with AMB) show a growth inhibition percentage in C. albicans not exceeding 50%. For the TTO-treated sample, the antifungal activity of the oil follows the pattern of „increasing concentration of the active substance, increasing antifungal activity,” similar to the AMB antifungal agent.
Based on the results presented in Figure 3B and Table 1, at a concentration of 2 μL/mL (1824.34 μg/mL), the TTO-treated sample caused a reduction of over 80% in optical density compared to the negative control, specifically inhibiting the growth of C. albicans by 87.4 ± 0.27%. This concentration is noted as MIC80. The survey results also recorded the MIC of tea tree oil against C. albicans as 0.1 μL/mL (91.217 μg/mL). Regarding the MFC criterion, only the sample treated with TTO at a concentration of 10 μL/mL (9121.7 μg/mL) showed no formation of fungal clusters on the surface of the Hansen medium after 48 hours of incubation.
Investigation of the biofilm formation capability of C. albicans. The experiment was conducted with three independent trials, and each trial included three parallel samples. The results of the experiment are shown in Figure S1, Table 2 and Table 3.
According to the data presented in Table 2 and Table 3, all three experimental trials indicate that the addition of varying percentages of glucose (w/v) to the Hansen medium can stimulate C. albicans to form biofilm (refer to Table 3). Analyzing the results from Table 3, NT1 exhibits the most proficient biofilm-forming capacity of C. albicans, achieving a moderate level of biofilm formation, in contrast to the other two trials which demonstrate weaker biofilm formation. This disparity could potentially be attributed to the elevated levels of supplemental D-glucose in the Hansen medium at 5% and 10%, inadvertently acting as an inhibitory factor that impedes biofilm formation. Consequently, there is an observable tendency toward decreased biofilm formation under these conditions.
Biofilm formation by C. albicans on Hansen medium supplemented with 2.5% glucose was documented through electron microscopy (Figure 4). Consequently, the Hansen medium enriched with 2.5% glucose is characterized as having the capacity to foster a moderate level of biofilm formation, establishing a foundation for subsequent investigations.
Exploring the inhibitory impact of TTO on C. albicans biofilm formation. The outcomes presented above provide the basis for the selection of an optimal biofilm-forming environment for C. albicans in this experiment. Consequently, the „Hansen medium supplemented with 2.5% glucose (w/v)” has been chosen as the experimental condition for biofilm formation. This choice is informed by its superior biofilm-forming capability compared to the other conditions investigated. The trial was carried out concurrently with four parallel samples. The outcomes were assessed using two metrics: MBIC, presented in Figure 5 and Table 4, and MBEC, as demonstrated in Figure 6 and Table 4.
We performed t-tests to compare the inhibitory effects of AMB and TTO (following MBIC) on C. albicans against the control (CK) at different concentrations. The statistical analysis revealed significant differences between the control and various concentrations of both substances. For AMB, a significant inhibitory effect was observed as early as 0.1 μL/mL (p ~ 0.022), with the significance increasing at higher concentrations, and a highly significant difference at 10 μL/mL (p ~ 0.012). Similarly, for TTO, starting at 2 μg/mL, there was a statistically significant difference compared to the control (p < 0.05). At 5 μg/mL (p = 0.010) and 10 μg/mL (p = 0.012), the difference was highly significant (p < 0.01), indicating a strong inhibitory effect at higher concentrations.. These results demonstrate that both AMB and TTO exhibit substantial antifungal activity against C. albicans, particularly at higher concentrations. (see Table S1)
The statistical analysis using t-tests to compare the inhibitory effects of AMB and TTO (following MBEC) compared to the control at different concentrations on C. albicans revealed varying degrees of effectiveness at different concentrations. For AMB, there were no significant differences at the concentrations of 0.1 μg/mL and 1 μg/mL (p > 0.05), indicating no notable inhibitory effect at these lower concentrations. At 2 μg/mL, although the difference was not statistically significant (p = 0.052), the result was close to significance, suggesting that inhibitory effects might begin to manifest at this concentration. The concentration of 5 μg/mL also did not show a significant difference (p = 0.101), but at the highest concentration of 10 μg/mL, a statistically significant difference was observed (p = 0.019), indicating a strong inhibitory effect. For TTO, the concentrations of 0.1 μL/mL and 1 μL/mL did not result in significant differences (p > 0.05), suggesting no strong inhibitory effect at these levels. At 2 μL/mL, although no significant difference was observed (p = 0.130), the data indicate that inhibitory effects might start appearing. The result for 5 μL/mL was close to significance (p = 0.089), while at 10 μL/mL, although not statistically significant, the difference approached significance (p = 0.067), suggesting a considerable inhibitory potential at this higher concentration. In summary, AMB demonstrated a significant inhibitory effect at 10 μg/mL, and while TTO did not reach statistical significance at 10 μL/mL, the data suggest a notable inhibitory effect at this concentration. (see Table S2)
Regarding the MBEC, the findings depicted in Figure 6 reveal an incremental trend in the eradication capacity of pre-formed C. albicans biofilm with rising concentrations of the bioactive agents (AMB and TTO). However, neither AMB nor TTO exhibited a specific MBEC value, as the biofilm mass at all tested concentrations surpassed 50% when compared to the untreated control. In essence, the biofilm eradication efficacy of both bioactive compounds remained below 50%, precluding the determination of a definitive MBEC value. At a concentration of 10 μg/mL, AMB eradicated only 49.28% of the C. albicans biofilm mass, while for TTO treatment at the highest concentration of 10 μL/mL, the reduction in biofilm mass after 24 hours of pre-culture was recorded at 62.43 ± 17.82% compared to the untreated control.
4. Discussion
Investigation of the antifungal activity against C. albicans of TTO. The experiments demonstrated that adding sterile distilled water containing 0.5% Tween 80 to Mueller-Hinton medium promotes the growth of C. albicans, as opposed to the medium without this supplementation. This observation is consistent with the findings of Carson and Riley, who suggested that Tween 80 serves as a stimulant due to its oleic acid content [38]. Consequently, the Mueller-Hinton medium supplemented with sterile distilled water containing 0.5% Tween 80 is designated as the negative control for comparison with samples treated with the antifungal agent AMB and TTO.
The MIC of TTO against C. albicans was found to be 0.1 μL/mL (91.217 μg/mL) in our study. However, other studies, such as Thosar et al. (2013) [39], reported a MIC of 0.5 μL/mL for C. albicans. Rosato et al. (2008) [40] demonstrated MIC values ranging from 1.75 to 3.5 mg/mL for various Candida strains treated with TTO. In contrast, Oro et al. (2015) [41] reported a lower average MIC of 20.02 μg/mL for C. albicans when exposed to TTO. These variations suggest differing sensitivities among C. albicans strains.
MFC values for TTO against C. albicans exhibit variability among studies. Oro et al. (2015) reported an MFC of 25.33 μg/mL [41], while Francisconi et al. (2015) found it to be 0.25% TTO [8]. Noumi et al. (2011) observed different MFC values across C. albicans strains, ranging from 5 to >10 mg/mL [42]. This variation highlights the diverse sensitivity of C. albicans strains to TTO, mirroring trends seen in MIC values.
In terms of mechanism, AMB acts by creating pores in the fungal cell membrane, altering permeability, and causing cell death. TTO is thought to penetrate the cell wall and plasma membrane of C. albicans, disrupting these structures and leading to cell leakage due to its lipophilic nature.
TTO analysis identified eucalyptol (or 1,8-cineole) as the predominant component, comprising 29.41% of the oil. Other notable constituents include terpinolene (8.63%), γ-terpinene (7.92%), α-terpineol (5.67%), and various trace components. The antifungal efficacy of TTO is attributed to the presence of 1,8-cineole, which has been shown in studies to enhance fungal cell membrane permeability [12,13]. This facilitates the penetration of other oil components into the cells, influencing internal cellular processes [12].
Comparison with Previous TTO Formulations. The antimicrobial activity of TTO against C. albicans and its ability to inhibit biofilm formation have been extensively studied. However, our study utilizes a modified TTO formulation with altered concentrations of key active components, such as terpinen-4-ol and α-terpineol. These components are known to contribute significantly to the antimicrobial efficacy of TTO. For instance, previous research has shown that terpinen-4-ol, one of the major constituents of TTO, plays a crucial role in the inhibition of fungal growth and biofilm formation [13]. Higher concentrations of terpinen-4-ol in TTO formulations have been associated with stronger antifungal activity, while formulations with lower concentrations of this component demonstrate reduced efficacy [7]. In our formulation, the increased concentration of terpinen-4-ol likely contributes to the enhanced biofilm inhibition observed in our study compared to other formulations.
Impact of Specific Components on Antimicrobial Activity. In addition to terpinen-4-ol, α-terpineol is another important component of TTO that contributes to its antimicrobial properties. It has been reported that α-terpineol enhances the overall antifungal effects of TTO by increasing the disruption of microbial cell membranes [43]. The synergistic effects of terpinen-4-ol and α-terpineol may explain the high efficacy of our modified TTO formulation in inhibiting C. albicans biofilm.
Toxicity Considerations. Although this study focuses primarily on the antimicrobial effects of TTO, the potential toxicity of its components should also be considered. Research has shown that terpinen-4-ol and α-terpineol can exhibit cytotoxic effects at high concentrations, particularly in skin and mucosal tissues [44]. However, these cytotoxic effects tend to occur at concentrations higher than those used in typical antimicrobial applications [45]. While we did not directly assess the toxicity of our modified TTO formulation, future studies would be necessary to evaluate its safety, especially for therapeutic applications.
Investigation of C. albicans biofilm formation. The study conducted by Brambilla et al. (2015) revealed an increased biofilm formation of C. albicans with the rising percentage of D-glucose in the Trypticase-Soy broth (TSB) medium, ranging from 1.25 to 10% (w/v) [46]. This finding contrasts with the survey results, given the varying glucose concentrations in the growth media used for C. albicans biofilm formation. Specifically, TSB medium contains 2.5 g/L of glucose, while Hansen medium boasts a glucose concentration of up to 50 g/L. Consequently, this discrepancy may have impacted C. albicans’ biofilm formation in the study, leading to a tendency of reduced biofilm formation in NT2 and NT3 due to the exceptionally high sugar content in the experimental environment, inadvertently resulting in inhibitory effects on C. albicans.
Investigating the inhibition of C. albicans biofilm formation by TTO. The study found a MBIC of the antifungal agent AMB at 2 μg/mL (Figure 5, and Figure S2). Zarei et al. (2014) investigated MBIC on 120 C. albicans strains, revealing that 65% of them had an MBIC below 10 μg/mL, indicating the overall sensitivity of C. albicans strains to AMB. The observed MBIC of 2 μg/mL aligns with the known effectiveness of AMB in inhibiting C. albicans biofilm formation under in vitro conditions [47].
For the TTO-treated sample, the investigation identified an MBIC value of 2 μL/mL (Figure 5, and Figure S2). The diverse terpenes present in tea tree oil modify cell membrane permeability by infiltrating between the fatty-acyl chains of the cell membrane bilayer. This induces changes in membrane permeability, leading to alterations in cell surface and morphology and diminishing adhesion capabilities to C. albicans’ substrate [48]. This contributes to the observed reduction in biofilm formation of C. albicans when treated with TTO.
The absence of recorded MBEC values for both biological agents (AMB and TTO) in the survey (Table 4) may be attributed to the tea tree oil concentration examined not being sufficiently high to disrupt more than 50% of the pre-cultured biofilm mass within 24 hours. At the highest concentration of tea tree oil in the survey (10 μL/mL), it was observed to disrupt 37.57% of the C. albicans biofilm mass compared to the untreated sample, as indicated in Figure 6.
There are key limitations to consider in our study interpretation. Firstly, our focus was solely on tea tree oil’s antifungal and biofilm inhibitory effects on C. albicans, without comparison to other treatments or examination of its long-term impact. Secondly, our testing methods may not reflect real-world conditions accurately. Thirdly, the study used only one C. albicans (ATCC 24433) strain, potentially limiting generalizability. Fourthly, we didn’t address the purity of the tea tree oil used, which could affect results. Finally, synergistic effects with other antifungals weren’t explored, potentially impacting effectiveness. Hence, acknowledging inevitable measurement errors is crucial.
5. Conclusion
In this study, we investigated the antifungal efficacy of a modified formulation of TTO against C. albicans, focusing on both its planktonic and biofilm states. Our findings establish the MIC of TTO at 0.1 μL/mL and the MFC at 10 μL/mL, demonstrating its strong antifungal activity. Notably, we identified a MBIC of 2 μL/mL, highlighting TTO’s potent ability to inhibit biofilm formation. This study is among the first to provide a comprehensive evaluation of TTO’s effects on C. albicans, particularly in the context of biofilm formation, which is a critical factor in persistent fungal infections.
The modified TTO formulation used in this study, characterized by increased concentrations of terpinen-4-ol and α-terpineol, exhibited enhanced antifungal and biofilm inhibitory properties compared to previously studied formulations. These findings underscore the potential of this modified TTO formulation as a novel therapeutic agent for managing C. albicans infections, particularly those associated with biofilm formation. Further research into the toxicity profile and clinical applications of this formulation is warranted to fully explore its potential as a natural antifungal treatment.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
This research received no external funding.
Compliance with Ethical Standards
This article does not involve any studies conducted by the authors that included human participants.
Acknowledgments
The completion of this research work was made possible through the collaborative efforts and dedication of a multidisciplinary team. We extend our sincere appreciation to each member for their invaluable contributions.
Conflicts of Interest
The authors declare no conflicts of in-terest.
References
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single-and multi-species Candida albicans biofilms. Nature Reviews Microbiology 2018, 16, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Figueiral, M.H.; Azul, A.; Pinto, E.; Fonseca, P.; Branco, F.M.; Scully, C. Denture-related stomatitis: identification of aetiological and predisposing factors–a large cohort. Journal of oral rehabilitation 2007, 34, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Grando, T.; De Sá, M.; Baldissera, M.; Oliveira, C.; De Souza, M.; Raffin, R.; Santos, R.; Domingues, R.; Minho, A.; Leal, M. In vitro activity of essential oils of free and nanostructured Melaleuca alternifolia and of terpinen-4-ol on eggs and larvae of Haemonchus contortus. Journal of Helminthology 2016, 90, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Catalán, A.; Pacheco, J.G.; Martínez, A.; Mondaca, M.A. In vitro and in vivo activity of Melaleuca alternifolia mixed with tissue conditioner on Candida albicans. Oral Surgery, Oral Medicine, Oral Pathology, Oral Radiology, and Endodontology 2008, 105, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.; Dry, L.; Johnson, M.; Michalak, E.; Carson, C.; Riley, T. Susceptibility of oral bacteria to Melaleuca alternifolia (tea tree) oil in vitro. Oral microbiology and immunology 2003, 18, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.D.; Markham, J. Susceptibility and intrinsic tolerance of Pseudomonas aeruginosa to selected plant volatile compounds. Journal of Applied Microbiology 2007, 103, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Carson, C.F.; Hammer, K.A.; Riley, T.V. Melaleuca alternifolia (tea tree) oil: a review of antimicrobial and other medicinal properties. Clinical microbiology reviews 2006, 19, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Francisconi, R.S.; Bordini, E.A.F.; Nogueira, M.N.M.; Fontana, A.; Bedran, T.B.L.; Correia, M.F.; Spolidorio, D.M.P. Effect of Melaleuca alternifolia and its components on Candida albicans and Candida tropicalis. Journal of US-China Medical Science 2015, 12, 91–98. [Google Scholar]
- Maquera Huacho, P.M.; Rodriguez Herrero, E.; Verspecht, T.; Pauwels, M.; Marcantonio Jr, E.; Palomari Spolidorio, D.M.; Teughels, W. Terpinen-4-ol and carvacrol affect multi-species biofilm composition. Biofouling 2019, 35, 561–572. [Google Scholar] [CrossRef]
- Maquera-Huacho, P.M.; Tonon, C.C.; Correia, M.F.; Francisconi, R.S.; Bordini, E.A.F.; Marcantonio, É.; Spolidorio, D.M.P. In vitro antibacterial and cytotoxic activities of carvacrol and terpinen-4-ol against biofilm formation on titanium implant surfaces. Biofouling 2018, 34, 699–709. [Google Scholar] [CrossRef]
- Ramage, G.; Milligan, S.; Lappin, D.F.; Sherry, L.; Sweeney, P.; Williams, C.; Bagg, J.; Culshaw, S. Antifungal, cytotoxic, and immunomodulatory properties of tea tree oil and its derivative components: potential role in management of oral candidosis in cancer patients. Frontiers in microbiology 2012, 3, 220. [Google Scholar] [CrossRef] [PubMed]
- Brun, P.; Bernabè, G.; Filippini, R.; Piovan, A. In vitro antimicrobial activities of commercially available tea tree (Melaleuca alternifolia) essential oils. Current microbiology 2019, 76, 108–116. [Google Scholar] [CrossRef]
- Hammer, K.; Carson, C.; Riley, T. Antifungal activity of the components of Melaleuca alternifolia (tea tree) oil. Journal of applied microbiology 2003, 95, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Rodrı́guez-Tudela, J.L.; Cuenca-Estrella, M.; Dı́az-Guerra, T.M.; Mellado, E. Standardization of antifungal susceptibility variables for a semiautomated methodology. Journal of clinical microbiology 2001, 39, 2513–2517. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: quantification, structure and matrix composition. Sabouraudia 2009, 47, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Vaňková, E.; Paldrychová, M.; Kašparová, P.; Lokočová, K.; Kodeš, Z.; Maťátková, O.; Kolouchová, I.; Masák, J. Natural antioxidant pterostilbene as an effective antibiofilm agent, particularly for gram-positive cocci. World Journal of Microbiology and Biotechnology 2020, 36, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Van Nguyen, H.; Vu, T.T.; Chu-Ky, S.; Sarter, S. Interaction effects of Litsea cubeba essential oil and antibiotics on antibacterial activity against pathogenic bacteria in aquaculture. Vietnam Journal of Science and Technology 2017, 55, 66–73. [Google Scholar] [CrossRef]
- Chen, S.; Li, Z.; Gu, Z.; Ban, X.; Hong, Y.; Cheng, L.; Li, C. A new micro-agar dilution method to determine the minimum inhibitory concentration of essential oils against microorganisms. Journal of Microbiological Methods 2023, 211, 106791. [Google Scholar] [CrossRef] [PubMed]
- Hulankova, R. The influence of liquid medium choice in determination of minimum inhibitory concentration of essential oils against pathogenic bacteria. Antibiotics 2022, 11, 150. [Google Scholar] [CrossRef]
- Otajevwo, D.F.; Osawaru, O.E. Testing the efficacy of Mueller Hinton agar over Nutrient agar for optimal antibiotic sensitivity testing response by selected clinical bacterial pathogens. GSC Advanced Research and Reviews 2020, 5, 061–074. [Google Scholar] [CrossRef]
- Casagrande Pierantoni, D.; Corte, L.; Roscini, L.; Cardinali, G. High-Throughput rapid and inexpensive assay for quantitative determination of low cell-density yeast cultures. Microorganisms 2019, 7, 32. [Google Scholar] [CrossRef]
- Makambi, W.K.; Ikonomova, S.P.; Karlsson, A.J. Quantifying the antifungal activity of peptides against Candida albicans. JoVE (Journal of Visualized Experiments), 2023; e64416. [Google Scholar]
- Rouf, A.; Kanojia, V.; Naik, H.; Naseer, B.; Qadri, T. An overview of microbial cell culture. Journal of Pharmacognosy and Phytochemistry 2017, 6, 1923–1928. [Google Scholar]
- Xie, J.L.; Singh-Babak, S.D.; Cowen, L.E. Minimum inhibitory concentration (MIC) assay for antifungal drugs. Bio-protocol 2012, 2, e252–e252. [Google Scholar] [CrossRef]
- Vu, K.; Buckley, B.J.; Bujaroski, R.S.; Blumwald, E.; Kelso, M.J.; Gelli, A. Antifungal activity of 6-substituted amiloride and hexamethylene amiloride (HMA) analogs. Frontiers in Cellular and Infection Microbiology 2023, 13, 1101568. [Google Scholar] [CrossRef] [PubMed]
- Blanc, A.R.; Sortino, M.A.; Butassi, E.; Svetaz, L.A. Synergistic effects of Thymus vulgaris essential oil in combination with antifungal agents and inhibition of virulence factors of Candida albicans. Phytomedicine Plus 2023, 3, 100481. [Google Scholar] [CrossRef]
- Salem, M.Z.; Elansary, H.O.; Ali, H.M.; El-Settawy, A.A.; Elshikh, M.S.; Abdel-Salam, E.M.; Skalicka-Woźniak, K. Bioactivity of essential oils extracted from Cupressus macrocarpa branchlets and Corymbia citriodora leaves grown in Egypt. BMC complementary and alternative medicine 2018, 18, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Silva, S.; Ribeiro, B.; Henriques, M.; Azeredo, J. Influence of glucose concentration on the structure and quantity of biofilms formed by Candida parapsilosis. FEMS yeast research 2015, 15, fov043. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Tudela, J.L.; Cuenca-Estrella, M.; Diaz-Guerra, T.M.; Mellado, E. Standardization of antifungal susceptibility variables for a semiautomated methodology. J Clin Microbiol 2001, 39, 2513–2517. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: quantification, structure and matrix composition. Med Mycol 2009, 47, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Franklin, M.J. Physiological heterogeneity in biofilms. Nature Reviews Microbiology 2008, 6, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Merritt, J.H.; Kadouri, D.E.; O'Toole, G.A. Growing and analyzing static biofilms. Current protocols in microbiology 2011, 22, 1B–1. [Google Scholar] [CrossRef]
- Alves, A.M.C.V.; Lopes, B.O.; Leite, A.C.R.d.M.; Cruz, G.S.; Brito, É.H.S.d.; Lima, L.F.d.; Černáková, L.; Azevedo, N.F.; Rodrigues, C.F. Characterization of Oral Candida spp. Biofilms in Children and Adults Carriers from Eastern Europe and South America. Antibiotics 2023, 12, 797. [Google Scholar] [CrossRef] [PubMed]
- Purwasena, I.A.; Astuti, D.I.; Taufik, I.; Putri, F.Z. The Potential of Clove Essential Oil Microemulsion as an Alternative Biocide Against Pseudomonas aeruginosa Biofilm. Journal of Pure & Applied Microbiology 2020, 14. [Google Scholar]
- Kulišová, M.; Rabochová, M.; Lorinčík, J.; Brányik, T.; Hrudka, J.; Scholtz, V.; Jarošová Kolouchová, I. Exploring Non-Thermal Plasma and UV Radiation as Biofilm Control Strategies against Foodborne Filamentous Fungal Contaminants. Foods 2024, 13, 1054. [Google Scholar] [CrossRef]
- Krueger, Q.; Phippen, B.; Reitzel, A. Antibiotics alter development and gene expression in the model cnidarian Nematostella vectensis. PeerJ 2024, 12, e17349. [Google Scholar] [CrossRef] [PubMed]
- Morales-Medina, W.R. Assessing the Microbial Ecology of Sewer Biofilms for Pathogens, Antibiotic Resistance, and SARS-CoV-2 and Drinking Water Biofilters for Manganese Removal. Rutgers The State University of New Jersey, School of Graduate Studies, 2021.
- Carson, C.; Riley, T. Susceptibility of Propionibacterium acnes to the essential oil of Melaleuca alternifolia. Letters in Applied Microbiology 1994, 19, 24–25. [Google Scholar] [CrossRef]
- Thosar, N.; Basak, S.; Bahadure, R.N.; Rajurkar, M. Antimicrobial efficacy of five essential oils against oral pathogens: An in vitro study. European journal of dentistry 2013, 7, S071–S077. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.; Vitali, C.; Gallo, D.; Balenzano, L.; Mallamaci, R. The inhibition of Candida species by selected essential oils and their synergism with amphotericin B. Phytomedicine 2008, 15, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Oro, D.; Heissler, A.; Rossi, E.M.; Scapin, D.; da Silva Malheiros, P.; Boff, E. Antifungal activity of natural compounds against Candida species isolated from HIV-positive patients. Asian Pacific Journal of Tropical Biomedicine 2015, 5, 781–784. [Google Scholar] [CrossRef]
- Noumi, E.; Snoussi, M.; Hajlaoui, H.; Trabelsi, N.; Ksouri, R.; Valentin, E.; Bakhrouf, A. Chemical composition, antioxidant and antifungal potential of Melaleuca alternifolia (tea tree) and Eucalyptus globulus essential oils against oral Candida species. J. Med. Plants Res 2011, 5, 4147–4156. [Google Scholar]
- Mondello, F.; De Bernardis, F.; Girolamo, A.; Cassone, A.; Salvatore, G. In vivo activity of terpinen-4-ol, the main bioactive component of Melaleuca alternifolia Cheel (tea tree) oil against azole-susceptible and-resistant human pathogenic Candida species. BMC infectious diseases 2006, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.; Carson, C.; Riley, T. In vitro activity of Melaleuca alternifolia (tea tree) oil against dermatophytes and other filamentous fungi. Journal of Antimicrobial Chemotherapy 2002, 50, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.; Mann, C.; Markham, J. Interactions between components of the essential oil of Melaleuca alternifolia. Journal of applied microbiology 2001, 91, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Brambilla, E.; Ionescu, A.C.; Cazzaniga, G.; Ottobelli, M.; Samaranayake, L.P. Levorotatory carbohydrates and xylitol subdue Streptococcus mutans and Candida albicans adhesion and biofilm formation. Journal of basic microbiology 2016, 56, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudabadi, A.Z.; Zarrin, M.; Kiasat, N. Biofilm formation and susceptibility to amphotericin B and fluconazole in Candida albicans. Jundishapur journal of microbiology 2014, 7. [Google Scholar]
- Dalleau, S.; Cateau, E.; Berges, T.; Berjeaud, J.M.; Imbert, C. In vitro activity of terpenes against Candida biofilms. Int J Antimicrob Agents 2008, 31, 572–576. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Experimental sequences for this study. (A) General overview of the survey experimental sequence. (B) Diagram investigating the antifungal activity of TTO against Candida albicans. (C) Diagram investigating biofilm formation in Candida albicans. (D) Diagram determining MBIC in the study inhibiting biofilm formation of Candida albicans using TTO. (E) Diagram establishing MBEC in the study inhibiting biofilm formation of Candida albicans using TTO.
Figure 1.
Experimental sequences for this study. (A) General overview of the survey experimental sequence. (B) Diagram investigating the antifungal activity of TTO against Candida albicans. (C) Diagram investigating biofilm formation in Candida albicans. (D) Diagram determining MBIC in the study inhibiting biofilm formation of Candida albicans using TTO. (E) Diagram establishing MBEC in the study inhibiting biofilm formation of Candida albicans using TTO.
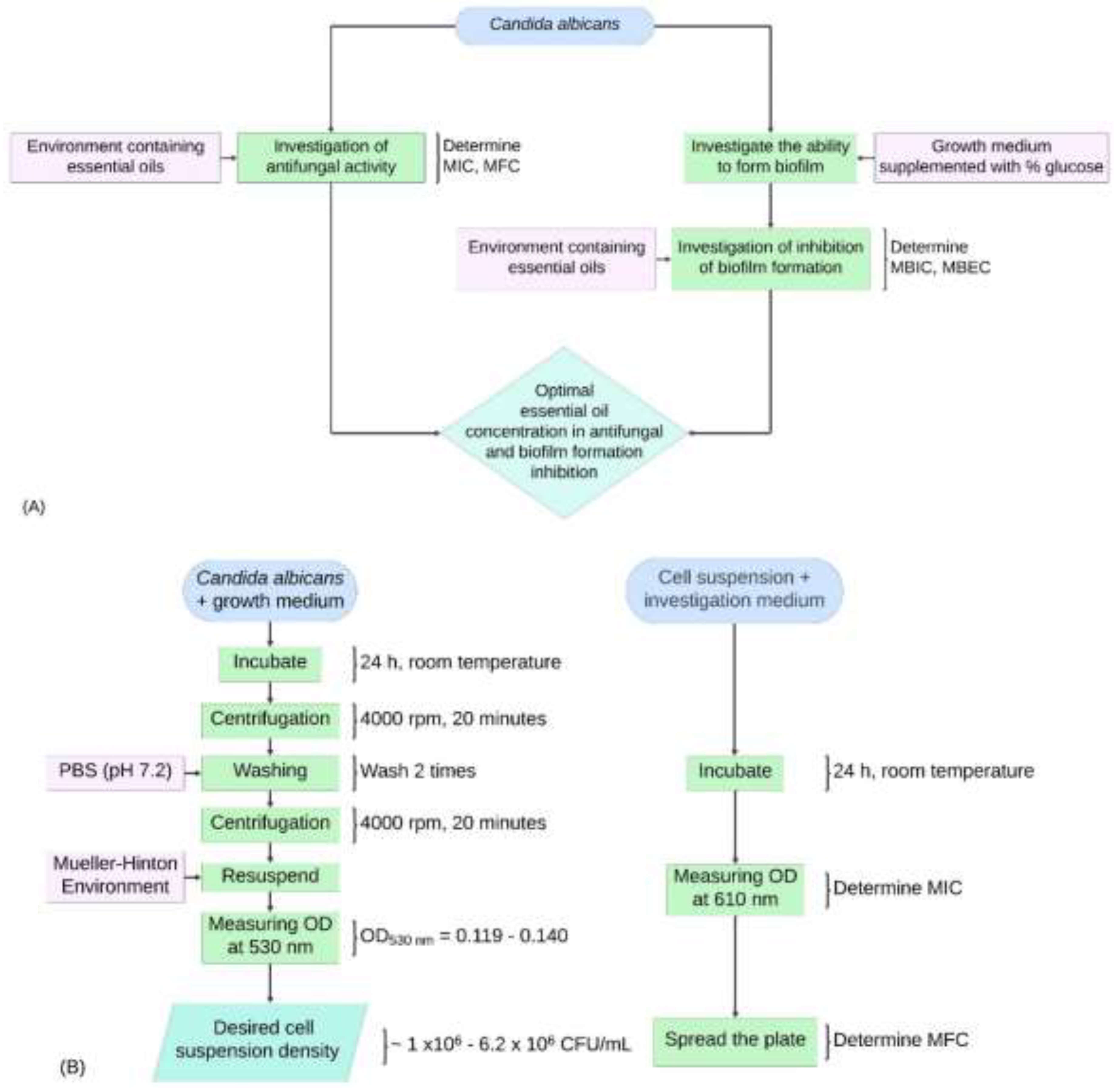
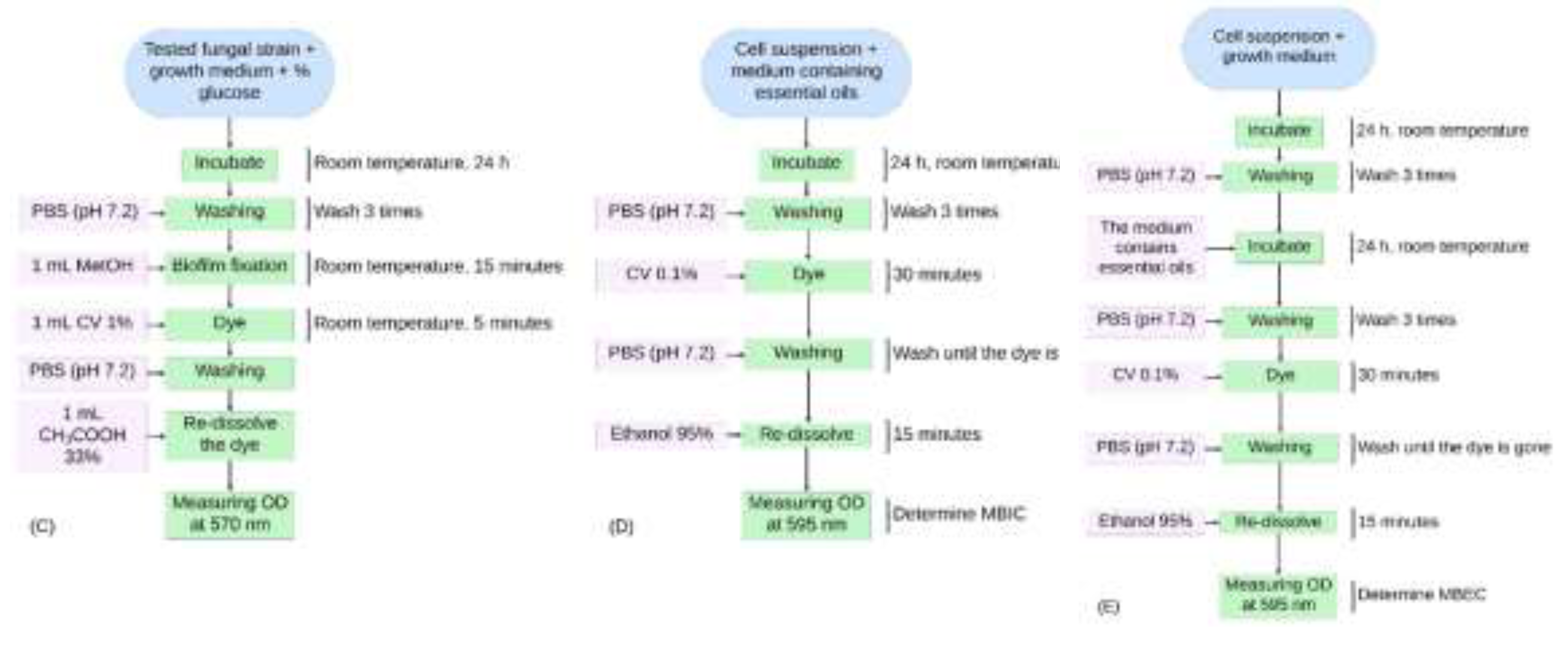
Figure 2.
Morphology of Candida albicans. (a) Microscopic morphology of Candida albicans using a simple staining method. (b) Cluster morphology of Candida albicans after A) 24 h and B) 48 h.
Figure 2.
Morphology of Candida albicans. (a) Microscopic morphology of Candida albicans using a simple staining method. (b) Cluster morphology of Candida albicans after A) 24 h and B) 48 h.
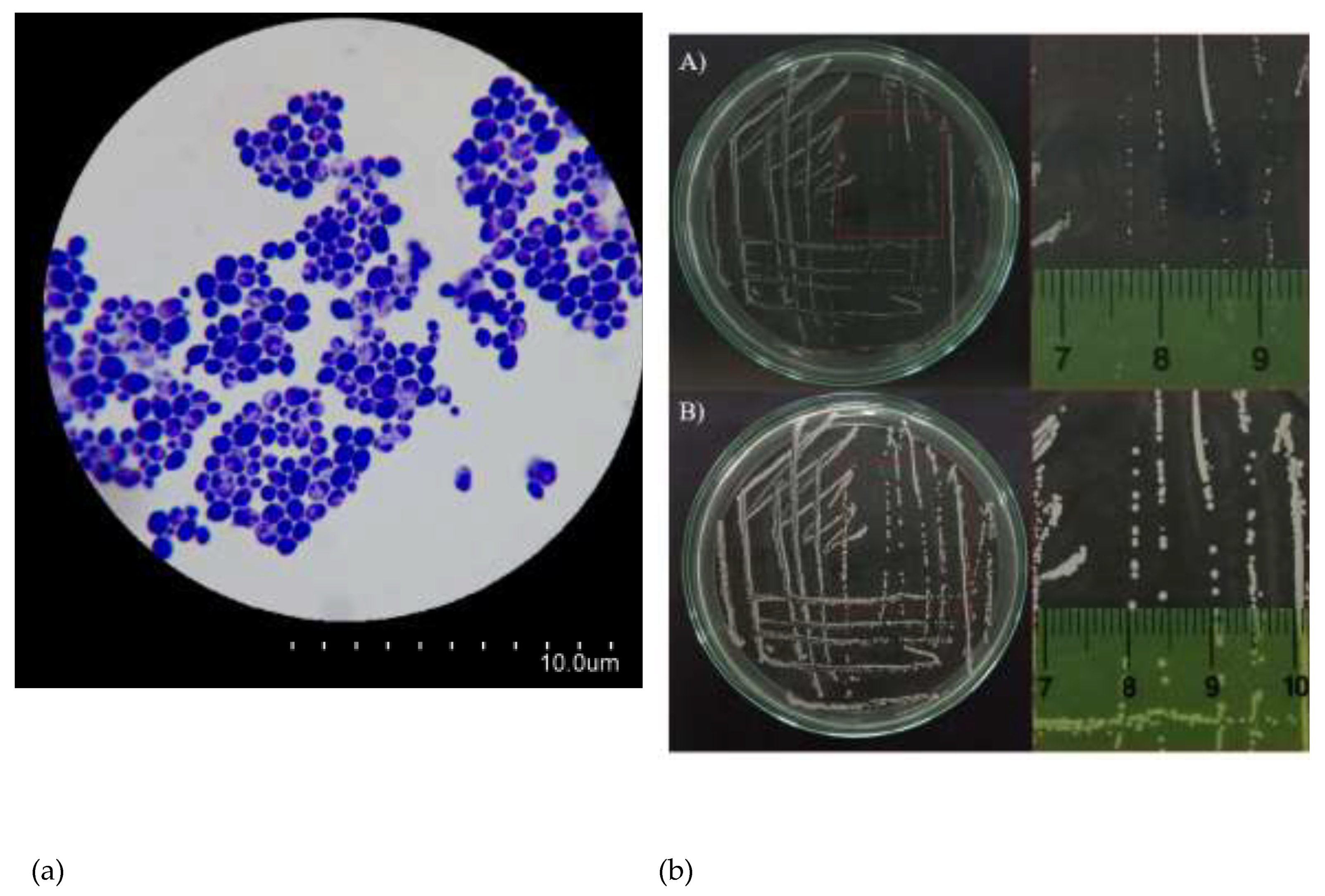
Figure 3.
Antifungal activity against Candida albicans of the antifungal agent AMB and TTO at various test concentrations. (A) MFC results are presented as follows: Images labeled a), b), c), d), e) show samples treated with the antifungal agent AMB at concentrations of 0.1, 1, 2, 5, and 10 μg/mL respectively; and f), g), h), i), k) display samples treated with TTO at concentrations of 0.1, 1, 2, 5, and 10 μL/mL respectively. (B) Graphical representation of concentration thresholds inhibiting fungal growth.
Figure 3.
Antifungal activity against Candida albicans of the antifungal agent AMB and TTO at various test concentrations. (A) MFC results are presented as follows: Images labeled a), b), c), d), e) show samples treated with the antifungal agent AMB at concentrations of 0.1, 1, 2, 5, and 10 μg/mL respectively; and f), g), h), i), k) display samples treated with TTO at concentrations of 0.1, 1, 2, 5, and 10 μL/mL respectively. (B) Graphical representation of concentration thresholds inhibiting fungal growth.
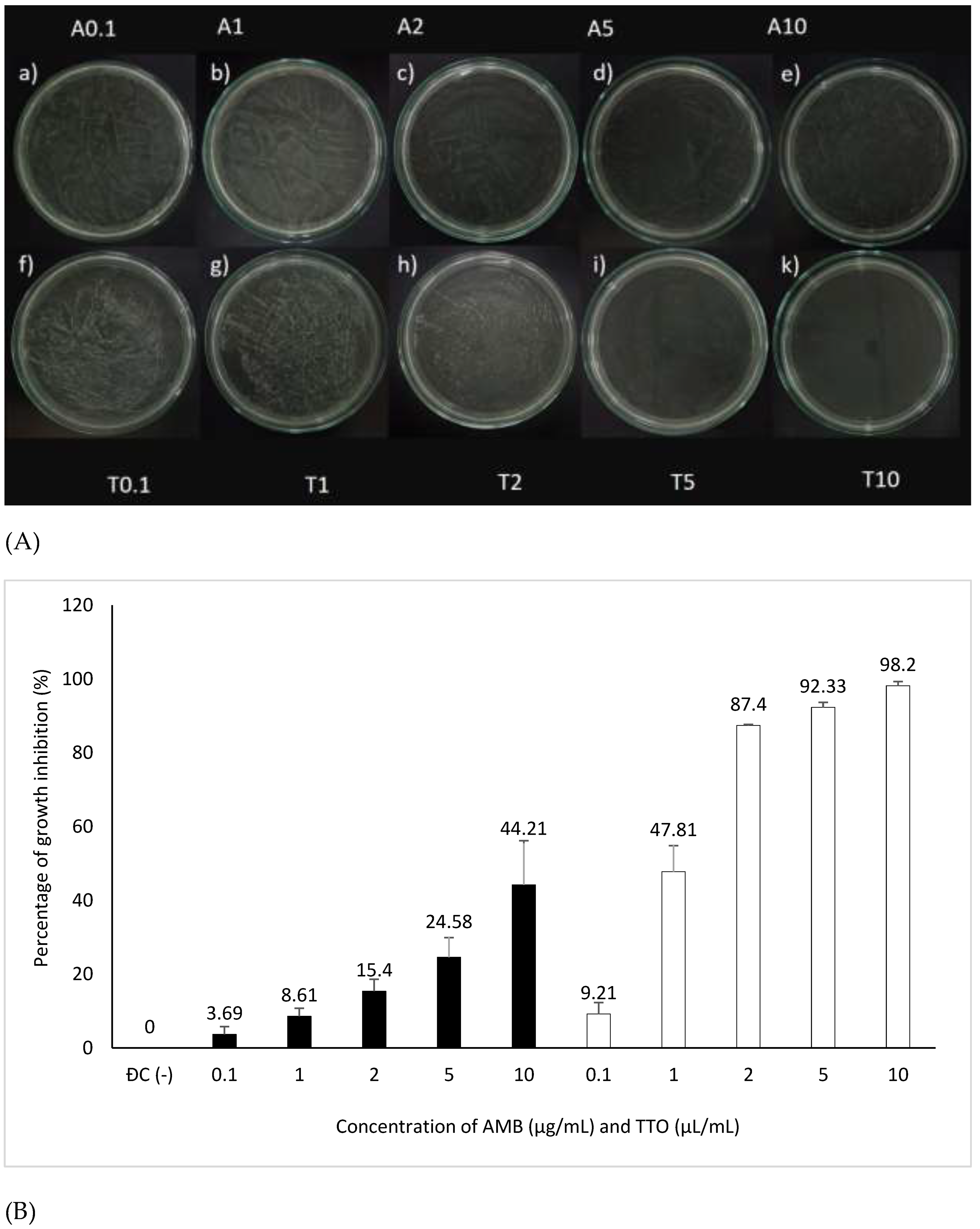
Figure 4.
Scanning Electron Microscopy (SEM) image of Candida albicans biofilm in the experimental condition of Hansen medium + 2.5% glucose. The marked region (green circle) highlights the biofilm formation.
Figure 4.
Scanning Electron Microscopy (SEM) image of Candida albicans biofilm in the experimental condition of Hansen medium + 2.5% glucose. The marked region (green circle) highlights the biofilm formation.
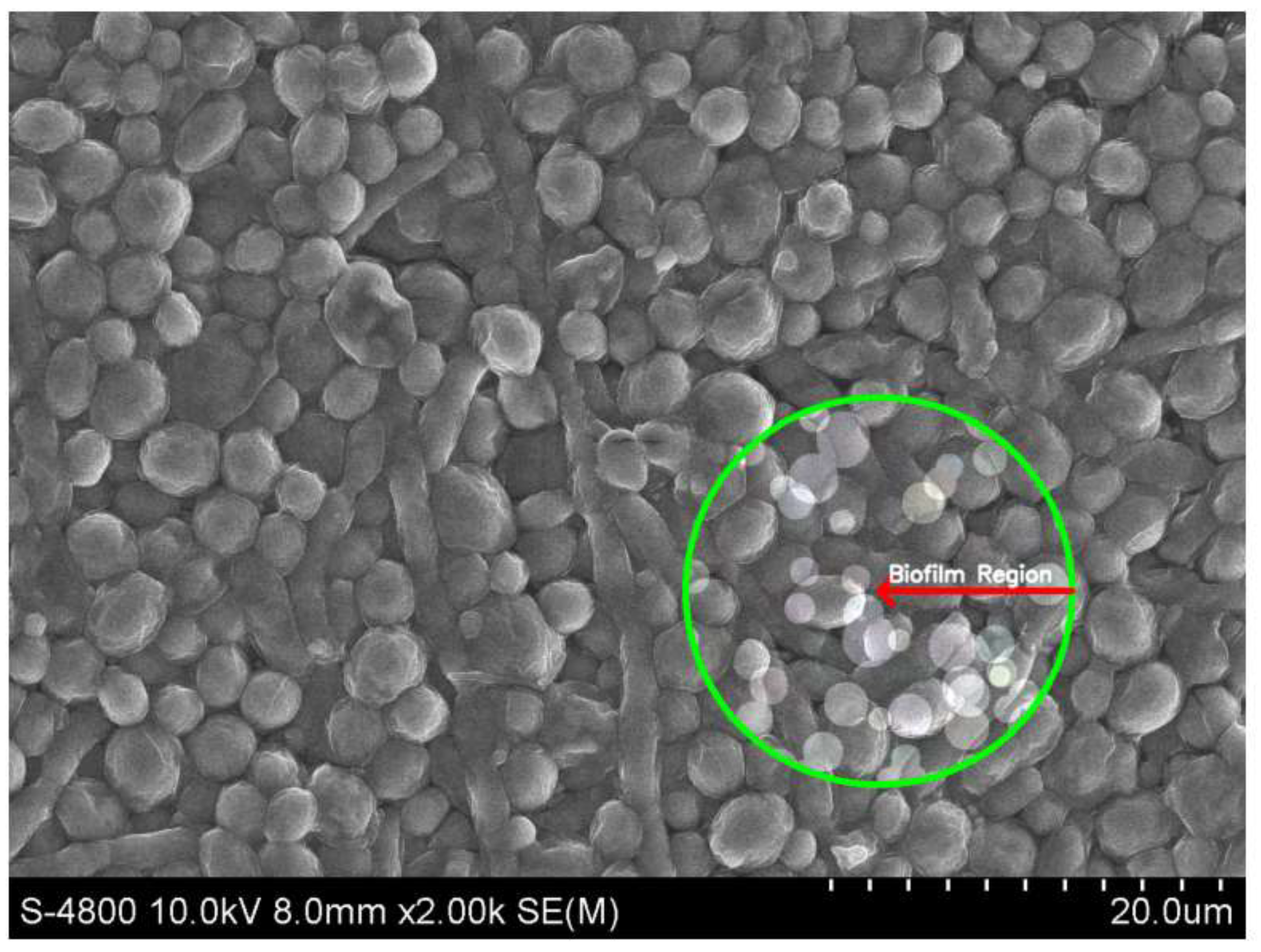
Figure 5.
Suppression of Candida albicans biofilm formation by AMB and TTO at different investigated concentrations (following MBIC). For AMB, statistically significant differences (p < 0.05) compared to the control included concentrations (μg/mL) of 0.1, 1, 2, 5, and 10; For TTO, statistically significant differences (p < 0.05) included concentrations (μL/mL) of 2, 5, and 10.
Figure 5.
Suppression of Candida albicans biofilm formation by AMB and TTO at different investigated concentrations (following MBIC). For AMB, statistically significant differences (p < 0.05) compared to the control included concentrations (μg/mL) of 0.1, 1, 2, 5, and 10; For TTO, statistically significant differences (p < 0.05) included concentrations (μL/mL) of 2, 5, and 10.
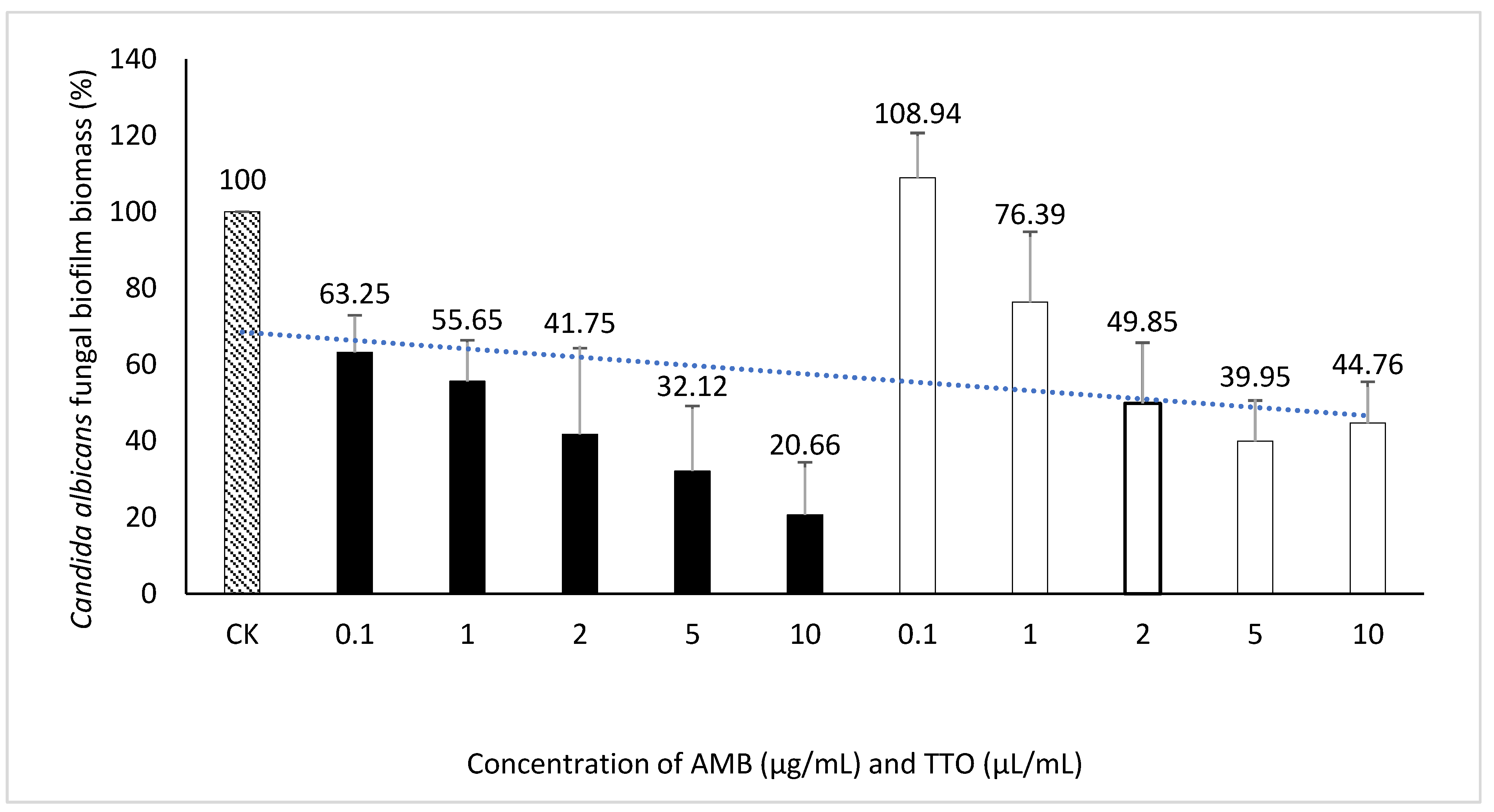
Figure 6.
Inhibition of Candida albicans biofilm formation by AMB and TTO at different test concentrations (following MBEC). For AMB, the statistically significant difference (p < 0.05) compared to the control had a threshold of only 10; For TTO, all concentrations were not statistically significant (p > 0.05).
Figure 6.
Inhibition of Candida albicans biofilm formation by AMB and TTO at different test concentrations (following MBEC). For AMB, the statistically significant difference (p < 0.05) compared to the control had a threshold of only 10; For TTO, all concentrations were not statistically significant (p > 0.05).
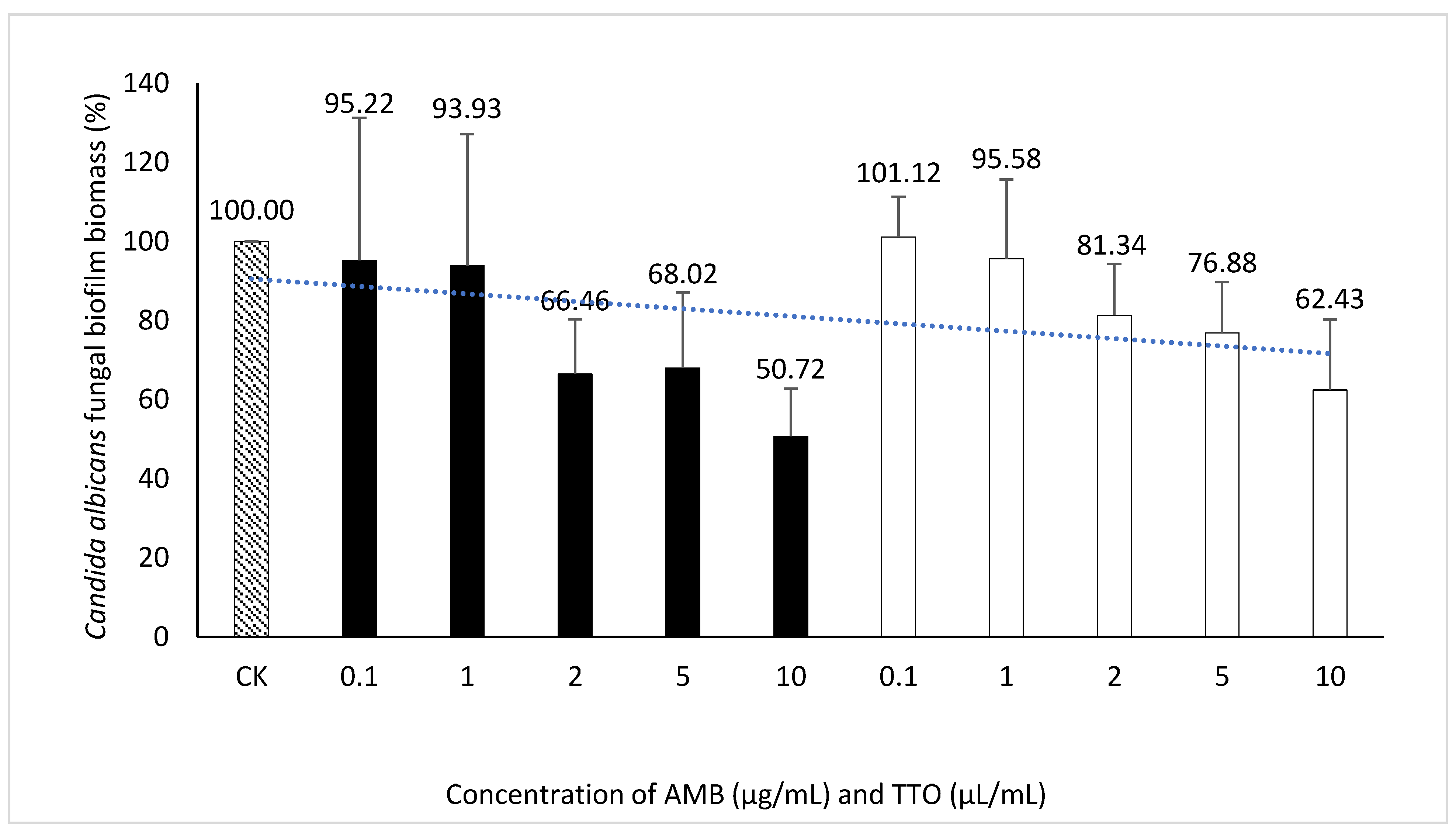

Table 1.
MIC and MFC of the AMB antifungal agent and TTO.
| Bioactive compounds | MIC (Mean ± SD) | MIC80 (Mean ± SD) | MFC (Mean ± SD) |
|---|---|---|---|
| AMB | 0.1 /mL (3.69 ± 2.09) | - | - |
| TTO | 0.1 /mL (9.21 ± 3.11) | 2 /mL (87.4 ± 0.27) | 10 /mL (98.2 ± 1.13) |
Note: (-): Not detected. The MIC and MFC values were determined as the average of three independent experiments, and the results are presented as mean ± standard deviation (SD) to ensure reproducibility and accuracy.
Table 2.
The mean ODs value at a wavelength of 570 nm.
| Trial | Symbol | The mean ODs value at a wavelength of 570 nm | ODc value |
|---|---|---|---|
| Hansen + 2.5% glucose | NT1 | 2.6390 1.6245 | 1.31270.3875 |
| Hansen + 5% glucose | NT2 | 2.4337 1.5600 | |
| Hansen + 10% glucose | NT3 | 2.2874 1.5124 |
Note: When comparing, the ODc value is taken to approximately two decimal places, ODc = 1.31.
Table 3.
The extent of biofilm formation by Candida albicans in the Hansen medium supplemented with varying percentages of glucose (w/v).
Table 3.
The extent of biofilm formation by Candida albicans in the Hansen medium supplemented with varying percentages of glucose (w/v).
| The extent of biofilm formation | Trials | ||
|---|---|---|---|
|
Hansen + 2.5% glucose (NT1) |
Hansen + 5% glucose (NT2) |
Hansen + 10% glucose (NT3) |
|
|
No biofilm producer ODs 1.31 |
- | - | - |
|
Weak biofilm producer 1.31 ODs 2.62 |
- | + | + |
|
Moderate biofilm producer 2.62 ODs 5.24 |
+ | - | - |
|
Strong biofilm producer 5.24 ODs |
- | - | - |
Note: (+): Biofilm formation at the corresponding level; (-): No biofilm formation at the corresponding level.
Table 4.
The MBIC and MBEC values for the antifungal agent AMB and TTO were determined in this study.
Table 4.
The MBIC and MBEC values for the antifungal agent AMB and TTO were determined in this study.
| Bioactive compounds | MBIC (Mean ± SD) | MBEC (Mean ± SD) |
|---|---|---|
| AMB | 2 /mL (41.75 ± 22.54) | - |
| TTO | 2 /mL (49.85 ± 15.91) | - |
Note: (-): Not detected. The MBIC and MBEC values were determined as the average of four independent experiments, and the results are presented as mean ± standard deviation (SD) to ensure reproducibility and accuracy.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



