
Submitted:
05 September 2024
Posted:
05 September 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
tRNA-derived fragments function as markers additionally to playing the role key of signalling molecules in a number of disorders. It is known that the repertoire of these molecules differs greatly in different cell types and varies depending on the physiological condition. The aim of our research was to compare the pattern of tRF expression in the main blood cell types and to determine how the composition of these molecules changes during COVID-19-induced cytokine storm. Erythrocytes, monocytes, lymphocytes, neutrophils, basophils, and eosinophils from control donors and patients with severe COVID-19 were obtained by fluorescence sorting. We extracted RNA from FACS sorted cells and performed NGS of short RNAs. The composition of tRNA-derived fragments was analysed applying a semi-custom bioinformatic pipeline. In this study, we assessed the length and type distribution of TRFs and reported the 150 most prevalent TRF sequences across all cell types. Additionally we demonstrate a significant (p<0.05, fold change>16) change in the pattern of tRFs in erythrocytes (21 downregulated, 12 upregulated), monocytes (53 downregulated, 38 upregulated) and lymphocytes (49 upregulated) in patients with severe COVID-19. Thus, different blood cell types exhibit a significant variety of TRFs and react to cytokine storm by dramatically changing their differential expression patterns. We suppose that the observed phenomenon occurs due to the regulation of nucleotide modifications and alterations in activity of various Rnases.
Keywords:
tRNA fragment
; blood cells
; severe COVID-19
; sRNA NGS
; RBC
; ymphocytes
; monocytes
1. Introduction
tRNA-derived fragments (tRFs) are a relatively recently discovered class of small RNA molecules that are shown to be involved in the pathogenesis of various diseases, particularly cancer and viral infections. Processed from transfer RNAs, tRFs regulate gene expression through various mechanisms like miRNA-like silencing of target mRNAs [1,2,3,4].
There are several types of classified tRFs based on the cleavage site within the tRNA molecule: tRF-1: Cleaved from the 3' trailer sequence of pre-tRNAs. tRF-2: Cleaved from the D-loop of mature tRNAs. tRF-3: Cleaved from the 3' end of mature tRNAs. tRF-5: Cleaved from the 5' end of mature tRNAs. i-tRF: Internal tRFs derived from the internal region of mature tRNAs [5].
Different tissues and organs possess a different set of tRNA fragments [2]. Nevertheless, it has not been studied how the pattern of these molecules differs in different blood cells. Supposably, the repertoire of these molecules changes in correspondence with changes in the immunological state of patients. In particular, it is not clear how these molecules will change in blood cells during the cytokine storm induced by COVID-19.
It has been previously reported by Wu et. al. that tRFs were the most significantly affected small non-coding RNAs in nasopharyngeal swabs of COVID-19 patients, they had also observed that SARS-CoV-2-infected airway epithelial cells exhibit the same tendency [4]. In an attempt to additionally shed light on tRNA-derived fragments and their involvement in host-virus interactions it seems relevant to study tRFs in different pathophysiological circumstances. In the case of this study our aim was to reveal the differential expression of tRFs in fluorescence activated sorted cells of healthy control donors and SARS-CoV-2 infected patients, as well as to observe and characterise tRFs during viral infections.
2. Materials and Methods
2.1. Patients and Data Collection
Six healthy donors and five RT-PCR confirmed SARS-CoV-2 positive patients made up the two groups of research participants. Three patients were hospitalised to the critical care unit and two patients were moved to the infectious disease unit. Severe patients satisfied the following criteria for admission to the intensive care unit (ICU): body temperature ≥ 39 °C, respiration rate ≥ 30/min, and oxygen saturation (SpO2) ≤ 93%.
2.2. Cell Sorting
Сell sorting was performed on a MoFlow Astrios EQ device (Beckman Coulter). Erythrocytes were isolated using antibodies against CD235 protein (Beckman Coulter , IM2212U). To isolate leukocytes, erythrocytes were first lysed with VersaLyse lysing solution (Beckman Coulter). Granulocytes (neutrophils, basophils and eosinophils) were isolated using duraclone IM granulocyte antibody panel (B88651, Beckman Coulter). Lymphocytes and monocytes were isolated using antibodies CD45 (A79392 Beckman Coulter), CD16 (6607118 Beckman Coulter), CD14 (A70204 Beckman Coulter, USA). Cell sample purity for all cell populations was >95% according to flow cytometry data. A detailed description of cell sorting protocols is provided in the following article [6].
2.3. RNA Separation, Library Preparation and Next Generation Sequencing
RNA was isolated\extracted using ExtractRNA reagent (Evrogen) according to the provided protocol. After extraction, RNA was dissolved in 10 µl RNAse free water. The quality of the obtained RNA was checked on a TapeStation (Agilent). Only samples with RIN>5 were taken for sequencing. Short RNA libraries were prepared using the Small RNA Library Prep Kit (BGI, 1000006383). Sequencing was performed on a DNBSEQ-G400 instrument (BGI)
2.4. Bioinformatics
A general design of the bioinformatic processing pipeline is presented in Figure 1.
Read Quality Control
For quality control we applied FastQC [7]. All datasets qualified quality control standards. Each FastQ file contained between 17 and 27 million reads.
Adapter Trimming
Adapter trimming was executed using Trimmomatic [8]. Long clipping sequence was set as AGTCGGAGGCCAAGCGGTCTTAGGAAGACAA. We opted to use optimal run settings for SE (single-end) reads (TruSeq3-SE:2:30:10 LEADING:3 TRAILING:3 SLIDINGWINDOW:4:15 MINLEN:20) which allowed us to minimise execution time with no data loss. As an input, trimmomatic takes the adapter.fa file, in which our adapter sequence is registered. For each FastQ file from 50 to 68% of the input reads were marked as dropped after running Trimmomatic.
Read Mapping, Count and Normalisation
Full genome alignment and transriptome-based pseudoalignment methods are not applicable for small-RNA-seq identification due to tRNA cleavage events, which led us to utilising "BLAST"(Blast Local Alignment Search Tool) [9].
We implemented BLAST through “tRNAExplorer" [10], a Python-based pipeline, optimised for tRF-profile analysis. In order to utilise tRNAExplorer, we needed a *.bed genome annotation file as a reference, which we would BLAST our reads *.fa files against. While there is an option to generate a custom annotation file, we used an already compiled grch.38 database which came within the tRNAExplorer package. To run the pipeline, we created project directories with sample lists individually for each type of cells and custom configuration file, with data directories paths and launch options. While tRNAExplorer supports trimming and QC, we omitted these steps due to previous data processing. As an input each run of the pipeline takes all the *.fa files with read data from the specified directory, list of samples, and path to database *.bed file. We automated these runs with another custom Python script. As a final result of data processing we obtained *.csv and *.tsv text files, containing BLAST run results for every sample, cleavage sites data and a collective read count table for all provided samples with annotations.
This data was already sufficient for differential expression analysis and visualisation, but we applied another extra step, running a custom Python script which organises data and performs statistical tests.
2.5. Statistics
Wilcoxon–Mann–Whitney test was used to perform statistical analysis on the data. P-values less than 0.05 were considered statistically significant. The p-values for the RNAseq volcano graphics were log converted to log10 (1.31) for data visualisation.
3. Results
3.1. tRF Length Distribution in Main Cell Types
Analysing tRF length distribution in erythrocytes, monocytes, lymphocytes, neutrophils, basophils and eosinophils we observed a remarkable similarity between control and severe COVID-19 patient’s cells (Figure 2). This data is additionally supported by our heatmap, which demonstrated a distinguishable pattern of tRF expression depending on cell type. Erythrocytes are presented as the most distinguishable group of cells in terms of tRF length and tRF expression patterns, which may be due to the fact that the erythrocyte is an inactive cell in terms of synthetic processes.
3.2. tRF Type Composition in Main Cell Types
Analysing the main tRF types in sorted lymphocytes of control donors and severe COVID-19 patients we observed the prevalence of tRF-5 in 7 out of 8 samples. 1 severe COVID-19 patient exhibited dominating above all presence of i-tRF and tRF-3 types (Figure 3). It is worth noting that this patient had extremely high levels of IL-6 - 2 398 (pg/ml) on the same day that this sample was taken.
3.3. tRF Expression in Control Donors and Severe COVID-19 Patients
To compare tRF expression in different cell types we calculated the 150 most abundant tRFs based on RAW read counts, log normalised for data presentation. The heatmap demonstrates a relatively vivid frontier for erythrocytes (Figure 4).
3.4. tRF Differential Expression in Erythrocytes
3.5. tRF Differential Expression in Lymphocytes
3.6. tRF Differential Expression in Monocytes
4. Discussion
This pilot study aimed at demonstrating a distinguishable tRF pattern for different sorted cell types. Given the fact that tRFs have been discovered relatively recently, research aimed at understanding their origin can help elucidate their biological purposes.
Analysing tRF length distribution in erythrocytes, monocytes, lymphocytes, neutrophils, basophils and eosinophils we observed a remarkable similarity between control and severe COVID-19 patient’s cells (Figure 2). This data is additionally supported by our heatmap (Figure 4), which demonstrated a distinguishable pattern of tRF expression depending on cell type. Erythrocytes are presented as the most distinguishable group of cells in terms of tRF length and tRF expression patterns, which may be due to the fact that the erythrocyte is an inactive cell in terms of synthetic processes.
Regarding tRF type composition, It has been previously noted that 3-tRFs are produced in response to various cellular stresses like oxidative stress, hypoxia, and viral infection [11]. Some 3-tRFs can inhibit viral replication by interfering with viral gene expression or packaging. They can act as signalling molecules to mediate stress responses [12,13].
Given the fact that IL-6 is a key mediator of the "cytokine storm" that leads to acute respiratory distress syndrome (ARDS) and multi-organ dysfunction in severe COVID-19 [14], it is assumable that such distribution of tRF types may be due the condition the patient underwent. Other cell types did not exhibit any significant alterations in tRF composition.
Overlooking tRF expression in control donors and severe COVID-19 patients, it is worth noting how the expression is increased in the majority of tRF in granulocytes (neutrophils, basophils, eosinophils). A similar tendency to increased expression is observed in monocytes. Granulocytes and monocytes both originate from CFU-GM (Colony Forming Unit–Granulocyte–Macrophage), also known as granulocyte–macrophage progenitor (GMP) [15].
The aforementioned suggests that tRFs may dominantly persist from the progenitor stages of cell development, and are only slightly modified by environmental or physiological factors.
In our study we encountered fragments which have been previously associated with several types of cancer.
Lymphocytic tRFs: It is noted that tRF#31-R29P4P9L5HJVE previously has been acknowledged as a marker for lung cancer prediction among smokers 10 years prior to being diagnosed [19].
tRF#19-VKS4I71Z has been mentioned as an abundant novel-trf in a 2016 study of RNA-seq data from human prostate tissue [20].
In a 2023 research dedicated to studying tRFs in cancer, a high level tRF#34-5QKDN6QQ1362HQ has been mentioned as a predictor of breast cancer improved survival [2].
Monocytic tRFs: tRF#22-WEK6S1852 in previous research was found to be significantly downregulated and associated with human malignant mesothelioma [16].
A 2019 colon cancer study revealed significantly differentially expressed tRFs between colon cancer tissues and peritumor tissues, whereas another upregulated tRF that we observed - tRF#22-9LON4VN11 was mentioned, demonstrating downregulation in colon cancer tissues with a log2 fold change of -1.26 [17].
Another fragment we observed - tRF#18-HSQSD2D2, downregulation of which has been previously associated with early-stage breast cancer [2].
In a study devoted to examining the dysregulation of different tRFs in chronic lymphocytic leukemia - tRF-20-RK9P4P9L was amongst the top 15 differentially expressed sRNAs in aggressive chronic lymphocytic leukemia vs. normal controls, with the linear fold change being −76.64 and −258.08, respectively. Samples were composed of CD5+/CD19+ B cells. It is worth noting that in our study, the same tRF was also significantly downregulated but only in CD14++ CD16− monocytes [18].
tRF#22-WEPSJR852 was found in peripheral blood of fibromyalgia patients in a dissertation dedicated to the search of morphological substrate to fibromyalgia [21].
Erythrocytes appeared to be the only cell type which showed differential expression of tRFs that have not previously been associated with any type of cancer or disease. The fact that the erythrocyte is an inactive cell in terms of synthetic processes may elucidate these findings, however, regarding different cell types, tRFs and their interrelations with the conditions mentioned earlier it is more likely to suppose a coincidence, rather than some significant finding.
Differences in the length, type, and composition of specific tRFs between different cells are supposedly due to the specificity of the tRNA cleavage system. The same can be said about changes that occur during cytokine storm. The regulation system of tRNA processing is still poorly understood.
Nonetheless, we can say that it occurs at several “levels”. First of all - the regulation of nucleotide modifications in certain molecules [22]. Another stage is dependent on the regulation of RNAses that cleave tRNAs at specific sites.
The main types of nucleases involved in tRF biogenesis are: Dicer - cleaves 5' ends of mature tRNAs. Angiogenin (ANG) - cleaves mature tRNAs at anticodon loops. RNase Z/ELAC2 - cleaves 3' trailer sequences of pre-tRNAs [11,23,24].
RNase P: excision of external transcribed spacer (ETS) and internal transcribed spacers (ITS) from pre-tRNA transcripts [25].
Regulation of these enzymes can occur at the level of transcription initiation, as well as post-translational modifications of the enzyme
It is also likely that tRNA cleavage is regulated by proteins that bind this molecule [26] and make certain sites inaccessible for the aforementioned process. Unfortunately, we cannot state by what mechanism the difference between tRFs in blood cells is regulated.
In summary, it should be noted that TRF profiles significantly differ in different types of blood cells and demonstrate dramatic differential expression (sometimes more than 500-fold) during cytokine storm. Such profound differences suggest a major role of tRNA-derived fragments in the functioning of blood cells.
5. Conclusions
The composition of tRFs in erythrocytes monocytes, lymphocytes, neutrophils, basophils and eosinophils was analysed on the basis of sRNA-SEQ. We demonstrate notable alterations in the length and types of these molecules in main blood cell populations. We additionally observed a significant change in the profile of tRFs in the erythrocytes, monocytes and lymphocytes of patients infected with SARS-CoV-2.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. All RAW read data and nucleotide composition of tRFs are presented in two supplementary files: “Control cells” for control donors and “COVID cells” for patients with severe COVID-19.
Funding
This work was supported by St. Petersburg State University, project ID: 95412780.
Institutional Review Board Statement
This study was conducted according to the guidelines of the declaration of Helsinki and approved by the ethics committee.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, V.N. Small RNAs: Classification, Biogenesis, and Function. Molecules and Cells 2005, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Gu, J.; Wang, M.; Zhang, J.; Chen, Y.; Jiang, P.; Zhu, T.; Zhang, X. Emerging Roles of tRNA-Derived Fragments in Cancer. Molecular Cancer 2023, 22, 30. [Google Scholar] [CrossRef] [PubMed]
- Krishna, S.; Raghavan, S.; DasGupta, R.; Palakodeti, D. tRNA-Derived Fragments (tRFs): Establishing Their Turf in Post-Transcriptional Gene Regulation. Cell. Mol. Life Sci. 2021, 78, 2607–2619. [Google Scholar] [CrossRef]
- Wu, W.; Choi, E.-J.; Wang, B.; Zhang, K.; Adam, A.; Huang, G.; Tunkle, L.; Huang, P.; Goru, R.; Imirowicz, I.; et al. Changes of Small Non-Coding RNAs by Severe Acute Respiratory Syndrome Coronavirus 2 Infection. Front. Mol. Biosci. 2022, 9. [Google Scholar] [CrossRef]
- Yu, X.; Xie, Y.; Zhang, S.; Song, X.; Xiao, B.; Yan, Z. tRNA-Derived Fragments: Mechanisms Underlying Their Regulation of Gene Expression and Potential Applications as Therapeutic Targets in Cancers and Virus Infections. Theranostics 2021, 11, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Kondratov, K.A.; Artamonov, A.A.; Nikitin, Y.V.; Velmiskina, A.A.; Mikhailovskii, V.Y.; Mosenko, S.V.; Polkovnikova, I.A.; Asinovskaya, A.Y.; Apalko, S.V.; Sushentseva, N.N.; et al. Revealing Differential Expression Patterns of piRNA in FACS Blood Cells of SARS-CoV−2 Infected Patients. BMC Medical Genomics 2024, 17, 212. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. [Online] 2010. Available online at: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J Mol Biol 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sun, Z.; Tan, J.; Zhao, M.; Peng, Q.; Zhou, M.; Zuo, S.; Wu, F.; Li, X.; Dong, Y.; Xie, M.; et al. Integrated Genomic Analysis Reveals Regulatory Pathways and Dynamic Landscapes of the tRNA Transcriptome. Sci Rep 2021, 11, 5226. [Google Scholar] [CrossRef]
- Zhang, S.; Yu, X.; Xie, Y.; Ye, G.; Guo, J. tRNA Derived Fragments:A Novel Player in Gene Regulation and Applications in Cancer. Front Oncol 2023, 13, 1063930. [Google Scholar] [CrossRef]
- Kumar, P.; Kuscu, C.; Dutta, A. Biogenesis and Function of Transfer RNA Related Fragments (tRFs). Trends Biochem Sci 2016, 41, 679–689. [Google Scholar] [CrossRef]
- Gong, M.; Deng, Y.; Xiang, Y.; Ye, D. The Role and Mechanism of Action of tRNA-Derived Fragments in the Diagnosis and Treatment of Malignant Tumors. Cell Communication and Signaling 2023, 21, 62. [Google Scholar] [CrossRef]
- McGonagle, D.; Sharif, K.; O’Regan, A.; Bridgewood, C. The Role of Cytokines Including Interleukin-6 in COVID-19 Induced Pneumonia and Macrophage Activation Syndrome-Like Disease. Autoimmun Rev 2020, 19, 102537. [Google Scholar] [CrossRef]
- Guzzi, N.; Cieśla, M.; Ngoc, P.C.T.; Lang, S.; Arora, S.; Dimitriou, M.; Pimková, K.; Sommarin, M.N.E.; Munita, R.; Lubas, M.; et al. Pseudouridylation of tRNA-Derived Fragments Steers Translational Control in Stem Cells. Cell 2018, 173, 1204–1216.e26. [Google Scholar] [CrossRef]
- Filetti, V.; La Ferlita, A.; Di Maria, A.; Cardile, V.; Graziano, A.C.E.; Rapisarda, V.; Ledda, C.; Pulvirenti, A.; Loreto, C. Dysregulation of microRNAs and tRNA-Derived ncRNAs in Mesothelial and Mesothelioma Cell Lines after Asbestiform Fiber Exposure. Sci Rep 2022, 12, 9181. [Google Scholar] [CrossRef]
- Xiong, W.; Wang, X.; Cai, X.; Xiong, W.; Liu, Y.; Li, C.; Liu, Q.; Qin, J.; Li, Y. Identification of tRNAderived Fragments in Colon Cancer by Comprehensive Small RNA Sequencing. Oncology Reports 2019, 42, 735–744. [Google Scholar] [CrossRef]
- Veneziano, D.; Tomasello, L.; Balatti, V.; Palamarchuk, A.; Rassenti, L.Z.; Kipps, T.J.; Pekarsky, Y.; Croce, C.M. Dysregulation of Different Classes of tRNA Fragments in Chronic Lymphocytic Leukemia. Proc Natl Acad Sci U S A 2019, 116, 24252–24258. [Google Scholar] [CrossRef]
- Umu, S.U.; Langseth, H.; Zuber, V.; Helland, Å.; Lyle, R.; Rounge, T.B. Serum RNAs Can Predict Lung Cancer up to 10 Years Prior to Diagnosis. eLife 2022, 11, e71035. [Google Scholar] [CrossRef]
- Olvedy, M.; Scaravilli, M.; Hoogstrate, Y.; Visakorpi, T.; Jenster, G.; Martens-Uzunova, E.S. A Comprehensive Repertoire of tRNA-Derived Fragments in Prostate Cancer. Oncotarget 2016, 7, 24766–24777. [Google Scholar] [CrossRef]
- Erbacher, C. Systemic and Local Mechanisms of Small Fiber Pathology in Female Patients with Fibromyalgia Syndrome, Universität Würzburg, 2023.
- Lyons, S.M.; Fay, M.M.; Ivanov, P. The Role of RNA Modifications in the Regulation of tRNA Cleavage. FEBS Letters 2018, 592, 2828–2844. [Google Scholar] [CrossRef]
- Kazimierczyk, M.; Wojnicka, M.; Biała, E.; Żydowicz-Machtel, P.; Imiołczyk, B.; Ostrowski, T.; Kurzyńska-Kokorniak, A.; Wrzesinski, J. Characteristics of Transfer RNA-Derived Fragments Expressed during Human Renal Cell Development: The Role of Dicer in tRF Biogenesis. International Journal of Molecular Sciences 2022, 23, 3644. [Google Scholar] [CrossRef]
- Liu, B.; Cao, J.; Wang, X.; Guo, C.; Liu, Y.; Wang, T. Deciphering the tRNA-Derived Small RNAs: Origin, Development, and Future. Cell Death Dis 2021, 13, 1–13. [Google Scholar] [CrossRef]
- Li, Z.; Stanton, B.A. Transfer RNA-Derived Fragments, the Underappreciated Regulatory Small RNAs in Microbial Pathogenesis. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Karanasios, E.; Simos, G. Building Arks for tRNA: Structure and Function of the Arc1p Family of Non-Catalytic tRNA-Binding Proteins. FEBS Letters 2010, 584, 3842–3849. [Google Scholar] [CrossRef]
Figure 1.
NGS data analysis pipeline.
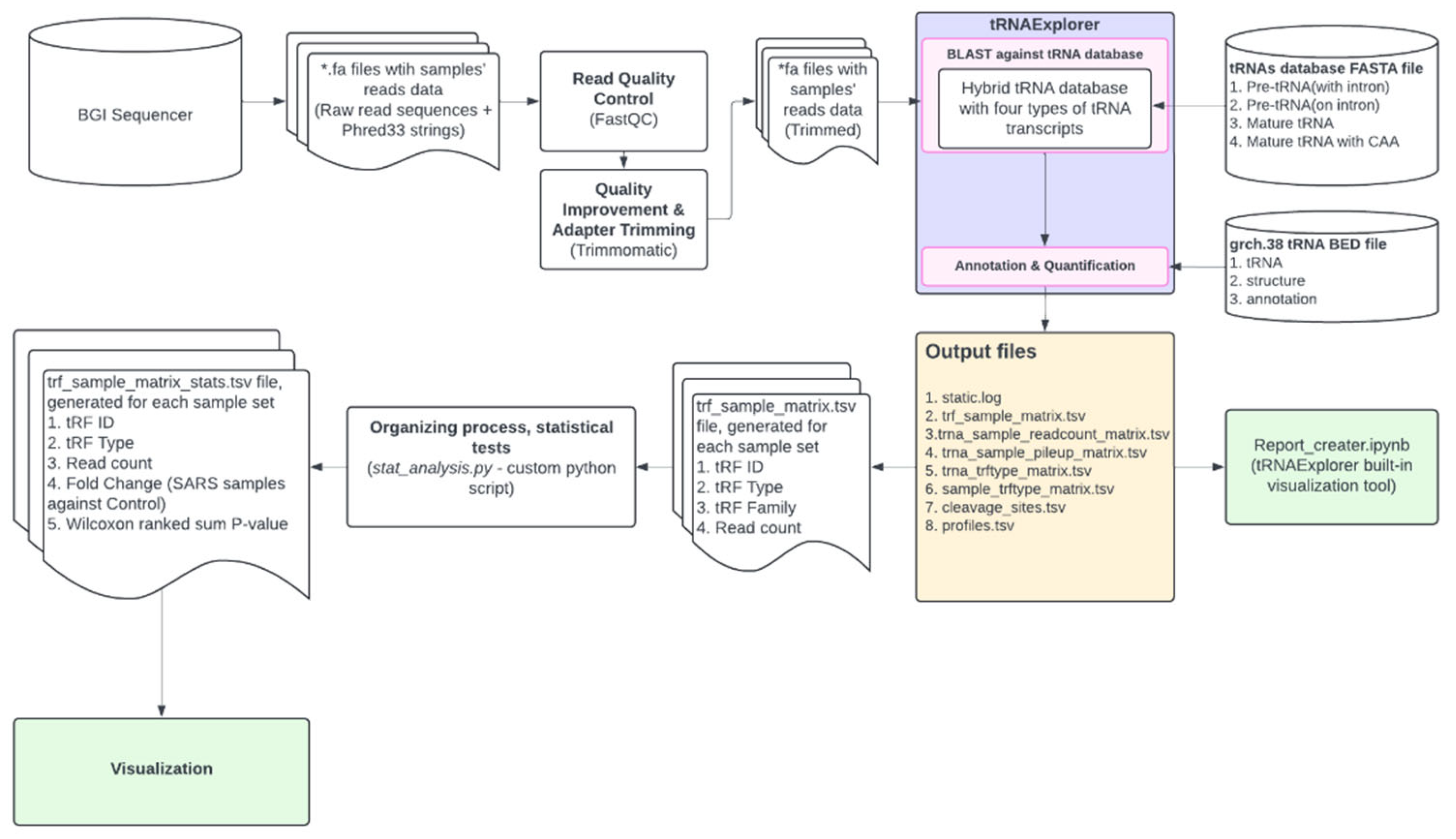
Figure 2.
tRF length distribution in main cell types (red line represents median value).
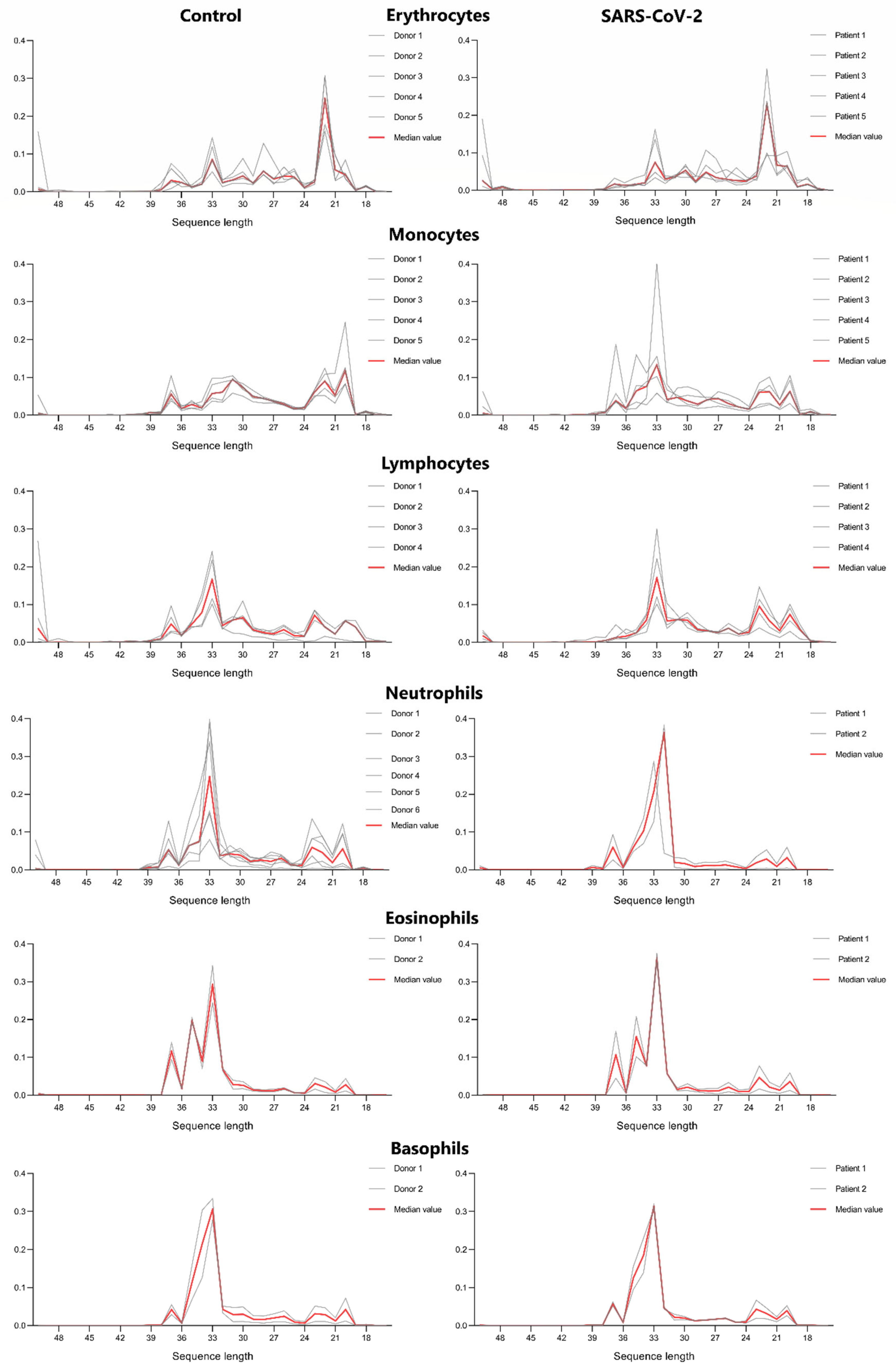
Figure 3.
tRF type composition in main cell types (colour matches tRF types).
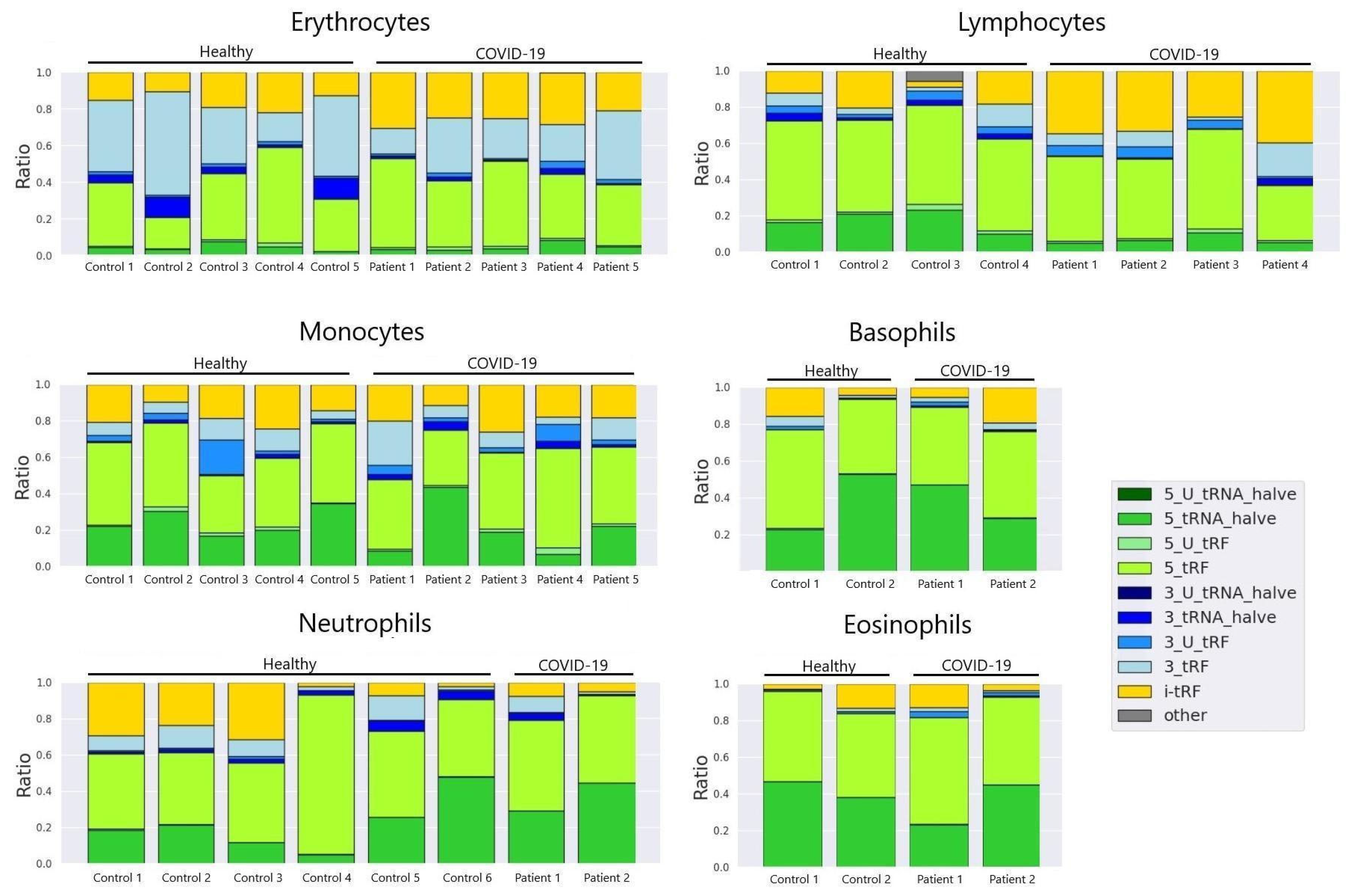
Figure 4.
tRF expression in control donors (C) and severe COVID-19 (S) patients. (Lym - lymphocytes, Mon - monocytes, Ery - erythrocytes, Neu - neutrophils, Bas - basophils, Eos - eosinophils).
Figure 4.
tRF expression in control donors (C) and severe COVID-19 (S) patients. (Lym - lymphocytes, Mon - monocytes, Ery - erythrocytes, Neu - neutrophils, Bas - basophils, Eos - eosinophils).
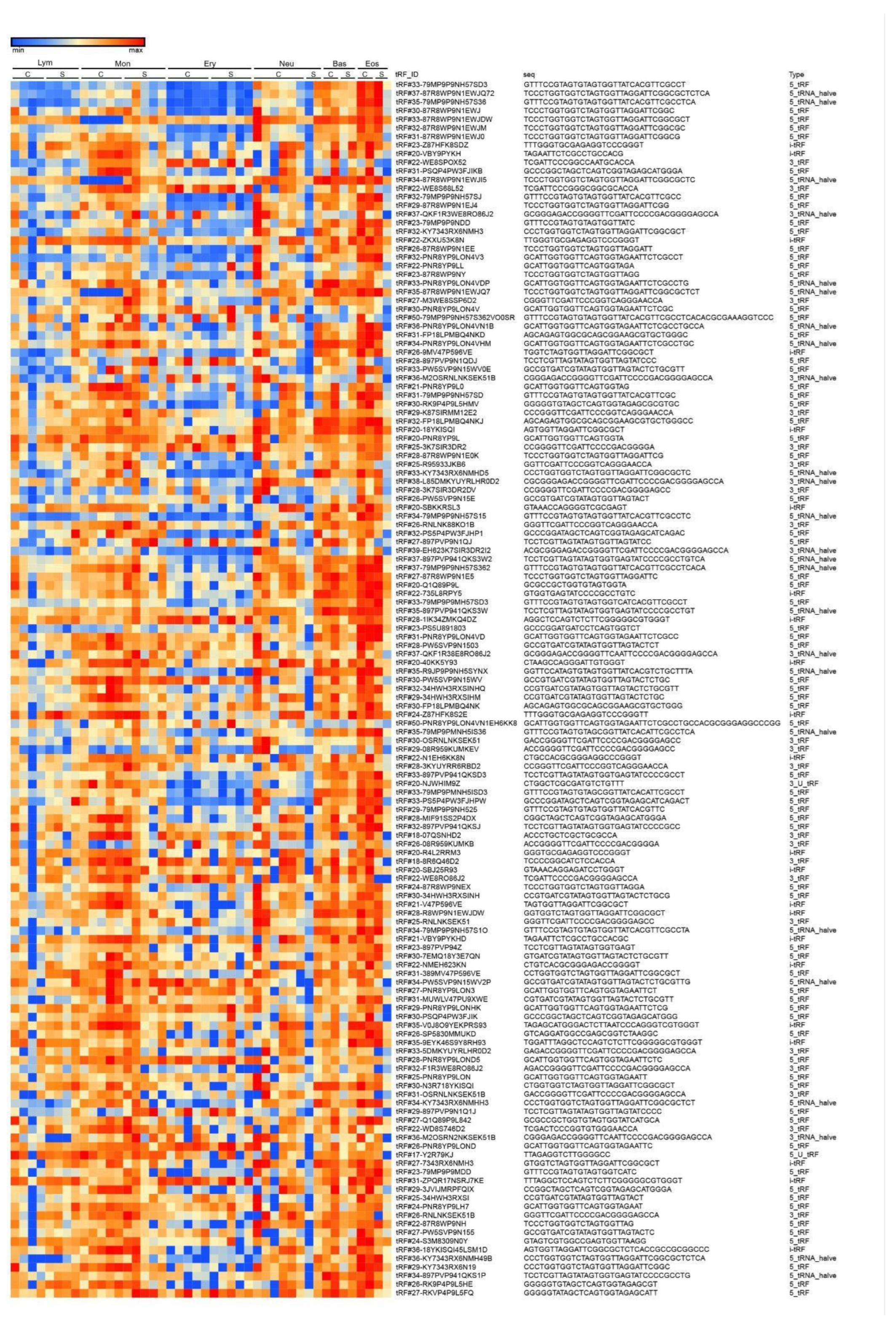
Figure 5.
tRF differential expression in erythrocytes (For all volcano plots we applied log4 (instead of log2) for fold change presentation.).
Figure 5.
tRF differential expression in erythrocytes (For all volcano plots we applied log4 (instead of log2) for fold change presentation.).
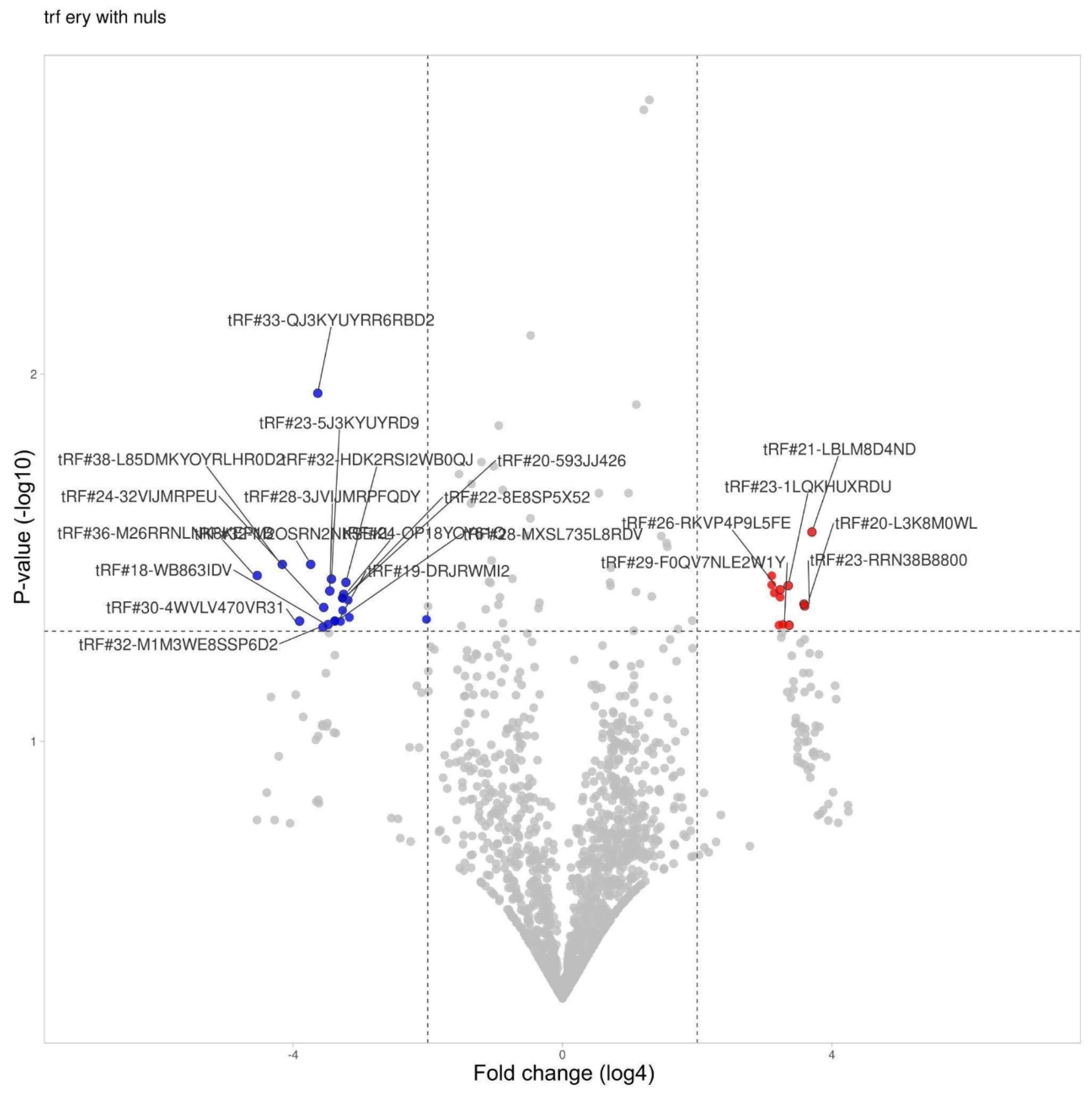
Figure 6.
tRF differential expression in lymphocytes.
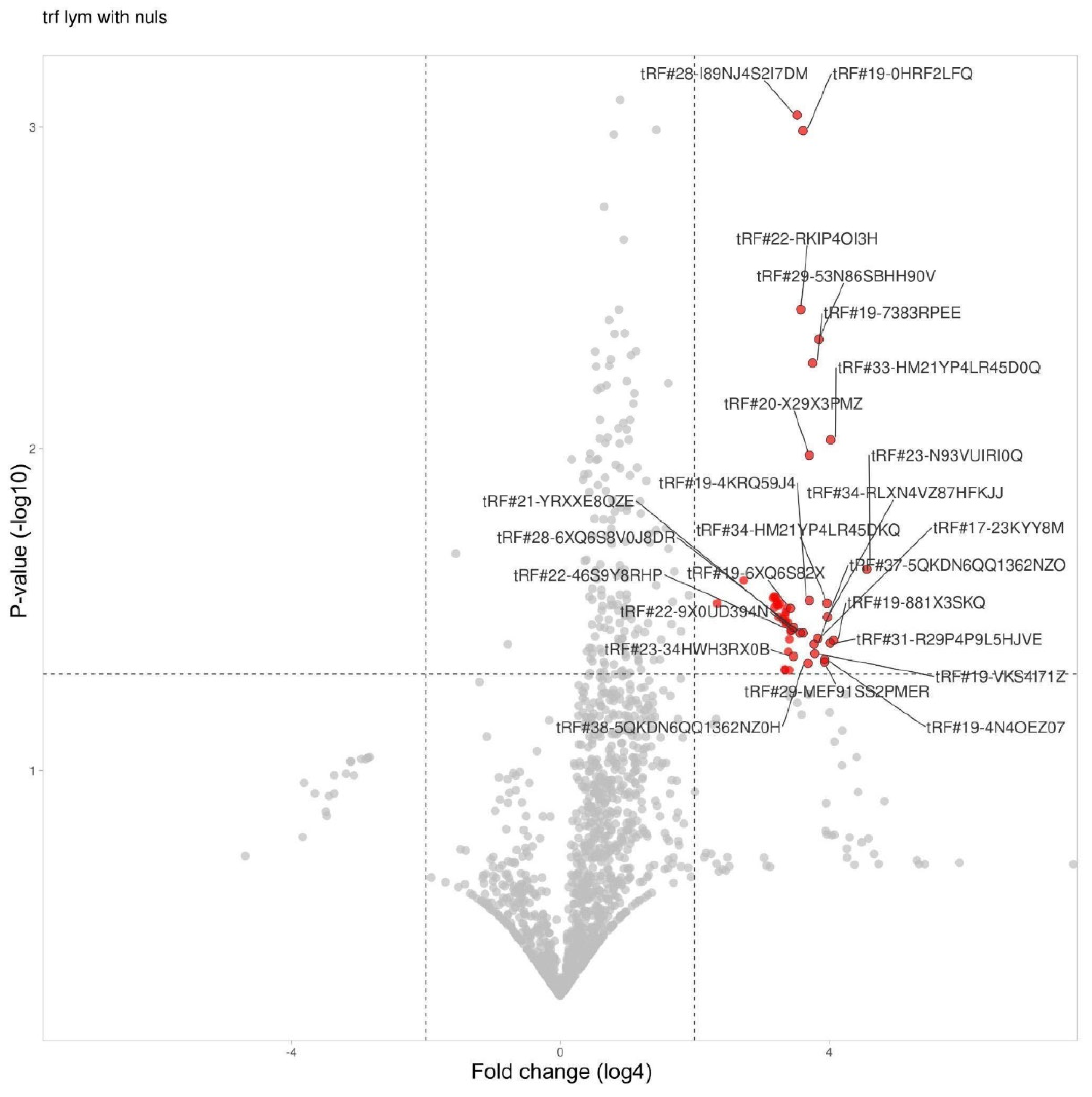
Figure 7.
tRF differential expression in monocytes.
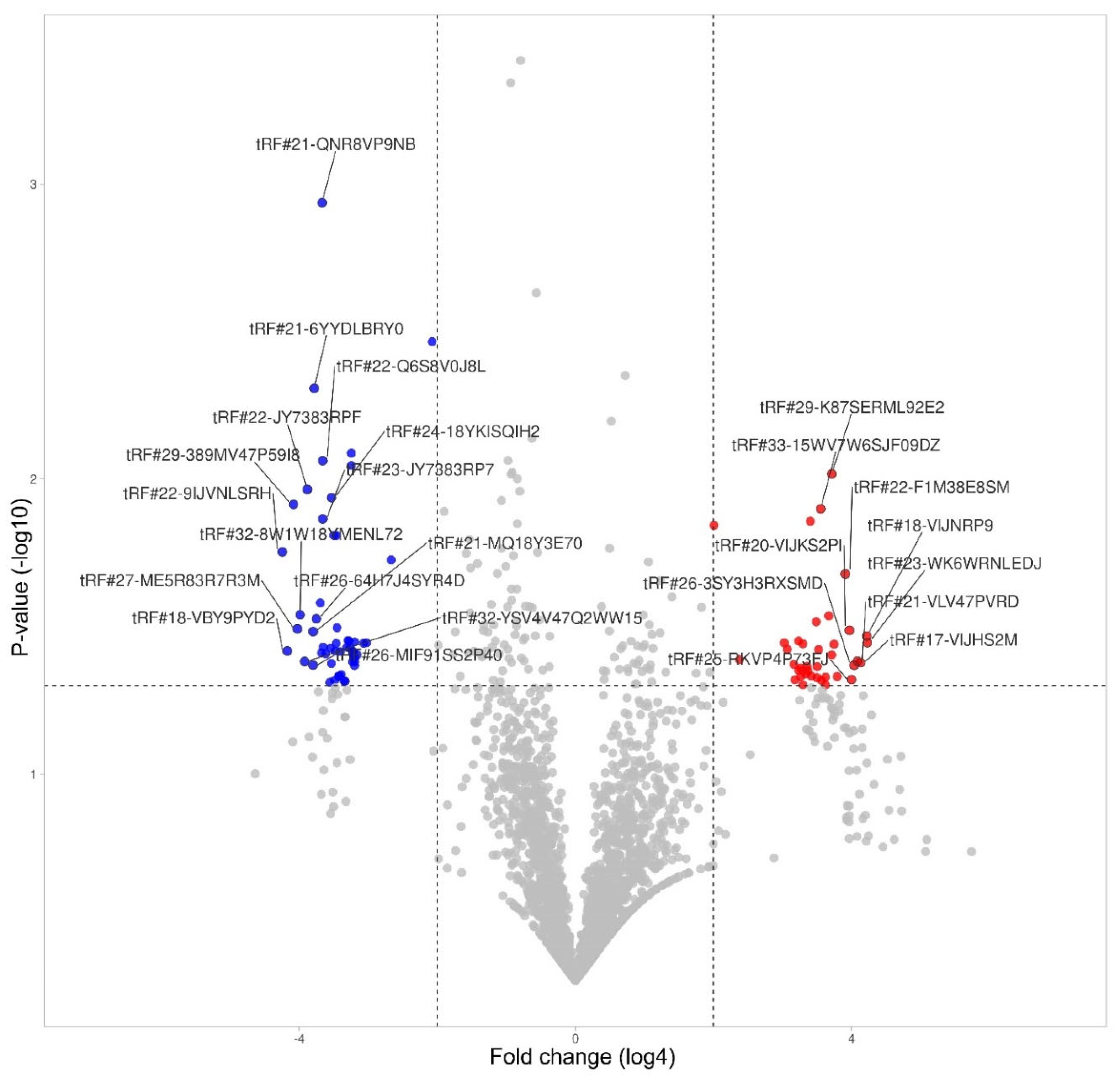

Table 1.
List of differentially expressed tRNA-derived fragments in erythrocytes during severe COVID-19.
Table 1.
List of differentially expressed tRNA-derived fragments in erythrocytes during severe COVID-19.
| Table 21. | Change | Fold change | P-value |
|---|---|---|---|
| tRF#21-LBLM8D4ND | Increased | 170 | 0,027 |
| tRF#23-RRN38B8800 | Increased | 146 | 0,043 |
| tRF#20-L3K8M0WL | Increased | 144 | 0,042 |
| tRF#29-F0QV7NLE2W1Y | Increased | 106 | 0,048 |
| tRF#23-1LQKHUXRDU | Increased | 104 | 0,038 |
| tRF#21-SRRN2EK8B | Increased | 94 | 0,048 |
| tRF#26-RKVP4P9L5FE | Increased | 88 | 0,039 |
| tRF#17-M3WE8SN | Increased | 88 | 0,040 |
| tRF#21-8PR9DM3WE | Increased | 86 | 0,048 |
| tRF#23-6KK87SIRD4 | Increased | 78 | 0,039 |
| tRF#33-73NK7F6Z2DNLDW | Increased | 74 | 0,035 |
| tRF#31-79MP9P9MH57SD | Increased | 74 | 0,037 |
| tRF#36-M26RRNLNK8KEP1B | Decreased | 536 | 0,035 |
| tRF#38-L85DMKYOYRLHR0D2 | Decreased | 320 | 0,033 |
| tRF#30-4WVLV470VR31 | Decreased | 224 | 0,047 |
| tRF#32-M2OSRN2NKSEKL | Decreased | 178 | 0,033 |
| tRF#33-QJ3KYUYRR6RBD2 | Decreased | 154 | 0,011 |
| tRF#32-M1M3WE8SSP6D2 | Decreased | 138 | 0,049 |
| tRF#24-32VIJMRPEU | Decreased | 136 | 0,043 |
| tRF#18-WB863IDV | Decreased | 124 | 0,048 |
| tRF#28-3JVIJMRPFQDY | Decreased | 120 | 0,039 |
| tRF#23-5J3KYUYRD9 | Decreased | 116 | 0,036 |
| tRF#19-DRJRWMI2 | Decreased | 108 | 0,047 |
| tRF#28-MXSL735L8RDV | Decreased | 108 | 0,047 |
| tRF#29-YP9LON4VN1EM | Decreased | 96 | 0,047 |
| tRF#20-593JJ426 | Decreased | 92 | 0,041 |
| tRF#22-8E8SP5X52 | Decreased | 92 | 0,041 |
| tRF#20-PW5SVP9N | Decreased | 92 | 0,044 |
| tRF#24-OP18YOY61Q | Decreased | 90 | 0,040 |
| tRF#32-HDK2RSI2WB0QJ | Decreased | 86 | 0,037 |
| tRF#21-1MQ8YUY60 | Decreased | 82 | 0,041 |
| tRF#37-WYL1M3WE8S68L52 | Decreased | 80 | 0,046 |
| tRF#28-3K76IR3DR2DV | Decreased | 16 | 0,047 |
Table 2.
List of differentially expressed tRNA-derived fragments in lymphocytes during severe COVID-19.
Table 2.
List of differentially expressed tRNA-derived fragments in lymphocytes during severe COVID-19.
| tRF id | Change | Fold change | P-value |
|---|---|---|---|
| tRF#23-N93VUIRI0Q | Increased | 557 | 0,024 |
| tRF#19-881X3SKQ | Increased | 280 | 0,039 |
| tRF#33-HM21YP4LR45D0Q | Increased | 265 | 0,009 |
| tRF#31-R29P4P9L5HJVE | Increased | 263 | 0,040 |
| tRF#34-RLXN4VZ87HFKJJ | Increased | 247 | 0,033 |
| tRF#34-HM21YP4LR45DKQ | Increased | 245 | 0,030 |
| tRF#29-MEF91SS2PMER | Increased | 233 | 0,046 |
| tRF#19-4N4OEZ07 | Increased | 233 | 0,045 |
| tRF#29-53N86SBHH90V | Increased | 207 | 0,005 |
| tRF#17-23KYY8M | Increased | 203 | 0,039 |
| tRF#19-VKS4I71Z | Increased | 190 | 0,043 |
| tRF#37-5QKDN6QQ1362NZO | Increased | 188 | 0,040 |
| tRF#19-7383RPEE | Increased | 183 | 0,005 |
| tRF#19-4KRQ59J4 | Increased | 170 | 0,030 |
| tRF#20-X29X3PMZ | Increased | 170 | 0,010 |
| tRF#38-5QKDN6QQ1362NZ0H | Increased | 165 | 0,046 |
| tRF#28-6XQ6S8V0J8DR | Increased | 150 | 0,037 |
| tRF#19-0HRF2LFQ | Increased | 150 | 0,001 |
| tRF#22-RKIP4OI3H | Increased | 143 | 0,004 |
| tRF#22-9X0UD394N | Increased | 140 | 0,037 |
| tRF#28-I89NJ4S2I7DM | Increased | 133 | 0,001 |
| tRF#21-YRXXE8QZE | Increased | 123 | 0,036 |
| tRF#23-34HWH3RX0B | Increased | 123 | 0,044 |
| tRF#22-46S9Y8RHP | Increased | 118 | 0,037 |
| tRF#19-P7M84I2Q | Increased | 115 | 0,037 |
| tRF#19-6XQ6S82X | Increased | 115 | 0,031 |
| tRF#20-1QKS3W2V | Increased | 113 | 0,049 |
| tRF#19-MIF91S2H | Increased | 113 | 0,039 |
| tRF#21-WRD81H93E | Increased | 110 | 0,043 |
| tRF#21-9L5H52NL0 | Increased | 110 | 0,035 |
| tRF#22-W60XY9BIQ | Increased | 108 | 0,031 |
| tRF#23-R9J89O9N9 | Increased | 105 | 0,035 |
| tRF#19-8NWE6WIZ | Increased | 105 | 0,035 |
| tRF#28-Z3R918VBY9DV | Increased | 105 | 0,032 |
| tRF#23-9M8O90Q4DZ | Increased | 103 | 0,049 |
| tRF#22-Z3FJ6KEWH | Increased | 103 | 0,049 |
| tRF#23-9N1QKS3WD1 | Increased | 103 | 0,033 |
| tRF#22-7EMQ18Y31 | Increased | 98 | 0,034 |
| tRF#21-7O3B1NR8E | Increased | 93 | 0,030 |
| tRF#22-282K63ZNQ | Increased | 90 | 0,033 |
| tRF#18-7383RP7 | Increased | 90 | 0,031 |
| tRF#23-RPM8309M0F | Increased | 88 | 0,030 |
| tRF#22-6LQ6S8V02 | Increased | 88 | 0,030 |
| tRF#35-S4I7LZM3Q01M3K | Increased | 85 | 0,029 |
| tRF#25-QSD2NSWWDZ | Increased | 83 | 0,031 |
| tRF#23-1E6SF8WOD9 | Increased | 83 | 0,029 |
| tRF#22-2EJ1OWZIQ | Increased | 80 | 0,029 |
| tRF#34-5QKDN6QQ1362HQ | Increased | 44 | 0,026 |
| tRF#35-5QKDN6QQ1362NZ | Increased | 25 | 0,030 |
Table 3.
List of differentially expressed tRNA-derived fragments in monocytes during severe COVID-19.
Table 3.
List of differentially expressed tRNA-derived fragments in monocytes during severe COVID-19.
| tRF id | Change | Fold change | P-value |
|---|---|---|---|
| tRF#23-WK6WRNLEDJ | Increased | 352 | 0,036 |
| tRF#18-VIJNRP9 | Increased | 348 | 0,034 |
| tRF#17-VIJHS2M | Increased | 306 | 0,042 |
| tRF#21-VLV47PVRD | Increased | 288 | 0,041 |
| tRF#26-3SY3H3RXSMD | Increased | 270 | 0,043 |
| tRF#25-RKVP4P73FJ | Increased | 256 | 0,048 |
| tRF#20-VIJKS2PI | Increased | 246 | 0,033 |
| tRF#22-F1M38E8SM | Increased | 226 | 0,021 |
| tRF#44-K8HJ83ML5F82NZD7HY | Increased | 192 | 0,047 |
| tRF#25-RKILQ673FJ | Increased | 180 | 0,036 |
| tRF#28-R1RXQ678Y2D8 | Increased | 172 | 0,039 |
| tRF#29-K87SERML92E2 | Increased | 172 | 0,010 |
| tRF#31-ZPQR16ZSIJ7KE | Increased | 162 | 0,029 |
| tRF#22-WEK6S1852 | Increased | 152 | 0,047 |
| tRF#23-K8HJ83MLDS | Increased | 152 | 0,050 |
| tRF#18-VIJKS2DU | Increased | 140 | 0,048 |
| tRF#33-15WV7W6SJF09DZ | Increased | 138 | 0,013 |
| tRF#25-P21MNKYUPR | Increased | 132 | 0,038 |
| tRF#38-Y2R79K3BEE4O3Q03 | Increased | 128 | 0,043 |
| tRF#24-PS5U8918JP | Increased | 128 | 0,047 |
| tRF#21-W60XY9BIE | Increased | 126 | 0,030 |
| tRF#30-6Q46D6PUMZQZ | Increased | 114 | 0,046 |
| tRF#22-9LON4VN11 | Increased | 112 | 0,014 |
| tRF#34-IEWS7YRR50SRIZ | Increased | 106 | 0,045 |
| tRF#18-HSQSD2D2 | Increased | 104 | 0,043 |
| tRF#32-ZPQR17NSRJ7KQ | Increased | 102 | 0,046 |
| tRF#26-R3HJ83RPFQE | Increased | 96 | 0,036 |
| tRF#19-K876IR19 | Increased | 96 | 0,050 |
| tRF#37-9EUK46S9Y8RH93Q | Increased | 92 | 0,044 |
| tRF#20-9VWVEH93 | Increased | 92 | 0,047 |
| tRF#28-WS3V2VR0PSDZ | Increased | 88 | 0,035 |
| tRF#20-SVKKN27F | Increased | 88 | 0,044 |
| tRF#32-FN5KYUSRYWRSJ | Increased | 82 | 0,048 |
| tRF#27-WN1Q18Y3HRK | Increased | 80 | 0,042 |
| tRF#20-3K7SIR3D | Increased | 70 | 0,038 |
| tRF#22-WEPSJR852 | Increased | 66 | 0,036 |
| tRF#21-MXSL73VLE | Increased | 27 | 0,041 |
| tRF#25-QNR8ZP9LON | Increased | 16 | 0,014 |
| tRF#22-9IJVNLSRH | Decreased | 360 | 0,018 |
| tRF#18-VBY9PYD2 | Decreased | 326 | 0,038 |
| tRF#29-389MV47P59I8 | Decreased | 288 | 0,012 |
| tRF#27-ME5R83R7R3M | Decreased | 266 | 0,032 |
| tRF#32-8W1W18YMENL72 | Decreased | 252 | 0,029 |
| tRF#32-YSV4V47Q2WW15 | Decreased | 230 | 0,042 |
| tRF#22-JY7383RPF | Decreased | 218 | 0,011 |
| tRF#26-MIF91SS2P40 | Decreased | 194 | 0,043 |
| tRF#21-MQ18Y3E70 | Decreased | 194 | 0,033 |
| tRF#21-6YYDLBRY0 | Decreased | 190 | 0,005 |
| tRF#26-64H7J4SYR4D | Decreased | 182 | 0,030 |
| tRF#21-MUWLV47PE | Decreased | 168 | 0,026 |
| tRF#21-R84QPYVMD | Decreased | 164 | 0,039 |
| tRF#21-QNR8VP9NB | Decreased | 162 | 0,001 |
| tRF#22-Q6S8V0J8L | Decreased | 160 | 0,009 |
| tRF#23-JY7383RP7 | Decreased | 160 | 0,014 |
| tRF#33-1MN0YU09FKRFD2 | Decreased | 158 | 0,037 |
| tRF#22-9P9NH57SJ | Decreased | 150 | 0,039 |
| tRF#23-RXSINHZ4DV | Decreased | 138 | 0,049 |
| tRF#24-941QKS3WF8 | Decreased | 136 | 0,037 |
| tRF#20-J87383RP | Decreased | 134 | 0,042 |
| tRF#24-18YKISQIH2 | Decreased | 134 | 0,012 |
| tRF#38-ML5F924ZDRJKW4DZ | Decreased | 128 | 0,016 |
| tRF#21-9P4P9L5HE | Decreased | 126 | 0,048 |
| tRF#24-Z3RQ18YJFH | Decreased | 124 | 0,038 |
| tRF#23-J87383RP7 | Decreased | 122 | 0,036 |
| tRF#20-RK9P4P9L | Decreased | 122 | 0,016 |
| tRF#22-MQ18Y3E7M | Decreased | 120 | 0,032 |
| tRF#28-RUPLQVNRDF0E | Decreased | 116 | 0,047 |
| tRF#22-73H3RXPLM | Decreased | 114 | 0,046 |
| tRF#24-94SX73V2KK | Decreased | 112 | 0,038 |
| tRF#21-MIF91SS20 | Decreased | 110 | 0,046 |
| tRF#24-04SXQ3V2KJ | Decreased | 104 | 0,048 |
| tRF#21-9LV470JPD | Decreased | 102 | 0,048 |
| tRF#20-1PSJPM17 | Decreased | 98 | 0,037 |
| tRF#34-JY7383RPD9W1JV | Decreased | 96 | 0,035 |
| tRF#24-YDLBRY73JL | Decreased | 94 | 0,035 |
| tRF#21-3P47M26YB | Decreased | 92 | 0,037 |
| tRF#22-6YR29P4PP | Decreased | 90 | 0,008 |
| tRF#21-WB8689SVD | Decreased | 90 | 0,009 |
| tRF#21-WLV47PU9E | Decreased | 88 | 0,039 |
| tRF#21-N1EH6KK80 | Decreased | 88 | 0,042 |
| tRF#18-SR99RHD2 | Decreased | 86 | 0,038 |
| tRF#33-5F924ZDRJKW4DZ | Decreased | 86 | 0,039 |
| tRF#22-5721V98B3 | Decreased | 84 | 0,036 |
| tRF#26-MY73H3RXPL0 | Decreased | 84 | 0,040 |
| tRF#22-J4S2I7L7M | Decreased | 84 | 0,042 |
| tRF#28-RKVP4P9L5F0Q | Decreased | 84 | 0,043 |
| tRF#26-94SL735FVI0 | Decreased | 80 | 0,039 |
| tRF#18-6M0Y1MY | Decreased | 70 | 0,036 |
| tRF#24-MY73H3RXII | Decreased | 66 | 0,036 |
| tRF#33-1N3KYUSR681SD2 | Decreased | 40 | 0,019 |
| tRF#25-R8VP9NFQFY | Decreased | 18 | 0,003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



