
Submitted:
06 September 2024
Posted:
07 September 2024
You are already at the latest version
Abstract
Cytokines play a substantial role in the pathophysiology of infectious diseases. Consequently, genetic polymorphisms in cytokine genes can have profound effect on the susceptibility to or protection from infections and also influence the clinical manifestations, disease severity as well as outcome. The objective of this brief narrative review is to provide an update on the recent research findings on the role of genetic polymorphisms of IL-18, a recently discovered cytokine on certain infectious diseases of public health relevance. The current review suggests there is a significant impact of IL-18 polymorphisms on different infectious diseases. The presence of G and T alleles at position -656 is suggested to have a probable protective effect against VL. The −137G/−607C (GC) and −137C/−607C (CC) haplotypes in the promoter region are associated with increased susceptibility to severe malarial anemia. SNPs rs544354 and rs574429 (GC) are linked to a higher risk of P. falciparum infection and increased parasitemia. The -137 C/C genotype and -137 G allele (with G/G genotype) are associated with faster progression to AIDS. In contrast, the IL-18 genotype does not seem to affect overall HIV susceptibility but influences disease progression. The G and GG genotypes at position -137 and allele A at position -607 are likely associated with protection against HBV infection. Conversely, the C allele at position -137 is linked to increased susceptibility to HBV infections. The GG genotype of IL-18 -137 G > C variant is associated with increased susceptibility to pulmonary tuberculosis (PTB). The AC genotype of IL-18 -607A>C has been linked to PTB susceptibility, particularly in patients with co-morbid diabetes mellitus. SNPs −137G/C, 113T/G, and +127C/T are associated with susceptibility to H. pylori infection, indicating that these genetic variants may influence the risk of H. pylori-related diseases. There are conflicting results in studies conducted in different populations (e.g., genetic polymorphism, studies conducted in India and China against HBV infection) highlighting the need for further research with larger sample sizes to resolve discrepancies. Due to the variability in findings, extensive studies in diverse populations are needed to better understand the impact of IL-18 polymorphisms.
Keywords:
cytokines
; IL-18
; gene polymorphism
; infectious diseases
; precision medicine
Introduction:
Interleukin -18 (IL-18):
Host and pathogen genetics significantly influence the pathogenesis and outcome of infectious diseases. After the discovery of CCR5 delta 32 deletion mutation rendering protection from Human Immunodeficiency Virus (HIV) infection, there is a great interest in exploring cytokine and chemokine polymorphisms and their influence on susceptibility and course of infections as well as protection from infections. IL-18 is a newly identified proinflammatory cytokine and is a key regulator of both innate and acquired immune responses [1]. It is a member of the IL-1 cytokine family, a group of 11 cytokines that stimulate the innate immune system [2,3]. In the presence of IL-12, IL-18 stimulates the production of interferon-gamma (IFN- γ) by acting on T helper 1 (Th 1) cells, macrophages, natural killer (NK) cells, natural killer T (NKT) cells, B cells, dendritic cells (DCs), and non-polarized T cells. IL-18 has a pleiotropic effect based on the cytokine environment, indicating its significant role in both health and disease [4]. IL-18 levels are typically high in patients with psoriasis (Gangemi S et al. 2003), systemic lupus erythematosus (SLE) [5], hypertension, chronic renal disease [6] multiple sclerosis (MS) [7] and certain infectious diseases like COVID-19 [8,9]. Pregnant women with recurrent miscarriage have significantly increased IL-18 gene expression [10,39]. IL-18 cytokine plays a role in the pathogenesis of several inflammatory and autoimmune diseases [11]. IL-18 has demonstrated value in providing protective immunity against mycobacteria through IFN- γ induction in cases of Mycobacterium avium infection [12]. A study reported that children with severe Mycoplasma pneumoniae infection expressed similar IL-18 and IFN- γ [13]. In another study, daily injections of IL-12 and IL-18 provided protective immunity against re-infection and prevented the spread of Leishmania major infection in mice infected with the intracellular protozoon [14]. Furthermore, in a mouse model of Herpes simplex virus (HSV), it was found that IL-18 functions as a protective factor against viral infection [15].
IL-18 was previously termed “IFN- γ -inducing factor” owing to its ability to promote IFN-γ release in CD3-stimulated Th1 cells. The human IL-18 protein has 193 amino acids, while the mouse IL-18 protein has 192 [16]. In humans, the IL-18 gene is located on chromosome 11 and in mice, it is found on chromosome 9. The gene has 7 exons with two different promoters on exons 1 and 2, which include an interferon consensus sequence binding protein and PU.1 binding sites (a hematopoietic-specific transcription factor) [1]. IL-18 is synthesized in the cytoplasm as an inactive precursor (pro-IL-18) of 193 amino acids and 24 kDa. Binding of Toll-like receptors (TLRs) to the Pathogen Associated Molecular Patterns (PAMPs) and activation of the NF-kB pathway, results in the transcription of IL-18 precursor. Secretion of IL-18 requires proteolytic processing to produce physiologically active IL-18 [17]. Caspase-1, an internal cysteine protease found in the NACHT-LRR and pyrin domain-containing protein 3 (NLRP3) inflammasome, is responsible for converting pro-IL-18 into mature IL-18, similar to pro-IL-1b [4] & [18]. Caspase 1 is activated by a variety of inflammasomes, including AIM2-like receptors, Nod-like receptors and TRIM family members with PYD or CARD domains. When Caspase 1 is activated, cells undergo pyroptosis, resulting in the formation of membrane holes and the release of mature IL-1b and IL-18. Figure 1 depicts the IL-18 signal transduction and biological effects of IL-18 gene.
The IL-18 receptor (IL-18R) is required for IL-18 signaling. T cells and NK cells produce IL-18R, which promotes IFN- γ production through STAT4 signaling [19]. The IL-18R consists of two subunits: the IL-18Ra chain (also known as IL-1R-related protein or IL-1R5) and the IL-18Rb chain (also known as IL-1R-associated protein-like or IL-R7). IL-18Ra and IL-18Rb chains belong to the IL-1R family and share a TIR domain with TLRs. When activated by IL-18, IL-18Ra forms a high-affinity binding heterodimer with IL-18Rb, enabling downstream signal transduction. IL-18 interacts with IL-12 and activate the innate immune system by inducing NK cells to respond to infections and cancers. Mice deficient in IL-18 exhibit greater susceptibility to infections and decreased NK cell activity, indicating the critical function IL-18 plays in establishing NK cell activity [20]. In the adaptive immune system, IL-18 endorses T cells’ activation and differentiation to upregulate the production of IFN- γ [18].
Different types of cells - hematopoietic as well as non-hematopoietic, can produce IL-18. It is generally produced by immature dendritic cells, monocytes, and macrophages during the acute immune response and it shares structural similarity with interleukin-1β (IL-1β). It is also found to be released by intestinal epithelial cells, endothelial cells, osteoblasts, keratinocytes, and mesenchymal cells [18].
Genetic Polymorphisms of IL-18
Genetic variation that effects a cytokine’s structure or expression to change, can have profound clinical consequences. Impact of cytokine gene polymorphisms on infectious diseases, including risk of infection, disease development, chronicity, response to vaccination and treatment, and vertical transmission, has been the subject recent research. There are several known distinct polymorphisms in IL-18. Most commonly, single nucleotide polymorphisms (SNPs) are located in the promoter region (656 G/T, 607 C/A, 137 G/C), whereas SNPs like 1113 T/G, 1127 C/T are located in the 50-untranslated region [21]. A single polymorphism at position 105 in the IL-18 gene’s coding area has also been discovered [22]. The polymorphisms at positions 607 and 137, in particular, are associated with significant differences in IL-18 expression, even though the functional importance of the polymorphisms mentioned above has not yet been thoroughly confirmed [21,23]. Functionally active regions of this promoter are impacted by C to A transversion at position -607 and G to C transversion at position -137. These components comprise binding sites for the cAMP response element binding protein (CREB) and transcriptional histone 4 transcription factor 1 (H4TF-1), respectively. Consequently, mutations at these locations may have an impact on IL-18 expression and affect the quantity of IL-18 produced. Therefore, the differences in IL-18 gene expression and production appear to be impacted up on by these two polymorphisms -607C/A and -137G/C and their haplotypes. Genotype G/G, at position -137 has been linked to higher IL-18 production and transcription activity [21,24].
IL-18 Polymorphisms and Infectious Diseases
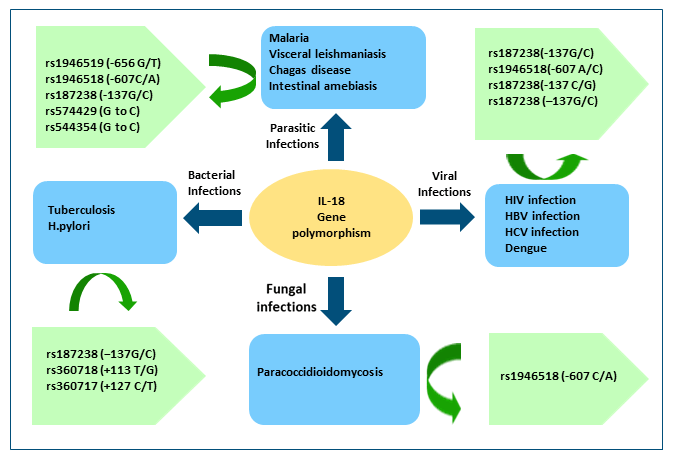
Multiple studies have reported an association between polymorphisms in the IL-18 gene and susceptibility to or protection from various infectious diseases – parasitic, bacterial, viral, and non-infectious diseases. We aim to highlight the various known genetic polymorphisms of the IL18 gene and their association with certain infectious diseases of public health importance. IL-18 gene and SNPs associated with infectious diseases are shown in Figure 2.
Parasitic Infections
Malaria
Malaria is caused by the unicellular protozoan, Plasmodium and is mostly spread by female Anopheles mosquitoes. Malaria continues to remain a serious global health challenge, especially in low- and middle-income nations, despite intense control efforts. An estimated 247 million new cases of malaria are reported every year, with an estimated 881,000 deaths from the disease, 91% of which happen in Africa and 85% of which occur in children under the age of five [25].
IL 18 has an important role in the pathophysiology of malarial infection. This cytokine is essential for controlling parasitemia and resolving malarial illness. Interleukin 12 (IL-12) and 18 (IL-18) together generate interferon-γ (IFN-γ), which mediates the first immunological response to P. falciparum infection, by inducing TNFα and increasing the release of reactive nitrogen and oxygen radicals. IL18 has a pro-inflammatory function in patients with uncomplicated P. falciparum infection [26]. A study reported that children with moderate malaria had a higher range of IL-18 and IL-12 than children with a severe form of the disease [27]. In murine model, it stimulates IFN- γ, which helps in defense against pathogenic organisms like Plasmodia [28]. It works along with IL-12 in the innate immune pathway against Plasmodium infection. In severe malaria cases, IL-18 levels remained high throughout the course of the disease which indicate the significant association between IL-18 levels in severe malaria patients and parasitemia [26]. When compared to healthy control subjects, SNPs rs5744292 and rs544354 were associated with parasitemia in patients infected with P. falciparaum. Increased parasite density in infected patients has been associated with SNP rs5744292. Significant age-dependent correlations were found between SNPs rs544354 and rs360714 and parasite density; the risk alleles were more prevalent in patients at a younger age (1–9 years old) [25].
Visceral Leishmaniasis (VL)
Leishmania donovani and Leishmania infantum (chagasi) are the intracellular protozoan parasites, which are responsible for the potentially lethal infectious disease Visceral Leishmaniasis. There is significant VL-related morbidity and mortality in Brazil and the Indian subcontinent [29]. Several researchers investigated the vital relation between IL-18 gene polymorphisms and the predisposition to VL.
A study was conducted in Bihar, India, to examine the IL-18 gene SNPs in the promoter region (rs187238 - 137 G/C), and (rs1946519 -656 G/T) and link with VL. Significantly, the G allele at rs1946519 (− 656 G/T) and codon region (rs549908 + 105 A/C) was linked to protection from VL [30]. In another study by Moravej A et al., it was observed that in Iranian VL patients, the T allele frequency was decreased, while it was higher in controls (− 656). Resistance to VL was associated with a T allele at – 656 [31]. Ahmadpour E et al., on the other hand, showed that in the East Azerbaijan, Iran people, the IL-18 gene promoter polymorphisms at positions -137 and -607 are not linked to VL [32].
Chagas Disease
Chagas disease is a chronic, parasitic infection caused by the protozoan Trypanosoma cruzi, and the disease affects around 8 million people in Latin America, with 30-40% having or developing cardiomyopathy. Strauss, M. et al. examined the association of three IL18 genetic variations rs2043055, rs1946518, and rs360719, and the development of chronic Chagas cardiomyopathy (CCC) and the susceptibility to Trypanosoma cruzi infection in various Latin American populations [33]. An SNP (rs2043055) was found to associated with an increased risk of Chagas cardiomyopathy (CC), according to a study conducted in 2015 by Nogueira, L.G. et al. [34]. This study also found a significant variation in genotype frequencies among patients with moderate and severe CCC. In contrast, the findings of Gomes dos Santos, A. et al., IL18 s1946518 AA genotypes reduced risk of developing cardiomyopathy and decrease the severity of cardiomyopathy in Chagas disease [35].
Intestinal Amebiasis
The protozoan Entamoeba histolytica causes intestinal amebiasis and extra-intestinal symptoms. Ninety percent of E. histolytica infections are asymptomatic, but over 50 million develop symptoms, and each infection results in roughly 100,000 fatalities annually. Al-Sultany, A.K. et al. 2023 investigated the genetic polymorphisms of IL-18 and its role in susceptibility to infection by E. histolytica [36]. Findings indicated that mutations in SNPs such as SNP 1 (rs1866694757), SNP 4 (rs 1946518), SNP 6 (rs 1946519), and SNP 7 (rs 1215648807) may be associated with an increased risk of intestinal infections. Whereas, other SNPs, such as SNP 2 (rs 940255648), SNP 3 (rs 1037707423), SNP 5 (rs 1213044637), SNP 8 (rs 1866697972), SNP 9 (rs 1866698066), and SNP 10 (rs 186668286), showed the protective activity against disease progression. However, this study is limited by its small sample size and to confirm these results, further research is needed, with large number of participants from a range of populations.
Viral Infections
HIV Infection:
Globally, HIV has infected over 75 million people and is a leading cause of morbidity and mortality. Untreated HIV disease increases the risk of opportunistic infections and malignancies by causing gradual depletion of CD4+ T cells and a variety of immunological abnormalities. An increasing body of evidence indicates the importance of host gene variations in both the progression of AIDS and an individual’s susceptibility to HIV-1 infection [37]. IL-18-137 the G/G genotype and G allele appears to play a role in the pathogenesis of HIV-1 infection in North Indians. HIV-1 patients carrying the G/G genotype and -137 G allele might have a faster progression of their illness to acquired immunodeficiency syndrome (AIDS) than individuals with the C/C genotype. The G/G genotype and G allele at position -137 of the IL-18 gene promoter polymorphism may be involved in the progress of HIV-1 infection [38].
HBV Infection:
The hepatitis B virus (HBV) affects more than 350 million populations worldwide. Hepatitis B is a prominent cause of chronic hepatitis, cirrhosis, and hepatocellular carcinoma and is responsible for 1 million deaths per annum. Certain IL-18 polymorphisms have been linked to an increased risk of developing chronic hepatitis B, changes in the severity and course of the illness. In the Chinese population, Jiang et al. 2018 reported that the -137 G allele and the GG genotype are associated with the development of chronic hepatitis B (CHB) infection and decreased blood levels of IL-18, respectively [40]. Zhang et al.‘s earlier study revealed no association between disease susceptibility and polymorphism at position -607. Zhang et al. have also shown the protective functions of the -137 C allele against HBV infection [41].
HCV Infection:
Hepatotropic RNA virus known as the hepatitis C virus (HCV) gradually damages the liver, potentially leading to hepatocellular carcinoma and liver cirrhosis. China has the largest number of HCV patients (29.8 million), whereas Egypt has the highest prevalence of HCV at >10% of the general population [42,43].
Similar to Hep B, IL18 has a significant role in chronic hepatitis C infection as well. More than 50% of patients progress from having an acute infection to developing a persistent infection, of those 10–20% are tending toward hepatocellular carcinoma (HCC) and cirrhosis [44,45]. IL-18 genetic polymorphisms and plasma levels of IL-18 have been stated to impact HCV infection. Farid S et al. stated that, the polymorphism at the -607 location with the AA allele is greater in healthy individuals than in Chronic hepatitis C virus (CHCV) patients, the polymorphism at the -607 location with the AA allele is a promising protective marker [46]. In contrast, Said EM et al. evaluated the frequencies of two functional IL-18 gene variations (-607 C/A and -137 G/C) and quantitatively assessed the IL-18 plasma levels to ascertain their influence on the severity of CHC and revealed that individuals with CHC have higher levels of IL-18 plasma, which positively correlates with the severity of liver disease. Milder liver disease is linked to the A/A allele at position -607 of the IL-18 gene promoter [47]. In another investigation, the haplotypes of the IL18 promoter polymorphisms 607C/A and 137G/C were studied about the outcomes of HCV persistence or clearance in European Americans (EA) and African Americans (AA). A strong association was found in haplotype carrying -607A and -137C with viral clearance [48] Yue M et al. have shown that three IL-18 variants −656G > T, +105A > C and −137G > C, are also associated with protection against vulnerability to HCV infection in the high-risk Chinese Han population. Additionally, he discovered that the GCC haplotype was significantly linked to a lower risk of being susceptible to HCV infection. In contrast, the GGC haplotype was strongly related to protection against HCV infection that persisted [49].
Hirankarn et al. have reported an association between the -607 AA genotype and the development of CHB [50]. The CC and GC genotypes of the polymorphism in the -137 position were linked to a higher risk of HCC in CHB patients in the Chinese population [51]. According to Bao et al., patients with HBV infection who had the GC genotype and the C allele of the polymorphism at location -137 had a considerably lower chance of developing HCC [52]. A subsequent investigation demonstrated a correlation between the GG genotype and the G allele of the -137 polymorphism with the severity of necro inflammatory activity and liver fibrosis [53]. Li et al., on the other hand, showed that the AA genotype and the A allele at the -607-promoter region may be linked to CHB resistance [54]. Kim et al. found that in CHB patients, the IL-18 gene’s +13925 C +8925 G and -148 C alleles were linked to the development of HCC [55]. Further, a strong correlation between the alleles for -148 C, +8925 G, and +13925 C and HBV clearance was noted [56]. Zhu et al., reported that in CHB patients, the polymorphisms in the -607 and 137 promoter region of IL-18 were not linked to the development of HCC ([57]. Dai ZJ et al., (2017) study shows that IL-18 polymorphisms rs187238 GG genotype increases the risk of hepatocellular carcinoma in healthy individuals while it is associated with cirrhosis in chronic hepatitis B carriers in the Northwest Chinese population [58].
Dengue:
Dengue fever is a mosquito-borne disease, widespread in tropical and subtropical areas and caused by the dengue virus belonging to the Flavivirus genus of the Flaviviridae family. A wide spectrum of clinical manifestations ranging from asymptomatic to mild febrile illness, dengue fever (DF) and severe dengue hemorrhagic fever (DHF) are unknown.
Pohan Herdiman T et al. investigated the association between hospitalized DF and DHF patients IL-18 levels and the severity of their sickness. Research revealed that IL-18 levels were considerably greater in DHF than DF. Furthermore, a strong association was found between IL-18 levels with low platelet values and hematocrit and This work supports the potential contribution of IL-18 in the etiology of adult DHF [25]. Another study showed that high levels of IL-18 at the later stages of the disease and during severe illness indicate that this cytokine may influence the pathophysiology of DHF by causing a change in the Th1-to Th2-type response [59]. However, we could not find any studies specifically investigating the role of IL-18 polymorphisms and dengue fever.
Bacterial Infections
Several studies investigated IL-18 gene polymorphism and its crucial role in various bacterial infections like Helicobacter pylori (H. pylori) infections, tuberculosis, etc.
Tuberculosis (TB):
Tuberculosis (TB) is a significant global health issue caused by Mycobacterium tuberculosis (MTB). It usually affects the lungs, but it can also damage other organs such as the kidneys, spine, or brain. Every year, it causes morbidity in millions of people and is the second biggest cause of mortality from an infectious disease, next to the human immunodeficiency virus (HIV)/AIDS. According to a global TB survey, Asia and Africa account for the majority of the projected (8.7 million) cases worldwide [60]. An estimated 30% of the general population is thought to be infected with MTB and 10% of those infected will develop eventually active TB [61].
A meta-analysis study conducted by He C et al. revealed that the polymorphisms IL-18 rs1946518 IL-18 rs187238 and rs1800795 may increase a person’s risk of tuberculosis, particularly in Asians. [62]. An additional investigation into the relationship between IL-18 and TB susceptibility analysis revealed that patients in the TB group had greater IL-18 levels than those in the control group [63]. In the Chinese Han population, the connection between SNPs of the IL-18 gene and susceptibility to pulmonary tuberculosis was investigated by [64].
H. pylori Infections:
Helicobacter pylori (H. pylori) is a bacteria that infects the stomach and duodenum. H. pylori infection is a primary causative factor for gastritis and peptic ulcers (Pacifico L et al. 2010). It has been estimated that 50% of the global population is infected with this bacteria. Rezaeifar A. et al. showed how IL-18 promoter polymorphisms affected serum levels of IL-18, in patients with H. pylori-infected duodenal ulcers (DU) and also elucidated potential correlation between the H. pylori virulence factors CagA and VacA antibodies with IL-18 serum level. It revealed that a higher risk of developing DU was linked to the IL-18 -607C variation. Moreover, CagA-positive individuals with high levels of IL-18 are more susceptible to DU [65]. Myung DS et al. 2015 explored that, IL-18 gene polymorphisms at positions −137G/C (rs187238), +113T/G (rs360718), and +127C/T (rs360717) demonstrated protective activity against H. pylori infection by decreasing its susceptibility in the Korean population [66].
Fungal Infections
Paracoccidioidomycosis (PCM):
Paracoccidioidomycosis (PCM) is a widespread endemic illness in Latin America, paracoccidioides species are the causative agent of the disease. High morbidity and sequelae have been associated with this mycosis. Sato PK et al. 2020 found that polymorphism in the IL18 gene’s promoter region and its association with the severity of PCM by demonstrating, the risk related to the IL18-607 A-allele in the acute form (AF) and multifocal chronic (MC) groups, along with the protective effect of the C-allele in unifocal chronic (UC), and its possible association with the greater levels of IL-18 at various stages of the disease’s progression [67]
The results of the various studies on IL-18 genetic polymorphisms and their association with different infectious diseases of public health significance have been summarized in Table 1.
Summary
The current review suggests there is a significant impact of IL-18 polymorphisms on different infectious diseases. The presence of G and T alleles at position -656 is suggested to have a probable protective effect against VL. The −137G/−607C (GC) and −137C/−607C (CC) haplotypes in the promoter region are associated with increased susceptibility to severe malarial anemia. SNPs rs544354 and rs574429 (GC) are linked to a higher risk of P. falciparum infection and increased parasitemia. The -137 C/C genotype and -137 G allele (with G/G genotype) are associated with faster progression to AIDS. In contrast, the IL-18 genotype does not seem to affect overall HIV susceptibility but influences disease progression. The G and GG genotypes at position -137 and allele A at position -607 are likely associated with protection against HBV infection. Conversely, the C allele at position -137 is linked to increased susceptibility to HBV infections. The GG genotype of IL-18 -137 G > C variant is associated with increased susceptibility to pulmonary tuberculosis (PTB). The AC genotype of IL-18 -607A>C has been linked to PTB susceptibility, particularly in patients with co-morbid diabetes mellitus. SNPs −137G/C, 113T/G, and +127C/T are associated with susceptibility to H. pylori infection, indicating that these genetic variants may influence the risk of H. pylori-related diseases. There are conflicting results in studies conducted in different populations (e.g., genetic polymorphism, studies conducted in India and China against HBV infection) highlighting the need for further research with larger sample sizes to resolve discrepancies. Due to the variability in findings, extensive studies in diverse populations are needed to better understand the impact of IL-18 polymorphisms.
Future Direction:
Population studies are crucial to understanding how IL-18 genetic polymorphisms vary across different ethnic groups and how this diversity contributes to differential susceptibility to infections worldwide. In regions with high infectious disease burdens, understanding genetic predispositions can inform public health strategies and interventions. It is also noted that studies in different populations often report contradicting findings. Hence, future studies should define the target disease and controls very well, should be adequately powered and a multidisciplinary approach, integrating genetics, immunology, bioinformatics, and clinical studies to elucidate underlying mechanisms and translate findings into improved healthcare practices. Epigenetic alterations (e.g., DNA methylation, histone modifications) may impact IL-18 gene expression and these modifications interact with genetic polymorphisms in the context of infections. Longitudinal studies to track how IL-18 polymorphisms influence infection outcomes over time can provide insights into long-term immune modulation and potential implications for chronic infections. Genetic studies assume much significance in the current context of personalized medicine or precision medicine as targeted immunotherapy that enhances IL-18 signaling to boost immune responses or its inhibition to contain hyperinflammatory responses may come up in the future.
Acknowledgments
Dr Vijesh Sreedhar K is supported by an extramural grant of ICMR (Project ID EM 2229; Grant number: IRIS ID No- 2020-1428; No.6/9(257)2020/ECD-II).
References
- K. Nakanishi, T. Yoshimoto, H. Tsutsui, H. Okamura, Interleukin-18 regulates both Th1 and Th2 responses, Annu Rev Immunol. 19 (2001) 423–474. https://doi: 10.1146/annurev.immunol.19.1.423. [CrossRef]
- J.E. Sims, D.E. Smith, The IL-1 family: regulators of immunity, Nat. Rev. Immunol. 10 (2010) 89–102 https://doi.org/10.1038/nri2691.
- J.E. Sims, M.j. Nicklin, J.F. Bazan, J.L. Barton, S.J. Busfield, J.E. Ford, A new nomenclature for IL-1-family genes, Trends Immunol. 22 (2001) 536–537. https://doi:10.1016/S1471-4906(01)02040-3. [CrossRef]
- K. Yasuda, K. Nakanishi, H. Tsutsui, Interleukin-18 in health and disease, Int J Mol Sci. 20 (2019) 649–701. https://doi.org/10.1038/nri2691.
- M.C. Park, Y.B. Park, S.K. Lee, Elevated interleukin-18 levels correlated with disease activity in systemic lupus erythematosus, Clin Rheumatol 23 (2004) 225-229. https://doi: 10.1007/s10067-004-0867-x. [CrossRef]
- J. M. Thomas, B.M. Huuskes, C.G. Sobey, G.R. Drummond, A. Vinh, The IL-18/ IL-18R1 signalling axis: Diagnostic and therapeutic potential in hypertension and chronic kidney disease, Pharmacol Ther. 239 (2022) 108191. https://doi: 10.1016/j.pharmthera.2022.108191. [CrossRef]
- F. Nicoletti, R. Di Marco, K. Mangano, F. Patti, E. Reggio, A. Nicoletti, Increased serum levels of interleukin-18 in patients with multiple sclerosis, J. Neurol. 57 (2001) 342–344. https://doi.org/10.1212/WNL.57.2.342. [CrossRef]
- T. Nakahashi, M.K. Ellingson, P. Wong, B. Israelow, C. Lucas, J. Klein, Sex differences in immune responses that underlie COVID-19 disease outcomes, Nature. 588 (2020) 315–320. https://doi: 10.1038/s41586-020-2700-3. [CrossRef]
- T.S. Rodrigues, K.S.G. de Sa, A.Y. Ishimoto, A. Becerra, S. Oliveira, L. Almeida, Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J Exp Med. 218 (2021) e20201707. https://doi: 10.1084/jem.20201707. [CrossRef]
- S. Lob, B. Ochmann, Z. Ma, T. Vilsmaier, C. Kuhn, E. Schmoeckel, The role of interleukin-18 in recurrent early pregnancy loss, J Reprod Immunol. 148 (2021) 103432. https://doi: 10.1016/j.jri.2021.103432. [CrossRef]
- S.K. Sedimbi, T. Hagglof, M.C. Karlsson, IL-18 in inflammatory and autoimmune disease, Cell Mol Life Sci 70 (2013) 4795–4808. https://doi: 10.1007/ s00018-013-1425-y. [CrossRef]
- K. Kobayashi, N. Nakata, M. Kai, T. Kasama, Y. Hanyuda, Y. Hatano, Decreased Expression of Cytokines That Induce Type 1 Helper T Cell/Interferon-γ Responses in Genetically Susceptible Mice Infected with Mycobacterium avium, J. Clin. immunol. 85 (1997) 112-116. https://doi.org/10.1006/clin.1997.4421. [CrossRef]
- R. Zhu, S. Mao, W. Shi, L. Wu, J. Zhang, A prediction study of IL-18 and IFN-γ in glucocorticoid treatment response in infants and young children with severe Mycoplasma pneumoniae pneumonia, Transl Pediatr. 11(2022) 738-747. https://doi.org/10.21037/tp-22-139. [CrossRef]
- K. Ohkusu, T. Yoshimoto, K. Takeda, T. Ogura, S. Kashiwamura, Y. Iwakura, S. Akira, H. Okamura, K. Nakanishi, Potentiality of Interleukin-18 as a Useful Reagent for Treatment and Prevention of Leishmania major Infection, Infect Immun. 68 (2000) 2449-2456. https://doi.org/10.1128/iai.68.5.2449-2456.2000. [CrossRef]
- N. Fujioka, R. Akazawa, K. Ohashi, M. Fujii, M. Ikeda, M. Kurimot, Interleukin-18 Protects Mice against Acute Herpes Simplex Virus Type 1 Infection. J Virol 73 (1999) 2401-2409.https://doi.org/10.1128/jvi.73.3.2401-2409.1999. [CrossRef]
- S. Ushio, M. Namba, T. Okura, K. Hattori, Y. Nukada, K. Akita K, Cloning of the cDNA for human IFN-gamma-inducing factor, expression in escherichia coli, and studies on the biologic activities of the protein, J Immunol. 156 (1996) 4274–4279. https://doi.org/10.4049/jimmunol.156.11.4274. [CrossRef]
- M. Tomura, S. Maruo, J. Mu, X.Y. Zhou, H.J. Ahn, T. Hamaoka, Differential capacities of CD4+, CD8+, and CD4-CD8- T cell subsets to express IL-18 receptor and produce IFN-gamma in response to IL-18. J Immunol 160(1998): 3759–65. https://doi.org/10.4049/jimmunol.160.8.3759. [CrossRef]
- G. Kaplanski, Interleukin-18: Biological properties and role in disease pathogenesis. Immunol Rev 281(2018):13853 https://doi.org/10.1111/imr.12616. [CrossRef]
- [19] T. Yoshimoto, K. Takeda, T. Tanaka, K. Ohkusu, S. Kashiwamura, H. Okamura, IL-12 up-regulates IL-18 receptor expression on T cells, Th1 cells, and b cells: synergism with IL-18 for IFN-gamma production. J Immunol 161(1998):3400–7. https://doi.org/10.4049/jimmunol.161.7.3400. [CrossRef]
- K. Hoshino, H. Tsutsui, T. Kawai, K. Takeda, K. Nakanishi, Y. Takeda, Cutting edge: generation of IL-18 receptor-deficient mice: evidence for IL-1 receptor-related protein as an essential IL-18 binding receptor. J Immunol (1999) 162(9):5041–4. https://doi.org/10.4049/jimmunol.162.9.5041. [CrossRef]
- V. Giedraitis, B. He, W.X. Huang, J. Hillert, Cloning and mutation analysis of the human IL-18 promoter: a possible role of polymorphisms in expression regulation, J. Neuroimmunol. 112 (2001) 146-152, https://doi.org/10.1016/S0165-5728(00)00407-0. [CrossRef]
- S. Higa, T. Hiran, M. Mayumi, M. Hiraoka, Y. Ohshima, M. Nambu, E. Yamaguchi, N. Hizawa, N. Kondo, E. Matsui, Y. Katada, Association between interleukin-18 gene polymorphism 105A/C and asthma. Clinical & Experimental Allergy, 33 (2003) 1097-1102, https://doi.org/10.1046/j.1365-2222.2003.01739.x. [CrossRef]
- S.R. Thompson, and S.E. Humphries, Interleukin-18 genetics and inflammatory disease susceptibility, Genes & Immunity. 8 (2007) 91-99, https://doi.org/10.1038/sj.gene.6364366. [CrossRef]
- J. Arimitsu, T. Hirano, S. Higa, M. Kawai, T. Naka, A. Ogata, Y. Shima, M. Fujimoto, T. Yamadori, K. Hagiwara, T. Ohgawara, IL-18 gene polymorphisms affect IL-18 production capability by monocytes, Biochem. Biophys. Res. Commun. 342 (2006) 1413-1416, https://doi.org/10.1016/j.bbrc.2006.02.096. [CrossRef]
- A.U. AlRuwaisan, M.R. Al-Anazi, M.I. Shafeai, F.H. Rudiny, A.M. Motaen, S.M. Bin Dajem, H. Alothaid, K. Morsy, S. Alkahtani, A.A. Al-Qahtani, Associations of single nucleotide polymorphisms in Il-18 gene with plasmodium falciparum-associated malaria, J. Inflamm. Res. 14 (2021) 3587-3619, https://doi.org/10.2147/JIR.S314638. [CrossRef]
- D. Torre, M. Giola, F. Speranza, A. Matteelli, C. Basilico, and G. Biondi, Serum levels of interleukin-18 in patients with uncomplicated Plasmodium falciparum malaria, Eur. Cytokine Netw. 12 (2001) 361-364.
- L. Malaguarnera, S. Pignatelli, M. Musumeci, J. Simporè, and S. Musumeci, Plasma levels of interleukin-18 and interleukin-12 in Plasmodium falciparum malaria. Parasite immunology, 24(2002) 489-492, https://doi.org/10.1046/j.1365-3024.2002.00485.x. [CrossRef]
- R.P. Singh, S.I. Kashiwamura, P. Rao, H. Okamura, A. Mukherjee, V.S. Chauhan. The role of IL-18 in blood-stage immunity against murine malaria Plasmodium yoelii 265 and Plasmodium berghei ANKA. The Journal of Immunology. 168 (2002) 4674-81.
- A.S. Mustafa, E.A. Elbishbishi, R. Agarwal, and U.C. Chaturvedi, Elevated levels of interleukin-13 and IL-18 in patients with dengue hemorrhagic fever, FEMS Microbiol. Immunol. 30 (2001) 229-233, https://doi.org/10.1111/j.1574-695X.2001.tb01575.x. [CrossRef]
- D. Kumar, P. Tiwary, J. Chakravarty, S. Sundar, Association of interleukin-18 gene polymorphism with susceptibility to visceral leishmaniasis in endemic area of Bihar, an Indian population, Sci. World J. 1 (2014) 852104, http://dx.doi.org/10.1155/2014/852104. [CrossRef]
- Moravej, M. Rasouli, S. Asaei, M. Kalani, and Y. Mansoori, Association of interleukin-18 gene variants with susceptibility to visceral leishmaniasis in Iranian population, Mol. Biol. Rep. 40 (2013) 4009-4014, https://doi.org/10.1007/s11033-012-2479-x. [CrossRef]
- E. Ahmadpour, A. Bazmani, M.H. Kohansal, A. Kazemi, Z. Babaloo, IL-18 gene polymorphism in patients with visceral leishmaniasis in East Azarbaijan, Iran, J. Parasit. Dis. 40 (2016) 981–985, https://doi.org/10.1007/s12639-014-0619-z. [CrossRef]
- M. Strauss, M. Acosta-Herrera, A. Alcaraz, D. Casares-Marfil, P. Bosch-Nicolau, M.S. Lo Presti, I. Molina, C.I. González, C.I., Chagas Genetics CYTED Network and J. Martín, Association of IL18 genetic polymorphisms with Chagas disease in Latin American populations, PLOS Negl. Trop. Dis. 13 (2019) e0007859, https://doi.org/10.1371/journal.pntd.0007859. [CrossRef]
- L.G. Nogueira, A.F. Frade, B.M. Ianni, L. Laugier, C.W. Pissetti, S. Cabantous, M. Baron, G. Lima Peixoto, A. de Melo Borges, E. Donadi and J.A. Marin-Neto, Functional IL18 polymorphism and susceptibility to Chronic Chagas Disease, Cytokine. 73 (2015) 79-83, https://doi.org/10.1016/j.cyto.2015.01.037. [CrossRef]
- Gomes dos Santos, E.H. Watanabe, D.T. Ferreira, J. Oliveira, É.S. Nakanishi, C.S. Oliveira, E. Bocchi, C.T.G. Novaes, F. Cruz, N.B. Carvalho, P.K. Sato, A specific IL6 polymorphic genotype modulates the risk of Trypanosoma cruzi parasitemia while IL18, IL17A, and IL1B variant profiles and HIV infection protect against cardiomyopathy in Chagas disease, Front. immunol. 11 (2020) 521409, https://doi.org/10.3389/fimmu.2020.521409. [CrossRef]
- A.K. Al-Sultany, K.A.H. Al-Morshidy, Single nucleotide polymorphism of IL-18 gene and resistin gene in children infection with Entamoeba histolytica, Med. j. Babylon. 20 (2023) 697-704, https://doi.org/10.4103/MJBL.MJBL_70_23. [CrossRef]
- Deeks SG, Overbaugh J, Phillips A, Buchbinder S. HIV infection. Nature reviews Disease primers. 2015 Oct 1;1(1):1-22. https://doi.org/10.1038/nrdp.2015.35. [CrossRef]
- R.C. Sobti, V.L. Sharma, A.M. Abitew, N. Berhane, S.A. Mahdi, M. Askari, V.S. Kuttiat, V.S. and A. Wanchu, The -137G/C polymorphism of interleukin 18 promoter and risk of HIV-1 infection and its progression to AIDS, Acta virologica. 55 (2011) 353, https://doi.org/10.4149/av_2011_04_353. [CrossRef]
- G.M. Al-Khateeb, M.S. Sater, R.R. Finan, F.E. Mustafa, A.S. Al-Busaidi, M.A. Al-Sulaiti, W.Y. Almawi, Analysis of interleukin-18 promoter polymorphisms and changes in interleukin-18 serum levels underscores the involvement of interleukin-18 in recurrent spontaneous miscarriage, Fertil. Steril. 96 (2011) 921-926, https://doi.org/10.1016/j.fertnstert.2011.06.079. [CrossRef]
- H. Jiang, F. Cao, H. Cao H, Q. Rao, Y. Yang, Associations of human leukocyte antigen and interleukin-18 gene polymorphisms with viral load in patients with hepatitis B infection, Medicine. 97 (2018) 11249, http://dx.doi.org/10.1097/MD.0000000000011249. [CrossRef]
- P.A. Zhang, J.M. Wu, Y. Li and X.S. Yang, X.S, Relationship of interleukin-18 gene promoter polymorphisms with chronic hepatitis B in Chinese Han population. Chin. J. Genet. 22 (2005) 528-532. https://doi.org/10.1128/jvi.73.3.2401-2409.1999. [CrossRef]
- Hajarizadeh, B., Grebely, J. & Dore, G. Epidemiology and natural history of HCV infection. Nat Rev Gastroenterol Hepatol 10, 553–562 (2013). https://doi.org/10.1038/nrgastro.2013.107. [CrossRef]
- Manns, M., Buti, M., Gane, E. et al. Hepatitis C virus infection. Nat Rev Dis Primers 3, 17006 (2017). https://doi.org/10.1038/nrdp.2017.6. [CrossRef]
- K. Kiyosawa, E. Tanaka, Hepatitis C Virus in the Etiology of Hepatocellular Carcinoma, Perspect. Med. Virol. 6 (2002) 31–42, https://doi.org/10.1016/S0168-7069(02)06064-0. [CrossRef]
- F.V. Chisari, Unscrambling Hepatitis C Virus–Host Interactions, Nature. 436 (2005) 930–932, https://doi.org/10.1038/nature04076. [CrossRef]
- S. Farid, L. Rashid, S. Swelam, The Role of Interleukin-18 Promoter Polymorphisms (-607 C/A and 137 G/C) in Determining HCV Clearance or Persistence, Egypt. J. Hosp. Med. 50 (2018) 141-149, https://doi.org/10.21608/ejhm.2018.16083. [CrossRef]
- E.M. Said, M.S. Soliman, H.I. Shousha, M.S. Rashed, A.A. Elazm, R.Z. Aamer, M.H. Kamel and F.M. Abdelsalam, Interleukin-18 and its gene single nucleotide polymorphisms (SNPs) influence chronic hepatitis C progression, J. Infect. Dev. Ctries. 12 (2018) 257-264. https://doi.org/10.3855/jidc.9813. [CrossRef]
- P. An, C.L. Thio, G.D. Kirk, S. Donfield, J.J. Goedert, C.A. Winkler, Regulatory polymorphisms in the interleukin-18 promoter are associated with hepatitis C virus clearance, J. Infect. Dis. 198 (2008) 1159-1165, https://doi.org/10.1086/592047. [CrossRef]
- M. Yue, J.J. Wang, L. Feng, Y. Zhang, Y. Liu, J. Wang, X.Z. Deng, K. Xu and J. Zhang, J, Association of interleukin-18 gene polymorphisms with the outcomes of hepatitis C virus infection in high-risk Chinese Han population, Immunol. Lett. 154 (2013) 54-60, https://doi.org/10.1016/j.imlet.2013.08.007. [CrossRef]
- N. Hirankarn, C. Manonom, P. Tangkijvanich, Y. Poovorawan, Association of interleukin-18 gene polymorphism (− 607A/A genotype) with susceptibility to chronic hepatitis B virus infection, Tissue Antigens. 70 (2007) 160-163. https://doi.org/10.1111/j.1399-0039.2007.00865.x. [CrossRef]
- H.K. Lau, M.J. Hsieh, S.F. Yang, H.L. Wang, W.H. Kuo, H.L. Lee, and C.B. Yeh, Association between interleukin-18 polymorphisms and hepatocellular carcinoma occurrence and clinical progression, Int. J. Med. Sci. Public Health. 13 (2016) 556, https://doi.org/10.7150/ijms.15853. [CrossRef]
- J. Bao, Y. Lu, Y. Deng, C. Rong, Y. Liu, X. Huang, L. Song, S. Li, X. Qin, Association between IL-18 polymorphisms, serum levels, and HBV-related hepatocellular carcinoma in a Chinese population: a retrospective case–control study, Cancer. Cell. Int. 15 (2015) 1-8, https://doi.org/10.1186/s12935-015-0223-z. [CrossRef]
- S.D.C. Ferreira, S.G.F. Chachá, F.F. Souza, A.C. Teixeira, R. de Carvalho Santana, N.H.S. Deghaide, S. Rodrigues, Marano, L.A., Mendes-Junior, C.T., Zucoloto, S. and Donadi, E.A, IL-18, TNF, and IFN-γ alleles and genotypes are associated with susceptibility to chronic hepatitis B infection and severity of liver injury, J. Med. Virol. 87 (2015) 1689-1696. https://doi.org/10.1002/jmv.24225. [CrossRef]
- N. Li, Y.F. Gao, T.C. Zhang, P. Chen, X. Li, F. Su, Relationship between interleukin 18 polymorphisms and susceptibility to chronic hepatitis B virus infection, World J Hepatol. 4 (2012) 105, https://doi.org/10.4254/wjh.v4.i3.105. [CrossRef]
- YS. Kim, JY. Cheong, SW. Cho, KM. Lee, JC. Hwang, B. Oh, K. Kimm, JA. Lee, BL. Park, HS. Cheong, HD. Shin, JH. Kim, A functional SNP of the interleukin-18 gene is associated with the presence of hepatocellular carcinoma in hepatitis B virus-infected patients, J. Dig. Dis. Sci. 54 (2009) 2722–2728, https://doi.org/10.1007/s10620-009-0970-6. [CrossRef]
- J.Y. Cheong, S.W. Cho, B. Oh, K. Kimm, K.M. Lee, S.J. Shin, J.A. Lee, B.L. Park, H.S. Cheong, H.D. Shin, and B.Y. Cho, Association of interleukin-18 gene polymorphisms with hepatitis B virus clearance, Dig. Dis. Sci. 55 (2010) 1113-1119, https://doi.org/10.1007/s10620-009-0819-z. [CrossRef]
- Zhu R, Mao S, Shi W, Wu L, Zhang J. A prediction study of IL-18 and IFN-γ in glucocorticoid treatment response in infants and young children with severe Mycoplasma pneumoniae pneumonia. Transl Pediatr. 2022 May;11(5):738-747. https://doi.org/10.21037/tp-22-139. [CrossRef]
- Z.J. Dai, X.H. Liu, M. Wang, Y. Guo, W. Zhu, X. Li, S. Lin, T. Tian, K. Liu, Y. Zheng, P. Xu, IL-18 polymorphisms contribute to hepatitis B virus-related cirrhosis and hepatocellular carcinoma susceptibility in Chinese population: a case-control study, Oncotarget, 8 (2017) 81350. https://doi.org/10.18632/oncotarget.18531. [CrossRef]
- H.T. Pohan, S. Suhendro, R. Bur, A. Matondang, S. Djauzi, K. Inada and S. Endo, Interleukin-18 levels in adult dengue fever and dengue hemorrhagic fever, Med. J. Indones. 13 (2004) 86-9, https://doi.org/10.13181/mji.v13i2.136. [CrossRef]
- L.B. Zhen, Y.P. Sun, Y.Y. Chen, L.S. Yin, IL-18 polymorphisms and tuberculosis susceptibility: a meta-analysis. Afr. Health Sci., 19(2019)1311-1320. https://doi.org/10.4314/ahs.v19i1.2. [CrossRef]
- C. He, L. Liu, Associations of polymorphisms in IL-6 and IL-18 with tuberculosis: evidence from a meta-analysis, Microb. Pathog. 139 (2020) 103823, https://doi.org/10.1016/j.micpath.2019.103823. [CrossRef]
- B. Zhang, L. Xiao, Q. Qiu, L. Miao, S. Yan and S. Zhou, Association between IL-18, IFN-γ and TB susceptibility: a systematic review and meta-analysis, Ann. Palliat. Med. 10 (2021) 108780886-108710886, https://doi.org/10.21037/apm-21-2582. [CrossRef]
- M. Han, J. Yue, Y.Y. Lian, Y.L. Zhao, H.X. Wang, L.R. Liu, Relationship between single nucleotide polymorphism of interleukin-18 and susceptibility to pulmonary tuberculosis in the Chinese Han population, Microbiol. Immunol. 55 (2011) 388-393, https://doi.org/10.1111/j.1348-0421.2011.00332.x. [CrossRef]
- Pacifico L, Anania C, Osborn JF, Ferraro F, Chiesa C. Consequences of Helicobacter pylori infection in children. World J Gastroenterol. 2010 16(41):5181-94. https://doi.org/ 10.3748/wjg.v16.i41.5181. [CrossRef]
- Rezaeifar, E. Eskandari-Nasab, M. Moghadampour, E. Kharazi-Nejad, S.S.A. Hasani, A. Asadi-Saghandi, M. Hadadi-Fishani, A. Sepanjnia, and B. Sadeghi-Kalani, The association of interleukin-18 promoter polymorphisms and serum levels with duodenal ulcer, and their correlations with bacterial CagA and VacA virulence factors, Scand. J. Infect. Dis. 45 (2013) 584-592, https://doi.org/10.3109/00365548.2013.794301. [CrossRef]
- D.S. Myung, W.S. Lee, Y.L. Park, N. Kim, H.H. Oh, M.Y. Kim, C.Y. Oak, C.Y. Chung, H.C. Park, J.S. Kim, and S.B. Cho. Association between interleukin-18 gene polymorphism and Helicobacter pylori infection in the Korean population, Scientific reports, 5 (2015) 11535. https://doi.org/10.1038/srep11535. [CrossRef]
- P.K. Sato, F.D. Busser, F.M.D.C. Carvalho, A. Gomes dos Santos, A. Sadahiro, C.L. Diogo, A.S.G. Kono, M.L. Moretti, O.D.C. Luiz and M.A. Shikanai-Yasuda, Polymorphism in the Promoter Region of the IL18 Gene and the Association with Severity on Paracoccidioidomycosis, Front. immunol. 11 (2020) 542210, https://doi.org/10.3389/fimmu.2020.542210. [CrossRef]
- S.B. Anyona, P. Kempaiah, E. Raballah, C. Ouma, T. Were, G.C. Davenport, S.N. Konah, J.M. Vulule, J.B. Hittner, C.W. Gichuki, J.M. Ong’echa. Functional promoter haplotypes of interleukin-18 condition susceptibility to severe malarial anemia and childhood mortality. Infect. Immun, 79(2011), 4923-4932. https://doi.org/10.1128/iai.05601-11. [CrossRef]
- S.I. Oyedeji, H.O. Awobode, and J.F. Kun, Interleukin-18 Gene Promoter Polymorphisms in Children with Uncomplicated Malaria in Lafia, North-central Nigeria, FJPAS, 6 (2021) 10-19.
- VK. Karra, PK. Gumma, SJ. Chowdhury, R. Ruttala, SK. Polipalli, A. Chakravarti, P. Kar, IL-18 polymorphisms in hepatitis B virus related liver disease, Cytokine. 73 (2015) 277-282, https://doi.org/10.1016/j.cyto.2015.02.015. [CrossRef]
- K. Manohar, P.V. Suneetha, N.T. Sukriti, Pati, A.C. Gupta, S. Hissar, P. Sakhuja, P. and S.K.Sarin, Association of IL-18 promoter polymorphism with liver disease severity in HCV-infected patients, Hepatol. Int. 3 (2009) 371-377, https://doi.org/10.1007/s12072-009-9127-0. [CrossRef]
- N. Bouzgarrou, E. Hassen, E. Schvoerer, F. Stoll-Keller, O. Bahri, S. Gabbouj, I. Cheikh, N. Maamouri, N. Mammi, H. Saffar, A. Trabelsi, Association of interleukin-18 polymorphisms and plasma level with the outcome of chronic HCV infection, J. Med. Virol. 80 (2008) 607-614, https://doi.org/10.1002/jmv.21079. [CrossRef]
- M. Harishankar, P. Selvaraj, DN. Rajeswari, SP. Anand, PR. Narayanan, Promoter polymorphism of IL-18 gene in pulmonary tuberculosis in South Indian population, Int. J. Immunogenet. 34 (2007) 317-320, https://doi.org/10.1111/j.1744-313X.2007.00714.x. [CrossRef]
- C. Zhou, N. Ouyang, Q.H. Li, S.H. Luo, Q. He, S. Lei, H. and Q. Liu, The-137G/C single nucleotide polymorphism in IL-18 gene promoter contributes to tuberculosis susceptibility in Chinese Han population, Infect. Genet. Evol. 36 (2015) 76-380. https://doi.org/10.1016/j.meegid.2015.10.014. [CrossRef]
- M. Ponnana, R. Sivangala, L. Joshi, V. Valluri and S. Gaddam, IL-6 and IL-18 cytokine gene variants of pulmonary tuberculosis patients with co-morbid diabetes mellitus and their household contacts in Hyderabad, Gene. 627 (2017) 298-306, https://doi.org/10.1016/j.gene.2017.06.046. [CrossRef]
- Urazova, E.G. Churina, R.R. Hasanova, V.V. Novitskiy and V.S.Poletika, Association between polymorphisms of cytokine genes and secretion of IL-12p70, IL-18, and IL-27 by dendritic cells in patients with pulmonary tuberculosis, Tuberculosis. 115 (2019) 56-62, https://doi.org/10.1016/j.tube.2019.02.003. [CrossRef]
Figure 1.
IL-18 signal transduction and its biological effects.
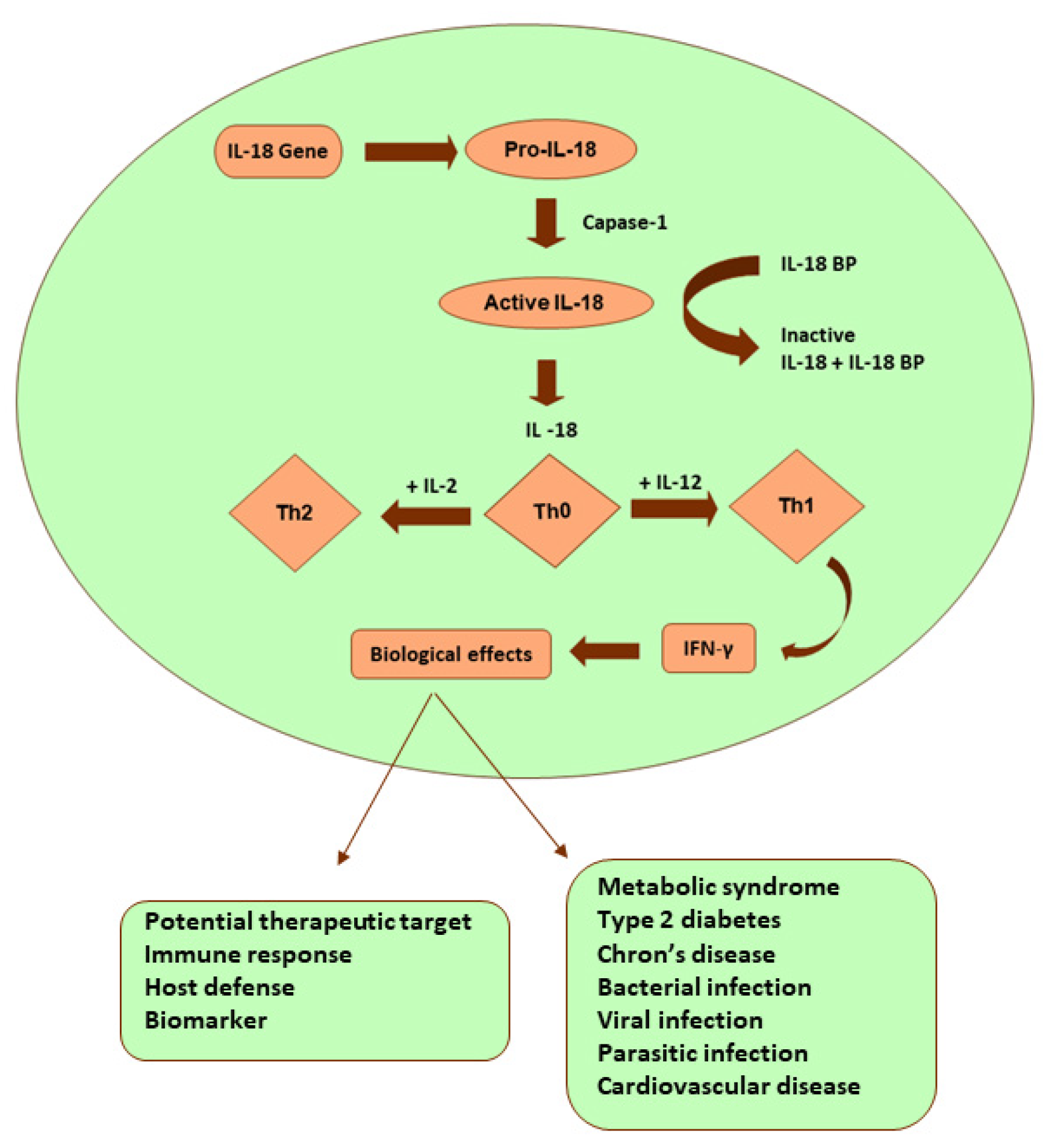
Figure 2.
Selected IL-18 polymorphisms associated with infectious diseases.
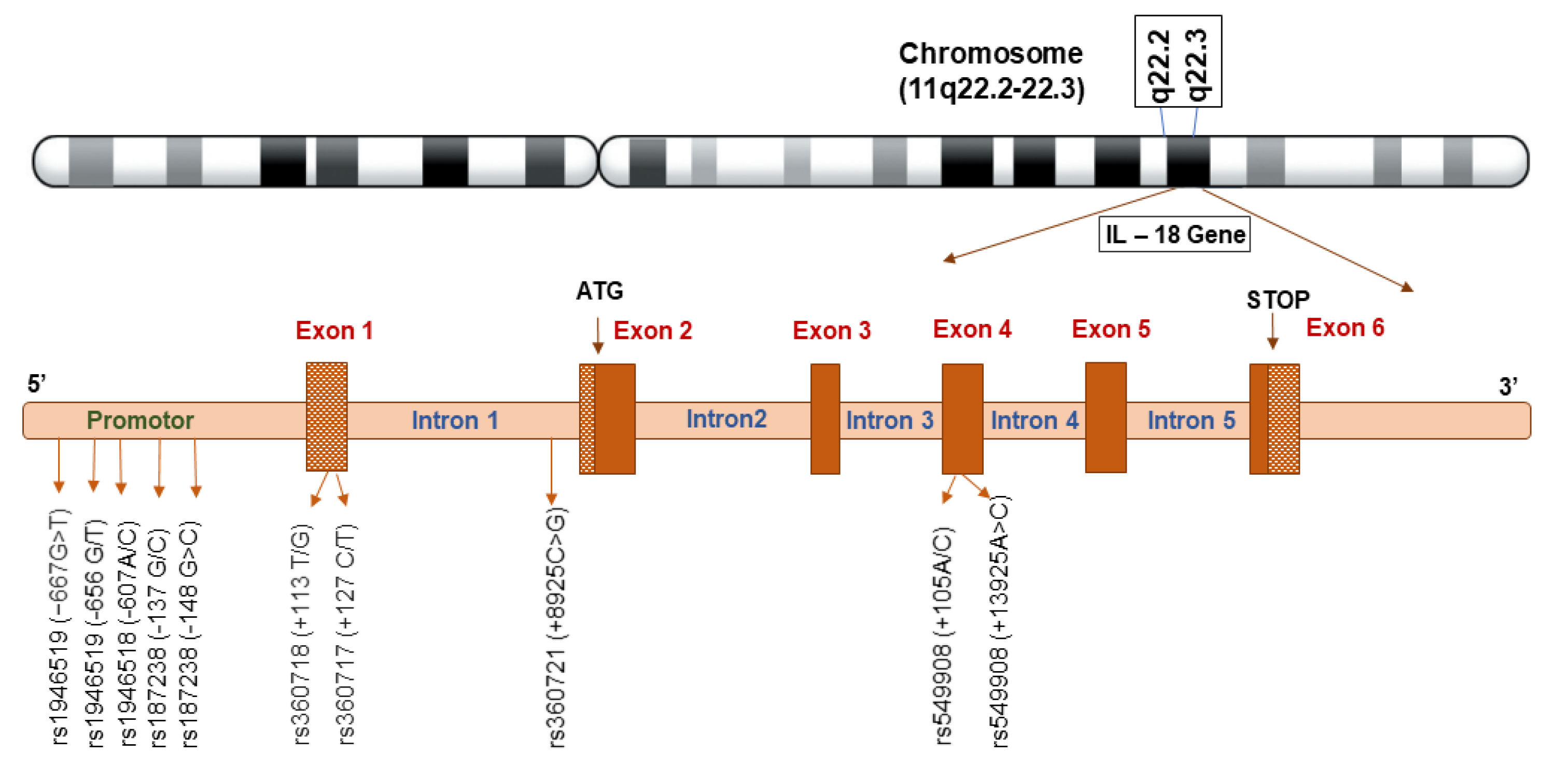

Table 1.
Studies on IL-18 polymorphisms and various infectious diseases.
| Infections | Population | SNPs | Location | Clinical impact | Reference |
|---|---|---|---|---|---|
| Visceral Leishmaniasis | Iranian 118 patients 156 controls |
rs1946519 (-656 G/T) rs187238 (-137 G/C) |
Promoter region | G allele at the position −656, a protective allele against VL. | [31]. |
| rs549908 (+105A/C) | Codon region | ||||
| Visceral Leishmaniasis | Indian 204 patients 267 controls |
rs1946519 (-656 G/T) rs187238 (-137 G/C) |
Promoter region | T allele at position−656, a protective against VL. | [30]. |
| rs549908 (+105A/C) | Codon region | ||||
| Visceral Leishmaniasis |
Iranian (East Azarbaijan) 91 patients, 185 controls |
rs1946518 (-607A/C) rs187238 (-137G/C) |
Promoter region | No significant association |
[32]. |
| Malaria | Saudi Arabian 250 patients 200 controls |
rs574429 (G to C) rs544354 (G to C) |
Promoter region | Increased susceptibility to P. falciparum infection and related parasitemia | [25]. |
| Severe malarial anemia (SMA) | Western Kenyan 123 patients 400 controls |
rs1946518 (-607C/A) rs187238 (-137G/C) |
Promoter region | −137G/−607C (GC) haplotype associated with increased susceptibility to SMA. −137C/−607C (CC) significantly associated with childhood mortality |
[68]. |
| Uncomplicated Malaria |
Nigerian 171 patients 166 controls |
rs1946518 (-607C/A) rs187238(-137G/C) |
Promoter region | No significant association |
[69]. |
| Chronic Chagas Disease |
Brazilian 849 patients 202 controls |
rs2043055 (A/G) | Promoter region |
No significant association |
[34]. |
| Intestinal amoebiasis |
Iraq 25 patients 25 controls |
rs1866694757(CC /CA) rs1946518(TT /TG) rs1946519(CC/AC) rs1215648807(AA /AC) |
Promoter region |
Increased susceptibility and associated with progression of E. histolytica infection |
[36]. |
| rs940255648 (GG /GC) rs1037707423 (CC /CT) rs1213044637 (GG /GA) rs1866697972 (GG /GT) rs1866698066 (GG /GT) rs1866698286 (AA /TT) |
Promoter region | Protection against progression of infection | |||
| HIV infection | Indian 500 patients 500 controls |
rs187238(-137G/C) | Promoter region |
Associated with the progression of HIV-1/AIDS | [38]. |
| HBV infection | Indian 271 patients 280 controls |
rs1946518(-607 A/C) rs187238(-137 C/G) |
Promoter region |
Allele A at position -607: a protective allele against HBV infection Allele C at position -137 is associated with increased susceptibility to HBV infections. |
[70]. |
| HBV infection | Chinese 376 patients 254 controls |
rs187238 (-137 G allele and GG genotype -137 G/C) |
Promotor region |
Protection against HBV infection |
[40]. |
| HBV infection | Chinese Han 231 patients 300 controls |
rs187238 (-137 G/C) rs1946518 (-607C/A) |
Promotor region | -137C allele protective against HBV infection -607AA genotype associated with increased susceptibility to HBV |
[41]. |
| HBV infection | Thailand 140 patients 140 controls |
rs1946518 (-607 A/A) |
Promotor region | Associated with progression of HBV infection. | [50]. |
| HBV infection | Korean 730 patients 320 controls |
rs1946519 (−667G>T) rs187238 (-148 G>C) |
Promotor region |
Associated with HBV clearance |
[56]. |
| rs360721 (+8925C>G) | Intron 1 | ||||
| rs549908 (+13925A>C) | Exon 4 | ||||
| HBV-related HCC* |
Chinese 153 Patients 165 controls |
rs1946518 (-607 C/A) rs187238 (-137 G/C) |
Promoter region | Protective allele against HBV-related HCC | [52]. |
| HCV infection |
Indian 204 Patients 350 controls |
rs1946518 (-607C/A) |
Promoter region | −607 position with A/A allele associated with protection against disease severity |
[71]. |
| HCV infection |
Tunisian 81 patients 82 controls |
rs187238 (-137G/C) | Promoter region | Associated with disease severity |
[72]. |
| Tuberculosis |
Indian 165 patients 173 controls |
rs1946518 (–607(C/A) rs187238 (–137G/A) |
Promoter region | No significant association |
[73]. |
| Tuberculosis | Chinese 407 patients 469 controls |
rs1946518 (–607(C/A) rs187238 (–137G/C) |
Promoter region | Increased susceptibility to tuberculosis | [74]. |
| Tuberculosis | Indian 505 patients 200 controls |
rs1946518 (–607(C/A) rs187238 (–137G/C) rs1800795 (-174G>C) |
Promoter region | AC genotype of IL-18 -607A>C increased susceptibility to PTB in the patients affected with co-morbid diabetes mellitus | [75]. |
| Tuberculosis | Russia 334 patients 183 Controls |
rs549908 (105C/A) | Promoter region | C allele and CC genotype of 105 A/C predisposes to the progression of disseminated TB |
[76]. |
| H-Pylori infection |
Korean 456 Patients 222 controls |
rs1946519 (−656 G/T) rs1946518 (−607 C/A) rs187238 (−137G/C) rs360718 (+113 T/G) rs360717 (+127 C/T) |
Promoter region |
−137G/C, 113T/G, +127C/T associated with protection against H. pylori infection | [66]. |
Note: VL - Visceral Leishmaniasis, SME - Severe malarial anemia, HIV – Human Immunodeficiency Virus, HBV – Hepatitis B virus, HCV – Hepatitis C virus, HCC – Hepatocellular carcinoma, TB – Tuberculosis, H-Pylori - Helicobacter pylori.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



