Submitted:
09 September 2024
Posted:
10 September 2024
You are already at the latest version
Abstract
Cell-free DNA (cfDNA) analysis is a pivotal tool in non-invasive diagnostics, including cancer monitoring and prenatal testing. However, the preanalytical phase, particularly the choice of anticoagulant, significantly impacts cfDNA integrity and yield. This study aims to compare cfDNA yield, stability, and DNase activity across plasma-citrate, plasma-heparin, plasma-EDTA, and serum, to identify the most suitable specimen for cfDNA analysis among the tested anticoagulants. Blood samples from 15 healthy volunteers were collected in four types of tubes (citrate, heparin, EDTA, and serum). cfDNA was extracted and quantified using qPCR, and endogenous DNase activity was assessed through hydrolysis probe assays. Samples were incubated at 37°C for 24 hours to evaluate cfDNA degradation rates. Statistical analyses were performed using repeated measures ANOVA and Sidak's multiple comparison tests. Baseline cfDNA levels were highest in serum, followed by heparin-plasma, while EDTA and citrate-plasma exhibited the lowest levels. DNase activity was fully inhibited in EDTA-plasma, partially inhibited in citrate-plasma, and active in both heparin-plasma and serum. cfDNA degradation was most pronounced in heparin-plasma (85.3%) and serum (55.6%), compared to minimal degradation in EDTA (8%) and citrate-plasma (13.3%). Among the tested anticoagulants, EDTA-plasma proved to be the most suitable specimen for cfDNA analysis due to its low DNase activity and minimal gDNA contamination. Citrate-plasma, with intermediate DNase inhibition, could be an alternative if the citrate concentration were increased. Heparin-plasma and serum showed significant cfDNA degradation, rendering them unsuitable for cfDNA analysis. These findings highlight the importance of anticoagulant selection in preserving cfDNA integrity for diagnostic applications.
Keywords:
Cell-Free DNA
; Serum
; EDTA Plasma
; Heparin Plasma
; Citrate Plasma
; Nuclease Activity
; DNase Activity
1. Introduction
Cell-free DNA (cfDNA) analysis has revolutionized non-invasive diagnostics, offering genetic insights from blood samples without the need for invasive tissue biopsies [1]. In oncology, circulating tumor DNA (ctDNA) allows for the detection of tumor-specific mutations, aiding in early cancer diagnosis, monitoring treatment response, and identifying minimal residual disease [2,3]. This method is particularly beneficial as it captures tumor heterogeneity and allows for repeat sampling to track disease progression [1].
In prenatal care, cfDNA analysis has enabled non-invasive prenatal testing (NIPT) for fetal chromosomal abnormalities like trisomy 21, 18, and 13, with high accuracy. This technology has reduced the need for invasive procedures like amniocentesis, offering a safer option for both mother and fetus [4,5].
The preanalytical phase is critical in cfDNA analysis, as it affects cfDNA yield, quality, and integrity [6]. Key factors include the blood collection tube type and time to processing [7]. Additionally, DNase activity and the interaction of tube additives can significantly impact cfDNA stability [8,9], making proper management of these variables essential for reliable analysis.
Specialized tubes like Streck and PAXgene effectively stabilize leukocytes and prevent genomic DNA (gDNA) contamination by preserving cell integrity [8,10,11]. Streck tubes, with their formaldehyde-releasing agent, better prevent cell lysis and maintain cfDNA stability for up to 7 days at room temperature, while inhibiting DNase activity [8]. Although PAXgene tubes offer similar DNase inhibition, they are less effective in long-term prevention of cell lysis [10,11]. Both tubes, however, are specialized and may not always be available for cfDNA studies.
The American Society of Clinical Oncology and the College of American Pathologists recommend using plasma-EDTA for cfDNA collection, with processing within 6 hours to minimize leukocyte lysis and gDNA contamination [12]. Plasma-EDTA benefits from EDTA-mediated inhibition of blood DNases, preserving cfDNA during the preanalytical phase when immediate processing is not feasible [9]. When specialized tubes like Streck or PAXgene are unavailable, plasma-EDTA, with strict time management, serves as a viable alternative.
Serum, on the other hand, presents challenges for cfDNA analysis. While it may appear to yield higher cfDNA levels, this is primarily due to contamination with gDNA from lysed leukocytes during clotting, which compromises the sensitivity and specificity of the analysis [4,13]. Additionally, DNase activity in serum accelerates cfDNA degradation [9], making it less suitable for cfDNA analysis compared to plasma collected in EDTA or specialized tubes designed to preserve cfDNA integrity.
Other common anticoagulants, such as heparin and citrate, are less well-characterized for cfDNA analysis. Although these anticoagulants are typically chosen based on specific test requirements [14], their impact on cfDNA yield, gDNA contamination, and DNase activity remains insufficiently understood, highlighting a gap in their evaluation for cfDNA studies [15].
This study aims to compare cfDNA yield, stability, and DNase activity across plasma-citrate, plasma-heparin, plasma-EDTA, and serum specimens. We will evaluate the initial cfDNA amount in plasma-citrate and plasma-heparin, using plasma-EDTA and serum as positive and negative controls, respectively. Plasma-EDTA is expected to show low gDNA contamination and DNase inhibition, while serum is anticipated to have high gDNA contamination and DNase activity. Additionally, we will assess whether plasma-citrate and plasma-heparin could be viable alternatives to plasma-EDTA for preserving cfDNA integrity. By identifying the best specimen for cfDNA analysis among the tested ones, this study may also reveal key cellular and molecular mechanisms influencing cfDNA stability.
2. Materials and Methods
2.1. Subjects, Blood Collection, and Processing
This experimental study enrolled 15 healthy volunteers (7 men and 8 women). Blood samples were simultaneously collected in four 4 mL tubes: Vacuette K3EDTA (coated with 1.8 mg of K3EDTA per mL of blood), Vacuette Sodium Citrate (containing 0.109 mol/L (3.2%) sodium citrate), Vacuette Sodium Heparin (14 IU of heparin/mL of blood), and Vacuette Z Serum with Clot Activator (plain tubes coated with micronized silica particles), all from Greiner Bio-One. Within 10-15 minutes after venipuncture, the tubes were centrifuged at 2,000 g for 10 minutes at room temperature. Subsequently, 1,000 μL of the supernatants were transferred into polypropylene tubes and stored at −20°C until further use. After thawing, the samples were centrifuged again at 14,000 g for 10 minutes at room temperature, and paired supernatants were used for subsequent experiments.
2.2. Endogenous DNase Activity Assay
Endogenous DNase activity was assessed as previously described, and detailed description of the method can be found elsewhere [9]. Briefly, the assay included 20 μL of the crude samples, 250 nM of a hydrolysis probe (5'-6-FAM-CTCCAGCTC/ZEN/CACCTGAACGGCC-IABFQ-3'), 10 μL of 2X Maxima Probe qPCR Master Mix (Thermo Fisher Scientific), and nuclease-free water (Thermo Fisher Scientific) to a final volume of 35 μL. The reactions were incubated at a constant temperature of 37°C for 24 hours using the Step-One qPCR System (Thermo Fisher Scientific). Fluorescence was measured at the beginning of the reaction and then at 30-minute intervals throughout the incubation period.
The Maxima Probe qPCR Master Mix was utilized as a source of the passive reference dye (ROX) to normalize the fluorescence readings. The data were presented as delta Rn (ΔRn), calculated by dividing the FAM fluorescence by the ROX fluorescence, followed by subtraction of the baseline value, which was established using the initial fluorescence measurement. This approach allowed us to monitor the degradation of the single-stranded DNA probe by endogenous DNase activity. Nuclease-free water and DNase I (Thermo Fisher Scientific) were employed as negative and positive controls, respectively.
2.3. Specimen Treatment
Serum and plasma samples were incubated at 37°C for 24 hours to investigate the effect of endogenous nucleases on cfDNA. Treatments were conducted prior to DNA extraction, and cfDNA levels were measured as described below.
2.4. DNA Extraction
DNA was extracted from 500 μL of each sample using the generic protocol 2.0.1 of the NucliSens EasyMAG system (BioMérieux). The extraction involved 50 μL of magnetic silica particle suspension, and the DNA was eluted in 25 μL.
2.5. cfDNA Quantification
cfDNA was quantified using qPCR, targeting a 60 base pair sequence from the RNAse P gene. The reactions were performed on the Step-One qPCR System (Thermo Fisher Scientific) under the following conditions: 12.5 μL of 2X Maxima Probe qPCR Master Mix (Thermo Fisher Scientific), 2.5 μL of 10X PrimeTime qPCR Assay (Integrated DNA Technologies), consisting of the primers RNAse P-F (5'-AGATTTGGACCTGCGAGCG-3'), RNAse P-R (5'-GAGCGGCTGTCTCCACAAGT-3'), and the RNAse P probe (5'-HEX-TTCTGACCT/ZEN/GAAGGCTCTGCGCG-IABFQ-3'). A total of 5 μL of extracted DNA was used in a final reaction volume of 25 μL. The thermal cycling conditions were as follows: an initial pre-incubation at 95°C for 10 minutes, followed by 40 cycles of 15 seconds at 95°C and 15 seconds at 60°C. The RNAse P calibration curve was described by the equation Y = -3.306X + 36.613 (efficiency = 100.68% and R² = 0.998). All samples were run in duplicate, and each run included a no-template control.
The quantity of cfDNA in Genomic Equivalents (GE) per reaction was determined by comparison with a 5-fold dilution series of a well-characterized DNA sample (ranging from 150,000 to 5 GE per reaction) extracted from whole blood using the QIAamp DNA Blood Mini Kit (QIAGEN). A conversion factor of 6.6 pg of DNA per cell (GE) was applied. The RNAse P concentration in GE/mL was calculated as previously described [16]. This measurement in GE/mL was chosen because it offers a direct estimation of the number of cells. To convert GE/mL to ng/mL, multiply the GE/mL value by 6.6×10⁻³. The qPCR experiments and their description followed the Minimum Information for qPCR Experiments (MIQE) guidelines [17].
2.6. Statistical Analysis
The statistical methods employed included the D'Agostino & Pearson omnibus normality test to assess data distribution, followed by repeated measures one-way ANOVA for comparing multiple conditions. Sidak’s multiple comparisons test was used for post-hoc pairwise comparisons to control for type I error. The significance level was set at p < 0.05 for all analyses. Data that passed the normality test were further analyzed using the repeated measures ANOVA. All graphs and statistical analyses were performed using Prism 6.0h software.
3. Results
3.1. Basal cfDNA Yields in Serum, Heparin-Plasma, Citrate-Plasma, and EDTA-Plasma
First, we measured the basal cfDNA levels (15 minutes after blood draw) in serum, heparin-plasma, citrate-plasma, and EDTA-plasma. The basal cfDNA yields (mean ± SD) were highest in serum, at 1473 ± 797 GE/mL, significantly exceeding those in the other specimens (p < 0.0001 compared to heparin, citrate, and EDTA). Heparin-plasma presented intermediate cfDNA levels of 472 ± 190 GE/mL, which were distinct from those in serum (p < 0.0001), citrate-plasma (p < 0.0001), and EDTA-plasma (p < 0.0001). Citrate-plasma and EDTA-plasma had the lowest cfDNA quantities, 147.4 ± 41.6 GE/mL and 154.8 ± 37.3 GE/mL, respectively, and these quantities were not significantly different from each other (p > 0.99) (Figure 1).
3.2. Direct Measurement of the Endogenous DNase Activity in Serum, Heparin-Plasma, Citrate-Plasma, and EDTA-Plasma
Next, we measured the endogenous DNase activity in serum, heparin-plasma, citrate-plasma, and EDTA-plasma using a fluorescent DNase activity assay based on hydrolysis probe degradation, to evaluate the effect of each tube additive on blood DNase activity. Sustained hydrolysis probe degradation (indicating DNase activity) was observed over time in serum, heparin-plasma, and most of citrate-plasma samples. The kinetics of probe degradation in serum and heparin-plasma were similar, indicating active DNase presence; however, serum consistently exhibited lower delta Rn levels at each time point compared to heparin-plasma.
In contrast, the kinetics of probe degradation in citrate-plasma differed from those in serum and heparin-plasma, suggesting a partial inhibitory effect on DNase activity. On the other hand, in EDTA-plasma, the probe degradation plateaued at the beginning of the reaction, with no further increase observed, confirming an extensive inhibitory effect of EDTA on blood DNase activity (a representative depiction of endogenous DNase activity in a paired set of tested plasmas and serum can be found in Figure 2A).
Considering the last reading point of the DNase activity assay, the mean (SD) delta Rn observed in serum and heparin-plasma was [1.277 ± 0.32 delta Rn] and [1.479 ± 0.43 delta Rn], respectively, with no significant difference between them (p = 0.17). Citrate-plasma exhibited an intermediate DNase activity level [0.85 ± 0.53 delta Rn], which was significantly different from heparin-plasma (p < 0.0001), serum (p = 0.0002), and EDTA-plasma (p < 0.0001). DNase activity in EDTA-plasma was highly inhibited [0.09 ± 0.009 delta Rn], and significantly different from heparin-plasma, serum, and citrate-plasma (p < 0.0001 for all) (Figure 1B).
3.3. Effect of Endogenous DNase Activity on cfDNA Levels in Plasma and Serum
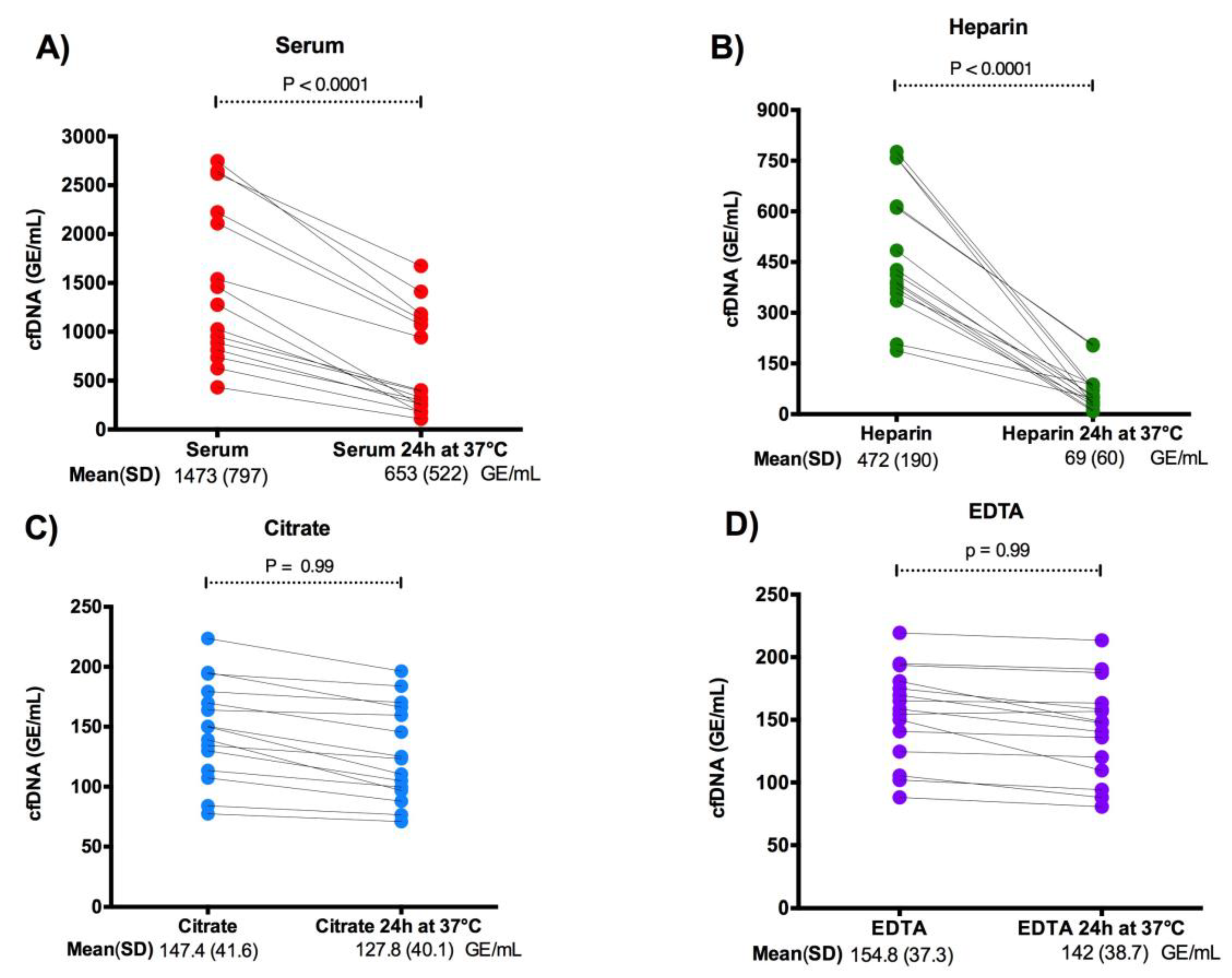
Next, to observe the effect of blood DNase activity on cfDNA ex-vivo, we measured the degradation of cfDNA by quantifying its levels after incubation at 37°C for 24 hours. In serum, cfDNA levels decreased by a mean of 55.6% [from 1473 ± 797 GE/mL to 653 ± 522 GE/mL, p < 0.0001] (Figure 3A). Heparin-plasma showed a mean reduction of 85.3% [from 472 ± 190 GE/mL to 69 ± 60 GE/mL, p < 0.0001] (Figure 3B). Conversely, in citrate-plasma, cfDNA levels declined by 13.3% [from 147.4 ± 41.6) GE/mL to 127.8 ± 40.1 GE/mL, p = 0.99] (Figure 3C), and in EDTA-plasma, there was an 8.0% reduction [from 154.8 ± 37.3 GE/mL to 142.4 ± 38.7 GE/mL, p > 0.99] (Figure 3D).
4. Discussion
In this study, we comprehensively examined the baseline levels of cfDNA, endogenous DNase activity, and the impact on cfDNA stability across four sample types: serum, heparin-plasma, citrate-plasma, and EDTA-plasma. Our findings reveal three key points:
(a) EDTA-plasma and citrate-plasma exhibited significantly lower initial cfDNA concentrations compared to serum and heparin-plasma;
(b) DNase activity was almost completely inhibited in EDTA-plasma, partially inhibited in citrate-plasma, and remained active in both heparin-plasma and serum; and
(c) DNase-driven cfDNA degradation was substantial in serum and heparin-plasma but modest to minimal in citrate-plasma and EDTA-plasma.
Our quantitative analysis, conducted within 15 minutes of blood draw, showed that EDTA-plasma and citrate-plasma had lower baseline cfDNA levels compared to heparin-plasma and serum. These findings indicate that EDTA-plasma and citrate-plasma introduce minimal ex-vivo cfDNA, with values (~75-225 GE/mL or ~0.5-1.485 ng/mL) comparable to those reported in the circulation of healthy individuals (0.06 to 22.5 ng/mL) [18].
The elevated cfDNA levels observed in serum, as expected, were attributed to leukocyte lysis during coagulation [6]. Unexpectedly, heparin-plasma showed higher cfDNA levels than EDTA-plasma and citrate-plasma, despite the assumption that anticoagulation would prevent the release of additional gDNA into cfDNA.
These results suggest that different anticoagulants have distinct pre-analytical effects on baseline cfDNA levels. Heparin, citrate, and EDTA anticoagulate through different mechanisms.
Heparin works by binding to antithrombin, a naturally occurring inhibitor of blood clotting. Once bound, heparin significantly enhances antithrombin's ability to inhibit key clotting factors, such as thrombin (factor IIa), factor Xa, and factor IXa. This prevents the formation of fibrin, the protein that forms the structural framework of a blood clot [19].
On the other hand, EDTA and citrate inhibit coagulation by acting as divalent ion chelators. They bind to calcium ions (Ca²⁺), which are essential cofactors in the coagulation cascade. Calcium ions facilitate the activation of several clotting factors, including prothrombin (factor II) and factor X, which are crucial for converting fibrinogen to fibrin. By sequestering calcium, EDTA and citrate effectively disrupt this cascade, preventing blood clot formation [19].
The observed cfDNA levels in paired citrate-plasma, EDTA-plasma, and heparin-plasma indicate that different anticoagulants have distinct pre-analytical effects on baseline cfDNA levels due to their varied influences on blood coagulation and cell stability.
Corroborating this observed effect, in vitro research has shown that heparin can induce leukocyte lysis in a dose-dependent manner, with concentrations of 10 IU/mL causing 10-30% cell lysis [20]. Another in vitro study demonstrated that heparin at 10 IU/mL can also promote the formation of neutrophil extracellular traps (NETs) within 15 minutes, which are often associated with cell lysis, death, and the release of extracellular DNA [21]. We hypothesize that the slightly higher ex-vivo cfDNA levels observed in heparin-plasma shortly after blood draw (15 minutes) could be linked to this in-vitro phenomenon, as the amount of heparin in collection tubes typically ranges from 14-18 IU/mL [19].
These observations suggest that divalent ion chelators, such as EDTA and citrate, are preferable for cfDNA analysis, as they minimize unwanted gDNA contamination from leukocytes. This prevention reduces pre-analytical interference from non-tumor or non-fetal DNA sequences, preserving assay sensitivity in applications like liquid biopsies and fetal DNA analysis.
Our second major finding, that DNase activity is highly inhibited in EDTA-plasma, partially inhibited in citrate-plasma, and active in both heparin-plasma and serum, is supported by two key observations: (1) the direct measurement of DNase activity using a hydrolysis probe assay, and (2) cfDNA levels assessed after incubation at 37°C for 24 hours to promote DNA degradation [22].
DNase activity was highest in heparin-plasma and serum, intermediate in citrate-plasma, and lowest in EDTA-plasma. After incubation at 37°C for 24 hours, cfDNA degradation was most pronounced in heparin-plasma (85.3%) and serum (55.6%), compared to citrate (13.3%) and EDTA (8%).
These results can be explained by the fact that EDTA and citrate chelate divalent ions (Mg²⁺ and Ca²⁺), rendering them unavailable for DNase activity, as these ions are essential for most blood DNases [23,24]. This dual effect—preventing both coagulation and DNase activity—makes EDTA the preferred anticoagulant for cfDNA analysis, as it minimizes gDNA contamination and reduces DNase activity.
Citrate-plasma showed intermediate DNase inhibition, with cfDNA degradation rates similar to EDTA-plasma after 24 hours at 37°C (13% versus 8%). While citrate interferes with DNase activity, the concentration tested (3.2%) may be insufficient to achieve the higher inhibition seen with EDTA. Despite this intermediate DNase activity, the baseline cfDNA amount did not significantly change after 24 hours at 37°C, making citrate-plasma the second-best option for cfDNA analysis among the specimens tested when plasma separation is performed soon after blood draw. The main drawback remains the intermediate DNase activity. Indeed, we found that increasing the citrate concentration to 8% in collection tubes improves its effectiveness, making it comparable to EDTA for cfDNA analyses (preliminary results) [25].
A previous study evaluated the cfDNA quality of citrate-plasma and EDTA-plasma in terms of gDNA contamination after 72 hours of storage at 4°C. It demonstrated that citrate-plasma (3.2%) provided superior cfDNA quality, with minimal gDNA contamination, compared to EDTA-plasma, which exhibited larger cfDNA fragments due to gDNA contamination. The cfDNA quality and quantity in citrate tubes were maintained throughout this period, making the results comparable to those obtained with cell-stabilizing tubes like Streck [26].
In the above-cited study, better results from citrate-plasma were observed only after 72 hours of contact with leukocytes, and they were comparable to EDTA before this time. Storage at 4°C is less favorable for blood DNase activity and could lead to its full inhibition. While the pre-analytical conditions in our study and the previously mentioned one were different and not directly comparable, taken together, these observations suggest that citrate-plasma is a viable option for cfDNA analysis. Storing samples at 4°C may compensate for the intermediate DNase activity observed, thereby helping preserve cfDNA quality.
Heparin-plasma and serum, with fully active DNases, present significant drawbacks due to gDNA contamination and cfDNA degradation, making these specimens unsuitable for cfDNA analysis. Despite this, several studies on cfDNA have used heparin-plasma. The results from these prior cfDNA studies should be interpreted cautiously, considering the preanalytical effects observed in our study.
Although DNase activity in heparin-plasma was not significantly different from serum, cfDNA degradation was higher in heparin-plasma (85.3% vs. 55.6%). This could be attributed to serum DNase inhibition by G-actin released from lysed cells [27], which is more pronounced due to higher gDNA contamination. On the other hand, heparin at 10 IU/mL has been shown to form polymeric complexes with cell-free DNA in whole blood [28] and facilitates DNase I-mediated digestion of cfDNA, independent of its anticoagulant properties [29].
Since heparin-plasma exhibited higher cfDNA levels compared to citrate-plasma and EDTA-plasma, which seem to represent the basal levels of cfDNA in circulation, our observations support the hypothesis that heparin, at least to some extent, directly induces ex-vivo cell death and the subsequent release of gDNA into plasma. The simultaneous occurrence of G-actin inhibition in serum and heparin stimulation of DNase I activity could have contributed to the observed cfDNA degradation levels after 24 hours of incubation at 37°C in our experiments.
Indeed, heparin is not the sample of choice for cfDNA studies due to its inhibitory effects on polymerase chain reaction (PCR) assays. Heparin is known to inhibit DNA polymerase activity, which can significantly interfere with the amplification of cfDNA, leading to unreliable results [30,31]. However, the addition of heparinase, an enzyme that degrades heparin, has been shown to enable reliable quantification of circulating tumor DNA (ctDNA) from heparinized plasma samples using droplet digital PCR [31].
We cannot exclude the possibility that heparin inhibited the qPCR reactions observed in this study. However, it is unlikely that modern DNA extraction methods, which result in highly purified DNA templates, are significantly affected by this issue. Although we did not observe any qPCR inhibition, the experiments were not specifically designed to test for this effect. We speculate that the observed inhibitory effect of heparin on cfDNA could be due to its high DNase activity and cfDNA degradation, potentially being confounded with PCR inhibition.
Additionally, several studies have consistently shown that heparin use in pregnant women poses a preanalytical problem for non-invasive prenatal testing (NIPT) due to increased degradation of cell-free fetal DNA to the production of shorter cfDNA fragments and lower fetal fractions [32,33]. These factors lead to more frequent retests and a significantly higher likelihood of test failure, with an odds ratio of 21.87 for nonreportable results in heparin-treated patients [34].
This underscores the need for careful consideration of heparin use in NIPT testing. We hypothesize that the increased degradation of cell-free fetal DNA and non-reportable NIPT results in women using heparin could be secondary to increased blood DNase activity in-vivo. This hypothesis should be tested in specific experiments evaluating DNase activity in individuals using heparin.
5. Conclusions
In conclusion, among the tested anticoagulants, EDTA-plasma proved to be the most suitable for cfDNA analysis due to its minimal DNase activity and low gDNA contamination. Citrate-plasma showed intermediate DNase inhibition and low gDNA contamination, and increasing the citrate concentration in collection tubes could further improve its effectiveness. In contrast, heparin-plasma and serum exhibited the highest DNase activity and gDNA contamination, resulting in significant cfDNA degradation, making them less ideal for cfDNA analysis. These findings underscore the importance of selecting the appropriate anticoagulant for accurate cfDNA analysis, particularly in applications such as liquid biopsies and prenatal testing. Extreme caution must be exercised when interpreting results from previous cfDNA studies that utilized serum and heparin-plasma.
Author Contributions
Conceptualization, G.B.B. and T.S.H.R.; methodology, G.B.B. and RHJ; formal analysis, G.B.B., T.S.H.R. and RHJ; resources, L.F.A.N; data curation, G.B.B.; writing—original draft preparation, T.S.H.R.; writing—review and editing, G.B.B. and R.H.J; supervision, R.H.J.; project administration, G.B.B.; funding acquisition, R.H.J. and L.F.A.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and was approved by the Ethics Committee of the Federal District Faculty of Health Sciences (FEPECS) under protocol code CAAE 35684414.5.0000.5553.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
All data supporting the results reported in this study are included within the article.
Acknowledgments
We thank Janete Ana Ribeiro Vaz and Sandra Santana Soares Costa for their support in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Stewart, C.M.; Kothari, P.D.; Mouliere, F.; Mair, R.; Somnay, S.; Benayed, R.; Zehir, A.; Weigelt, B.; Dawson, S.-J.; Arcila, M.E.; et al. The Value of Cell-Free DNA for Molecular Pathology. J Pathol 2018, 244, 616–627. [Google Scholar] [CrossRef]
- De, S. Signatures Beyond Oncogenic Mutations in Cell-Free DNA Sequencing for Non-Invasive, Early Detection of Cancer. Front Genet 2021, 12, 759832. [Google Scholar] [CrossRef] [PubMed]
- Dudley, J.C.; Diehn, M. Detection and Diagnostic Utilization of Cellular and Cell-Free Tumor DNA. Annu Rev Pathol 2021, 16, 199–222. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, K.; Weimer, E.T. Characteristics, Properties, and Potential Applications of Circulating Cell-Free Dna in Clinical Diagnostics: A Focus on Transplantation. J Immunol Methods 2018, 463, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Szilágyi, M.; Pös, O.; Márton, É.; Buglyó, G.; Soltész, B.; Keserű, J.; Penyige, A.; Szemes, T.; Nagy, B. Circulating Cell-Free Nucleic Acids: Main Characteristics and Clinical Application. Int J Mol Sci 2020, 21, 6827. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Pan, M.; Zhou, Z.; Chen, C.; Xing, X.; Cheng, S.; Zhang, S.; Zheng, H.; Qian, K. The Impact of Preanalytical Variables on the Analysis of Cell-Free DNA from Blood and Urine Samples. Front Cell Dev Biol 2024, 12, 1385041. [Google Scholar] [CrossRef]
- Ungerer, V.; Bronkhorst, A.J.; Holdenrieder, S. Preanalytical Variables That Affect the Outcome of Cell-Free DNA Measurements. Crit Rev Clin Lab Sci 2020, 57, 484–507. [Google Scholar] [CrossRef]
- Medina Diaz, I.; Nocon, A.; Mehnert, D.H.; Fredebohm, J.; Diehl, F.; Holtrup, F. Performance of Streck cfDNA Blood Collection Tubes for Liquid Biopsy Testing. PLoS One 2016, 11, e0166354. [Google Scholar] [CrossRef]
- Barra, G.B.; Santa Rita, T.H.; de Almeida Vasques, J.; Chianca, C.F.; Nery, L.F.A.; Santana Soares Costa, S. EDTA-Mediated Inhibition of DNases Protects Circulating Cell-Free DNA from Ex Vivo Degradation in Blood Samples. Clin Biochem 2015, 48, 976–981. [Google Scholar] [CrossRef]
- Toro, P.V.; Erlanger, B.; Beaver, J.A.; Cochran, R.L.; VanDenBerg, D.A.; Yakim, E.; Cravero, K.; Chu, D.; Zabransky, D.J.; Wong, H.Y.; et al. Comparison of Cell Stabilizing Blood Collection Tubes for Circulating Plasma Tumor DNA. Clin Biochem 2015, 48, 993–998. [Google Scholar] [CrossRef]
- Schmidt, B.; Reinicke, D.; Reindl, I.; Bork, I.; Wollschläger, B.; Lambrecht, N.; Fleischhacker, M. Liquid Biopsy - Performance of the PAXgene® Blood ccfDNA Tubes for the Isolation and Characterization of Cell-Free Plasma DNA from Tumor Patients. Clin Chim Acta 2017, 469, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Merker, J.D.; Oxnard, G.R.; Compton, C.; Diehn, M.; Hurley, P.; Lazar, A.J.; Lindeman, N.; Lockwood, C.M.; Rai, A.J.; Schilsky, R.L.; et al. Circulating Tumor DNA Analysis in Patients With Cancer: American Society of Clinical Oncology and College of American Pathologists Joint Review. J Clin Oncol 2018, 36, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Wong, F.C.K.; Sun, K.; Jiang, P.; Cheng, Y.K.Y.; Chan, K.C.A.; Leung, T.Y.; Chiu, R.W.K.; Lo, Y.M.D. Cell-Free DNA in Maternal Plasma and Serum: A Comparison of Quantity, Quality and Tissue Origin Using Genomic and Epigenomic Approaches. Clin Biochem 2016, 49, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Vignoli, A.; Tenori, L.; Morsiani, C.; Turano, P.; Capri, M.; Luchinat, C. Serum or Plasma (and Which Plasma), That Is the Question. J Proteome Res 2022, 21, 1061–1072. [Google Scholar] [CrossRef]
- Ayala-Lopez, N.; Conklin, S.E.; Tenney, B.J.; Ness, M.; Marzinke, M.A. Comparative Evaluation of Blood Collection Tubes for Clinical Chemistry Analysis. Clin Chim Acta 2021, 520, 118–125. [Google Scholar] [CrossRef]
- Lo, Y.M.; Tein, M.S.; Lau, T.K.; Haines, C.J.; Leung, T.N.; Poon, P.M.; Wainscoat, J.S.; Johnson, P.J.; Chang, A.M.; Hjelm, N.M. Quantitative Analysis of Fetal DNA in Maternal Plasma and Serum: Implications for Noninvasive Prenatal Diagnosis. Am J Hum Genet 1998, 62, 768–775. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin Chem 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Ivancic-Jelecki, J.; Brgles, M.; Santak, M.; Forcic, D. Isolation of Cell-Free DNA from Plasma by Chromatography on Short Monolithic Columns and Quantification of Non-Apoptotic Fragments by Real-Time Polymerase Chain Reaction. J Chromatogr A 2009, 1216, 2717–2724. [Google Scholar] [CrossRef]
- Lima-Oliveira, G.; Brennan-Bourdon, L.M.; Varela, B.; Arredondo, M.E.; Aranda, E.; Flores, S.; Ochoa, P. Clot Activators and Anticoagulant Additives for Blood Collection. A Critical Review on Behalf of COLABIOCLI WG-PRE-LATAM. Crit Rev Clin Lab Sci 2021, 58, 207–224. [Google Scholar] [CrossRef]
- Adachi, I.; Iwaki, H.; Adachi, H.; Ueno, M.; Horikoshi, I. Heparin-Induced Leukocyte Lysis in Vitro. J Pharmacobiodyn 1986, 9, 207–210. [Google Scholar] [CrossRef]
- Lelliott, P.M.; Momota, M.; Shibahara, T.; Lee, M.S.J.; Smith, N.I.; Ishii, K.J.; Coban, C. Heparin Induces Neutrophil Elastase-Dependent Vital and Lytic NET Formation. Int Immunol 2020, 32, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Fasco, M.J.; Kaminsky, L.S. Optimization of Dnase I Removal of Contaminating DNA from RNA for Use in Quantitative RNA-PCR. Biotechniques 1996, 20, 1012–1014. [Google Scholar] [CrossRef] [PubMed]
- Guéroult, M.; Picot, D.; Abi-Ghanem, J.; Hartmann, B.; Baaden, M. How Cations Can Assist DNase I in DNA Binding and Hydrolysis. PLoS Comput Biol 2010, 6, e1001000. [Google Scholar] [CrossRef] [PubMed]
- Barra, G. Blood Nucleases Affecting Circulating DNA in Serum and Plasma. In Cell-Free Circulating DNA; WORLD SCIENTIFIC, 2022; pp. 175–208 ISBN 9789811244674.
- Barra, G.B.; Santa Rita, T.H.; Jácomo, R.H.; Nery, L.F.A. Sodium Citrate at 8% Is Equivalent to EDTA as Anticoagulant of Choice for Circulation Cell-Free DNA Analysis: Low Contamination by Blood Cells Genomic DNA and Inhibition of Blood Nulcease Activity. In Proceedings of the Clinical Chemistry; Chicago, July 6 2014; Vol. 60, p. S190.
- Sato, A.; Nakashima, C.; Abe, T.; Kato, J.; Hirai, M.; Nakamura, T.; Komiya, K.; Kimura, S.; Sueoka, E.; Sueoka-Aragane, N. Investigation of Appropriate Pre-Analytical Procedure for Circulating Free DNA from Liquid Biopsy. Oncotarget 2018, 9, 31904–31914. [Google Scholar] [CrossRef] [PubMed]
- Dinsdale, R.J.; Hazeldine, J.; Al Tarrah, K.; Hampson, P.; Devi, A.; Ermogenous, C.; Bamford, A.L.; Bishop, J.; Watts, S.; Kirkman, E.; et al. Dysregulation of the Actin Scavenging System and Inhibition of DNase Activity Following Severe Thermal Injury. Br J Surg 2020, 107, 391–401. [Google Scholar] [CrossRef]
- de Vries, J.C.; Barendrecht, A.D.; Clark, C.C.; Urbanus, R.T.; Boross, P.; de Maat, S.; Maas, C. Heparin Forms Polymers with Cell-Free DNA Which Elongate Under Shear in Flowing Blood. Sci Rep 2019, 9, 18316. [Google Scholar] [CrossRef]
- Sohrabipour, S.; Muniz, V.S.; Sharma, N.; Dwivedi, D.J.; Liaw, P.C. Mechanistic Studies of DNase I Activity: Impact of Heparin Variants and PAD4. Shock 2021, 56, 975–987. [Google Scholar] [CrossRef]
- Holodniy, M.; Kim, S.; Katzenstein, D.; Konrad, M.; Groves, E.; Merigan, T.C. Inhibition of Human Immunodeficiency Virus Gene Amplification by Heparin. J Clin Microbiol 1991, 29, 676–679. [Google Scholar] [CrossRef]
- Sefrioui, D.; Beaussire, L.; Clatot, F.; Delacour, J.; Perdrix, A.; Frebourg, T.; Michel, P.; Di Fiore, F.; Sarafan-Vasseur, N. Heparinase Enables Reliable Quantification of Circulating Tumor DNA from Heparinized Plasma Samples by Droplet Digital PCR. Clin Chim Acta 2017, 472, 75–79. [Google Scholar] [CrossRef]
- Nakamura, N.; Sasaki, A.; Mikami, M.; Nishiyama, M.; Akaishi, R.; Wada, S.; Ozawa, N.; Sago, H. Nonreportable Rates and Cell-Free DNA Profiles in Noninvasive Prenatal Testing among Women with Heparin Treatment. Prenat Diagn 2020, 40, 838–845. [Google Scholar] [CrossRef]
- Wardrop, J.; Dharajiya, N.; Boomer, T.; McCullough, R.; Monroe, T.; Khanna, A. Low Molecular Weight Heparin and Noninvasive Prenatal Testing [22C]. Obstetrics & Gynecology 2016, 127, 32S. [Google Scholar] [CrossRef]
- Nitsche, J.F.; Lovell, D.; Stephens, N.; Conrad, S.; Bebeau, K.; Brost, B.C. The Effects of Heparin, Aspirin, and Maternal Clinical Factors on the Rate of Nonreportable Cell-Free DNA Results: A Retrospective Cohort Study. Am J Obstet Gynecol MFM 2023, 5, 100846. [Google Scholar] [CrossRef] [PubMed]
- Wardrop, J.; Dharajiya, N.; Boomer, T.; McCullough, R.; Monroe, T.; Khanna, A. Low Molecular Weight Heparin and Noninvasive Prenatal Testing [22C]. Obstetrics & Gynecology 2016, 127, 32S. [Google Scholar] [CrossRef]
- Nitsche, J.F.; Lovell, D.; Stephens, N.; Conrad, S.; Bebeau, K.; Brost, B.C. The Effects of Heparin, Aspirin, and Maternal Clinical Factors on the Rate of Nonreportable Cell-Free DNA Results: A Retrospective Cohort Study. Am J Obstet Gynecol MFM 2023, 5, 100846. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Basal cfDNA levels in genomic equivalents per mL (GE/mL) in serum, heparin-plasma, citrate-plasma, and EDTA-plasma samples, measured 15 minutes after blood draw. The cfDNA yield [mean (SD)] was highest in serum, followed by heparin-plasma, citrate-plasma, and EDTA-plasma (n = 15). Statistical analysis revealed that all comparisons were highly significant (P < 0.0001), except for the comparison between EDTA-plasma and citrate-plasma (P = 0.99).
Figure 1.
Basal cfDNA levels in genomic equivalents per mL (GE/mL) in serum, heparin-plasma, citrate-plasma, and EDTA-plasma samples, measured 15 minutes after blood draw. The cfDNA yield [mean (SD)] was highest in serum, followed by heparin-plasma, citrate-plasma, and EDTA-plasma (n = 15). Statistical analysis revealed that all comparisons were highly significant (P < 0.0001), except for the comparison between EDTA-plasma and citrate-plasma (P = 0.99).
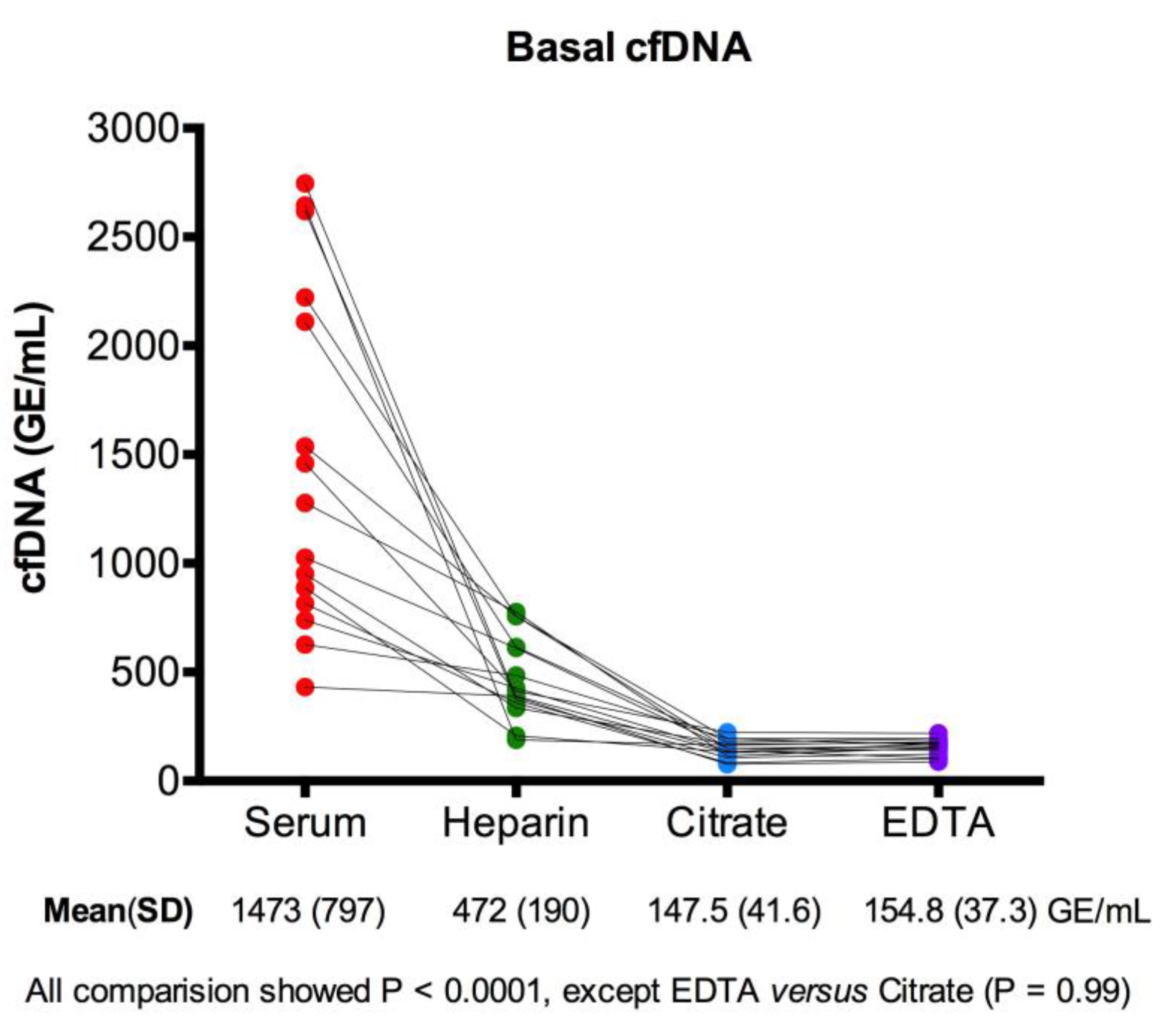
Figure 2.
A) Representative curves showing the endogenous DNase activity over time in serum, heparin-plasma, citrate-plasma, and EDTA-plasma. The curves illustrate the hydrolysis of a DNA probe, with activity represented as delta Rn (ΔRn) values over a 24-hour period. DNase activity was evident in serum, heparin-plasma, and citrate-plasma, with serum and heparin-plasma showing the highest activity. EDTA-plasma exhibited minimal activity, indicating strong inhibition of DNase. B) Final delta Rn (ΔRn) values from the DNase activity assay for individual samples (n = 15). Each line connects measurements from the same sample across different plasma and serum types. The mean (SD) delta Rn values are provided for each sample type, showing that DNase activity was highest in heparin-plasma and serum, partially inhibited in citrate-plasma, and fully inhibited in EDTA-plasma. Statistical analysis showed all comparisons to be significant (P < 0.0001) except for the comparison between serum and heparin-plasma (P = 0.17).
Figure 2.
A) Representative curves showing the endogenous DNase activity over time in serum, heparin-plasma, citrate-plasma, and EDTA-plasma. The curves illustrate the hydrolysis of a DNA probe, with activity represented as delta Rn (ΔRn) values over a 24-hour period. DNase activity was evident in serum, heparin-plasma, and citrate-plasma, with serum and heparin-plasma showing the highest activity. EDTA-plasma exhibited minimal activity, indicating strong inhibition of DNase. B) Final delta Rn (ΔRn) values from the DNase activity assay for individual samples (n = 15). Each line connects measurements from the same sample across different plasma and serum types. The mean (SD) delta Rn values are provided for each sample type, showing that DNase activity was highest in heparin-plasma and serum, partially inhibited in citrate-plasma, and fully inhibited in EDTA-plasma. Statistical analysis showed all comparisons to be significant (P < 0.0001) except for the comparison between serum and heparin-plasma (P = 0.17).
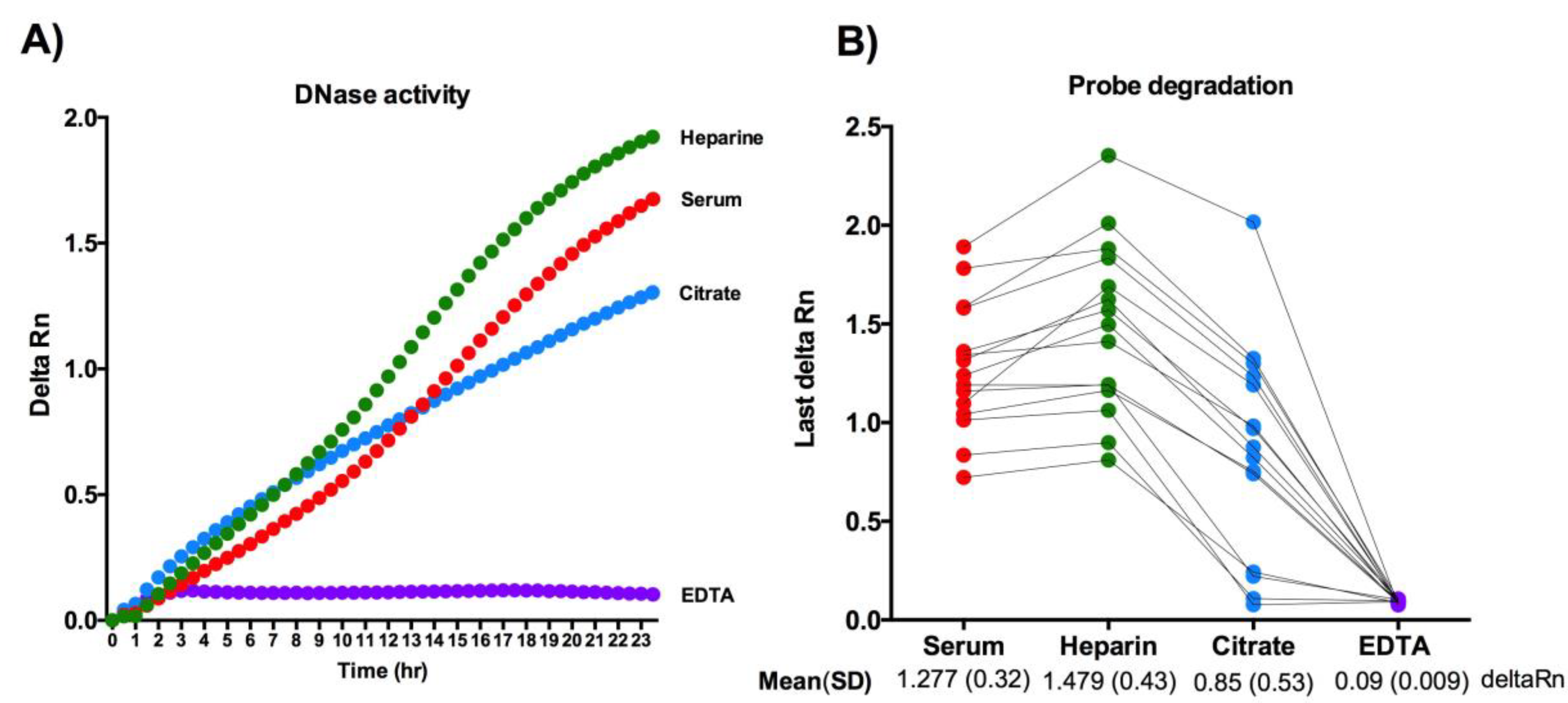
Figure 3.
Changes in cfDNA levels (in genomic equivalents per mL, GE/mL) after 24 hours of incubation at 37°C across different sample types: A) Serum cfDNA levels decreased significantly by 55.6%. B) Heparin-plasma showed a significant reduction of 85.3%. C) EDTA-plasma exhibited an 8.0% reduction, with no significant difference. D) Citrate-plasma levels decreased by 13.3%, with no significant difference. In conclusion, heparin-plasma and serum samples showed the most substantial cfDNA degradation, likely due to active blood DNase activity, whereas EDTA-plasma and citrate-plasma demonstrated greater cfDNA stability, indicating inhibited blood DNase activity.
Figure 3.
Changes in cfDNA levels (in genomic equivalents per mL, GE/mL) after 24 hours of incubation at 37°C across different sample types: A) Serum cfDNA levels decreased significantly by 55.6%. B) Heparin-plasma showed a significant reduction of 85.3%. C) EDTA-plasma exhibited an 8.0% reduction, with no significant difference. D) Citrate-plasma levels decreased by 13.3%, with no significant difference. In conclusion, heparin-plasma and serum samples showed the most substantial cfDNA degradation, likely due to active blood DNase activity, whereas EDTA-plasma and citrate-plasma demonstrated greater cfDNA stability, indicating inhibited blood DNase activity.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



