
Submitted:
09 September 2024
Posted:
10 September 2024
You are already at the latest version
Abstract
Background. The multisystemic features of Down syndrome (DS) in children are accompanied by an immunodeficiency, making them susceptible to infections and immune dysregulation with autoimmune, allergic, inflammatory, and hematological complications. This study was aimed at a better understanding of the abnormalities within the B and T cell compartments and their correlations with clinical immunophenotypes. Methods. Medical records of 35 DS children were retrospectively reviewed, referring to clinical symptomatology including history of infections, immune dysregulation disorders, and humoral and cellular immune response. Results. While the etiology of respiratory tract infections included typical viral and bacterial pathogens, SARS-CoV2-induced inflammatory disease and syndromic immunodeficiency contributed significantly to deterioration of the clinical course. Allergic diseases in the form of asthma, allergic rhinitis, and alimentary allergy were the most frequent manifestations of immune dysregulation and were followed by autoimmune disorders, such as Crohn disease, celiac disease, autoimmune thyroiditis, and alopecia, as well as inflammatory disorders, balanitis xerotica obliterans and lymphadenopathy, and a hematological disorder of myelopoiesis. Deficiency of serum immunoglobulin levels, reduced numbers of naïve B cells, and non-switched memory B cells along with low naïve T helper cells and significantly reduced regulatory T helper cells were the most prominent immune abnormalities. Conclusions. The loss of naïvete in B and T lymphocyte compartments with a deficiency of regulatory T cells may be underpinning pathomechanisms for the skewed immune response. The clinical immunophenotype in DS is complex and represents syndromic inborn error of immunity with immune dysregulation.
Keywords:
down syndrome
; immunodeficiency
; immune dysregulation
; inborn error of immunity
; regulatory T cells
Introduction
Trisomy 21 (OMIM 190685), aka Down syndrome (DS), is a multisystemic condition characterized by a significant clinical, epidemiological, and societal burden. With its prevalence ranging from 5,7 through 8,27 to 12,9 live births per 10000 due to differences in geographic distributions, maternal age, prenatal testing trends, and Down syndrome-related elective terminations associated with the global tendency to reduce the overall population size [1,2,3], the syndrome remains one of the commonest genetic disorders in humans. The genetic complexity in DS is caused either by the presence of an extra copy of chromosome 21 resulting in trisomy or, in a small percentage of cases, Robertsonian translocation and attachment of the long arm of chromosome 21 to another chromosome, isochromosome or ring chromosome [4,5]. DS is characterized by a wide range of clinical phenotypic features, such as intellectual disability, craniofacial dysmorphism, congenital heart disease, most frequently atrioventricular septal defect (AVSD), congenital gastrointestinal abnormalities including Hirschprung disease, duodenal atresia or imperforated anus, decreased muscle tone, increased risk of neurological complications, predominantly early development of Alzheimer dementia, and epilepsy, endocrinopathies, such as short stature and hypothyroidism [4]. Anatomical developmental abnormalities and organ dysfunctions with airway anomalies, structural lung disease, pulmonary hypertension, obstructive sleep apnea, and aspirations are overlapping disorders and show a mutual relationship with an increased predisposition to respiratory infections in affected individuals [6,7,8]. Furthermore, innate and adaptive immune response alterations, and immune-mediated autoimmune, autoinflammatory, myelodysplastic, and lymphoproliferative disorders contribute to the syndromic immunodeficiency in DS [9,10]. It has been hypothesized that this complex phenotype is determined by a subset of dosage-sensitive genes driving alterations in diverse cellular processes. This Down Syndrome Critical Region (DSCR) is mapped to the long arm of chromosome 21 and covers 5,4 Mb containing genes regulating cellular processes, such as splicing (RBM1 (RNA-Binding Motif 1), U2AF1 (U2 Small Nuclear RNA Auxiliary Factor 1), U2AF1L5 (U2 Small Nuclear RNA Auxiliary Factor 1 Like 5)), DNA methylation (PRMT2 (Protein Arginine Methyltransferase 2), N6AMT1 (N(6)-Adenine Specific DNA Methyltransferase)), and metabolism (SOD1 (Superoxide Dismutase 1)). Alternatively, another hypothesis argues against the role of the critical dosage-sensitive region and suggests the multiple genetic abnormalities responsible for the clinical phenotype in DS [11]. Complementing this view, it has been proposed that dysregulated cellular homeostasis caused by additional genetic material leads to a constitutive upregulation of genes triggering aneuploidy-associated stress and resulting in genome-wide changes in gene expression and alterations in cellular pathways. Consequently, impaired mitochondrial function, altered metabolism, epigenetic deregulation, endocytic defects, altered cell specifications, increased levels of oxidative stress, transformed proteostasis and autophagy, elevated DNA damage, and activation of the immune response play a crucial role in the pathophysiology of DS [11,12,13]. It may, therefore, be assumed that heterogeneity of deviated cellular immunoregulatory pathways on genomic, proteomic, and metabolomic levels may underpin syndromic immunodeficiency and immune dysregulation with autoimmune, inflammatory, allergic, lymphoproliferative, and myelodysplastic disorders [14,15,16]. Whereas the genetic and pathophysiological mechanisms determining the immunological defects are complex and multidirectional, it has been postulated, that overexpression of genes such as RCAN1 (Regulator of Calcineurin 1) impairs the inhibitory effect on signal transduction mediated by the nuclear factor of activated T cells (NFAT) and has the potential to reduce the inflammatory response by stabilizing the activity of nuclear factor kappa B (NF-κB) [17].
Aim of the Study
The study was aimed at a better understanding of the abnormalities within the B and T lymph cell compartments in children with DS. We also sought to determine correlations between deficiencies in specific B and T lymphocyte subsets and the DS patients’ clinical and immunological phenotypes.
Materials and Methods
The Study Group
We retrospectively reviewed the medical records of a cohort of pediatric patients in whom a definitive clinical and genetic diagnosis of trisomy 21 had been established. All the children studied were referred to our Department of Pediatric Pneumonology, Allergy, and Clinical Immunology due to suspicion of inborn errors of immunity (IEI) for an in-depth evaluation of clinical phenotypes and immune parameters, from January 2022 by December 2023. The study group consisted of 35 children, 25 boys (71%) and 10 girls (29%), aged from 2 to 204 months (17 years) (mean age 45 months). A discerning retrospective analysis of medical conditions, including history of thymectomy, upper and lower respiratory tract disorders, infectious episodes, congenital heart defects, endocrine disorders, and other malformations and organ dysfunctions that may have an impact on infectious complications has been performed in all the study participants. In all the children studied, airway aspirates for RT-PCR tests for RNA or DNA of 25 respiratory viruses and also for bacterial cultures. Immunological characteristics of the participating children included immunoglobulin levels and flow cytometric analysis of the B and T cell compartments. Antibody levels and relative and absolute counts of lymphocyte subpopulations were interpreted using age-matched reference values.
Flow Cytometric Peripheral Blood (PB) Lymphocyte Immunophenotyping
Cells were labeled with the following murine fluorochrome-stained monoclonal antibodies: anti-CD45 FITC (fluorescein isothiocyanate), anti-CD14 PE (phycoerythrin), anti-CD19 PE, anti-CD19 PerCP (peridinin chlorophyll protein), anti-IgM FITC, anti-IgD FITC, anti-CD38 APC (allophycocyanin), anti-CD27 PE, anti-CD21 FITC, as well as anti-CD3 FITC, anti-CD4 FITC, CD45RA FITC, CD127 FITC, CD185 FITC, anti-CD8 PE, anti-CD16+CD56 PE, CD25 PE, CD31 PE, CD45RO PE, anti-CD3 PerCP, CD197 PerCP, anti-CD4 APC, and anti-CD8 APC (all Beckton-Dickinson Biosciences, USA). The acquisition of cells and analysis were carried out with the use of the flow cytometer FACSCanto and FACSDiva software (Beckton-Dickinson, USA). With sequential gating on biparametric scattering CD45+CD14- lymphocytes, the following lymphocyte subpopulations were identified:
- -
- CD19+ B cells, immature CD19+CD21lo, immature activated CD19+CD38loCD21lo, transitional CD19+CD38hisIgMhi, naïve CD19+CD27-sIgD+, non-switched memory CD19+CD27+sIgD+, switched memory CD19+CD27+IgD- B cells, and CD19+CD38hisIgM- plasmablasts
- -
- CD3+ T cells, CD3+CD4+ T helper cells, CD3+CD4+CD31+CD45RA+ recent thymic emigrants, naïve CD3+CD4+CD27+CD45RA+, regulatory CD3+CD4+CD25++CD27-, central memory CD3+CD4+CD27+CD45RO+, effector memory CD3+CD4+CD27-CD45RO+, terminally differentiated CD3+CD4+CD27-CD45RA+, follicular CD3+CD4+CD185+CD45RO+, and regulatory CD3+CD4+CD45RO+CD127-CD25++ T helper cells.
- -
- among CD3+CD8+ cytotoxic T cells, the following subsets were distinguished: naïve CD3+CD8+CD197+CD27+CD45RA+, central memory CD3+CD8+CD197+CD27+CD45RO+, effector memory CD3+CD8+CD197-CD27-CD45RO+, and terminally differentiated CD3+CD8+CD197-CD27-CD45RA+ cells.
The relative values of PB B and T lymphocytes of the total lymphocyte population as well as B and T cell subsets were calculated. The absolute counts of all cell subsets were calculated from the PB leukocyte counts. A comparative analysis was done with the reference cut-off values of B and T cell subsets for pediatric populations of different age groups [18,19].
Results
Clinical symptomatology. Congenital cardiac malformations occurred in 25 DS-affected children and were the most frequent developmental abnormalities among patients in the study group. They were represented by atrioventricular septal defect (AVSD) in 10, atrial septal defect type II (ASD II) in 8, ventricular septal defect (VSD) in 6 out of 35 reported children, and ASD I, coarctation of aorta (CoA), Fallot tetralogy (ToF) and total anomalous pulmonary vein return (TAPVR), each occurring in 1 patient in the study group. As many as 13 children were thymectomized during corrective cardiac surgery.
Intestinal aganglionosis aka Hirschprung disease was found in three of the patients, and an intrapulmonary sequestration was found in one. Both these congenital malformations and cardiovascular abnormalities contribute to increasing the risk of infections and affect developmental milestones together with immune response.
Infections.The predominating symptomatology indicative of immunodeficiency in the study group were recurrent episodes of respiratory tract infections, most frequently pneumonia, bronchitis, and bronchiolitis in children below the age of two years old. Airway aspirates were taken in all the children studied and the viral DNA or RNA of 9 respiratory viruses were identified by RT-PCR in as many as 17 children (49%). Most frequently, RSV and Rhinovirus-RNA were identified in airway aspirates, both in 11 children studied. Whereas in five out of 35 children, respiratory viruses were not found in the airway aspirates, in two of them, aged 14 and 17 years old (170 and 204 months, respectively), Epstein-Barr virus (EBV)-DNA was found. In twelve out of 35 children, a SARS-CoV2 infection was diagnosed on RT-PCR examination, rendering this infection the most frequent viral etiology overall. All the children with COVID-19 presented with fever, severe respiratory disease, and deterioration of their cardiovascular disorders. Other coronaviruses of non-SARS serotypes, such as OC43 and HKU1, were identified in the children studied as a co-infection with other respiratory viruses.
Airway aspirates were also taken for cultures that were positive in as many as 15 children, in many of them as coinfections. The big three bacteria were Streptococcus pneumoniae, Haemophilus influenzae, and Staphylococcus aureus. The type and frequency of respiratory viruses and the bacterial etiology of respiratory infections in the study group is displayed in Table 1. Beyond respiratory tract infections, two children suffered from recurrent abscesses – perianal abscesses occurred in one patient, and hydradenitis suppurativa in another one.
Immune dysregulation was observed in the study group in the form of allergic, autoimmune, inflammatory, and hematopoietic disorders. The frequency of immune dysregulation disorders and organ-specific immunopathology are summarized in Table 2.
Immunodiagnostics. Antibody deficiency was the most commonly identified immunological abnormality among the reported DS patients and was observable beyond the age of 48 months when transient hypogammaglobulinemia can not be definitely ruled out. Of note, the predominant antibody production defect was IgM deficiency, and IgM serum levels were below the age-matched reference values in as many as 23 children (66%) whereas 12 (34%) children had normal or borderline IgM levels. IgA deficiency was the second most frequent antibody deficit, occurring in 13 of 35 children (37%). Low IgG levels were identified in five children (14%) only and all of them were below the age of 48 months, whereas no patients beyond the age of four years old had IgG hypogammaglobulinemia. The distribution of serum immunoglobulin levels related to the patients’ age is displayed in Figure 1.
Within the B cell compartment, flow cytometric immunophenotyping showed a low absolute count of the total B cell pool which occurred in 14 (40%) out of 35 whereas the relative B cell count was low in 12 (34%) children with Down syndrome studied. The most prominent developmental abnormalities regarding B cell subsets reported in the study group included low numbers of naïve B cells in 13 (37%) children, low memory B cell counts, among them low non-switched memory/ marginal zone B cells in 15 (43%), and low switched memory B cells in 13 (37%) children, and finally, plasmablasts were low in 13 (37%) children. Relative numbers and absolute counts of B cell subsets analyzed in the children studied are shown in Figure 2.
In Figure 3, the distribution of the total T CD3+ pool and T CD4+ helper cell subsets is displayed. Whereas the number of T CD3+ lymph cells was low in 12 children, the absolute counts of T CD4+ helper cells were below reference values in 14 children (40%). For recent thymic emigrants, both their relative counts and absolute numbers were low in 8 (23%) children. A substantial reduction in the production of naïve CD4+CD45RA+CD27+ T helper cells was seen in a significant number of the children studied, with low relative values in 18 (51%) and low absolute counts in 17 (49%) of them.
Meaningful defects were also assessed in the production of regulatory CD4+CD25++CD127- T helper cells, their low relative count occurred in 18 (51%) children and the low absolute number was observable in as many as 25 (71%) patients in the study group. The regression curve displayed in Figure 3 shows the naïve CD4+CD45RA+ to memory CD4+CD45RO+ T helper cell ratio. Of note, as many as 18 children were found to have their results under the curve. In 8 children, the naïve to memory cell ratio was lower than 1, indicating a loss of T helper cell naïvete.
As opposed to T helper cell deficiencies shown in significant proportions of children with DS, abnormal distribution of T CD8+ cytotoxic / suppressor pool was observable in a few children. The total CD8+ T cell absolute numbers were low in 3, and the relative counts in 2 children. Interestingly, relative counts of two different T CD8+ cell subsets, namely terminally differentiated and effector memory T CD8+ cells, were low in 19 (54%) and in 24 (69%) children, respectively. The results of T CD8+ cell immunophenotyping are displayed in Figure 4.
Discussion
The results of our study in a pediatric group of patients with DS show a multiplicity of immunological disorders, such as severe, recurrent viral and bacterial respiratory tract infections, autoimmune manifestations, and immunodeficiency with impaired antibody biosynthesis and disturbances within B and T cell compartments. Down syndrome children present as a syndromic immunodeficiency with anatomical developmental defects and dysfunctions affecting many organs and systems including the cardiovascular, respiratory, endocrine, gastrointestinal, and neuromuscular disorders significantly overlapping with immunodeficiency, autoimmune, lymphoproliferative, and inflammatory manifestations.
Our study shows that low serum levels of IgM antibodies are the most prominent feature of humoral immunodeficiency in DS, observable in 66% of the children reported. IgM deficiency was accompanied by IgA hypogammaglobulinemia in 37% of the children studied. These results are consistent with other studies, in which low levels of IgM and IgA antibodies were demonstrated, making affected children susceptible to infections [14,20,21] and predisposing them to developing immune dysregulation disorders [17].
B cell lymphopenia was assessed in as many as 15 out of 35 children with DS. The reduced total B cell pool in patients with DS may be attributed to a decreased bone marrow development, output, poor peripheral survival, and disturbed homeostasis. In contrast to other inborn errors of immunity, there is no compensatory homeostatic B cell proliferation as a response to a reduced B cell pool [22]. The results of our study showing abnormal B cell differentiation and maturation at different stages of their development are consistent with other studies [23,24]. Whereas the transitional B cell pool, which reflects the development of B cell precursors and the output from the bone marrow was not affected in our study, naïve mature B cells were significantly decreased. Since maturation of the naïve B cell pool occurs at the periphery and is an antigen and T-cell independent process, defective generation of B cells at this stage suggests an intrinsic B cell developmental defect. Referring to the low serum IgM levels in the children studied, it is worth noting that the IgM deficiency correlates with a reduced non-switched memory B cell pool, which is a germinal center-independent, predominantly IgM-producing subpopulation. Importantly, non-switched memory B cells are essential in early primary immune response, providing protection against infections caused by encapsulated bacteria and viruses, as well as in the response to vaccine antigens [25]. Although in our study switched memory B cell pool was not strikingly reduced and sustained normal in 23 out of 35 (66%) children in the study group, alterations in switched memory B cell generation were observed as a cardinal feature of impaired adaptive immunity in children with DS [22,24,26]. The development of this long-living, highly differentiated B cell subset requires T cell-dependent processes of immunoglobulin isotype switching, antigen affinity maturation, and somatic hypermutations in germinal centers. Therefore, deficiency of the switched memory B cells in DS has been suggested to be reminiscent of common variable immunodeficiency (CVID) [23,27,28].
In our study, significant developmental disorders of T CD4+ helper cells were observable. These alterations included a reduction of the total T CD4+ helper cell pool in 40% of the children studied and low naïve CD4+CD45RA+CD27+ T helper cell relative numbers and absolute counts in 51% and 46% of DS children studied. In as many as 23% of children participating in our study, the CD45RA: CD45RO ratio was lower than 1, reflecting the loss of naïvete and decline in the size of newly generated naïve T CD4+ helper cells which may implicate their functional consequences of immunosenescence observable in DS [29,30]. It has been proposed that induction of cellular senescence may contribute to early thymic involution and immune dysregulation. This hypothesis has been supported by the analysis of senescence and cellular damage markers in thymocytes and peripheral T cells, such as increased beta-galactosidase activity, increased levels of the cell cycle inhibitor p16, telomere length, and increased levels of reactive oxygen species reflecting increased oxidative stress [30]. The further considered pathophysiological mechanism of T cell dysfunction in DS is T cell exhaustion, a state of altered immunological profile induced by chronic inflammation due to chronic infection or malignancy related to inflammatory cytokine signaling cascades [31]. In DS, the immune phenotype with increased expression of the programmed death receptor, PD-1, immune checkpoint protein CD160, and immunoregulatory receptor SLAM (signaling lymphocyte activation molecule) or CD244, T cell inhibitory molecules, has been associated with T cell exhaustion [31,32]. Noteworthy, the evidence for T cell exhaustion has also been found in other inborn errors of immunity (IEI) characterized by impaired T cell functions, such as proliferation and cytokine expression, and increased susceptibility to apoptosis, in which genetic underpinnings predispose to immune dysregulation, such as variants in LRBA (Lipopolysaccharide (LPS) Responsive Beige-Like Anchor Protein), CTLA-4 (Cytotoxic T Cell-Associated Protein 4), PI3KR1 (Phosphoinositide-3-Kinase Regulatory Subunit 1), and PIK3CD (Phosphatidylinosito-4,5-Biphosphate 3-Kinase Catalytic Subunit Delta) [33].
It may, therefore, be assumed, that the above-mentioned alterations within T and B cell compartments, including features of lymphocyte senescence and exhaustion, contribute significantly to an increased susceptibility to infections, and predisposition to autoimmune, autoinflammatory, and malignant disorders associated with DS patients in our study group. Another aspect of symptomatology related to immune dysregulation in DS is an aberrant inflammatory cytokine profile with elevated serum levels of acute phase markers, such as interleukin (IL)-1β, IL-6, tumor necrosis factor (TNF)-α, and interferon (IFN)-γ [34,35,36]. The IFN hyperactivity has been hypothesized to be a background for an autoimmunity-prone state, phenotypically presented in our patients with autoimmune thyroiditis, alopecia, celiac, and Crohn disease [37,38]. Interferonopathy in Down syndrome children was also reported to be associated with the development of acute immune dysregulation, life-threatening clinical condition, hemophagocytic lymphohistiocytosis treated with favorable outcomes targeting interferon pathway with emapalumab [39].
Referring to the numerousness and multiplicity of respiratory tract viral infections in the children studied, the results of viral studies clearly pointed to a high frequency of respiratory viruses as etiological factors of infectious airway episodes in DS children. The spectrum of etiologies included the big three of respiratory viruses, with respiratory syncytial virus (RSV), rhinovirus and, surprisingly, bocavirus. Of note, the most frequent infection among children studied was COVID-19 and it had the most devastating effect on the respiratory and cardiovascular condition, with systemic inflammatory response syndrome (SIRS) [40,41,42]. Children with Down syndrome are particularly vulnerable to a severe course of COVID-19 due to syndromic immunodeficiency associated with immune dysregulation [43], yet in immunocompromised children the susceptibility to pneumonia due to other coronaviruses, such as OC43 and HKU1 is also increased [44,45].
In adult DS patients, an epidemiological study [46] has demonstrated a low risk of viral infections, while in children with DS, respiratory viruses are the leading etiological factors of airway infections, contributing to a high rate of hospitalizations, respiratory failure, and fatalities, as well as having a strong impact on neurodevelopmental scores [47]. Beyond defects in immunity, meaningful contributory factors make DS children vulnerable to frequent and severe infectious respiratory episodes, such as neuromuscular disorders, psychomotor delay, gastroesophageal reflux, upper airway abnormalities with macroglossia and dysfunctional swallowing, structural lung disease, congenital heart disease with altered pulmonary blood flow [6,7,8,47,48]. While increased expression of genes for alpha/beta or type I interferon type I (IFN-I) receptors, IFNAR1 and IFNAR2, and enhanced signaling of type I interferon pathways might be assumed to serve as protective factors due to their crucial role in controlling viral infections, defective regulation of IFN-I mechanisms may be an additional explanation for impaired response to respiratory viruses in children with DS [49,50].
T helper 2 (Th2)-skewed immune response with increased levels of IL-4 and IL-13, cytokines that regulate various aspects of allergic inflammation [51], which was observed in a proportion of participants in our study. This may be another pathogenetic factor contributing to the susceptibility to infections in children with DS.
Predisposition to abnormal hematopoiesis in children with DS is perceived as another manifestation of the immune dysregulation phenotype, resulting from mutual interactions between epigenetic, oncogenic, chromosomal, and cellular homeostasis mechanisms [52]. Children with DS have a significantly increased risk of both myeloid and lymphoblastic leukemia compared to the general pediatric population [53,54]. A unique neonatal myeloproliferative disorder, observable in as many as 4 patients in our study group, transient abnormal myelopoiesis (TAM), manifests with circulating blasts having morphological and immunological features of the megakaryocytic lineage, posing a risk of developing myeloid leukemia associated with Down syndrome (ML-DS) [55,56]. The preleukemic, myelodysplastic phase of TAM in blasts bearing trisomy 21 is driven by variants in the key hematopoietic transcription factor GATA1 gene. While in the majority of infants with DS, TAM resolves spontaneously, 5 to 10% of them are found to have variants in cohesin, protein CCCTC-binding factor CTCF gene, oncogenes belonging to the RAS family, genes of the Janus Kinase and Signal Transducer and Activator of Transcription JAK/STAT4 pathway that induce progressing to ML-DS [53,54,55,56]. Consequently, in DS., an integrated multidisciplinary approach and care as well as the development of further comprehensive research strategies with special emphasis on immunodeficiency and immune dysregulation are required [57,58].
Conclusions
In the last decades, significant progress has been made in the field of immunodiagnostic cell phenotyping and immunogenetics contributing to a better understanding of the immunodeficiency and immune dysregulation phenomena in children with DS. Their immunophenotypes show multiple and complex lymphoproliferative and myeloproliferative alterations due to chromosomal aneuploidy, single gene variants, epigenetic dysregulation, and molecular dysfunctions. Consequently, immunodeficiency with infectious complications and immune dysregulation in the form of autoimmune, allergic, inflammatory, and hematopoietic disorders are observable in pediatric DS patients. IgM hypogammaglobulinemia with B and T cell lymphopenia and reduced numbers of naïve B and T helper cells in children with DS along with low numbers of cytotoxic effector memory T cells are crucial immunological abnormalities predisposing to infectious diseases. In this study, we have also demonstrated that in addition to the loss of naïvete of T helper cells, low numbers of regulatory T cells may contribute to immune dysregulation disorders in children with DS. Non-immunological phenotyping features, being integral parts of DS and having a significant impact on immune deficiency and immune dysregulation disorders, place this multisystemic condition among syndromic inborn errors of immunity.
Author Contributions
Aleksandra Szczawińska-Popłonyk was responsible for the principal design of the review and its intellectual content, coordinated and supervised data collection, drafted, and critically revised the final manuscript. The first draft of the manuscript was written by Natalia Popłonyk and Karina Awdi. These two authors equally contributed to this work. All authors read and approved the final manuscript.
Funding support
No funds, grants, or other funding support were received during preparation of this manuscript
Ethics approval
NA. This research was conducted retrospectively from data obtained for clinical purposes. Ethical approval was waived by the Bioethical Committee of Poznań University of Medical Sciences. In view of the retrospective nature of the study, and all the procedures being performed were part of the routine care and did not need ethical approval
Informed consent to participate/ publication:
NA.
Availability of data and material/data transparency
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request
Conflict of interest/Competing interest
All authors declare no existing financial and non-financial conflict of interest regarding this manuscript
Abbreviations
| APC | Allophycocyanin |
| AVSD | Atrioventricular septal defect |
| CoA | Coarctation of aorta |
| CVID | Common variable immunodeficiency |
| CTCF | CCCT-Binding Factor |
| CTLA-4 | Cytotoxic T Cell-Associated Protein 4 |
| DS | Down Syndrome |
| DSCR | Down Syndrome Critical Region |
| EBV | Epstein-Barr virus |
| FITC | Fluorescein isothiocyanate |
| GATA1 | Globin Transcription Factor 1 |
| IEI | Inborn errors of immunity |
| IFN | Interferon |
| IFNAR | Interferon alpha/beta receptor |
| JAK | Janus Kinase |
| LRBA | Lipopolysaccharide Responsive Beige-Like Anchor Protein |
| ML-DS | Myeloid leukemia associated with Down syndrome |
| NFAT | Nuclear factor of activated T cells |
| NF-κB | Nuclear factor kappa B |
| N6AMT1 | N(6)-Adenine Specific DNA Methyltransferase |
| PD-1 | Programmed death receptor |
| PE | Phycoerythrin |
| PerCP | Peridinin chlorophyll protein |
| PIK3CD | Phosphatidylinositol-4,5-Biphosphate 3-Kinase Catalytic Subunit Delta |
| PIK3R1 | Phosphoinositide-3-Kinase Regulatory Subunit 1 |
| PRMT2 | Protein Arginine Methyltransferase 2 |
| RBM1 | RNA-Binding Motif 1 (human spermatogenesis candidate gene) |
| RCAN1 | Regulator of Calcineurin 1 |
| SLAM | Signaling lymphocyte activation molecule |
| SOD1 | Superoxide Dismutase 1 |
| STAT4 | Signal Transducer and Activator of Transcription 4 |
| TAM | Transient abnormal myelopoiesis |
| TAPVR | Total anomalous pulmonary vein return |
| TNF | Tumor necrosis factor |
| ToF | Tetralogy of Fallot |
| U2AF1 | U2 Small Nuclear RNA Auxiliary Factor 1 |
| U2AF1L5 | Small Nuclear RNA Auxiliary Factor 1 Like 5 |
| VSD | Ventricular septal defect |
References
- De Graaf, G.; Buckley, F.; Skotko, B.G. Estimation of the number of people with Down syndrome in Europe. Eur J Human Genet 2021, 29, 402–410. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, G.; Buckley, F.; Skotko, B.G. Estimates of live births, natural losses, and elective terminations with Down syndrome in the United States. Am J Med Genet 2015, 167A, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Presson, A.P.; Partyka, G.; Jensen, K.M.; Devine, O.J.; Rasmussen, S.A.; McCabe, L.L.; et al. Current estimates of Down syndrome population prevalence in the United States. J Pediatr 2013, 163, 1163–1168. [Google Scholar] [CrossRef] [PubMed]
- Asim, A.; Kumar, A.; Muthuswamy, S.; Jain, S.; Agarwal, S. Down syndrome: an insight of the disease. J Biomed Sci 2015, 22, 41. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, A.; Raja, K.; Venugopalan, M.; Chandrasekaran, B.; Kovanur Sampath, K.; Muthusamy, H.; et al. Down syndrome-a narrative review with a focus on anatomical features. Clin Anat 2016, 29, 29,568–577. [Google Scholar] [CrossRef]
- De Lausnay, M.; Ides, K.; Wojciechowski, M.; Boudewyns, A.; Verhulst, S.; Van Hoorenbeek, K. Pulmonary complications in children with Down syndrome: a scoping review. Paediatr Resp Rev 2021, 40, 65–72. [Google Scholar] [CrossRef]
- Alsubie, H.S.; Rosen, D. The evaluation and management of respiratory disease in children with Down syndrome (DS). Paediatr Resp Rev 2018, 26, 49–54. [Google Scholar] [CrossRef]
- Danopoulos, S.; Deutsch, G.H.; Dumortier, C.; Mariani, T.J.; Al Alam, D. Lung disease manifestations in Down syndrome. Am J Physiol Lung Cell Mol Physiol 2021, 321, 892–899. [Google Scholar] [CrossRef]
- Verstegen, R.H.J.; Chang, K.J.J.; Kusters, M.A.A. Clinical implications of immune-mediated diseases in children with Down syndrome. Pediatr Allergy Immunol 2020, 31, 117–123. [Google Scholar] [CrossRef]
- Szczawinska-Poplonyk, A.; Begier, K.; Dorota, A.; Dabrowska, M.; Galecka, D.; Wawrzeniak, K.; Wroblewski, K. Syndromic immunodeficiencies: a pediatrician’s perspective on selected diseases. Allergol Immunopathol 2021, 49, 117–136. [Google Scholar] [CrossRef]
- Krivega, M.; Stiefel, C.M.; Storchova, Z. Consequences of the chromosome gain: a new view on trisomy syndromes. Am J Human Genet 2022, 109, 2126–2140. [Google Scholar] [CrossRef] [PubMed]
- Chunduri, N.K.; Storchova, Z. The diverse consequences of aneuploidy. Nat Cell Biol 2019, 21, 2010–2021. [Google Scholar] [CrossRef] [PubMed]
- Antonarakis, S.E.; Skotko, B.G.; Rafii, M.S.; Strydom, A.; Pape, S.E.; Bianchi, D.W.; et al. Down syndrome. Nat Rev Dis Primers 2020, 6, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Huggard, D.; Doherty, D.G.; Molloy, E.J. Immune dysregulation in children with Down syndrome. Front Pediatr 2020, 8, 73. [Google Scholar] [CrossRef]
- Ferrari, M.; Stagi, S. Autoimmunity and genetic syndromes: a focus on Down syndrome. Genes 2021, 12, 268. [Google Scholar] [CrossRef]
- Satge D, Seidel MG, The pattern of malignancies in Down syndrome and its potential context with the immune system. Front Immunol 2018, 9, 3058. [CrossRef]
- Ram, G.; Chinen, J. Infections and ammunodeficiency in Down syndrome. Clin Exp Immunol 2011, 164, 9–16. [Google Scholar] [CrossRef]
- Piatosa, B.; Wolska-Kuśnierz, B.; Pac, M.; Siewiera, K.; Gałkowska, E.; Bernatowska, E. B cell subsets in healthy children: reference values for evaluation of B cell maturation process in peripheral blood. Cytometry B Clin Cytom 2010, 78, 372–381. [Google Scholar] [CrossRef]
- Schatorje, E.J.H.; Gemen, E.F.A.; Driessen, G.J.A.; Leuvenink, J.; van Hout, R.W.N.M.; de Vries, E. Paediatric reference values for the peripheral T cell compartment. Scand J Immunol 2012, 75, 436–444. [Google Scholar] [CrossRef]
- Huggard, D.; Worrall, A.P.; Kirkham, C.; McGrane, F.; Mandira, R.; Casey, L.; et al. Immune screening in children with Down syndrome. Acta Paediatr 2022, 111, 2025–2028. [Google Scholar] [CrossRef]
- Eissa, E.; Afifi, H.H.; Abo-Shanab, A.M.; Thomas, M.M.; Taher, M.B.; Kandil, R.; et al. Importance of TREC and KREC as molecular markers for immunological evaluation of Down syndrome children. Sci Rep 2023, 13, 15445. [Google Scholar] [CrossRef] [PubMed]
- Verstegen, R.H.J.; Kusters, M.A.A. Inborn errors of adaptive immunity in Down syndrome. J Clin Immunol 2020, 40, 791–806. [Google Scholar] [CrossRef] [PubMed]
- Verstegen, R.H.J.; Borte, S.; Bok, L.A.; van Zwieten, P.H.T.; von Dobeln, U.; Hammarstrom, L.; et al. Impact of Down syndrome on the performance of neonatal screening assays for severe primary immunodeficiency diseases. J Allergy Clin Immunol 2014, 133, 1208–1211. [Google Scholar] [CrossRef] [PubMed]
- Carsetti, R.; Valentini, D.; Marcellini, V.; Scarsella, M.; Marasco, E.; Giustini, F.; et al. Reduced numbers of switched memory B cells with high terminal differentiation potential in Down syndrome. Eur J Immunol 2015, 45, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Akkaya, M.; Kwa Kk Pierce, S.K. B cell memory: building two walls of protection against pathogens. Nat Rev Immunol 2020, 20, 229–238. [Google Scholar] [CrossRef]
- Verstegen, R.H.J.; Driessen, G.J.; Bartol, S.J.W.; van Noesel, C.J.M.; Boon, L.; van der Burg, M.; et al. Defective B cell memory in patients with Down syndrome. J Allergy Clin Immunol 2014, 134, 1346–1353. [Google Scholar] [CrossRef]
- Verstegen, R.H.J.; Kusters, M.A.A.; Gemen, E.F.A.; de Vries, E. Down syndrome B lymphocyte subpopulations, intrinsic defect or decreased T lymphocyte help. Pediatr Res 2010, 67, 563–569. [Google Scholar] [CrossRef]
- Szczawińska-Popłonyk, A.; Schwartzmann, E.; Bukowska-Olech, E.; Biernat, M.; Gattner, S.; Korobacz, T.; et al. The pediatric common variable immunodeficiency – from genetics to therapy: a review. Eur J Pediatr 2022, 181, 1371–1383. [Google Scholar] [CrossRef]
- Gensous, N.; Bacalini, M.G.; Franceschi, C.; Garagnani, P. Down syndrome, accelerated aging and immunosenescence. Semin Immunopathol 2020, 42, 635–645. [Google Scholar] [CrossRef]
- Marcovecchio, G.E.; Ferrua, F.; Fontana, E.; Beretta, S.; Genua, M.; Bortolomai, I.; et al. Premature senescence and increased oxidative stress in the thymus of Down syndrome patients. Front Immunol 2021, 12, 669893. [Google Scholar] [CrossRef]
- Campos, J.S.; Henrickson, S.E. Defining and targeting patterns of T cell dysfunction in inborn errors of immunity. Front Immunol 2022, 13, 932715. [Google Scholar] [CrossRef] [PubMed]
- Peeters, D.; Pico-Knijnenburg, I.; Wieringa, D.; Rad, M.; Cuperus, R.; Ruige, M.; et al. AKT hyperphosphorylation and T cell exhaustion in Down syndrome. Front Immunol 2022, 13, 724436. [Google Scholar] [CrossRef] [PubMed]
- Delmonte, O.M.; Castagnoli, R.; Calzoni, E.; Notarangelo, L.D. Inborn errors of immunity with immune dysregulation: from bench to bedside. Front Pediatr 2019, 7, 353. [Google Scholar] [CrossRef] [PubMed]
- Huggard, D.; Kelly, L.; Ryan, E.; McGrane, F.; Lagan, N.; Roche, E.; et al. Increased systemic inflammation in children with Down syndrome. Cytokine 2020, 127, 154938. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Che, M.; Yuan, J.; Yu, Y.; Cao, C.; Qin, X.Y.; et al. Aberrations in circulating inflammatory cytokine levels in patients with Down syndrome: a meta-analysis. Oncotarget 2017, 8, 84489–84496. [Google Scholar] [CrossRef]
- Araya, P.; Waugh, K.A.; Sullivan, K.D.; Nunez, N.G.; Roselli, E.; Smith, K.P.; et al. Trisomy 21 dysregulates T cell lineages toward an autoimmunity-prone state associated with interferon hyperactivity. Proc Natl Acad Sci USA 2019, 116, 24231–24241. [Google Scholar] [CrossRef]
- Aversa, T.; Crisafulli, G.; Zirilli, G.; De Luca, F.; Gallizi, R.; Valenzise, R. Epidemiological aspects of autoimmune thyroide disease in children with Down’s syndrome. Ital J Pediatr 2018, 44, 39. [Google Scholar] [CrossRef]
- Guaraldi, F.; Rossetto Giaccherino, R.; Lanfranco, F.; Motta, G.; Gori, D.; Arvat, E.; et al. Endocrine autoimmunity in Down’s syndrome. Front Horm Res 2017, 48, 133–146. [Google Scholar] [CrossRef]
- Guild, A.; Fritch, J.; Patel, S.; Reinhardt, A.; Acquazzino, M. Hemophagocytic lymphohistiocytosis in trisomy 21: successful treatment with interferon inhibition. Pediatr Rheumatol Online J 2022, 20, 104. [Google Scholar] [CrossRef]
- Leung, C.; Su Li Simoes-E-Silva, A.C.; Arocha, L.S.; de Paiva, K.M.; Haas, P. Risk for severe illness and death among pediatric patients with Down syndrome hospitaliza for COVID-19, Brazil. Emerg Infect Dis 2023, 29, 126–35. [Google Scholar] [CrossRef]
- Pitchan Velammal, P.N.K.; Balasubramanian, S.; Ayoobkhan, F.S.; Mohan, G.V.K.; Aggarval, P.; Rabaan, A.A.; et al. COVID-19 in patients with Down syndrome: a systematic review. Immun Inflamm Dis 2024, 12, 1219. [Google Scholar] [CrossRef] [PubMed]
- Kołtan, S.; Ziętkiewicz, M.; Becht, R.; Berdej-Szczot, E.; Cienkusz, M.; Ewertowska, M.; et al. COVID-19 in unvaccinated patients with inborn errors of immunity – Polish experience. Front Immunol 2022, 13, 953700. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, J.M. Down syndrome and COVID-19: a perfect storm? Cell Rep Med 2020, 1, 100019. [Google Scholar] [CrossRef] [PubMed]
- Szczawińska-Popłonyk, A.; Jończyk-Potoczna, K.; Bręborowicz, A.; Bartkowska-Śniatkowska, A.; Figlerowicz, M. Fatal respiratory distress syndrome due to coronavirus infection in a child with severe combined immunodeficiency. Influenza Other Respir Viruses 2013, 7, 634–636. [Google Scholar] [CrossRef] [PubMed]
- Ogimi, C.; Englund, J.A.; Bradford, M.C.; Qin, X.; Boeckh, M.; Waghmare, A. Characteristics and outcomes of coronavirus infection in children: the role of viral factors and an immunocompromised state. J Pediatr Infect Dis Soc 2019, 8, 21–28. [Google Scholar] [CrossRef]
- Fitzpatrick, V.; Rivelli, A.; Chaudhari, S.; Chicoine, L.; Jia, G.; Rzhetsky, A.; et al. Prevalence of infectious diseases among 6078 individuals with Down syndrome in the United States. J Patient Cent Res Rev 2022, 9, 64–69. [Google Scholar] [CrossRef]
- Ghezzi, M.; Garancini, N.; De Santis, R.; Gianolino, L.; Zirpoli, S.; Mandelli, A.; et al. Recurrent respiratory infections in children with Down syndrome: a review. Children 2014, 11, 246. [Google Scholar] [CrossRef]
- Eijsvoogel, N.B.; Verstegen, R.H.J.; van Well, G.T.J.; van Hout, R.W.N.M.; de Vries, E. Increased rate of respiratory symptoms in children with Down syndrome: a 2-year web-based parent-reported prospective study. Eur J Pediatr 2022, 181, 4079–4089. [Google Scholar] [CrossRef]
- Notarangelo, L.D.; Bosticardo, M. Interferons in Down syndrome: when more is less. Immunity 2022, 55, 1967–1969. [Google Scholar] [CrossRef]
- Malle, L.; Bogunovic, D. Down syndrome and type I interferon: not so simple. Curr Opin Immunol 2022, 72, 196–205. [Google Scholar] [CrossRef]
- Malle, L.; Patel, R.S.; Martin-Fernandez, M.; Stewart, O.; Philippot, Q.; Buta, S.; et al. Autoimmunity in Down’s syndrome via cytokines, CD4 T cells and CD11+ B cells. Nature 2023, 615, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Biselli, J.M.; Zampieri, B.L.; Biselli-Chicote, P.M.; de Souza, J.E.S.; Burger, M.C.; da Silva, W.A., Jr.; et al. Differential microRNA expression profile in blood of children with Down syndrome suggests a role in immunological dysfunction. Human Cell 2022, 35, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Baruchel, A.; Bourquin, J.; Crispino, J.; Cuartero, S.; Hasle, H.; Hitzler, J.; et al. Down syndrome and leukemia: from basic mechanisms to clinical advances. Haematologica 2023, 108, 2570–2581. [Google Scholar] [CrossRef] [PubMed]
- Peroni, E.; Gottardi, M.; D’Antona, L.; Randi, M.L.; Rosato, A.; Coltro, G. Hematologic neoplasms associated with Down syndrome: cellular and molecular heterogeneity of the disease. Int J Mol Sci 2023, 24, 15325. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K. Recent advances in the understanding of transient abnormal myelopoiesis in Down syndrome. Pediatrics Int 2019, 61, 222–229. [Google Scholar] [CrossRef]
- Li, J.; Kalev-Zylinska, M.L. Advances in molecular characterization of myeloid proliferations associated with Down syndrome. Front Genet 2022, 13, 891214. [Google Scholar] [CrossRef]
- Hendrix, J.A.; Amon, A.; Abbeduto, L.; Agiovlasitis, S.; Alsaied, T.; Anderson, H.A.; et al. Opportunities, barriers, and recommendations in Down syndrome research. Transl Sci Rare Dis 2021, 5, 99–129. [Google Scholar] [CrossRef]
- Van Gameren-Oosterom, H.B.M.; Weijerman, M.E.; van Wieringen, H.; de Winter, J.P.; van Wermeskerken, A. Clinical practice-latest insights in optimizing the care of children with Down syndrome. Eur J Pediatr 2023, 182, 2027–2039. [Google Scholar] [CrossRef]
Figure 1.
The distribution of serum immunoglobulins G, M, and A in the children studied. The immunoglobulin concentrations expressed in mg/dL in relation to the patients’ age given in months, and to age-matched reference values. Reference minimal and maximal values  , patients’ values
, patients’ values  .
.
, patients’ values .
Figure 1.
The distribution of serum immunoglobulins G, M, and A in the children studied. The immunoglobulin concentrations expressed in mg/dL in relation to the patients’ age given in months, and to age-matched reference values. Reference minimal and maximal values , patients’ values .
, patients’ values .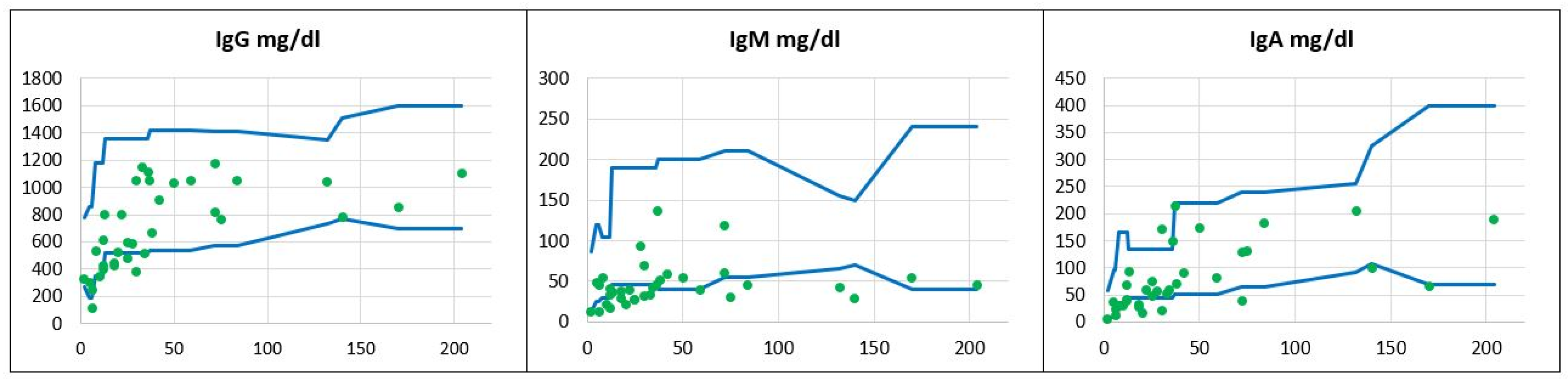
Figure 2.
The relative counts and absolute numbers of B cell subpopulations in a group of children with Down syndrome, in relation to age-matched reference values. Reference minimal and maximal values , patients’ relative counts (%)  , absolute numbers (cc)
, absolute numbers (cc)  .
.
, patients’ relative counts (%) , absolute numbers (cc) .
Figure 2.
The relative counts and absolute numbers of B cell subpopulations in a group of children with Down syndrome, in relation to age-matched reference values. Reference minimal and maximal values , patients’ relative counts (%) , absolute numbers (cc) .
, patients’ relative counts (%) , absolute numbers (cc) .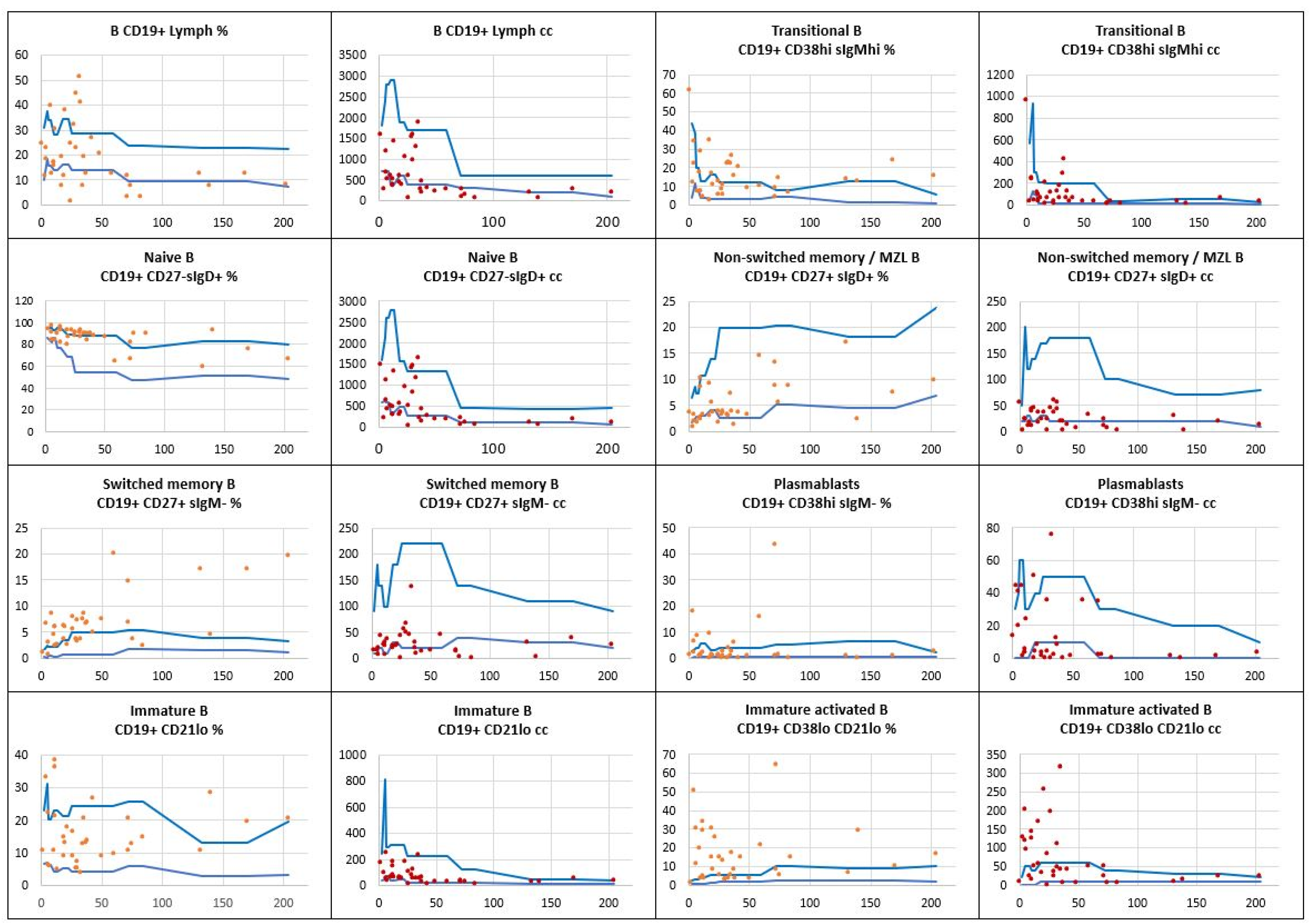
Figure 3.
The relative counts and absolute numbers of T CD4+ helper cell subpopulations in a group of children with Down syndrome, in relation to age-matched values. The regression curve shows naïve CD4+CD45RA+ to memory CD4+CD45RO+ T helper cell ratio. Reference minimal and maximal values , patients’ relative counts (%) , absolute numbers (cc) .
, patients’ relative counts (%) , absolute numbers (cc) .
Figure 3.
The relative counts and absolute numbers of T CD4+ helper cell subpopulations in a group of children with Down syndrome, in relation to age-matched values. The regression curve shows naïve CD4+CD45RA+ to memory CD4+CD45RO+ T helper cell ratio. Reference minimal and maximal values , patients’ relative counts (%) , absolute numbers (cc) .
, patients’ relative counts (%) , absolute numbers (cc) .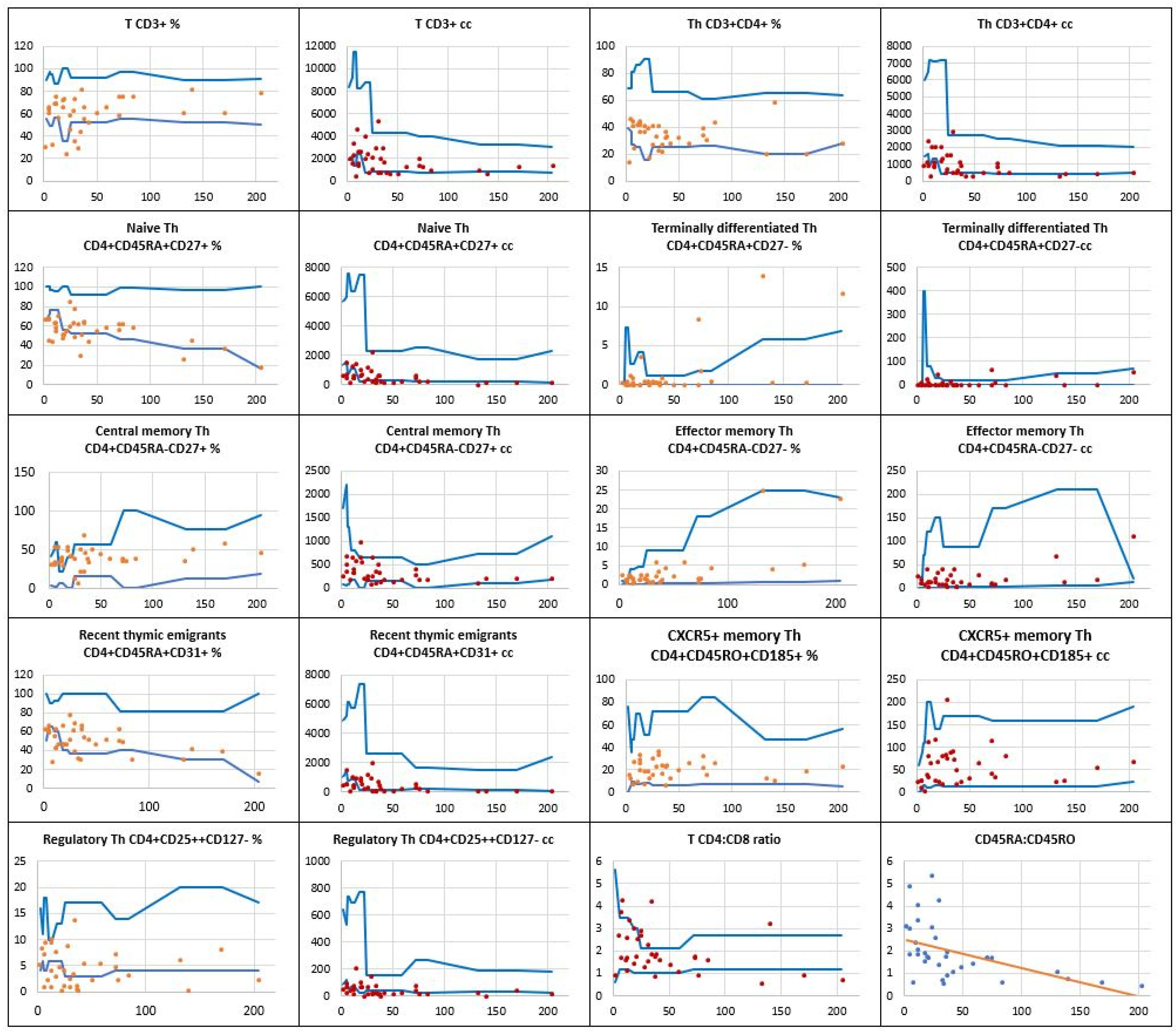
Figure 4.
The relative counts and absolute numbers of T CD8+ cytotoxic / suppressor cell subpopulations in a group of children with Down syndrome, in relation to age-matched values. Reference minimal and maximal values , patients’ relative counts (%) , absolute numbers (cc) .
, patients’ relative counts (%) , absolute numbers (cc) .
Figure 4.
The relative counts and absolute numbers of T CD8+ cytotoxic / suppressor cell subpopulations in a group of children with Down syndrome, in relation to age-matched values. Reference minimal and maximal values , patients’ relative counts (%) , absolute numbers (cc) .
, patients’ relative counts (%) , absolute numbers (cc) .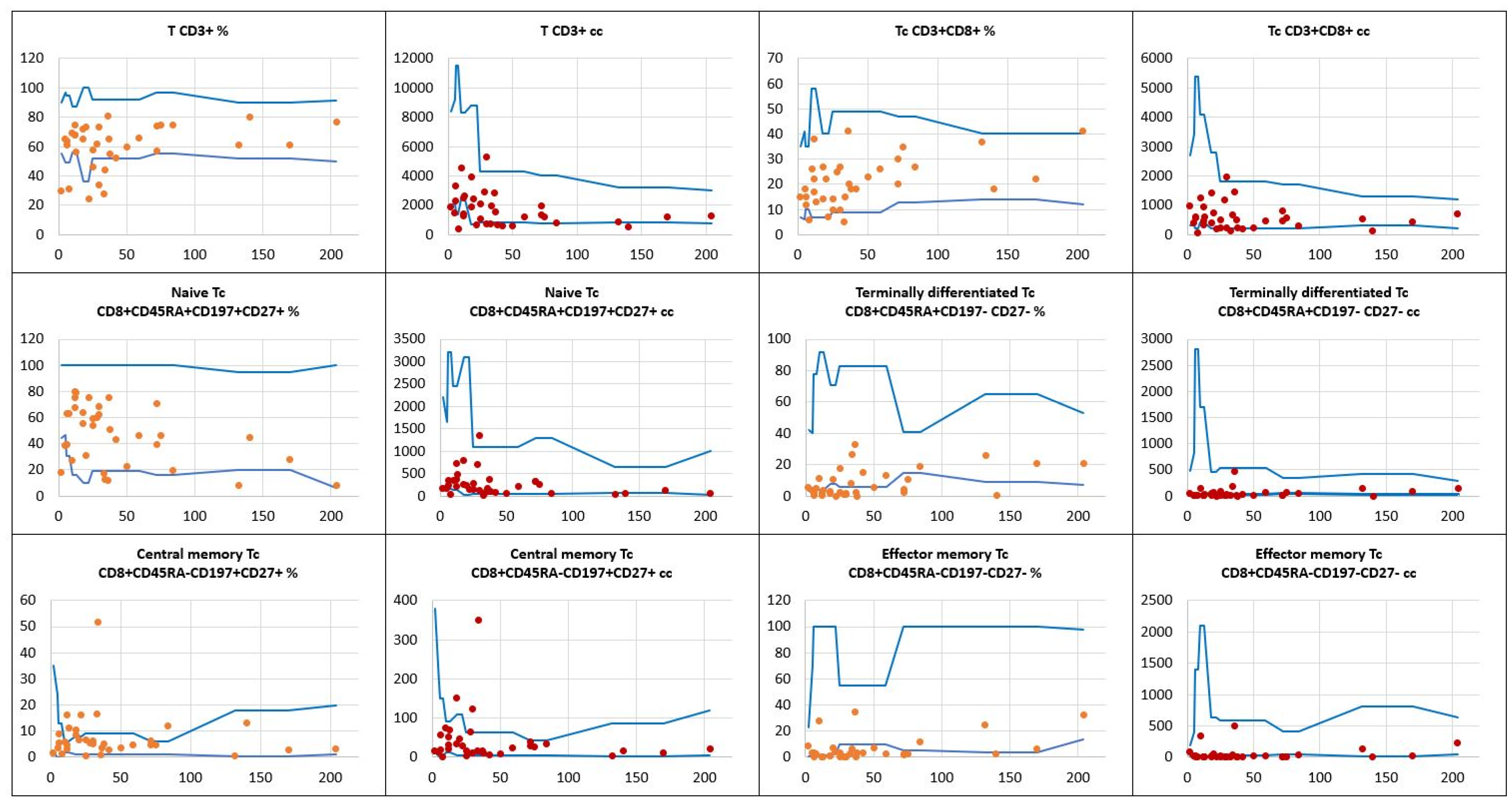

Table 1.
Infectious etiologies of respiratory tract diseases and their frequencies in DS children.
| Infectious etiology | |||
|---|---|---|---|
| Type of infection | Frequency | ||
| Viral |
SARS-CoV2 RSV Rhinovirus Bocavirus Adenovirus Coronavirus OC43 Influenza virus A/B Parainfluenza 3 and 4 Coronavirus HKU1 |
N=12 (34%) N=11 (31%) N=11 (31%) N=6 (17%) N=3 (9%) N=3 (9%) N=2 (6%) N=2 (6%) N=1 (3%) |
Total number of infected children N=17 (49%) |
| Bacterial |
Streptococcus pneumoniae Haemophilus influenzae Staphylococcus aureus Escherichia coli Klebsiella pneumoniae Pseudomonas aeruginosa |
N=12 (34%) N=9 (26%) N=8 (23%) N=6 (18%) N=4 (11%) N=1 (3%) |
Total number of infected children N=15 (43%) |
Table 2.
Frequency of immune dysregulation disorders and organ-specific immunopathology in DS children.
Table 2.
Frequency of immune dysregulation disorders and organ-specific immunopathology in DS children.
| Immune dysregulation disorders | |||
|---|---|---|---|
| Type of disorder | Frequency | ||
| Allergic |
Asthma Food allergy Allergic rhinitis |
N=8 (23%) N=8 (23%) N=1 (3%) |
Total N=17 (49%) |
| Autoimmune |
Thyroiditis Alopecia Celiac disease Crohn disease |
N=1 (3%) N=1 (3%) N=1 (3%) N=1 (3%) |
Total N=4 (11%) |
| Inflammatory |
Balanitis xerotica obliterans Hydradenitis suppurativa Lymphadenopathy |
N=2 (6%) N=1 (3%) N=2 (6%) |
Total N=6 (18%) |
| Hematopoietic | Transient abnormal myelopoiesis | N=4 (11%) | Total N=4 (11%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



