
Submitted:
09 September 2024
Posted:
10 September 2024
You are already at the latest version
Abstract
Plant growth-promoting rhizobacteria (PGPR) are beneficial bacteria that play a crucial role in sustainable agriculture by enhancing plant growth through various mechanisms. This review examines the contributions of PGPR in improving nutrient availability, producing phytohormones, providing biocontrol against pathogens, and enhancing abiotic stress tolerance. By reducing the necessity for chemical fertilizers and pesticides, PGPR mitigate environmental impacts, enhance soil health, and support long-term agricultural productivity. However, challenges such as inconsistent performance across various soils, regulatory barriers, and limited farmer awareness hinder their widespread adoption. Recent advancements in nano-encapsulation technology, genetic engineering, and bioinformatics present promising solutions for overcoming these obstacles and enhancing PGPR efficacy. The incorporation of PGPR into biofertilizers, biopesticides, and integrated pest management (IPM) strategies offers a sustainable resolution to global agricultural challenges. This review addresses the current state of PGPR research, applications, and future directions for optimizing their use in promoting sustainable agriculture.
Keywords:
PGPR
; IPM
; Sustainable agriculture
; Biofertilizer
; Biopesticide
; Plant fitness tetrahedron
1. Introduction
Plant growth-promoting rhizobacteria (PGPR) are beneficial bacteria residing in the rhizosphere, the soil region contacted by plant roots [1]. These bacteria play a vital role in promoting plant growth and health through various mechanisms, making them a key component of sustainable agriculture [2,3]. PGPR can promote plant growth directly by improving nutrient acquisition and producing growth-mediating hormones (phytohormones) or indirectly by protecting plants from pathogens (biotic stresses) and mitigating abiotic stresses (such as drought and salinity) [4,5,6].
Different PGPR have different mechanisms of action. Among them, conserved mechanisms are nutrient solubilization and fixation [7,8], phytohormone production [4,6], biocontrol of pathogens [4,5,6], and abiotic stress tolerance [5]. Specifically, PGPR enhance nutrient availability by solubilizing phosphorus and fixing atmospheric nitrogen, thus reducing the need for chemical fertilizers, which are not sustainable [9,10]. PGPR produce phytohormones such as auxins, gibberellins, and cytokinins, which stimulate root and shoot development, enhancing overall plant growth [5,6,11,12]. PGPR protect plants from pathogens by producing antimicrobial compounds, competing for resources, and inducing systemic resistance in plants, which can reduce the reliance on chemical pesticides [4,6,9]. These bacteria also help plants withstand abiotic stresses like drought and salinity by enhancing stress response mechanisms and detoxifying harmful substances [4,5,9].
The need for sustainable agricultural practices has become increasingly urgent due to the environmental and health concerns associated with conventional farming methods. Traditional agriculture often relies heavily on synthetic fertilizers and pesticides, which can lead to soil degradation, water pollution, and the development of resistant pest strains. These conventional practices contribute to environmental issues such as eutrophication and biodiversity loss, posing significant threats to ecosystems and human health [9,13]. By integrating PGPR into agricultural practices, farmers can achieve higher crop yields and quality while reducing the environmental impact of farming. This approach aligns with the goals of sustainable agriculture, promoting ecological balance and long-term productivity [9,11].
PGPR are increasingly recognized as a sustainable alternative to conventional agricultural practices. Their ability to enhance crop productivity while minimizing environmental impacts makes them a promising solution for future agricultural challenges. As research advances, the development of more effective PGPR strains and application methods is expected to further their roles in sustainable agriculture [14].
The integration of PGPR in agricultural practices offers several benefits: 1) Reduced chemical inputs: By improving nutrient availability and providing natural pest control, PGPR reduce the need for synthetic fertilizers and pesticides, contributing to more sustainable agricultural practices [9,10]. 2) Improved soil health: PGPR enhance soil fertility and structure, supporting long-term agricultural productivity and sustainability [9]. 3) Environmental benefits: The reduction in chemical inputs leads to lower environmental pollution and a smaller carbon footprint, aligning with the goals of sustainable agriculture [11,15]. 4) Additionally, PGPR play a pivotal role in augmenting soil fertility and structure by promoting organic matter breakdown and nutrient cycling, thereby fostering overall soil health [7,9,15].
2. Mechanisms of Action
2.1. Nutrient Solubilization and Fixation
PGPR play a crucial role in enhancing plant nutrition by solubilizing phosphorus and fixing atmospheric nitrogen, making these essential nutrients more available to plants. Phosphorus is a vital nutrient for plant growth, but it often exists in forms that are not readily accessible to plants. Some PGPR can solubilize these insoluble phosphates through the production of organic acids like gluconic acid and citric acid, which lower the pH and chelate cations bound to phosphate group, thereby releasing it into a form that plants can absorb [16]. This process is facilitated by enzymes like phytases and phosphatases, which further aid in the mineralization of organic phosphorus compounds [17]. Recent studies have highlighted the effectiveness of PGPR strains from genera such as Bacillus, Pseudomonas, and Enterobacter in solubilizing phosphate and promoting plant growth [18].
In addition to phosphorus solubilization, PGPR are instrumental in fixing atmospheric nitrogen, a process that converts inert nitrogen gas into ammonia, which plants can incorporate. This biological nitrogen fixation is primarily carried out by diazotrophic bacteria, which possess the nitrogenase enzyme complex capable of reducing atmospheric nitrogen to ammonia [19]. PGPR such as Azospirillum and Rhizobium are well-known for their nitrogen-fixing capabilities, particularly in association with leguminous plants where they form symbiotic relationships in root nodules [20]. These interactions not only enhance nitrogen availability but also improve soil fertility and reduce the necessity for chemical nitrogen fertilizers, contributing to more sustainable agricultural practices [19].
2.2. Phytohormone Production
PGPR are known for their ability to produce phytohormones such as auxins, gibberellins, and cytokinins, which play a crucial role in enhancing plant growth and development. Auxins, particularly indole-3-acetic acid (IAA), are among the most studied phytohormones produced by PGPR [4,5]. IAA is primarily produced in the rhizosphere and is instrumental in promoting root elongation, root hair formation, and lateral root development, which collectively enhance the plant’s ability to absorb water and nutrients [21,22]. This hormone is produced by various PGPR species, including Azospirillum, Pseudomonas, and Agrobacterium. IAA’s production is also considered a key mechanism through which PGPR facilitate plant growth [23].
Gibberellins (GAs) are another group of phytohormones produced by PGPR that significantly influence plant growth. These hormones are involved in promoting seed germination, stem elongation, and flowering. PGPR such as Bacillus and Pseudomonas species have been reported to produce gibberellins, which can enhance plant growth even under stressful conditions [24,25]. The application of PGPR that produce gibberellins can be particularly beneficial in improving crop yield and resilience to unfavorable stresses, making them a valuable component of sustainable agricultural practices [26].
Cytokinins are another class of phytohormones produced by PGPR that play a vital role in cell division, shoot initiation, and leaf expansion. These phytohormones interact with auxins to regulate various aspects of plant growth and development, including delaying leaf senescence and enhancing chlorophyll production [27,28]. The production of cytokinins by PGPR can lead to improved plant vigor and productivity, as demonstrated in studies involving Bacillus megaterium and other cytokinin-producing rhizobacteria [29].
2.3. Biocontrol of Pathogens
PGPR protect plants from pathogens through several mechanisms, including the production of antimicrobial compounds and competition for resources. These bacteria reside in the rhizosphere and play a pivotal role in enhancing plant health by naturally suppressing plant diseases. One of the primary mechanisms by which PGPR exert their protective effects is through the production of antimicrobial compounds like antibiotics, siderophores, and enzymes that degrade pathogen cell walls [4,6]. For instance, PGPR like Pseudomonas and Bacillus species produce antibiotics that inhibit the growth of harmful pathogens, thereby reducing disease incidence in plants [30].
In addition to producing antimicrobial compounds, PGPR compete with pathogens for nutrients and ecological niches, effectively limiting the resources available to harmful microbes [31]. This competition is crucial in the rhizosphere, where nutrients can be scarce, and the ability of PGPR to efficiently utilize these resources can outcompete and suppress pathogenic organisms [32]. By rapidly colonizing plant roots and establishing themselves in the rhizosphere, PGPR can effectively prevent the colonization and proliferation of pathogens, thereby acting as a natural biocontrol agent [33].
Furthermore, PGPR can induce systemic resistance in plants, enhancing their innate immune responses against a broad spectrum of pathogens [4,6]. This induced resistance is often mediated by signaling molecules such as salicylic acid, jasmonic acid, and ethylene, which activate defense pathways in plants, providing them with enhanced protection against future pathogen attacks [34]. The integration of PGPR into agricultural practices not only helps in reducing the reliance on chemical pesticides but also contributes to sustainable agriculture by promoting plant health and resilience in an environmentally friendly manner [9].
2.4. Abiotic Stress Tolerance
PGPR play a significant role in helping plants cope with abiotic stresses such as drought, salinity, and heavy metal toxicity. These stresses are exacerbated by climate change and pose significant challenges to agricultural productivity. PGPR mitigate these stresses through different mechanisms such as the production of phytohormones, modulation of antioxidant systems, and enhancement of nutrient uptake [35,36].
In the context of drought stress, PGPR enhance plant tolerance by producing phytohormones like indole-3-acetic acid (IAA) and gibberellic acid, which promote root growth and increase water uptake efficiency [37,38]. Additionally, PGPR can produce 1-aminocyclopropane-1-carboxylate deaminase (ACC), an enzyme that lowers ethylene levels in plants [4]. Since ethylene can inhibit root growth under stress conditions, reducing its concentration helps in maintaining root growth and function during drought [39].
For salinity stress, PGPR improve plant resilience by enhancing ionic balance and osmotic adjustment. They achieve this by producing osmoprotectants and modulating ion transporters that help maintain a favorable potassium to sodium ratio, which is necessary for cellular function under saline conditions [40,41]. PGPR also enhance the antioxidant capacity of plants, reducing oxidative damage caused by salt-induced stress [35].
In the case of heavy metal toxicity in plants and soil, PGPR assist in phytoremediation by secreting chelating agents and enzymes that transform metals into less toxic forms. They are also capable of immobilizing heavy metals in the rhizosphere, which helps prevent these metals from being absorbed by plants [42,43]. This not only protects plants from metal toxicity but also improves soil health by reducing metal bioavailability [44].
3. Applications in Agriculture
3.1. Biofertilizers
PGPR have emerged as a promising alternative to synthetic fertilizers, offering a sustainable approach to improve soil fertility and plant growth. These beneficial bacteria are naturally occurring soil microorganisms that colonize plant roots (or even other parts of the plant) and promote growth through various mechanisms. Recent studies have shown that PGPR can improve soil biological activity [45,46]. The formulation of PGPR biofertilizers involves isolating effective strains from the rhizosphere, mass-producing them through fermentation, and developing stable formulations such as liquid suspensions, powders, or granules [47].
The application of PGPR biofertilizers can be tailored to different crops and farming practices. Methods such as seed inoculation, soil drenching, foliar sprays, and root dipping are commonly used to deliver PGPR to plants. These applications enhance root colonization, improve nutrient uptake, and protect plants from pathogens [45,48]. PGPR biofertilizers offer several advantages over conventional synthetic fertilizers. PGPR biofertilizers are environmentally sustainable, reducing soil and water pollution while maintaining soil health and fertility [47]. PGPR also improve nutrient efficiency by enhancing the availability of nitrogen, phosphorus, and other essential nutrients, thereby reducing the need for chemical fertilizers [45].
Moreover, PGPR enhance plant resilience to abiotic stresses by producing stress-relieving compounds like ACC-deaminase and inducing systemic resistance , contributing to healthier plant growth [45,48].. While the initial costs of PGPR biofertilizers may be higher, they can lead to long-term cost savings by reducing the need for chemical inputs and improving crop yields [47].
3.2. Seed Treatments and Soil Amendments
PGPR are applied to crops through various methods, each tailored to optimize their effectiveness in different environments and crop types. One common method is seed inoculation, where seeds are coated with PGPR before planting. This technique promotes early root colonization, leading to improved seedling vigor and nutrient uptake. Seed inoculation has been shown to be particularly effective in enhancing the growth of crops like maize and cowpea, as it facilitates the establishment of healthy soil microbiome [49].
Another effective method is soil amendment, which involves incorporating PGPR into the soil to improve its fertility and structure. This approach enhances nutrient availability and promotes root growth, making it suitable for various crops, including lettuce and poplar. Recent studies have demonstrated that soil amendments with PGPR can significantly improve plant growth and yield by enhancing nutrient uptake and reducing the need for chemical fertilizers [50].
Additionally, PGPR can be applied through foliar sprays and root dipping, which are particularly useful in environments where soil conditions are suboptimal or where rapid microbial colonization is needed [49].
The effectiveness of PGPR applications varies depending on the crop and environmental conditions. For instance, in hydroponic systems, PGPR have been used to reduce mineral fertilizer use while maintaining or even improving crop yield and quality. This is particularly beneficial in regions where mineral fertilizers are costly or difficult to obtain [51].
4. Challenges and Limitations
4.1. Consistency and Survival
The inconsistent performance of PGPR in different soils and environmental conditions presents a significant challenge to their widespread adoption in sustainable agriculture. One of the primary reasons for this variability is the complex interaction between PGPR and native soil microbiome. These interactions can influence the colonization and efficacy of PGPR, as native microbes often compete with introduced strains for resources and niches [52]. Additionally, the physiochemical properties of the soil, such as pH, nutrient content, and moisture levels, can affect the survival and activity of PGPR. This makes it difficult to predict their performance across different environments [53].
Another factor contributing to the inconsistent performance of PGPR is the specificity of plant-microbe interactions. Different plant species and even cultivars within a species can exhibit varying responses to the same PGPR strains. This host specificity can result in variable plant growth promotion and disease resistance outcomes, depending on the plant genotype and the PGPR strain used [9]. Moreover, environmental stressors such as temperature fluctuations can further complicate the effectiveness of PGPR, as these conditions can alter plant physiology and microbial community dynamics in the rhizosphere [54].
To address these challenges, several strategies have been proposed, with one being to select robust PGPR strains that are adaptable to local environmental conditions and compatible with specific crops is crucial. This involves screening for PGPR that can withstand stress factors and that have shown effectiveness with local plant species [54]. Additionally, developing advanced formulations that enhance the viability and stability of PGPR during storage, transport, and application can improve their persistence in the soil [53]. Utilizing consortia of different PGPR strains that have synergistic effects can also enhance nutrient acquisition, disease suppression, and stress tolerance [55].
4.2. Compatibility and Interaction
The compatibility of PGPR with target crops and indigenous soil microbiomeis crucial for their successful application in sustainable agriculture. This compatibility ensures that PGPR can effectively colonize plant roots, promote growth, and provide protection against pathogens. One of the primary challenges in achieving this compatibility is the strain specificity of PGPR, as certain strains may only benefit specific plants or perform optimally under particular environmental conditions [51]. This specificity necessitates careful selection of PGPR strains that are well-suited to the target crop and its unique growing environment.
The interactions between PGPR and native soil microbiome also play a significant role in determining the effectiveness of PGPR applications. Indigenous microorganisms can compete with introduced PGPR strains for resources and ecological niches, potentially limiting their colonization and persistence in the rhizosphere [52]. It is essential to understand these interactions, as they can influence the overall efficacy of PGPR in promoting plant growth and health. For example, PGPR must be able to survive and proliferate in the presence of native soil microbes to exert their beneficial effects [33].
To address these challenges, it is important to select PGPR strains that are not only effective with the target crop but also compatible with the existing soil microbiome. This involves screening for strains that can thrive in the specific soil conditions and that have demonstrated positive interactions with the target plant species [55]. Additionally, developing formulations that enhance the stability and viability of PGPR during storage and application can improve their performance in diverse environments [56].
4.3. Commercialization and Adoption
The widespread adoption of PGPR technologies faces several significant barriers, including regulatory challenges and a lack of farmer awareness. Regulatory issues are particularly daunting because each country has its own set of regulations governing the use of microbial products in agriculture. These regulations can be complex and costly to navigate, often requiring significant investment in time and resources to ensure compliance. For example, the high costs associated with the development and registration of new biocontrol agents (BCAs) have been identified as a barrier in countries like Australia, where regulatory frameworks demand rigorous evaluation to ensure environmental safety and efficacy [57,58]. This complexity is compounded by the lack of standardized international regulations, which can hinder the global commercialization of PGPR products [33,59].
In addition to regulatory hurdles, there is a critical need to increase farmer awareness and education regarding the benefits and application of PGPR technologies. Many farmers may be unfamiliar with PGPR or lack the necessary knowledge to integrate these biological solutions into their existing farming practices. This lack of awareness can lead to slow adoption rates, as farmers may perceive chemical fertilizers and pesticides as more reliable and predictable compared to PGPR [58]. Educational programs and workshops that demonstrate the economic and environmental benefits of PGPR can play a crucial role in overcoming this barrier, helping farmers understand how to effectively use PGPR to enhance crop productivity and sustainability [57,60].
Moreover, the scalability and cost-effectiveness of PGPR strategies are critical for their widespread adoption. PGPR technologies must be adaptable to different crops, environments, and farming practices to be effective. This adaptability requires ongoing research and development to tailor PGPR solutions to specific agricultural contexts [55]. Additionally, efforts must be made to reduce production costs and improve accessibility, particularly for farmers in developing countries. Collaboration among researchers, farmers, governments, and industry stakeholders is essential to develop cost-effective and sustainable PGPR products and practices [55].
5. Advances and Future Directions
5.1. Nano-Encapsulation Technology
Nano-encapsulation offers a promising method to increasing the efficacy and stability of PGPR formulations, which can address some of the challenges associated with their application in agriculture. By encapsulating PGPR in nanoparticles, it is possible to protect these beneficial microbes from environmental stressors, such as UV radiation, desiccation, and temperature fluctuations, thereby improving their survival and functionality [61]. This protective mechanism ensures that PGPR can be delivered more effectively to plant roots, enhancing their colonization and promoting better plant growth and resilience [62].
Recent studies have demonstrated the potential of various encapsulation materials, such as alginate, silica nanoparticles, and carbon nanotubes, to improve the delivery and performance of PGPR. For instance, the encapsulation of Pseudomonas sp. in alginate beads with salicylic acid and zinc oxide nanoparticles has shown enhanced antifungal activity and superior plant growth-promoting effects on rice seedlings compared to non-encapsulated strains [62]. Similarly, nano-encapsulated Bacillus subtilis using sodium alginate, starch, and bentonite has been effective in controlling the proliferation of Rhizoctonia solani and increasing bean vegetative growth parameters [63].
The use of nano-encapsulation not only improves the stability and efficacy of PGPR formulations but also allows for controlled and sustained release of the bacteria into the soil. This ensures a more consistent and prolonged interaction between PGPR and the plant roots, which is crucial for maximizing their growth-promoting effects [63,64]. Moreover, nano-encapsulation can enhance the resilience of PGPR to abiotic stresses, such as drought and salinity, by providing a stable microenvironment that supports their metabolic activity [65].
5.2. Biotechnological Approaches
The integration of genetic engineering and bioinformatics in the development of PGPR strains holds significant promise for enhancing their effectiveness in agriculture. Genetic engineering allows for the modification of PGPR strains to enhance desirable traits such as nutrient solubilization, phytohormone production, and pathogen resistance. By employing techniques such as CRISPR-Cas9, researchers can precisely edit the genomes of PGPR to introduce or enhance specific plant growth-promoting traits, thereby improving their efficacy under various environmental conditions [66,67].
Bioinformatics plays a crucial role in this process by enabling the analysis and interpretation of large genomic datasets, which is essential for identifying genes responsible for beneficial traits in PGPR. Through genome mining and comparative genomics, researchers can uncover biosynthetic gene clusters and regulatory networks that contribute to the plant growth-promoting capabilities of PGPR [68]. This information is invaluable for designing genetically engineered strains that are more robust and effective in promoting plant health and productivity [69].
Moreover, bioinformatics tools facilitate the prediction and modeling of PGPR interactions with plant hosts and soil microbiome, allowing for the optimization of strain selection and application strategies. By understanding these complex interactions, scientists can develop PGPR strains that are better customized to given crops and environmental conditions, thereby enhancing their performance in the field [70]. This approach not only improves the efficacy of PGPR but also supports sustainable agriculture by lowering the need for chemical fertilizers and pesticides [71].
5.3. Integrated Pest Management (IPM)
Incorporating PGPR into Integrated Pest Management (IPM) strategies offers a comprehensive approach to enhancing crop health and yield while reducing reliance on chemical pesticides. By incorporating PGPR into IPM, farmers can leverage these natural processes to create more resilient agricultural systems that are less dependent on synthetic chemicals, thereby minimizing environmental impact and promoting sustainable practices [32].
PGPR contribute to IPM by suppressing plant diseases through various mechanisms, including the production of pathogen-antagonizing compounds [6] and the stimulation of systemic resistance in plants [4,6]. These actions not only help control pathogens but also enhance the plant’s innate defense responses, making them more resistant to pest attacks [33]. For example, PGPR can produce antibiotics and enzymes that inhibit the growth of harmful microbes, while also triggering plant immune responses that bolster the plant’s ability to withstand pest pressures [34].
Moreover, PGPR improve nutrient uptake and root architecture, indirectly contributing to pest management. By enhancing root growth and nutrient acquisition, PGPR help plants become more vigorous and less susceptible to pest infestations. This improved plant health can lead to increased tolerance to pest damage and a reduction in the need for chemical interventions [55].
6. Conclusions
The integration of PGPR into agricultural systems as biofertilizers offers a sustainable alternative to chemical fertilizers, promoting nutrient cycling and improving plant health and yield (Figure 1A). By solubilizing phosphorus and fixing nitrogen, PGPR enhance nutrient availability and uptake, leading to improved plant growth and productivity [18]. Moreover, the use of PGPR can mitigate environmental impacts associated with excessive fertilizer use, such as soil degradation and water pollution, thereby supporting sustainable agriculture (Table 1) [16,20]. PGPR biofertilizers represent a viable alternative to synthetic fertilizers, providing sustainable solutions for enhancing crop productivity and soil health. Their ability to improve nutrient uptake, enhance stress tolerance, and suppress diseases makes them an invaluable tool in the pursuit of sustainable agriculture, particularly as the global demand for food continues to rise [45,47]. As research continues to advance, the development of more effective PGPR strains and formulations will be crucial in maximizing their benefits for crop production and environmental health [17].
The production of auxins, gibberellins, and cytokinins by PGPR significantly contributes to their ability to promote plant growth and development (Table 1). These phytohormones enhance root and shoot growth, improve nutrient uptake, and increase plant resilience to environmental stresses which makes PGPR an integral part of modern sustainable agriculture (Figure 1B) [26,124]. As research continues to explore the complex interactions between PGPR-produced phytohormones and plant physiology, the potential for optimizing PGPR applications in agriculture remains promising [125].
The integration of PGPR into agricultural practices offers a sustainable approach to managing abiotic stresses (Figure 1C). By enhancing plant growth and resilience, PGPR contribute to improved crop yields and stability, even under adverse environmental conditions [39,126]. PGPR offer a promising approach to biopesticide development, providing an eco-friendly and effective means of controlling plant diseases and pests (Figure 1C). Their ability to enhance plant resilience and promote growth makes them an invaluable tool in the pursuit of sustainable agricultural practices [55]. As research continues to advance, the development of more effective PGPR strains and formulations will be crucial in maximizing their benefits for stress management in agriculture (Table 1) [127].
The application of PGPR through methods like seed inoculation and soil amendments offers a sustainable approach to improving crop productivity and resilience (Figure 1D). By enhancing nutrient uptake and stress tolerance, PGPR contribute to more sustainable agricultural practices, reducing the reliance on chemical inputs and supporting global food security [9].While the inconsistent performance of PGPR in different soils and environmental conditions poses challenges, ongoing research and innovation in PGPR selection, formulation, and application strategies hold promise for overcoming these obstacles.
By optimizing PGPR inoculum efficacy in field conditions, researchers and farmers can enhance crop productivity, soil health, and environmental sustainability, contributing to more resilient agricultural systems [9,53]. Despite those promising advancements, further research is needed to optimize nano-encapsulation techniques and assess their long-term impacts on soil health and plant growth. The scalability and cost-effectiveness of these technologies remain challenges that need to be addressed to facilitate their widespread adoption in agriculture [128]. Nonetheless, the integration of nanotechnology with PGPR formulations represents a significant step forward in developing more efficient and sustainable agricultural practices (Table 1) [129].
Ensuring the compatibility of PGPR with target crops and indigenous soil microflora is essential for maximizing their benefits in sustainable agriculture (Figure 2). By selecting appropriate strains and understanding the complex interactions within the soil ecosystem, researchers and practitioners can enhance the effectiveness of PGPR applications, leading to improved crop productivity and resilience (Table 1) [51,52].While PGPR technologies hold significant potential for improving sustainable agriculture, overcoming regulatory hurdles and increasing farmer awareness are crucial for their widespread adoption. By addressing these challenges, PGPR can become a more integral part of agricultural systems, contributing to enhanced crop productivity and environmental sustainability [48].
Despite these advancements, challenges remain in the large-scale application of genetically engineered PGPR. Regulatory hurdles and public concerns about genetically modified organisms (GMOs) can impede the commercialization of these strains. However, ongoing research and dialogue among scientists, policymakers, and stakeholders are essential to address these issues and realize the full potential of genetic engineering and bioinformatics in developing effective PGPR strains for sustainable agriculture [130,131].
The integration of PGPR into IPM strategies requires careful selection of compatible strains that can thrive in specific environmental conditions and work synergistically with other IPM components. This approach not only enhances the effectiveness of pest management but also contributes to overall crop productivity and sustainability [55]. As research continues to advance, the development of tailored PGPR formulations and application methods will be crucial in maximizing their benefits within IPM frameworks, ultimately leading to more resilient and productive agricultural systems (Figure 3) [32]. PGPR protect plants from pathogens through a combination of antimicrobial production, resource competition, and induction of systemic resistance. These mechanisms make PGPR a valuable component of integrated pest management strategies, offering a sustainable alternative to chemical-based disease control methods (Table 1) [133]. As research continues to advance, the development of more effective PGPR strains and formulations will be crucial in maximizing their benefits for crop protection and productivity [11].
Author Contributions
Conceptualization, Supervision, Writing—original draft preparation, Writing—review & editing, Visualization, P.Y.; Writing—review & editing, A.C., S.S., C.H., L.L., M.A.A. All authors have read and approved the final version of the manuscript.
Funding
This research received no external funding
Acknowledgments
We acknowledge Huang Lab members for the fruitful discussion on this topic.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mohanty, P.; Singh, P.K.; Chakraborty, D.; Mishra, S.; Pattnaik, R. Insight Into the Role of PGPR in Sustainable Agriculture and Environment. Front. Sustain. Food Syst. 2021, 5. [Google Scholar] [CrossRef]
- Yang, P.; Bokros, N.; Debolt, S.; Zhao, Z.; Xia, Y. Genome Sequence Source of Bacillus Amyloliquefaciens Strain GD4a, a Bacterial Endophyte Associated with Switchgrass Plants. Phytobiomes Journal 2022, 6, 354–357. [Google Scholar] [CrossRef]
- Sahib, M.R.; Yang, P.; Bokros, N.; Shapiro, N.; Woyke, T.; Kyrpides, N.C.; Xia, Y.; DeBolt, S. Improved Draft Genome Sequence of Microbacterium Sp. Strain LKL04, a Bacterial Endophyte Associated with Switchgrass Plants. Microbiol Resour Announc 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Yuan, P.; Liu, W.; Zhao, Z.; Bernier, M.C.; Zhang, C.; Adhikari, A.; Opiyo, S.O.; Zhao, L.; Banks, F.; et al. Plant Growth Promotion and Plant Disease Suppression Induced by Bacillus Amyloliquefaciens Strain GD4a. Plants 2024, 13, 672. [Google Scholar] [CrossRef]
- Yang, P.; Liu, W.; Yuan, P.; Zhao, Z.; Zhang, C.; Opiyo, S.O.; Adhikari, A.; Zhao, L.; Harsh, G.; Xia, Y. Plant Growth Promotion and Stress Tolerance Enhancement through Inoculation with Bacillus Proteolyticus OSUB18. Biology 2023, 12, 1495. [Google Scholar] [CrossRef]
- Yang, P.; Zhao, Z.; Fan, J.; Liang, Y.; Bernier, M.C.; Gao, Y.; Zhao, L.; Opiyo, S.O.; Xia, Y. Bacillus Proteolyticus OSUB18 Triggers Induced Systemic Resistance against Bacterial and Fungal Pathogens in Arabidopsis. Frontiers in Plant Science 2023, 14. [Google Scholar] [CrossRef]
- Zhu, Y.-G.; Peng, J.; Chen, C.; Xiong, C.; Li, S.; Ge, A.; Wang, E.; Liesack, W. Harnessing Biological Nitrogen Fixation in Plant Leaves. Trends in Plant Science 2023, 0. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Lee, T.; Schiessl, K.; Oldroyd, G.E.D. Processing of NODULE INCEPTION Controls the Transition to Nitrogen Fixation in Root Nodules. Science 2021, 374, 629–632. [Google Scholar] [CrossRef]
- de Andrade, L.A.; Santos, C.H.B.; Frezarin, E.T.; Sales, L.R.; Rigobelo, E.C. Plant Growth-Promoting Rhizobacteria for Sustainable Agricultural Production. Microorganisms 2023, 11, 1088. [Google Scholar] [CrossRef]
- Wang, D.; Wang, C.; Chen, Y.; Xie, Z. Developing Plant-Growth-Promoting Rhizobacteria: A Crucial Approach for Achieving Sustainable Agriculture. Agronomy 2023, 13, 1835. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability—A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.N.; Jha, D.K. Plant Growth-Promoting Rhizobacteria (PGPR): Emergence in Agriculture. World J Microbiol Biotechnol 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, D.; Singh, B. Plant Growth Promoting Rhizobacteria-an Efficient Tool for Agriculture Promotion. Adv Plants Agric Res 2016, 4. [Google Scholar] [CrossRef]
- Hasan, A.; Tabassum, B.; Hashim, M.; Khan, N. Role of Plant Growth Promoting Rhizobacteria (PGPR) as a Plant Growth Enhancer for Sustainable Agriculture: A Review. Bacteria 2024, 3, 59–75. [Google Scholar] [CrossRef]
- Mekonnen, H.; Kibret, M. The Roles of Plant Growth Promoting Rhizobacteria in Sustainable Vegetable Production in Ethiopia. Chemical and Biological Technologies in Agriculture 2021, 8, 15. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate Solubilizing Microbes: Sustainable Approach for Managing Phosphorus Deficiency in Agricultural Soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Frontiers in Microbiology 2017, 8. [Google Scholar] [CrossRef]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial Features of Plant Growth-Promoting Rhizobacteria for Improving Plant Growth and Health in Challenging Conditions: A Methodical Review. Science of The Total Environment 2020, 743, 140682. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Klingl, A.; Lin, Y.-Y.; Boul, E.; Thomas-Oates, J.; Marín, M. A Subcompatible Rhizobium Strain Reveals Infection Duality in Lotus. Journal of Experimental Botany 2019, 70, 1903–1913. [Google Scholar] [CrossRef]
- Ahemad, M.; Kibret, M. Mechanisms and Applications of Plant Growth Promoting Rhizobacteria: Current Perspective. Journal of King Saud University - Science 2014, 26, 1–20. [Google Scholar] [CrossRef]
- Poveda, J.; Eugui, D.; Abril-Urías, P.; Velasco, P. Endophytic Fungi as Direct Plant Growth Promoters for Sustainable Agricultural Production. Symbiosis 2021, 85, 1–19. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.-J.; Zhang, J.-Z. Aux/IAA Gene Family in Plants: Molecular Structure, Regulation, and Function. International Journal of Molecular Sciences 2018, 19, 259. [Google Scholar] [CrossRef]
- Grobelak, A.; Napora, A.; Kacprzak, M. Using Plant Growth-Promoting Rhizobacteria (PGPR) to Improve Plant Growth. Ecological Engineering 2015, 84, 22–28. [Google Scholar] [CrossRef]
- Nett, R.S.; Montanares, M.; Marcassa, A.; Lu, X.; Nagel, R.; Charles, T.C.; Hedden, P.; Rojas, M.C.; Peters, R.J. Elucidation of Gibberellin Biosynthesis in Bacteria Reveals Convergent Evolution. Nat Chem Biol 2017, 13, 69–74. [Google Scholar] [CrossRef]
- Rizza, A.; Jones, A.M. The Makings of a Gradient: Spatiotemporal Distribution of Gibberellins in Plant Development. Current Opinion in Plant Biology 2019, 47, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Adeleke, B.S.; Babalola, O.O. The Endosphere Microbial Communities, a Great Promise in Agriculture. Int Microbiol 2021, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ren, G.; Zhang, K.; Li, Z.; Miao, Y.; Guo, H. Leaf Senescence: Progression, Regulation, and Application. Molecular Horticulture 2021, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Hönig, M.; Plíhalová, L.; Husičková, A.; Nisler, J.; Doležal, K. Role of Cytokinins in Senescence, Antioxidant Defence and Photosynthesis. International Journal of Molecular Sciences 2018, 19, 4045. [Google Scholar] [CrossRef]
- Salamone, G. de; Eugenia, I. Direct Beneficial Effects of Cytokinin-Producing Rhizobacteria on Plant Growth. 2000.
- Lobhi, D.; Patil, N.P.; Sansinenea, E.; Sayyed, R.Z. Plant Growth-Promoting Rhizobacteria (PGPR): An Overview. In Secondary Metabolites and Volatiles of PGPR in Plant-Growth Promotion; Sayyed, R.Z., Uarrota, V.G., Eds.; Springer International Publishing: Cham, 2022; pp. 1–19. ISBN 978-3-031-07559-9. [Google Scholar]
- Yang, P.; Zhao, L.; Gao, Y.G.; Xia, Y. Detection, Diagnosis, and Preventive Management of the Bacterial Plant Pathogen Pseudomonas Syringae. Plants 2023, 12, 1765. [Google Scholar] [CrossRef]
- Wang, H.; Liu, R.; You, M.P.; Barbetti, M.J.; Chen, Y. Pathogen Biocontrol Using Plant Growth-Promoting Bacteria (PGPR): Role of Bacterial Diversity. Microorganisms 2021, 9, 1988. [Google Scholar] [CrossRef]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant Associated Rhizobacteria for Biocontrol and Plant Growth Enhancement. Front Plant Sci 2021, 12, 634796. [Google Scholar] [CrossRef] [PubMed]
- Al-Turki, A.; Murali, M.; Omar, A.F.; Rehan, M.; Sayyed, R.Z. Recent Advances in PGPR-Mediated Resilience toward Interactive Effects of Drought and Salt Stress in Plants. Front Microbiol 2023, 14, 1214845. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.A.; Kumar, V.; Bhat, M.A.; Wani, I.A.; Dar, F.L.; Farooq, I.; Bhatti, F.; Koser, R.; Rahman, S.; Jan, A.T. Mechanistic Insights of the Interaction of Plant Growth-Promoting Rhizobacteria (PGPR) With Plant Roots Toward Enhancing Plant Productivity by Alleviating Salinity Stress. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Ilangumaran, G.; Smith, D.L. Plant Growth Promoting Rhizobacteria in Amelioration of Salinity Stress: A Systems Biology Perspective. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Sheikh, I.; Kumar, V.; Yadav, A.N.; Dhaliwal, H.S.; Saxena, A.K. Alleviation of Drought Stress and Plant Growth Promotion by Pseudomonas Libanensis EU-LWNA-33, a Drought-Adaptive Phosphorus-Solubilizing Bacterium. Proc. Natl. Acad. Sci., India, Sect. B Biol. Sci. 2020, 90, 785–795. [Google Scholar] [CrossRef]
- Jochum, M.D.; McWilliams, K.L.; Borrego, E.J.; Kolomiets, M.V.; Niu, G.; Pierson, E.A.; Jo, Y.-K. Bioprospecting Plant Growth-Promoting Rhizobacteria That Mitigate Drought Stress in Grasses. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of Drought Stress Tolerance in Crops by Plant Growth Promoting Rhizobacteria. Microbiological Research 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Nawaz, A.; Shahbaz, M.; Asadullah; Imran, A. ; Marghoob, M.U.; Imtiaz, M.; Mubeen, F. Potential of Salt Tolerant PGPR in Growth and Yield Augmentation of Wheat (Triticum Aestivum L.) Under Saline Conditions. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Abbas, R.; Rasul, S.; Aslam, K.; Baber, M.; Shahid, M.; Mubeen, F.; Naqqash, T. Halotolerant PGPR: A Hope for Cultivation of Saline Soils. Journal of King Saud University - Science 2019, 31, 1195–1201. [Google Scholar] [CrossRef]
- Dehghani Bidgoli, R.; Azarnezhad, N.; Akhbari, M.; Ghorbani, M. Salinity Stress and PGPR Effects on Essential Oil Changes in Rosmarinus Officinalis L. Agriculture & Food Security 2019, 8, 2. [Google Scholar] [CrossRef]
- Goswami, M.; Deka, S. Plant Growth-Promoting Rhizobacteria—Alleviators of Abiotic Stresses in Soil: A Review. Pedosphere 2020, 30, 40–61. [Google Scholar] [CrossRef]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; AL-Harrasi, A. Plant Growth Promoting Bacteria as an Alternative Strategy for Salt Tolerance in Plants: A Review. Microbiological Research 2018, 209, 21–32. [Google Scholar] [CrossRef] [PubMed]
- García-López, J.V.; Redondo-Gómez, S.; Flores-Duarte, N.J.; Zunzunegui, M.; Rodríguez-Llorente, I.D.; Pajuelo, E.; Mateos-Naranjo, E. Exploring through the Use of Physiological and Isotopic Techniques the Potential of a PGPR-Based Biofertilizer to Improve Nitrogen Fertilization Practices Efficiency in Strawberry Cultivation. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Jian, Q.; Yao, X.; Guan, L.; Li, L.; Liu, F.; Zhang, C.; Li, D.; Tang, H.; Lu, L. Plant Growth-Promoting Rhizobacteria (PGPR) Improve the Growth and Quality of Several Crops. Heliyon 2024, 10, e31553. [Google Scholar] [CrossRef] [PubMed]
- Aloo, B.N.; Tripathi, V.; Makumba, B.A.; Mbega, E.R. Plant Growth-Promoting Rhizobacterial Biofertilizers for Crop Production: The Past, Present, and Future. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Kumar, M.; Giri, V.P.; Pandey, S.; Gupta, A.; Patel, M.K.; Bajpai, A.B.; Jenkins, S.; Siddique, K.H.M. Plant-Growth-Promoting Rhizobacteria Emerging as an Effective Bioinoculant to Improve the Growth, Production, and Stress Tolerance of Vegetable Crops. Int J Mol Sci 2021, 22, 12245. [Google Scholar] [CrossRef]
- Mahmood, A.; Turgay, O.C.; Farooq, M.; Hayat, R. Seed Biopriming with Plant Growth Promoting Rhizobacteria: A Review. FEMS Microbiology Ecology 2016, 92, fiw112. [Google Scholar] [CrossRef] [PubMed]
- Romero-Estonllo, M.; Ramos-Castro, J.; San Miguel del Río, Y.; Rodríguez-Garrido, B.; Prieto-Fernández, Á.; Kidd, P.S.; Monterroso, C. Soil Amendment and Rhizobacterial Inoculation Improved Cu Phytostabilization, Plant Growth and Microbial Activity in a Bench-Scale Experiment. Front. Microbiol. 2023, 14. [Google Scholar] [CrossRef]
- Ikiz, B.; Dasgan, H.Y.; Gruda, N.S. Utilizing the Power of Plant Growth Promoting Rhizobacteria on Reducing Mineral Fertilizer, Improved Yield, and Nutritional Quality of Batavia Lettuce in a Floating Culture. Sci Rep 2024, 14, 1616. [Google Scholar] [CrossRef]
- Samain, E.; Duclercq, J.; Ait Barka, E.; Eickermann, M.; Ernenwein, C.; Mazoyon, C.; Sarazin, V.; Dubois, F.; Aussenac, T.; Selim, S. PGPR-Soil Microbial Communities’ Interactions and Their Influence on Wheat Growth Promotion and Resistance Induction against Mycosphaerella Graminicola. Biology (Basel) 2023, 12, 1416. [Google Scholar] [CrossRef]
- Haskett, T.L.; Tkacz, A.; Poole, P.S. Engineering Rhizobacteria for Sustainable Agriculture. ISME J 2021, 15, 949–964. [Google Scholar] [CrossRef] [PubMed]
- Thakur, R.; Dhar, H.; July 2024, S.M. PGPR Strategies for Climate-Resilient Agriculture. Available online: https://www.the-microbiologist.com/features/pgpr-strategies-for-climate-resilient-agriculture/3389.article (accessed on 25 August 2024).
- Agbodjato, N.A.; Babalola, O.O. Promoting Sustainable Agriculture by Exploiting Plant Growth-Promoting Rhizobacteria (PGPR) to Improve Maize and Cowpea Crops. PeerJ 2024, 12, e16836. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Yadav, V.K.; Chundawat, R.S.; Soltane, R.; Awwad, N.S.; Ibrahium, H.A.; Yadav, K.K.; Vicas, S.I. Enhancing Plant Growth Promoting Rhizobacterial Activities through Consortium Exposure: A Review. Front. Bioeng. Biotechnol. 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Barratt, B.I.P.; Moran, V.C.; Bigler, F.; van Lenteren, J.C. The Status of Biological Control and Recommendations for Improving Uptake for the Future. BioControl 2018, 63, 155–167. [Google Scholar] [CrossRef]
- Begum, M.; Lees, E.; Ampt, P.; Mansfield, S. Development of Australian Commercial Producers of Invertebrate Biological Control Agents from 1971 to 2014. BioControl 2017, 62, 525–533. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.-P. Advances in Plant Growth-Promoting Bacterial Inoculant Technology: Formulations and Practical Perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Heimpel, G.E.; Yang, Y.; Hill, J.D.; Ragsdale, D.W. Environmental Consequences of Invasive Species: Greenhouse Gas Emissions of Insecticide Use and the Role of Biological Control in Reducing Emissions. PLOS ONE 2013, 8, e72293. [Google Scholar] [CrossRef]
- Balla, A.; Silini, A.; Cherif-Silini, H.; Chenari Bouket, A.; Alenezi, F.N.; Belbahri, L. Recent Advances in Encapsulation Techniques of Plant Growth-Promoting Microorganisms and Their Prospects in the Sustainable Agriculture. Applied Sciences 2022, 12, 9020. [Google Scholar] [CrossRef]
- Ravichandran, M.; Samiappan, S.C.; Rangaraj, S.; Murugan, K.; Al-Dhabi, N.A.; Karuppiah, P. Chapter 13 - Nanoemulsion Formulations with Plant Growth Promoting Rhizobacteria (PGPR) for Sustainable Agriculture. In Bio-Based Nanoemulsions for Agri-Food Applications; Abd-Elsalam, K.A., Murugan, K., Eds.; Nanobiotechnology for Plant Protection; Elsevier, 2022; pp. 207–223 ISBN 978-0-323-89846-1.
- Pour, M.M.; Saberi-Riseh, R.; Mohammadinejad, R.; Hosseini, A. Nano-Encapsulation of Plant Growth-Promoting Rhizobacteria and Their Metabolites Using Alginate-Silica Nanoparticles and Carbon Nanotube Improves UCB1 Pistachio Micropropagation. Journal of Microbiology and Biotechnology 2019, 29, 1096–1103. [Google Scholar] [CrossRef]
- Pour, M.M.; Riseh, R.S.; Ranjbar-Karimi, R.; Hassanisaadi, M.; Rahdar, A.; Baino, F. Microencapsulation of Bacillus Velezensis Using Alginate-Gum Polymers Enriched with TiO2 and SiO2 Nanoparticles. Micromachines 2022, 13, 1423. [Google Scholar] [CrossRef]
- Yang, K.; Wang, Q.; Wang, Y.; Li, S.; Gu, Y.; Gao, N.; Zhang, F.; Lei, P.; Wang, R.; Xu, H. Poly(γ-Glutamic Acid) Nanocoating To Enhance the Viability of Pseudomonas Stutzeri NRCB010 through Cell Surface Engineering. ACS Appl. Mater. Interfaces 2021, 13, 39957–39966. [Google Scholar] [CrossRef] [PubMed]
- Fahde, S.; Boughribil, S.; Sijilmassi, B.; Amri, A. Rhizobia: A Promising Source of Plant Growth-Promoting Molecules and Their Non-Legume Interactions: Examining Applications and Mechanisms. Agriculture 2023, 13, 1279. [Google Scholar] [CrossRef]
- Chieb, M.; Gachomo, E.W. The Role of Plant Growth Promoting Rhizobacteria in Plant Drought Stress Responses. BMC Plant Biology 2023, 23, 407. [Google Scholar] [CrossRef] [PubMed]
- Paterson, J.; Jahanshah, G.; Li, Y.; Wang, Q.; Mehnaz, S.; Gross, H. The Contribution of Genome Mining Strategies to the Understanding of Active Principles of PGPR Strains. FEMS Microbiology Ecology 2017, 93, fiw249. [Google Scholar] [CrossRef]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Saqib, S.; Ullah, F.; Ayaz, A.; Zaman, W. The Key Roles of ROS and RNS as a Signaling Molecule in Plant–Microbe Interactions. Antioxidants 2023, 12, 268. [Google Scholar] [CrossRef]
- Yao, K.; Wang, Y.; Li, X.; Ji, H. Genome-Wide Identification of the Soybean LysM-RLK Family Genes and Its Nitrogen Response. International Journal of Molecular Sciences 2023, 24, 13621. [Google Scholar] [CrossRef]
- Agrawal, R.; Satlewal, A.; Varma, A. Characterization of Plant Growth-Promoting Rhizobacteria (PGPR): A Perspective of Conventional Versus Recent Techniques. In Heavy Metal Contamination of Soils: Monitoring and Remediation; Sherameti, I., Varma, A., Eds.; Springer International Publishing: Cham, 2015; pp. 471–485. ISBN 978-3-319-14526-6. [Google Scholar]
- Stassinos, P.M.; Rossi, M.; Borromeo, I.; Capo, C.; Beninati, S.; Forni, C. Amelioration of Salt Stress Tolerance in Rapeseed (Brassica Napus) Cultivars by Seed Inoculation with Arthrobacter Globiformis. Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology 2022, 156, 370–383. [Google Scholar] [CrossRef]
- Ultanbekova, G.; Mukhatayeva, K.; Mamytova, N.; Kanalbek, G.; Kazizmurat, M. Isolation and Selection of Nodulating Bacteria from the Rhizosphere of Legume (Mash - Vigna Radiata) and Bean (Phaseolus Lunatus) Plants and to Enhance Specific Properties through Induced Mutagenesis Methods. BIO Web Conf. 2024, 100, 02033. [Google Scholar] [CrossRef]
- Sun, Y.; Sun, P.; Xue, J.; Du, Y.; Yan, H.; Wang, L.; Yi, X.; Sun, J.; Zhang, X.; Gao, J. Arthrobacter Wenxiniae Sp. Nov., a Novel Plant Growth-Promoting Rhizobacteria Species Harbouring a Carotenoids Biosynthetic Gene Cluster. Antonie van Leeuwenhoek 2022, 115, 353–364. [Google Scholar] [CrossRef]
- Spaepen, S.; Bossuyt, S.; Engelen, K.; Marchal, K.; Vanderleyden, J. Phenotypical and Molecular Responses of Arabidopsis Thaliana Roots as a Result of Inoculation with the Auxin-Producing Bacterium Azospirillum Brasilense. New Phytologist 2014, 201, 850–861. [Google Scholar] [CrossRef]
- El-Ballat, E.M.; Elsilk, S.E.; Ali, H.M.; Ali, H.E.; Hano, C.; El-Esawi, M.A. Metal-Resistant PGPR Strain Azospirillum Brasilense EMCC1454 Enhances Growth and Chromium Stress Tolerance of Chickpea (Cicer Arietinum L.) by Modulating Redox Potential, Osmolytes, Antioxidants, and Stress-Related Gene Expression. Plants 2023, 12, 2110. [Google Scholar] [CrossRef] [PubMed]
- Abdel Latef, A.A.H.; Abu Alhmad, M.F.; Kordrostami, M.; Abo–Baker, A.-B.A.-E.; Zakir, A. Inoculation with Azospirillum Lipoferum or Azotobacter Chroococcum Reinforces Maize Growth by Improving Physiological Activities Under Saline Conditions. J Plant Growth Regul 2020, 39, 1293–1306. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Omer, A.M.; Badawy, A.A.; Osman, M.S.; Ragaey, M.M. Strategy of Salt Tolerance and Interactive Impact of Azotobacter Chroococcum and/or Alcaligenes Faecalis Inoculation on Canola (Brassica Napus L.) Plants Grown in Saline Soil. Plants 2021, 10, 110. [Google Scholar] [CrossRef]
- Zalila-Kolsi, I.; Ben-Mahmoud, A.; Al-Barazie, R. Bacillus Amyloliquefaciens: Harnessing Its Potential for Industrial, Medical, and Agricultural Applications—A Comprehensive Review. Microorganisms 2023, 11, 2215. [Google Scholar] [CrossRef]
- Qi, H.-Y.; Wang, D.; Han, D.; Song, J.; Ali, M.; Dai, X.-F.; Zhang, X.-J.; Chen, J.-Y. Unlocking Antagonistic Potential of Bacillus Amyloliquefaciens KRS005 to Control Gray Mold. Frontiers in Microbiology 2023, 14. [Google Scholar] [CrossRef]
- Luo, Z.; Yan, Y.; Du, S.; Zhu, Y.; Pan, F.; Wang, R.; Xu, Z.; Xu, X.; Li, S.; Xu, H. Recent Advances and Prospects of Bacillus Amyloliquefaciens as Microbial Cell Factories: From Rational Design to Industrial Applications. Critical Reviews in Biotechnology 2023, 43, 1073–1091. [Google Scholar] [CrossRef]
- Xu, F.; Liao, H.; Zhang, Y.; Yao, M.; Liu, J.; Sun, L.; Zhang, X.; Yang, J.; Wang, K.; Wang, X.; et al. Coordination of Root Auxin with the Fungus Piriformospora Indica and Bacterium Bacillus Cereus Enhances Rice Rhizosheath Formation under Soil Drying. ISME J 2021, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Yu, P.; Wang, J.; Li, C.; Guo, H.; Liu, C.; Kong, L.; Yu, L.; Wu, S.; Lei, T.; et al. A Study on Prevalence and Characterization of Bacillus Cereus in Ready-to-Eat Foods in China. Front. Microbiol. 2020, 10. [Google Scholar] [CrossRef]
- Kulkova, I.; Dobrzyński, J.; Kowalczyk, P.; Bełżecki, G.; Kramkowski, K. Plant Growth Promotion Using Bacillus Cereus. International Journal of Molecular Sciences 2023, 24, 9759. [Google Scholar] [CrossRef]
- Zhou, C.; Zhu, J.; Qian, N.; Guo, J.; Yan, C. Bacillus Subtilis SL18r Induces Tomato Resistance Against Botrytis Cinerea, Involving Activation of Long Non-Coding RNA, MSTRG18363, to Decoy miR1918. Front. Plant Sci. 2021, 11, 634819. [Google Scholar] [CrossRef]
- Tian, T.; Sun, B.; Shi, H.; Gao, T.; He, Y.; Li, Y.; Liu, Y.; Li, X.; Zhang, L.; Li, S.; et al. Sucrose Triggers a Novel Signaling Cascade Promoting Bacillus Subtilis Rhizosphere Colonization. ISME J 2021, 15, 2723–2737. [Google Scholar] [CrossRef] [PubMed]
- Pohare, M.B.; Wagh, S.G.; Udayasuriyan, V. Bacillus Thuringiensis as Potential Biocontrol Agent for Sustainable Agriculture. In Current Trends in Microbial Biotechnology for Sustainable Agriculture; Yadav, A.N., Singh, J., Singh, C., Yadav, N., Eds.; Environmental and Microbial Biotechnology; Springer: Singapore, 2021; pp. 439–468. ISBN 9789811569494. [Google Scholar]
- Kahn, T.W.; Duck, N.B.; McCarville, M.T.; Schouten, L.C.; Schweri, K.; Zaitseva, J.; Daum, J. A Bacillus Thuringiensis Cry Protein Controls Soybean Cyst Nematode in Transgenic Soybean Plants. Nat Commun 2021, 12, 3380. [Google Scholar] [CrossRef] [PubMed]
- Malik, L.; Sanaullah, M.; Mahmood, F.; Hussain, S.; Shahzad, T. Co-Application of Biochar and Salt Tolerant PGPR to Improve Soil Quality and Wheat Production in a Naturally Saline Soil. Rhizosphere 2024, 29, 100849. [Google Scholar] [CrossRef]
- de Almeida, J.R.; Bonatelli, M.L.; Batista, B.D.; Teixeira-Silva, N.S.; Mondin, M.; dos Santos, R.C.; Bento, J.M.S.; de Almeida Hayashibara, C.A.; Azevedo, J.L.; Quecine, M.C. RZ2MS9, a Tropical Plant Growth-Promoting Rhizobacterium, Colonizes Maize Endophytically and Alters the Plant’s Production of Volatile Organic Compounds during Co-Inoculation with Ab-V5. Environmental Microbiology Reports 2021, 13, 812–821. [Google Scholar] [CrossRef]
- Madala, N.E.; Molinaro, A.; Dubery, I.A. Distinct Carbohydrate and Lipid-Based Molecular Patterns within Lipopolysaccharides from Burkholderia Cepacia Contribute to Defense-Associated Differential Gene Expression in Arabidopsis Thaliana. Innate immunity 2012, 18, 140–154. [Google Scholar] [CrossRef]
- Heo, A.Y.; Koo, Y.M.; Choi, H.W. Biological Control Activity of Plant Growth Promoting Rhizobacteria Burkholderia Contaminans AY001 against Tomato Fusarium Wilt and Bacterial Speck Diseases. Biology 2022, 11, 619. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, K.; Mitra, S.; Sarkar, A.; Maiti, T.K. Alleviation of Phytotoxic Effects of Cadmium on Rice Seedlings by Cadmium Resistant PGPR Strain Enterobacter Aerogenes MCC 3092. Journal of Hazardous Materials 2018, 351, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Sun, S.; Wang, R.; Ma, X.; Shen, S.; Luo, Y.; Ma, X.; Wu, T.; Li, S.; Yang, Z.; et al. Study on the Mechanism of Salt Relief and Growth Promotion of Enterobacter Cloacae on Cotton. BMC Plant Biol 2023, 23, 656. [Google Scholar] [CrossRef]
- Seo, H.; Kim, J.H.; Lee, S.-M.; Lee, S.-W. The Plant-Associated Flavobacterium: A Hidden Helper for Improving Plant Health. Plant Pathol J 2024, 40, 251–260. [Google Scholar] [CrossRef]
- Kraut-Cohen, J.; Shapiro, O.H.; Dror, B.; Cytryn, E. Pectin Induced Colony Expansion of Soil-Derived Flavobacterium Strains. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Sommer, A.; Wenig, M.; Knappe, C.; Kublik, S.; Foesel, B.U.; Schloter, M.; Vlot, A.C. A Salicylic Acid-Associated Plant-Microbe Interaction Attracts Beneficial Flavobacterium Sp. to the Arabidopsis Thaliana Phyllosphere. Physiologia Plantarum 2024, 176, e14483. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, K.; Mitra, S.; Sarkar, A.; Soren, T.; Maiti, T.K. Characterization of Cadmium-Resistant Klebsiella Pneumoniae MCC 3091 Promoted Rice Seedling Growth by Alleviating Phytotoxicity of Cadmium. Environ Sci Pollut Res 2017, 24, 24419–24437. [Google Scholar] [CrossRef] [PubMed]
- Kusale, S.P.; Attar, Y.C.; Sayyed, R.Z.; Malek, R.A.; Ilyas, N.; Suriani, N.L.; Khan, N.; El Enshasy, H.A. Production of Plant Beneficial and Antioxidants Metabolites by Klebsiellavariicola under Salinity Stress. Molecules 2021, 26, 1894. [Google Scholar] [CrossRef] [PubMed]
- Dubey, A.; Kumar, A.; Khan, M.L.; Payasi, D.K. Plant Growth-Promoting and Bio-Control Activity of Micrococcus Luteus Strain AKAD 3-5 Isolated from the Soybean (Glycine Max (L.) Merr.) Rhizosphere. [CrossRef]
- Badawy, I.H.; Hmed, A.A.; Sofy, M.R.; Al-Mokadem, A.Z. Alleviation of Cadmium and Nickel Toxicity and Phyto-Stimulation of Tomato Plant L. by Endophytic Micrococcus Luteus and Enterobacter Cloacae. Plants 2022, 11, 2018. [Google Scholar] [CrossRef]
- Kabiraj, A.; Halder, U.; Chitikineni, A.; Varshney, R.K.; Bandopadhyay, R. Insight into the Genome of an Arsenic Loving and Plant Growth-Promoting Strain of Micrococcus Luteus Isolated from Arsenic Contaminated Groundwater. Environ Sci Pollut Res 2024, 31, 39063–39076. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Zaid, A.; Abo-Baker, A.-B.A.-E.; Salem, W.; Abu Alhmad, M.F. Mitigation of Copper Stress in Maize by Inoculation with Paenibacillus Polymyxa and Bacillus Circulans. Plants 2020, 9, 1513. [Google Scholar] [CrossRef]
- E, Y.; Yuan, J.; Yang, F.; Wang, L.; Ma, J.; Li, J.; Pu, X.; Raza, W.; Huang, Q.; Shen, Q. PGPR Strain Paenibacillus Polymyxa SQR-21 Potentially Benefits Watermelon Growth by Re-Shaping Root Protein Expression. AMB Expr 2017, 7, 104. [Google Scholar] [CrossRef]
- Langendries, S.; Goormachtig, S. , A Jack of All Trades. Environmental Microbiology 2021, 23, 5659–5669. [Google Scholar] [CrossRef]
- Liu, H.; Li, Y.; Ge, K.; Du, B.; Liu, K.; Wang, C.; Ding, Y. Interactional Mechanisms of Paenibacillus Polymyxa SC2 and Pepper (Capsicum Annuum L.) Suggested by Transcriptomics. BMC Microbiol 2021, 21, 70. [Google Scholar] [CrossRef]
- Paulsen, I.T.; Press, C.M.; Ravel, J.; Kobayashi, D.Y.; Myers, G.S.A.; Mavrodi, D.V.; DeBoy, R.T.; Seshadri, R.; Ren, Q.; Madupu, R.; et al. Complete Genome Sequence of the Plant Commensal Pseudomonas Fluorescens Pf-5. Nat. Biotechnol. 2005, 23, 873–878. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Trotel-Aziz, P.; Villaume, S.; Rabenoelina, F.; Schwarzenberg, A.; Nguema-Ona, E.; Clément, C.; Baillieul, F.; Aziz, A. Bacillus Subtilis and Pseudomonas Fluorescens Trigger Common and Distinct Systemic Immune Responses in Arabidopsis Thaliana Depending on the Pathogen Lifestyle. Vaccines 2020, 8, 503. [Google Scholar] [CrossRef] [PubMed]
- Costa-Gutierrez, S.B.; Adler, C.; Espinosa-Urgel, M.; de Cristóbal, R.E. Pseudomonas Putida and Its Close Relatives: Mixing and Mastering the Perfect Tune for Plants. Appl Microbiol Biotechnol 2022, 106, 3351–3367. [Google Scholar] [CrossRef]
- Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Salt Stress Alleviation in Citrus Plants by Plant Growth-Promoting Rhizobacteria Pseudomonas Putida and Novosphingobium Sp. Plant Cell Rep 2018, 37, 1557–1569. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xu, Z.; Xie, J.; Hesselberg-Thomsen, V.; Tan, T.; Zheng, D.; Strube, M.L.; Dragoš, A.; Shen, Q.; Zhang, R.; et al. Bacillus Velezensis Stimulates Resident Rhizosphere Pseudomonas Stutzeri for Plant Health through Metabolic Interactions. ISME J 2021, 1–14. [Google Scholar] [CrossRef]
- Lami, M.J.; Adler, C.; Caram-Di Santo, M.C.; Zenoff, A.M.; de Cristóbal, R.E.; Espinosa-Urgel, M.; Vincent, P.A. Pseudomonas Stutzeri MJL19, a Rhizosphere-colonizing Bacterium That Promotes Plant Growth under Saline Stress. Journal of Applied Microbiology 2020, 129, 1321–1336. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Mushtaq, Z.; Nazir, A.; Jaffar, M.T.; Asghar, H.N.; Alzuaibr, F.M.; Alasmari, A.; Alqurashi, M. Growth Response of Cowpea (Vigna Unguiculata L.) Exposed to Pseudomonas Fluorescens, Pseudomonas Stutzeri, and Pseudomonas Gessardii in Lead Contaminated Soil. Plant Stress 2023, 10, 100259. [Google Scholar] [CrossRef]
- Jiang, S.; Li, J.; Wang, Q.; Yin, C.; Zhan, Y.; Yan, Y.; Lin, M.; Ke, X. Maize Growth Promotion by Inoculation with an Engineered Ammonium-Excreting Strain of Nitrogen-Fixing Pseudomonasstutzeri. Microorganisms 2022, 10, 1986. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Guinel, F.C.; Glick, B.R. Rhizobium Leguminosarum Biovar Viciae 1-Aminocyclopropane-1-Carboxylate Deaminase Promotes Nodulation of Pea Plants. Appl Environ Microbiol 2003, 69, 4396–4402. [Google Scholar] [CrossRef] [PubMed]
- Yadav, J.; Verma, J.P. Effect of Seed Inoculation with Indigenous Rhizobium and Plant Growth Promoting Rhizobacteria on Nutrients Uptake and Yields of Chickpea (Cicer Arietinum L.). European Journal of Soil Biology 2014, 63, 70–77. [Google Scholar] [CrossRef]
- Zahir, Z.A.; Zafar-ul-Hye, M.; Sajjad, S.; Naveed, M. Comparative Effectiveness of Pseudomonas and Serratia Sp. Containing ACC-Deaminase for Coinoculation with Rhizobium Leguminosarum to Improve Growth, Nodulation, and Yield of Lentil. Biol Fertil Soils 2011, 47, 457–465. [Google Scholar] [CrossRef]
- Suárez-Moreno, Z.R.; Vinchira-Villarraga, D.M.; Vergara-Morales, D.I.; Castellanos, L.; Ramos, F.A.; Guarnaccia, C.; Degrassi, G.; Venturi, V.; Moreno-Sarmiento, N. Plant-Growth Promotion and Biocontrol Properties of Three Streptomyces Spp. Isolates to Control Bacterial Rice Pathogens. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and Interactions in Plant Growth Promotion. Appl Microbiol Biotechnol 2019, 103, 1179–1188. [Google Scholar] [CrossRef]
- Dev, A.S.R.; Harish, S.; Karthikeyan, G.; Nivedha, M.; Sangeetha, C. Consortia of Streptomyces Spp. Triggers Defense/PAMP Genes during the Interaction of Groundnut Bud Necrosis Orthotospovirus in Tomato. 3 Biotech 2024, 14, 196. [Google Scholar] [CrossRef]
- Lu, L.; Liu, N.; Fan, Z.; Liu, M.; Zhang, X.; Tian, J.; Yu, Y.; Lin, H.; Huang, Y.; Kong, Z. A Novel PGPR Strain, Streptomyces Lasalocidi JCM 3373T, Alleviates Salt Stress and Shapes Root Architecture in Soybean by Secreting Indole-3-Carboxaldehyde. Plant, Cell & Environment 2024, 47, 1941–1956. [Google Scholar] [CrossRef]
- Wang, M.; Xue, J.; Ma, J.; Feng, X.; Ying, H.; Xu, H. Streptomyces Lydicus M01 Regulates Soil Microbial Community and Alleviates Foliar Disease Caused by Alternaria Alternata on Cucumbers. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Devi, S.; Sharma, M.; Manhas, R.K. Investigating the Plant Growth Promoting and Biocontrol Potentiality of Endophytic Streptomyces SP. SP5 against Early Blight in Solanum Lycopersicum Seedlings. BMC Microbiol 2022, 22, 285. [Google Scholar] [CrossRef] [PubMed]
- Antar, M.; Gopal, P.; Msimbira, L.A.; Naamala, J.; Nazari, M.; Overbeek, W.; Backer, R.; Smith, D.L. Inter-Organismal Signaling in the Rhizosphere. In Rhizosphere Biology: Interactions Between Microbes and Plants; Gupta, V.V.S.R., Sharma, A.K., Eds.; Springer: Singapore, 2021; pp. 255–293. ISBN 9789811561252. [Google Scholar]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Leontidou, K.; Genitsaris, S.; Papadopoulou, A.; Kamou, N.; Bosmali, I.; Matsi, T.; Madesis, P.; Vokou, D.; Karamanoli, K.; Mellidou, I. Plant Growth Promoting Rhizobacteria Isolated from Halophytes and Drought-Tolerant Plants: Genomic Characterisation and Exploration of Phyto-Beneficial Traits. Sci Rep 2020, 10, 14857. [Google Scholar] [CrossRef]
- Kumar, A.; Patel, J.S.; Meena, V.S.; Ramteke, P.W. Plant Growth-Promoting Rhizobacteria: Strategies to Improve Abiotic Stresses under Sustainable Agriculture. Journal of Plant Nutrition 2019, 42, 1402–1415. [Google Scholar] [CrossRef]
- Campos, E.V.R.; Pereira, A.D.E.S.; Aleksieienko, I.; Do Carmo, G.C.; Gohari, G.; Santaella, C.; Fraceto, L.F.; Oliveira, H.C. Encapsulated Plant Growth Regulators and Associative Microorganisms: Nature-Based Solutions to Mitigate the Effects of Climate Change on Plants. Plant Science 2023, 331, 111688. [Google Scholar] [CrossRef]
- Akhtar, N.; Ilyas, N.; Meraj, T.A.; Pour-Aboughadareh, A.; Sayyed, R.Z.; Mashwani, Z.-R.; Poczai, P. Improvement of Plant Responses by Nanobiofertilizer: A Step towards Sustainable Agriculture. Nanomaterials 2022, 12, 965. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Song, I.-G.; Yoon, H.; Park, J.-W. Sub-Micron Microplastics Affect Nitrogen Cycling by Altering Microbial Abundance and Activities in a Soil-Legume System. Journal of Hazardous Materials 2023, 460, 132504. [Google Scholar] [CrossRef] [PubMed]
- Enebe, M.C.; Babalola, O.O. The Influence of Plant Growth-Promoting Rhizobacteria in Plant Tolerance to Abiotic Stress: A Survival Strategy. Appl Microbiol Biotechnol 2018, 102, 7821–7835. [Google Scholar] [CrossRef] [PubMed]
- Fry, W.E. Principles of Plant Disease Management; Academic Press: Cambridge, MA, USA, 2012; ISBN 978-0-08-091830-3. [Google Scholar]
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms. Int J Mol Sci 2021, 22, 10529. [Google Scholar] [CrossRef]
Figure 1.
Roles of PGPR in sustainable agriculture. A. Nutrient availability. B. Phytohormone production. Abbreviations: Indole-3-Acetic Acid (IAA); Gibberellins (GA); Abscisic Acid (ABA). C. Biocontrol and stress tolerance. D. Soil health improvement. Created with BioRender.com.
Figure 1.
Roles of PGPR in sustainable agriculture. A. Nutrient availability. B. Phytohormone production. Abbreviations: Indole-3-Acetic Acid (IAA); Gibberellins (GA); Abscisic Acid (ABA). C. Biocontrol and stress tolerance. D. Soil health improvement. Created with BioRender.com.
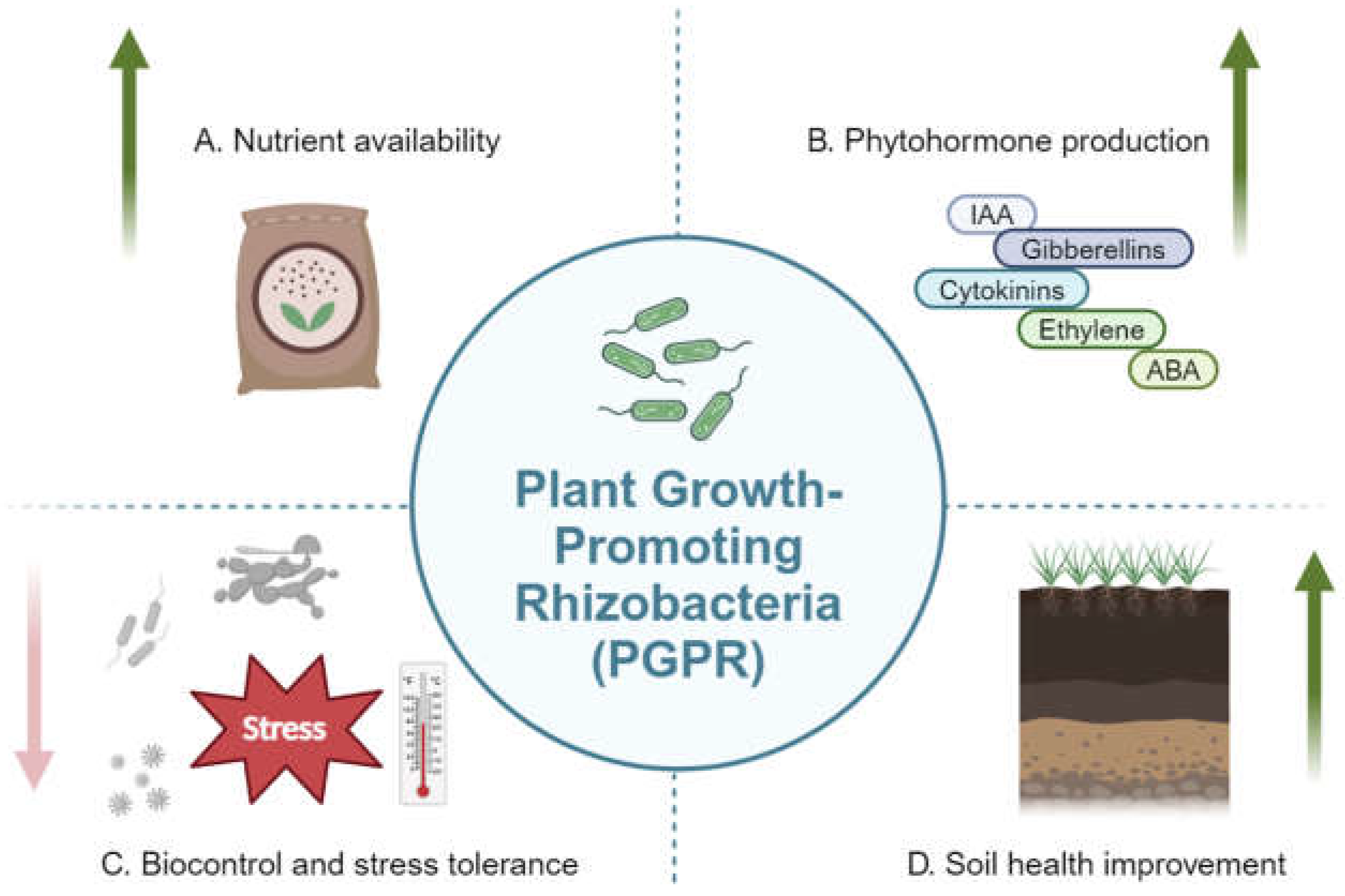
Figure 2.
The plant fitness tetrahedron for sustainable agriculture. Adapted from [31]. PGPR play a crucial role in maintaining the health and vitality of plants. Created with BioRender.com.
Figure 2.
The plant fitness tetrahedron for sustainable agriculture. Adapted from [31]. PGPR play a crucial role in maintaining the health and vitality of plants. Created with BioRender.com.
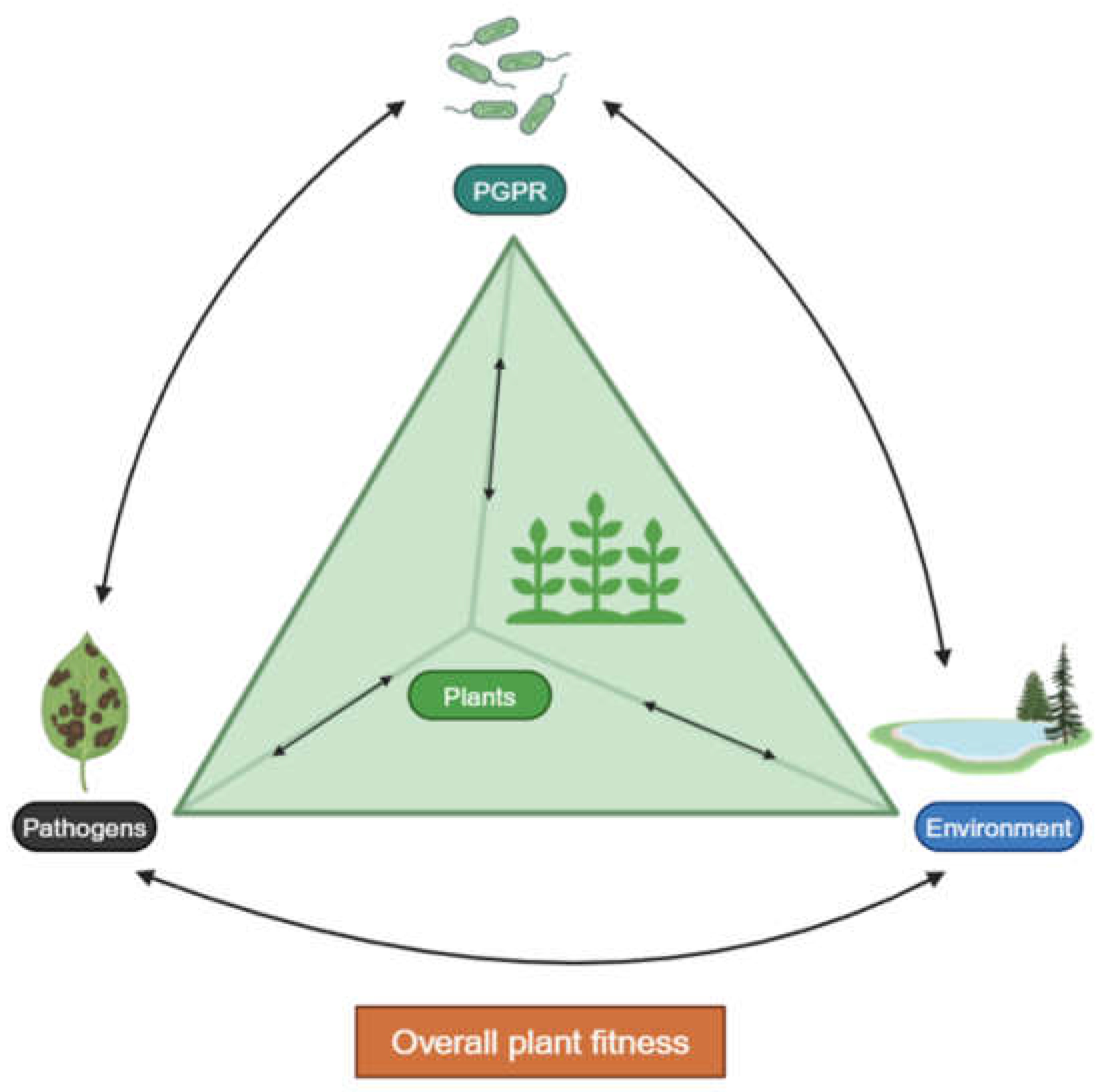
Figure 3.
Integrated pest management (IPM) with PGPR. IPM has six directions: protection, resistance, therapy, avoidance, exclusion, and eradication. Directions integrated with PGPR are highlighted in green. Adapted from [31,132]. Created with BioRender.com.
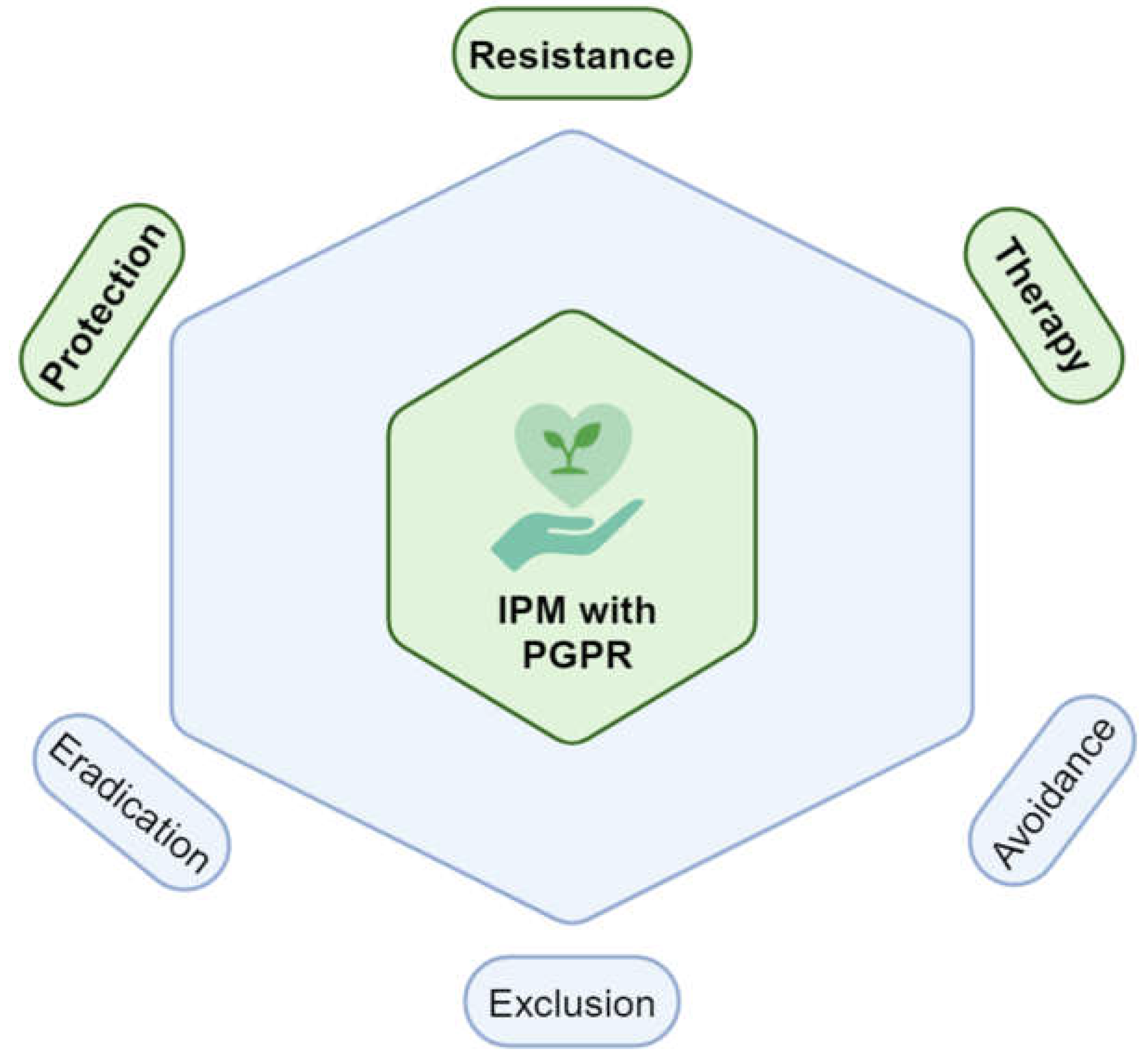

Table 1.
Potentials of PGPR from soil microbiome for sustainable agriculture.
| Functional PGPR Species | Source Isolated From | Beneficial Roles | Mechanisms of Action | Applicable Plants | References |
|---|---|---|---|---|---|
| Arthrobacter globiformis | Soil, rhizosphere | Nutrient solubilization, pathogen control | Phosphate solubilization, biocontrol of pathogens | Vegetables, cereals | [72,73,74] |
| Azospirillum brasilense | Soil, plant roots | Enhanced root growth, nitrogen supply | Nitrogen fixation, phytohormone production (auxins) | Cereals (e.g., maize, wheat), grasses | [75,76] |
| Azotobacter chroococcum | Soil, rhizosphere | Nitrogen fixation, soil fertility improvement | Nitrogen fixation, produces growth-promoting substances | Cereals, vegetables | [77,78] |
| Bacillus amyloliquefaciens | Soil, plant roots | Pathogen suppression, growth enhancement | Antifungal activity, induced systemic resistance | Fruits, vegetables | [4,79,80,81] |
| Bacillus cereus | Soil, plant roots | Disease control, plant vigor improvement | Biocontrol of pathogens, induced systemic resistance | Vegetables, cereals | [82,83,84] |
| Bacillus subtilis | Soil, plant roots | Disease suppression, nutrient uptake enhancement | Produces antibiotics, induces systemic resistance, solubilizes phosphorus | Vegetables, cereals, legumes | [85,86] |
| Bacillus thuringiensis | Soil, plant roots | Insect pest suppression, plant protection | Pest control, biocontrol of insects | Vegetables, fruits | [87,88,89,90] |
| Burkholderia cepacia | Soil, rhizosphere | Pathogen control, plant growth promotion | Produces antifungal compounds, competes with pathogens | Vegetables, ornamentals | [91,92] |
| Enterobacter cloacae | Soil, plant roots | Disease suppression, nutrient enhancement | Phosphate solubilization, biocontrol of pathogens | Vegetables, cereals | [93,94] |
| Flavobacterium johnsoniae | Soil, rhizosphere | Disease suppression, growth enhancement | Biocontrol of pathogens, growth promotion | Vegetables, cereals | [95,96,97] |
| Klebsiella pneumoniae | Soil, rhizosphere | Nitrogen supply, growth enhancement | Nitrogen fixation, growth promotion | Vegetables, cereals | [98,99] |
| Micrococcus luteus | Soil, plant roots | Nutrient availability, plant growth support | Phosphate solubilization, growth promotion | Vegetables, cereals | [100,101,102] |
| Paenibacillus macerans | Soil, plant roots | Nutrient availability, growth promotion | Produces enzymes, solubilizes phosphorus | Vegetables, cereals | [103,104] |
| Paenibacillus polymyxa | Soil, plant roots | Nutrient solubilization, disease suppression | Nitrogen fixation, phosphate solubilization | Cereals, vegetables | [105,106] |
| Pseudomonas fluorescens | Soil, plant roots, rhizosphere | Disease resistance, improved nutrient acquisition | Produces siderophores, antibiotics, induces systemic resistance | Vegetables, fruits, cereals | [107,108] |
| Pseudomonas putida | Soil, rhizosphere | Enhanced nutrient uptake, disease control | Siderophore production, biocontrol of pathogens | Vegetables, ornamentals | [109,110] |
| Pseudomonas stutzeri | Soil, rhizosphere | Nitrogen fixation, disease suppression | Nitrogen fixation, biocontrol of pathogens | Vegetables, cereals | [111,112,113,114] |
| Rhizobium leguminosarum | Soil, legume roots | Symbiotic nitrogen fixation, plant growth promotion | Nitrogen fixation, nodule formation | Legumes (e.g., peas, beans) | [115,116,117] |
| Streptomyces griseoviridis | Soil, plant roots | Disease control, enhanced plant health | Produces antibiotics, competes with pathogens | Vegetables, ornamentals | [118,119,120] |
| Streptomyces lydicus | Soil, rhizosphere | Disease control, enhanced plant health | Antifungal activity, biocontrol of pathogens | Vegetables, ornamentals | [121,122,123] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



