
Submitted:
10 September 2024
Posted:
11 September 2024
You are already at the latest version
Abstract
Drought and water scarcity are significant factors limiting the development of China's maize industry. Deep sowing, as a method to mitigate drought and preserve soil moisture and seedlings, can effectively mitigate the adverse effects of drought stress on their growth. However, deep sowing can influence the emergence of maize seedlings and has a certain impact on their growth and development. To investigate the physiological and transcriptomic changes in maize inbred lines under deep sowing stress, and the alleviation effects of exogenous hormones, this study investigated the response of mesocotyl in deep-seeding tolerant maize inbred line Qi319 and deep-seeding sensitive maize inbred line Zi330 to deep-seeding stress under exogenous MeJA treatment. The physiological and transcriptomic mechanisms underlying the alleviation of deep-seeding stress by exogenous MeJA were elucidated. Results showed that compared to normal sowing depth, mesocotyl length (MES), mesocotyl and coleoptile length (MESCOL) significantly increased under deep-seeding stress for both inbred lines, while seedling length (SDL) was significantly inhibited. However, after adding 1.5 μmol·L-1 MeJA, MES, MESCOL, and SDL significantly increased compared to deep-seeding stress alone. Exogenous MeJA promoted cell elongation, enhanced mesocotyl length, and improved seedling emergence ability. RNA-Seq sequencing revealed differentially expressed genes (DEGs). Under deep-seeding stress compared to normal sowing depth conditions, 1248 DEGs were identified in Qi319 and 1288 DEGs were identified in Zi330. After adding exogenous MeJA under deep-seeding stress compared to normal sowing depth with exogenous MeJA treatment conditions, 538 DEGs were identified in Qi319 and 2284 DEGs were identified in Zi 330. GO enrichment analysis showed that differentially expressed genes were mainly concentrated in biological processes, cell components and molecular functions. KEGG enrichment analysis showed that differentially expressed genes were mainly concentrated in transport and catabolism, signal transduction, amino acid metabolism and lipid metabolism processes. Compared with the deep-seeded treatment, 12 differentially expressed genes were co-expressed in the two inbred lines under exogenous MeJA treatment, and 10 key genes, including ornithine decarboxylase, terpene synthase 7 and ethylene reaction transcription factor 11, were screened out. Our findings elucidate the molecular mechanism underlying maize's tolerance response to deep seeding and offer valuable insights into candidate gene resources for improving deep seeding tolerance in maize.
Keywords:
Maize
; Exogenous methyl jasmonate
; Deep sowing stress
; Mitigation effect
1. Introduction
Maize (Zea mays L.) is an important crop for both food and feed. It is very important to ensure the high and stable yield of maize for food production safety. Drought and water shortage is one of the most important abiotic stresses that cause global maize yield reduction. Most of the maize planting areas in China are located in arid and semi-arid areas. According to statistics, the annual yield reduction caused by drought and water shortage is about 20%-50%[1,2]. In recent years, in order to solve the problem of grain yield reduction caused by drought, many scholars have also proposed different solutions, mainly including cultivating new drought tolerant varieties[3] and deep sowing seeds[4]. However, because the breeding of new drought tolerant varieties is related to its genetic mechanism, and its drought tolerance is controlled by a variety of micro effect genes, its regulation mechanism is still unclear and it is difficult to be applied on a large scale. Therefore, it is a relatively simple and feasible method to reduce drought stress by deep sowing of maize seeds. At the same time, deep sowing is also an efficient measure for drought resistance and seedling protection of maize and a new way for drought resistance cultivation of maize[5].
At present, the research on deep sowing tolerance of maize mainly focuses on the selection of deep sowing quality and characteristics, while the research on the role of exogenous hormones in alleviating deep sowing stress in the regulation of hypocotyl is less, and the research results are not the same, so a systematic evaluation system cannot be formed. Studies have shown that with the increase of sowing depth, the main reason for maize tolerance to deep sowing is the significant elongation of mesocotyl, and the change of mesocotyl length is closely related to hormone regulation[6]. It was found that the mesocotyl length was significantly correlated with the contents of auxin (IAA) and abscisic acid (ABA)[7]. Although there have been a lot of transcriptome studies on maize tolerance to deep sowing, there are few reports on the mechanism of exogenous hormones regulating mesocotyl growth in response to deep sowing stress.
Methyl jasmonate (MeJA), as a hormone in response to plant damage, is widely used in plant resistance to damage stress, such as plant growth and development, metabolic regulation, disease tolerance and stress resistance[8,9]. Studies have shown that MeJA, as an internal signal transduction molecule, has the characteristics of easy transport, non-ionization and easy membrane penetration [10]. However, the current research on alleviating deep sowing stress by exogenous MeJA has not been reported, and the mechanism of alleviating deep sowing stress is still unclear. In this study, deep sowing tolerant maize inbred line Qi319 and deep sowing sensitive maize inbred line Zi330 were used as materials. The differences of mesocotyl of different inbred lines under deep sowing and exogenous MeJA treatment were studied from aspects of phenotype, cell structure, physiology and biochemistry, and the metabolic regulatory networks and regulatory genes related to deep sowing were studied from transcriptomics. To elucidate the metabolic pathway and key genes of MeJA alleviating deep sowing effect, and reveal the molecular mechanism of deep sowing tolerance related traits in maize, laying a foundation for the exploration of deep sowing tolerance genes and the study of drought resistance mechanism in maize.
2. Results
2.1. Physiological Characteristics of Exogenous MeJA in Alleviating Deep Sowing Stress in Maize Inbred Lines
2.1.1. Morphological Analysis of Exogenous MeJA Alleviating Deep Sowing Stress in Maize Inbred Lines
As can be seen from Table 1, at 3 cm sowing depth, the addition of exogenous hormone MeJA promoted the biomass accumulation of the two maize inbred lines. Compared with CK, the root length, seedling length and seedling fresh weight all reached significant differences. At the sowing depth of 15 cm, the growth of the two maize inbred lines was significantly affected. After the addition of MeJA treatment, the material accumulation of the seedlings of both inbred lines increased in DM treatment. The seedling length (SDL), root length (RL), seedling fresh weight (SDW) and root fresh weight (RW) of deep sowing inbred line Qi319 were significantly increased by 34.26%,13.97%, 7.48% and 17.54%, respectively. The seedling length (SDL), seedling fresh weight (SDW) and root fresh weight (RW) of deep sowing sensitive inbred lines from Zi330 increased by 8.96%, 7.69% and 33.33%, respectively. The results showed that MeJA was the main factor in the enhancement of seed stress resistance under deep sowing. When MeJA was treated with exogenous hormone at the same time, the morphological indexes of the two inbred lines were significantly increased, which significantly promoted the growth of different maize inbred lines, and the increase rate of the inbred lines tolerant to deep sowing was higher than that of the inbred lines sensitive to deep sowing.
It can be seen from Table 2, compared with CK, after the treatment of adding exogenous hormone MeJA, the mesocotyls length (MES), coleoptile length (COL), mesocotyl weight + coleoptile weight (MESW+COLW), and mesocotyl length + coleoptile length (MES+COL) of the two inbred lines in CM treatment and DM treatment were significantly higher than those in CK and DS treatment, indicating that the application of exogenous hormone MeJA can significantly promote the elongation of mesocotyls. Compared with DS, the length of Hypocotyl of deep seeded sensitive inbred line Zi330 increased by 26.51%, while that of deep seeded tolerant inbred line Qi319 increased by 7.5%. It can be seen that MeJA has different effects on the improvement of deep sowing tolerance of different varieties, indicating that the deep sowing of seeds is regulated by both the genetic characteristics of varieties and exogenous hormone MeJA, which shows a cumulative effect. The application of exogenous hormone at the same time of deep sowing can adjust the deep sowing tolerance of maize seeds.
2.1.2. Morphological Analysis of Exogenous MeJA Alleviating Deep Sowing Stress in Maize Inbred Lines
It can be seen from Figure 1, after the addition of exogenous MeJA, the endogenous hormones of Zi330 and Q319 changed significantly, and the endogenous hormone changes were particularly significant under deep sowing conditions. Compared to DS treatment, the growth hormone (IAA) and gibberellin (GA3) in the mesocotyl of deep-sowing sensitive inbred lines increased significantly from Zi330 under DM treatment, suggesting that MeJA may have a synergistic effect with IAA and GA3, while abscisic acid (ABA) and brasinolide (EBR) contents decreased significantly in response to deep-sowing stress. It is suggested that MeJA may have antagonistic regulation with ABA and EBR. At the sowing depth of 3 cm and 15 cm, the endogenous MeJA content in the mesocotyl of the two maize inbred lines decreased to different degrees after MeJA was applied, which may be due to the low content of the optimal hormone for MeJA response to deep sowing of maize. It is speculated that when exogenous MeJA is applied, the content of endogenous MeJA in the mesocotyl of the two maize inbred lines decreases to different degrees. Endogenous MeJA produced by plant mesocotyl can alleviate the damage caused by deep sowing stress. These results indicated that endogenous hormones could stabilize the internal environment balance and promote the growth and development of plants by regulating their own levels under deep sowing conditions.
2.1.3. Cytological Observation of Exogenous MeJA Alleviating Deep Sowing Stress in Maize Inbred Lines
As shown in Figure 2, the mesocotyl of the two maize inbred lines seedlings at 15 cm sowing depth increased significantly compared with 3 cm sowing depth, indicating that the response of maize seedlings to deep sowing stress was closely related to mesocotyl length. The length of mesocotyl cells of the two inbred lines increased after the addition of 1.5 μmol.L-1 exogenous MeJA treatment at the sowing depth of 3 cm and 15 cm, and the length of mesocotyl cells increased most significantly under the treatment of DM. Compared with CK treatment, the mesocotyl cell length of Zi330 and Qi319 increased by 25.98% and 19.69% under CM treatment. Compared with DS, the mesocotyl cell length of Zi330 and Qi319 increased by 46.44% and 31.99% under DM treatment.
2.2. Transcriptome Analysis of Exogenous MeJA Alleviating Deep Sowing Stress under Deep Sowing Stress
2.2.1. Analysis of Sequencing Results
The constructed library was sequenced with the DNBSEQ platform for transcriptome, 24 samples were sequenced, and a total of 31384 genes were detected. Each sample obtained an average of 45.64M original Reads. After filtering and quality control of the original Reads, each sample obtained an average of approximately 44.18M valid data, with a Q20% value range of 96.41%-96.65%. The amount of sample sequencing data is shown in Table 3.
2.2.2. Identification and Functional Analysis of Differentially Expressed Genes
In order to comprehensively study the deep sowing tolerance of maize and the mitigation effect of exogenous MeJA on deep sowing stress of maize, we used FPKM to calculate gene expression, and genes with differencing multiple of gene expression |log2FC|≥1 and P≤0.05 were defined as DEGs. As can be seen in Figure 3A, compared with normal sowing depth, 1248 and 1288 DEGs were identified under deep sowing stress, accounting for 48.79% and 40.06% of the total differential genes respectively, while 1310 and 1927 genes were down-regulated, respectively. They accounted for 51.21% and 59.94% of the total differential genes respectively. Compared with the addition of exogenous MeJA under normal sowing depth, 538 DEGs were up-regulated and 94 DEGs were down-regulated after exogenous MeJA was applied to Qi319 under deep sowing stress. However, 2284 DEGs were up-regulated and 3601 DEGs were down-regulated after the addition of exogenous MeJA under deep-sowing stress, indicating that the deep-sowing tolerance of Zi330 was enhanced by more down-regulated DEGs under exogenous MeJA. Similarly, analysis of the distribution of common and specific differential genes in Venn diagram also fully demonstrated the mitigation of exogenous hormone MeJA on maize inbred lines with different deep-sowing tolerance characteristics. As shown in Figure 3B, 225 different genes were co-expressed between the two inbred lines under deep sowing stress compared with normal sowing depth. There were 42 differential genes that were co-expressed after exogenous MeJA was applied under deep sowing stress, compared with that under normal sowing depth. With distilled water applied under normal sowing depth as control, there were 6 differential genes co-expressed after exogenous MeJA was applied. In contrast, with the application of distilled water at a depth of 15 cm, there were 3 differential genes that were co-expressed after the application of exogenous MeJA (Figure 3C).
2.2.3. GO Analysis of Differentially Expressed Genes
GO functional annotations can be divided into three main categories: Biological processes, Cellular components, and Molecular functions. GO functional analysis of DEGs in 2 maize inbred lines subjected to deep sowing stress was conducted. Under deep sowing stress, the main biological process (BP) of DEGs enrichment in deep-sowing inbred line Qi319 was cellular process, followed by metabolic process, biological regulation; The main enriched cell component (CC) was cell, followed by cell part, organelle and mebrane. The main enriched molecular functions (MF) are catalytic activity, binding, and transporter activity. The term of significant enrichment of DEGs in BP and CC of the deep-sowing sensitive inbred lines from Zi330 was basically the same as that of Qi319. The main enriched MF is binding, catalytic activity and structural molecule activity, which is different from Qi319. It can be seen that the DEGs identified in this study mainly focus on metabolic regulation, cell metabolism, active molecule binding, and transport (Figure 4).
GO functional annotation analysis was carried out on the DEGs identified by two inbreeding lines under the treatment of exogenous MeJA alleviating deep sowing stress (Figure 5). Biological processes, cell components and molecular functions were the main enrichment processes of differentially expressed genes in the two inbred lines treated with exogenous MeJA. The BP enrichment of DEGs identified by deep-sowing inbred line Qi319 was mainly cellular process, followed by metabolic process and biological regulation. CC was mainly enriched in cells, followed by cell parts, organelles and membranes. The main enrichment of MF is catalytic activity, binding activity, transport activity. The BP enriched in DEGs from Zi330 were metabolic process, cellular process and stress response. In addition, the term of significant enrichment of DEGs from Zi330 sensitive inbred lines in CC and MF was basically the same as that of Qi319. However, since Zi330 has more DEGs in the same enrichment term, it indicates that exogenous MeJA mainly enhances the deep sowing tolerance of maize by up-regulating or down-regulating more DEGs.
2.2.4. Pathway Enrichment Analysis of Differentially Expressed Genes
In order to further understand the biological functions of differential genes, pathway enrichment analysis was conducted on differential genes of two maize inbred lines under deep sowing stress. It mainly involves cellular processes, environmental information processing, and genetic information processing, metabolism and organismal systems. Compared to the control group, the significant enrichment pathways noted in the inbred line Qi319 under deep sowing stress mainly include transport and catabolism, global and overview maps, and signal transduction, amino acid metabolism, energy metabolism and environmental adaptation; Among them, the enrichment of carbohydrate metabolism pathway is the most significant (Figure 5A). The significant enrichment pathway noted by the deep seeding sensitive inbred lines from 330 under deep seeding stress, Including transport and catabolism, global and overview atlas, amino acid metabolism, energy metabolism, carbohydrate metabolism, environmental adaptation and other pathways, among which the enrichment of translation pathway is the most significant (Figure 5B). In both inbred lines, multiple enrichment pathways were screened, including transport and catabolism, amino acid metabolism, energy metabolism, carbohydrate metabolism, signal transduction, transcription, global and overview maps.
The pathway enrichment analysis was conducted on the differential genes treated with exogenous MeJA at the sowing depth of 15cm, which mainly involved five branches: cell process, environmental information processing, genetic information processing, metabolism, and biological system. Compared with deep-sowing treatment, the significant enrichment pathways noted in deep-sowing tolerance inbreeding line Qi319 under exogenous MeJA treatment mainly included transport and catabolism, global and overview maps, signal transduction, amino acid metabolism and carbohydrate metabolism. Among them, the enrichment degree of translation pathway was the most significant (Figure 5C). The significant enrichment pathways noted in the deep seeding sensitive inbred line Zi330 include carbohydrate metabolism, amino acid metabolism, energy metabolism, metabolism of cofactor and vitamins, metabolism of terpenoids and polyketides, nucleotide metabolism (Figure 5D). The pathways of carbohydrate metabolism, amino acid metabolism, global and overview maps, lipid metabolism and signal transduction were simultaneously screened in the two inbred lines.
Compared with normal sowing depth, the differential genes whose expression levels were up-regulated and down-regulated in different comparison groups reflected the molecular mechanism of mesocotyl response to deep sowing stress in the two maize inbred lines, while the differential genes whose expression levels were opposite might reflect the reason of deep sowing tolerance of maize inbred line Qi319. Taking deep sowing stress as the control, it was speculated that the differential genes whose expression levels were up-regulated and down-regulated in different comparison groups under exogenous MeJA treatment might reflect the molecular mechanism of the mesocotyl response of the two maize inbred lines to exogenous hormones to alleviate deep sowing stress. Compared with the control, 704 different genes were co-expressed in the two maize inbred lines with different tolerance to deep sowing stress. Using |log2FC|≥2 and P≤0.05 as the criteria, 76 differentially expressed genes were identified. As can be seen from Figure 6, these significantly different genes are mainly involved in biological processes such as plant-pathogen interaction, unsaturated fatty acid biosynthesis, fatty acid metabolism, regulation of photomorphogenesis, and regulation of intima systems. The differentially expressed genes in both inbred lines were up-regulated and down regulated when MeJA was applied under deep sowing stress, which reflected the molecular mechanism of the alleviation effect of exogenous MeJA on maize inbred lines under deep sowing stress. Compared with deep sowing treatment, we used | log2FC | ≥ 2 and P ≤ 0.05 as the criteria to screen the differentially expressed genes in the two inbred lines under exogenous MeJA remission treatment, and obtained 12 differentially expressed genes with significant co-expression. It can be seen from Fig. 6 that these significantly different genes are mainly involved in arginine and proline metabolism, glutathione metabolism, terpene biosynthesis, mitochondrial calcium homeostasis, sesquiterpene biosynthesis, terpene compounds biosynthesis and other biological processes.
Compared with the control, the differentially expressed genes that were up-regulated in the deep sowing tolerant inbred line Qi319 but down-regulated in the deep sowing sensitive inbred line Zi330 under deep sowing stress were screened using |log2FC|≥2 and P≤0.05 as the criteria, and 6 significantly differentially expressed genes were obtained. From Table 5, it can be seen that these significantly different genes are mainly involved in biological processes such as plant hormone signal transduction, amino acid metabolism, alkaloid synthesis, transcriptional regulation, and amine metabolism.
2.2.6. Validation of DEGs by qRT-PCR Analysis
qRT-PCR was used to verify 10 common DEGs of the two inbred lines under deep sowing stress. The results are shown in Figure 7. The relative expression of genes Zm00001d029630, Zm00001d022517, Zm00001d018229, Zm00001d023859, Zm00001d017696, Zm00001d033071, Zm00001d039468, Zm00001d021089, Zm00001d008837 and Zm00001d052759 is consistent with the expression trend of RNA-seq sequencing results of the two inbred lines under deep sowing stress, Based on the above analysis, the expression trends of 10 genes were consistent with the results of RNA-seq sequencing, which fully demonstrated the reliability of our sequencing results.
2.2.7. Gene Co-Expression Network Analysis
WGCNA is a method that can analyze gene expression patterns in multiple samples and identify gene sets (i.e., module modules) with similar expression patterns. To identify co expression patterns among DEGs, we used WGCNA to cluster genes with similar expression patterns into modules. We constructed a co- expression network using expression data of 24 sample genes with FPKM values >1 (Figure 8A). Hierarchical clustering tree analysis was used to identify co-expression modules in WGCNA. Each leaf on the clustering tree represents a gene, and a total of 15 co-expression modules were constructed. From the heat map of the gene co-expression network, it can be seen that the colors between the same modules are darker, while the colors between modules are lighter. This indicates that the correlation between modules is small, and our gene co-expression module clustering results are reliable and can be further analyzed (Figure 8B). Then, we analyzed the relationship between modules and specific traits/phenotypes of plants under normal and deep sowing depth, deep sowing stress, and exogenous hormone treatment. Previous studies have shown that the tolerance of maize to deep sowing is mainly related to the mesocotyl elongation. Therefore, our interest was focused on analyzing modules with which mesocotyl elongation was significantly positively correlated with the royalblue and bisque4 modules (Figure 8C, D).
2.2.8. Analysis of Hub Genes Interaction Network in the Module
To determine specific genes most likely to be important in deep sowing tolerance, we additionally used the expression data to generate a co-expression network. The network was used to identify “hub genes” for treatment groups. This term referred to genes with high connectivity in a gene interaction network. In this study, the five genes with the highest kME values (feature gene connectivity) in the royalblue and bisque4 modules were classified as hub genes. The hub gene and its interacting genes were used to draw a gene co-expression network diagram (Figure 9). In the royalblue module (Figure 9A), the central gene 103640916 is a member of the histone H2A family, which regulates gene expression and cell differentiation by interacting with DNA. The 103628268 gene is a member of the ubiquitin conjugating enzyme E2T (UBE2T) family, which mediates the ubiquitin—proteasome system and regulates gene expression. The genes 100217314, 103638413 and 100274062 are functionally unknown genes. In the bisque4 module (Figure 9B), the central gene 103643355 is a recombinant protein of streptococcus pyogenes serotype M6rpsE, and the primary ribosomal protein S5 is crucial for the 30S ribosome biosynthesis of eukaryotic bacteria. The 103625782 gene, as an effector, can interact with chloroplast proteins, regulate chloroplast signaling pathways, and is a GLKs target gene encoding enzymes in the chlorophyll biosynthesis pathway. GLKs are positive regulatory transcription factors that dominate chloroplast development in plant nuclei. The 100285290 gene is a FK506 binding protein (FKBPs) that plays different roles in many key processes of plant growth, development, and response to abiotic stress. The 542277 gene is a Z-type nucleic acid binding protein 1 that activates receptor interacting protein kinase 3, causing cell death during RNA and DNA viral infections. The 100283885 gene is a functionally unknown gene. Therefore, we speculate that transcriptional regulation, histone, ubiquitin protease, protein binding, and chlorophyll biosynthesis play important roles in maize deep sowing tolerance and the exogenous hormone MeJA in alleviating deep sowing stress.
3. Discussion
3.1. Phenotypic Analysis of Exogenous MeJA to Alleviate Deep Sowing Stress
Drought is one of the major abiotic stresses in China. Increasing the sowing depth of maize can make maize use the deep soil water efficiently, so as to avoid the problem of maize yield reduction caused by drought due to the loss of soil surface water [11,12]. Studies have shown that within a certain range, with the increase of sowing depth, the emergence time will be extended, the emergence rate will be reduced, MES, COL and RL will be increased, and the overall degree of seedlings will be significantly deteriorate[11].In this study, compared with normal sowing depth, MES and MES+COL of maize inbred lines Zi330 and Qi319 were significantly increased (P < 0.05), while SDL was significantly shortened under deep sowing stress. In addition, compared with deep-sowing stress, MES, MES+COL and SDL of maize inbred line Qi319 and Zi330 were significantly increased after the addition of exogenous MeJA with 1.5 μmol·L-1 under deep-sowing stress. This indicates that exogenous MeJA treatment of maize materials with different deep sowing characteristics under deep sowing stress can effectively alleviate the persecution of maize under deep sowing stress.
3.2. Analysis of Relative Hormone Content in Maize Mesocotyl Under Exogenous MeJA Treatment
Studies have shown that under deep sowing stress, the elongation of maize mesocotyl is largely regulated by various plant endogenous hormones such as IAA, GA, CTK, ABA and ETH [13,14]. The contents of GA3 and IAA in the mesocotyl of maize increased when GA3 and UCZ(Gibberellin synthesis inhibitor tenobuzole) were applied at the same time under deep sowing, indicating that when exogenous GA3 was applied to alleviate deep sowing stress, the mesocotyl of maize promoted the elongation of mesocotyl by increasing IAA content. IAA can acidify the mesocotyl epidermal cell wall and relax it, thereby regulating mesocotyl cell elongation [15]. CTK mainly promotes mesocotyl elongation by dividing rice mesocotyl cells [16]. In this study, after exogenous MeJA was added, IAA increased, while ABA and EBR decreased, which may be due to the synergistic effect of EBR and IAA. The increase of IAA content inhibited the synthesis of EBR and led to the decrease of its content. There was no significant difference in EBR content after exogenous MeJA was applied under deep sowing stress, indicating that exogenous MeJA promoted the elongation of mesocotyl, mainly promoting the elongation or enlargement of cell volume under deep sowing stress. Different from this study, Wu et al. [17] believed that the elongation of rice mesocotyl was mainly due to the promotion of cell division by ABA, and ABA and GA had a positive superposition effect on mesocotyl elongation. Therefore, whether the regulation mechanism of ABA on mesocotyl elongation of different crops is the same needs to be further studied and demonstrated.
3.3. Cytological Analysis of Maize Mesocotyl Under Exogenous MeJA Treatment
Mesocotyl elongation is cytologically divided into an increase in the number of cells and an increase in the volume of cells. Zhao et al. [18] showed that the mesocotyl of maize alleviated deep sowing stress mainly by promoting the increase of mesocotyl cell length, rather than the elongation of mesocotyl caused by the change of cell number, which was consistent with the results of this study. The results of this study showed that the cytological reason for promoting maize mesocotyl under deep sowing conditions was mainly the increase of cell volume rather than the increase of cell number, and exogenous MeJA could promote cell elongation to increase mesocotyl length and enhance seedling exhume ability. It was speculated that exogenous MeJA induced changes of different hormone contents in the mesocotyl of maize seedlings, thereby regulating cell growth. Moreover, there were differences in the relief effects of exogenous hormones between deep-sowing tolerant inbred lines and deep-sowing sensitive inbred lines. Therefore, in actual production, the particularity of materials should be fully considered if exogenous hormones should be added to alleviate deep sowing stress.
3.4. Analysis of Key Differential Expressed Genes Under Deep Sowing Stress and Exogenous MeJA Mitigation Treatment
The statistical analysis of differential genes showed that 704 differential genes were co-expressed in the mesocotyl of the two inbred lines under deep sowing stress, and 276 differential genes were up-expressed in the deep sowing tolerant inbred line Qi319 but down-expressed in the deep sowing sensitive inbred line Qi330. Using | log2FC | ≥ 2 and P ≤ 0.05 as criteria, 76 differentially expressed genes with co-upregulation were screened, and 6 differentially expressed genes with co-upregulation were upregulated in deep-sowing tolerant inbred line Qi319 but down-expressed in deep-sowing sensitive inbred line Zi330. These significant genes are mainly involved in unsaturated fat acid biosynthesis, fatty acid metabolism, endometrial system regulation, plant hormone signal transduction, amino acid metabolism and other processes.
Lignin is a kind of aromaticity polymer, which is mainly involved in the secondary wall of plant cells. When the lignin content in plant tissue is too high, the cell elongation and growth of the tissue will be inhibited, mainly because of the low cell wall relaxation caused by the high lignin content [19]. Some studies also believe that peroxidase (POD) is induced by hydrogen peroxide (H2O2) and synthesizes too much lignin, which leads to the hardening of cell wall and the reduction of relaxation, resulting in the inhibition of cell elongation and growth [20,21]. In this process, reducing nicotinamide adenine dinucleotide phosphate oxidase (NADPH), POD and polyamine oxidase (PAO) are the three main sources of H2O2 [22,23]. Genes involved in the redox process, such as GRMZM2G359298, were up-regulated in both inbred lines after treatment with exogenous MeJA. It is suggesting that peroxidase may be induced by exogenous MeJA, thereby eliminating the accumulation of H2O2 to reduce the oxidation and polymerization of monosaccharide alcohols in the cell wall into lignin and reducing the degree of cell elongation inhibition. These genes may be responsible for cell wall synthesis and cell elongation, suggesting that they may play an important role in mesocotyl elongation of maize under deep sowing stress induced by MeJA.
Most terpenoids in plants belong to secondary metabolic compounds, which play a positive role in the growth and development of plants, physiological functions and interactions between plants and environmental factors. Plant hormones such as gibberellin, abscisic acid, brassinolactone and strigolactone (SLs) are also terpenoid derivatives [24]. At present, terpenoids are mainly studied in maize salt stress and resistance to disease, while few are studied under deep sowing stress. In this study, transcripome sequencing was performed on maize inbred lines with different deep sowing tolerance characteristics, and it was found that some genes were enriched in terpenoid biosynthesis under deep sowing conditions when MeJA was applied. The synthesis of terpenoids may play an important role in alleviating the adaptability of maize under deep seeding stress.
At present, the interaction between hormones related to the elongation of maize hypocotyl cells is still unclear. Zhao and Wang [25] believe that IAA mainly regulates the elongation of maize hypocotyls by increasing the synthesis and transportation of IAA, and found that increasing IAA concentration under deep sowing conditions can effectively promote the elongation of maize mesocotyls. In addition, under the condition of deep sowing, auxin binding proteins actively participate in promoting the elongation of maize mesocotyl [26]. Kutschera etal. [27] applied exogenous IAA treatment to maize mesocotyls and coleoptiles cultured in vitro, and found that both tissues were significantly elongated after treatment. They also found that BR and IAA promoted cell elongation and growth by acidification and relaxation of the epidermal cell wall. Pan et al. [28] showed that exogenous GA3 also promoted the elongation of mesocotyls under deep sowing conditions. In addition, the "auxin elongation hypothesis" has been confirmed and expanded. It is reported that auxin binding protein 1 and IAA receptor are involved in the maize auxin/light signal network [29]. Compared with deep seeding treatment, differential genes that were upregulated in the hypocotyls of two maize inbred lines under exogenous MeJA treatment were screened using |log2FC|≥2 and P≤0.05 as the standard. Twelve differential genes were identified, ten key genes including ornithine decarboxylase gene, terpene synthase 7 gene, ethylene response transcription factor 11 gene, calcium one-way transporter 4 mitochondrial gene, protein exordium gene, peroxidase 64 gene, adenine nucleotide α hydrolase like superfamily protein gene, glutamine decarboxylase 1 gene, resistance protein rps2 gene and extensin like protein gene were preliminarily screened out, which laid the foundation for functional identification and cloning of related genes in the later stage. Among them, the expression of LOC103632960 gene is up-regulated, which is ethylene response transcription factor 11. Ethylene response factor (ERF) is an important transcription factor, which is involved in the response of plants to ethylene and the regulation of plant growth and development. ERF family members widely exist in various tissues of plants and play an important role in histogenesis, flowering regulation, stress resistance response and other aspects [30]. Studies have shown that the overexpression of tomato terf2/leerf2 gene in tobacco and tomato regulates the induced expression of cold resistance related genes and enhances the cold resistance of transgenic plants [31]. The overexpression of ptaerf194 gene in poplar improves the resistance of plants to drought stress by improving water use efficiency and limiting water loss [32]. The expression of LOC10363309 gene was up-regulated, which was mitochondrial calcium one-way transporter 4. Mitochondrial calcium one-way transporter MCU was a highly selective calcium channel, located in the inner membrane of mitochondria, and was the main medium for calcium ions to enter the mitochondrial matrix. Duan et al. [33] found that mitochondrial calcium one-way transporter ghmcu4 negatively regulates cell elongation, inhibits ghmcu4 gene expression, significantly increases ca2+ content, activates calcium signaling pathway, further promotes H2O2 accumulation, and enhances plant physiological and biochemical metabolism, thereby promoting plant cell growth and fiber elongation. The expression of LOC107546763 gene was up-regulated, which was an extensin like protein. Extensin was the main structural protein in plant cell wall, and it was a glycoprotein rich in hydroxyproline. It widely existed in the cell wall of various plants, accounting for 1% to 15% of the primary wall dry weight in dicotyledons. It plays an important role in plant growth and development. Its activity affects plant morphogenesis and growth and development, and plays an important role in cell wall formation, cell elongation, stress resistance and other aspects [34]. In this study, the cell length of MES, mescol, SDL and mesocotyl tissue of the two inbred lines increased after exogenous MeJA treatment, indicating that exogenous MeJA can promote cell elongation, increase the length of mesocotyl, and improve the ability of seedling emergence. Therefore, it is speculated that these genes can be induced to express after treatment with exogenous hormone MeJA, and may also be involved in the regulation of mesodermal elongation under deep seeding stress.
4. Materials and Methods
4.1. Experiment Materials
The deep sowing tolerant maize inbred line Qi319 and deep sowing sensitive maize inbred line Zi330 were provided by the maize Research Group of Gansu Agricultural University. According to the previous results of our research group, the sowing depth of deep sowing stress was 15 cm, and the concentration of MeJA was 1.5 μmol·L-1.
MeJA solution: Methyl Jasmonate (C13H20O3) raw powder was dissolved in distilled water and prepared into 100.00 μmol·L-1 mother liquor, diluted to 1.50 μmol·L-1, CK (distilled water).
4.2. Experiment method
4.2.1. Seedling culture and treatment
Select plump, uniform and undamaged maize seeds, disinfect them with 0.5% NaClO solution for 10 min, rinsed 3-5 times with ddH2O, and soaked the seeds in distilled water for 12 h. After water was blotted out with sterilized filter paper, the seeds were sown in PVC tubes (17 cm in diameter and 50 cm in height) filled with sterilized vermiculite and distilled water in the ratio of 5 g:1 mL, and the bottom of the tubes were sealed with nylon mesh. Ten seeds were sown per PVC tube and 6 replications per treatment. Seeds were covered with 3 cm of soil for the normal sown control and 15 cm of soil for the deep sowing stress. Cultivate in a dark incubator at a temperature of 25/20°C during the day/night, and water each PVC pipe with 50 mL of the corresponding treatment solution every 2 days. Measurements of indices related to deep sowing tolerance were made 12 days after germination. In addition, the concentration of exogenous hormone treatment was the best concentration selected in the early stage. Three biological replicates were set for each treatment.
Experimental treatment: Treatment 1 (CK): 3cm sowing depth + distilled water treatment; Treatment 2 (CM): 3cm sowing depth +1.5μmol·L-1 MeJA treatment; Treatment 3 (DS): 15cm sowing depth + distilled water treatment; Treatment 4 (DM): 15cm sowing depth +1.5 μmol·L-1 MeJA treatment.
4.2.2. Determination of Related Indexes of Deep Sowing
The determination of phenotypic traits follows the method of Peng Yunling et al. [35]: Mesocotyl length (MES), coleoptile length (COL), seedling length (SDL), root length (RL), seedling fresh weight (SDW), root fresh weight (RW), mesocotyl weight (MESW) and coleoptile weight(COLW) were measured with scale and analytical balance after 12 days of seed germination. Take 10 plants to calculate the average value. The paraffin section technique with safranin-fixed green staining was used to observe the histological structure of the middle part of maize mesocotyl [36]. The extraction method of endogenous hormones is based on the method of Wang et al. [37]. The technique was repeated 3 times by liquid chromatography-mass spectrometry (HPLC-MS).
4.2.3. Transcriptomic analysis of deep seeding stress in maize
After 12 days of seed germination, the vermiculite attached to the corn seedlings was washed, the residual water on the surface was sucked up, the mesocotyl was removed and placed in a 5 mL condensing tube, and the seeds were frozen in liquid nitrogen and stored at -80℃.
RNA extraction, library construction, and sequencing: Total RNA was isolated and purified from 24 samples following the TRIzol (Invitrogen, CA, USA) reagent procedure. RNA-seq library was constructed with total RNA of each treated sample whose purity met the criteria for building the library, which was completed by BGI.
Quality assessment of sequencing results: clean reads were obtained by filtering the raw reads obtained by sequencing, and subsequent analysis was based on clean reads. The filtering software SOAPnuke independently developed by BTU was used for filtering. The specific steps are as follows: 1) Remove reads containing adapter (adapter contamination); 2) reads with N content greater than 5% were removed; 3) Low-quality reads were removed (reads with a quality score of less than 15 bases were defined as those with a total number of bases greater than 20%). The filtered Clean Reads are saved in FASTQ format. Subsequently, each sample was obtained clean reads and compared with the fourth version of the B73 maize reference genome using HISAT [38] software (GCF_000005005.2_B73_RefGen_v4). RSEM software was used to detect gene expression levels.
4.2.4. Analysis of Differentially Expressed Genes
The method of Audic S et al. [39] was adopted to screen DEGs. DEGs were screened using | log2Fold change (FC) | ≥ 1 and FDR ≤ 0.001 as criteria among different comparison groups. WEGO software [40] was used to classify the functional notes of selected DEGs in the GO database. Based on KEGG pathways significant enrichment analysis (said 49-year-old kyoko Encyclopedia of Genes and Genomes) public database (HTTP:/iwww. Genome.jp/keg/pathway.html), using hypergeometric inspection, The pathway of significant enrichment was identified. Pathways with Q value ≤ 0.05 were defined as those that were significantly enriched in DEGs. Significant enrichment of pathway can identify the most important biochemical metabolic pathways and signal transduction pathways involved in DEGs.
4.2.5. Weighted Gene Co-Expression Network Analysis (WGCNA)
Using the gene expression data obtained by RNA-seq, the FPKM threshold is set to 1, the module similarity threshold is 0.25, the minimum number of genes in the module is 30, and the co-expressed gene module is constructed using the WGCNA R package. The co-expression modules were associated with phenotypic traits, and genes with kME > 0.5 were selected as module members (kME is the characteristic gene connectivity for screening hub genes). Cytoscape_v3.9.1 software was used to visualize the gene interaction network of the core module. Finally, the hub genes in the core modules were determined according to the kME value and gene connectivity [41].
4.3. qRT-PCR of Differentially Expressed Genes
In order to ensure the accuracy and reliability of the experimental results, we used total RNA from each treatment material for library construction. The cDNA was reverse transcribed with RNA simple total RNA Kit (Tiangen, Shanghai, China). Ten DEGs were randomly selected from two inbred lines under deep sowing stress, and their specific primers were designed by Primer-BLAST on NCBI. qPCR amplification was performed on quantum Studio 5 real-time PCR system (Thermo Fisher Scientific, MA, United States) using super real premix plus (SYBR Green) (Tiangen, Shanghai, China). Each treatment had three techniques replicates. In addition, each real time PCR was performed at 20 µL. The reaction volume included 10 µL SuperReal PreMix Plus, 6 µL ddH2O, 0.8 µL forward primer (10 µmol/L), 0.8 µL reverse primer, 0.4 µL Rox reference dye and 2 µL template cDNA. The amplification procedure was described with reference to Chen et al. [42]. Using the actin gene as the internal reference gene, the gene expression level was analyzed using the 2−∆∆CT calculation method.
4.4. Statistics Analysis of Data
Statistical analysis and plotting of the data were conducted using Microsoft Excel 2010, PCA analysis was performed using IBM SPSS Statistics 26, and slice cell length measurements were performed using Image Pro Plus 6.0. The mean ± standard deviation (SE) represents the experimental results.
5. Conclusions
Based on the above analysis, MES, MESCOL and SDL of the two inbred lines were significantly increased after treatment with exogenous MeJA under deep sowing stress, indicating that exogenous MeJA can promote cell elongation, increase the length of mesodermal axis and improve the ability of seedling emergence. We established the following molecular models to explain the response mechanism of two different inbred lines to deep sowing stress and the mechanism of exogenous MeJA alleviating deep sowing stress in Maize (Figure 10). Compared with the deep sowing treatment, exogenous MeJA alleviated the deep sowing stress of maize mainly through transportation and catabolism, signal transduction, amino acid metabolism, lipid metabolism, translation, intimal system regulation, terpene biosynthesis and other pathways. After the addition of exogenous MeJA mitigation treatment, there were 12 significantly differentially expressed genes in the two inbred lines, and 10 key genes, such as ornithine decarboxylase, terpene synthase 7 and ethylene response transcription factor 11, were screened out. By regulating the elongation of hypocotyls under deep sowing stress, the deep sowing tolerance of maize was enhanced. The results laid a foundation for the candidate gene resources for the improvement of deep sowing tolerance of maize.
Author Contributions
YP and FW designed the experiments. XY, GZ and ZF performed the experiments. XY, ZF and FW analyzed the data. FW, XY, ZF and GZ wrote the manuscript. All authors read and approved the final manuscript.
Funding
This research was supported by the industrial support plan for National level scientific research and innovation platform project, China (No. 2024CXPT-01), the National Natural Science Foundation project, China (No. 32060487), and the Key Project of Natural Science Foundation of Gansu Province (No. 23JRRA1405).
Data Availability Statement
The datasets and materials used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Wang C.T.; Li S.K. Assessment of limiting factors and techniques prioritization for maize production in China.Scientia Agricultura Sinica, 2010, 43(6): 1136-1146. [CrossRef]
- Sun, C.X.; Shen X.Y. Research progress on identification indicators and quantitative analysis methods for crop drought resistance.Chinese Journal of Agriculture, 2002, 18 (1): 49-51.
- Avramova, V.; Abdelgawad, H.; Vasileva, I.; Petrova, A.S.; Holek, A., Mariën, J., Asard, H., Beemster, G.T.S. High antioxidant activity facilitates maintenance of cell division in leaves of drought tolerant maize hybrids. Frontiers in Plant Science, 2017(8):84-99. [CrossRef]
- Ribaut, J.M.; Hoisington, D. A.; Deutsch, J. A.; Jiang, C.; Gonzalez-de-Leon, D. Identification of quantitative trait loci under drought conditions in tropical maize. 1. Flowering parameters and the anthesis-silking interval. Theoretical and Applied Genetics, 1996, 92: 905-914.
- Wang, Y.T.; Zhou, Y.F.; Li, F.X.; Yi B.; Bai, W.; Yan, T.; Xu, W.J.; Gao, M.C.; Huang, R.D. Identification and classification of drought resistance of sorghum varieties during germination period based on principal component analysis and SOM clustering analysis. Journal of Crops, 2014, 40 (01): 110-121. [CrossRef]
- Zhao, X.Q.; Zhong, Y. Research progress on the physiological response mechanism and molecular genetic mechanism of maize seeds to deep sowing stress. Molecular Plant Breeding, 2021, 19 (07): 2381-2390. [CrossRef]
- Li L. The regulatory mechanism of hypocotyl elongation and growth in weedy rice. Shenyang Agricultural University, 2013.
- Xue, R.H.; Jin, S.A. Methyl jasmonate: an important plant signal transduction molecule. Biotechnology Communications, 2006,17 (06): 985-988.
- Swiatck, A.; Dongen, W. V.; Onckelen, H.V. Metabolic fate of jasmonates in tobacco bright yellow-2 cells. Plant Physiol, 2004, 135(1): 161. DOI:10.2307/4281736.
- Shi, Y.M.; Tao, Y.W.; Qin, W.Y.; Fei, X.N. The effects of low temperature and plant growth regulators on the flowering of vanilla. Journal of Horticulture, 1997, 24 (2): 185-188.
- Du, Z.M.; Hao, J.S.; Zhou, J.; Li, H.X.; Gao, Q.Y.; Cui, H.B.; Zhen, X.B.; Ma, C.; Liang, J. N. Field in situ remediation of copper and cadmium compound polluted soil with four improvers. Journal of Soil Science, 2012, 49 (3): 508-517.
- Gao, Y. J.; Yang, J.L.; Chen, L.; Zhu, G.; Zhe, X.Y.; Liu, W.; Li, S.X. Soil water use status of dry land winter wheat under different cultivation modes, Nitrogen Application Rates, and Planting Density. Agricultural Research in Arid Areas, 2007, 25 (3): 45-50.
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant, Cell & Environment, 2010, 33(4): 453-467. [CrossRef]
- Kutschera, U.; Wang, Z.Y. Growth-limiting proteins in maize coleoptiles and the auxin brassinosteroid hypothesis of mesocotyl elongation. Protoplasma, 2016, 253(1): 3-14. [CrossRef]
- Hu, Z.; Yamauchi, T.; Yang, J.; Yusuke, J.; Tomoko, T.M.; Hiroaki, I.; Itsuro, T.; Yoshiaki, N.; Nobuhiro, T.; Shinjiro, Y. Strigolactone and cytokinin act antagonistically in regulating rice mesocotyl elongation in darkness[J]. Plant & cell physiology, 2014, 55(1): 30-41. [CrossRef]
- Liang, Q.; Wang, C.; Ma, D.R.; Li, L.; Cui, Z.B.; Wang, X.X.; Qian, Q.; Cai, B.D.; Feng, Y.Q.; Chen, W.F. Cortical microtubule disorganized related to an endogenous gibberellin increase plays an important role in rice mesocotyl elongation. Plant Biotechnology, 2016, 33(2): 59-69. [CrossRef]
- Wu, S.Q.; Ding, R.; Li X.S. Effects of gibberellin and Abscisic acid on the elongation and growth of mesocotyl in etiolated seedlings of black rice. Amino Acids and Biological Resources, 2002, (03): 44-56. [CrossRef]
- Zhao, G.W.; Ma P.; Wang J.H.; Wang, G.Y. Identification of tolerance to deep sowing and physiological response to deep sowing stress in different maize inbred lines. Journal of Maize Sciences, 2009, 17 (05): 9-13. [CrossRef]
- Zhou, D.B. Plant hormone and Cytoskeleton alignment. Communication of Plant physiology, 2005 (02): 224-228.
- Cosgrove, D.J. Expansive growth of plant cell walls. Plant physiology and biochemistry: PPB, 2000, 38(1-2): 109-124.
- Karpinska, B.; Karlsson, M.; Schinkel, H.; Streller, S.; Süss, K.H.; Melzer, M.; Wingsle, G. A novel superoxide dismutase with a high isoelectric point in higher plants. expression, regulation, and protein localization. Plant physiology, 2001, 126(4): 1668-1677. www.plantphysiol.org © 2001 American Society of Plant Biologists.
- Jiang, M.; Zhang, J. Involvement of plasma-membrane NADPH oxidase in abscisic acid- and water stress-induced antioxidant defense in leaves of maize seedlings. Planta, 2002, 215(6):1022-1030. [CrossRef]
- Frahry, G.; Schopfer, P. NADH-stimulated, cyanide-resistant superoxide production in maize coleoptiles analyzed with a tetrazolium-based assay. Planta, 2001, 212(2): 175-183. [CrossRef]
- Yoda, H.; Yamaguchi, Y.; Sano, H. Induction of hypersensitive cell death by hydrogen peroxide produced through polyamine degradation in tobacco plants. Plant physiology, 2003, 132(4): 1973-1981. [CrossRef]
- Wang, L.J.; Fang, X.; Yang, C.Q.; Li, J.W.; Chen, X.Y. Secondary metabolism and regulation of terpenoids in plants. Chinese Science: Life Sciences, 2013, 43 (12): 1030-1046.
- Zhao,G.; Wang, J. Effect of auxin on mesocotyl elongation of dark-grown maize under different seeding depths. Russian Journal of Plant Physiology, 2010, 57(1): 79-86. [CrossRef]
- Kutschera, U.; Khanna, R. Auxin action in developing maize coleoptiles: challenges and open questions. Plant signaling & behavior, 2020,15(6). [CrossRef]
- Pan, B.; Zhong, T.; Zhao, G. Promoting deep-sowing germinability of corn (Zea mays) by seed soaking with gibberellic acid[J]. Archives of Agronomy and Soil Science, 2017, 63(9): 1314-1323. [CrossRef]
- Fan, M.; Bai, M.Y.; Kim, J.G.; Wang, T.; Eunkyoo, O.; Chen, L.; Ho Park, C.; Seung-Hyun, S.; Kim, S.K.; Mudgett, M. B.; Wang, Z.Y. The bHLH transcription factor HBI1 mediates the trade-off between growth and pathogen-associated molecular pattern-triggered immunityin Arabidopsis. The Plant cell, 2014, 26(2): 828-841. [CrossRef]
- Wan, F.X.; Ye, K.J.; Wei, Q.T.; Zhao, Z.J.; Zhang, Y.H.; Wang, X.H. Cloning and expression analysis of ERF transcription factor in eggplant. Molecular Plant Breeding, 2024. https://link.cnki.net/urlid/46.1068.S.20240518.2018.002.
- Zhang, Z.; Huang, R. Enhanced tolerance to freezing in tobacco and tomato overexpressing transcription factor TERF2/LeERF2 is modulated by ethylene biosynthesis. Plant Mol Biol, 2010, 73(3): 241-249. [CrossRef]
- Wang, S.J.; Fan, Y.; Du, S.H.; Zhao, K.; Liu, Q.; Yao, W.J.; Zheng, T.C.; Han, Y.Z. PtaERF194 inhibits plant growth and enhances drought tolerance in poplar. Tree Physiology, 2022, 42(8): 1678-1692. [CrossRef]
- Duan, Y.J.; Shang, X.G.; Tian, R.P.; Li, W.X.; Song, X.H.; Zhang, D.Y.; Guo, W.Z. Suppressing a mitochondrial calcium uniporter activates the calcium signaling pathway and promotes cell elongation in cotton. The Crop Journal, 2024, 12: 411-421. [CrossRef]
- Humphrey, T.V.; Haasen, K.E.; Aldea-Brydges, M.G.; Sun, H.; Zayed, Y.; Indriolo, E.; Goring, D.R. PERK-KIPK-KCBP signalling negatively regulates root growth in Arabidopsis thaliana. J Exp Bot, 2015, 66 (1): 71-83. [CrossRef]
- Peng, Y.L.; Yang, F.L.; Zhao, X.Q.; Ren, X.W. Analysis of differences in deep sowing tolerance among different maize inbred lines. Agricultural Research in Arid Areas, 2014, 32 (01): 25-33+51.
- Tian, C.X.; Zhang, Y.M.; Ma, H.L. Preliminary study on the method of making embryonic structure paraffin slices of poa pratensis. Grassland Science, 2013, 30 (12): 1980-1986.
- Wang, S.P.; Ruan, X.F. Isolation and purification of four plant hormone. Communication of Plant physiology, 1987, (05): 50-53. [CrossRef]
- Shahzad, A.; Gul, h.; Ahsan,M.; Wang, D.P.; Fahad, S. Comparative genetic evaluation of maize imported lines at seeding and maturity stages under drought stress. Journal of Plant Growth Regulation, 2023, (42): 989-1005. [CrossRef]
- Audio, S.; Claverie, J.M. The significance of digital gene expression profiles. Genome Research, 1997, 7 (10): 986-995.
- Ye, J.; Fang, L.; Zheng, H.K.; Zhang, Y.; Chen, J.; Zhang, Z.J.; Wang, J.; Li, S.T.; i, R.Q.; Bolund, L.; Wang, J. WEGO: a web tool for plotting GO annotations. Nuclear Acids Research, 2006, 34 (2): 293-297.
- Yao, Y.; Xiong, E.; Qu, X.; Li, J.; Liu, H.; Quan, L.; Lu, W.; Zhu, X.; Chen, M.; Li, K.; et al. WGCNA and transcriptome proffling reveal hub genes for key development stage seed size/oil content between wild and cultivated soybean. BMC Genom.2023, 24, 494. [CrossRef]
- Li, S.T.; Chen, H.; Hou, Z.; Li Y.; Yang C.L.; Wang D.J.; Song C.P. Screening of abiotic stress-responsive cotton genes using a cotton full length cDNA overexpressing Arabidopsis library. Journal of Integrative Plant Biology, 2020, 62(07): 998-1016. [CrossRef]
Figure 1.
Effects of exogenous MeJA on endogenous hormones of maize inbred lines seedlings under deep sowing stress. CK: Distilled water treatment under 3cm sowing depth; CM:3cm sowing depth with 1.5 μmol·L-1 exogenous MeJA treatment; DS: Distilled water treatment under 15cm sowing depth; DM: 15cm sowing depth with 1.5 μmol·L-1 exogenous MeJA treatment.
Figure 1.
Effects of exogenous MeJA on endogenous hormones of maize inbred lines seedlings under deep sowing stress. CK: Distilled water treatment under 3cm sowing depth; CM:3cm sowing depth with 1.5 μmol·L-1 exogenous MeJA treatment; DS: Distilled water treatment under 15cm sowing depth; DM: 15cm sowing depth with 1.5 μmol·L-1 exogenous MeJA treatment.
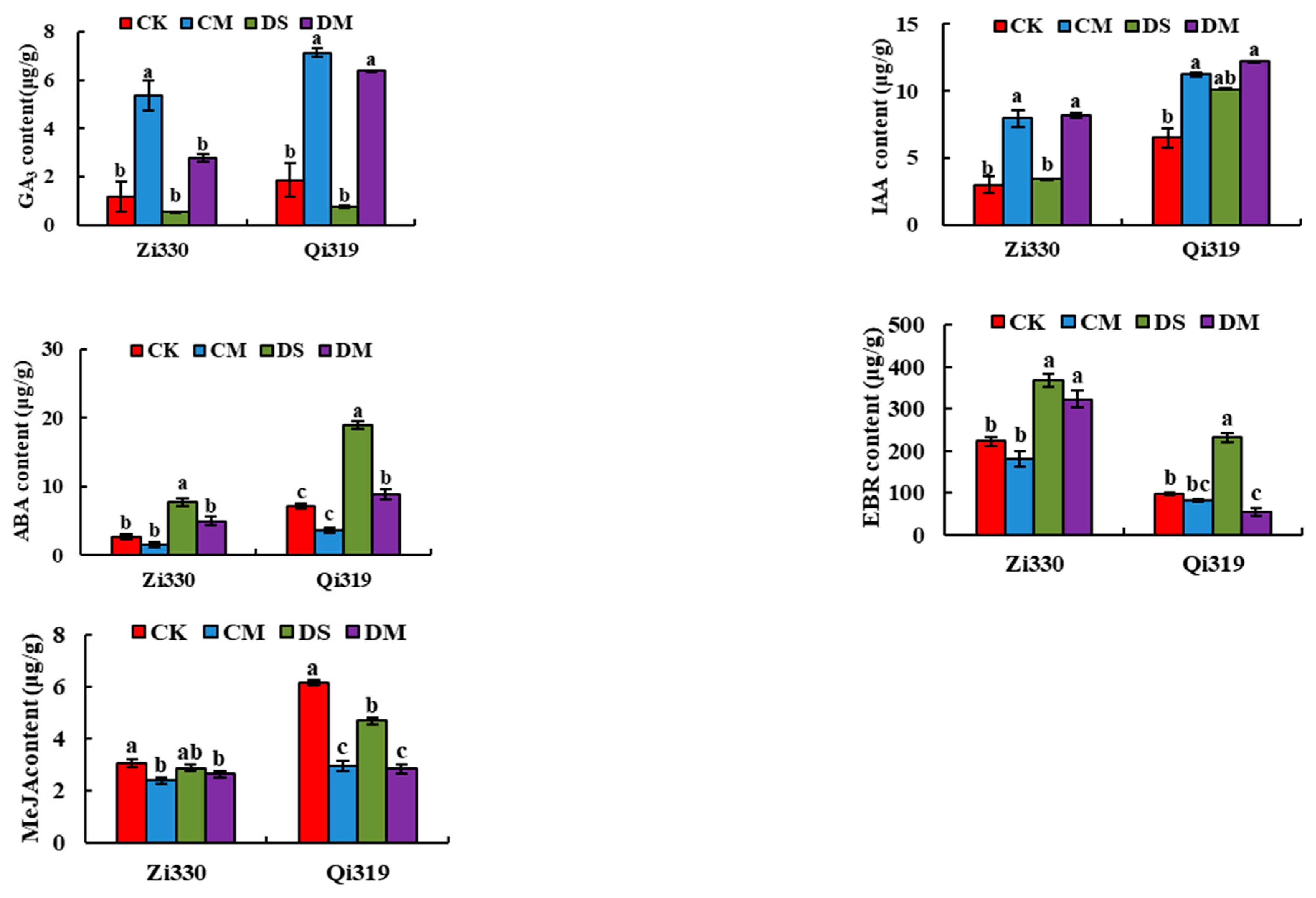
Figure 2.
Longitudinal structure and cell length of mesocotyl cells treated by exogenous MeJA under deep sowing stress of maize inbred lines.A:3 cm sowing depth+distilled water treatment (Qi319); B:3 cm sowing depth+MeJA treatment (Qi319); C: 15 cm sowing depth+distilled water treatment (Qi319); D:15 cm sowing depth+MeJA treatment (Qi319); E:3 cm sowing depth+distilled water treatment (Zi330); F: 3 cm seeding depth+MeJA treatment (Zi330); G: 15 cm sowing depth+distilled water treatment (Zi330); H: 15 cm seeding depth+MeJA treatment (Zi330); I: Cell length of mesocotyl. The treatments and abbreviations are the same as those given in Figure 1.
Figure 2.
Longitudinal structure and cell length of mesocotyl cells treated by exogenous MeJA under deep sowing stress of maize inbred lines.A:3 cm sowing depth+distilled water treatment (Qi319); B:3 cm sowing depth+MeJA treatment (Qi319); C: 15 cm sowing depth+distilled water treatment (Qi319); D:15 cm sowing depth+MeJA treatment (Qi319); E:3 cm sowing depth+distilled water treatment (Zi330); F: 3 cm seeding depth+MeJA treatment (Zi330); G: 15 cm sowing depth+distilled water treatment (Zi330); H: 15 cm seeding depth+MeJA treatment (Zi330); I: Cell length of mesocotyl. The treatments and abbreviations are the same as those given in Figure 1.
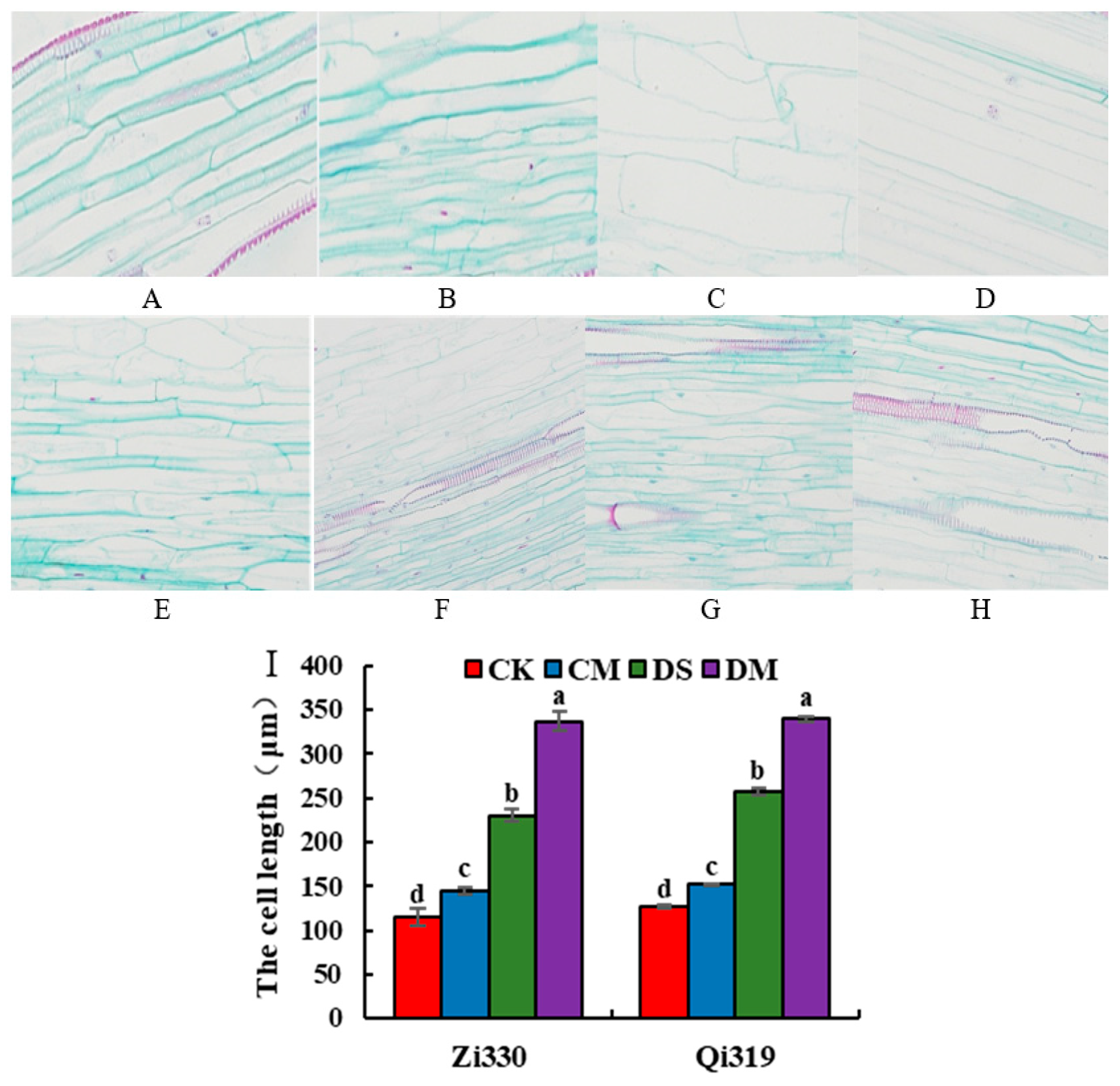
Figure 3.
Differential gene expression analysis. A: Number distribution of down-regulated DEGs in different comparison groups; B, C: Veen diagram analysis of normal sowing depth, deep sowing stress, normal sowing depth and deep sowing stress with exogenous MeJA. Qi_CK3: Qi319 under 3cm sowing depth of distilled water treatment; Qi_CK15: Qi319 distilled water treatment at 15 cm sowing depth;Qi_T3: Qi319 under 3cm sowing depth MeJA treatment; Qi_T15: Qi319 under 15cm sowing depth MeJA treatment; Zi_CK3: distilled water treatment Zi330 at 3cm sowing depth; Zi_CK15: distilled water treatment Zi330 at 15cm sowing depth; Zi_T3: MeJA treatment Zi330 at 3cm sowing depth; Zi_T15: MeJA treatment since Zi330 at 15cm sowing depth.
Figure 3.
Differential gene expression analysis. A: Number distribution of down-regulated DEGs in different comparison groups; B, C: Veen diagram analysis of normal sowing depth, deep sowing stress, normal sowing depth and deep sowing stress with exogenous MeJA. Qi_CK3: Qi319 under 3cm sowing depth of distilled water treatment; Qi_CK15: Qi319 distilled water treatment at 15 cm sowing depth;Qi_T3: Qi319 under 3cm sowing depth MeJA treatment; Qi_T15: Qi319 under 15cm sowing depth MeJA treatment; Zi_CK3: distilled water treatment Zi330 at 3cm sowing depth; Zi_CK15: distilled water treatment Zi330 at 15cm sowing depth; Zi_T3: MeJA treatment Zi330 at 3cm sowing depth; Zi_T15: MeJA treatment since Zi330 at 15cm sowing depth.
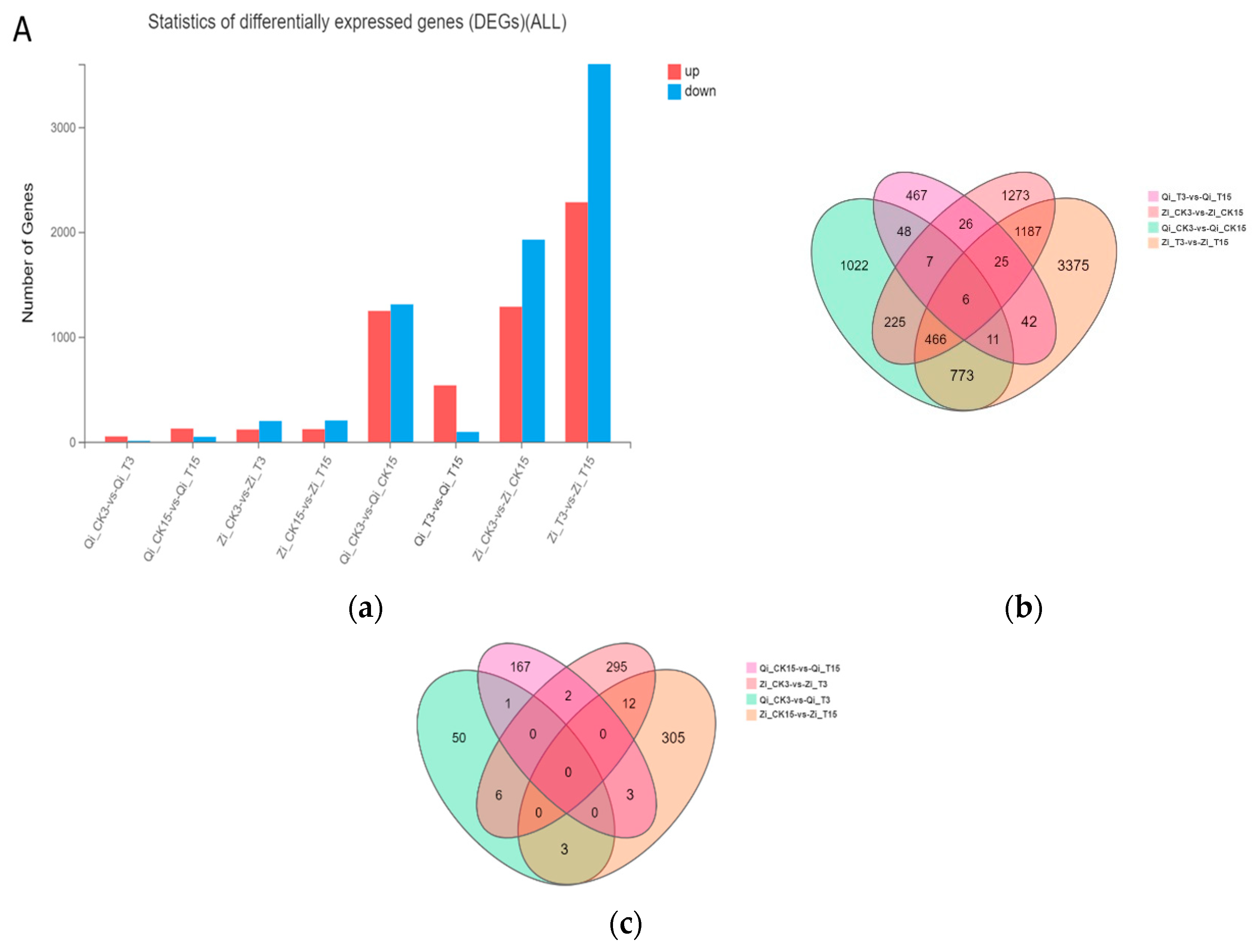
Figure 4.
GO analysis of two inbred lines in different comparison groups. A: GO analysis of maize inbred line Qi319 under normal sowing depth and deep sowing stress; B: GO analysis of maize inbred line Zi330 under normal sowing depth and deep sowing stress. C: GO analysis of maize inbred line Qi319 after adding exogenous MeJA under deep seeding stress; D: GO analysis of maize inbred line Zi330 after adding exogenous MeJA under deep seeding stress. The treatments and abbreviations are the same as those given in Figure 1.
Figure 4.
GO analysis of two inbred lines in different comparison groups. A: GO analysis of maize inbred line Qi319 under normal sowing depth and deep sowing stress; B: GO analysis of maize inbred line Zi330 under normal sowing depth and deep sowing stress. C: GO analysis of maize inbred line Qi319 after adding exogenous MeJA under deep seeding stress; D: GO analysis of maize inbred line Zi330 after adding exogenous MeJA under deep seeding stress. The treatments and abbreviations are the same as those given in Figure 1.
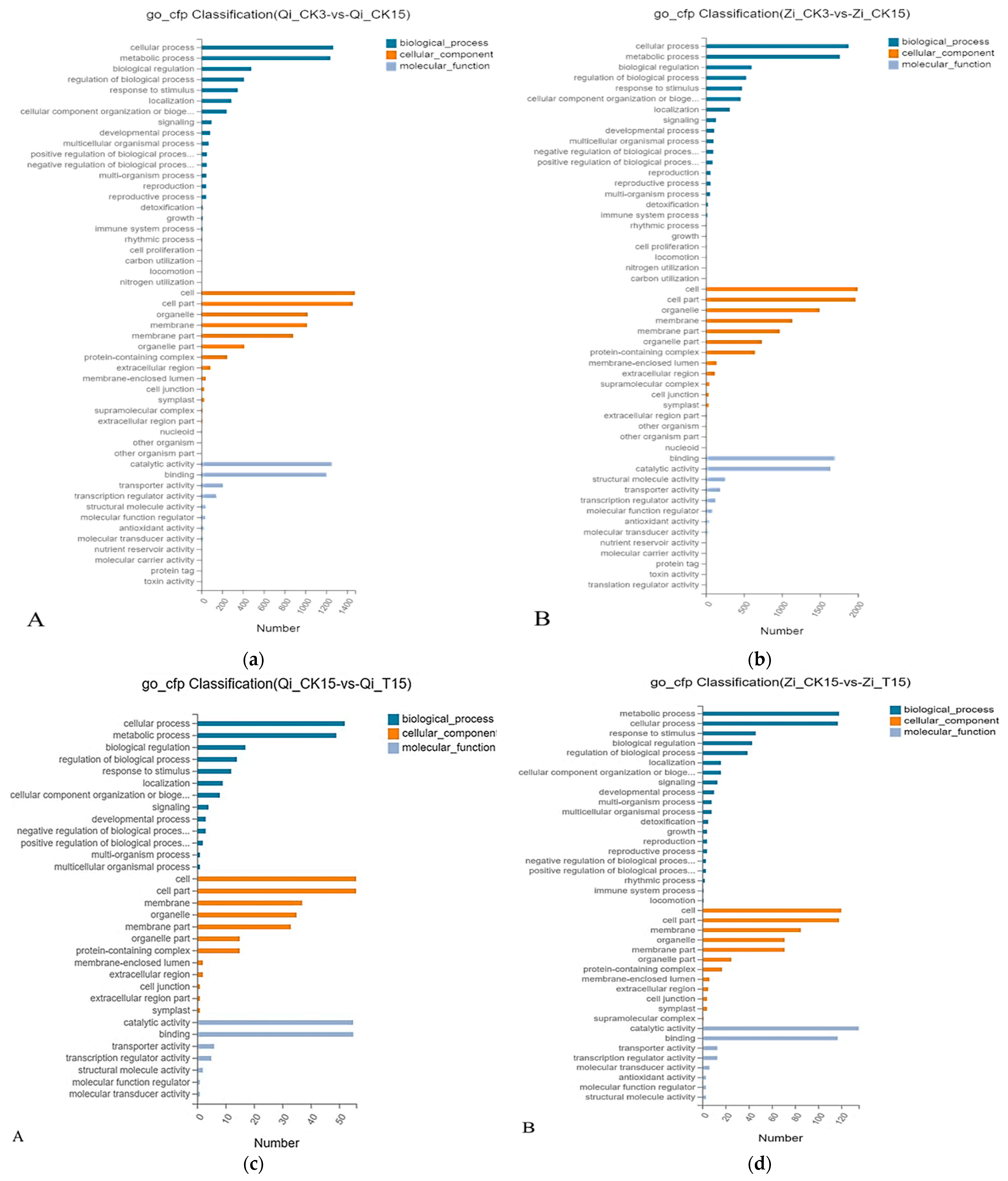
Figure 5.
Pathway enrichment analysis of the two inbred lines in different comparison groups. A: Pathway enrichment analysis of maize inbred line Qi319 under normal sowing depth and deep sowing stress; B: Pathway enrichment analysis of maize inbred line Zi330 under normal sowing depth and deep sowing stress. C: Pathway enrichment analysis of maize inbred line Qi319 after adding exogenous MeJA under deep seeding stress; D: Pathway enrichment analysis of maize inbred line Zi330 adding exogenous MeJA under deep seeding stress. The treatments and abbreviations are the same as those given in Figure 1.
Figure 5.
Pathway enrichment analysis of the two inbred lines in different comparison groups. A: Pathway enrichment analysis of maize inbred line Qi319 under normal sowing depth and deep sowing stress; B: Pathway enrichment analysis of maize inbred line Zi330 under normal sowing depth and deep sowing stress. C: Pathway enrichment analysis of maize inbred line Qi319 after adding exogenous MeJA under deep seeding stress; D: Pathway enrichment analysis of maize inbred line Zi330 adding exogenous MeJA under deep seeding stress. The treatments and abbreviations are the same as those given in Figure 1.
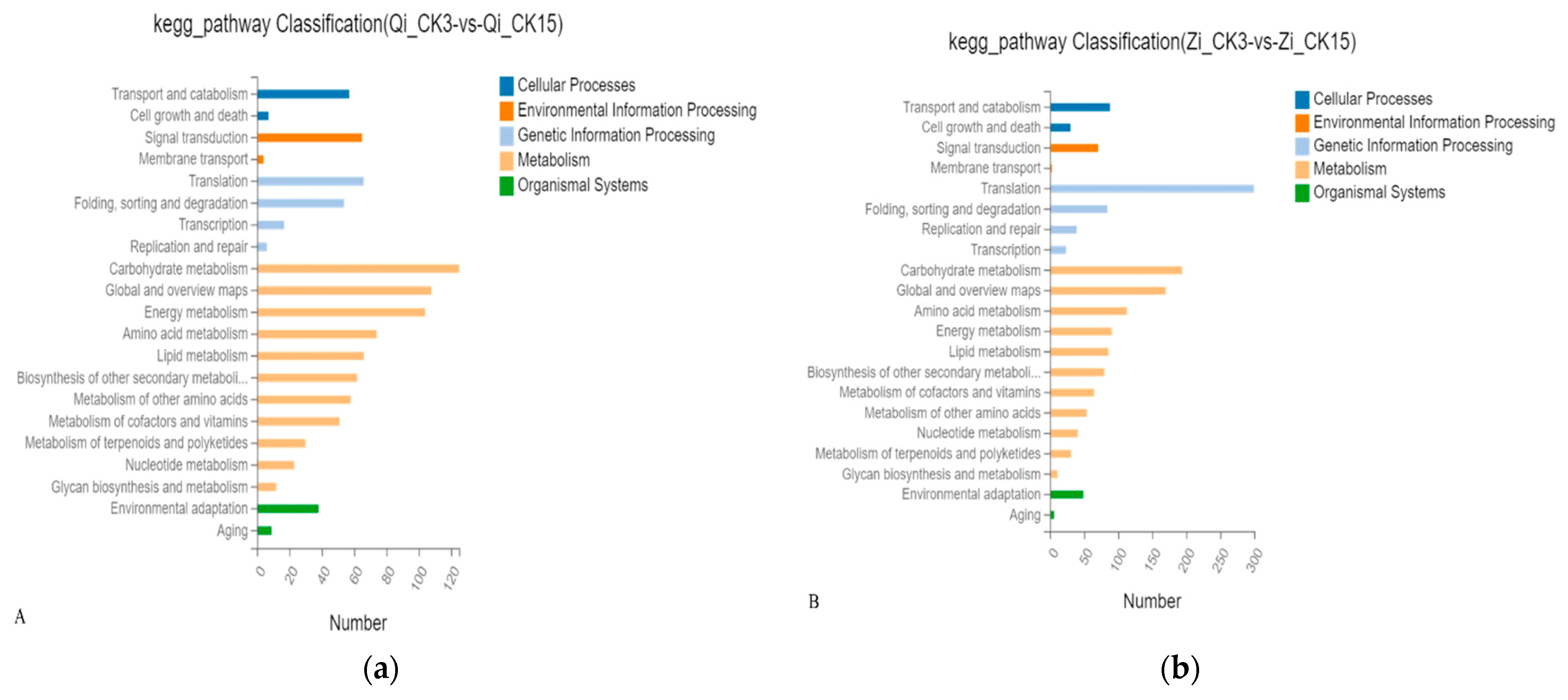
Figure 6.
Functional analysis of genes with significantly up-regulated expression in different comparison groups. A: Pathway enrichment analysis of two cultivars under normal and deep sowing stress; B: GO analysis of two varieties under normal and deep sowing stress; C: Pathway enrichment analysis of two cultivars under deep sowing stress after adding exogenous MeJA; D: GO analysis of two varieties under deep sowing stress after adding exogenous MeJA.
Figure 6.
Functional analysis of genes with significantly up-regulated expression in different comparison groups. A: Pathway enrichment analysis of two cultivars under normal and deep sowing stress; B: GO analysis of two varieties under normal and deep sowing stress; C: Pathway enrichment analysis of two cultivars under deep sowing stress after adding exogenous MeJA; D: GO analysis of two varieties under deep sowing stress after adding exogenous MeJA.
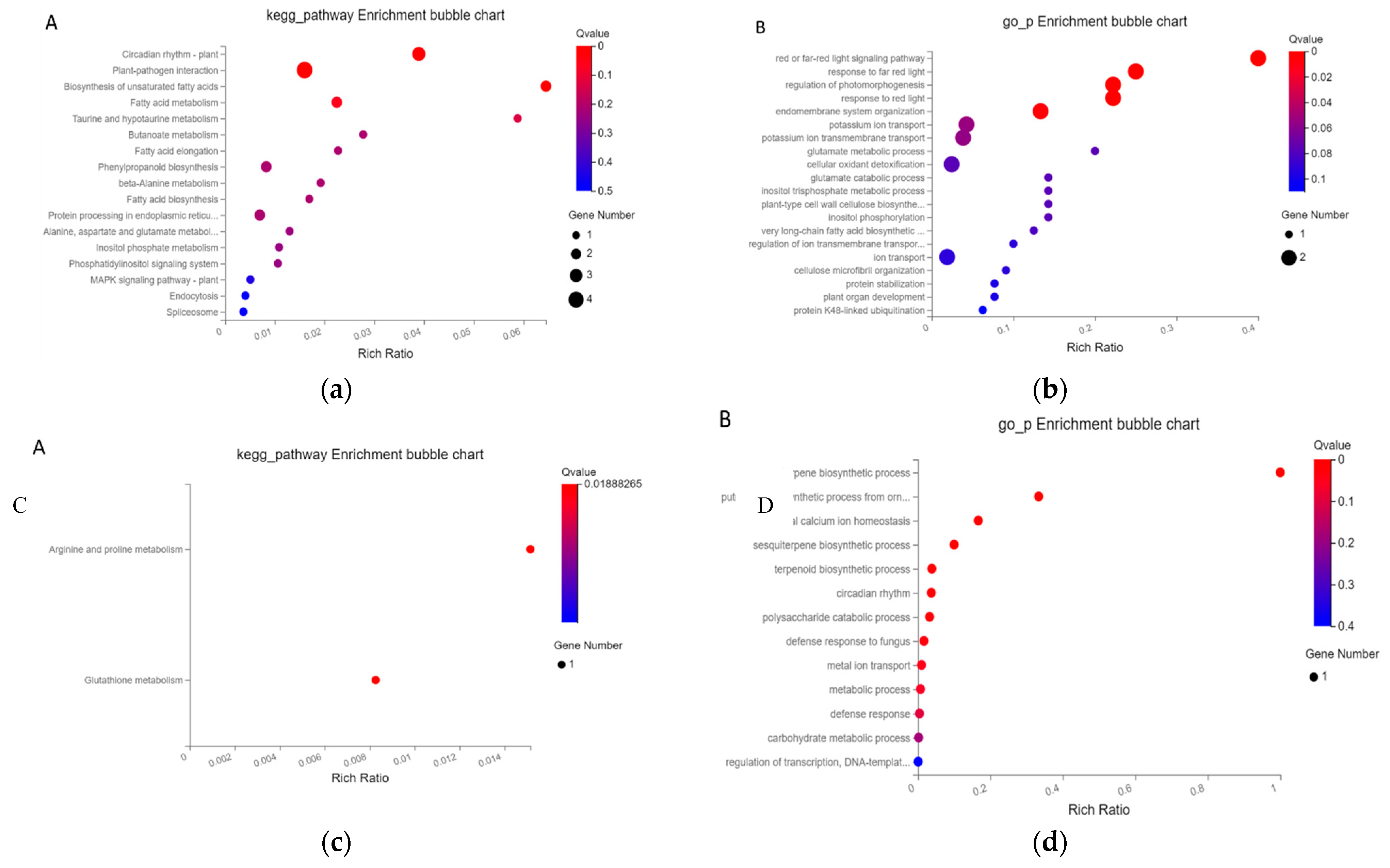
Figure 7.
Real time quantitative PCR validation of significantly upregulated differentially expressed genes between two varieties under deep sowing stress treatment. (A) the expression changes in response to the QCK, QDS, ZCK and ZDS treatments for each candidate gene as measured by qRT-PCR, (B) Scatter plot showing the changes in the expression (logy-fold change) of selected genes based on RNA-seq via qRT-PCR. QCK: Distilled water treatment under 3cm sowing depth of Qi319; QDS: Distilled water treatment under 15cm sowing depth of Qi319; ZCK: Distilled water treatment under 3cm sowing depth of Zi330; ZDS: Distilled water treatment under 15cm sowing depth of Zi330.
Figure 7.
Real time quantitative PCR validation of significantly upregulated differentially expressed genes between two varieties under deep sowing stress treatment. (A) the expression changes in response to the QCK, QDS, ZCK and ZDS treatments for each candidate gene as measured by qRT-PCR, (B) Scatter plot showing the changes in the expression (logy-fold change) of selected genes based on RNA-seq via qRT-PCR. QCK: Distilled water treatment under 3cm sowing depth of Qi319; QDS: Distilled water treatment under 15cm sowing depth of Qi319; ZCK: Distilled water treatment under 3cm sowing depth of Zi330; ZDS: Distilled water treatment under 15cm sowing depth of Zi330.
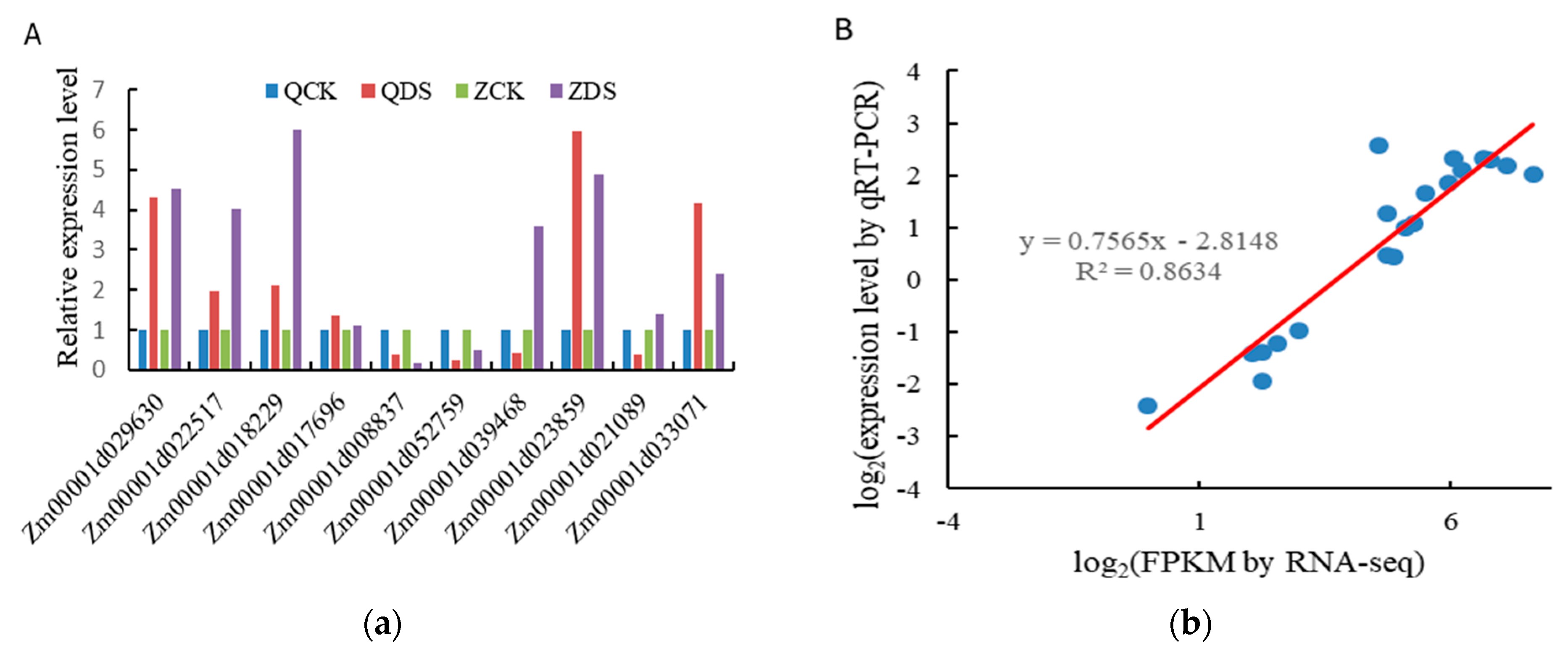
Figure 8.
Gene cluster analysis and correlation analysis between phenotype and module. (A)Hierarchical clustering analysis of co-expression genes. (B) Correlated heat maps between modules. (C) Correlation between gene module and phenotype. (D) Heat map of correlation between gene module and phenotype. MES: mesocotyl length; COL: coleoptile length; MESCOL: mesocotyl length and coleoptile length; MEWCOW: mesocotyl weight and coleoptile weight. RL: root length; RW: root fresh weight; SDL: seedling length; SDW: seedling fresh weight.
Figure 8.
Gene cluster analysis and correlation analysis between phenotype and module. (A)Hierarchical clustering analysis of co-expression genes. (B) Correlated heat maps between modules. (C) Correlation between gene module and phenotype. (D) Heat map of correlation between gene module and phenotype. MES: mesocotyl length; COL: coleoptile length; MESCOL: mesocotyl length and coleoptile length; MEWCOW: mesocotyl weight and coleoptile weight. RL: root length; RW: root fresh weight; SDL: seedling length; SDW: seedling fresh weight.
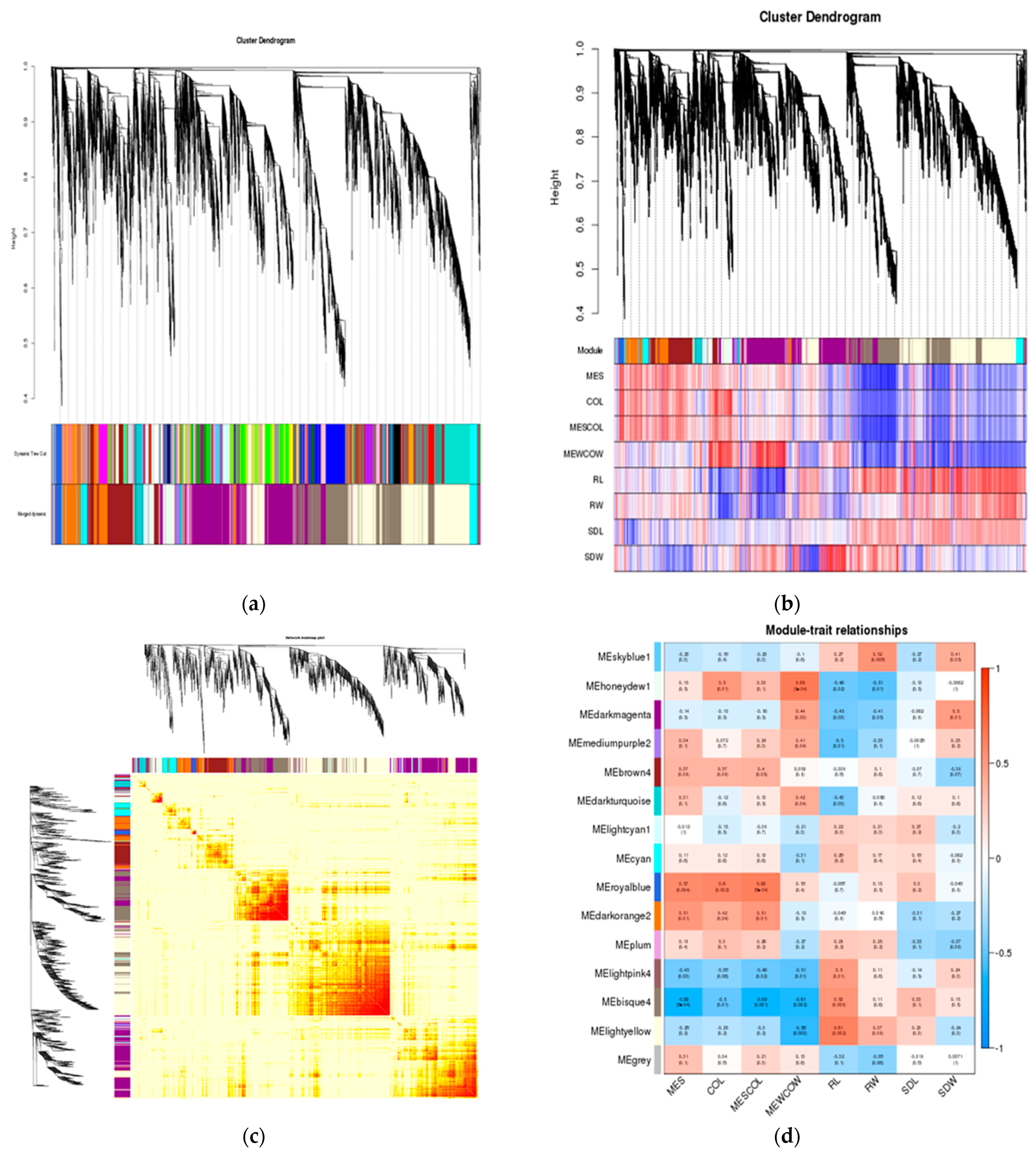
Figure 9.
Analysis of hub genes network interaction in phenotypic significant enrichment module. (A) Network interaction analysis of hub genes in royalblue module. (B) Network interaction analysis of hub genes in bisque4 module. The color gradient of the dots represent the high or low soft threshold of connectivity, with the redder color of the dots representing a higher soft threshold of connectivity.
Figure 9.
Analysis of hub genes network interaction in phenotypic significant enrichment module. (A) Network interaction analysis of hub genes in royalblue module. (B) Network interaction analysis of hub genes in bisque4 module. The color gradient of the dots represent the high or low soft threshold of connectivity, with the redder color of the dots representing a higher soft threshold of connectivity.
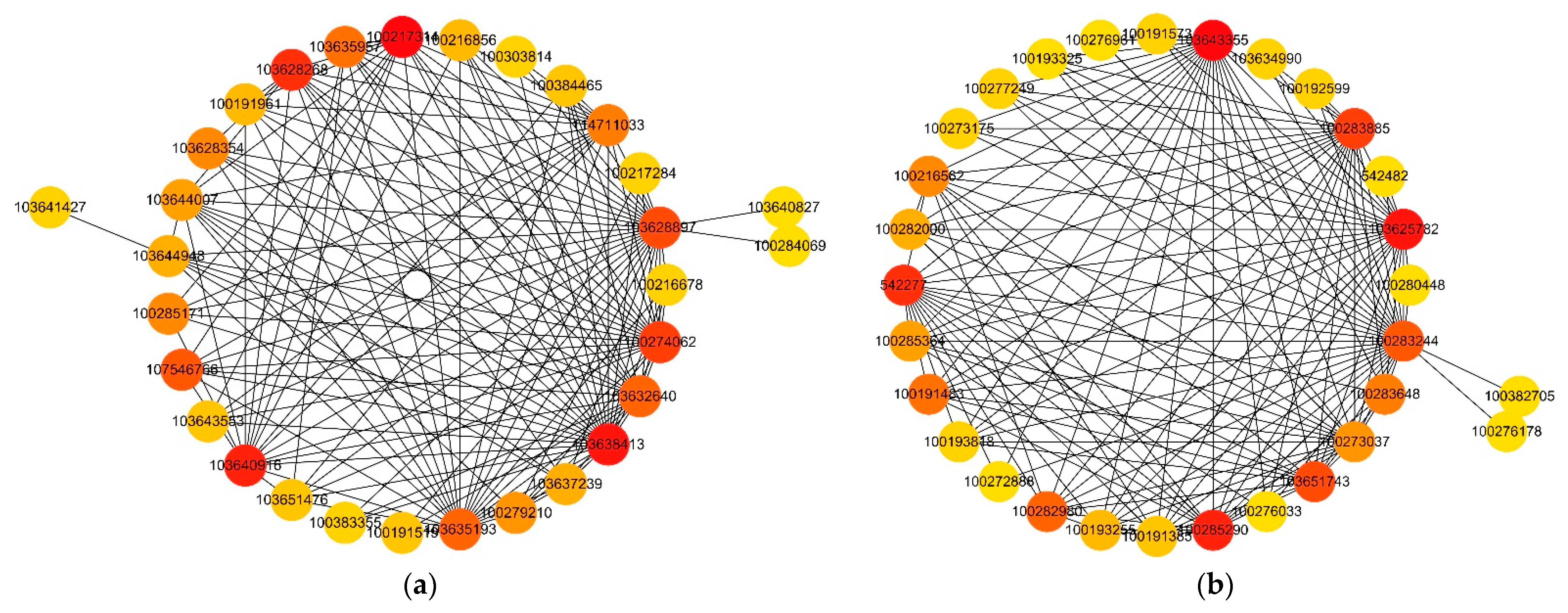
Figure 10.
Model of the molecular mechanisms underlying deep sowing tolerance and MeJA mitigation of deep sowing stress-induced damage in maize.
Figure 10.
Model of the molecular mechanisms underlying deep sowing tolerance and MeJA mitigation of deep sowing stress-induced damage in maize.
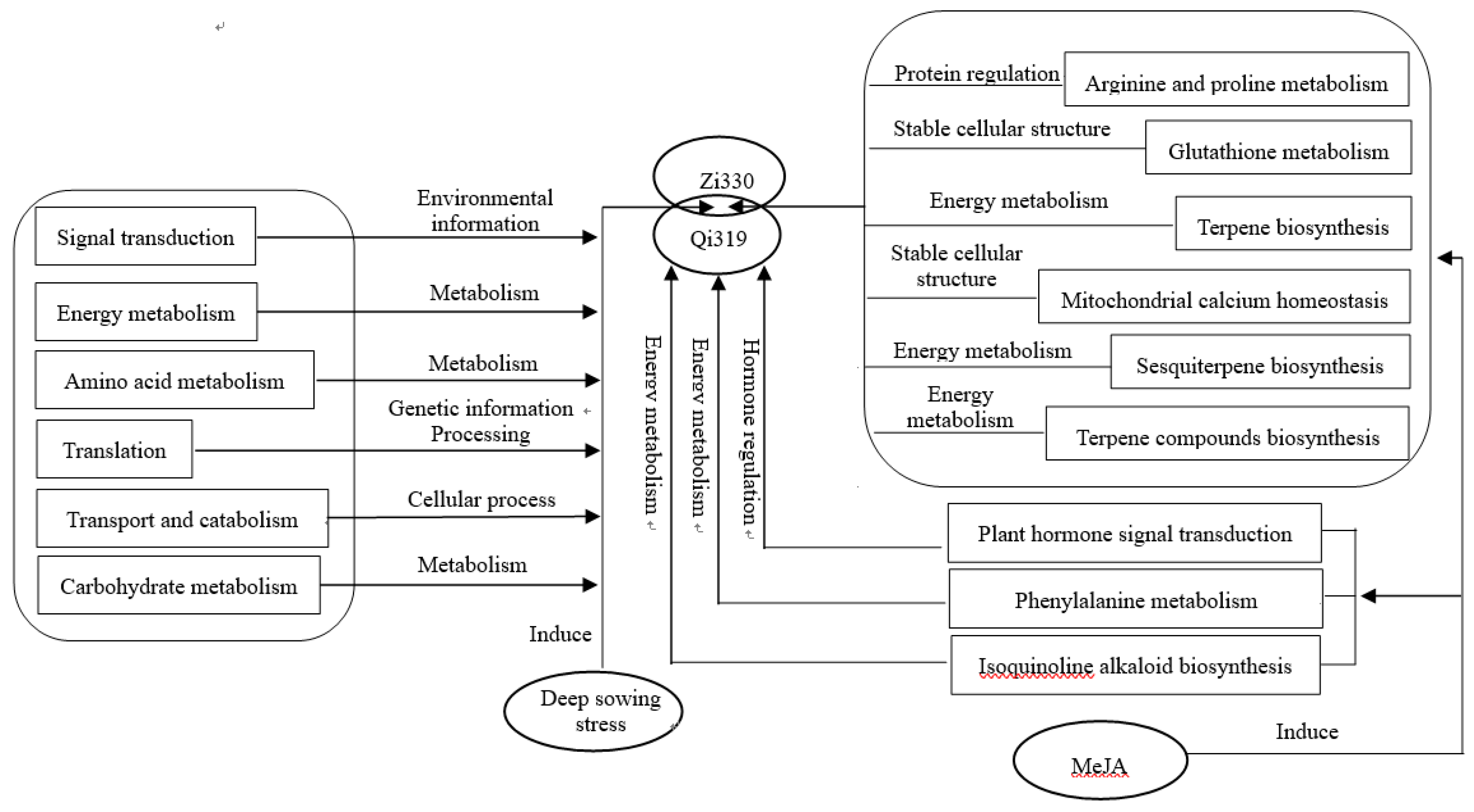

Table 1.
Seedling growth indexes of maize inbred lines treated with exogenous MeJA.
| Material | Treatment | SDL(cm) | RL(cm) | SDW(g) | RW(g) |
|---|---|---|---|---|---|
| Zi330 | CK | 26.60±0.81c | 33.10±0.64a | 1.57±0.06a | 1.38±0.12a |
| CM | 33.73±1.18a | 29.00±0.21b | 1.29±0.02b | 1.20±0.07ab | |
| DS | 29.23±0.86bc | 20.27±0.48c | 1.17±0.04b | 0.96±0.03b | |
| DM | 31.85±1.55ab | 18.80±0.30c | 1.26±0.05b | 1.28±0.01a | |
| Qi319 | CK | 34.10±2.17a | 34.93±0.64a | 1.21±0.09a | 1.32±0.03a |
| CM | 27.47±1.26ab | 34.67±0.79a | 0.86±0.04b | 1.33±0.06a | |
| DS | 25.25±5.05b | 23.40±1.20c | 1.07±0.10ab | 1.14±0.07b | |
| DM | 33.90±0.70a | 26.67±0.64b | 1.15±0.10ab | 1.34±0.03a |
Different lowercase letters represent the same inbred line with significant difference under different treatments (P<0.05), the following is the same. CK: Distilled water treatment under 3cm sowing depth; CM:3cm sowing depth with 1.5 μmol·L-1 exogenous MeJA treatment; DS: Distilled water treatment under 15cm sowing depth; DM: 15cm sowing depth with 1.5 μmol·L-1 exogenous MeJA treatment. RL: root length; RW: root fresh weight; SDL: seedling length; SDW: seedling fresh weight.
Table 2.
Growth characteristics of mesocotyl of maize inbred lines treated with exogenous MeJA.
| Material | Treatment | MES(cm) | COL(cm) | MES+COL(cm) | MESW+COLW(g) |
|---|---|---|---|---|---|
| Zi330 | CK | 2.67±0.17c | 2.57±0.20b | 5.23±0.35c | 0.33±0.04c |
| CM | 2.67±0.33c | 2.23±0.09b | 4.90±0.25c | 0.25±0.01c | |
| DS | 5.53±0.29b | 6.30±0.71a | 11.83±0.43a | 0.75±0.02a | |
| DM | 7.00±0.60a | 2.80±0.30b | 9.80±0.30b | 0.62±0.05b | |
| Qi319 | CK | 2.40±0.21d | 3.20±0.21b | 5.60±0.21c | 0.24±0.02c |
| CM | 3.13±0.09c | 2.50±0.40b | 5.63±0.32c | 0.23±0.03c | |
| DS | 8.00±0.20b | 3.53±1.90b | 13.30±1.00b | 0.37±0.06b | |
| DM | 8.60±0.12a | 7.10±0.35a | 15.70±0.26a | 0.52±0.03a |
Different lowercase letters represent the same inbred line with significant difference under different treatments (P<0.05). The treatments and abbreviations are the same as those given in Table 1. MES: mesocotyl length; COL: coleoptile length; MES+COL: mesocotyl length and coleoptile length; MESW+COLW: mesocotyl weight and coleoptile weight.
Table 3.
Summary of sequencing data.
| Sample | Total Raw Reads (M) | Total Clean Reads (M) | Total Clean Bases (Gb) | Clean Reads Q20 (%) | Clean Reads Q30 (%) | Clean Reads Ratio (%) |
|---|---|---|---|---|---|---|
| Zi_3_CK1 | 45.57 | 44.02 | 6.60 | 96.47 | 91.25 | 96.60 |
| Zi_3_CK2 | 45.57 | 44.01 | 6.60 | 96.50 | 91.33 | 96.56 |
| Zi_3_CK3 | 45.57 | 44.25 | 6.64 | 96.55 | 91.34 | 97.09 |
| Zi_3_T1 | 45.57 | 44.37 | 6.66 | 96.41 | 91.05 | 97.37 |
| Zi_3_T2 | 45.57 | 43.91 | 6.59 | 96.61 | 91.56 | 96.34 |
| Zi_3_T3 | 45.57 | 44.24 | 6.64 | 96.53 | 91.38 | 97.08 |
| Zi_15_CK1 | 45.57 | 44.51 | 6.68 | 96.42 | 91.08 | 97.66 |
| Zi_15_CK2 | 45.57 | 44.40 | 6.66 | 96.54 | 91.34 | 97.43 |
| Zi_15_CK3 | 45.57 | 44.29 | 6.64 | 96.53 | 91.36 | 97.17 |
| Zi_15_T1 | 45.57 | 44.26 | 6.64 | 96.59 | 91.50 | 97.13 |
| Zi_15_T2 | 45.57 | 44.1 | 6.62 | 96.55 | 91.35 | 96.77 |
| Zi_15_T3 | 45.57 | 44.38 | 6.66 | 96.51 | 91.31 | 97.37 |
| Qi_3_CK1 | 45.56 | 44.28 | 6.64 | 96.54 | 91.42 | 97.19 |
| Qi_3_CK2 | 45.57 | 44.18 | 6.63 | 96.56 | 91.45 | 96.95 |
| Qi_3_CK3 | 45.57 | 44.21 | 6.63 | 96.52 | 91.34 | 97.01 |
| Qi_3_T1 | 45.57 | 44.17 | 6.63 | 96.52 | 91.38 | 96.92 |
| Qi_3_T2 | 45.57 | 44.32 | 6.65 | 96.44 | 91.13 | 97.25 |
| Qi_3_T3 | 45.57 | 44.17 | 6.63 | 96.47 | 91.26 | 96.92 |
| Qi_15_CK1 | 45.57 | 44.15 | 6.62 | 96.44 | 91.17 | 96.88 |
| Qi_15_CK2 | 45.57 | 44.11 | 6.62 | 96.52 | 91.33 | 96.78 |
| Qi_15_CK3 | 47.33 | 44.12 | 6.62 | 96.65 | 91.72 | 93.23 |
| Qi_15_T1 | 45.57 | 44.07 | 6.61 | 96.59 | 91.55 | 96.7 |
| Qi_15_T2 | 45.57 | 43.95 | 6.59 | 96.64 | 91.65 | 96.43 |
| Qi_15_T3 | 45.57 | 43.93 | 6.59 | 96.62 | 91.65 | 96.40 |
Table 5.
Functional analysis of differential genes with different expression trends in the two varieties under deep sowing treatment.
Table 5.
Functional analysis of differential genes with different expression trends in the two varieties under deep sowing treatment.
| Gene ID | KEGG Pathway Desc | GO_P Desc |
|---|---|---|
| 100272950 | Plant hormone signal transduction | transcription, DNA-templated; regulation of transcription, DNA-templated |
| 100281647 | NA | systemic acquired resistance |
| 100284161 | NA | NA |
| 100284641 | NA | systemic acquired resistance |
| 103638673 | NA | NA |
| 103641531 | Glycine, serine and threonine metabolism; Tyrosine metabolism;Phenylalanine metabolism; beta-Alanine metabolism; Isoquinoline alkaloid biosynthesis; Tropane, piperidine and pyridine alkaloid biosynthesis | amine metabolic |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



