
Submitted:
11 September 2024
Posted:
11 September 2024
You are already at the latest version
Abstract
Avian Metapneumovirus (aMPV) is a significant poultry pathogen with a global presence, primarily causing respiratory issues in turkeys. It also affects chickens, although the severity of its impact is often lessened in this species. In Morocco, aMPV has been detected in broiler flocks, prompting the need to deeply analyze circulating strains to understand better the epidemiology and develop control measures accordingly. This research focuses on the sequencing and molecular characterization of aMPV in these flocks. Additionally, aMPV isolated from turkeys displaying Turkey Rhinotracheitis (TRT) signs, were included in the study to compare the findings. RNA extracted from positive swabs was subjected to nested PCR, targeting the attachment protein of the G gene, followed by gel electrophoresis. Amplicons were purified and sequenced using the Sanger method. Bioinformatics tools facilitated sequence analyses, including BLAST for similarity searches and Mega® for phylogenetic analysis using the maximum likelihood method with 1000 bootstrap replicates. The investigation unveiled the existence of three distinct clades of the same aMPV/B strain, two of which originated from used vaccines and one from field recombination, all circulating in broilers and turkeys and indicating potential virus transmission between both poultry species.
This article presents the first-ever molecular characterization of aMPV isolated from Moroccan broilers and turkeys, encompassing comprehensive investigations on its presence and subtype, and genetic characterization.
Keywords:
Avian Metapneumovirus
; Subtype B
; Morocco
; characterization
; sequencing
; phylogenetic analysis
1. Introduction
Avian Metapneumovirus (aMPV) is a globally recognized pathogen causing respiratory diseases, leading to substantial economic losses in the poultry industry. Also known as Metapneumovirus avis, it belongs to the Pneumoviridae family, specifically the genus Metapneumovirus [1].
So far, aMPV has been classified into four subtypes: A, B, C, and D [2]. These subtypes have been shown to exhibit varying effects on their avian hosts [3]. The recent disclosure of two new divergent viruses has also raised the possibility of further subtype candidates [4,5]. Traditionally, turkeys have been considered the most susceptible hosts to aMPV, with often more severe clinical symptoms displayed [6,7] and documented susceptibility to all four subtypes described [8,9]. However, recent reports have emphasized the infection’s impact on broilers, highlighting the role of aMPV as a primary infectious agent [10].
While aMPV’s global presence is established [11], specific epidemiological data at the local level are essential to understanding the virus dissemination, particularly in regions like North Africa, with high poultry density and diverse pathogens’ circulation. Unfortunately, background data on avian Metapneumovirus in this area is scarce and limited to only subtype B detection [12,13,14].
In Morocco, poultry production is diversified, with broiler farms established in various regions with different farm densities. Unlike turkeys, broilers in Morocco are generally not vaccinated against aMPV, except in some cases, where vaccination is adopted but not regularly. Despite frequent field reports of Swollen Head Syndrome (SHS), often attributed to aMPV, its real prevalence and impact remain unclear. Recent studies on Moroccan broiler flocks with varying health status have detected aMPV, thus considered a potential etiological agent of clinical respiratory cases [15,16].
Consequently, our study aimed to comprehensively investigate the molecular epidemiology of aMPV in Moroccan broiler farms. Additionally, the potential circulation of aMPV amongst turkeys was evaluated by sampling flocks displaying suggestive symptoms of Turkey Rhinotracheitis (TRT). This research represents the first-ever report on the molecular characterization of aMPV in Morocco.
2. Materials and Methods
2.1. Sample Collection and RNA Extraction
Twenty-two samples (n = 22) of aMPV RNA were extracted from field swabs, collected between March and June 2022 from six different broiler chicken flocks, and all of them were confirmed to be positive for aMPV by real-time RT-PCR [16]. Eighteen of them were obtained from a longitudinal field study, and four from broiler flocks affected by SHS.
Additionally, five (n = 5) freshly collected heads from one broiler flock showing SHS symptoms and three (n = 3) from one flock of turkeys displaying clinical signs suggestive of TRT, were analyzed. After sectioning the heads, a mixture of different tissues collected from skin, sub-cutis, and bones, was crushed and blended, using a homogenizer to ensure proper mixing and uniform distribution of the viral antigen. The aim was to detect aMPV of viruses obtained in turkey tissues and compare their sequences with those already isolated and characterized from broilers.
It should be pointed out that all the studied broilers of this study were strictly unvaccinated against SHS.
Table 1 outlines the diverse RNA samples and their respective sources, indicating whether they originated from broilers or turkeys’ flocks. These flocks were located in different geographic regions of Morocco, as shown in Figure 1.
These flocks were located in different geographic regions of Morocco, as per Figure 1.
A live subtype-B vaccine, the 1062 strain of chicken origin, was used as a positive reference control (Laboratorios HIPRA S.A., Amer, Spain). Its RNA was thus extracted, using the Kylt® RNA/DNA purification kit, following the manufacturer’s instructions (AniCon Labor GmbH, Emstek, Germany).
2.2. Reverse-Transcriptase and Double Conventional PCR Amplification
Using the kit Applied Biosystems® Reverse-Transcription Kit (ThermoFisher Scientific Waltham, Massachusetts, USA), 10µL of RNA from a 100µL dilution was mixed with 10µL of RT mix, made of MM RT Buffer (2µL), RT random primers (2µL), dNTP (0.8µL), multi-scribe RT (1µL) and H2O (4.2µL), with all the steps followed in compliance with the manufacturer’s instructions. The obtained cDNA was subjected to a double conventional PCR amplification protocol, often referred to as “nested PCR” or “nested amplification” to detect and subtype aMPV. This technique involved using two sets of primers in two successive and separate PCR reactions, where the second set of primers is located within the region amplified by the first set of primers. The sequences of these primers, used for a nested conventional PCR and designed in the attachment protein G gene [17,18], are reported in Table 2.
The primary reason for using this nested PCR method is to increase the specificity and sensitivity of cDNA amplifications. Specific primers, targeting the conserved regions of aMPV subtypes A and B, were used in separate reactions for each tested sample.
Table 3 illustrates the steps of the nested PCR for aMPV cDNA amplification.
2.3. Gel Electrophoresis
After a double conventional PCR amplification, the products were separated by gel electrophoresis. Thus, 0.8g of Agarose Top Vision Agarose® (Thermo Fisher Scientific Waltham, Massachusetts, USA) was added to 50mL of an in-house prepared Tris-Acetate-EDTA (TAE) solution. Containing 242g of Tris base, 57.1mL of glacial acetic acid, and 100mL of EDTA for every 1L of the preparation, this solution was adjusted to a pH of 8 and diluted to a 2% concentration (10mL of the prepared solution mixed with 490mL of distilled water). Additionally, 5µL of cDNA was included in the mixture with a green stain Xpert Green DNA Stain® (20.000X) (GRiSP, Lda, Porto, Portugal); such fluorescent dye allowed DNA visualization within the gel after electrophoresis.
Bands of the expected size would confirm aMPV detection and its subtype. The differentiation between the A and B subtypes was possible based on their respective base sizes of 268bp and 361bp.
2.4. Pre-Sequencing Samples Purification and Sequencing
Following gel electrophoresis, amplicons displaying the most robust band intensity were chosen for purification and sequencing. Gel extraction was performed using a solution extraction kit and a gel to ensure optimal retrieval. The resulting sequence products from solution extraction and gel were subsequently denoted at the end of the sequence, named “P” and “G”, respectively. The purification process was carried, out using the NucleoSpin® Gel and PCR Clean-up XS kit (Macherey Nagel, Düren, Germany), following the manufacturer’s guidelines.
For sequencing, the purified DNA fragments were sent to Plateau de Génomique Get-Purpan, UDEAR UMR 5165 CNRS/UPS, CHU PURPAN, Toulouse, France. Sanger sequencing was performed, using the forward and reverse primers used for PCR amplification. This allowed us to obtain the nucleotide sequence of the aMPV isolates.
2.5. Bioinformatics Analysis and Genetic Characterization
The obtained sequences were analyzed, using bioinformatics tools to determine the genetic characteristics and relationships of the Moroccan aMPV isolates. Using the BLAST program (National Center for Biotechnology Information, Bethesda, Maryland, USA; https://blast.ncbi.nlm.nih.gov/Blast.cgi), sequence similarities were sought with documented reference for aMPV strains, in the database, from North Africa, Europe, and the Mediterranean basin. Subsequently, multiple sequence alignment was performed using BioEdit® 7.2 (Ibis Therapeutics, Carlsbad, California, USA; https://bioedit.software.informer.com/7.2/), and phylogenetic analysis was conducted with Mega® 11, using the maximum likelihood (ML) method, with 1000 bootstrap replicates (Biodesign Institute, Arizona State University, USA; https://www.megasoftware.net/) and Figtree (Institute of Evolutionary Biology, University of Edinburgh, UK; http://tree.bio.ed.ac.uk/software/figtree/). Data were then analyzed using the neighbor-joining method.
Partial nucleotide sequences of the G gene of the Moroccan aMPV viruses were compared with published aMPV sequences, using a BLAST search within the EMBL/GenBank database. Sequences were investigated through phylogenetic analysis, comparing partial G sequences of the Moroccan aMPV subtype B detections with reference strains in the GenBank. Notably, the inclusion of reference strains in the phylogenetic tree was delimited to one strain per country, per year and per bird type to mitigate excessive condensation. These data are shown in Supplemental Tables S1, S2, and S3.
Additionally, two unpublished sequences, referred to as aMPV/Morocco/SF1 and aMPV/Morocco/SF2 and provided by the Avian Pathology Unit of the Agronomic and Veterinary Institute Hassan II, were included for comparative analysis with the study outcomes of this. It should be highlighted that the aMPV/Morocco/SF2 strain did not exhibit significant similarity with the reference strains available in GenBank; and for this reason, it was not considered for further accession number assignment.
Herewith, eight nucleotide sequences were submitted to GenBank, representing the Moroccan, under accession numbers PQ202991-PQ202998, as per Table 4.
3. Results
3.1. Double Conventional PCR Amplification
Although aMPV subtype B was detected in all analyzed samples, sequencing analysis was specifically conducted only on those displaying highly intense bands during gel electrophoresis. Thus, only one sequence of aMPV RNA extracts was identified (aMPV/Morocco/5P) and 2 out of 5 samples from broilers with SHS of which one yielded two different sequences after extraction (aMPV/Morocco/727P and G, aMPV/Morocco/731G). Similarly, 2 out of 3 RNA extracts from turkeys showing TRT signs were identified, of which only one exhibited two sequences after extraction in gel and solution (aMPV/Morocco/764P, aMPV/Morocco/766P and aMPV/Morocco/766G).
No samples were found positive for aMPV subtype A.
3.2. Sequencing and Genetic Characterization
The partial G gene sequences of the Moroccan isolates were 350 nucleotides in length. These isolates have a nucleotide sequence identity between 91.8% (aMPV/Morocco/727P and aMPV/Morocco/SF1, GenBank accession numbers PQ202991and PQ202997, respectively) and 99.7% (aMPV/Morocco/731G, and aMPV/Morocco/766P, GenBank accession numbers PQ202994 and PQ202996, respectively), and when compared to each other and from 78.2% (aMPV/Morocco/766G, GenBank accession number PQ202995, and aMPV-B/BR/1890/E1/19, GenBank accession number OP572408.1) to 90.76% (aMPV/Morocco/727G, GenBank accession number PQ202992 and aMPV/B/Romania/Ty/67/17, GenBank accession number MT432878.1), when compared to non-Moroccan aMPV isolates. The reference strains aforementioned are classified as derived vaccine ones.
Table 5 presents the highest percentage of similarities observed between the Moroccan isolates and the documented reference strains.
All Moroccan isolates showed a substantial likelihood, ranging from 90.76% to 99.70%, to reference strains, reported as vaccine-derived or associated with commercial subtype B vaccines.
Phylogenetic trees were constructed from the nucleotide and deduced amino-acid sequences of the partial G glycoprotein genes of Moroccan aMPV strains and non-Moroccan aMPV referenced strains. The ML phylogenetic analysis revealed that Moroccan aMPV isolates were separated into three main clusters (Figure 2).
The first cluster, related to documented 11/94 derived strains, included four Moroccan isolates: aMPV/Morocco/727G, aMPV/Morocco/P5, aMPV/Morocco/SF1 (GenBank accession numbers PQ202992, PQ202993 and PQ202997, respectively), besides the omitted isolate previously referred as aMPV/Morocco/SF2. These viruses grouped as well with chicken and turkey Nigerian and Algerian strains (aMPV/chicken/Nigeria/NIR89/2006; GenBank accession number AM490057.1, and aMPV/B/turkey/Algeria/26; GenBank accession number KP892758.1), respectively.
The second cluster, is represented by four viruses: aMPV/Morocco/731G, aMPV/Morocco/764P, aMPV/Morocco/766G, and 766P (GenBank accession numbers PQ202994, PQ202998, PQ202995 and PQ202996, respectively). It is grouped with vaccine strain pl21 (GenBank accession number MZ574139.1) and its derived strains.
The presumed third group contains only one Moroccan virus, aMPV/Morocco/727P (GenBank accession number PQ202991), clustering with field strains reported in Europe, but also the Tunisian vaccine-derived strain TN1015/17 (GenBank accession number MH411239.1).
4. Discussion
Avian Metapneumovirus has been implicated as the causal agent of swollen head syndrome since its first clinical description in the 1990s [24]. While the circulation of aMPV has been suspected in Morocco, it has often been considered a minor co-infecting agent in clinical respiratory problems, particularly in broiler flocks, for which its significance has been traditionally overlooked [15]. This has resulted in a lack of knowledge regarding its prevalence and incidence in broilers. This paper represents the first comprehensive report on the molecular characterization of aMPV strains circulating in Morocco, revealing three clusters of the aMPV/B subtype, two of them being vaccine-derived. The observed genetic diversity underscored the presence of different Metapneumovirus strains of subtype B among broiler and turkey farmings in Morocco, possibly having diverse sources.
Per prior research’s focus on aMPV subtypes, our study has been limited to investigating aMPV/A and aMPV/B circulation [15,16].
So far, only subtype B has been detected and sequenced, from either stored or freshly collected samples. These findings align with previous investigations, emphasizing the dominance of subtype B, especially in broiler chickens [16]. The prevalence of aMPV/B has shown a substantial increase across different regions of the world, including a very recent description in the USA [23], and aligning with previous research in neighboring countries from North Africa and the Mediterranean areas, emphasizing the dominance of subtype B [10,12,13,14,20,25,26,27,28,29]. It is worth noting that subtype B spreads more widely through respiratory routes, unlike subtype A, which is more limited to the oro-fecal route [30] and with low excretion in chickens [31].
The constructed phylogenetic tree and nucleotide likelihood identity highlighted distinct clades of aMPV subtype B in Morocco. In the first clade, Moroccan isolates formed clusters with strains documented in various countries, notably those identified as vaccine-derived ones [19,20,21], and exhibited grouping with the vaccine strain 11/94. While previous reports pointed out the possibility of the vaccine spread from nearby vaccinated flocks, presumably turkeys [19,20], the detection of aMPV in our case resulted as well from flocks with clinical expression of the SHS, albeit far from any other poultry production types mandatorily vaccinated against aMPV [16]. This finding suggests the hypothesis of potential virus transmission from Europe to North Africa, possibly facilitated by migratory birds or trade [13]. It is crucial to acknowledge the limitations of our study due to the scarcity of research and data available from North African countries, except Tunisia. This limitation may introduce bias and distort the understanding of aMPV subtypes spread from wild birds [32]. Given the insufficient public databases, further investigations are necessary to comprehensively elucidate the epidemiological patterns of aMPV infection and pathogenicity. Nevertheless, it is noteworthy that this hypothesis contradicts investigations from Italy that have downplayed the role of wild birds in the spread of aMPV subtypes A and B [33].
Notably, the Algerian strain (GenBank accession number KP892758.1) [14] demonstrated substantial phylogenetic proximity to the Moroccan isolates within this clade, highlighting consistent and comparable epidemiological context across North African countries.
Within the second clade, which predominantly encompasses strains detected in turkeys, a convergence of viruses was observed, originating from widely employed commercial vaccines and Italian turkey field isolates. This pattern aligns with previous findings that grouped these viruses, whether vaccines, vaccine-derived, or old strains, with aMPV/B vaccine, strains VCO3 or 1062, identified in vaccinated turkeys in Italy [19]. Previous reports on isolates with high phylogenetic proximity to our second cluster confirmed their unrelatedness to vaccine strains 1062 or pl21 [34]. However, the documented strains with similarity with the Moroccan isolates all showed closeness to vaccine-derived strain pl21 [22,23,31]., which can plausibly explain the observed similarities with the Tunisian isolate, linked to the vaccine strain pl21 [13], and that might have reverted to virulence. Interestingly, this Tunisian exhibits a 99% genetic similarity with the Algerian and Nigerian strains (GenBank accession numbers KP892758 and AM490058, respectively), classified as vaccine-derived strains [13]. Our isolates of the first group also clustered, on the phylogenetic tree, with these vaccine-derived strains. These observations reinforce the hypothesis that the North African isolates share a common origin.
It is plausible that detection of aMPV/B in turkeys, looked up cautiously, given the limited number of samples tested (three individual birds originated from the same flock) might be related to the extensive use, in the past and potentially still, of heterologous vaccines against aMPV/A for many years, exerting vaccine pressure and generating alternation of aMPV/B [35].
Remarkably, the cluster of isolates solely from turkey flocks affected by TRT also includes one aMPV strain detected in chickens (aMPV/Morocco/731G, GenBank accession number PQ202994), showing a high percentage of nucleotide identity to the Brazilian and Italian strains, aMPV-B/BR/1890/E1/19 (GenBank accession number OP572408.1) and aMPV/B/Italy/Ty/742-01/17 (GenBank accession number MT436229.1) respectively. The notable similarity existing between broiler and turkey aMPV/B local strains holds significance, indicating the potential for aMPV transmission between these two species. This finding suggests that the aMPV strains, detected in Moroccan broilers, also circulate in turkeys and vice versa, implying a form of cross-species transmission [36].
This outcome is particularly pertinent for the specificities of farm complexes in Morocco, which are granted authorization for rearing either broiler chickens or fattening turkeys in the same complex, leading to the alternation of both production types throughout the year. On a larger scale, this holds for regions with high-density poultry, highlighting the need for vigilant biosecurity and disease management practices to curb the potential impact of diverse aMPV strains within poultry populations. In high-risk areas, vaccination is an indispensable part of the strategic control of aMPV, alongside strict biosecurity measures [37].
The aMPV/Morocco/727P (GenBank accession number PQ202991), detected in broilers, presumably formed a distinct third clade with field strains. It displayed nucleotide sequence similarities with the Italian-related isolates, previously documented in chickens and various Euro-Mediterranean areas [10,25,27,29] and capable of escaping vaccine-induced immunity [38], just like the Tunisian strain TN1000/19 (GenBank accession number MZ229769.1) reported as field one and plausibly originating from Spain [13]. Nevertheless, performing nucleotide identity analysis with reference strains showed that aMPV/Morocco/727P disclosed high similarity with both the Nigerian and the Algerian strains, aMPV/chicken/Nigeria/NIR89/2006 (GenBank accession number AM490057.1) and Algeria/26/aMPVB/turkey (GenBank accession number KP892758.1), suggesting its clustering within the first group. This finding likely confirms that aMPV/Morocco/727P and aMPV/Morocco/727G could represent the same isolate.
Although our research coincided with a significant incidence of swollen head syndrome reported in the field, it does not rule out the possible circulation of either subtype B or subtype A or both in Morocco, without prior documentation. Further research is required to enhance our comprehension of aMPV/B dissemination in Morocco through larger-scale sampling. Additionally, despite the description of virus isolation from apparently ill birds, the analysis could not necessarily provide definitive insights into the pathogenicity of the locally identified strains. Last but not least, it is essential to emphasize the significance of a surveillance program in other poultry industrial species to gain a more comprehensive understanding of the prevalence of the disease.
5. Conclusions
As the first endeavor of its kind, this research sets the stage for future investigations and serves as a vital reference point for ongoing monitoring and surveillance efforts to better understand the dynamics of aMPV circulation in the Moroccan poultry population and to develop effective control strategies for preventing and managing aMPV infections, with emphasis on broilers and turkeys. Ultimately, a comprehensive understanding of aMPV epidemiology is pivotal to mitigating its impact on poultry flocks and ensuring sustainable poultry production in Morocco.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: North African aMPV/B strains obtained or retrieved from GenBank; Table S2: Mediterranean aMPV/B strains obtained or retrieved from GenBank; Table S3: Other aMPV/B strains obtained or retrieved from GenBank.
Author Contributions
Conceptualization, A.M.; Methodology, A.M., A.G., S.N. and S.F.; Software, O.A., and S.F.; Validation, A.G., E.C., and C.L.; Formal Analysis, S.F.; Investigation, A.M., and M.M.; Resources, M.D, and H.F.; Data Curation, O.A., S.F. and C.L.; Writing – Original Draft Preparation, A.M.; Writing – Review & Editing, E.C., C.L. and A.G.; Visualization, A.M. and J.C.; Supervision, S.N.; Project Administration, H.F., M.B. and M.D.; Funding Acquisition, E.C.S. All authors have read and agreed to the published version of the manuscript.
Funding
The Agronomic and Veterinary Institute Hassan II and Laboratorios HIPRA, S.A., Spain funded the present study. Laboratorios HIPRA, S.A., Spain funded the APC.
Institutional Review Board Statement
Ethical review and approval were waived for this study due to its conduction before the implementation of the local Committee of Ethics (a detailed justification is provided as “Attestation of Ethics Committee”).
Acknowledgments
The authors wish to thank The Avian Pathology Unit of the Agronomic and Veterinary Institute Hassan II for technical assistance in the sequence analysis. Thanks go also to Dr. Jihene Lachheb and Dr. Zied Bouslama from Institut Pasteur de Tunis, University of Tunis El Manar (UTM), Tunisia, for providing some sequences used in this study.
Conflicts of Interest
The authors declare no conflict of interest. The funders did not influence the study’s design; the collection, analysis, or interpretation of the data; the writing of the manuscript; or the decision to publish the results.
References
- International Committee on Taxonomy of Viruses. Virus taxonomy: 2022, [released, 2022; modified, March 2023; cited, Sept. 2023]. Available from:https://talk.ictvonline.org/taxonomy/.
- Cook, J.K.A.; Cavanagh, D. Detection and differentiation of avian pneumoviruses (metapneumoviruses). Avian Pathol. 2002, 31, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.A.; Allée, C.; Courtillon, C.; Szerman, N.; Lemaitre, E.; Toquin, D.; Mangart, J.M.; Amelot, M.; Eterradossi, N. Host specificity of avian metapneumoviruses. Avian Pathol. 2019, 48, 311–318. [Google Scholar] [CrossRef]
- Canuti, M.; Kroyer, A.N.K.; Ojkic, D.; Whitney, H.G.; Robertson, G.J.; Lang, A.S. Discovery and characterization of novel rna viruses in aquatic North American wild birds. Viruses 2019, 11, 1–14. [Google Scholar] [CrossRef]
- Retallack, H.; Clubb, S.; DeRisi, J.L. Genome Sequence of a Divergent Avian Metapneumovirus from a Monk Parakeet (Myiopsitta monachus). Microbiol. Resour. Announc. 2019, 8, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Bexter, F.; Rüger, N.; Sid, H.; Herbst, A.; Gabriel, G.; Osterhaus, A.; Rautenschlein, S. In Vitro Investigation of the Interaction of Avian Metapneumovirus and Newcastle Disease Virus with Turkey Respiratory and Reproductive Tissue. Viruses 2023, 15(4) 907, 1–19. [Google Scholar] [CrossRef]
- Hartmann, S.; Sid, H.; Rautenschlein, S. Avian metapneumovirus infection of chicken and turkey tracheal organ cultures: comparison of virus–host interactions. Avian Pathol. 2015, 44, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Kaboudi, K.; Lachheb, J. Avian metapneumovirus infection in turkeys: a review on turkey rhinotracheitis. J Appl Poult Res 2021, 100211, 1–17. [Google Scholar] [CrossRef]
- Lupini, C.; Legnardi, M.; Graziosi, G.; Cecchinato, M.; Listorti, V.; Terregino, C.; Catelli, E. Vaccine Interaction and Protection against Virulent Avian Metapneumovirus (aMPV) Challenge after Combined Administration of Newcastle Disease and aMPV Live Vaccines to Day-Old Turkeys. Vaccines 2023, 708, 1–10. [Google Scholar] [CrossRef]
- Tucciarone, C.M.; Franzo, G.; Lupini, C.; Alejo, C.T.; Listorti, V.; Mescolini, G.; Brandão, P.E.; Martini, M.; Catelli, E.; Cecchinato, M. Avian Metapneumovirus circulation in Italian broiler farms. Poult. Sci. 2018, 97, 503–509. [Google Scholar] [CrossRef]
- Suarez, D.L.; Miller, P.J.; Koch, G.; Mundt, E.; Rautenschlein, S. Newcastle disease, other avian paramyxoviruses, and avian metapneumovirus infections. In Diseases of Poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., de Wit, S., Grimes, T., Johnson, D., Kromm, M., Prajitno, T.Y., Rubinoff, I., Zavala, G. Eds.; Wiley-Blackwell. Hoboken New Jersey, USA, 2020; pp. 111–166. [CrossRef]
- Jbenyeni, A.; Croville, G.; Cazaban, C.; Guérin, J.L. Predominance of low pathogenic avian influenza virus H9N2 in the respiratory co-infections in broilers in Tunisia: a longitudinal field study, 2018 – 2020. Vet. Res. 2023, 1–14. [Google Scholar] [CrossRef]
- Lachheb, J.; Bouslama, Z.; Nsiri, J.; Badr, C.; Al Gallas, N.; Souissi, N.; Khazri, I.; Larbi, I.; Kaboudi, K.; Ghram, A. Phylogenetic and phylodynamic analyses of subtype-B metapneumovirus from chickens in Tunisia. Poult. Sci. 2022, 102, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sid, H.; Benachour, K.; Rautenschlein, S. Co-infection with Multiple Respiratory Pathogens Contributes to Increased Mortality Rates in Algerian Poultry Flocks. Avian Dis. 2015, 59, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Mernizi, A.; Bouziane, S.; Fathi, H.; Criado, J.; Bouslikhane, M.; Ghram, A.; Catelli, E.; Mouahid, M.; Nassik, S. First seroepidemiological and risk factor survey of avian metapneumovirus circulation in Moroccan broiler farms. Vet. Glas. 2022, 77, 1–16. [Google Scholar] [CrossRef]
- Mernizi, A.; Kadiri, O.; Criado, J.L.; Bouslikhane, M.; Ghram, A.; Mouahid, M.; Catelli, E.; Nassik, S. Detection of Avian Metapneumovirus Subtypes A and B in Moroccan Broiler. Iran J Vet Med 2023, 1–28. [Google Scholar] [CrossRef]
- Juhasz, K.; Easton, A.J. Extensive sequence variation in the attachment (G) protein gene of avian pneumovirus: Evidence for two distinct subgroups. J. Gen. Virol. 1994, 75, 2873–2880. [Google Scholar] [CrossRef] [PubMed]
- Cecchinato, M.; Catelli, E.; Lupini, C.; Ricchizzi, E.; Clubbe, J.; Battilani, M.; Naylor, C.J. Avian metapneumovirus (AMPV) attachment protein involvement in probable virus evolution concurrent with mass live vaccine introduction. Vet. Microbiol. 2010, 146, 24–34. [Google Scholar] [CrossRef]
- Mescolini, G.; Lupini, C.; Franzo, G.; Quaglia, G.; Legnardi, M.; Cecchinato, M.; Tucciarone, C.M.; Blanco, A.; Turblin, V.; Biarnés, M.; Tatone, F.; Falchieri, M.; Catelli, E. What is new on molecular characteristics of Avian metapneumovirus strains circulating in Europe? Transbound. Emerg. Dis. 2021, 68, 1314–1322. [Google Scholar] [CrossRef]
- Bayraktar, E.; Umar, S.; Yilmaz, A.; Turan, N.; Franzo, G.; Tucciarone, C.M.; Cecchinato, M.; Cakan, B.; Iqbal, M.; Yilmaz, H. First molecular characterisation of avian metapneumovirus (aMPV) in Turkish broiler flocks. Avian Dis. 2018, 62, 425–430. [Google Scholar] [CrossRef]
- Owoade, A.A.A.; Ducatez, M.F.; Hübschen, J.M.; Sausy, A.; Chen, H.; Guan, Y.; Muller, C.P. Avian Metapneumovirus Subtype A in China and Subtypes A and B in Nigeria. Avian Dis. 2008, 52, 502–506. [Google Scholar] [CrossRef]
- Kariithi, H.M.; Volkening, J.D.; Veiga Alves, V.; Reis-cunha, J.L.; Cury Rocha Veloso Arantes, L.; Santos Fernando, F.; Fernandes Filho, T.; da Silva Martins, N.R.; Lemière, S.; de Freitas Neto, O.C.; Decanini, E.L.; Afonso, C.L.; Suarez, D.L. Complete Genome Sequences of Avian Metapneumovirus. Microbiol. Resour. Announc. 2023, 7, 8–11. [Google Scholar] [CrossRef]
- Luqman, M.; Duhan, N.; Temeeyasen, G.; Selim, M.; Jangra, S.; Mo, S.K. Geographical Expansion of Avian Metapneumovirus Subtype B: First Detection and Molecular Characterization of Avian Metapneumovirus Subtype B in US Poultry. Viruses 2024, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- El Houafdi, M.; Hamam, J.; Vanmarche, J.; Cook, J.K.A. Swollen head syndrome in broiler chicken in Morocco. In Proceedings of 40th Western Poultry Disease Conference, Acapulco, Mexico, Mar 10-13; Jensen, M.; Sarfati, D., Eds; Asociación Nacional de Especialistas en Ciencias Avícolas, Cancun, Mexico, 1991, p. 126–127.
- Andreopoulou, M.; Franzo, G.; Tucciarone, C.M.; Prentza, Z.; Koutoulis, K.C.; Cecchinato, M.; Chaligianni, I. Molecular epidemiology of infectious bronchitis virus and avian metapneumovirus in Greece. Poult. Sci. 2019, 98, 5374–5384. [Google Scholar] [CrossRef]
- Banet-Noach, C.; Simanov, L.; Perk, S. Characterization of Israeli avian metapneumovirus strains in turkeys and chickens. Avian Pathol. 2005, 34, 220–226. [Google Scholar] [CrossRef]
- Franzo, G.; Tucciarone, C.M.; Enache, M.; Bejan, V.; Ramon, G.; Koutoulis, K.C.; Cecchinato, M. First Report of Avian Metapneumovirus Subtype B Field Strain in a Romanian Broiler Flock during an Outbreak of Respiratory Disease. Avian Dis. 2017, 61, 250–254. [Google Scholar] [CrossRef]
- Lupini, C.; Tucciarone, C.M.; Mescolini, G.; Quaglia, G.; Graziosi, G.; Turblin, V.; Brown, P.; Cecchinato, M.; Legnardi, M.; Delquigny, T.; Lemière, S.; Perreul, S.; Catelli, E. Longitudinal Survey on aMPV Circulation in French Broiler Flocks following Different Vaccination Strategies. Animals 2023, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tucciarone, C.M.; Andreopoulou, M.; Franzo, G.; Prentza, Z.; Chaligiannis, I.; Cecchinato, M. First Identification and Molecular Characterization of Avian metapneumovirus Subtype B from Chickens in Greece. Avian Dis. 2017, 61, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.A.; Allée, C.; Courtillon, C.; Szerman, N.; Lemaitre, E.; Toquin, D.; Mangart, J.M.; Amelot, M.; Eterradossi, N. Host specificity of avian metapneumoviruses. Avian Pathol. 2019, 48, 311–318. [Google Scholar] [CrossRef]
- Loor-Giler, A.; Muslin, C.; Santander-Parra, S.; Coello, D.; de la Torre, D.; Abad, H.; Nuñez, L. Simultaneous detection of infectious bronchitis virus and avian metapneumovirus genotypes A, B, and C by multiplex RT-qPCR assay in chicken tracheal samples in Ecuador. Front. vet. sci. 2024, 11, 1–10. [Google Scholar] [CrossRef]
- Graziosi, G.; Lupini, C.; Catelli, E. Disentangling the role of wild birds in avian metapneumovirus (aMPV) epidemiology: A systematic review and meta-analysis. Transbound. Emerg. Dis. 2022, 69, 3285–3299. [Google Scholar] [CrossRef]
- Tucciarone, C.M.; Franzo, G.; Legnardi, M.; Pasotto, D.; Lupini, C.; Catelli, E.; Quaglia, G.; Graziosi, G.; Dal Molin, E.; Gobbo, F.; Cecchinato, M. Molecular Survey on A, B, C and New Avian Metapneumovirus (aMPV) Subtypes in Wild Birds of Northern-Central Italy. Vet. Sci. 2022, 9, 1–11. [Google Scholar] [CrossRef]
- Nguyen, V.G.; Chung, H.C.; Do, H.Q.; Nguyen, T.T.; Cao, T.B.P.; Truong, H.T.; Mai, T.N.; Le, T.T.; Nguyen, T.H.; Le, T.L.; Huynh, T.M.L. Serological and molecular characterization of avian metapneumovirus in chickens in Northern Vietnam. Vet. Sci. 2021, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Salles, G.B.C.; Pilati, G.V.T.; Savi, B.P.; Muniz, E.C.; Dahmer, M.; Vogt, J.R.; de Lima Neto, A.J.; Fongaro, G. Surveillance of Avian Metapneumovirus in Non-Vaccinated Chickens and Co-Infection with Avian Pathogenic Escherichia coli. Microorganisms 2024, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lupini, C.; Cecchinato, M.; Ricchizzi, E.; Naylor, C.J.; Catelli, E. A turkey rhinotracheitis outbreak caused by the environmental spread of a vaccine-derived avian metapneumovirus. Avian Pathol. 2011, 40, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Salles, G.B.C.; Pilati, G.V.T.; Muniz, E.C.; de Lima Neto, A.J.; Vogt, J.R.; Dahmer, M.; Savi, B.P.; Padilha, D.A.; Fongaro, G. Trends and Challenges in the Surveillance and Control of Avian Metapneumovirus. Viruses 2023, 15, 1–11. [Google Scholar] [CrossRef]
- Catelli, E.; Lupini, C.; Cecchinato, M.; Ricchizzi, E.; Brown, P.; Naylor, C.J. Field avian Metapneumovirus evolution avoiding vaccine induced immunity. Vaccine 2010, 28, 916–921. [Google Scholar] [CrossRef]
Figure 1.
Map of Morocco, outlining the origins of the different samples studied. The sample types are indicated as follows: △: longitudinal study sampling, ◯: one-off sampling, *: fresh broiler head sampling, ☆: fresh turkey head sampling from a slaughterhouse.
Figure 1.
Map of Morocco, outlining the origins of the different samples studied. The sample types are indicated as follows: △: longitudinal study sampling, ◯: one-off sampling, *: fresh broiler head sampling, ☆: fresh turkey head sampling from a slaughterhouse.
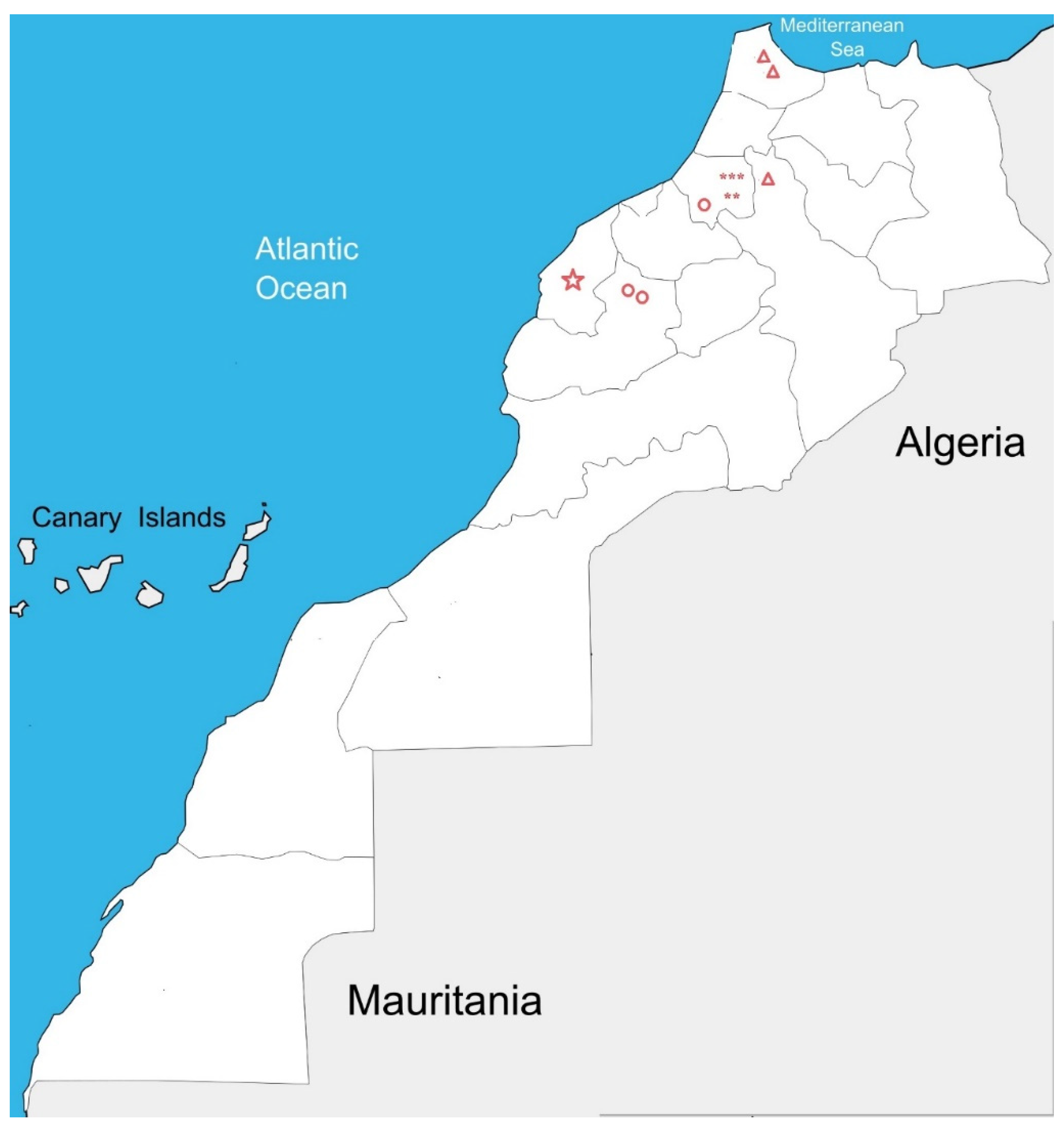
Figure 2.
Phylogenetic analysis of the partial nucleic acid sequence of the G gene. Detections of aMPV/B from Morocco are shaded in yellow whereas vaccine strains are framed in blue. Data were analyzed using the maximum likelihood method.
Figure 2.
Phylogenetic analysis of the partial nucleic acid sequence of the G gene. Detections of aMPV/B from Morocco are shaded in yellow whereas vaccine strains are framed in blue. Data were analyzed using the maximum likelihood method.
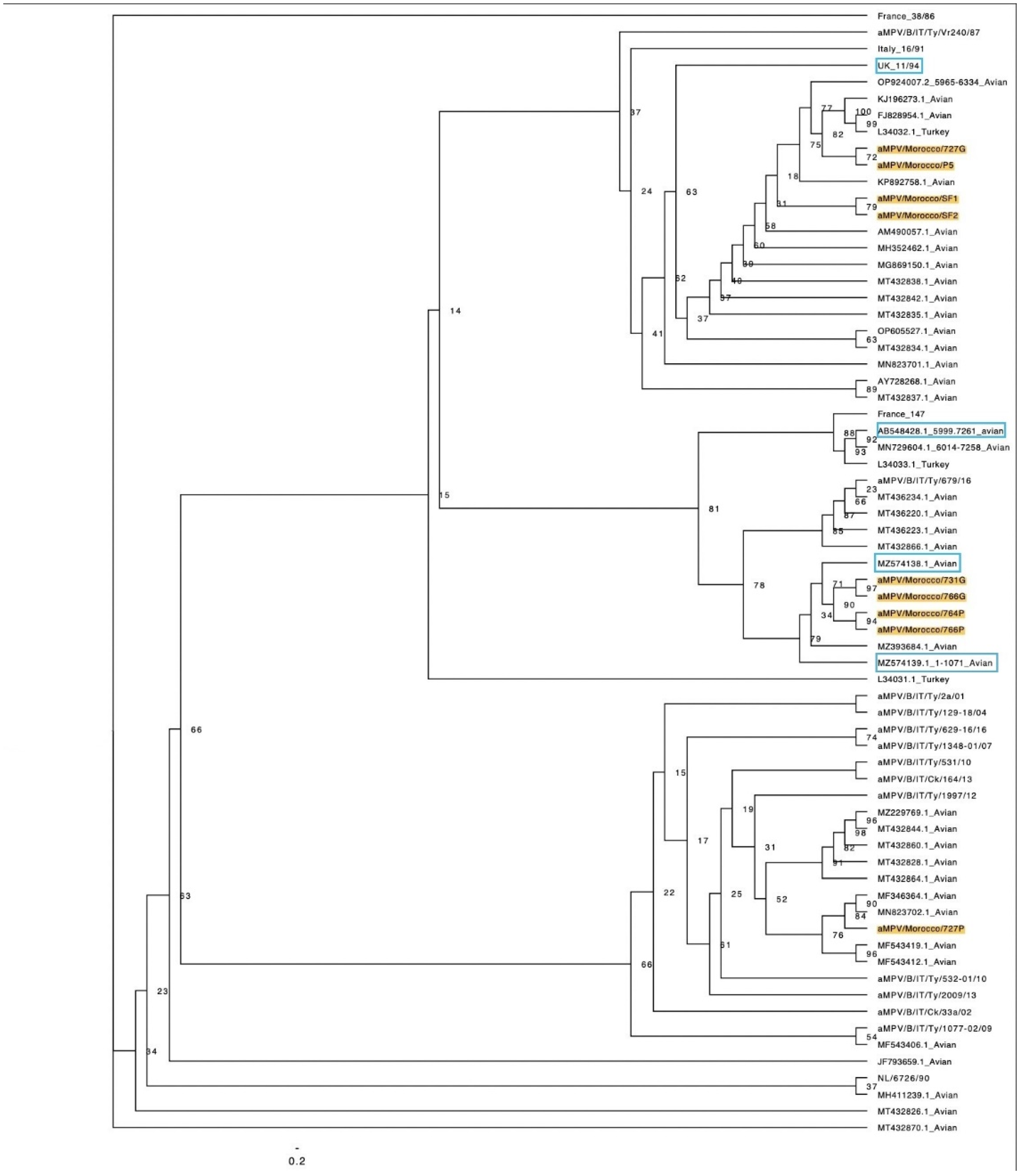

Table 1.
RNA Samples taken between March 2022 and April 2023, and subject to PCR and sequencing.
| Sample identification | Number of samples | Origin | Nature of RNA samples |
|---|---|---|---|
| P1 to P22 | 22 | Broilers | Extracted from aMPV-positive swabs [16] |
| 727 to 731 | 05 | Broilers | Extracted from birds with SHS |
| 764 to 766 | 03 | Turkeys | Extracted from birds with TRT |
Table 2.
Sequences of G gene primers used for nested conventional PCR.
| Primers | Sequence (5’-3’) | References |
|---|---|---|
| G Start+ | CAAGTATCCAGATGGGGTC | [17] |
| G5- | CAAAGAA/G CCAATAAGCCCA | [18] |
| G6- | CTGACAAATTGGTCCTGATT | [18] |
| G8+A | CACTCACTGTTAGCGTCATA | [18] |
| G9+B | TAGTCCTCAAGCAAGTCCTC | [18] |
Table 3.
Protocol of the aMPV Nested PCR and reagent details.
| 1st amplification | 2nd amplification | |||
|---|---|---|---|---|
| Steps | Composition | Volume (µl) | Composition | Volume (µl) |
| Preparation of the PCR mix | Mastermix DreamTaq® Green PCR | 25 | Mastermix DreamTaq® Green PCR | 25 |
| Forward G6- | 2 | Forward G5- | 2 | |
| Reverse G START+ | 2 | Reverse G8+A Reverse G9+B |
2 2 |
|
| H2O | 11 | H2O | 9 | |
| cDNA | 10 | cDNA | 10 | |
| Total | 40 | Total | 40 | |
| Hybridization | 57°C | 58°C | ||
Table 4.
Nucleotide sequences submitted with their respective accession numbers.
| Code | Sequence name on the submission | Accession Number |
|---|---|---|
| 727G | aMPV/B/Morocco/Ck/MA-1/2023 | PQ202992 |
| 727P | aMPV/B/Morocco/Ck/MA-2/2023 | PQ202991 |
| 731G | aMPV/B/Morocco/Ck/MA-3/2023 | PQ202994 |
| 764P | aMPV/B/Morocco/Ty/MA-4/2023 | PQ202998 |
| 766G | aMPV/B/Morocco/Ty/MA-5/2023 | PQ202995 |
| 766P | aMPV/B/Morocco/Ty/MA-6/2023 | PQ202996 |
| P5 | aMPV/B/Morocco/Ck/MA-7/2023 | PQ202993 |
| SF1 | aMPV/B/Morocco/Ck/FS-1/2023 | PQ202997 |
Table 5.
Percentages of similarity between aMPV Moroccan isolates and reference strains.
| Code | Documented GenBank strains | Similarity (%) |
Accession Number | Ref. |
|---|---|---|---|---|
|
727G |
Avian Metapneumovirus strain aMPV/B/Romania/Ty/67/17 glycoprotein (G) gene, partial cds Avian Metapneumovirus isolate 2018_0404_Chicken_Turkey_2018 attachment protein (G) gene, partial cds |
90.76 |
MT432878.1 MH352465.1 |
[19] [20] |
|
727P |
Avian Metapneumovirus isolate Algeria/26/aMPVB/turkey attachment protein gene, partial cds Avian Metapneumovirus partial mRNA for attachment protein (G gene), strain aMPV/chicken/Nigeria/NIR89/2006 |
96.79 96.09 |
KP892758.1 AM490057.1 |
[14] [21] |
|
731G |
Avian Metapneumovirus isolate aMPV-B/BR/1890/E1/19, complete genome Avian Metapneumovirus strain aMPV/B/Italy/Ty/742-01/17 glycoprotein (G) gene, partial cds Avian Metapneumovirus isolate 101/2011 attachment protein (G) gene, partial cds |
99.05 |
OP572408.1 MT436229.1 KC954647.1 |
[22] [19] |
|
764P |
Avian Metapneumovirus strain aMPV/B/France/GuineaFowl/1060/18 glycoprotein (G) gene, partial cds Avian Metapneumovirus strain aMPV/B/Romania/Ty/85/17 glycoprotein (G) gene, partial cds |
98.96 98.63 |
MT432904.1 MT432883.1 |
[19] [19] |
|
766G |
Avian Metapneumovirus isolate aMPV-B/BR/1890/E1/19, complete genome Avian Metapneumovirus strain aMPV/B/Italy/Ty/742-01/17 glycoprotein (G) gene, partial cds Avian Metapneumovirus isolate 101/2011 attachment protein (G) gene, partial cds |
99.70 |
OP572408.1 MT436229.1 KC954647.1 |
[22] [19] |
|
766P |
Avian Metapneumovirus isolate aMPV-B/BR/1890/E1/19, complete genome Avian Metapneumovirus strain aMPV/B/Italy/Ty/742-01/17 glycoprotein (G) gene, partial cds Avian Metapneumovirus isolate 101/2011 attachment protein (G) gene, partial cds |
98.70 |
OP572408.1 MT436229.1 KC954647.1 |
[22] [19] |
|
P5 |
Avian Metapneumovirus isolate aMPV/Chicken/PCRLAB/HG/2010 attachment protein (G) gene, partial cds Avian Metapneumovirus isolate 101/2011 attachment protein (G) gene, partial cds |
96.76 |
MN108496.1 KC954647.1 |
|
|
SF1 |
Avian Metapneumovirus strain aMPV/B/Spain/Ty/4954-1/15 glycoprotein (G) gene, partial cds Avian Metapneumovirus strain aMPV/B/France/Ck/785/17 glycoprotein (G) gene, partial cds Avian Metapneumovirus isolate aMPV-B/turkey/VA/USA/ADRDL-5, complete genome |
99.10 |
MT432835.1 MT432891.1 PP273460.1 |
[19] [19] [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



