
Submitted:
12 September 2024
Posted:
13 September 2024
You are already at the latest version
Abstract
Nitrogen is essential for plant growth, yield and crop quality, and its deficiency limits food production worldwide. In addition, excessive fertilization and inefficient use of N can increase production costs and cause environmental problems. A possible solution to this problem is the application of biofertilizers, which improve N assimilation and enhance biomass and yield. Therefore, the objective of this research was to evaluate the impact of the application of the combination of green and red algal (Ulva lactuca, Solieria spp.), Rhizobium sp., Trichoderma asperel-lum and the combination of the above three biofertilizers on N assimilation. A completely ran-domized design was carried out with 10 plants per treatment and five treatments: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Analyses showed that the Rhi-zobium sp. treatment resulted in higher nitrate reductase enzyme activity quantified in maize leaves, which possibly favored more efficient phosphosynthetic activity, reflecting higher bio-mass accumulation and increased yield. The use of Rhizobium sp. showed increases in biomass (13.4%), yield (11.82%), SPAD values (12%), total chlorophyll (18.4%), carotenoids (13.6%), num-ber of leaves (11.4%) and plant height (11.27%) compared to the control. This results in nitrogen assimilation promises to be a key mechanism for sustainable agricultural practices in the future.
Keywords:
algal extracts
; biofertilizers
; nitrogen assimilation
; Rhizobium sp.
; Trichoderma asperellum
; Zea mays L.
1. Introduction
Maize (Zea mays L.) is the cereal most used for human consumption in Mexico, being a fundamental food item in the daily diet [1]. The largest maize producer in the world is the USA with 348.75 million tons, while Mexico ranks sixth with 26.63 million tons [2]. To achieve high yields, chemical nitrogen (N) fertilizers are widely used in intensive maize production systems, due to the crop’s high N demand [3,4].
Nitrogen (N) is essential for plant growth, yield and crop quality and N deficiency constrains food production worldwide [5]. N is crucial for the synthesis of amino acids, proteins and nucleic acids and a key element in many physiological processes [6]. N assimilation is a complex process which begins with the uptake of nitrate (NO3-) and ammonium (NH4+), the main inorganic forms that plants can absorb. Once accumulated inside the root, the nitrate reductase enzyme (NR) reduces (NO3-) to (NO2-), then the nitrite reductase enzyme (NiR) reduces (NO2-) to (NH4+), which is then converted in organic N compounds [7,8]. Among all the enzymes that participate in N assimilation, NR is the key player in regulating N incorporation in plants [9]. Thus, improving N uptake and assimilation can boost yields and crop quality. However, excessive N fertilization and low N use efficiency can lead to increased production costs and environmental concerns [10].
A potential solution to this issue is the application of biofertilizers, which improve soil fertility [11] and enhance plant growth and yield [12]. Biofertilizers contain microorganisms or natural substances [13] that enhance soil properties, influence plant growth and increase productivity both under optimal conditions and stress, without harming the environment [14]. An example of these biofertilizers, Plant Growth Promoting Fungi (PGPF) and Plant Growth Promoting Bacteria (PGPB) have beneficial microorganisms often used as biofertilizers aiding in plant growth [15], and can work as biological control agents [16,17,18]. Previous studies have demonstrated the effective use of biofertilizers as a sustainable strategy to improve plant N uptake [19,20,21]. PGBG can facilitate the absorption of crucial nutrients such as N, phosphorous (P) and iron (Fe) or regulate plant hormones including auxin, cytokinin to promote plant growth [22]. Some of the most important genera of PGPB are Rhizobium, Azotobacter, Bacillus and Azospirillum [23,24].
Algal, essential photosynthetic microorganisms abundant in N and potassium, are present in all terrestrial environments. They are used in agriculture as biobased fertilizers to enhance soil nutrients, thereby promoting plant growth and increasing yield [25,26]. The levels (2, 4 and 6 kg L-1 of water) of seaweed extract (brown seaweed Ecklonia maxima) significantly affected all growth traits (plant height, stem diameter, number of cobs per plant, cob seeds weight) and yield of maize in both the spring and autumn seasons [27]. Several authors reported the use of seaweed extract (Gracilaria edulis, Laminaria sp., A. nodosum, and K. alvarezii) in maize crops, with increases in seed germination percentage and rate, seedling vigor, shoot and root growth, net carbon assimilation, and total grain yield [28,29,30]. No information is available in the published literature on the foliar application of seaweed extracts of Ulva lactuca and Solieria spp.
Several species of the genus Rhizobium produce nodules on the roots of legumes, and endophytes that can reside within the interior tissues of plants [31]. Under certain conditions, these bacteria act as phosphate solubilizers, fix N and produce hormones in association with non-legume crops [32]. According to [33] Rhizobium can enhance plant growth directly by regulating the before mentioned plant hormones, which modulate root architecture and growth. Indeed, N absorption and utilization is dependent on root length, root surface area and root biomass [32]. The inoculation of maize crop with Rhizobium sp. from various rhizospheres (rice rhizosphere isolate, maize rhizosphere isolate, soybean rhizosphere isolate, peanut rhizosphere isolate, and edamame rhizosphere isolate) showed a significant effect on plant height, stem diameter, husked ear weight, and dry ear weight compared to controls [33]. Additionally, significant increases in maize grain yield were reported when the crop was inoculated with several species of PGPB (Azospirillum brasilense, Rhizobium tropici, Bradyrhizobium sp. and Rhizobium sp.) [34,35].
Species of the genus Trichoderma are commonly found across the globe [36]. These microorganisms inhabit nearly all soil types and frequently function as plant mycoparasites, saprotrophs and symbionts [37,38]. In a similar way to PGBG, Trichoderma inoculation improved N accessibility [39] by releasing organic acid in the soil. It has been observed that certain Trichoderma species provide benefits to crops, including enhanced plant growth, better seedling emergence and improved seed germination [40]. As plant growth regulators, Trichoderma spp. can produce phytohormones [41,42] such as gibberellic acid (GA3) [43] and indole-3-acetic acid (IAA) [44], which are essential for plant growth. These phytohormones increase flowering, photosynthesis efficiency, yield quality, and improve root condition and structure, enhancing seed germination and viability [45]. Earlier studies in maize utilized T. asperellum and Bacillus amyloliquefaciens to treat seeds and plants in greenhouse, resulting in increased plant growth (shoot, root and seedling length) [46]. In another experiment in maize, authors used T. harzianum and B. subtilis to treat seeds and plants, observing increased plant growth and yield [47].
In general, there is scarce literature on the use of biofertilizers based on algal extracts (Ulva lactuca and Solieria spp.), Rhizobium sp. and Trichoderma asperellum in hybrid maize. Therefore, the objective of the present study was to evaluate the impact of the application of the combination of green and red algal (Ulva lactuca, Solieria spp.), Rhizobium sp., Trichoderma asperellum and the combination of the above three biofertilizers on N assimilation, biomass and yield in hybrid maize.
2. Materials and Methods
2.1. Crop Management
The maize crop was grown in open field. The location was in Lomas del Consuelo Meoqui, Chihuahua, Mexico, from August to December 2022. Hybrid maize seeds (ASGROU A 75-73) were sown in loam soil, in 80 cm furrows, with 3 seeds per linear meter (33.3 cm between plants). Three fertilizations were applied, the first two a month apart and the third 22 days later. The first fertilization was 300 kg ha-1 of urea and 50 kg ha-1 of commercial MAP® 11-52-00, the second one of 20 L ha-1 of commercial Nitro-Sul® and the third one of 150 kg ha-1 of urea dissolved in water and applied in irrigation. The plants were irrigated every 15 days using a rolling irrigation system.
2.2. Experimental Design and Treatments
A completely randomized design was done with five treatments: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=Trichoderma asperellum; T5=T2+T3+T4 using 10 replications per treatment (Figure 1).
Treatment 1 was the control, it was not under the effects of any biofertilizer, it only received the commercial fertilizers.
Treatment 2 was a combination of three green and red algal products commercial, whose composition were Algomel Push® (10 mL L-1 of water); Algomel Proact® (5 mL L-1 of water) and Oceamax® (10 mL L-1 of water), which were foliar applied every 15 days, after the appearance of the first pair of true leaves.
Treatment 3 the bacterium (Rhizobium sp.) was cultured in Potato Dextrose Agar (PDA) medium 72 h to six days, and it was used at 109 colony-forming units - CFU mL−1. These products were provided by Dr. Micah Royan Isaac of the Olmix Company. Rhizobium sp. strains were isolated from root nodules of beans plants (Phaseolus vulgaris L.) collected in Delicias, Chihuahua, México. The disinfection of nodules and the bacterium culture were done by the methodology of [48]. The bacterium isolate was grown in liquid medium for 72 h at 30°C and it was applied at density of 106–107 CFU mL-1.
Treatment 4 T. asperellum biofertilizer was described by [49] and the fungi concentration used was 1×1010 UFM mL-1. The T. asperellum strain used in this work is part of the Facultad de Ciencias Agrícolas y Forestales of Chihuahua, México strain collection and is registered in the GeBank under the accession number MN950427.
Treatment 5 was a combination of T2+T3+T4, the Trichoderma asperellum and Rhizobium sp. was applied to the soil near the rhizosphere, immediately followed by foliar application of the algal extracts.
In all treatments except the control, Trichoderma asperellum and Rhizobium sp. were applied to the soil near the roots of the plants at a rate of 20 L ha-1 every 15 days.
2.3. Plant Sampling
At physiological maturity of the plants, 100 days after sowing, plant samples were collected and the roots, aerial parts and cob were separated from the 10 plants of each treatment, then carefully washed with distilled water. The fresh plant material was used for analysis of biomass, yield, nitrate reductase enzyme activity and photosynthetic pigments.
2.4. Plant Analysis
2.4.1. Total Biomass and Yield
After 24 h of washing with distilled water, the roots, aerial parts and cob of each plant were weighed on a precision balance (Luna LBL 34001e Precision Balance, Milton Keynes, UK). Biomass was expressed as the sum of all plant organs (g plant-1 f.w.). Yield was expressed as the fresh weight of ears per plant (g plant-1 f.w.).
2.4.2. Nitrate Reductase Enzyme Activity (NR Activity) “In Vivo”
The nitrate reductase enzyme (NR, EC 1.7.1.1) activity was determined by the methodology suggested by [50], where 0.1 g of fresh 7 mm diameter leaves discs was weighted and placed in 10 mL of incubation buffer (100 mM K-phosphate buffer, pH 7.5 and 1% (v/v) propanol). The samples were infiltrated at a pressure of 0.8 bars (NAPCO 5851 vacuum oven, Winchester, VA, USA) and they were incubated at 30 °C in the dark for 1 h (WIG-50 digital incubator, DAIHAN SCIENTIFIC, Seoul, Republic of Korea) after that, placed in a (FELISA FE-371 boiling water bath, Zapopan, Jalisco, México) for 15 min to stop NR activity. Then, 1 mL of enzyme extract was taken and 2 mL of 1% (w/v) sulphanilamide in 1.5 M HCl and 2 mL of 0.02% (w/v) N-(1-naphthyl)-ethylenediamine dichlorohydrate was added (20 mg of NNEDA dissolved in 100 mL of distilled water). Finally, the resulting nitrite concentration was determined spectrophotometrically (Thermo Fisher Scientific, GENESYS™ 10S, Madison, WI, USA) at 540 nm against a NO2− standard curve. The results were expressed as µmol NO2− formed g−1 f.w. h−1.
2.4.3. SPAD Values
SPAD index (Soil Plant Analysis Development) values were determined using the SPAD-502 portable chlorophyllometer (Konica Minolta Sensing, Inc., Osaka, Japan), which provides a quantitative assessment of leaf green intensity [51], measurements were conducted around midday (during peak light hours), with three random readings taken per plant. Two readings were made with the SPAD-502 meter, one on October 15, 2022, before grain filling stage and the second one on November 10, 2022, before maturation stage.
2.4.4. Photosynthetic Pigments
The methodology of [52] was used for photosynthetic pigments analysis. Leaves sample stalks of 7 mm in diameter, with approximately 0.125 g of weight from each treatment, were placed in test tubes and 10 mL of methanol were added to each sample. The samples were left to rest for 24 h in the dark. After this time the samples were read at wavelengths of 470, 653 and 666 nm in a Genesis 10S UV-VIS spectrophotometer (Thermo Fisher Scientific, GENESYS™ 10S, Madison, WI, USA). The results were expressed in μg cm2 of fresh weight and the following formulas were used (Equations 1, 2, 3 and 4):
where Vf: final volume; W1: weight per leaf disc; W2: total weight of leaf discs; r: radius of the leaf discs; n: number of leaf discs.
Chl a* = [15.65(A666) - 7.34(A653)]
Chl a = (Chl a* x Vf x W1)/(W2 x π x r2 x n)
Chl a = (Chl a* x Vf x W1)/(W2 x π x r2 x n)
Chl b* = [27.05(A653) -11.21(A666)]
Chl b = (Chl b* x Vf x W1)/(W2 x π x r2 x n)
Chl b = (Chl b* x Vf x W1)/(W2 x π x r2 x n)
Carotenoids* = [(1000*A470) - 2.86(Chl a) -129.2(Chl b)]/ (221)
Carotenoids = (Carotenoids* x V x W1)/(W2 x π x r2 x n)
Carotenoids = (Carotenoids* x V x W1)/(W2 x π x r2 x n)
Total chlorophyll = Chl a + Chl b
2.4.5. Number of Leaves and Plant Height
The number of leaves was determined by manual counting. The plant height refers to the measurement taken from the highest point to the base of naturally growing plants, and it was determined with a measuring tape. The results were expressed as cm. Both variables were measured before flowering.
2.5. Statistical Analysis
The normal distribution of the data was examined using the Shapiro-Wilk test (p ≤ 0.05), and the equality of variances among the analyzed variables was assessed via Levene's test, considering significance at (p ≤ 0.05). Following this, ANOVA analysis was conducted at a significance level of 0.05. The variables were then subjected to Duncan's test to compare the means of the two factor levels (dose and application time) at a significance level of (p≤ 0.05). The statistical program SAS version 9.0 was used.
3. Results and Discussion
3.1. Biomass and Yield
Biomass is a crucial indicator for crops, as it reflects the total amount of organic matter accumulated by plants, reflecting crop yield potential [53]. In this experiment, all treatments displayed significant differences of biomass compared to control. The highest biomass occurred in T3 (Rhizobium sp.) followed by T2 (algal extracts), T4 (T. asperellum) and T5 (combination of the three biofertilizers) (Figure 2). The application of the bacterium Rhizobium sp. induced an increment of the results of 13.4 % compared to the control. The other treatments (algal extracts, T. asperellum and the combination of all biofertilizers) had increases of 11.39 %, 11.18 % and 11.52 %, compared to the control respectively. Several researchers have reported the application of Rhizobium sp. as PGPR (Plant Growth Promoting Rhizobacteria) in other crops [24,54,55] because Rhizobium sp. like other rhizobacteria, effectively colonizes the roots of non-leguminous plants. It can solubilize phosphorus, produces siderophores, plant hormones, and also shows antagonistic effects against plant pathogens [56,57]. In the present study, the application of Rhizobium sp. improved the biomass. In physiological terms, this phenomenon may arise in maize because, by receiving an additional supply of N through biological N fixation, it can increase its rate of photosynthesis and its capacity to produce more plant matter. The combined treatment (T5) also showed high biomass, suggesting that synergistic interactions between the biofertilizers were optimizing the uptake of N and other nutrients, thus maximizing biomass production. As reported by [58,59,60], the use of biofertilizers promotes biological N fixation, making a more efficient use of N and increases nutrient uptake, generating more biomass.
Crop yields are widely recognized as a key indicator for global food security [61]. In this experiment, the highest yields were obtained with (T3) Rhizobium sp., and the combination of algal extracts, Rhizobium sp. and T. asperellum (T5) as a management practice for hybrid maize cultivation. These treatments were significantly different from the rest. Treatments 2 (algal extracts) and 4 (T. asperellum) were not different to the control (Figure 3). Additionally, when applying Rhizobium sp. (T3) on hybrid maize crop, an increase in yields of 11.82 % was obtained and when combining the three biofertilizers the increase was 11.60 % compared to the control, respectively. In the case of maize, which has a high N demand, the presence of Rhizobium sp. could be a relevant factor that allows plants to avoid N deficiencies and favor fruit filling [62]. Furthermore, according to [63] the application of Rhizobium sp. reduces the internal competition of resources between vegetative and reproductive growth, optimizing both processes and favoring the final yield in terms of grain production. According to [64,65], the use of biofertilizers and their synergy promote nutrient uptake and thus improve yield.
3.2. Nitrate Reductase Enzyme Activity (NR Activity) “In Vivo”
The NR enzyme catalyzes the first step of nitrate assimilation in higher plants, so its activity is a key and limiting process for N assimilation in plants [66]. In this experiment (Figure 4) showed the NR activity in different treatments (T1, T2, T3, T4 and T5), measured in μmol NO₂- g-¹ f.w. h-¹. The control, algal extracts treatment and Rhizobium sp. present similar activities, above 20 μmol NO₂- g-¹ f.w. h-¹, with no significant differences between them. This represents approximately 20 % more activity compared to T4 and T5. Treatments T4 (T. asperellum) and T5 (algal extracts, Rhizobium sp. and T. asperellum) showed a NR activity below 20 μmol NO₂- g-¹ f.w. h-¹, representing a 20 % reduction in activity compared to treatments T1, T2 and T3, being significantly lower.
In this research it was observed that, as NR activity decreases (particularly at T5, where algal extracts, Rhizobium sp., and Trichoderma asperellum are combined), biomass and yield are not compromised; in fact, significant improvements in these parameters are observed (Figure 2 and Figure 3). This suggests that treatments T4 and T5 had an effect in which the uptake of inorganic N could be facilitated, making the process more efficient in the pathway where the NR enzyme is key. This agrees with that reported by [67], where Trichoderma spp. applied to Sebacinales spp. and their interaction with other biofertilizers, can improve N uptake in plants. This may be due to several factors related to how microorganisms modify the rhizosphere environment and nitrogen assimilation pathways in plants. T. asperellum is a fungus known for its ability to promote plant growth and improve nutrient uptake efficiency through phosphate solubilization and the production of enzymes that degrade organic matter, thus providing organic nitrogen sources to the plant [98]. The availability of organic forms of nitrogen, such as amino acids or peptides, may reduce the need for inorganic nitrate as the main source of N, which would explain the lower activity of NR in this treatment [68]. In addition, the combination of algal extracts, Rhizobium sp., and T. asperellum (T5) could be providing increased availability of organic nitrogen and promoting direct assimilation of ammonium or reduced forms of nitrogen, which decreases the need for the plant to use nitrate and, therefore, reduces NR activity. This synergy between biofertilizers may favor alternative metabolic pathways that are more efficient in nitrogen incorporation, which optimizes biomass production without relying so much on nitrate reduction [69]. In the case of algal [70], he examines how algal extracts provide not only alternative sources of N, but also bioactive compounds that improve N assimilation efficiency and promote growth and yield in crops, such as maize, by overexpression of N transporter genes. In the case of T3 (Rhizobium sp.), the highest NR activity was obtained in this work, although there is no significant difference with the control, where it is possible to observe that the fungi positively favored N assimilation in the metabolic pathway of NR. Rhizobium sp. not only acts as an atmospheric nitrogen fixer but is also a biostimulant to produce auxins, plant hormones essential for root growth. According to what was reported by [71], auxins promote cell elongation and the formation of lateral roots, which increases the nutrient uptake area in plants. These findings reinforce the results obtained in this experiment, where inoculation with Rhizobium sp. led to a significant increase in nitrate reductase (NR) activity. Similarly, previous research has shown that different Rhizobium species, such as Mesorhizobium ciceri and Rhizobium phaseoli, promote not only the growth of legumes, but also of non-leguminous crops such as maize. These studies show that improved nutrient uptake and plant yield are related to the ability of these bacteria to stimulate root growth mechanisms and the assimilation of inorganic N [72,73,74].
3.3. SPAD Values
The chlorophyll index is a fast and non-destructive way to detect chlorophyll levels in plants and thus form an idea of their nutritional situation [75].The index of chlorophyll in hybrid maize (ASGROU A 75-73) was affected by the application of the biofertilizers used (Figure 5). This variable allows to know the levels of chlorophyll in the leaves and thus give an idea of the efficiency of the photosynthetic process.
The higher result was observed with the application of Rhizobium sp. compared to the other treatments, with a significant increase of 12% over the control. The treatments with T. asperellum and the combination of all biofertilizers did not show significant differences between them, although they were significantly different to the control, showing increases of 11.45 % (T4) and 11.35 % (T5) over it, respectively. The lowest SPAD values were observed in the control and the algal extracts. Previous research has documented the increment in chlorophyll content (SPAD values), photosynthetic pigments, biomass and yield after application of biofertilizers to maize crops [76,77,78,79]. Referring to the content of chlorophylls (SPAD values, the highest values in this research were obtained when applying Rhizobium sp., likewise, in terms of total chlorophylls and yield, the values were similar when applying the bacterium and the combination of the three biofertilizers. In this context [80] inoculated Rhizobium sp. in maize crops, resulting in an increase on shoot relative chlorophyll content, dry weight, photosynthetic rate, leaf area and grain yield. Hussain et al. [81] reported a rhizobial strain identified as Rhizobium phaseoli that was used in maize under drought conditions induced by polyethylene glycol and even under this situation the inoculation with rhizobia led to a significant increase in chlorophyll levels. Comparable results were described by [82] in fodder maize inoculated with rhizobia in field conditions, noting significant increases in chlorophyll content, photosynthesis, and transpiration rate.
3.4. Photosynthetic Pigments
Photosynthetic pigments are molecules that capture light, transfer it to the reaction centers to be converted chemical energy, enabling photosynthesis [83]. These pigments can be divided into total chlorophyll (chlorophyll a and chlorophyll b) and total carotenoids. Chlorophyll is the main pigment for light absorption meanwhile carotenoids have a support role. N is essential for the synthesis of photosynthetic pigments; thus, a continuous and adequate supply of N is necessary [84]. In this experiment, the application of Rhizobium sp. in maize plants and the combination of the three biofertilizers induced higher chlorophyll content with significant differences compared to the other treatments. Likewise, the highest values for carotenes were obtained with Rhizobium sp. In both cases, the lowest pigments content occurred in the control (Figure 6). The buildup in chlorophyll content observed in the Rhizobium sp. treatment and the combination of the three treatments compared to the control was 18.4 % and the treatment (T5) with combination of biofertilizers had an increase of 15.98 % compared to control. Regarding carotenes, treatment with Rhizobium sp., and biofertilizers combination (T5) had the highest significant values compared to control (13.6%). Photosynthesis is one of the most important processes in plants that determine biomass and yield [85,86]. This increase could be attributed to improved NR activity (Figure 4) but also boosted N fixation and uptake. It is also possible that siderophores (iron-chelating molecules) released by Rhizobium sp. increased Fe accessibility for plants [87]. Fe is also a key component for chlorophyll synthesis, redox reactions and several biochemical reactions, for example N fixation. Algal effects on crops include increased N, availability from soil and improved photosynthetic pigment content due to the presence of phytohormones that share similar regulatory roles with higher plants [88,89]. T. asperellum could up-regulate photosynthetic pigment concentration by the up-regulation of genes modulating chlorophyll and carotenoid biosynthesis or the response against reactive oxygen species [90,91]. The secondary metabolites released by Trichoderma sp. may influence plant nutrient uptake and growth by activating phytohormones consisting of indole acetic acid or auxin [92].
3.5. Number of Leaves and Plant Height
Leaves have key roles in photosynthesis, carbon fixation, respiration and transpiration [93]. The number of leaves in hybrid maize plants (ASGROU A 75-73) was higher with the application of Rhizobium sp. with significant differences compared to the other treatments (Figure 7).
The hybrid maize plants increased their leaf numbers in 11.4 % compared to the control when Rhizobium sp. was used (T3). The combination of the three biofertilizers (T5) increased this variable in a 11.1 % followed by the application of T. asperellum (T4) with a 10% increase. Regarding plant height, results did not show significant differences when using Rhizobium sp. (T3) and the three combined biofertilizers (T5) (Figure 8), obtaining increases of 11.27 % and 11.21 % respectively when compared to the control. The lowest value was obtained with the algal extracts (T2). In the other hand, the control (T1) and the plants treated with T. asperellum (T4) were statistically similar.
In this investigation, the application of Rhizobium sp. to the soil increased the number of leaves and height of maize plants, most likely because Rhizobium has different mechanisms to promote plant growth. These include direct mechanisms such as N fixation, phosphate solubilization and the production of phytohormones [82]. An increased N fixation due to Rhizobium can improve plant growth, leading to more biomass accumulation, as noted in alfalfa [94]. Another trait of Rhizobium is the capacity to transform soluble phosphate from their insoluble form via the release of different organic acids (acetic acid, oxalic acid and gluconic acid) or by proton (H+) extrusion. P is essential not only for the formation of organic compounds (DNA, RNA, ATP), but it also is involved in cell division and energy transfer processes [95]. The production of phytohormones by various bacteria is also an important trait and these comprise gibberellins, indole acetic acid, auxins, cytokinins and abscisic acid, which are crucial for plant and leaf growth and stem elongation in plants [82]. Apart from phytohormones, enzymes including phytase and protease can also be synthetized, enhancing nutrient uptake [96]. Moreover, Rhizobium can stimulate root growth, facilitating the absorption of crucial elements such as Fe and P. [97]. Similar results were found by [98] where application of Rhizobium (R. etli bv. Phaseoli, R. leguminosarum bv. Trifolii) and Sinorhizobium sp. improved plant height, corn growth, and grain yield of maize in several areas. In another research, the inoculation of maize with several Rhizobium species (Cp3, Lt2 and Br3), boosted plant height from 101.0 to 104.0 cm [99]. In addition, [33] upon the application of Rhizobium sp. to maize crops, reported a rise not only in plant height but also in stem diameter, ear dry weight and ear weight without husks compared to control. In the present experiment the biggest height of the plants ranged from 135-134 cm (T3-T5).
3.6. Correlation Analysis and Heat Map
The Pearson's heat map and correlation (Figure 9), allows us to observe the dependence or interdependence relationships between the variables analyzed. The color scale indicates that the shades closer to red are strong positive correlations (close to +1), while the bluish shades show negative correlations (close to -1) and white or light color suggests no or very low correlation.
The map shows that leaf number and yield have an almost perfect correlation (0.99), indicating that the higher the number of leaves, the higher the yield increases significantly. Chlorophyll a and carotenoids have a very high correlation (0.95), suggesting that the presence of one of these pigments is closely related to the other. Chlorophyll a and biomass also have a strong correlation (0.94), indicating that biomass accumulation is directly related to the amount of chlorophyll in plants. The number of leaves and plant height (0.9) are strongly correlated, indicating that the higher the number of leaves, the higher the plant height. Plant height and yield (0.86) and plant height and number of leaves (0.9) have strong positive correlations, suggesting that taller plants tend to generate higher yields and have more leaves. Biomass is also strongly correlated with yield (0.75), indicating that an increase in biomass is related to higher plant yield. In addition, in the case of nitrate reductase activity had weak negative correlations with variables such as plant height (-0.46), number of leaves (-0.13), yield (-0.14) and biomass (0.35), suggesting that an increase in the activity of this enzyme may be slightly related to a decrease in these variables, this is probably due to the effect of the biofertilizers applied.
3.7. Venn and Interaction Diagram
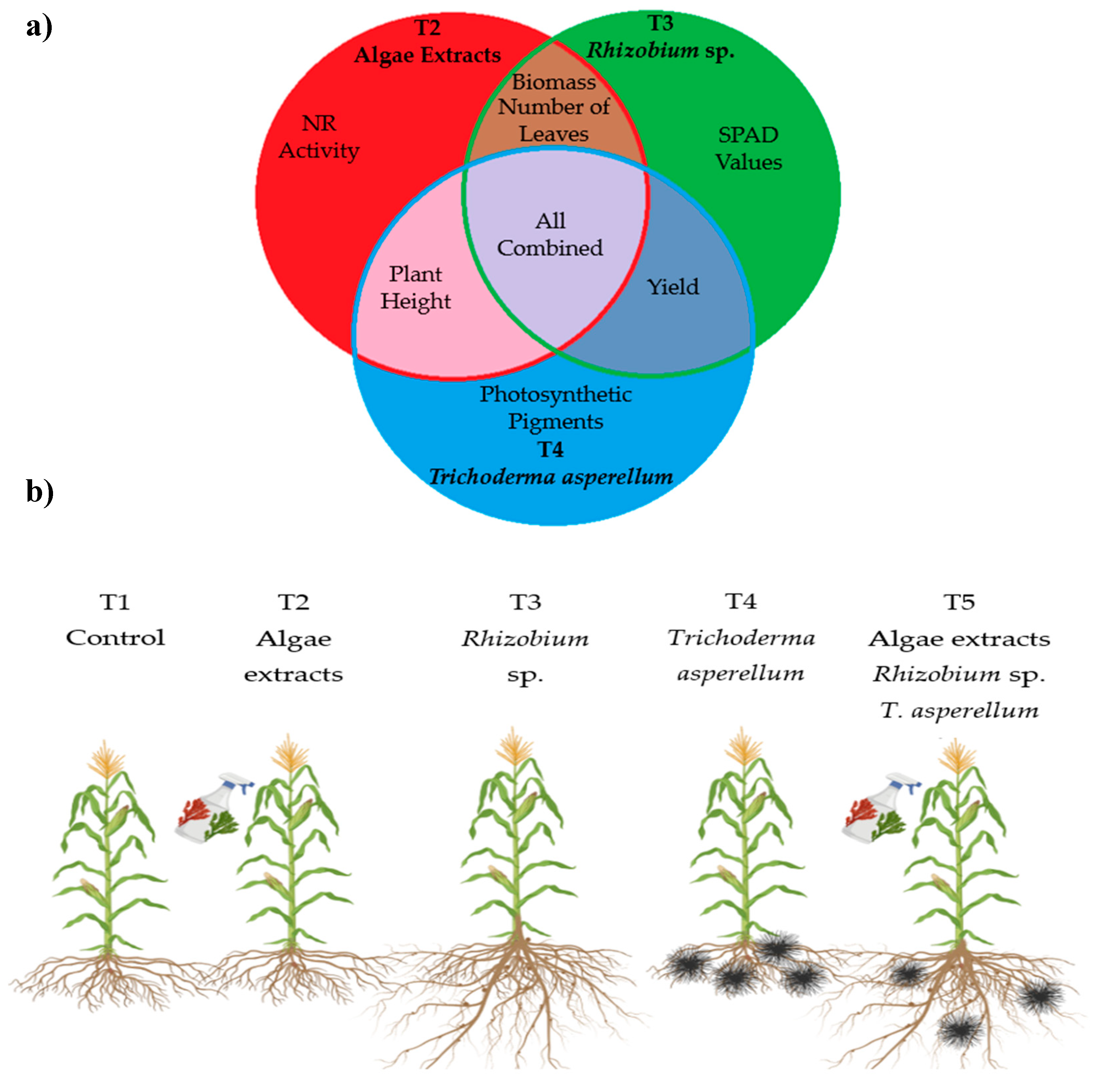
Figure 10 is a clear and concise representation of how different biofertilizer treatments (algal extracts, Rhizobium sp. and Trichoderma asperellum) directly impact the growth, physiology and yield of hybrid maize.
Part (a): Venn Diagram
This diagram illustrates the synergy between the different biological treatments and how, by combining them, outstanding effects on critical variables in plant growth are obtained. The intersections of the diagram show that T2 (algal extracts) is particularly associated with an increase in nitrate reductase activity (NR Activity), which is crucial for nitrogen assimilation and thus for plant growth.
T3 (Rhizobium sp.) stands out in the increase of SPAD values, which are related to the amount of chlorophyll in the leaves and, therefore, to photosynthetic efficiency. T4 (Trichoderma asperellum) favors the production of photosynthetic pigments, which is essential for light absorption and efficiency in the photosynthetic process. The central zone of the diagram, which combines the three treatments (T5), shows improvements in biomass, number of leaves, plant height and yield, suggesting that the joint application of these microorganisms and plant extracts could be more effective than the individual treatments. This combination not only optimizes resource use, but also tends to maximize plant growth and productivity.
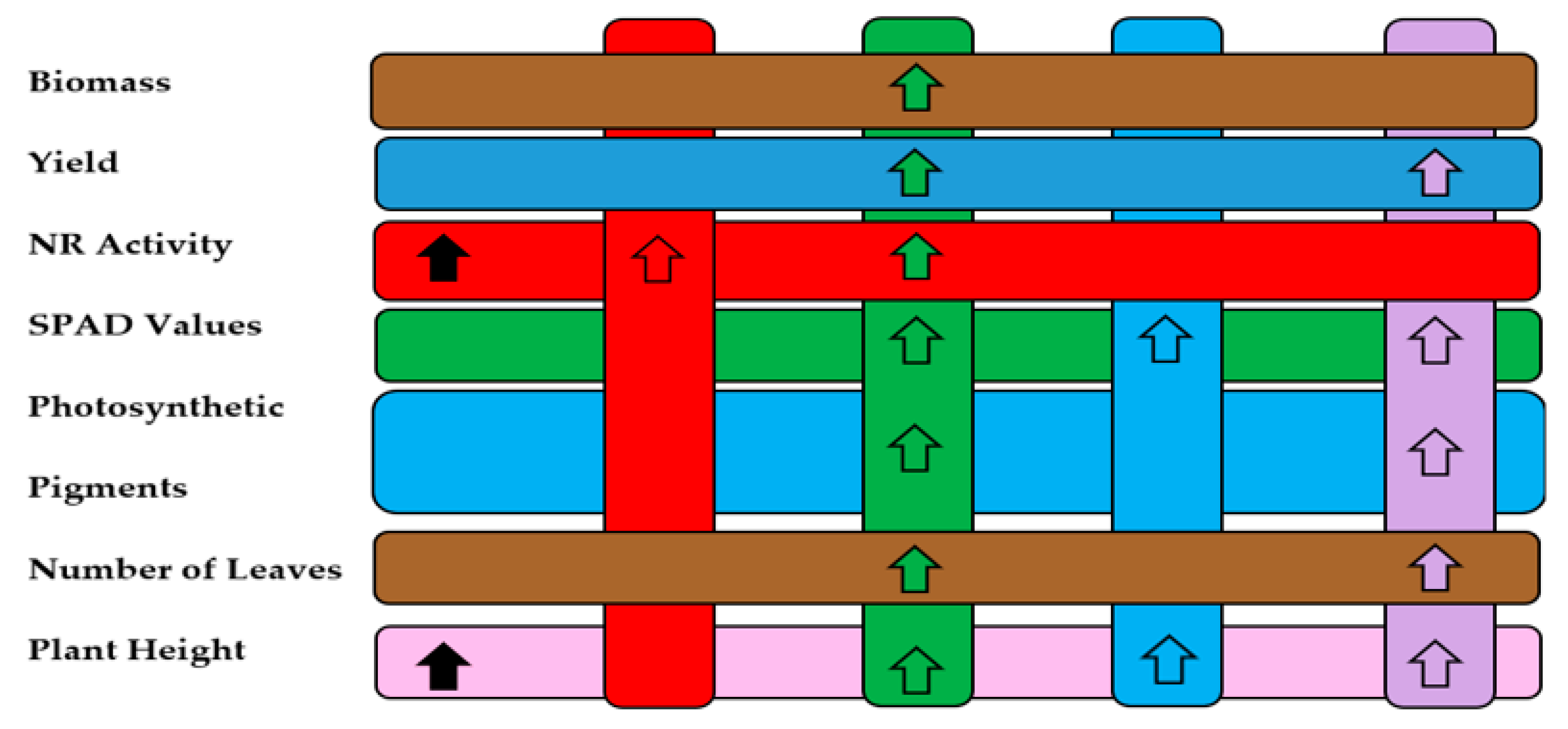
Part (b): Bar chart
This bar chart quantifies the impact of each treatment (T1 to T5) on key variables. Analyzing biomass, the combined treatment (T3) generates the greatest increase, implying that the application of Rhizobium sp. on hybrid corn significantly enhances overall plant growth. In yield, as with biomass, the T3 treatment offers the highest results, which is a crucial contribution to sustainable agricultural productivity. In NR activity, a notorious increase is observed with the T2 (algal extracts) and T3 (Rhizobium sp.) treatments, indicating that these biofertilizers improve the efficiency of nitrogen assimilation, an essential process for plant metabolism. In SPAD values: T3 (Rhizobium sp.) and T5 presented the greatest increases, suggesting an improvement in photosynthetic efficiency, key to biomass production and yield. The variable photosynthetic pigments, the treatments with Rhizobium sp. (T3) and the combination of all (T5) promote the production of essential pigments for photosynthesis, which also contributes to better growth and yield. As for the number of leaves and plant height, all biofertilizers, except T2 (algal extracts) increased considerably.
4. Conclusions
In this experiment, the application of Rhizobium sp. to the rhizosphere in hybrid maize plants had a greater effect on nitrogen assimilation, thanks to the symbiotic relationship generated with the maize. Analyses showed that the Rhizobium sp. treatment resulted in higher nitrate reductase enzyme activity quantified in maize leaves, which possibly favored more efficient photosynthetic activity, reflecting higher biomass accumulation and increased yield. The use of Rhizobium sp. showed increases in biomass (13.4%), yield (11.82%), SPAD values (12%), total chlorophyll content (18.4%), carotenoids (13.6%), number of leaves (11.4%) and plant height (11.27%) compared to the control. It is important to mention that treatment T5, which was the combination of algal extracts, Rhizobium sp. and Trichoderma asperellum also had very positive effects on nitrogen assimilation. However, more studies are needed on the synergistic relationship they form in terms of nitrogen assimilation and nitrogen use efficiency. Finally, the use of Rhizobium sp, and the combination of biofertilizers such as algal extracts, Rhizobium sp. and Trichoderma asperellum, in relation to nitrogen assimilation and nitrogen use efficiency have been little explored, and this research adds a new dimension to the field of study by demonstrating that these biofertilizers produce a very effective synergistic effect on nitrogen assimilation, which promises to be a key mechanism for sustainable agricultural practices in the future.
Author Contributions
Conceptualization, S.P.A. and E.S.; methodology, S.P.A, E.H.O.C. and J.C.A.-P.; formal analysis, C.M.E.B.; investigation, S.P.; writing—original draft preparation, S.P.A. and E.H.O.C.; writing—review and editing, J.R.S. and M.A.M.T.; visualization, R.H.C and E.H.O.C.; supervision, L.P.U.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Acknowledgments
I would like to thank Dr. Micah Royan Isaac of the Olmix Company for the algal extracts supplied for the completion of this work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rosas-Castor, J.M.; Guzmán-Mar, J.L.; Hernández-Ramírez, A.; Garza-González, M.T.; Hinojosa-Reyes, L. Arsenic Accumulation in Maize Crop (Zea mays): A Review. Sci. Total Environ. 2014, 488–489, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Statista. Available online: https://es.statista.com/estadisticas/613419/prinicpales-productores-de-maiz-en-el-mundo/ (accessed on 30 May 2024).
- Hou, P.; Gao, Q.; Xie, R.; Li, S.; Meng, Q.; Kirkby, E.A.; Chen, X. Grain yields in relation to N requirement: Optimizing nitrogen management for spring maize grown in China. Field Crops Res. 2012, 129, 1–6. [Google Scholar] [CrossRef]
- Lai, Z.; Zhang, H.; Ding, X.; Liao, Z.; Zhang, C.; Yu, J.; Fan, J. Ridge-furrow film mulch with nitrogen fertilization improves grain yield of dryland maize by promoting root growth, plant nitrogen uptake and remobilization. Soil Tillage Res. 2024, 241, 106118. [Google Scholar] [CrossRef]
- Mejías, J.H.; Salazar, F.; Pérez Amaro, L.; Hube, S.; Rodriguez, M.; Alfaro, M. Nanofertilizers: A cutting-edge approach to increase nitrogen use efficiency in grasslands. Front. Environ. Sci. 2021, 9, 635114. [Google Scholar] [CrossRef]
- Bai, F.; Qi, X.; Li, P.; Du, Z.; Guo, W. Groundwater depth and nitrogen application amount jointly regulate the water and residual soil nitrate accumulation in agricultural soil profile. Agronomy 2023, 13, 1163. [Google Scholar] [CrossRef]
- Wang, Y. Effect of NPK on NR and NiR activity of sugar beet. J. Nucl. Agric. Sci. 2012, 26, 0803–0808. [Google Scholar]
- Liu, X.; Hu, B.; Chu, C. Nitrogen assimilation in plants: current status and futurs prospects. J. Genet. Genomics 2022, 49, 394–404. [Google Scholar] [CrossRef]
- Zhang, W.; Ni, K.; Long, L.; Ruan, J. Nitrogen transport and assimilation in tea plant (Camellia sinensis): A review. Front. Plant Sci. 2023, 14, 1249202. [Google Scholar] [CrossRef]
- Yadav, M.R.; Kumar, S.; Lal, M.K.; Kumar, D.; Kumar, R.; Yadav, R.K.; Kumar, S.; Nanda, G.; Singh, J.; Udawat, P.; et al. Mechanistic Understanding of Leakage and Consequences and Recent Technological Advances in Improving Nitrogen Use Efficiency in Cereals. Agronomy 2023, 13, 527. [Google Scholar] [CrossRef]
- Nosheen, S.; Ajmal, I.; Song, Y. Microbes as Biofertilizers, a Potential Approach for Sustainable Crop Production. Sustainability 2021, 13, 1868. [Google Scholar] [CrossRef]
- Ammar, E.E.; Aioub, A.A.A.; Elesawy, A.E.; Karkour, A.M.; Mouhamed, M.S.; Amer, A.A.; EL-Shershaby, N.A. Algae as BioFertilizers: Between Current Situation and Future Prospective: The Role of Algae as a Biofertilizer in Serving of Eco-system. Saudi J. Biol. Sci. 2022, 29, 3083–3096. [Google Scholar] [CrossRef] [PubMed]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal Biostimulants and Biofertilisers in Crop Productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef]
- Tolisano, C.; Del Buono, D. Biobased: Biostimulants and Biogenic Nanoparticles Enter the Scene. Sci. Total Environ. 2023, 885, 163912. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of Action of Plant Growth Promoting Bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Y.; Ding, Z.; Bai, Y. Contribution of Microbial Inter-Kingdom Balance to Plant Health. Mol. Plant 2019, 12, 148–149. [Google Scholar] [CrossRef]
- El-Maraghy, S.S.; Tohamy, A.T.; Hussein, K.A. Plant Protection Properties of the Plant Growth Promoting Fungi (PGPF): Mechanisms and Potentiality. Curr. Res. Environ. Appl. Mycol. 2021, 11, 391–415. [Google Scholar] [CrossRef]
- Cao, M.; Narayanan, M.; Shi, X.; Chen, X.; Li, Z.; Ma, Y. Optimistic Contributions of Plant Growth-Promoting Bacteria for Sustainable Agriculture and Climate Stress Alleviation. Environ. Res. 2023, 15, 217. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of Phosphate-Solubilizing Microorganisms in Sustainable Agriculture—A Review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant Growth-Promoting Rhizobacteria (PGPR): Emergence in Agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Saharan, B.S.; Nehra, V. Plant Growth Promoting Rhizobacteria: A Critical Review. Life Sci. Med. Res. 2011, 2011, 1–30. [Google Scholar]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed]
- Deka, H.; Deka, S.; Baruah, C. Plant Growth Promoting Rhizobacteria for Value Addition: Mechanism of Action. In Plant-Growth-Promoting Rhizobacteria (PGPR) and Medicinal Plants, 1st ed.; Egamberdieva, D., Shrivastava, S., Varma, A., Eds.; Springer: New York, USA, 2015; pp. 305–321. [Google Scholar]
- Dodd, I.C.; Zinovkina, N.Y.; Safronova, V.I.; Belimov, A.A. Rhizobacterial Mediation of Plant Hormone Status. Ann. Appl. Biol. 2010, 157, 361–379. [Google Scholar] [CrossRef]
- Abdel-Raouf, N.; Al-Homaidan, A.A.; Ibraheem, I.B. Agricultural Importance of Algae. Afr. J. Biotechnol. 2012, 11, 11648–11658. [Google Scholar] [CrossRef]
- Barone, G.D.; Cernava, T.; Ullmann, J.; Liu, J.; Lio, E.; Germann, A.T.; Nakielski, A.; Russo, D.A.; Chavkin, T.; Knufmann, K.; Tripodi, F.; Coccetti, P.; Secundo, F.; Fu, P.; Pfleger, B.; Axmann, I.M.; Lindblad, P. Recent Developments in the Production and Utilization of Photosynthetic Microorganisms for Food Applications. Heliyon 2023, 9, e14708. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghazal, S.A.Y.; Aziz, M.M.; AL-Juheiehy, W.K.S. Response of Growth and Yield of Corn (Zea mays L.) to Biofertilizer and Sea-Algae Extract. Int. J. Agric. Stat. Sci. 2023, 19, 161–165. [Google Scholar]
- Basavaraja, P.K.; Yogendra, N.D.; Zodape, S.T.; Prakash, R.; Ghosh, A. Effect of Seaweed Sap as Foliar Spray on Growth and Yield of Hybrid Maize. J. Plant Nutr. 2018, 41, 1851–1861. [Google Scholar] [CrossRef]
- Layek, J.; Das, A.; Ramkrushna, G.I.; Ghosh, A.; Panwar, A.S.; Krishnappa, R.; Ngachan, S.V. Effect of Seaweed Sap on Germination, Growth and Productivity of Maize (Zea mays) in North Eastern Himalayas. Indian J. Agron. 2016, 61, 354–359. [Google Scholar] [CrossRef]
- Pal, A.; Dwivedi, S.K.; Maurya, P.K.; Kanwar, P. Effect of Seaweed Saps on Growth, Yield, Nutrient Uptake and Economic Improvement of Maize (Sweet Corn). J. Appl. Nat. Sci. 2015, 7, 970–975. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda, M.C.; Glick, B.R. Plant Growth-Promoting Bacterial Endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Zhou, S.; Xia, P.; Chen, J.; Xiong, Q.; Li, G.; Tian, J.; Wu, B.; Zhou, F. Optimizing Nitrogen Application Position to Change Root Distribution in Soil and Regulate Maize Growth and Yield Formation in a Wide–Narrow Row Cropping System: Pot and Field Experiments. Front. Plant Sci. 2024, 15, 1298249. [Google Scholar] [CrossRef]
- Widodo, T.W.; Muhklisin, I.; Nugroho, S.A.; Wardana, R.; Ummah, U.S.A. Growth and Yield of Maize Applicated by Rhizobium spp. from Legume and Non-legume Rhizosphere. J. Agric. Appl. Biol. 2023, 4, 151–160. [Google Scholar] [CrossRef]
- Marks, B.B.; Megías, M.; Ollero, F.J.; Nogueira, M.A.; Araujo, R.S.; Hungria, M. Maize Growth Promotion by Inoculation with Azospirillum brasilense and Metabolites of Rhizobium tropici Enriched on Lipo-chitooligosaccharides (LCOs). AMB Express 2015, 5, 71. [Google Scholar] [CrossRef] [PubMed]
- Pessoa Cavalcanti, M.I.; de Carvalho Nascimento, R.; Ribeiro, D.R.; Costa Escobar, I.E.; Resende, A.C.F.; Pereira, A.S.; Santiago, A.D.F.; Abrahão, R.S.N.; Fernandes-Júnior, P.V. Maize Growth and Yield Promoting Endophytes Isolated into a Legume Root Nodule by a Cross-Over Approach. Rhizosphere 2020, 15, 100211. [Google Scholar] [CrossRef]
- Kubiak, A.; Wolna-Maruwka, A.; Pilarska, A.A.; Niewiadomska, A.; Piotrowska-Cyplik, A. Fungi of the Trichoderma Genus: Future Perspectives of Benefits in Sustainable Agriculture. Appl. Sci. 2023, 13, 6434. [Google Scholar] [CrossRef]
- Alfiky, A.; Weisskopf, L. Deciphering Trichoderma–Plant–Pathogen Interactions for Better Development of Biocontrol Applications. J. Fungi 2021, 7, 61. [Google Scholar] [CrossRef]
- Chagas, L.F.B.; Castro, H.G.; Colonia, B.S.O.; Carvalho-Filho, M.R.; Miller, L.O.; Chagas-Junior, A.F. Efficiency of the Inoculation of Trichoderma asperellum UFT-201 in Cowpea Production Components under Growth Conditions in Field. Rev. Ciênc. Agrár. 2016, 39, 413–421. [Google Scholar] [CrossRef]
- Wei, Y.; Yang, H.; Hu, J.; Li, H.; Zhao, Z.; Wu, Y.; Li, J.; Zhou, Y.; Yang, K.; Yang, H. Trichoderma harzianum Inoculation Promotes Sweet Sorghum Growth in the Saline Soil by Modulating Rhizosphere Available Nutrients and Bacterial Community. Front. Plant Sci. 2023, 14, 1258131. [Google Scholar] [CrossRef]
- Cai, W.J.; Ye, T.T.; Wang, Q.; Cai, B.D.; Feng, Y.Q. A Rapid Approach to Investigate Spatiotemporal Distribution of Phytohormones in Rice. Plant Methods 2016, 12, 47. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Majewska, M.; Hanaka, A.; Tyśkiewicz, K.; Pawlik, A.; Janusz, G. Phytohormones (Auxin, Gibberellin) and ACC Deaminase In Vitro Synthesized by the Mycoparasitic Trichoderma DEMTkZ3A0 Strain and Changes in the Level of Auxin and Plant Resistance Markers in Wheat Seedlings Inoculated with This Strain Conidia. Int. J. Mol. Sci. 2019, 20, 4923. [Google Scholar] [CrossRef]
- Díaz, G.; Rodríguez, G.; Montana, L.; Miranda, T.; Basso, C.; Arcia, M. Efecto de la aplicación de bioestimulantes y Trichoderma sobre el crecimiento en plántulas de maracuyá (Passiflora edulis Sims) en vivero. Bioagro 2020, 32, 195–204. [Google Scholar]
- Sabre, W.I.; Ghoneem, K.M.; Rashad, Y.M.; Al-Askar, A.A. Trichoderma harzianum WKY1: An Indole Acetic Acid Producer for Growth Improvement and Anthracnose Disease Control in Sorghum. Biocontrol Sci. Technol. 2017, 27, 654–676. [Google Scholar] [CrossRef]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of Two Trichoderma Strains on Plant Growth, Rhizosphere Soil Nutrients and Fungal Community of Pinus sylvestris var. mongolica Annual Seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef]
- Karuppiah, V.; Vallikkannu, M.; Li, T.; Chen, J. Simultaneous and Sequential-Based Co-Fermentations of Trichoderma asperellum GDFS1009 and Bacillus amyloliquefaciens 1841: A Strategy to Enhance the Gene Expression and Metabolites to Improve the Biocontrol and Plant Growth Promoting Activity. Microb. Cell Factories 2019, 18, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Petcu, V.; Bubueanu, C.; Casarica, A.; Săvoiu, G.; Stoica, R.; Bazdoaca, C.; Lazăr, D.A.; Iordan, H.L.; Horhocea, D. Efficacy of Trichoderma harzianum and Bacillus subtilis as Seed and Vegetation Application Combined with Integrated Agroecology Measures on Maize. Romanian Agric. Res. 2023, 40, 1–10. [Google Scholar] [CrossRef]
- Abdel-Raouf, N.; Al-Homaidan, A.A.; Ibraheem, I.B. Agricultural Importance of Algae. Afr. J. Biotechnol. 2012, 11, 11648–11658. [Google Scholar] [CrossRef]
- Díaz, F.G.; Gutiérrez, R.T.; Mora, K.G.; García, M.C.N. Isolation and Characterization of Rhizobia from Crotalaria sp. in Southern Ecuador. Cultivos Tropicales 2016, 37, 40–47. [Google Scholar]
- Andrzejak, R.; Janowska, B.; Renska, B.; Kosiada, T. Effect of Trichoderma spp. and Fertilization on the Flowering of Begonia × tuberhybrida Voss. ‘Picotee Sunburst’. Agronomy 2022, 11, 1278. [Google Scholar] [CrossRef]
- Sánchez, E.; Rivero, R.M.; Ruiz, J.M.; Romero, L. Changes in Biomass, Enzymatic Activity and Protein Concentration in Roots and Leaves of Green Bean Plants (Phaseolus vulgaris L. cv. Strike) under High NH₄NO₃ Application Rates. Sci. Hortic. 2004, 99, 237–248. [Google Scholar] [CrossRef]
- Cunha, A.R.D.; Katz, I.; Sousa, A.D.P.; Martinez-Uribe, R.A. Índice SPAD en el Crecimiento y Desarrollo de Plantas de Lisianthus en Función de Diferentes Dosis de Nitrógeno en Ambiente Protegido. Idesia 2015, 33, 97–105. [Google Scholar] [CrossRef]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Bakuei, N.; Amini, G.; Njafpour, G.D.; Jahanshahi, M. Total and Sustainable Utilization of Biomass Resources: A Perspective. Front. Sustain. Energy 2015. [Google Scholar]
- Razafintsalama, H.; Trap, J.; Rabary, B.; Razakatiana, A.T.E.; Ramanankierana, H.; Rabeharisoa, L.; Becquer, T. Effect of Rhizobium Inoculation on Growth of Common Bean in Low-Fertility Tropical Soil Amended with Phosphorus and Lime. Sustainability 2022, 14, 4907. [Google Scholar] [CrossRef]
- Kandil, A.E.; Özdamar Ünlü, H. Effect of Rhizobium Inoculation on Yield and Some Quality Properties of Fresh Cowpea. Cogent Food Agric. 2023, 9, 2275410. [Google Scholar] [CrossRef]
- Díez-Méndez, A.; Menéndez, E. Rhizobium Presence and Functions in Microbiomes of Non-Leguminous Plants. In Symbiotic Soil Microorganisms, 1st ed.; Shrivastava, N., Mahajan, S., Varma, A., Eds.; Springer: Switzerland, 2021; pp. 241–266. [Google Scholar]
- Beltran-Medina, J.I.; Romero-Perdomo, F.; Molano-Chavez, L.; Silva, A.M.M.; Estrada-Bonilla, G.A. Differential Plant Growth Promotion Under Reduced Phosphate Rates in Two Genotypes of Maize by a Rhizobial Phosphate-Solubilizing Strain. Front. Sustain. Food Syst. 2022, 6, 955473. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Enhancing Nitrogen Use Efficiency in Crop Plants. Adv. Agron. 2005, 88, 97–185. [Google Scholar]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The Evolution and Future of Earth's Nitrogen Cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef]
- Chalk, P.M.; Souza, R.F. The Role of Biological Nitrogen Fixation in the Nitrogen Cycle of Sugarcane and Maize Cropping Systems. Symbiosis 2019, 78, 105–118. [Google Scholar]
- Albahri, G.; Alyamani, A.A.; Badran, A.; Hijazi, A.; Nasser, M.; Maresca, M.; Baydoun, E. Enhancing Essential Grains Yield for Sustainable Food Security and Bio-Safe Agriculture Through Latest Innovative Approaches. Agronomy 2023, 13, 1709. [Google Scholar] [CrossRef]
- Meng, L.; Zhang, A.; Wang, F.; Han, X.; Wang, D.; Li, S. Arbuscular Mycorrhizal Fungi and Rhizobium Facilitate Nitrogen Uptake and Transfer in Soybean/Maize Intercropping System. Front. Plant Sci. 2015, 6, 339. [Google Scholar] [CrossRef]
- Zheng, F.; Tan, D. Enhancing Maize Yield and Nutrient Utilization Through Improved Soil Quality Under Reduced Fertilizer Use: The Efficacy of Organic–Inorganic Compound Fertilizer. Agriculture 2024, 14, 1482. [Google Scholar] [CrossRef]
- Kumar, V.; Prasad, R. Role of Biofertilizers in Improving Soil Fertility and Crop Productivity: A Review. J. Pharmacogn. Phytochem. 2021, 10, 634–641. [Google Scholar]
- Xie, J.; Hu, L.; Wang, Z.; Yan, C.; Bao, F. Synergistic Effects of Biofertilizers on Crop Yield and Nutrient Uptake in Maize. Agron. J. 2017, 109, 2732–2740. [Google Scholar]
- Campbell, W.H. Nitrate Reductase Structure, Function and Regulation: Bridging the Gap Between Biochemistry and Physiology. Annu. Rev. Plant Biol. 1999, 50, 277–303. [Google Scholar] [CrossRef] [PubMed]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced Systemic Resistance and Plant Responses to Fungal Biocontrol Agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef]
- Altomare, A.; Burla, M.C.; Camalli, M.; Carrozzini, B.; Cascarano, G.L.; Giacovazzo, C.; Guagliardi, A.; Moliterni, A.G.G.; Polidori, G.; Rizzi, R. EXPO: A Program for Full Powder Pattern Decomposition and Crystal Structure Solution. J. Appl. Crystallogr. 1999, 32, 339–340. [Google Scholar] [CrossRef]
- Khan, A.R.; Ullah, I.; Khan, S.; Fahad, S.; Khan, A.H.; Ali, S.; Shah, A.; Zhou, W. Plant Growth-Promoting Rhizobacteria as an Alternative Tool for Boosting Crop Productivity in Stressful Environments: A Review. Front. Plant Sci. 2018, 9, 1801. [Google Scholar]
- Kaushal, M.; Wani, S.P. Rhizobacterial-Plant Interactions: Strategies Ensuring Plant Growth Promotion Under Drought and Salinity Stress. Agric. Ecosyst. Environ. 2016, 231, 68–78. [Google Scholar]
- Velasquez, S.M.; Barbez, E.; Kleine-Vehn, J.; Estevez, J.M. Auxin and Cellular Elongation. Plant Physiol. 2016, 170, 1206–1215. [Google Scholar] [CrossRef]
- Li, X.; Li, Z. What Determines Symbiotic Nitrogen Fixation Efficiency in Rhizobium: Recent Insights into Rhizobium leguminosarum. Arch. Microbiol. 2023, 205, 300. [Google Scholar] [CrossRef]
- Maitra, S.; Praharaj, S.; Brestic, M.; Sahoo, R.K.; Sagar, L.; Shankar, T.; Palai, J.B.; Sahoo, U.; Pramanick, B.; Nath, S.; Venugopalan, V.K.; Skalický, M.; Hossain, A. Rhizobium as Biotechnological Tools for Green Solutions: An Environment-Friendly Approach for Sustainable Crop Production in the Modern Era of Climate Change. Curr. Microbiol. 2023, 80, 219. [Google Scholar] [CrossRef]
- Mehboob, I.; Zahir, Z.A.; Arshad, M.; Tanveer, A.; Khalid, M. Comparative Effectiveness of Different Rhizobium sp. for Improving Growth and Yield of Maize (Zea mays L.). Soil Environ. 2012, 31, 37–46. [Google Scholar]
- Shrestha, S.; Brueck, H.; Asch, F. Chlorophyll Index, Photochemical Reflectance Index and Chlorophyll Fluorescence Measurements of Rice Leaves Supplied with Different N Levels. J. Photochem. Photobiol. B Biol. 2012, 113, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.S.; Li, Y.J.; Wu, P.T.; Zhao, X.N.; Gao, X.D.; Chen, X.L. Recovery Growth and Water Use of Intercropped Maize Following Wheat Harvest in Wheat/Maize Relay Strip Intercropping. Field Crops Res. 2020, 256, 107924. [Google Scholar] [CrossRef]
- Hu, W.; Lu, Z.; Meng, F.; Li, X.; Cong, R.; Ren, T.; Sharkey, T.D.; Lu, J. The Reduction in Leaf Area Precedes That in Photosynthesis Under Potassium Deficiency: The Importance of Leaf Anatomy. New Phytol. 2020, 227, 1749–1763. [Google Scholar] [CrossRef]
- Mehboob, I.; Zahir, Z.A.; Arshad, M.; Tanveer, A.; Khalid, M. Comparative Effectiveness of Different Rhizobium sp. for Improving Growth and Yield of Maize (Zea mays L.). Soil Environ. 2012, 31, 37–46. [Google Scholar]
- Hussein, M.H.; Eltanahy, E.; Al Bakry, A.F.; Elsafty, N.; Elshamy, M.N. Seaweed Extracts as Prospective Plant Growth Biostimulant and Salinity Stress Alleviator for Vigna sinensis and Zea mays. J. Appl. Phycol. 2021, 33, 1273–1291. [Google Scholar] [CrossRef]
- Kandil, A.E.; Özdamar Ünlü, H. Effect of Rhizobium Inoculation on Yield and Some Quality Properties of Fresh Cowpea. Cogent Food Agric. 2023, 9, 2275410. [Google Scholar] [CrossRef]
- Hussain, M.B.; Zahir, Z.A.; Asghar, H.A.; Mubaraka, R.; Naveed, M. Efficacy of Rhizobia for Improving Photosynthesis, Productivity and Mineral Nutrition of Maize. Clean Soil Air Water 2016, 44, 1564–1571. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd_Allah, E.F.; Hashem, A. Phytohormones and Beneficial Microbes: Essential Components for Plants to Balance Stress and Fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Zhao, J.; Chen, N.; Zhu, T.; Zhao, X.; Yuan, M.; Wang, Z.; Du, H. Simultaneous Quantification and Visualization of Photosynthetic Pigments in Lycopersicon esculentum Mill. Under Different Levels of Nitrogen Application with Visible-Near Infrared Hyperspectral Imaging Technology. Plants 2023, 12, 2956. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.S.; Li, Y.J.; Wu, P.T.; Zhao, X.N.; Gao, X.D.; Chen, X.L. Recovery Growth and Water Use of Intercropped Maize Following Wheat Harvest in Wheat/Maize Relay Strip Intercropping. Field Crops Res. 2020, 256, 107924. [Google Scholar]
- Hu, W.; Lu, Z.; Meng, F.; Li, X.; Cong, R.; Ren, T.; Sharkey, T.D.; Lu, J. The Reduction in Leaf Area Precedes That in Photosynthesis Under Potassium Deficiency: The Importance of Leaf Anatomy. New Phytol. 2020, 227, 1749–1763. [Google Scholar] [CrossRef] [PubMed]
- Gen-Jiménez, A.; Flores-Félix, J.D.; Rincón-Molina, C.I.; Manzano-Gomez, L.A.; Rogel, M.A.; Ruíz-Valdiviezo, V.M.; Rincón-Rosales, R. Enhance of Tomato Production and Induction of Changes on the Organic Profile Mediated by Rhizobium Biofortification. Front. Microbiol. 2023, 14, 1235930. [Google Scholar] [CrossRef]
- Lu, Y.; Xu, J. Phytohormones in Microalgae: A New Opportunity for Microalgal Biotechnology? Trends Plant Sci. 2015, 20, 273–282. [Google Scholar] [CrossRef]
- Ammar, E.E.; Aioub, A.A.A.; Elesawy, A.E.; Karkour, A.M.; Mouhamed, M.S.; Amer, A.A.; El-Shershaby, N.A. Algae as Bio-fertilizers: Between Current Situation and Future Prospective. Saudi J. Biol. Sci. 2022, 29, 3083–3096. [Google Scholar] [CrossRef]
- Doni, F.; Fathurrahman, F.; Mispan, M.S.; Suhaimi, N.S.M.; Yusoff, W.M.W.; Uphoff, N. Transcriptomic Profiling of Rice Seedlings Inoculated with the Symbiotic Fungus Trichoderma asperellum SL2. J. Plant Growth Regul. 2019, 38, 1507–1515. [Google Scholar] [CrossRef]
- Harman, G.; Khadka, R.; Doni, F.; Uphoff, N. Benefits to Plant Health and Productivity from Enhancing Plant Microbial Symbionts. Front. Plant Sci. 2021, 11, 610065. [Google Scholar]
- Vinale, F.; Sivasithamparam, K. Beneficial effects of Trichoderma secondary metabolites on crops. Phytotherapy Research 2020, 34(11), 2835–2842. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, L.; Qi, D. Variation in Leaf Traits at Different Altitudes Reflects the Adaptive Strategy of Plants to Environmental Changes. Ecol. Evol. 2020, 10, 8166–8175. [Google Scholar] [CrossRef]
- Fang, L.; Ju, W.; Yang, C.; Jin, X.; Liu, D.; Li, M.; Zhang, C. Exogenous Application of Signaling Molecules to Enhance the Resistance of Legume-Rhizobium Symbiosis in Pb/Cd-Contaminated Soils. Environ. Pollut. 2020, 265, 114744. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Gahtyari, N.C.; Chhabra, R.; Kumar, D. Role of Microbes in Improving Plant Growth and Soil Health for Sustainable Agriculture. In Advances in Plant Microbiome and Sustainable Agriculture: Diversity and Biotechnological Applications; Springer, 2020; pp. 207–256. [Google Scholar]
- Fahde, S.; Boughribil, S.; Sijilmassi, B.; Amri, A. Rhizobia: A Promising Source of Plant Growth-Promoting Molecules and Their Non-Legume Interactions: Examining Applications and Mechanisms. Agriculture 2023, 13, 1279. [Google Scholar] [CrossRef]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed]
- Hayat, R.; Ahmed, I.; Sheirdil, R.A. An Overview of Plant Growth Promoting Rhizobacteria (PGPR) for Sustainable Agriculture. In Crop Production; Springer, 2016. [Google Scholar]
- Qureshi, M.A.; Shahzad, H.; Imran, Z.; Mushtaq, M.; Akhtar, N.; Ali, M.A.; Mujeeb, F. Potential of Rhizobium Species to Enhance Growth and Fodder Yield of Maize in the Presence and Absence of L-Tryptophan. J. Anim. Plant Sci. 2013, 23, 1448–1454. [Google Scholar]
Figure 1.
Application of biofertilizers: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4.
Figure 1.
Application of biofertilizers: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4.
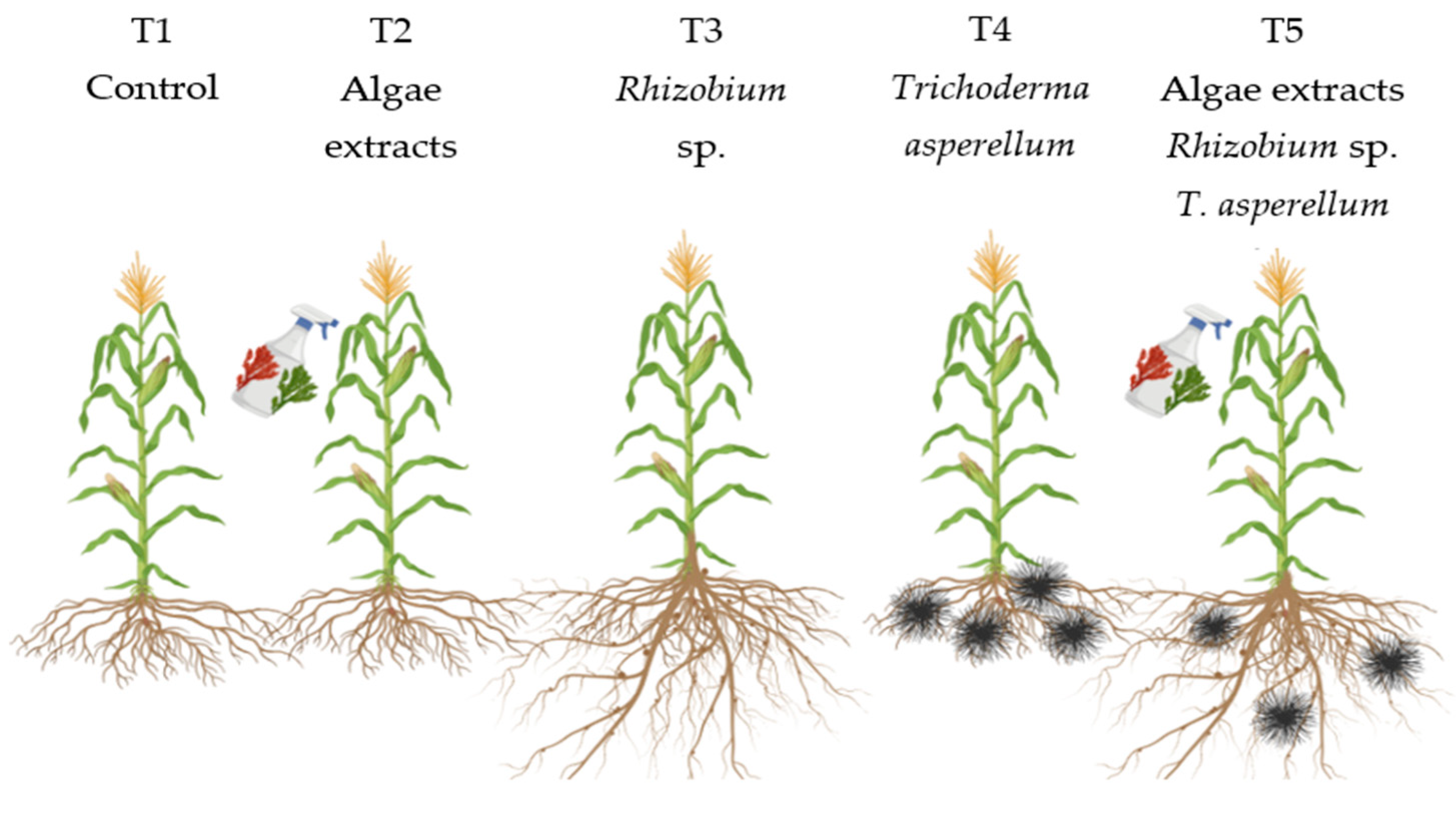
Figure 2.
Effect of biofertilizers application on total biomass where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤0.05).
Figure 2.
Effect of biofertilizers application on total biomass where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤0.05).
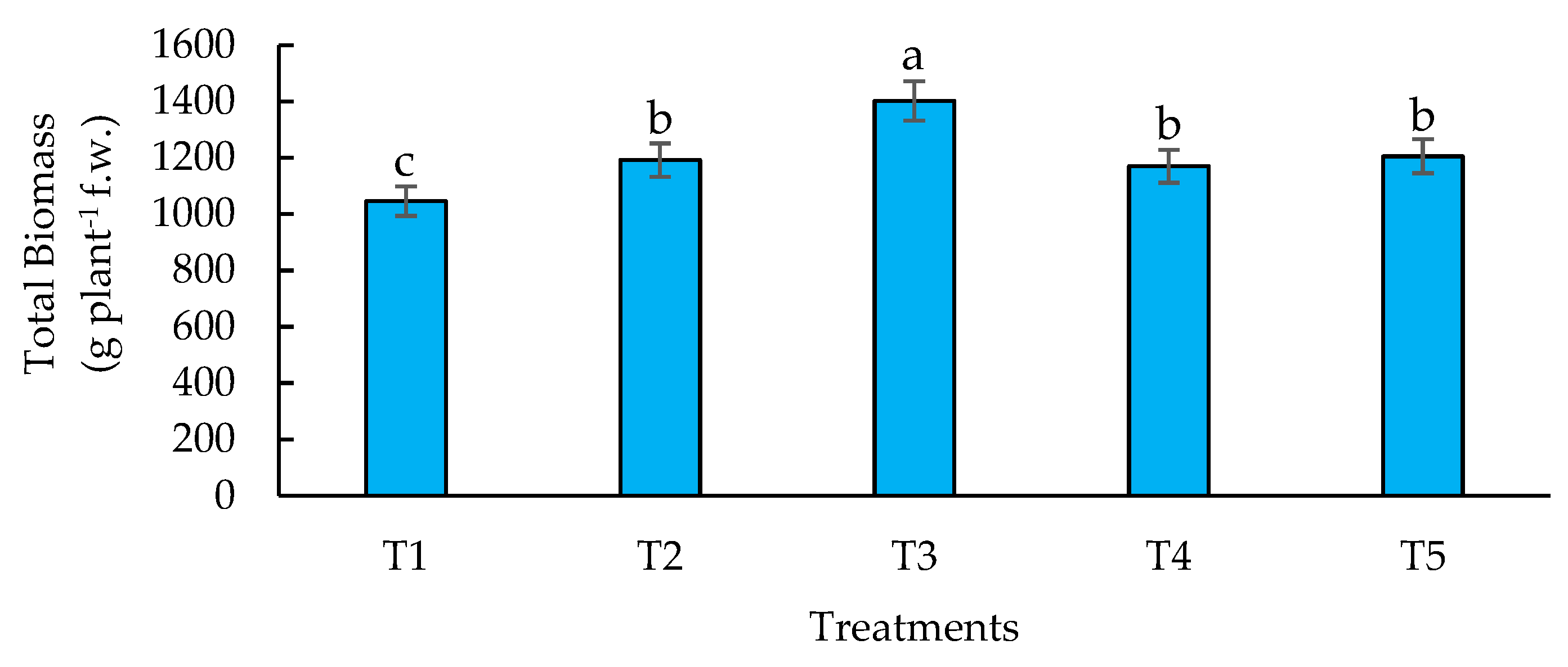
Figure 3.
Effect of biofertilizers application on yield where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤ 0.05).
Figure 3.
Effect of biofertilizers application on yield where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤ 0.05).
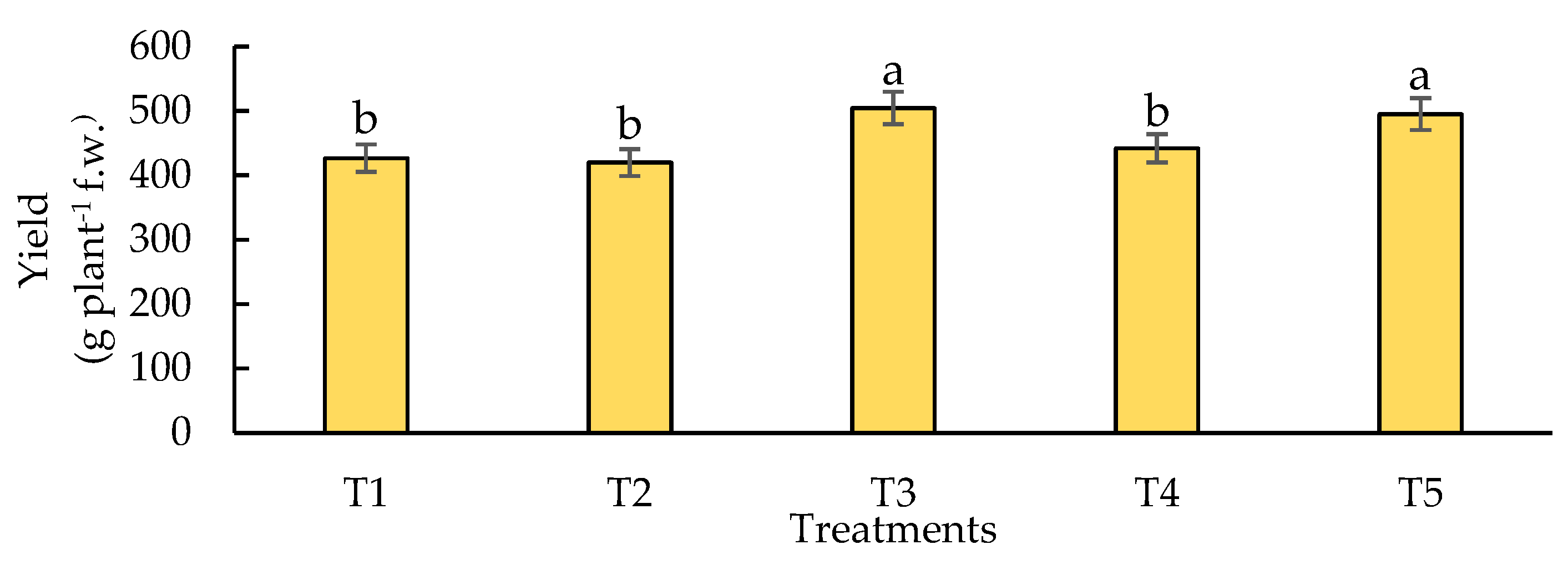
Figure 4.
Effect of biofertilizers application on Nitrate Reductase Activity “in vivo” where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤ 0.05).
Figure 4.
Effect of biofertilizers application on Nitrate Reductase Activity “in vivo” where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤ 0.05).
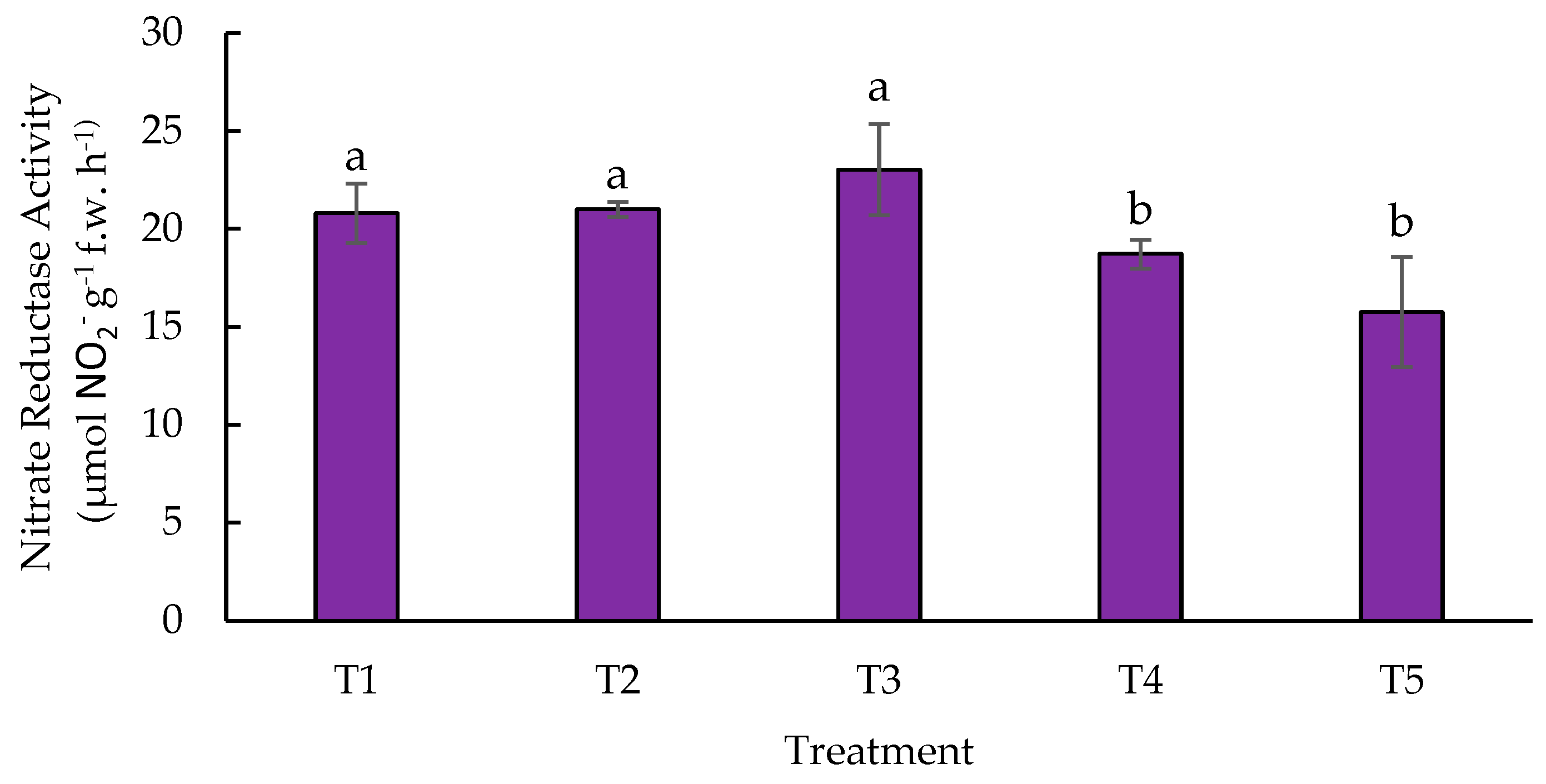
Figure 5.
Effect of biofertilizers application on SPAD values where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤0.05).
Figure 5.
Effect of biofertilizers application on SPAD values where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤0.05).
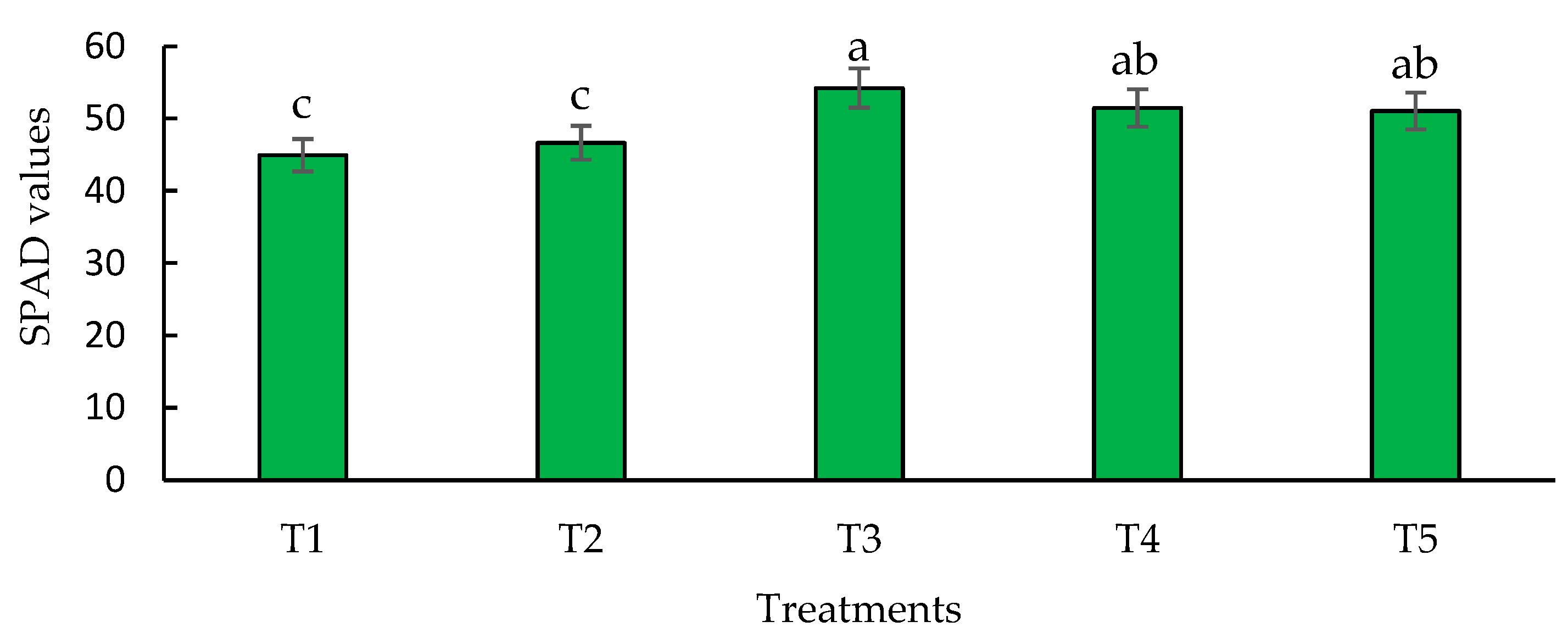
Figure 6.
Effect of application of biofertilizers on photosynthetic pigments activity. Where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤0.05).
Figure 6.
Effect of application of biofertilizers on photosynthetic pigments activity. Where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤0.05).
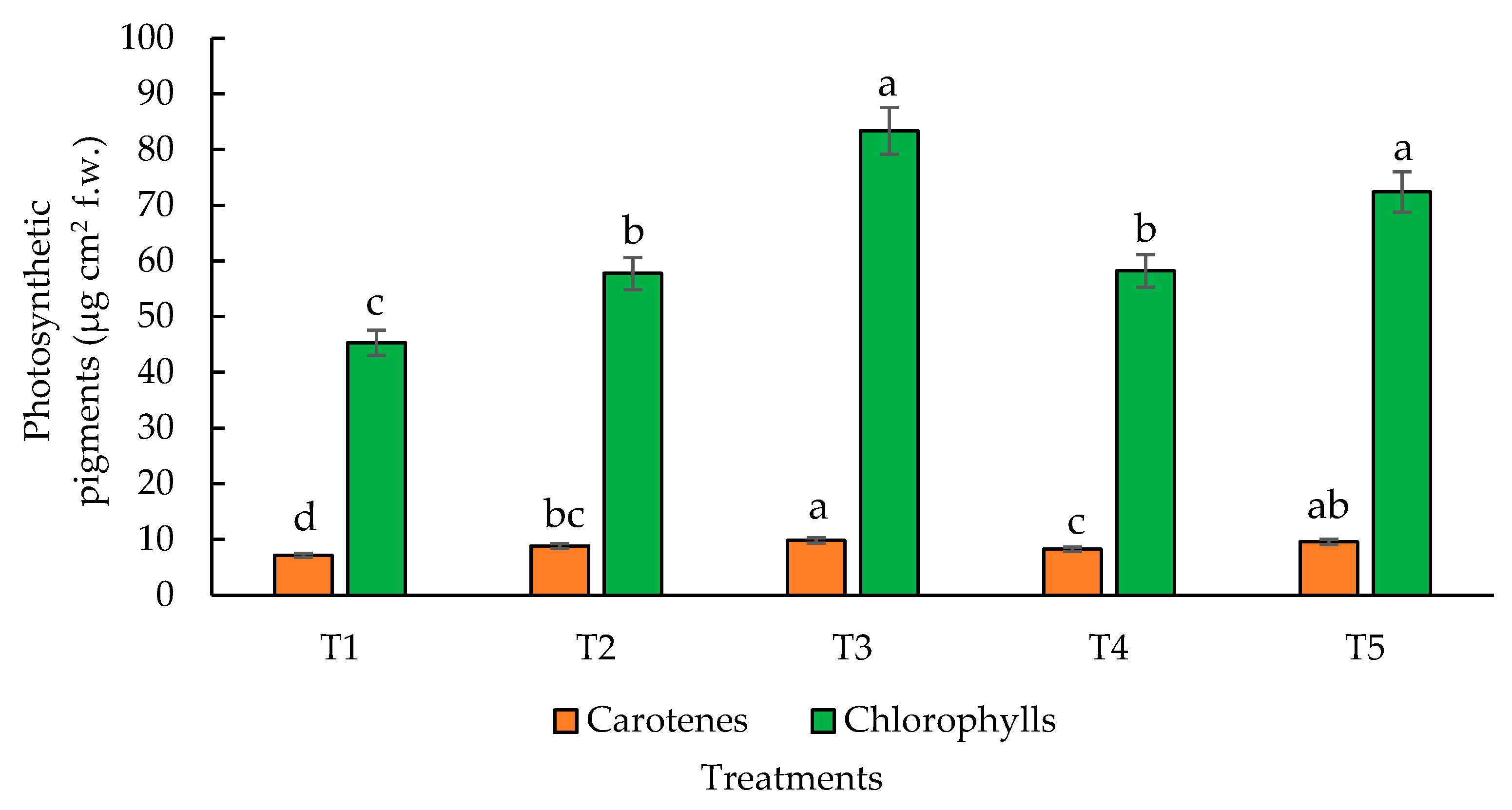
Figure 7.
Effect of biofertilizers application on leaf number where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan multiple range test (p ≤ 0.05).
Figure 7.
Effect of biofertilizers application on leaf number where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan multiple range test (p ≤ 0.05).
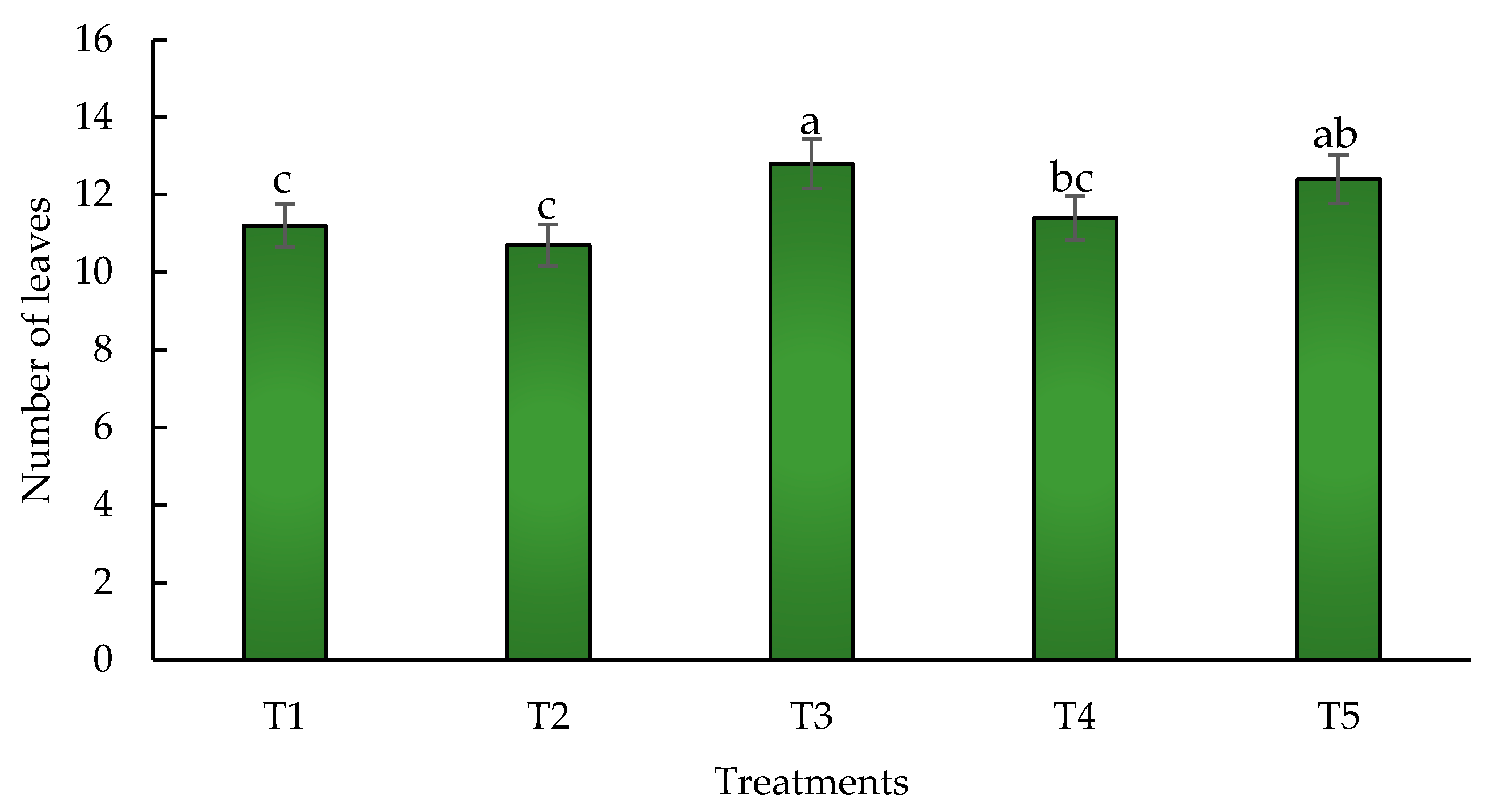
Figure 8.
Effect of biofertilizers application on plant height where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤ 0.05).
Figure 8.
Effect of biofertilizers application on plant height where: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4. Means with equal letters do not differ according to the Duncan Multiple Range Test (p ≤ 0.05).
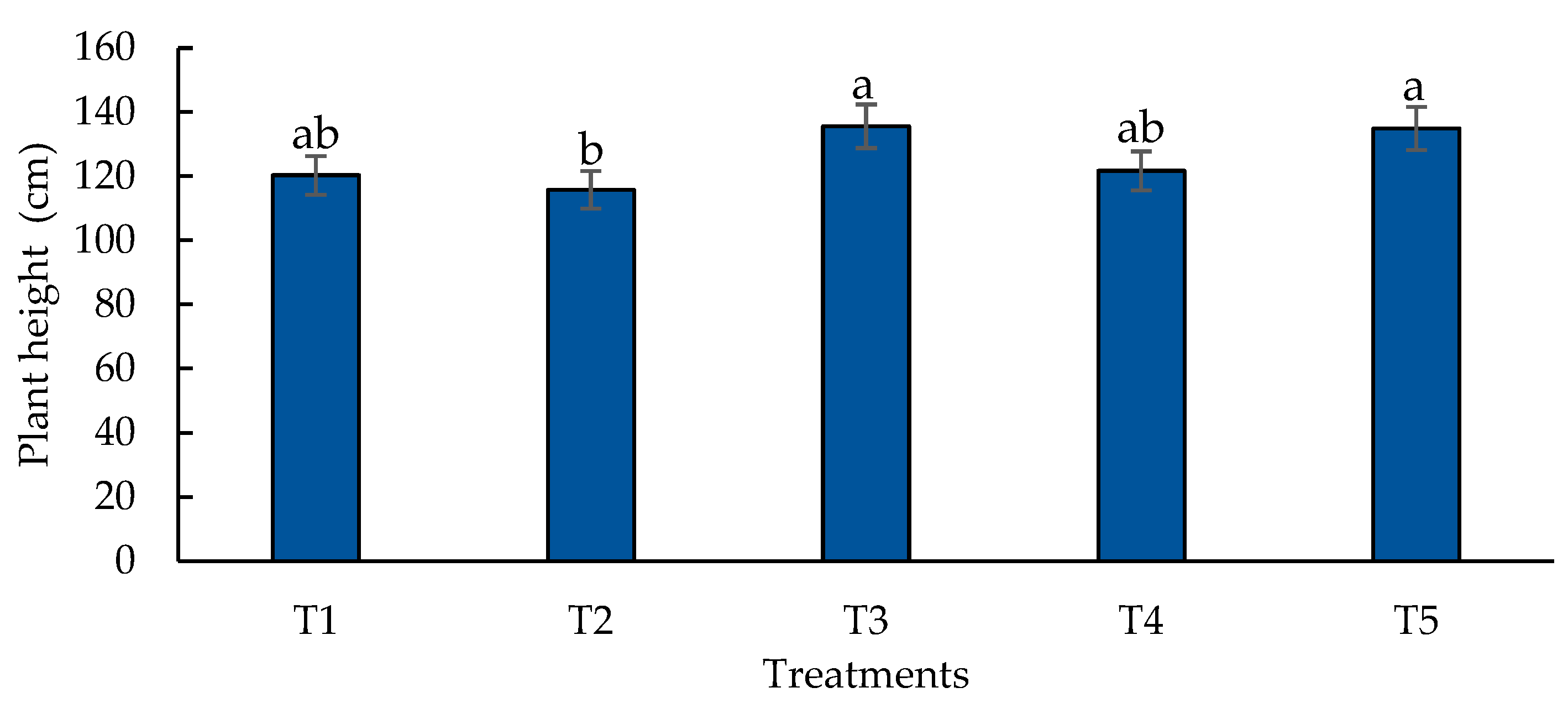
Figure 9.
Pearson’s correlation analysis represented in a heat map. The figure is made with the variables of biomass, yield, nitrate reductase activity, SPAD values, photosynthetic pigments, number of leaves, and plant height, where the effect of the biofertilizers: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4, applied to maize, can be seen.
Figure 9.
Pearson’s correlation analysis represented in a heat map. The figure is made with the variables of biomass, yield, nitrate reductase activity, SPAD values, photosynthetic pigments, number of leaves, and plant height, where the effect of the biofertilizers: T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4, applied to maize, can be seen.
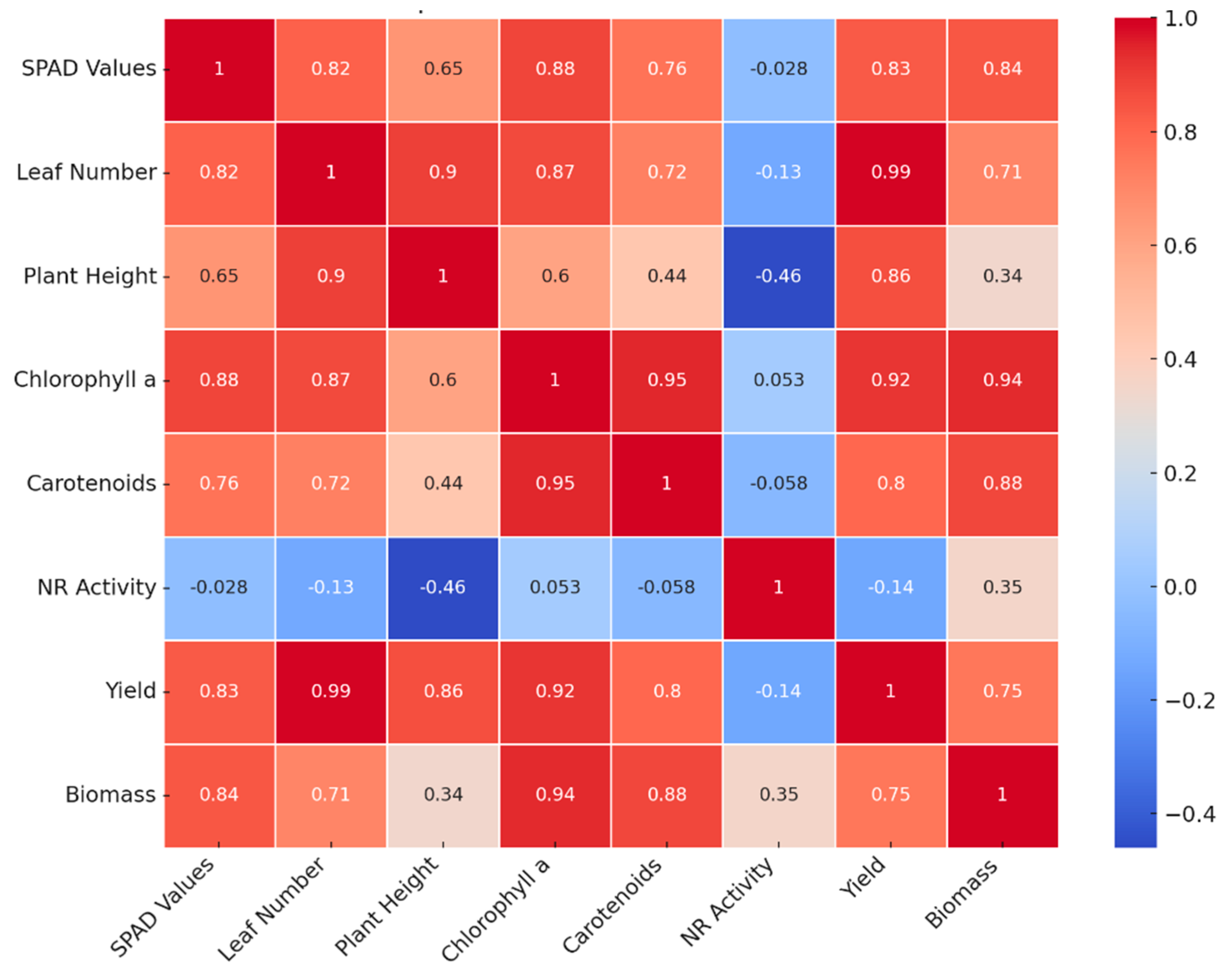
Figure 10.
Results showed in (a) Venn diagram represents the interaction of different treatments on plant growth and physiological variables.; (b) Interaction diagram. The bar chart shows how the different variables (biomass, yield, nitrate reductase activity, SPAD values, photosynthetic pigments, number of leaves, and plant height) respond to each treatment (T1 to T5). The arrows indicate whether there was an increase or decrease in these variables compared to the control (T1). T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4.
Figure 10.
Results showed in (a) Venn diagram represents the interaction of different treatments on plant growth and physiological variables.; (b) Interaction diagram. The bar chart shows how the different variables (biomass, yield, nitrate reductase activity, SPAD values, photosynthetic pigments, number of leaves, and plant height) respond to each treatment (T1 to T5). The arrows indicate whether there was an increase or decrease in these variables compared to the control (T1). T1=control; T2=algal extracts; T3=Rhizobium sp.; T4=T. asperellum; T5=T2+T3+T4.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



