
Submitted:
14 September 2024
Posted:
16 September 2024
You are already at the latest version
Abstract
Metallothioneins are multifunctional proteins implicated in various cellular processes. They have been used as biomarkers of heavy metal exposure and contamination due to their intrinsic ability to bind heavy metals and their transcriptional response to both physiological and noxious metal ions such as cadmium (Cd) and mercury (Hg). In this study, we aimed to clarify the role of iron and reactive oxygen species (ROSs) in the induction of the metallothionein system (Mtt) in the ciliate protozoan Tetrahymena thermophila. We investigated the relative mRNA abundances of the metallothionein genes Mtt1, Mtt2/4, and Mtt5, revealing for the first time their responsiveness to iron exposure. Furthermore, by using inhibitors of superoxide dismutase (SOD) and catalase (CAT), alone or in combination with iron, we highlighted the roles of superoxide ion and endogenous hydrogen peroxide, as well as the complex interplay between the metal and ROSs. These results enhance our understanding of the metallothionein system in ciliates and suggest that ROSs may be a primary evolutionary driver for the selection of these proteins in nature.
Keywords:
aquatic system
; heavy metals
; evolution
; oxidative stress
; pollution
; protist
; iron homeostasis
1. Introduction
Metallothionein (MT) is a superfamily of proteins widely distributed among both eukaryotic [1] and prokaryotic [2] organisms. They are distinguished by their exceptional affinity for heavy metal ions and metal binding capacity, attributed primarily to a unique content of reduced cysteine residues (20-35%) in the primary structure, giving rise to metal-tetrathiolate clusters [3].
Native purified MTs typically bind Zn(II) and Cu(I) ions, particularly under physiological conditions [4]. However, their binding affinities extend to other metals in vivo, including Cd(II) and Hg(II) [5]. Furthermore, monosubstituted derivative MTs, originating from the metal-free, known as MT apo form, can accommodate various metals such as Fe(II), an essential metal, along with Pb(II), Bi(III), Sn(II), Co(II), Ni(II), Tc(III), In(III), and Sb(III) [6]. These forms are practically artificial and do not occur in natural systems. Traditionally designated as reservoirs/buffers for physiological heavy metal ions and scavengers for toxic species like Cd, Cu and Hg [5], MTs have been often perceived to have an elusive primary function [7]. Knockout studies in fruit flies, in fact, revealed that MTs are non-essential for organismal development and survival [8]. Nevertheless, they confer significant advantages in coping with stressful situations, including exposure to heavy metals, low Zn conditions, infection, and various oxidative stress-related processes [9]. Moreover, beyond their metal ion affinity and binding capacity, MTs exhibit a substantial redox capacity, playing a pivotal role in diverse stress response processes, such as inflammation [10], gamma rays [11] or UVB irradiation [12], and challenges from xenobiotics triggering the formation of Reactive Oxygen Species (ROSs) [13]. Additionally, MT genes display transcriptional responsiveness to other non-physiological and physiological non-metal stimuli including glucocorticoids, and/or several polypeptide hormones [14]. Despite these broad functional capacities, no role in iron homeostasis has been attributed to MTs so far.
Tetrahymena spp. are ciliated protozoa that express multiple metallothionein genes, which fall into two discrete subfamilies, 7a and 7b, differing in their structural and evolutionary traits [15]. Tetrahymena MT genes coordinate expression with distinctive functions and respond diversely to environmental stressors. In T. thermophila, MTT1 (protein encoded by the Mtt1 gene) binds and responds preferentially to Cd(II), aiding in noxious metal homeostasis and detoxification. Nearly identical, Mtt2 and Mtt4 genes encode copper-MTs with the highest binding preference for Cu(I) [16]. Among its five metallothionein genes, the cadmium metallothionein Mtt5 gene is deemed essential [17]. The transcriptional regulation of MT genes in Tetrahymena has been a subject of investigation owing to its significance in understanding cellular responses to metal stress and environmental adaptation. Two seminal studies demonstrated the robust inducibility-repressibility of the Cd-inducible Mtt1 [18] and Cu-inducible Mtt2 [19] genes in T. thermophila. These studies revealed that Mtt1 or Mtt2 mRNA expression is tightly regulated by cadmium or copper ion concentration respectively, with rapid induction upon metal exposure and subsequent downregulation upon withdrawal. Moreover, both promoters were shown to be highly efficient in driving conditional expression of heterologous genes, under control of fully sub-toxic metal levels, offering a valuable tool for genetic manipulation and functional studies in T. thermophila, but also demonstrating definitively that Mtt1 and Mtt2 are primarily controlled by inducible promoters. Mt genes are transcriptionally regulated through multiple cis-acting elements known as Metal Responsive Elements (MRE) [14]. Tetrahymena MT promoters don’t appear to contain genuine MREs. Insights into the transcriptional regulation of Tetrahymena MT genes emerged from the work of Formigari et al., [20] showing the involvement of a proximal composite GATA cis-acting element in Cd inducibility of Mtt5 and those of Diaz et al., [15] and de Francisco et al., [21] who identified multiple Metallothionein Conserved Motif 1 (MTCM1) resembling AP-1 antioxidant responsive elements, suggesting a potential role for AP-1 (bZIP) transcription factors in regulation of Mtt gene expression. AP-1 binding sites have been identified in several MT vertebrate promoters, where they are part of a complex cis-acting element setting [22,23,24].
The aim of this study was to establish a central involvement of metallothioneins in the oxidative stress response within a relatively simpler and well-characterized metallothionein expressing system i.e., the ciliated protozoa T. thermophila. To achieve this objective, we employed quantitative real-time reverse transcription quantitative PCR (QPCR) in conjunction with multiplexed Taqman hydrolysis probes, allowing for precise assessment of the relative abundance levels of metallothionein mRNAs, specifically Mtt1, Mtt2/4, and Mtt5. In selecting the metallothionein genes Mtt1, Mtt2/4, and Mtt5 for analysis, we aimed to reduce complexity while ensuring a comprehensive understanding of MT gene regulation in T. thermophila. According to Diaz et al., [15] the coding sequences and amino acid compositions of Mtt2 and Mtt4 are nearly identical, indicating very close molecular evolution of the 7b Mtt sub-family. Due to the high nucleotide sequence similarity between them, we analyzed Mtt2 and Mtt4 together as a set. On the other hand, Mtt3 is similar to Mtt1, with both genes appearing to have evolved from the same ancestral gene. Although Mtt5 is distinct, it shares an evolutionary origin with Mtt1 and Mtt3 [15], providing a broader perspective on MT gene regulation for the 7a Mtt sub-family protein. Our investigation into the gene responsiveness of metallothionein involved the influence of superoxide and hydroxyl radicals, achieved through the use of inhibitors for superoxide dismutase (SOD) and catalase (CAT), as well as the introduction of iron to initiate the Fenton [25] and/or Haber-Weiss [26] reactions triggering superoxide and hydroxyl radicals’ formation. Unexpectedly, our findings highlighted a pivotal role of iron per se in modulating gene expression within T. thermophila metallothionein.
2. Materials and Methods
All chemicals were reagent grade and were purchased from Merck (Germany). All dehydrated culture media powders were purchase from DIFCO™ (Franklin Lakes, NJ, USA). Tetrahymena thermophila strain CU427, was kindly provided by the research group of Prof. Antonietta Laterza, University of Camerino, Italy.
2.1. Tetrahymena Thermophila Culture and Treatments
Cultures of T. thermophila (10 ml) were grown in axenic condition in PPY culture medium (10 g/L Difco BactoTM yeast extract, 1.5 g/L Difco BactoTM proteose peptone and 30 mg/L ethylenediaminetetraacetic acid ferric sodium salt) until the late-log phase at a density of 106 cells/ml, at 30°C, 150 rpm, using aerated 50 mL polypropylene plastic FalconTM tubes. Log-exponential cultures of T. thermophila were treated with the following chemicals: iron(III) chloride, sodium azide, sodium diethyldithiocarbamate, hydrogen peroxide. For the selection of testing concentrations, we used No Observed Effect Concentrations (NOECs) for swimming as a proxy for survival. Briefly, late log-exponential cultures of T. thermophila were settled in 96 multi-well plates in full medium with the addition of increasing concentrations of each individual chemical for 24-72 h. Cultures were then gathered and assessed for swimming behavior under light microscopy at 25-50X magnification. The concentration giving rise to an unequivocal change in overall T. thermophila motility was referred to as the Lowest Observed Effect Concentration (LOEC). The preceding concentration was designated as the NOEC and used for subsequent experiments. The designed concentrations used in the experiments are given in Table 2.
Cadmium in form of chloride was used at 20 μM as a positive control for Mtt1 and Mtt5 induction.
All reagents were obtained from Merck (Germany) and were of ultra-pure grade, with a purity equal or greater than 99%.
2.2. RNA Extraction
Total RNA was extracted with the GeneJet RNA Purification Kit (Thermo Fisher Scientific) according to the manufacturer instructions. This reagent system uses a silica-based membrane in a centrifuge column. Briefly, cell pellets (1-5 * 105 cells) were lysed and homogenized in lysis buffer containing guanidine thiocyanate and detergents. The lysate was then mixed with ethanol and loaded onto the silica gel purification column. The lysate was centrifuged at 10000 g at RT, the flowthrough was discarded while the RNA is retained by the membrane. DNA, proteins and salts were removed from the membrane in two different washing steps using a buffered hydroalcoholic (75% ethanol) proprietary solution. Total RNA was eventually eluted under low ionic strength conditions with nuclease-free water in a variable volume of 30-50 μL.
2.3. cDNA Synthesis
Before reverse-transcription, genomic DNA (gDNA) was removed using DNAse-I treatment [27]. Total RNA (up to 1 μg in μL nuclease free water) was mixed with 1 U DNase-I (Thermo Fisher Scientific) in reaction buffer containing MgCl2, 40 U RiboLock RNase Inhibitor reagent (Thermo Fisher Scientific) and incubated at 37°C for 30 min in a final volume 10 μl according to the manufacturer instructions. After DNase-I inactivation at 70°C for 10 min, 2 μL of the reaction mixture was used for first strand cDNA synthesis using a point mutated RNase-H deficient M-MuLV reverse transcriptase (RevertAid H-minus, Thermo Fisher Scientifics) in the presence of 125 ng random hexamers (Invitrogen), dithiothreitol, MgCl2 and enzyme buffer in a volume of 10 μL according to the manufacturer instructions.
2.4. Quantification of Relative mRNA Abundances of Mtt Genes
Real time quantitative PCR (QPCR) in combination with a TaqmanTM multiplex protocol was used evaluate relative mRNA abundances of the Mtt genes. This technique involves the use of hydrolysis probes with dual fluorescent labeling [28] (Table 2). The probe set designed for Mtt2 is 100% identical to Mtt4, therefore it recognizes the joint expression of both genes. A gene probe encoding the T. thermophila 17S ribosomal gene was used as reference for normalizing gene expression data between samples. All primers and probes were synthetized and purified by Eurofins Genomics (Ebersberg, Germany).

Table 1.
Primers and probes used for QPCR. NCBI GeneID is given in the name of the oligonucleotide/probe. All sequences are given in the 5’-3’ direction. Abbreviations. FAM, 6-carboxyfluorescein; Hex, hexachlorofluorescein; RED, Texas RedTM; BHQ1, Black Hole Quencer 1; BHQ2, Black Hole Quencer 2. F, Forward primer; R, Reverse primer.
Table 1.
Primers and probes used for QPCR. NCBI GeneID is given in the name of the oligonucleotide/probe. All sequences are given in the 5’-3’ direction. Abbreviations. FAM, 6-carboxyfluorescein; Hex, hexachlorofluorescein; RED, Texas RedTM; BHQ1, Black Hole Quencer 1; BHQ2, Black Hole Quencer 2. F, Forward primer; R, Reverse primer.
| NCBI ID | Sequence (5’-3’) | Oligo Type |
|---|---|---|
| AF537326-Mtt1 | FAM-TGC TGC ACA GAC CCT AAC AGC GGA-BHQ1 | Dual Labeled Probes |
| AY204351-Mtt2 | HEX-TGC TTG CAA TTG CAA ACC TTG CGA-BHQ1 | Dual Labeled Probes |
| AY884209-Mtt5 | RED-CTC ACT AGG GCA GCA GCA CCA CTT-BHQ2 | Dual Labeled Probes |
| M10932-17s | CY5-CCG CAG GTT CCA CTT CTG GTG TGC-BHQ2 | Dual Labeled Probes |
| AF537326-Mtt1F | TGC TGT GGT GTA AAT GCT AAG CC | Forward PCR Primer |
| AF537326-Mtt1R | TCC TGT ACC AGT GCA ACA TCC CT | Reverse PCR Primer |
| AY204351-Mtt2/4F | ACC TCT CTG CAA ATG TGG AAC TAC | Forward PCR Primer |
| AY204351-Mtt2/4R | CCA CAG CTT TCA GTA ACA CCA CAT | Reverse PCR Primer |
| AY884209-Mtt5F | TCT GGT GAA AGC ACT AAA ATT TGT | Forward PCR Primer |
| AY884209-Mtt5R | ATC AGA ATT GCA GCA ATT TTG GG | Reverse PCR Primer |
| M10932-17sF | CGA TCA GAT ACC GTC GTA GTC TTA | Forward PCR Primer |
| M10932-17sR | GAG TTT CCC CGT GTT GAG TCA | Reverse PCR Primer |
The QPCR reaction (7 μl final volume) was prepared in triplicate using 2 μl of diluted cDNA (1:10 dilution from the reverse-transcription reaction) and the iQ Multiplex Power master mix (Bio-Rad Laboratories) accounting for 3 min initial denaturation (95°C) and 45 cycles of two-step amplification: 15 sec denaturation (95°C), 1 min annealing plus extension at 59°C. Within each QPCR run, a standard curve was obtained for each probe set to estimate PCR efficiency (E) and linearity (R2) over at least 4 magnitude orders. The average E values for each target was comprised between 1.98 and 2.01, with a R2 higher than 0.99, therefore, for subsequent analysis of relative mRNA abundances an E value of 2 was adopted for all targets.
Statistics
Real time quantitative PCR data were analyzed in relative mode using the ΔΔCq method of Busting et al. [29] where Cq is the threshold cycle of QPCR reaction, using not exposed T. thermophila culture as relative control. GraphPad Prism® 9 (GraphPad Software, San Diego, CA, USA) was used to evaluate statistical significance of the Mtt mRNA relative abundances among groups by means of conventional or Welch’s ANOVA statistics in case of heteroscedastic data. For post-hoc multiple comparison the Tukey’s or Dunnet T3 test was performed accordingly (for details see captions to Figures). Statistical analysis was carried out on ΔCq values, i.e., Cq of GOI (Mtt1, Mtt2/4 and Mtt5) minus Cq of the housekeeping reference gene (17s rRNA).
3. Results
Figure 1 illustrates the effects of exogenous exposure to hydrogen peroxide and hydrogen peroxide with iron at 24 and 72 hours. We intentionally selected a concentration of hydrogen peroxide (Table 2) that was ineffective on its own to specifically evaluate its combined effect with iron on Mtt transcriptional activation under the current experimental conditions. When combined with iron, Mtt1 showed significant activation while a mild trend of activation was observed for Mtt5 and Mtt2 at 24 hours. No significant effects were seen at 72 hours.
The relative expression levels of Mtt1, Mtt2/4, and Mtt5 were, then, measured after 24 hours of exposure to various stressors (Table 2), including Cd (II) as a positive control, Fe (III), sodium azide, sodium diethyldithiocarbamate, and combinations of each inhibitor with iron (Figure 2a–c). Cadmium exposure resulted in the highest expression of the Mtt1 gene. Iron also induced Mtt1 expression (Figure 2a), though to a lesser extent. The inhibitors sodium azide and sodium diethyldithiocarbamate caused a significant effect on Mtt1 expression (Figure 2a). Interestingly, sodium azide plus Fe resulted in a trend of higher expression than sodium azide alone which, however, seemed to be driven by Fe, while diethyldithiocarbamate plus Fe showed a statistically significant lower expression level than diethyldithiocarbamate itself.
For Mtt2/4, (Figure 2b) iron exposure alone yielded substantial gene expression, relatively higher than that induced by cadmium, but generally lower than Mtt1, with differences from control values being only marginally statistically significant (p < 0.1). Sodium azide and diethyldithiocarbamate also elicited a marginal trend of Mtt2/4 induction similar to iron. Their combination with iron led to varied effects: sodium azide plus iron showed no additional effect, whereas diethyldithiocarbamate plus iron caused a sharp increase compared to diethyldithiocarbamate or iron alone.
Similar to Mtt1, cadmium exposure resulted in the highest expression of Mtt5 (Figure 2c). In general, the pattern of Mtt5 was very similar to that of Mtt1, with the same effect of iron on the co-exposure with each inhibitor but this time the difference between sodium azide and sodium azide plus Fe was statistically significant due to a lower uncertainty.
Table 2.
Substances tested for the effects on Mtt gene expression.
| Treatment | NOEC |
|---|---|
| Sodium azide (SAN) | 200 μM |
| Sodium diethyldithiocarbamate (DTN) | 100 μM |
| Cadmium chloride (Cd) | 20 μM |
| Ferric chloride (Fe) | 200 μM |
| Hydrogen peroxide (H2O2) | 100 μM |
After 72 h of exposure, the expression patterns of Mtt1 (Figure 2d), Mtt2/4 (Figure 2e), and Mtt5 (Figure 2f) genes were assessed under the same conditions as the 24h exposure. Relative mRNA abundances of Mtt1 fairly declined to new steady state levels, including with cadmium treatment, but remained substantial with iron whose level was significantly higher than any other analyzed condition. The treatments with sodium azide (and marginally diethyldithiocarbamate) rendered negative expression levels respect to control which may suggest repression of Mtt1 transcription. When combined with iron, the expression levels of Mtt1 with sodium azide or diethyldithiocarbamate treatments were significantly higher than the those of the individual inhibitors alone but lower than that of iron itself.
At 72 hours, iron exposure continued to induce substantial Mtt2/4 expression (Figure 2e), similar to the effects observed with sodium azide and diethyldithiocarbamate treatments. Combining these treatments with iron did not produce any notable additional effects, but a significant reduction for diethyldithiocarbamate plus iron below the 24h level and also below the 72h iron level. These findings suggest an interaction between iron and diethyldithiocarbamate and in same time an adaptive response of Mtt2 in this scenario.
The pattern for Mtt5 after 72 hours mirrored that of Mtt1, with significant relative mRNA abundances observed only with cadmium and iron exposures. Moreover, the effects of the two inhibitors were fully compensated, which may suggest faster adaptive kinetics for Mtt5 compared to Mtt1.
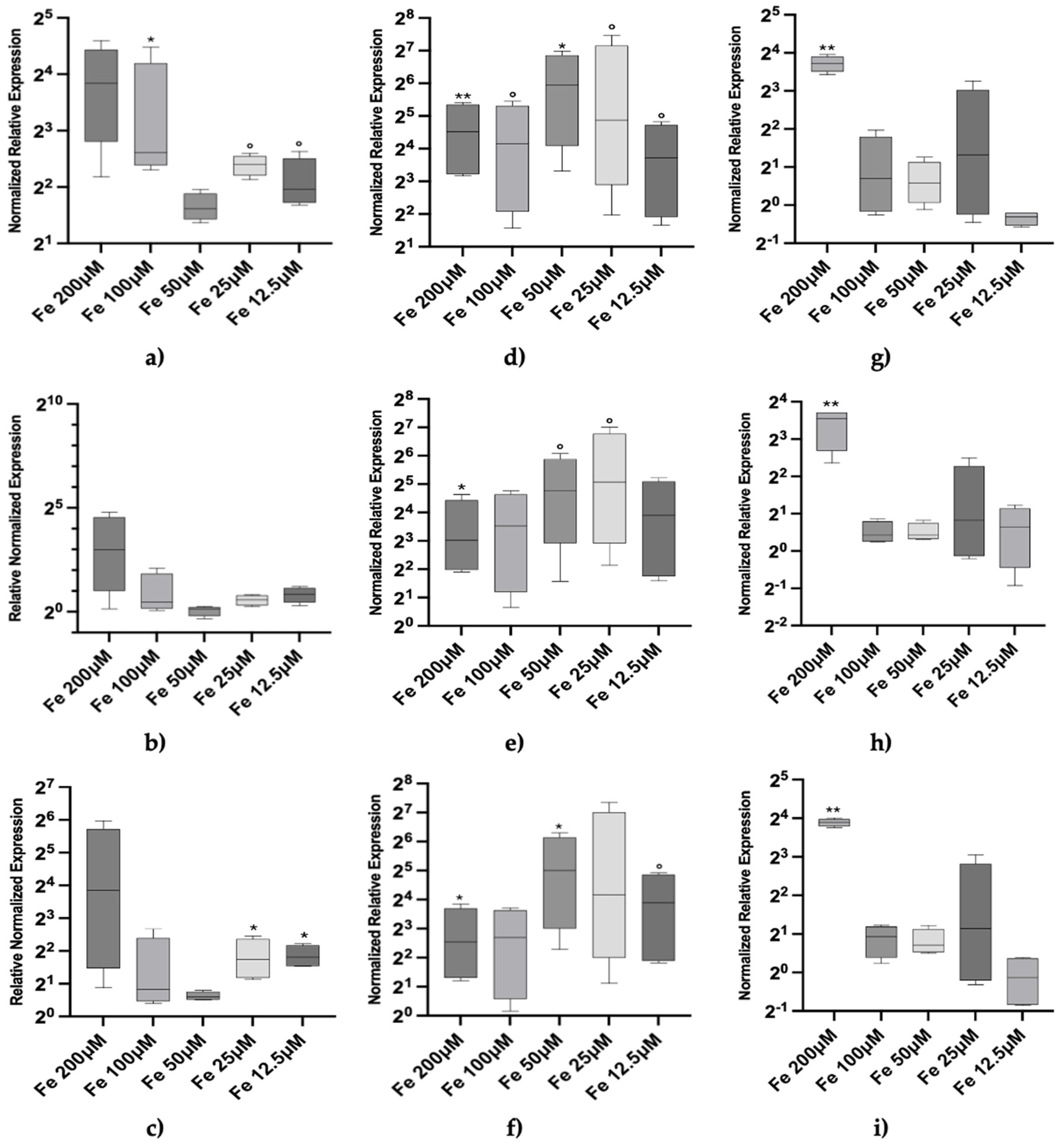
We, then, directed our focus towards solely iron as a potential novel Mtt inducer, examining dose-dependent effects ranging from 12.5 μM to 200 μM and kinetic responses spanning from 2h to 72h (Figure 3).
At 2 hours there was a positive transcriptional response of Mtt1 and Mtt5 at both high and low concentrations, although high uncertainty levels affected some conditions (Figure 3a–c). The gene Mtt2/4 showed only a trend of activation at 200 µM Fe. We analyzed the effects of same concentrations at 24h (Figure 3d–f) and 72 hours (Figure 3g–i), revealing that: no clear dose response pattern is seen for any Mtt genes; the response is adaptive peaking at 24h; the highest iron concentration (200 µM) is not compensated after 72h. The lack of a clear dose-response effect and the adaptive response observed over time may suggest that intracellular iron concentration is regulated in T. thermophila with a possible role for Mtt genes.
4. Discussion
Our initial hypothesis centered around confirming Tetrahymena Mtt’s involvement in ROS scavenging activity. Consequently, we utilized hydrogen peroxide and a combination of hydrogen peroxide with iron as exogenous sources of ROSs. Furthermore, we used inhibitors targeting key antioxidant enzymes, specifically sodium azide [30] and sodium diethyldithiocarbamate [31], to inhibit catalase (CAT) and superoxide dismutase (SOD) activities, respectively, thereby promoting the intracellular formation of ROS. Previous studies have demonstrated the efficacy of this strategy to raise levels of hydrogen peroxide [32] or superoxide ion [33] using micromolar levels of the same inhibitors employed in this work. To intensify this effect, we included iron as an additional treatment with the aim of stimulating the Fenton [25] and/or Haber-Weiss [26] reactions known to generate hydroxyl and/or hydroperoxyl radicals from hydrogen peroxide (H₂O₂) or superoxide (O₂⁻). Thus, we gathered and assessed the transcriptional modulation of the Mtt genes using QPCR quantifying relative abundances of T. thermophila metallothionein mRNAs (Mtt1, Mtt2/4, Mtt5).
Sodium azide was used due to its ability to inhibit catalase [30]. This enzyme is responsible for the conversion of hydrogen peroxide into molecular oxygen and water [34]. This is a fundamental defense mechanism that all organisms use to detoxify endogenous H2O2, and if this molecule is not taken care of, it reacts and degrade proteins, nucleic acids, and lipids. Sodium diethyldithiocarbamate was used because it is a known inhibitor of Super Oxide Dismutase (SOD) [31]. This enzyme is responsible for the dismutation reaction of the superoxide anion (O2-) into two different compounds (H2O2 and O2) [35]. SOD is a key enzyme in the cell’s defense against superoxide anion, a ROS species, which causes the inactivation of iron/sulphur clusters present in many enzymes and proteins that play a key role in metabolic processes.
In addition to exacerbating the intracellular oxidation we paired the two inhibitors with iron, therefore, three new treatments were obtained i.e., sodium azide plus iron, sodium diethyldithiocarbamate plus iron and H2O2 plus iron. Iron, in fact, must be tightly regulated intracellularly as it is the starting point of several reactions that create oxidative stress (sensu the Fenton [25] or Haber-Weiss reactions [26]). Previous data based on comparative QPCR analyses and protein-DNA interaction experiments supported the involvement of AP-1 transcription factors in mediating MT gene expression responses to metal stress in T. thermophila [21]. The latter finding is quite relevant in the context of gene expression results of our study since AP-1 transcription factors are responsible for the transcriptional control of oxidative stress related genes through the Antioxidant Response genetic Elements (AREs). In mammalian cells, these factors can bind to specific cis-acting elements rendering numerous cellular processes, including cell proliferation, apoptosis, development, but also triggering response to stress induced by exposure to UV or cytotoxic agents (e.g. cycloheximide, 4-nitroquinoline), iron-chelating complexes, oxidizing agents and several heavy metals including iron [36,37]. AP-1 transcription factors seem to be universal mediators as they are also relevant for abiotic stress response in yeast [38] and plants [39], moreover they have been already mapped in MT promoters of vertebrates [22,23,24]. In addition to transcription factors, epigenetic mechanisms such as microRNAs (miRNAs) have been implicated in post-transcriptional regulation of genes involved in metal stress responses. Amaro et al. [40] isolated and characterized miRNAs involved in the post-transcriptional regulation of transcripts linked to the response to cadmium stress in T. thermophila. These studies highlight the regulatory complexity underlying the cellular response to metals in modulating gene expression in Tetrahymena, even in a relatively simpler model organism. They also emphasize the inevitable interplay between heavy metal and oxidative stress responses, as has been previously described in vertebrates.
The activation of T. thermophila metallothionein genes by oxidants such as hydrogen peroxide is well-documented in the literature [41,42]. Our study, however, provides new insights into the response of metallothioneins to abiotic stimuli. Based on the hypothesis that the repression of catalase and SOD enzyme activities could increase the levels of H2O2 and superoxide radicals, we conducted a pioneering investigation into the influence of iron on metallothionein gene expression, focusing specifically on the T. thermophila Mtt system.
Our work represents a significant advancement in the understanding of the molecular regulation of metallothionein genes. By targeting key antioxidant enzymes with specific inhibitors and co-exposing protozoa to iron, the study aimed to simulate endogenous oxidative stress conditions conducive to MT gene induction. This innovative approach allowed us to explore the direct impact of reactive oxygen species (ROS) on MT gene expression. A key question is whether the two inhibitors could activate Mtt gene transcription, and this study provides clear evidence supporting that. This activation may result from either a direct effect of increased endogenous ROS levels or from indirect effects of ROS on intracellular metal levels (and their oxidation state). Another significant finding is that iron itself effectively modulated Mtt gene expression. The combination of iron with sodium azide or hydrogen peroxide never exceeded the effects of iron alone, suggesting non-significant interactions, at least by 24 hours (Figure 2a–c). Indeed, at 72 hours (Figure 2d–f), the effects of iron combined with sodium azide were significantly lower than those of iron alone for Mtt1 and Mtt5, but not for Mtt2/4. This finding suggests differential regulation between the 7a and 7b groups of Mtt genes, indicating that Mtt2/4 might have a unique regulatory mechanism compared to Mtt1 and Mtt5. Comprehensive mechanistic explanations for this behavior cannot be determined from gene expression data alone, necessitating further studies. However, for the combined effects of exogenous hydrogen peroxide and iron at 72 h a possible explanation can be drawn from an analogy with S. cerevisiae, where iron uptake involves a membrane-bound ferrireductase to reduce Fe(III) to Fe(II) and a ferrous transporter (Ftr1p) to import Fe(II) into the cell [43,44]. Exogenous hydrogen peroxide may reduce Fe(III) to Fe(II), interfering with iron’s availability to enter the cell and modulate Mtt gene expression. Simultaneously, the interaction between endogenously formed hydrogen peroxide, due to catalase inhibition by sodium azide, might alter the oxidation state of iron over time, affecting its ability to regulate Mtt1 and Mtt5 expression. This suggests that the observed effects at 72 hours are the result of a time-dependent process that requires further investigation to fully understand the underlying mechanisms.
A consistent pattern of iron inducibility was observed across all Mtt genes (Figure 3), characterized by an adaptive response that lowered its intensity over time, except at high, non-permissive levels (200 µM), where gene expression remained substantial throughout. Unlike Cd and Cu exposures, iron did not show dose dependency in Mtt gene transcriptional activity, likely due to the tight regulation of free iron intracellular concentrations [45].
Our results open up the intriguing hypothesis that the Mtt system participates in iron homeostasis. Despite a native Fe-bound metallothionein (MT) never being isolated so far, it is interesting to note that hepcidin binds one Fe(III) ion through a tetrahedral thiolate cluster [46], similar to how metallothionein binds multiple Cd/Cu atoms. Hepcidin, a liver hormone, is the overarching governor of iron homeostasis in the plasma of mammals [47]. Very little is known about iron homeostasis in ciliates; therefore, further research is essential to understand the potential roles and mechanisms involved.
Examining the effects of diethyldithiocarbamate, a superoxide dismutase (SOD) inhibitor, combined with iron (Figure 2), we observed a discordant pattern between the Mtt1/Mtt5 pair and Mtt2/4. Mtt1 and Mtt5 are phylogenetically related, with consistently aligned transcriptional patterns in our study. In contrast, diethyldithiocarbamate plus iron exhibited an antagonistic effect on Mtt1/Mtt5 expression while having an additive, if not cooperative, effect on Mtt2/4. This suggests that in co-exposures, iron alone can account for the transcriptional activation of Mtt1/Mtt5 but not Mtt2/4. The response observed indicates that either iron, or superoxide ions whose levels are elevated by SOD inhibition, have distinct modes of action on the Mtt2/4 promoter. Alternatively, this interaction may generate new effector(s), possibly through the Haber-Weiss reaction, which are more effective in activating Mtt2/4, i.e. the hydroperoxyl (HO₂•) and hydroxyl radical (OH•) are possible candidates.
Our findings highlight the complex interplay between iron, ROS, and the regulatory mechanisms governing MT gene expression. The evolution of metallothionein and heavy metal-inducible gene transcription systems is a fascinating topic that intertwines the roles of oxidative stress and heavy metal detoxification. While it is clear that this system is primarily known for its role in metal ion homeostasis and detoxification, we cannot exclude the possibility that oxidative stress response has driven their evolution. This study underscores the importance of considering both direct and indirect effects of oxidative stress and metal ions in understanding the molecular regulation of MT genes. Considering the previously described effects of cadmium and copper, it is evident that ROS alone cannot be regarded as the primary factor for MT gene activation. Metals can promptly induce and repress MT gene transcription, suggesting the involvement of a metal-sensitive transcription factor analogue to the MTF-1 [48] found in vertebrate systems. Rather than a zinc finger like MTF-1, we speculate the existence of a redox-sensitive transcription factor in T. thermophila, akin to the E. coli SoxR protein [49], or the S. cerevisiae Yap5 [50] which are indeed Fe-S (iron-sulfur) protein, known for their essential role in response to oxidative stress. The latter, in particular, is known for regulating iron storage and is a basic leucin zipper (bZIP) protein, the same family of transcription factors putatively identified by de Francisco [21] in Mtt promoters. This hypothetical factor may function as a redox sensor that recognizes superoxide and/or other ROS, while also being influenced by metal ions with high affinity for sulfur. An important question arises: what has driven the evolution of metallothionein and heavy metal-inducible gene transcription systems? Can we truly exclude the possibility that oxidative stress response mechanisms could have driven the evolution of inducible systems that incidentally respond to heavy metals? The dual functionality of MT in both metal detoxification and oxidative stress response suggests an evolutionary advantage in environments where organisms are exposed to both metals and oxidative stress. However, oxidative stress is a common challenge faced by aerobic organisms, leading to the production of ROS, which are highly reactive and can cause cellular damage. The evolution of MT may have been driven by the need to mitigate this damage. Additionally, MT, by binding metal ions, can reduce the catalytic activity of metals like copper or iron, which participate in Fenton and Haber-Weiss reactions, generating harmful hydroxyl radicals. The ability of MT to scavenge ROS directly provides an additional layer of protection and binding heavy metals may not represent the primary function of MT but rather a structural constraint to keep cysteine residues in a reduced state, acquiring redox potential and scavenging activity against oxidative radicals as already shown [11]. The incidental responsiveness to heavy metals could be an advantageous byproduct of this primary function. In mammals it has been shown that metals like cadmium and copper as well as ROS or disulfides like oxidized glutathione can displace zinc in MT, leading to changes in MT conformation and, consequently, increase gene transcription through the activation of the proper transcription factors [51].
In summary, while oxidative stress plays a significant role in Mtt gene activation, the primary regulatory mechanism likely involves a complex interplay between ROS and metal-responsive elements, mediated by specialized transcription factors that respond to both oxidative and metal-induced signals. The evolution of MT and its transcription system may have been driven by the need to protect against oxidative damage, with the incidental benefit of metal detoxification. The Mtt system’s response to iron suggests potential involvement in iron metabolism. Mtt2/4 promptly responded to the interactive effects of superoxide and iron and exhibited prolonged expression in response to elevated endogenous hydrogen peroxide and superoxide levels. Given its earlier molecular evolution [15] and responsiveness to Cu [16], we speculate that Mtt2/4 might better represent the antioxidant function of the ancestral MT gene, while the 7a group may have evolved later, acquiring specialized functions related to heavy metal homeostasis, including iron. Indeed, Mtt5, which is an essential gene [17], appears to be the best candidate for specifically controlling essential metals such as iron. Further research is needed to validate this hypothesis in view also of the limitation of this study (missing data on Mtt3, and no discrimination between Mtt2 and Mtt4 expression), identify the regulatory elements and factors involved, and better understand the evolutionary pressures that shaped these multifunctional proteins. The ciliate T. thermophila represents an appropriate experimental system for this purpose.
5. Conclusions
In conclusion, we have characterized the transcriptional response of T. thermophila metallothionein isoforms (Mtt1, Mtt2/4, and Mtt5) to iron and reactive oxygen species (ROS) using catalase and superoxide dismutase inhibitors. This study highlights the differential responses of the 7a and 7b Mtt groups revealing the complex interplay between these factors and their regulatory mechanisms. Our findings underscore the importance of further research to understand the specific roles of these isoforms in metal homeostasis -including iron- and oxidative stress response.
Author Contributions
Davide Gualandris: Methodology, Investigation, Formal Analysis, Data Curation, Statistical Analysis, Software, Writing—Original Draft, Review, Editing, Visualization. Davide Rotondo: Investigation. Candida Lorusso: Investigation. Antonio Calisi: Writing, Review, Editing. Francesco Dondero: Conceptualization, Methodology, Validation, Formal Analysis, Data Curation, Statistical Analysis, Software, Writing, Review, Editing, Visualization, Project administration, Funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the University of Eastern Piedmont with local research program FAR2015.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Capdevila, M., Atrian S. Metallothionein protein evolution: a miniassay. J Biol Inorg Chem. 2011 Oct;16(7):977-89. [CrossRef]
- Blindauer, C.A. Bacterial metallothioneins: past, present, and questions for the future. J Biol Inorg Chem. 2011 Oct;16(7):1011-24. [CrossRef]
- Drozd A., Wojewska D., Peris-Díaz M.D., Jakimowicz P., Krężel A., Crosstalk of the structural and zinc buffering properties of mammalian metallothionein-2, Metallomics, Volume 10, Issue 4, April 2018, Pages 595–613. [CrossRef]
- Vallee, B.L., Falchuk K.H., The biochemical basis of zinc physiology. Physiol Rev. 1993 Jan;73(1):79-118. [CrossRef]
- Kägi, J.H., Kojima Y. Chemistry and biochemistry of metallothionein. Experientia Suppl. 1987; 52:25-61. [CrossRef]
- Hunziker, P.E., and Kagi J.H.R., Metaltothionein. In: Metalloproteins: part 2. Metal Proteins with Non-redox roles. Harrison, P. (Edtor), pp. 149-181, Verlag Chemie, Basel, Switzerland, 1985.
- Palmiter, R.D. The elusive function of metallothioneins. Proc Natl Acad Sci U S A. 1998 Jul 21;95(15):8428-30. [CrossRef]
- Egli, D., Domènech J., Selvaraj A., Balamurugan K., Hua H., Capdevila M., Georgiev O., Schaffner W., Atrian S. The four members of the Drosophila metallothionein family exhibit distinct yet overlapping roles in heavy metal homeostasis and detoxification. Genes Cells. 2006 Jun;11(6):647-58. [CrossRef]
- Coyle, P., Philcox J.C., Carey L.C., Rofe A.M. Metallothionein: the multipurpose protein. Cell Mol Life Sci. 2002 Apr;59(4):627-47. [CrossRef]
- De, S.K., McMaster M.T., Andrews G.K. Endotoxin induction of murine metallothionein gene expression. J Biol Chem. 1990 Sep 5;265(25):15267-74. [PubMed]
- Thornalley, P.J., Vasák M. Possible role for metallothionein in protection against radiation-induced oxidative stress. Kinetics and mechanism of its reaction with superoxide and hydroxyl radicals. Biochim Biophys Acta. 1985 Jan 21;827(1):36-44. [CrossRef]
- Hanada, K., Baba T., Hashimoto I., Fukui R., Watanabe S. Possible role of cutaneous metallothionein in protection against photo-oxidative stress--epidermal localization and scavenging activity for superoxide and hydroxyl radicals. Photodermatol Photoimmunol Photomed. 1992-1993 Oct;9(5):209-13. [PubMed]
- Bauman, J.W., Madhu C., McKim J.M. Jr, Liu Y., Klaassen C.D. Induction of hepatic metallothionein by paraquat. Toxicol Appl Pharmacol. 1992 Dec;117(2):233-41. [CrossRef]
- Palmiter, R.D. Molecular biology of metallothionein gene expression. Experientia Suppl. 1987;52:63-80. [CrossRef]
- Díaz, S., Amaro F., Rico D., Campos V., Benítez L., Martín-González A., Hamilton E.P., Orias E., Gutiérrez J.C. Tetrahymena metallothioneins fall into two discrete subfamilies. PLoS One. 2007 Mar 14;2(3):e291. [CrossRef] [PubMed]
- Espart, A., Marín M., Gil-Moreno S., Palacios Ò., Amaro F., Martín-González A., Gutiérrez J.C., Capdevila M., Atrian S. Hints for metal-preference protein sequence determinants: different metal binding features of the five tetrahymena thermophila metallothioneins. Int J Biol Sci. 2015 Mar 18;11(4):456-71. [CrossRef] [PubMed]
- de Francisco P, Martín-González A, Turkewitz AP, Gutiérrez JC. Extreme metal adapted, knockout and knockdown strains reveal a coordinated gene expression among different Tetrahymena thermophila metallothionein isoforms. PLoS One. 2017 Dec 5;12(12):e0189076. [CrossRef]
- Shang, Y., Song X., Bowen J., Corstanje R., Gao Y., Gaertig J., Gorovsky M.A. A robust inducible-repressible promoter greatly facilitates gene knockouts, conditional expression, and overexpression of homologous and heterologous genes in Tetrahymena thermophila. Proc Natl Acad Sci U S A. 2002 Mar 19;99(6):3734-9. [CrossRef]
- Boldrin, F., Santovito G., Gaertig J., Wloga D., Cassidy-Hanley D., Clark T.G., Piccinni E. Metallothionein gene from Tetrahymena thermophila with a copper-inducible-repressible promoter. Eukaryot Cell. 2006 Feb;5(2):422-5. [CrossRef]
- Formigari, A., Boldrin F., Santovito G., Cassidy-Hanley D., Clark T.G., Piccinni E. Functional characterization of the 5’-upstream region of MTT5 metallothionein gene from Tetrahymena thermophila. Protist. 2010 Jan;161(1):71-7. [CrossRef]
- de Francisco, P., Amaro F., Martín-González A., Gutiérrez J.C. AP-1 (bZIP) Transcription Factors as Potential Regulators of Metallothionein Gene Expression in Tetrahymena thermophila. Front Genet. 2018 Oct 23;9:459. [CrossRef]
- Kling, P., Modig C., Mujahed H., Khalaf H., von Hofsten J., Olsson P.E. Differential regulation of the rainbow trout (Oncorhynchus mykiss) MT-A gene by nuclear factor interleukin-6 and activator protein-1. BMC Mol Biol. 2013 Dec 17;14:28. [CrossRef]
- Dalton, T., Palmiter R.D., Andrews G.K. Transcriptional induction of the mouse metallothionein-I gene in hydrogen peroxide-treated Hepa cells involves a composite major late transcription factor/antioxidant response element and metal response promoter elements. Nucleic Acids Res. 1994 Nov 25;22(23):5016-23. [CrossRef]
- Dalton, T.P., Li Q., Bittel D., Liang L., Andrews G.K. Oxidative stress activates metal-responsive transcription factor-1 binding activity. Occupancy in vivo of metal response elements in the metallothionein-I gene promoter. J Biol Chem. 1996 Oct 18;271(42):26233-41. [CrossRef]
- Fenton, H.J.H. Oxidation of tartaric acid in presence of iron. J Chem Soc, 1894 65, 899-910. [CrossRef]
- Haber F., andWeiss J. The catalytic decomposition of hydrogen peroxide by iron salts Proc. R. Soc. Lond.1934 A147332–35. [CrossRef]
- Kienzle, N., Young D., Zehntner S., Bushell G., Sculley T.B. DNaseI treatment is a prerequisite for the amplification of cDNA from episomal-based genes. Biotechniques. 1996 Apr;20(4):612-6. [CrossRef]
- Shi, X., Liu X., Wang Q., Das A., Ma G., Xu L., Sun Q., Peddireddi L., Jia W., Liu Y., Anderson G., Bai J., Shi J. A multiplex real-time PCR panel assay for simultaneous detection and differentiation of 12 common swine viruses. J Virol Methods. 2016 Oct;236:258-265. [CrossRef]
- Bustin, S.A., Benes V., Garson J.A., Hellemans J., Huggett J., Kubista M., Mueller R., Nolan T., Pfaffl M.W., Shipley G.L., Vandesompele J., Wittwer C.T. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem. 2009 Apr;55(4):611-22. [CrossRef]
- Sanchis-Segura, C., Miquel M., Correa M., Aragon C.M. The catalase inhibitor sodium azide reduces ethanol-induced locomotor activity. Alcohol. 1999 Aug;19(1):37-42. [CrossRef]
- Heikkila R.E., Cabbat F.S., Cohen G. In vivo inhibition of superoxide dismutase in mice by diethyldithiocarbamate. J Biol Chem. 1976 doi:10;251(7):2182-5.
- Zuo, Y., Hu J., Xu X., Gao X., Wang Y., Zhu S. Sodium azide induces mitochondria-mediated apoptosis in PC12 cells through Pgc-1α-associated signaling pathway. Mol Med Rep. 2019 Mar;19(3):2211-2219. [CrossRef]
- de Freitas Oliveira J.W., Torres T.M., Moreno C.J.G., Amorim-Carmo B., Damasceno I.Z., Soares A.K.M.C., da Silva Barbosa J., Rocha H.A.O., Silva M.S. Insights of antiparasitic activity of sodium diethyldithiocarbamate against different strains of Trypanosoma cruzi. Sci Rep. 2021 May 27;11(1):11200. [CrossRef]
- Gebicka, L., Krych-Madej J. The role of catalases in the prevention/promotion of oxidative stress. J Inorg Biochem. 2019 Aug;197:110699. [CrossRef]
- Wang, Y., Branicky R., Noë A., Hekimi S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J Cell Biol. 2018 Jun 4;217(6):1915-1928. [CrossRef]
- Karin, M., Liu Zg, Zandi E. AP-1 function and regulation. Curr Opin Cell Biol. 1997 Apr;9(2):240-6. [CrossRef]
- Valko, M., Morris H., Cronin M.T. Metals, toxicity and oxidative stress. Curr Med Chem. 2005;12(10):1161-208. [CrossRef]
- Chanarat, S., Svasti J. Stress-induced upregulation of the ubiquitin-relative Hub1 modulates pre-mRNA splicing and facilitates cadmium tolerance in Saccharomyces cerevisiae. Biochim Biophys Acta Mol Cell Res. 2020 Feb;1867(2):118565. [CrossRef]
- Sornaraj, P., Luang S., Lopato S., Hrmova M. Basic leucine zipper (bZIP) transcription factors involved in abiotic stresses: A molecular model of a wheat bZIP factor and implications of its structure in function. Biochim Biophys Acta. 2016;1860(1PtA):46-56. [CrossRef]
- Amaro, F., González D., Gutiérrez J.C. MicroRNAs in Tetrahymena thermophila: An epigenetic regulatory mechanism in the response to cadmium stress. Microbiol Res. 2024 Mar;280:127565. [CrossRef]
- Dondero, F., Cavaletto M., Ghezzi A.R., La Terza A., Banni M., Viarengo A. Biochemical characterization and quantitative gene expression analysis of the multi-stress inducible metallothionein from Tetrahymena thermophila. Protist. 2004 Jun;155(2):157-68. [CrossRef]
- Guo, R., Ma H., Gao F., Zhong L., Ren J. Metallothionein alleviates oxidative stress-induced endoplasmic reticulum stress and myocardial dysfunction. J Mol Cell Cardiol. 2009 Aug;47(2):228-37. [CrossRef]
- Martínez-Pastor, M.T., Perea-García A., Puig S. Mechanisms of iron sensing and regulation in the yeast Saccharomyces cerevisiae. World J Microbiol Biotechnol. 2017 Apr;33(4):75. [CrossRef]
- Askwith, C., Eide D., Van Ho A., Bernard P.S., Li L., Davis-Kaplan S., Sipe D.M., Kaplan J. The FET3 gene of S. cerevisiae encodes a multicopper oxidase required for ferrous iron uptake. Cell. 1994 Jan 28;76(2):403-10. [CrossRef]
- Mach, J., Sutak R. Iron in parasitic protists - from uptake to storage and where we can interfere. Metallomics. 2020 Sep 23;12(9):1335-1347. [CrossRef]
- Farnaud, S., Rapisarda C., Bui T., Drake A., Cammack R., Evans R.W. Identification of an iron-hepcidin complex. Biochem J. 2008 Aug 1;413(3):553-7. [CrossRef]
- Nemeth, E., Tuttle M.S., Powelson J., Vaughn M.B., Donovan A., Ward D.M., Ganz T., Kaplan J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science. 2004 Dec 17;306(5704):2090-3. [CrossRef]
- Laity, J.H., Andrews G.K. Understanding the mechanisms of zinc-sensing by metal-response element binding transcription factor-1 (MTF-1). Arch Biochem Biophys. 2007 Jul 15;463(2):201-10. [CrossRef]
- Greenberg, J.T., Demple B. A global response induced in Escherichia coli by redox-cycling agents overlaps with that induced by peroxide stress. J Bacteriol. 1989 Jul;171(7):3933-9. [CrossRef]
- Li, H., Mapolelo D.T., Dingra N.N., Naik S.G., Lees N.S., Hoffman B.M., Riggs-Gelasco P.J., Huynh B.H., Johnson M.K., Outten C.E. The yeast iron regulatory proteins Grx3/4 and Fra2 form heterodimeric complexes containing a [2Fe-2S] cluster with cysteinyl and histidyl ligation. Biochemistry. 2009 Oct 13;48(40):9569-81. [CrossRef]
- Bell, S.G., Vallee B.L. The metallothionein/thionein system: an oxidoreductive metabolic zinc link. Chembiochem. 2009 Jan 5;10(1):55-62. [CrossRef]
Figure 1.
Metallothionein mRNA relative abundances (box-plot) after exposure to 100 μM H2O2 and 100 μM H2O2 plus 200 μM FeCl3 at 24h (left column) and 72h (right column). (a) Mtt1 at 24h, (b) Mtt2/4 at 24h, (c) Mtt5 at 24h, (d) Mtt1 at 72h, (e) Mtt2/4 at 72h and (f) Mtt5 at 72h. The asterisks above each bar indicate the statistical significance of the difference in gene expression compared to control non-exposed cells. The inlet horizontal lines link the conditions that were further compared in contrasts using ANOVA. [**** p < 0.0001; (ANOVA, Tukey’s multiple comparison test)].
Figure 1.
Metallothionein mRNA relative abundances (box-plot) after exposure to 100 μM H2O2 and 100 μM H2O2 plus 200 μM FeCl3 at 24h (left column) and 72h (right column). (a) Mtt1 at 24h, (b) Mtt2/4 at 24h, (c) Mtt5 at 24h, (d) Mtt1 at 72h, (e) Mtt2/4 at 72h and (f) Mtt5 at 72h. The asterisks above each bar indicate the statistical significance of the difference in gene expression compared to control non-exposed cells. The inlet horizontal lines link the conditions that were further compared in contrasts using ANOVA. [**** p < 0.0001; (ANOVA, Tukey’s multiple comparison test)].
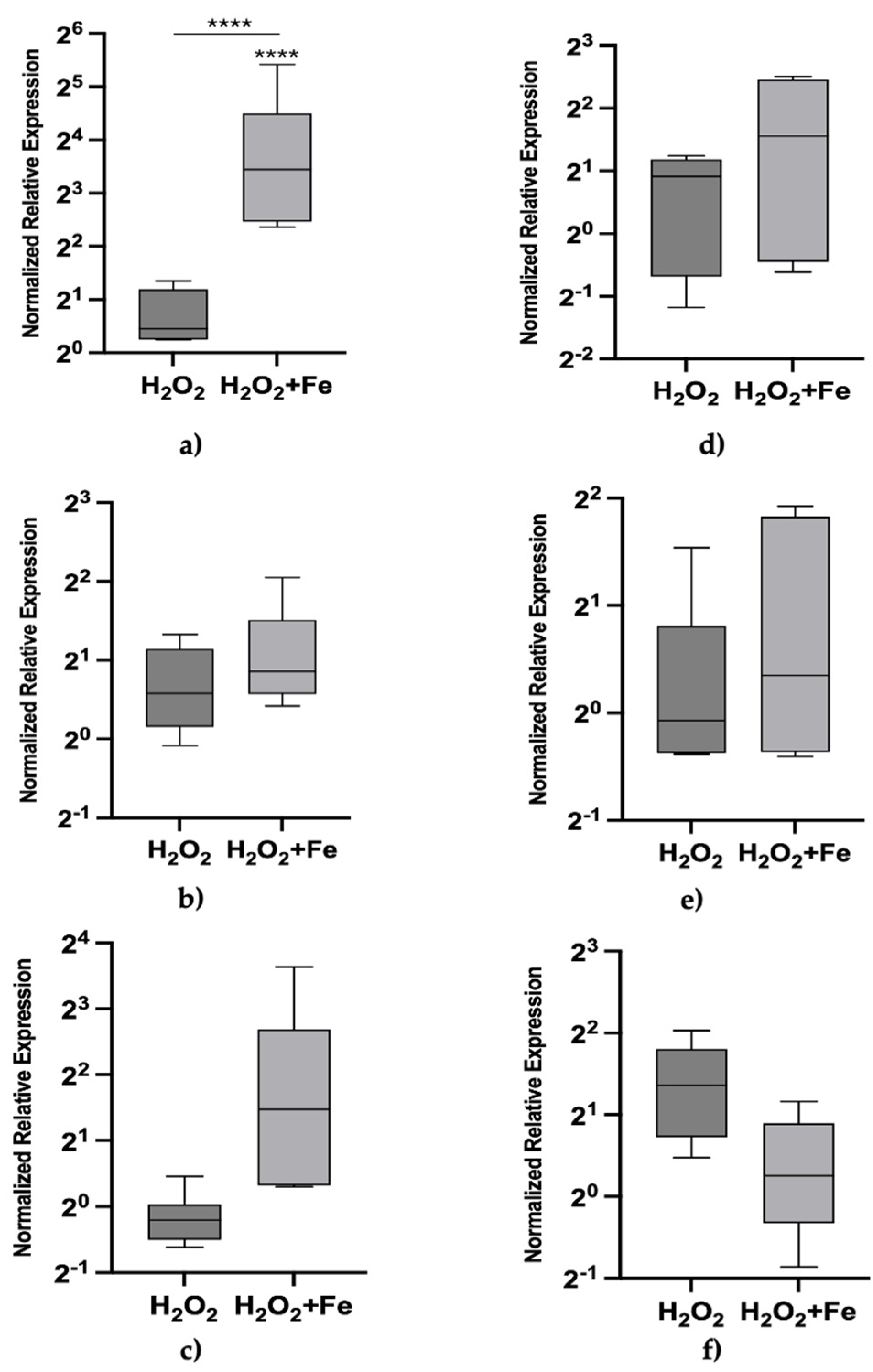
Figure 2.
Metallothionein mRNA relative abundances (box-plots) after 24 h (left column) and 72h (right colum) exposure to 20μM CdCl2, 200 μM FeCl3, 200 μM sodium azide (SAN), sodium azide plus iron (SAN+Fe), 100 μM sodium diethyldithiocarbamate (DTN) and sodium diethyldithiocarbamate plus FeCl3; (a) Mtt1 24 h; (b) Mtt2/4 24 h; (c) Mtt5 24 h; (d) Mtt1 72 h; (e) Mtt2/4 72 h and (f) Mtt5 72 h. The asterisks above each bar indicate the statistical significance of the difference in gene expression compared to control non-exposed cells. The inlet horizontal lines link the conditions that were further compared in contrasts using ANOVA. Significance levels: [**** p < 0.0001; *** p < 0.001; ** p < 0.01; * p < 0.05; ° p < 0.1 (a, b, c Welch’s ANOVA, Dunnet T3 multiple comparison test; e, f, g ANOVA, Tueky’s multiple comparison test)]. Cadmium was used as a positive control and was not included in contrasts.
Figure 2.
Metallothionein mRNA relative abundances (box-plots) after 24 h (left column) and 72h (right colum) exposure to 20μM CdCl2, 200 μM FeCl3, 200 μM sodium azide (SAN), sodium azide plus iron (SAN+Fe), 100 μM sodium diethyldithiocarbamate (DTN) and sodium diethyldithiocarbamate plus FeCl3; (a) Mtt1 24 h; (b) Mtt2/4 24 h; (c) Mtt5 24 h; (d) Mtt1 72 h; (e) Mtt2/4 72 h and (f) Mtt5 72 h. The asterisks above each bar indicate the statistical significance of the difference in gene expression compared to control non-exposed cells. The inlet horizontal lines link the conditions that were further compared in contrasts using ANOVA. Significance levels: [**** p < 0.0001; *** p < 0.001; ** p < 0.01; * p < 0.05; ° p < 0.1 (a, b, c Welch’s ANOVA, Dunnet T3 multiple comparison test; e, f, g ANOVA, Tueky’s multiple comparison test)]. Cadmium was used as a positive control and was not included in contrasts.
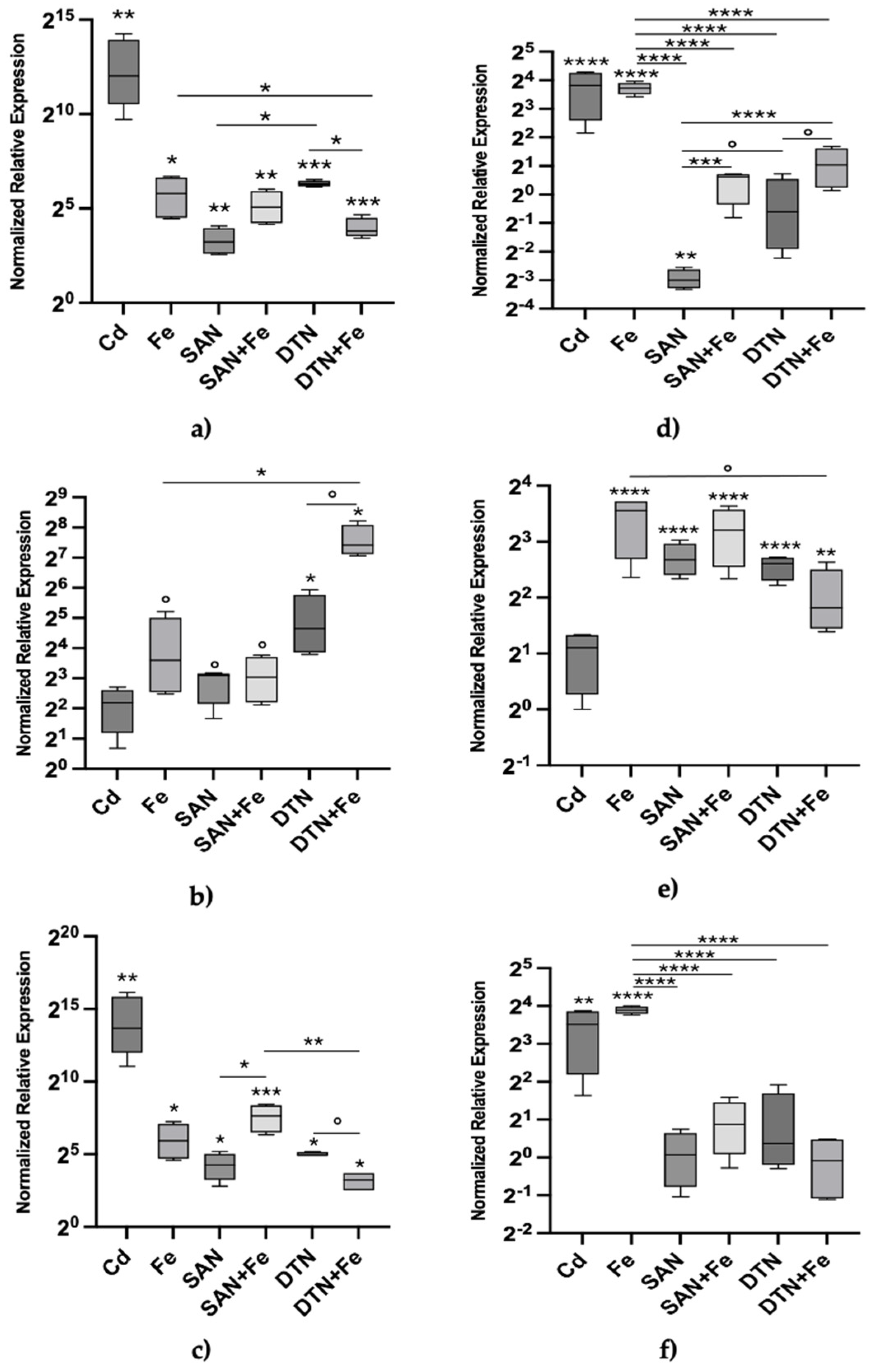
Figure 3.
Metallothionein mRNA relative abundances (box-plots) after 2h (left column), 24h (center column) & 72h (right column) exposure to iron. (a) Mtt1 2h; (b) Mtt2/4 2h; (c) Mtt5 2h; (d) Mtt1 24h; (e) Mtt2/4 24h; (f) Mtt5 24h; (g) Mtt1 72h; (h) Mtt2/4 72h & (i) Mtt5 72h. Significance levels: The asterisks above each bar indicate the statistical significance of the difference in gene expression compared to control non-exposed cells [** p < 0.001; * p < 0.05; ° p < 0.1 (Welch’s ANOVA, Dunnet T3 multiple comparison test vs not-exposed reference control)].
Figure 3.
Metallothionein mRNA relative abundances (box-plots) after 2h (left column), 24h (center column) & 72h (right column) exposure to iron. (a) Mtt1 2h; (b) Mtt2/4 2h; (c) Mtt5 2h; (d) Mtt1 24h; (e) Mtt2/4 24h; (f) Mtt5 24h; (g) Mtt1 72h; (h) Mtt2/4 72h & (i) Mtt5 72h. Significance levels: The asterisks above each bar indicate the statistical significance of the difference in gene expression compared to control non-exposed cells [** p < 0.001; * p < 0.05; ° p < 0.1 (Welch’s ANOVA, Dunnet T3 multiple comparison test vs not-exposed reference control)].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



