
Submitted:
14 September 2024
Posted:
17 September 2024
You are already at the latest version
Abstract
Concerns regarding the safety of ethoxyquin (EQ) have gradually increased, highlighting the necessity of identifying safer and more effective antioxidants for use in animal feed. This study investigates the effects of a compound antioxidant (CA) as an alternative to ethoxyquin on feed oxidative stability, growth performance, antioxidant capacity, and intestinal health in broilers. A total of 351 one-day-old Arbor Acres Plus male broilers were randomly assigned to three treatments with nine replicates of 13 birds each. The experimental treatments included: CON group (basal diet), EQ group (basal diet + 200 g/ton EQ at 60% purity), and CA group (basal diet + 200 g/ton compound antioxidant, which comprises 18% butylated hydroxytoluene, 3% citric acid, and 1% tertiary butylhydroquinone). The experiment lasted for 21 days. Compared to the control group, supplementation with CA significantly reduced the acid value, peroxide value, and malondialdehyde content in the feeds after storage. Moreover, CA supplementation significantly decreased feed intake and the feed conversion ratio (FCR) without affecting weight gain of boilers, while also improving liver antioxidant capacity (increased T-AOC and SOD levels), intestinal antioxidant capacity (increased CAT, HO-1, and GPX7 protein expression), and enhancing the duodenal villus height-to-crypt depth ratio and jejunal villus height. Additionally, CA treatment reduced intestinal barrier permeability (decreased diamine oxidase activity and D-lactate levels), potentially by promoting the expression of Occludin, ZO-1, and Mucin2 protein. Furthermore, the addition of CA elevated the protein levels of NF-κB p50 along with its inhibitory factors, IκB and NF-κB p105, in the intestines of broilers, thereby enhancing immune regulatory capacity. This supplementation also led to an increased abundance of beneficial gut microorganisms, including Lactobacillus, Alistipes, Candidatus Arthromitus, and Turicibacter. In comparison to the EQ group, CA supplementation enhanced feed oxidative stability, decreased the FCR, and improved the intestinal morphology, barrier functions, and inflammatory status in broilers. These findings suggest that this compound antioxidant may serve as a healthier and more effective alternative to ethoxyquin.
Keywords:
compound antioxidant
; ethoxyquin
; broiler
; antioxidant capacity
; intestinal health
1. Introduction
Animal diets that are rich in lipids, particularly polyunsaturated fatty acids, are prone to oxidation. This oxidation process may be exacerbated by factors such as high temperature, light and metal ions during feed production [1]. The oxidation of feed lipids not only diminishes the nutritional value of the feed [2], but also adversely affect the normal physiological functions of the animal [3,4]. The consumption of oxidized diets can destroy the intestinal villus morphology and intestinal barrier, reduce the antioxidant capacity of the body, promote intestinal inflammation and lead to disturbances in the intestinal microbiota, ultimately impacting the production performance of animals [5,6]. Incorporating antioxidants into feed to mitigate feed oxidation is an effective strategy. Ethoxyquin (EQ) is the most widely used antioxidant in feed due to its low cost and high antioxidant capacity [7]. However, the safety of ethoxyquin and its metabolites is of concern [8]. Recent studies have further characterized that ethoxyquin and its metabolites can remain in different tissues of animals, which may pose a potential hazard for consumers [9,10]. Consequently, there is an urgent need to identify safer and more effective antioxidant alternatives.
Butylated hydroxytoluene (BHT) is also a widely utilized antioxidant [11]. Recent studies indicate that its favorable safety profile enables it to significantly enhance plasma and liver lipid metabolism, as well as antioxidant and inflammatory responses, even when administered at ten times the maximum allowable additive dose of BHT (1500 mg/kg) to Micropterus salmoides [12]. Tertiary butylhydroquinone (TBHQ) is another antioxidant that can be used for antioxidant protection in food, and it is generally believed that its antioxidant capacity surpasses that of BHT [13]; however, its higher price limits its applicability. Recent research has highlighted the potential synergistic effects of combining antioxidants with varying properties [14,15]. A synergistic effect of BHT and TBHQ in stabilizing soybean biodiesel has been found in vitro [16]. Additionally, citric acid is frequently added as a synergist in feeds to chelate metal ions from containers and the environment, thereby mitigating the pro-oxidative effects of these metal ions [17,18]. Studies indicate that citric acid and TBHQ exhibit a strong synergistic effect, significantly reducing the peroxide value of peanuts during storage [19]. Given the potential synergistic effects of different antioxidants, along with the enhancement provided by chelating metal ions of citric acid, a compound antioxidant comprising BHT, TBHQ, and citric acid may demonstrate exhibit superior efficacy. This could be a healthier and more efficient solution for antioxidant protection than ethoxyquin. Current research on antioxidant combinations primarily emphasizes in vitro study, while compound antioxidants formulated based on antioxidant synergy are rarely incorporated into animal diets. This study aims to integrate a compound antioxidant (CA) blend of BHT, TBHQ, and citric acid into broiler diets to examine its protective antioxidant effects on the diets and its implications for broiler performance, as well as for antioxidant capacity and intestinal health. Additionally, the study compares this blend with ethoxyquin to evaluate its antioxidant properties.
2. Materials and Methods
2.1. Animals and Experimental Design
All animal procedures used in this experiment were approved by the Institutional Animal Care and Use Committee of the China Agricultural University, Beijing, China (Permit Number: AW52704202-1-1). A total of 351 one-day-old Arbor Acres Plus male broilers were randomly assigned to three groups, each consisting of nine replicates with 13 chickens per replicate. The treatments included: CON group (basal diet), EQ group (basal diet supplemented with 200 g/ton of EQ at 60% purity, effective content 120 g/ton), and CA group (basal diet supplemented with 200 g/ton of a compound antioxidant, containing 18% BHT, effective content 36 g/ton; 3% citric acid, effective content 6 g/ton; and 1% TBHQ, effective content 2 g/ton). EQ, BHT, citric acid, and TBHQ were purchased from Beijing Sunpu Biochemical and Technical CO., LTD. The experiment lasted for 21 days. Feed ingredients were crushed, mixed, and subsequently granulated into pellet form, and the nutritional analysis of the basal diet is detailed in Table 1.
All broilers were housed in cages and had ad libitum access to food and water throughout the feeding period. The animal management practices adhered to the standards established for Arbor Acres Plus broilers.
2.2. Feed Oxidation Stability Testing
After storing the feed for a designated period, oil samples were extracted to test the oxidation stability index. The specific procedures are as follows: Following the completion of feed preparation, 1.2 kg of each feed was allocated to three replicates situated at distinct locations within the chicken house. During the 3rd and 6th weeks, 200 g of feed was sampled, ground into a powder, passed through a 40-mesh screen, and preserved at -20°C. Subsequently, the oil was extracted to identify indicators associated with oxidation stability. The oil extraction process involved taking a suitably weighted sample and placing it in a wide-mouth bottle. Approximately four times the weight of petroleum ether was added and the mixture was stirred with a glass rod before tightly sealing the bottle was sealed tightly. The bottle was then shaken on a reciprocating oscillator for two hours. Subsequently, the solution and sample were transferred to a 50 mL centrifuge tube and centrifuged at 8000 r/min for five minutes. The supernatant was collected and transferred to a rotary evaporator with the water bath temperature maintained below 40°C. The solvent was evaporated under negative pressure, yielding the remaining liquid grease as the final sample. The determination of acid value (AV) and peroxide value (POV) were referred to Ma et al. [20]. The content of malondialdehyde (MDA) content was measured using commercial kits (# BC0025, Solarbio).
2.3. Growth Performance
On day 21, after a 5-hour fasting period, the body weight of the broilers and the remaining feed per cage were measured. Feed intake (FI) and body weight gain (BWG) were subsequently calculated to determine the feed conversion rate (FCR).
2.4. Sample Collection
At 21 days of age, a healthy chick close to the average weight, was selected from every replicate, electric shock fainting stunned and then bled through the carotid artery. A 4-mL blood sample was collected and placed in a coagulation tube. After centrifugation at 3,000 × g for 10 minutes at 4°C, the serum was separated and stored at -20°C. The small intestine was removed, and approximately 1 cm sections from the middle regions of the duodenum and jejunum were collected and fixed in a 4% paraformaldehyde solution for subsequent intestinal morphology measurements. Additionally, liver tissue, jejunal tissue, and cecal contents were collected and immediately frozen in liquid nitrogen, then stored at -80°C until further analysis.
2.5. Liver Antioxidant Capacity
The activities of total antioxidant capacity (T-AOC, #A015-2), superoxide dismutase (SOD), catalase (CAT, #A007-1), glutathione peroxidase (GSH-Px, #A005-1), glutathione (GSH, #A006-2), and malondialdehyde (MDA, #A003-1) in the liver were evaluated and normalized based on total protein content (BCA, #A045-4) using kits from the Nanjing Jiancheng Bioengineering Institute.
2.6. Intestinal Morphology
Tissue sections were fixed in 4% paraformaldehyde, embedded in paraffin, sliced, dehydrated, and stained with hematoxylin and eosin by Sevier Biotechnology Co., Ltd. (Wuhan, China). Measurements of villus height (VH), crypt depth (CD), and the ratio of villus height to crypt depth (VCR) were conducted using a Leica DM 750 optical microscope (Leica Microsystems, Wetzlar, Germany).
2.7. Intestinal Barrier Permeability
Diamine oxidase (DAO) activity (#BC1285) and D-lactate content (#BC5355) in serum were measured according to the protocols provided by kits from Beijing Solarbio.
2.8. Expression of Intestinal Barrier, Antioxidant, and Inflammation-Related Proteins
Jejunal mucosa (100 mg) was homogenized in 1 mL of RIPA lysis solution (Beijing Solarbio Biotech, China) with 1mM phenylmethylsulfonyl fluoride using a SCIENTZ-12 high-throughput tissue homogenizer (Xinzhi Biotech, China), followed by cell disruption with a tissue disruptor (Xinzhi Biotech, China). After incubation on ice for 30 minutes, the samples were centrifuged at 12,000 × g for 15 minutes to collect the supernatant. Soluble proteins were separated by SDS-PAGE gel electrophoresis (100 V, 2 hours), transferred to a nitrocellulose membrane (120 V, 40 minutes), and subsequently blocked with 5% skimmed milk powder (Sangon Biotech, China). The NC membrane was then probed with primary antibodies overnight at 4°C followed by incubation with secondary antibodies for 1 hour at room temperature. Western blot bands were scanned and analyzed using a western blot fully automated chemiluminescence imaging system, with band density normalized to β-actin content. The primary antibodies used were as follows: Occludin (#DF7504) and ZO-1 (#AF5154) were from Affinity Bioscience; MUC2 (#sc-515032) and HO-1 (#sc-136960) were form Santa Cruz; KEAP1 (#10503-2-AP), NRF2 (#16396-1-AP), GPX7 (#13501-1-AP), NQO1 (#11451-1-AP), TLR4 (#19811-1-AP), IκB (#10268-1-AP), and NF-κB1 (p50 and p105) (#14220-1-AP) were from Proteintech. β-actin (#I102) served as the internal reference protein (Bioworld Biotech). The secondary antibodies employed included Anti-rabbit HRP secondary antibody (#7074) and Anti-mouse HRP secondary antibody (#7076), both from Cell Signaling Technology.
2.9. Expression of Intestinal Inflammation-Related Genes
Total RNA was extracted using RNAiso Plus (#9108, Takara, Japan) and transcribed into cDNA with the Prime Script™ RT reagent Kit with gDNA Eraser (#RR047A, Takara, Japan). Real-time fluorescence quantitative reaction was performed with the SYBR Premix Ex Taq kit (#RR420A, Takara, Japan) on an ABI7500 fluorescence quantitative PCR instrument (Applied Biosystems, USA). All primer sequences for target genes are presented in Table 2. The relative expression levels of each gene were normalized to β-actin and analyzed using the 2-ΔΔCt method.
2.10. Cecal Microbiota
Cecal chyme samples were subjected to high-throughput sequencing of the 16S rRNA gene V3-V4 amplicon at Shanghai Paison Biotechnology Co., Ltd., utilizing the Illumina MiSeq platform (primer sequence: F: ACTCCTACGGGAGGCAGCA; R: GGACTACHVGGGTWTCTAAT). The raw sequencing data were denoised using the QIIME2 dada2 analysis process to obtain Amplicon Sequence Variants (ASVs) with 97% similarity. Subsequently, the ASVs were annotated with the Greengenes 13.8 database. Based on the species annotation results, both alpha and beta diversity were calculated, and differences between groups were compared to elucidate the distinct characteristics of microbial community structures under varying treatments.
2.11. Statistical Analysis
One-way ANOVA was conducted using SPSS 26.0 software, and multiple comparisons were performed using Ducan’s method. Percentages were compared with an inverse chord correction. Results are presented as mean ± standard error of the mean (SEM). Statistical significance was accepted at p < 0.05, while a statistical trend was considered when 0.05 ≤ p < 0.1. Graphs were generated by GraphPad Prism 10.
3. Results
3.1. Oxidative Stability of Feeds
Supplemental EQ or CA in the diet significantly decreased the AV and POV of feeds after three and six weeks of storage (p < 0.05) (Figure 1A,B). Notably, the CA group exhibited a lower AV than the EQ group at the sixth week (p < 0.05). Furthermore, supplementation with CA significantly reduced MDA content in the diets at both the third and sixth weeks compared to the control and EQ groups (p < 0.05) (Figure 1C).
3.2. Growth Performance
Supplemental CA or EQ all decreased FI of broilers (p = 0.044) (Figure 2B). Adding CA in the diet intended to increase the BWG compared to the EQ group (0.05 ≤ p < 0.1) (Figure 2A); however, there was no significant difference between the EQ and control groups. Additionally, supplemental CA to the diet significantly decreased the FCR compared to both the control and EQ groups (p = 0.002), with no differences observed between the latter two (Figure 2C).
3.3. Liver Antioxidant Capacity
Both dietary additions of CA and EQ significantly increased T-AOC (p < 0.001) and SOD (p = 0.006) activity in the livers of broilers compared to the control group (Figure 3A,B). However, no significant difference was observed between CA and EQ group.
3.4. Intestinal Morphology
In comparison to the control group, dietary supplementation with EQ tended to decrease duodenal VH (p = 0.067) (Table 3). Conversely, dietary CA supplementation significantly increased the VCR in the duodenum and jejunal VH when compared to both the control group and the EQ group (p < 0.001). In addition, compared with EQ group, the VCR in jejunum was significantly increased in the CA group (p = 0.001).
3.5. Intestinal Barrier Permeability and Expression of Related Proteins
Dietary CA supplementation significantly reduced serum DAO activity and D-lactate levels compared to both the control group and the EQ group (p < 0.05) (Figure 4A,B). Furthermore, the incorporation of CA markedly enhanced the relative protein expression of ZO-1, Occludin, and MUC2 (p < 0.05) (Figure 4C–F). Additionally, the inclusion of EQ led to a significant increase solely in the relative expression of Occludin (p = 0.020) (Figure 4C,E).
3.6. Intestinal Antioxidant Capacity
Compared with EQ group, the relative expressions of intestinal KEAP1 protein (p = 0.075) and NRF2 protein (p < 0.05) in the CA group were found to be lower (Figure 5A–C). In addition, the dietary addition of both CA and EQ significantly increased the relative expressions of CAT (p = 0.007) and GPX7 (p = 0.017) compared to the control group, with a trend towards increased HO-1 expression (p = 0.052) (Figure 5D–G). Notably, the expression of NQO1 protein in the CA group was higher than that observed in the EQ group (p = 0.014) (Figure 5E,H).
3.7. Expression of Intestinal Inflammation-Related Cytokines
In comparison to the control group, dietary supplementation with EQ significantly elevated the mRNA expression levels of IL-1β, IL-6, and IFN-γ (p < 0.05) (Figure 6A). Conversely, CA supplementation led to a significant reduction in TNF-α gene expression (p < 0.05). Both the EQ and CA diets significantly upregulated NF-κB p50 and NF-κB p105 when compared to the control group (p < 0.05) (Figure 6B–D). Additionally, the EQ group exhibited the highest expression levels of TLR4 protein and IκB protein among all groups (p < 0.05) (Figure 6E,F).
3.8. Cecal Microbiota
3.8.1. Microbial Diversity
The intestinal α diversity and β diversity of broilers under different treatments are presented in Figure 7. The Venn diagram illustrates that the control group, EQ group, and CA group contain 1685, 1150, and 982 unique ASVs, respectively, with 393 ASVs shared among all groups. Notably, the number of common ASVs between the CA and CON groups, as well as between the CA and EQ groups, is 112 and 107, respectively, which are much lower than the number of common ASVs (237) observed between the EQ and CON groups. No significant differences were found in the richness and diversity of cecal microbial communities among the groups (p > 0.05). The rarefaction curve based on the Chao1 index further indicates that, at the same sequencing depth, the Chao1 index for the CA group is lower than that of the control and EQ groups. Based on ASV level cluster analysis, principal coordinate analysis (PCoA) and non-metric multidimensional scaling (NMDS) results were plotted to evaluate beta diversity among all groups. The results of the PCoA and NMDS analyses indicated that the characteristic points of CA group were significantly separated from other groups.
3.8.2. Microbial Composition Analysis
The taxonomic composition of the top 10 phyla and the top 20 genera in the cecal microbial communities of broilers in each group was analyzed (Figure 8). At the phylum level, Firmicutes and Bacteroidetes were identified as the dominant bacterial groups in the cecal community of AA broilers, exhibiting average relative abundances of 80.64% and 17.39%, respectively. The heat map indicated that the CA group displayed a higher relative abundance of Firmicutes, Chlorobi, and Cyanobacteria, while the relative abundances of Actinobacteria, Spirochaetes, Bacteroidetes, and Chloroflexi were lower than those in the other groups. In comparison to the control group, the addition of compound antioxidants to the diet tended to increase the relative abundance of Firmicutes, while simultaneously reducing the relative abundance of Bacteroidetes and the ratio of Bacteroidetes to Firmicutes (0.05 ≤ p < 0.1). At the genus level, the dominant genera identified in broilers across all groups included Lactobacillus, Bacteroides, Faecalibacterium, Oscillospira, Enterococcus, Candidatus Arthromitus, and Parabacteroides, among others. The heat map analysis revealed that Lactobacillus, Candidatus Arthromitus, Alistipes, and Turicibacter were enriched in the CA group. Furthermore, compared to the control group, dietary supplementation with CA tended to increase the relative abundance of Lactobacilli in the cecum of broilers (p = 0.065). Additionally, it significantly reduced the relative abundance of Bacteroides and Coprococcus (p < 0.05) and also indicated a tendency to decrease the relative abundance of Anaeroplasma (p = 0.093).
4. Discussion
Lipid oxidation leads to the production of free fatty acids, with the acid value (AV) serving as an indicator of the concentration of these free fatty acids generated during the oxidation process [21]. Additionally, lipid oxidation is associated with the formation of hydroperoxides, which can be quantified by measuring the peroxide value (POV) [22]. Malondialdehyde (MDA), the end product of lipid oxidation, is widely used as a marker to assess the extent of lipid deterioration [23]. This study found that the addition of CA could effectively decreased the AV, POV, and MDA during storage, demonstrating greater efficacy than EQ after six weeks of storage.
The addition of BHT [24,25] and TBHQ [26] alone has a limited impact on broiler performance. Another study found that including citric acid can significantly enhance the weight gain of broilers; however, its effective dosage is as high as 5000 g/ton [27], and it remains unclear whether lower dosages would yield similar effects. Recent research indicates that these antioxidants can effectively enhance animal growth performance when incorporated in a complex form [28,29]. In this study, the inclusion of CA in the diet significantly reduced the feed conversion ratio compared to both the control and EQ groups, aligning with findings from previous studies. The improved performance of broilers with CA in this study may be attributed to the reduced degree of feed oxidation, improved liver and intestinal antioxidant capacity, and intestinal health [30,31].
The liver serves as the primary site for regulating oxidative stress, maintaining redox balance through a series of antioxidant enzymes [32]. Superoxide dismutase (SOD) promotes the degradation of superoxide anions and is the first line of defense of antioxidant systems. Additionally, total antioxidant capacity (T-AOC) can be utilized to assess the overall activity of antioxidant enzymes and substances present in tissues [33]. This study showed that dietary CA supplementation can significantly increase SOD activity and T-AOC in broiler livers. This is similar to the results of some previous studies that added BHT or citric acid alone [12,27].
The intestine, a vital organ responsible for the digestion and absorption of nutrients, is directly connected to the external environment and is particularly susceptible to oxidative damage from dietary sources and environmental factors. Maintaining redox balance is essential for optimal intestinal health [34]. As a transcriptional activator responsible for cellular redox regulation, NRF2 directly regulates the expression of numerous antioxidant proteins and enzymes that regulate biochemical reactions, including catalase (CAT), glutathione peroxidase (GPX7), heme oxygenase-1 (HO-1), and NAD(P)H quinone dehydrogenase 1 (NQO1) [35]. Both CAT and GPX7 play crucial roles in the elimination of peroxides within the body. In addition to its protective effects against oxidative damage, HO-1 has recently been recognized for its significant anti-inflammatory properties [36]. NQO1 contributes to the prevention of oxidative stress by detoxifying superoxide and facilitating the antioxidant forms of physiological substrates, including ubiquinone and vitamin E quinone [37]. This study demonstrates that dietary supplementation with CA or EQ all effectively enhances the expression of antioxidant protection genes, including CAT, HO-1, GPX7, and NQO1. Additionally, Kelch-like ECH-associated protein 1 (KEAP1) acts as a negative regulator of NRF2; its downregulation leads to increased translocation of NRF2 into the nucleus, subsequently inducing the transcription of genes responsible for antioxidant protection [38]. Notably, EQ appears to elevate the expression of its downstream genes by upregulating the relative levels of NRF2 protein, while CA promotes greater nuclear entry of NRF2 through the downregulation of KEAP1, thereby facilitating antioxidant protection within the organism. Further analysis is required to elucidate the detailed mechanisms involved.
The villus height (VH) and villi-to-crypt ratio (VCR) in the intestine are directly positively correlated with the digestive and absorptive capacity of the small intestine [39]. Compared with the control group, adding CA to the diet significantly increased duodenal VCR and jejunal VH. In addition, the duodenal VCR, jejunal VH and VCR in the CA group were higher than those in the EQ group. Citric acid, as the main components of CA, has been shown to improve intestinal villi morphology [40]. Broilers in CA group with more mature intestinal villi may contribute to the digestion and absorption of nutrients, and ultimately improve the growth performance of broilers. Serum levels of diamine oxidase (DAO) and D-Lactate serve as indicators of intestinal barrier permeability. Compared to the control and EQ groups, the administration of CA significantly reduced serum levels of DAO and D-Lactate in broilers, which suggests the reduced intestinal barrier permeability of broilers in CA group. Intestinal tight junction (TJ) proteins, including Occludin and ZO1, are essential components of the intestinal barrier [41]. TJ damages lead to an increase of the intestinal permeability and result in the access of intraluminal antigens or bacteria into the mucosa [42]. Additionally, MUC2 is an intestinal mucin secreted by goblet cells, forming a mucus layer on the intestinal epithelial surface that serves to resist microbial invasion [43]. This study found that the addition of CA significantly increases the expression of Occludin, ZO1, and MUC2 in the intestines of broilers, which may explain the observed improvement in intestinal barrier permeability.
Intestinal inflammation frequently leads to damage of intestinal cells [44], resulting in maldigestion and malabsorption of nutrients, which subsequently impairs host growth and development [45]. Compared to the control group, the inclusion of EQ in the diet significantly elevated the gene expression of IL-1β, IL-6, and IFN-γ. Conversely, dietary supplementation with CA decreased the relative expression of TNF-α. In alignment with these results, BHT and citric acid have been shown to mitigate inflammatory responses[12,40]. TLR4, a receptor of the innate immune system, is activated by lipopolysaccharide (LPS) produced by Gram-negative bacteria, leading to the stimulation of inflammatory cytokine production [46]. The high expression of TLR4 observed in the EQ group in this study may have induced the expression of IL-1β, IL-6, and IFN-γ. Furthermore, an intact intestinal barrier can prevent the leakage of LPS, thereby reducing TLR4 activation [47]. Conversely, the lower expression of inflammatory factors in the CA group may be attributed to reduced TLR4 expression and an improved intestinal barrier. NF-κB is a crucial regulator of inflammation that plays a protective role in intestinal epithelial cells, which is essential for maintaining epithelial integrity and intestinal immune homeostasis. A deficiency in NF-κB results in the apoptosis of intestinal epithelial cells, leading to tissue damage and bacterial translocation into the mucosa [48]. NF-κB p50, is a significant member of the NF-κB family [49]. The precursor of p50, NF-κB p105, exerts a negative regulatory function on p50 [50]. Mice deficient in NF-κB p105 but expressing mature NF-κB p50 spontaneously develop intestinal inflammation [51]. IκB is a protein that inhibits the overactivation of NF-κB by maintaining it in an inactive state within the cytoplasm, thereby preventing its binding to DNA [52]. The current study found that the addition of EQ or CA to the diet all up-regulated NF-κB p50 and its inhibitory factors NF-κB p105 and IκB, concurrently, which may enhance the regulatory capacity of immune homeostasis.
The large number of microorganisms present in the intestine forms a specific microecological system with the host, participating in the host’s metabolism, growth, and immunity [53]. Studies have demonstrated that antioxidants can influence host health by altering the composition of intestinal microorganisms [54,55]. To investigate the impact of CA on the intestinal microorganisms of broilers, we conducted an analysis of microbial differences across each group using 16S rDNA sequencing. The results indicated that the CA group shared fewer ASVs in common with the other, with distinct separation of its bacterial flora characteristics from those of the other groups. The CA group showed a higher abundance of Firmicutes and a lower abundance of Bacteroidetes compared to the control group, resulting in a decreased ratio of Bacteroidetes to Firmicutes. Firmicutes, primarily Gram-positive bacteria, play a crucial role in host nutrition and metabolism through the synthesis of short-chain fatty acids, particularly butyrate. Butyrate serves as an energy source for the development of intestinal epithelial cells, strengthens the intestinal barrier, and exhibits anti-inflammatory and antibacterial properties [56,57]. The growth performance of animals is positively correlated with the abundance of Firmicutes and negatively correlated with the ratio of Bacteroidetes to Firmicutes [58,59]. The increased abundance of Firmicutes observed in the broilers within the CA group in this study may enhance their production performance. In addition, Bacteroidetes preferentially consumes proteins in mucin to sustain its survival [60]. The reduced abundance of Bacteroidetes in the CA group may have contributed to an increased richness of mucin, thereby enhancing the integrity of the intestinal barrier. Moreover, the CA group was enriched in Lactobacillus, Candidatus Arthromitus, Alistipes, and Turicibacter at genus level. Candidatus Arthromitus plays a critical role in the maturation of the host gut immune barrier, inducing both innate and adaptive immune responses [61,62]. Furthermore, higher-performing flocks exhibit a greater proportion of Candidatus Arthromitus, suggesting that these bacteria may enhance intestinal health and protect commercial turkeys from pathogens that adversely affect production parameters [63]. Additionally, Turicibacter has been shown to promote host lipid metabolism by modifying bile acids [64]. Alistipes are important producers of short-chain fatty acids (SCFAs), recognized as growth promoters for broilers [65,66]. Moreover, some members of Alistipes possess anti-inflammatory and hyperuricemia-relieving benefits [67,68]. Further analysis comparing the CA group to the control group indicated that the relative abundance of Lactobacillus in the broilers of the CA group increased, whereas the relative abundance of Bacteroidetes, Coprococcus, and Anaeroplasma decreased. Lactobacilli, belonging to the phylum Firmicutes, are the most prevalent probiotics that produce lactic acid, acetic acid, and propionic acid, which can lower intestinal pH and inhibit the growth of various pathogenic bacteria, thereby maintaining the balance of intestinal flora [69]. Incorporating Lactobacillus into broiler diets has been shown to enhance the intestinal barrier of broilers, regulate intestinal microbiota, and provide immune protection [70,71]. Anaeroplasma is an opportunistic pathogen whose abundance significantly increases in mice with inflammation-driven colon cancer [72,73]. This study suggests that incorporating the compound antioxidants into the diet may increase the abundance of beneficial gut microbes, consequently exerting a favorable influence on the host.
5. Conclusions
This study demonstrates that a novel compound antioxidant, composed of 18% BHT, 3% citric acid, and 1% TBHQ, can effectively delay feed oxidation and enhance the growth performance of broilers by improving their antioxidant capacity and intestinal health (Figure 9). This compound antioxidant exhibits superior effects compared to ethoxyquin in both in vivo and in vitro experiments.
Author Contributions
Conceptualization, Y.X. and J.Y.; methodology, Y.X.; software, Y.X.; validation, Y.X. and X.G.; formal analysis, X.Y. and X.G.; investigation, Y.X.; resources, Y.X.; data curation, Y.X.; writing—original draft preparation, Y.X.; writing—review and editing, X.G. and J.Y.; visualization, Y.X.; supervision, J.Y.; project administration, J.Y.; funding acquisition, J.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Key Research and Development Program of China, grant number 2021YFD1300404.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Animal Care and Use Committee of the China Agricultural University (Permit Number: AW52704202-1-1).
Informed Consent Statement
Not applicable.
Data Availability Statement
The 16S rRNA amplicon sequencing reads from this dataset have been deposited in the National Center for Biotechnology Information under BioProjectID PRJNA1158563. Additionally, the data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shahidi, F.; Zhong, Y. Lipid Oxidation and Improving the Oxidative Stability. Chem. Soc. Rev. 2010, 39, 4067–4079. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Liu, Y.; Chen, F. Effect of Storage Temperature on Lipid Oxidation and Changes in Nutrient Contents in Peanuts. Food Sci. Nutr. 2019, 7, 2280–2290. [Google Scholar] [CrossRef]
- Guo, Q.; Zhang, L.; Yin, Y.; Gong, S.; Yang, Y.; Chen, S.; Han, M.; Duan, Y. Taurine Attenuates Oxidized Fish Oil-Induced Oxidative Stress and Lipid Metabolism Disorder in Mice. Antioxidants 2022, 11, 1391. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Guan, X.; Zhang, W.; Han, T.; Liu, X.; Shi, B. Oxidized Soybean Oil Evoked Hepatic Fatty Acid Metabolism Disturbance in Rats and Their Offspring. J. Agric. Food Chem. 2023, 71, 13483–13494. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Mahmood, T.; Tang, Z.; Wu, Y.; Yuan, J. Effects of Naturally Oxidized Corn Oil on Inflammatory Reaction and Intestinal Health of Broilers. Poult. Sci. 2022, 101, 101541. [Google Scholar] [CrossRef]
- Gao, F.; Liu, H.; Du, Y.; Fang, X.; Cheng, B.; Shi, B. Dietary Resveratrol Ameliorates Hepatic Fatty Acid Metabolism and Jejunal Barrier in Offspring Induced by Maternal Oxidized Soybean Oil Challenge. J. Agric. Food Chem. 2024, 72, 3730–3740. [Google Scholar] [CrossRef] [PubMed]
- Błaszczyk, A.; Augustyniak, A.; Skolimowski, J. Ethoxyquin: An Antioxidant Used in Animal Feed. Int. J. Food Sci. 2013, 2013, 1–12. [Google Scholar] [CrossRef]
- Błaszczyk, A.; Skolimowski, J. Cytotoxicity and Genotoxicity of Ethoxyquin Used As an Antioxidant. Food Rev. Int. 2015, 31, 222–235. [Google Scholar] [CrossRef]
- Zhang, C.; Gai, X.; Tian, Y.; Wang, J.; He, D.; Yang, W.; Zhang, L.; Chen, Y. Analysis of Ethoxyquin and Its Oxidation Products in Swine Tissues by Gas Chromatography-Tandem Mass Spectrometry for Evaluating the Feed-to-Animal Tissue Transfer of Ethoxyquin and Its Metabolites. J. Anim. Sci. Biotechnol. 2021, 12, 8. [Google Scholar] [CrossRef]
- Tan, X.Y.; Chin, Z.X.; Chua, S.L.; Lee, K.K.M.; Wu, Y.; Chan, J.S.H. Determination of Ethoxyquin by Ultra-High Performance Liquid Chromatography with Tandem Mass Spectrometry and a Singapore Survey of Ethoxyquin Residues in Eggs, Egg Products and Poultry. Food Addit. Contam. Part A 2024, 41, 261–270. [Google Scholar] [CrossRef]
- Yehye, W.A.; Rahman, N.A.; Ariffin, A.; Abd Hamid, S.B.; Alhadi, A.A.; Kadir, F.A.; Yaeghoobi, M. Understanding the Chemistry behind the Antioxidant Activities of Butylated Hydroxytoluene (BHT): A Review. Eur. J. Med. Chem. 2015, 101, 295–312. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.L.; Yu, H.H.; Liang, X.F.; Li, N.; Wang, X.; Li, F.H.; Wu, X.F.; Zheng, Y.H.; Xue, M.; Liang, X.F. Dietary Butylated Hydroxytoluene Improves Lipid Metabolism, Antioxidant and Anti-Apoptotic Response of Largemouth Bass (Micropterus Salmoides). Fish Shellfish Immunol. 2018, 72, 220–229. [Google Scholar] [CrossRef]
- Wong, Y.H.; Goh, K.M.; Nyam, K.L.; Nehdi, I.A.; Sbihi, H.M.; Tan, C.P. Effects of Natural and Synthetic Antioxidants on Changes in 3-MCPD Esters and Glycidyl Ester in Palm Olein during Deep-Fat Frying. Food Control 2019, 96, 488–493. [Google Scholar] [CrossRef]
- Bayram, I.; Decker, E.A. Underlying Mechanisms of Synergistic Antioxidant Interactions during Lipid Oxidation. Trends Food Sci. Technol. 2023, 133, 219–230. [Google Scholar] [CrossRef]
- Li, J.; Chen, J.; Bi, Y.; Yang, H. Insight into Synergistic Antioxidation Mechanisms of Butyl Hydroxyanisole with Common Synthetic Antioxidants. Grain Oil Sci. Technol. 2022, 5, 114–130. [Google Scholar] [CrossRef]
- Borsato, D.; Cini, J.R. de M.; Silva, H.C. da; Coppo, R.L.; Angilelli, K.G.; Moreira, I.; Maia, E.C.R. Oxidation Kinetics of Biodiesel from Soybean Mixed with Synthetic Antioxidants BHA, BHT and TBHQ: Determination of Activation Energy. Fuel Process. Technol. 2014, 127, 111–116. [Google Scholar] [CrossRef]
- Ghorbani Gorji, S.; Smyth, H.E.; Sharma, M.; Fitzgerald, M. Lipid Oxidation in Mayonnaise and the Role of Natural Antioxidants: A Review. Trends Food Sci. Technol. 2016, 56, 88–102. [Google Scholar] [CrossRef]
- Yu, Z.; Lu, L.; Lu, L.; Pan, L.; Qiu, X.; Tang, Y. Development and Antioxidation of Metal Ion Chelating Packaging Film. Food Packag. Shelf Life 2022, 32, 100846. [Google Scholar] [CrossRef]
- Ma, P.; Wen, H.; Chen, X.; Zhang, W.; Rong, L.; Luo, Y.; Xie, J. Synergistic Rosemary Extract with TBHQ and Citric Acid Improves Oxidative Stability and Shelf Life of Peanut. J. Food Sci. 2024, 89, 3591–3602. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, C.; Liu, C.; Tan, J.; Ma, H.; Wang, J. Physiochemical Responses of the Kernel Quality, Total Phenols and Antioxidant Enzymes of Walnut in Different Forms to the Low-Temperature Storage. Foods 2021, 10, 2027. [Google Scholar] [CrossRef]
- Mahesar, S.A.; Sherazi, S.T.H.; Khaskheli, A.R.; Kandhro, A.A.; Uddin, S. Analytical Approaches for the Assessment of Free Fatty Acids in Oils and Fats. Anal. Methods 2014, 6, 4956–4963. [Google Scholar] [CrossRef]
- Zhang, N.; Li, Y.; Wen, S.; Sun, Y.; Chen, J.; Gao, Y.; Sagymbek, A.; Yu, X. Analytical Methods for Determining the Peroxide Value of Edible Oils: A Mini-Review. Food Chem. 2021, 358, 129834. [Google Scholar] [CrossRef]
- Vandemoortele, A.; Heynderickx, P.M.; Leloup, L.; De Meulenaer, B. Kinetic Modeling of Malondialdehyde Reactivity in Oil to Simulate Actual Malondialdehyde Formation upon Lipid Oxidation. Food Res. Int. 2021, 140, 110063. [Google Scholar] [CrossRef] [PubMed]
- Farahat, M.; Abdallah, F.; Abdel-Hamid, T.; Hernandez-Santana, A. Effect of Supplementing Broiler Chicken Diets with Green Tea Extract on the Growth Performance, Lipid Profile, Antioxidant Status and Immune Response. Br. Poult. Sci. 2016, 57, 714–722. [Google Scholar] [CrossRef]
- Farahat, M.H.; Abdallah, F.M.; Ali, H.A.; Hernandez-Santana, A. Effect of Dietary Supplementation of Grape Seed Extract on the Growth Performance, Lipid Profile, Antioxidant Status and Immune Response of Broiler Chickens. animal 2017, 11, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.T.; Hanson, A.R.; Urriola, P.E.; Johnston, L.J.; Kerr, B.J.; Shurson, G.C. Addition of Tert-Butylhydroquinone (TBHQ) to Maize Oil Reduces Lipid Oxidation but Does Not Prevent Reductions in Serum Vitamin E in Nursery Pigs. J. Anim. Sci. Biotechnol. 2019, 10, 51. [Google Scholar] [CrossRef]
- Fikry, A.M.; Attia, A.I.; Ismail, I.E.; Alagawany, M.; Reda, F.M. Dietary Citric Acid Enhances Growth Performance, Nutrient Digestibility, Intestinal Microbiota, Antioxidant Status, and Immunity of Japanese Quails. Poult. Sci. 2021, 100, 101326. [Google Scholar] [CrossRef]
- Chen, X.; Zeng, D.; Zeng, X.; Zeng, Q. Effects of Complex Antioxidants Added to Chicken Diet on Growth Performance, Serum Biochemical Indices, Meat Quality, and Antioxidant Capacity. Animals 2024, 14, 360. [Google Scholar] [CrossRef]
- Zhao, W.; Li, J.; Xing, T.; Zhang, L.; Gao, F. Effects of Guanidinoacetic Acid and Complex Antioxidant Supplementation on Growth Performance, Meat Quality, and Antioxidant Function of Broiler Chickens. J. Sci. Food Agric. 2021, 101, 3961–3968. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative Stress Mitigation by Antioxidants - An Overview on Their Chemistry and Influences on Health Status. Eur. J. Med. Chem. 2021, 209, 112891. [Google Scholar] [CrossRef]
- Ayo, J.O.; Ogbuagu, N.E. Heat Stress, Haematology and Small Intestinal Morphology in Broiler Chickens: Insight into Impact and Antioxidant-Induced Amelioration. Worlds Poult. Sci. J. 2021, 77, 949–968. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.-Y.; Wang, N.; Zhang, Z.-J.; Lao, L.; Wong, C.-W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed]
- Tao, Q.; Zhang, Z.-D.; Qin, Z.; Liu, X.-W.; Li, S.-H.; Bai, L.-X.; Ge, W.-B.; Li, J.-Y.; Yang, Y.-J. Aspirin Eugenol Ester Alleviates Lipopolysaccharide-Induced Acute Lung Injury in Rats While Stabilizing Serum Metabolites Levels. Front. Immunol. 2022, 13, 939106. [Google Scholar] [CrossRef] [PubMed]
- Circu, M.L.; Aw, T.Y. Intestinal Redox Biology and Oxidative Stress. Semin. Cell Dev. Biol. 2012, 23, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [PubMed]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of Inflammation by the Antioxidant Haem Oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef]
- Ross, D.; Siegel, D. NQO1 in Protection against Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 67–72. [Google Scholar] [CrossRef]
- Onoki, T.; Izumi, Y.; Takahashi, M.; Murakami, S.; Matsumaru, D.; Ohta, N.; Wati, S.M.; Hatanaka, N.; Katsuoka, F.; Okutsu, M.; et al. Skeletal Muscle-Specific Keap1 Disruption Modulates Fatty Acid Utilization and Enhances Exercise Capacity in Female Mice. Redox Biol. 2021, 43, 101966. [Google Scholar] [CrossRef]
- Wang, M.; Yang, C.; Wang, Q.; Li, J.; Huang, P.; Li, Y.; Ding, X.; Yang, H.; Yin, Y. The Relationship between Villous Height and Growth Performance, Small Intestinal Mucosal Enzymes Activities and Nutrient Transporters Expression in Weaned Piglets. J. Anim. Physiol. Anim. Nutr. 2020, 104, 606–615. [Google Scholar] [CrossRef]
- Hu, P.; Yuan, M.; Guo, B.; Lin, J.; Yan, S.; Huang, H.; Chen, J.-L.; Wang, S.; Ma, Y. Citric Acid Promotes Immune Function by Modulating the Intestinal Barrier. Int. J. Mol. Sci. 2024, 25, 1239. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of Intestinal Epithelial Permeability by Tight Junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.; Turner, J.R. Cell Biology of Tight Junction Barrier Regulation and Mucosal Disease. Cold Spring Harb. Perspect. Biol. 2018, 10, a029314. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Wu, Y.; Wusiman, A.; Gu, P.; Mao, N.; Xu, S.; Zhu, T.; Feng, Z.; Liu, Z.; Wang, D. Alhagi Honey Polysaccharides Attenuate Intestinal Injury and Immune Suppression in Cyclophosphamide-Induced Mice. Food Funct. 2021, 12, 6863–6877. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Carbonell, R.; Yao, S.-J.; Das, S.; Guma, M. Dysregulation of Intestinal Epithelial Cell RIPK Pathways Promotes Chronic Inflammation in the IBD Gut. Front. Immunol. 2019, 10, 1094. [Google Scholar] [CrossRef]
- Nii, T.; Bungo, T.; Isobe, N.; Yoshimura, Y. Intestinal Inflammation Induced by Dextran Sodium Sulphate Causes Liver Inflammation and Lipid Metabolism Disfunction in Laying Hens. Poult. Sci. 2020, 99, 1663–1677. [Google Scholar] [CrossRef]
- Pelaseyed, T.; Bergström, J.H.; Gustafsson, J.K.; Ermund, A.; Birchenough, G.M.H.; Schütte, A.; van der Post, S.; Svensson, F.; Rodríguez-Piñeiro, A.M.; Nyström, E.E.L.; et al. The Mucus and Mucins of the Goblet Cells and Enterocytes Provide the First Defense Line of the Gastrointestinal Tract and Interact with the Immune System. Immunol. Rev. 2014, 260, 8–20. [Google Scholar] [CrossRef]
- Rohr, M.W.; Narasimhulu, C.A.; Rudeski-Rohr, T.A.; Parthasarathy, S. Negative Effects of a High-Fat Diet on Intestinal Permeability: A Review. Adv. Nutr. 2020, 11, 77–91. [Google Scholar] [CrossRef]
- Nenci, A.; Becker, C.; Wullaert, A.; Gareus, R.; van Loo, G.; Danese, S.; Huth, M.; Nikolaev, A.; Neufert, C.; Madison, B.; et al. Epithelial NEMO Links Innate Immunity to Chronic Intestinal Inflammation. Nature 2007, 446, 557–561. [Google Scholar] [CrossRef]
- Sun, S.-C.; Chang, J.-H.; Jin, J. Regulation of Nuclear Factor-κB in Autoimmunity. Trends Immunol. 2013, 34, 282–289. [Google Scholar] [CrossRef]
- BEINKE, S.; LEY, S.C. Functions of NF-κB1 and NF-κB2 in Immune Cell Biology. Biochem. J. 2004, 382, 393–409. [Google Scholar] [CrossRef]
- Chang, M.; Lee, A.J.; Fitzpatrick, L.; Zhang, M.; Sun, S.-C. NF-κB1 P105 Regulates T Cell Homeostasis and Prevents Chronic Inflammation1. J. Immunol. 2009, 182, 3131–3138. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.-C. Non-Canonical NF-κB Signaling Pathway. Cell Res. 2011, 21, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Guarner, F.; Malagelada, J.-R. Gut Flora in Health and Disease. The Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Jiang, L.; Lin, Y.; Liu, Z. Antioxidant Activity of Selenium-Enriched Chrysomyia Megacephala (Fabricius) Larvae Powder and Its Impact on Intestinal Microflora in D-Galactose Induced Aging Mice. BMC Complement. Med. Ther. 2020, 20, 264. [Google Scholar] [CrossRef] [PubMed]
- Riaz Rajoka, M.S.; Thirumdas, R.; Mehwish, H.M.; Umair, M.; Khurshid, M.; Hayat, H.F.; Phimolsiripol, Y.; Pallarés, N.; Martí-Quijal, F.J.; Barba, F.J. Role of Food Antioxidants in Modulating Gut Microbial Communities: Novel Understandings in Intestinal Oxidative Stress Damage and Their Impact on Host Health. Antioxidants 2021, 10, 1563. [Google Scholar] [CrossRef]
- Segain, J.-P.; Blétière, D.R. de la; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C.; Ferrier, L.; Bonnet, C.; Blottière, H.M.; Galmiche, J.-P. Butyrate Inhibits Inflammatory Responses through NFκB Inhibition: Implications for Crohn’s Disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef]
- Zhang, F.; Wan, Y.; Zuo, T.; Yeoh, Y.K.; Liu, Q.; Zhang, L.; Zhan, H.; Lu, W.; Xu, W.; Lui, G.C.Y.; et al. Prolonged Impairment of Short-Chain Fatty Acid and L-Isoleucine Biosynthesis in Gut Microbiome in Patients With COVID-19. Gastroenterology 2022, 162, 548–561.e4. [Google Scholar] [CrossRef]
- Han, G.G.; Lee, J.-Y.; Jin, G.-D.; Park, J.; Choi, Y.H.; Chae, B.J.; Kim, E.B.; Choi, Y.-J. Evaluating the Association between Body Weight and the Intestinal Microbiota of Weaned Piglets via 16S rRNA Sequencing. Appl. Microbiol. Biotechnol. 2017, 101, 5903–5911. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, W.; Zhang, H.; Wang, J.; Zhang, W.; Gao, J.; Wu, S.; Qi, G. Supplemental Bacillus Subtilis DSM 32315 Manipulates Intestinal Structure and Microbial Composition in Broiler Chickens. Sci. Rep. 2018, 8, 15358. [Google Scholar] [CrossRef]
- Zeng, X.; Xing, X.; Gupta, M.; Keber, F.C.; Lopez, J.G.; Lee, Y.-C.J.; Roichman, A.; Wang, L.; Neinast, M.D.; Donia, M.S.; et al. Gut Bacterial Nutrient Preferences Quantified in Vivo. Cell 2022, 185, 3441–3456.e19. [Google Scholar] [CrossRef]
- Yang, Y.; Torchinsky, M.B.; Gobert, M.; Xiong, H.; Xu, M.; Linehan, J.L.; Alonzo, F.; Ng, C.; Chen, A.; Lin, X.; et al. Focused Specificity of Intestinal TH17 Cells towards Commensal Bacterial Antigens. Nature 2014, 510, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V.; et al. Induction of Intestinal Th17 Cells by Segmented Filamentous Bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Danzeisen, J.L.; Calvert, A.J.; Noll, S.L.; McComb, B.; Sherwood, J.S.; Logue, C.M.; Johnson, T.J. Succession of the Turkey Gastrointestinal Bacterial Microbiome Related to Weight Gain. PeerJ 2013, 1, e237. [Google Scholar] [CrossRef]
- Lynch, J.B.; Gonzalez, E.L.; Choy, K.; Faull, K.F.; Jewell, T.; Arellano, A.; Liang, J.; Yu, K.B.; Paramo, J.; Hsiao, E.Y. Gut Microbiota Turicibacter Strains Differentially Modify Bile Acids and Host Lipids. Nat. Commun. 2023, 14, 3669. [Google Scholar] [CrossRef]
- Torok, V.A.; Hughes, R.J.; Mikkelsen, L.L.; Perez-Maldonado, R.; Balding, K.; MacAlpine, R.; Percy, N.J.; Ophel-Keller, K. Identification and Characterization of Potential Performance-Related Gut Microbiotas in Broiler Chickens across Various Feeding Trials. Appl. Environ. Microbiol. 2011, 77, 5868–5878. [Google Scholar] [CrossRef]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria With Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef]
- Xu, Y.-X.; Liu, L.-D.; Zhu, J.-Y.; Zhu, S.-S.; Ye, B.-Q.; Yang, J.-L.; Huang, J.-Y.; Huang, Z.-H.; You, Y.; Li, W.-K.; et al. Alistipes Indistinctus-Derived Hippuric Acid Promotes Intestinal Urate Excretion to Alleviate Hyperuricemia. Cell Host Microbe 2024, 32, 366–381.e9. [Google Scholar] [CrossRef] [PubMed]
- Dziarski, R.; Park, S.Y.; Kashyap, D.R.; Dowd, S.E.; Gupta, D. Pglyrp-Regulated Gut Microflora Prevotella Falsenii, Parabacteroides Distasonis and Bacteroides Eggerthii Enhance and Alistipes Finegoldii Attenuates Colitis in Mice. PLOS ONE 2016, 11, e0146162. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, S.; Guo, J.; Xie, Q.; Evivie, S.E.; Song, Y.; Li, B.; Huo, G. The Protective Effects of Lactobacillus Plantarum KLDS 1.0344 on LPS-Induced Mastitis In Vitro and In Vivo. Front. Immunol. 2021, 12, 770822. [Google Scholar] [CrossRef]
- Zhang, H.; Pertiwi, H.; Hou, Y.; Majdeddin, M.; Michiels, J. Protective Effects of Lactobacillus on Heat Stress-Induced Intestinal Injury in Finisher Broilers by Regulating Gut Microbiota and Stimulating Epithelial Development. Sci. Total Environ. 2024, 918, 170410. [Google Scholar] [CrossRef]
- Wang, L.; Lin, Z.; Ali, M.; Zhu, X.; Zhang, Y.; Li, S.; Li, K.; Kebzhai, F.; Li, J. Effects of Lactic Acid Bacteria Isolated from Tibetan Chickens on the Growth Performance and Gut Microbiota of Broiler. Front. Microbiol. 2023, 14, 1171074. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, H.; Chen, W.; Yao, X.; Xing, Y.; Wang, X.; Zhong, J.; Meng, G. Mycoplasma Hyorhinis Activates the NLRP3 Inflammasome and Promotes Migration and Invasion of Gastric Cancer Cells. PLOS ONE 2013, 8, e77955. [Google Scholar] [CrossRef] [PubMed]
- Gates, T.J.; Yuan, C.; Shetty, M.; Kaiser, T.; Nelson, A.C.; Chauhan, A.; Starr, T.K.; Staley, C.; Subramanian, S. Fecal Microbiota Restoration Modulates the Microbiome in Inflammation-Driven Colorectal Cancer. Cancers 2023, 15, 2260. [Google Scholar] [CrossRef] [PubMed]
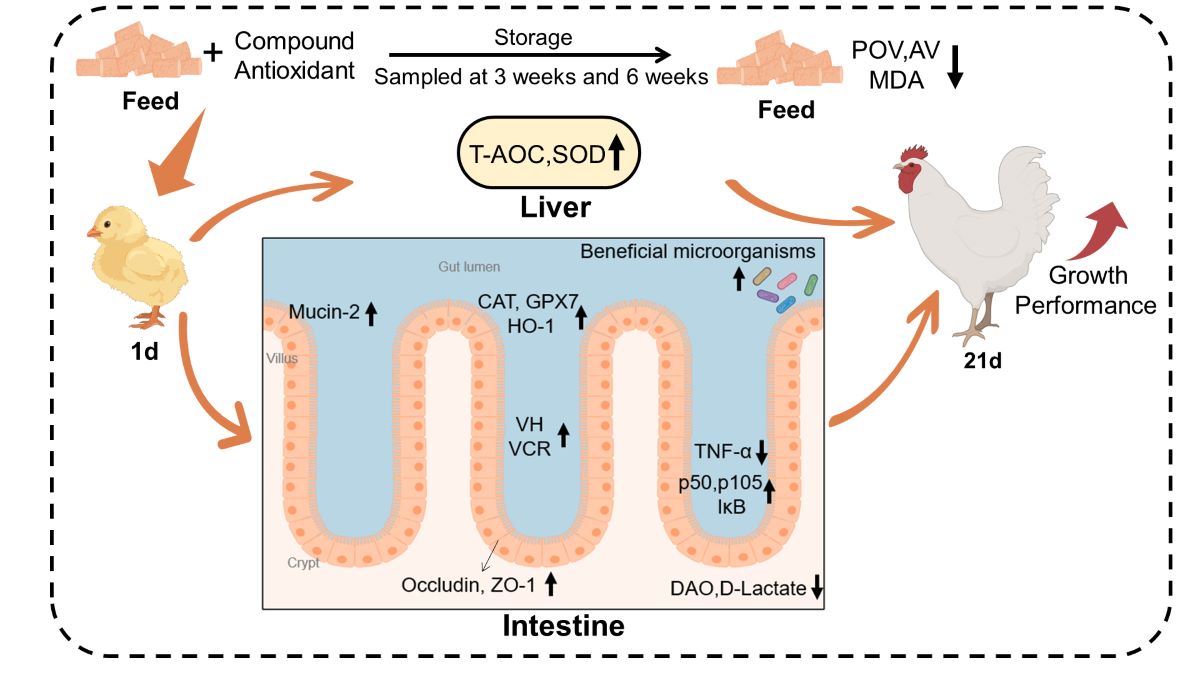
Figure 1.
Oxidative stability of feeds after 3 or 6 weeks of storage. N=3. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. Abbreviations: AV, acid value; POV, peroxide value; MDA, malondialdehyde. ab Different superscripts letters indicate significant differences (p < 0.05).
Figure 1.
Oxidative stability of feeds after 3 or 6 weeks of storage. N=3. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. Abbreviations: AV, acid value; POV, peroxide value; MDA, malondialdehyde. ab Different superscripts letters indicate significant differences (p < 0.05).
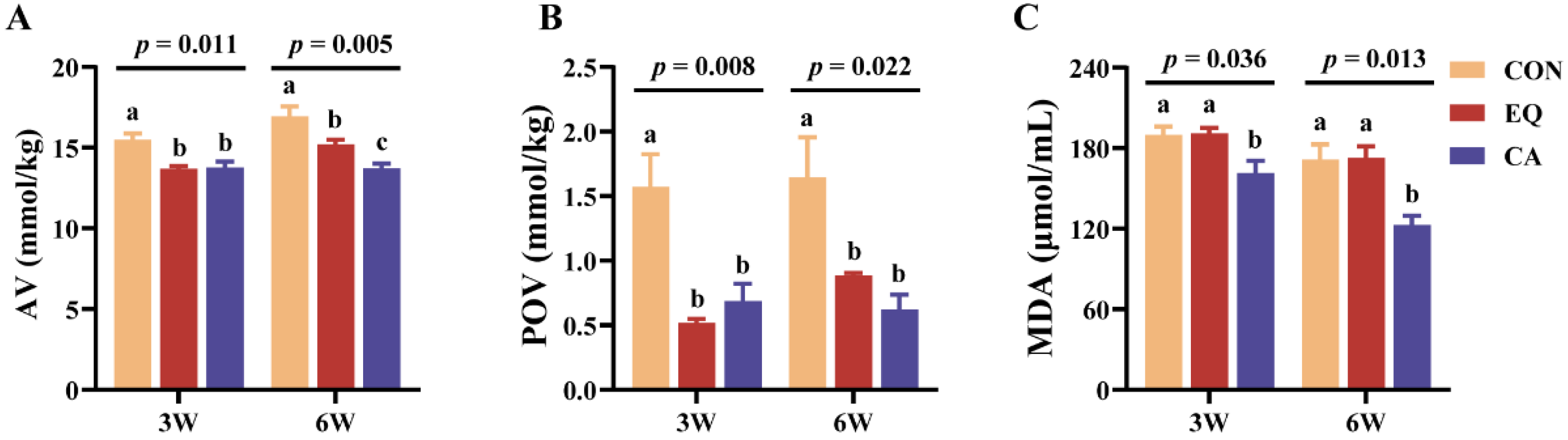
Figure 2.
Growth performance of broilers. N=9. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. Abbreviations: BWG, body weight gain; FI, feed intake; FCR, feed conversion ratio. ab Different superscripts letters indicate significant differences (p < 0.05).
Figure 2.
Growth performance of broilers. N=9. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. Abbreviations: BWG, body weight gain; FI, feed intake; FCR, feed conversion ratio. ab Different superscripts letters indicate significant differences (p < 0.05).
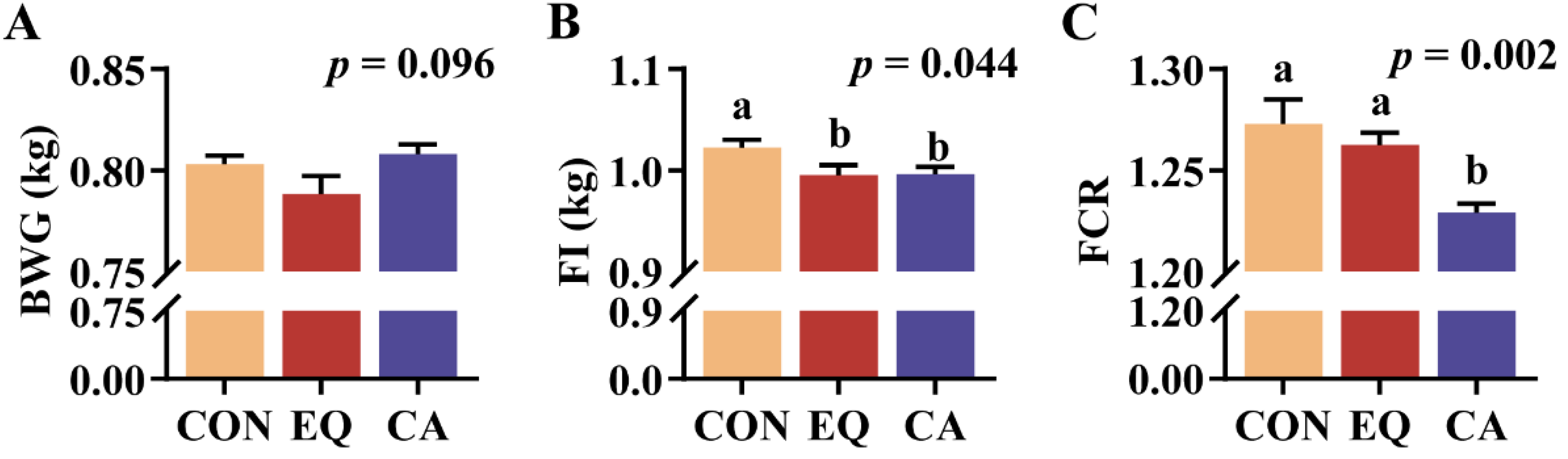
Figure 3.
Liver antioxidant capacity of broilers. N=8. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. Abbreviations: T-AOC, total antioxidant capacity; SOD, superoxide dismutase; CAT: catalase; GSH-Px, glutathione peroxidase; GSH, reduced glutathione; MDA, malondialdehyde. ab Different superscripts letters indicate significant differences (p < 0.05).
Figure 3.
Liver antioxidant capacity of broilers. N=8. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. Abbreviations: T-AOC, total antioxidant capacity; SOD, superoxide dismutase; CAT: catalase; GSH-Px, glutathione peroxidase; GSH, reduced glutathione; MDA, malondialdehyde. ab Different superscripts letters indicate significant differences (p < 0.05).
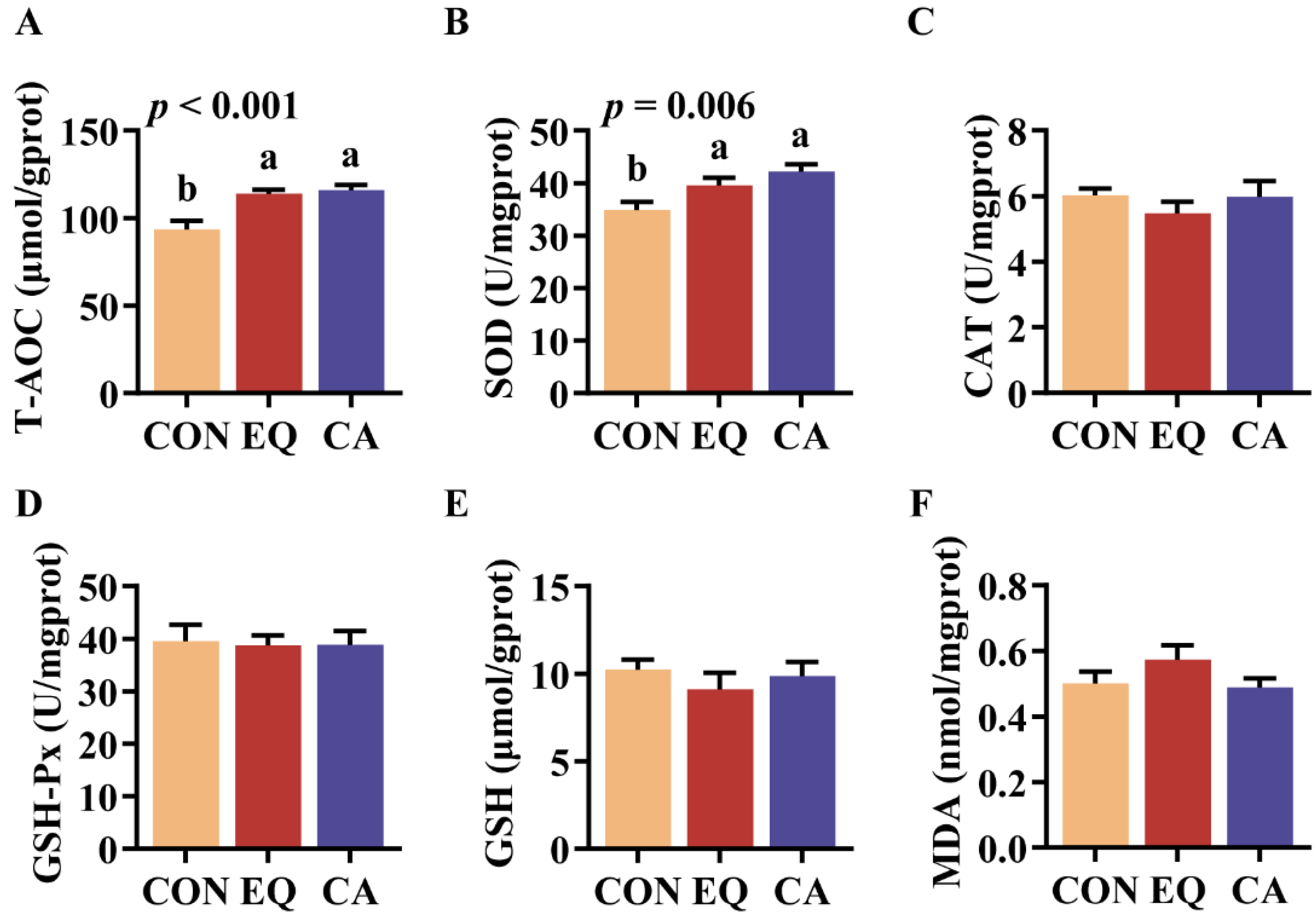
Figure 4.
Intestinal barrier permeability and intestinal barrier related protein expression. N=8. (A) Serum diamine oxidase activity and (B) D-lactate contents. (C-F) The relative protein expression of ZO-1, Occludin, and MUC2 in jejunum. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. Abbreviations: DAO, diamine oxidase. ab Different superscripts letters indicate significant differences (p < 0.05).
Figure 4.
Intestinal barrier permeability and intestinal barrier related protein expression. N=8. (A) Serum diamine oxidase activity and (B) D-lactate contents. (C-F) The relative protein expression of ZO-1, Occludin, and MUC2 in jejunum. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. Abbreviations: DAO, diamine oxidase. ab Different superscripts letters indicate significant differences (p < 0.05).
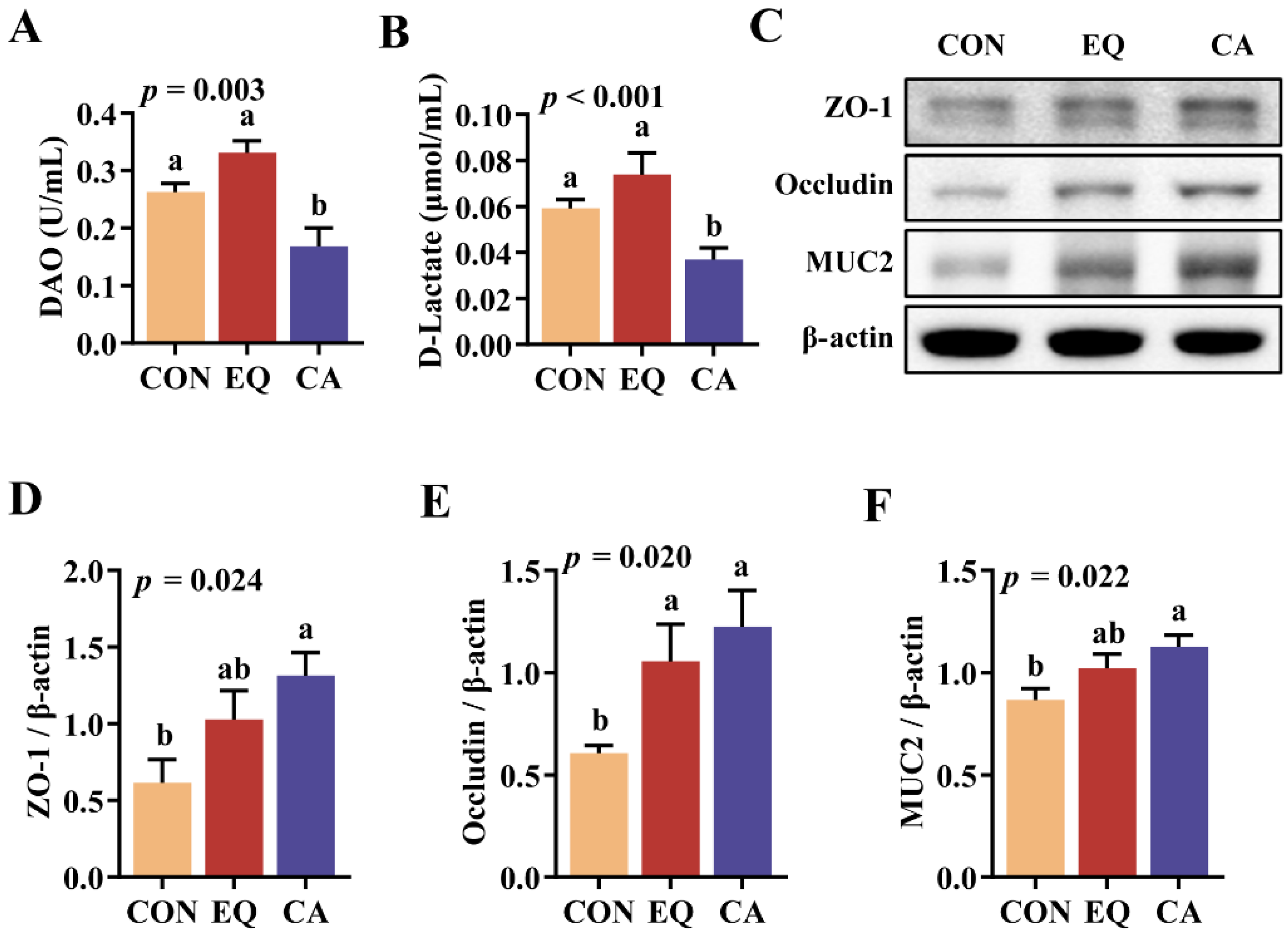
Figure 5.
Intestinal antioxidant related protein expression. N=8. (A-D) Relative protein levels of KEAP1, NRF2, and CAT in jejunum. (E-H) Relative protein levels of HO-1, GPX7 and NQO1 in jejunum. N=8. CON, basal diet; CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. ab Different superscripts letters indicate significant differences (p < 0.05).
Figure 5.
Intestinal antioxidant related protein expression. N=8. (A-D) Relative protein levels of KEAP1, NRF2, and CAT in jejunum. (E-H) Relative protein levels of HO-1, GPX7 and NQO1 in jejunum. N=8. CON, basal diet; CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. ab Different superscripts letters indicate significant differences (p < 0.05).
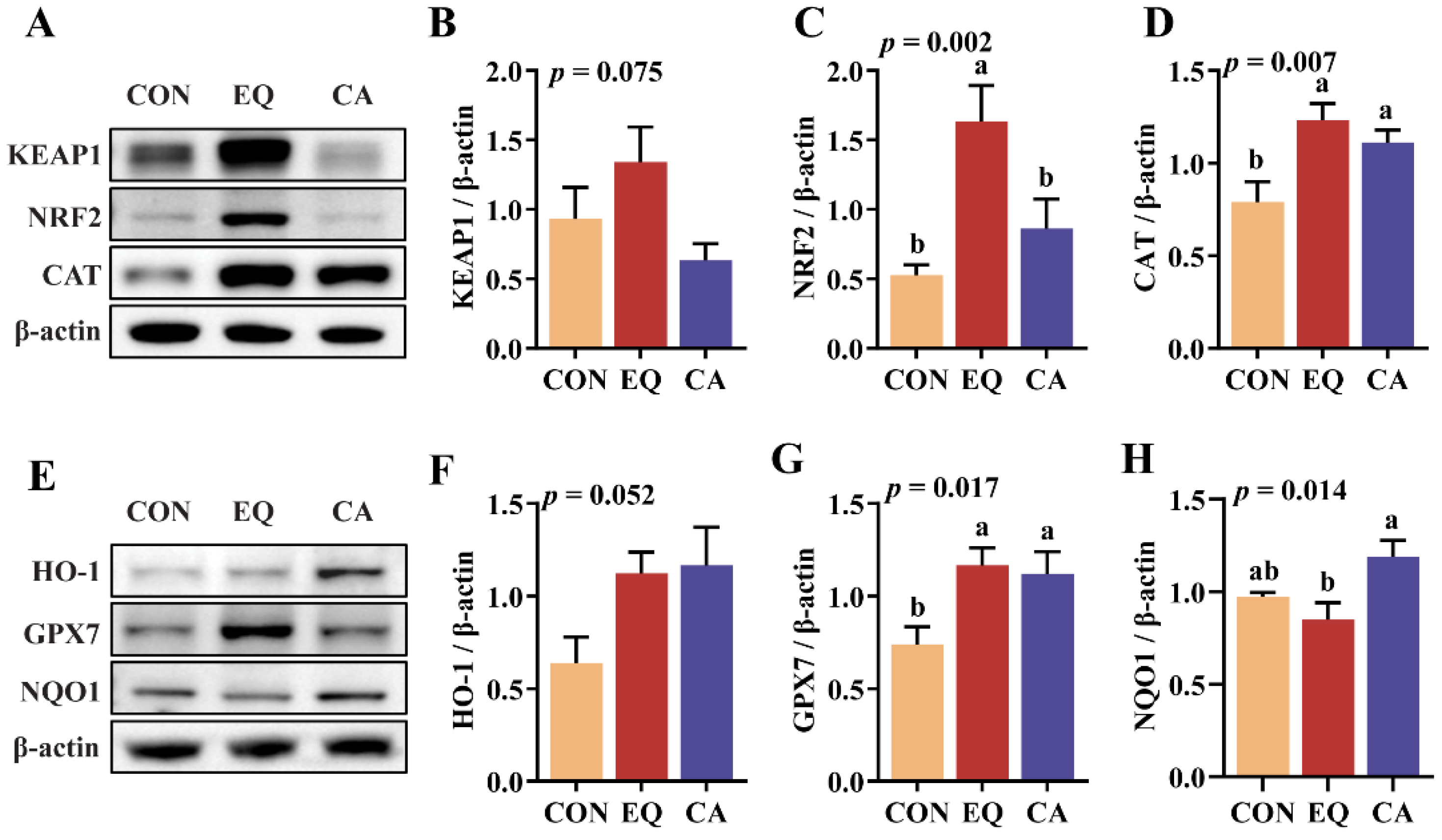
Figure 6.
Expression of inflammatory cytokines at gene level and protein level. N = 8. (A) Relative mRNA expression of IL-1β, IL-18, IL-6, IFN-γ, and TNF-α in jejunum. (B-F) Relative protein levels of NF-κB p50, NF-κB p105, IκB, and TLR4 in jejunum. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. ab Different superscripts letters indicate significant differences (p < 0.05).
Figure 6.
Expression of inflammatory cytokines at gene level and protein level. N = 8. (A) Relative mRNA expression of IL-1β, IL-18, IL-6, IFN-γ, and TNF-α in jejunum. (B-F) Relative protein levels of NF-κB p50, NF-κB p105, IκB, and TLR4 in jejunum. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. ab Different superscripts letters indicate significant differences (p < 0.05).
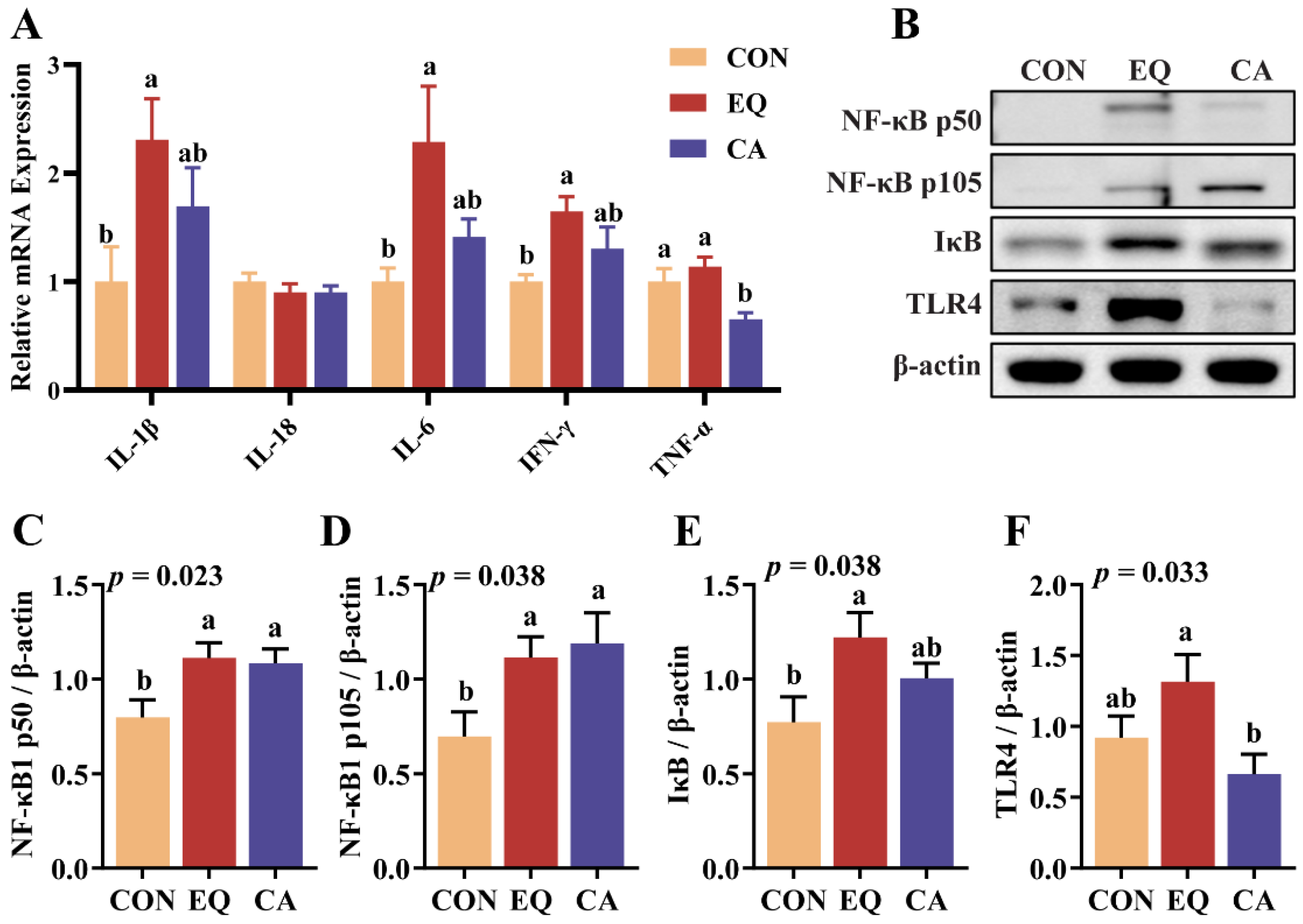
Figure 7.
Changes of microbial diversity in the cecum of broilers. N = 6. (A) Venn diagram of the operational taxonomic units (OTUs) among all groups. (B) Bacterial richness (Chao1 and Observed species) and diversity (Shannon and Simpson) were evaluated by Kruskal-Wallis rank sum test and Dunn’s test. (C) The rarefaction curve shows the chao1 index of each group under the same sampling depth. (D) Principal coordinate analysis (PCoA) was conducted at the ASV level. (E) Non-metric multidimensional scaling (NMDS) analysis was conducted at the ASV level. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant.
Figure 7.
Changes of microbial diversity in the cecum of broilers. N = 6. (A) Venn diagram of the operational taxonomic units (OTUs) among all groups. (B) Bacterial richness (Chao1 and Observed species) and diversity (Shannon and Simpson) were evaluated by Kruskal-Wallis rank sum test and Dunn’s test. (C) The rarefaction curve shows the chao1 index of each group under the same sampling depth. (D) Principal coordinate analysis (PCoA) was conducted at the ASV level. (E) Non-metric multidimensional scaling (NMDS) analysis was conducted at the ASV level. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant.
Figure 8.
Changes of microbial community composition in the cecum of broilers. N = 6. (A) Distribution of cecal microbiota at the phylum level. (B) Heatmap of species relative abundance of cecal microbiota at the phylum level. (C-E) Bacteria with differences in relative abundance in top 10 phylum, including Firmicutes, Bacteroidetes and the ratio of relative abundance of Bacteroidetes and Firmicutes. (F) Distribution of cecal microbiota at the genus level. (G) Heatmap of species relative abundance of cecal microbiota at the genus level. (H-K) Bacteria of bacteria with differences in relative abundance in top 20 genus, including Lactobacillus, Bacteroides, Coprococcus, and Anaeroplasma. The difference of microbial abundance between CON and EQ was analyzed by Wilcoxon rank sum test. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant.
Figure 8.
Changes of microbial community composition in the cecum of broilers. N = 6. (A) Distribution of cecal microbiota at the phylum level. (B) Heatmap of species relative abundance of cecal microbiota at the phylum level. (C-E) Bacteria with differences in relative abundance in top 10 phylum, including Firmicutes, Bacteroidetes and the ratio of relative abundance of Bacteroidetes and Firmicutes. (F) Distribution of cecal microbiota at the genus level. (G) Heatmap of species relative abundance of cecal microbiota at the genus level. (H-K) Bacteria of bacteria with differences in relative abundance in top 20 genus, including Lactobacillus, Bacteroides, Coprococcus, and Anaeroplasma. The difference of microbial abundance between CON and EQ was analyzed by Wilcoxon rank sum test. CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant.
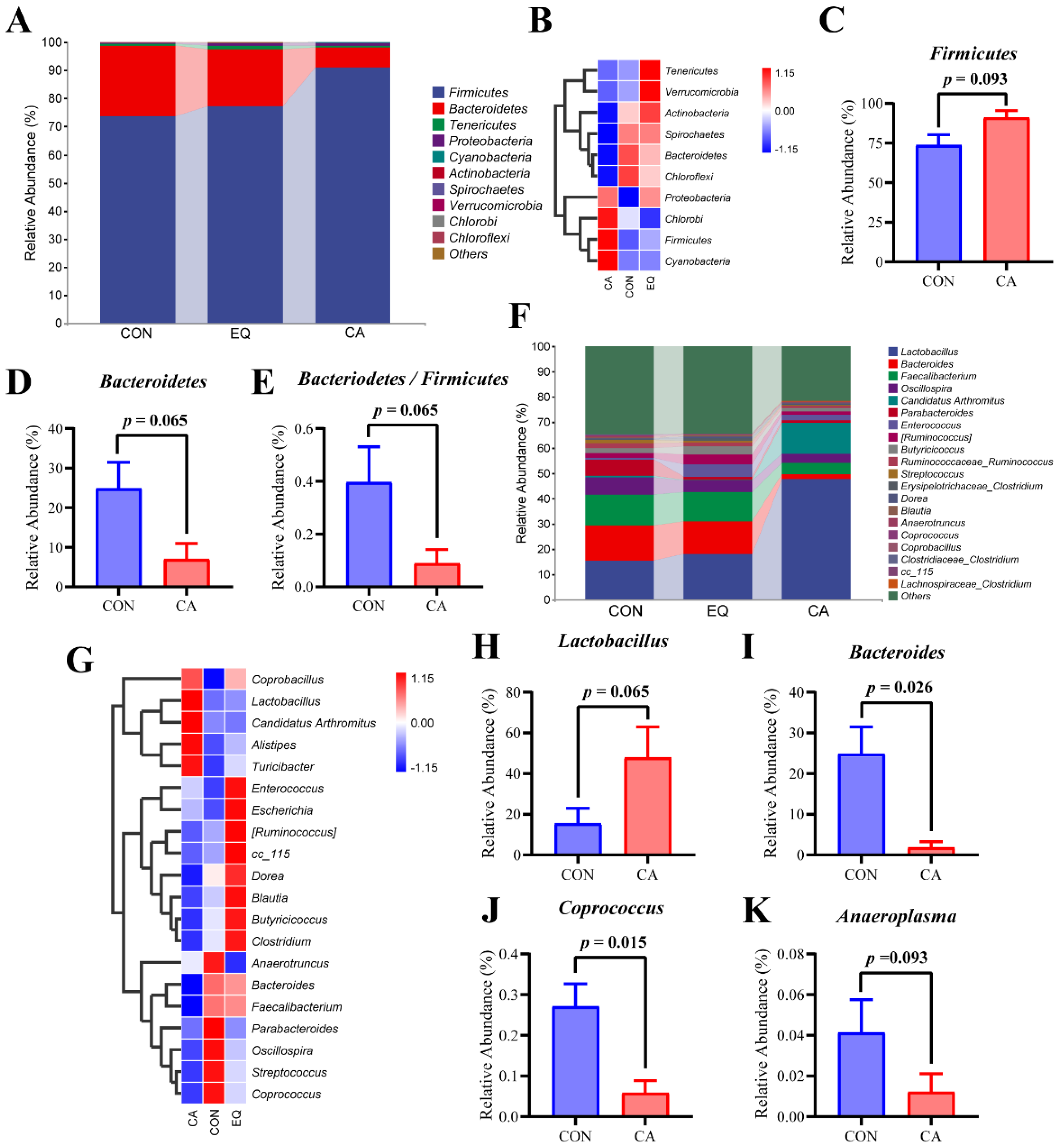
Figure 9.
Graphic summary of the effects of CA on feed antioxidant protection and broiler health. Compound Antioxidant: including 18% butylated hydroxytoluene, 3% citric acid, and 1% tertiary butylhydroquinone.
Figure 9.
Graphic summary of the effects of CA on feed antioxidant protection and broiler health. Compound Antioxidant: including 18% butylated hydroxytoluene, 3% citric acid, and 1% tertiary butylhydroquinone.
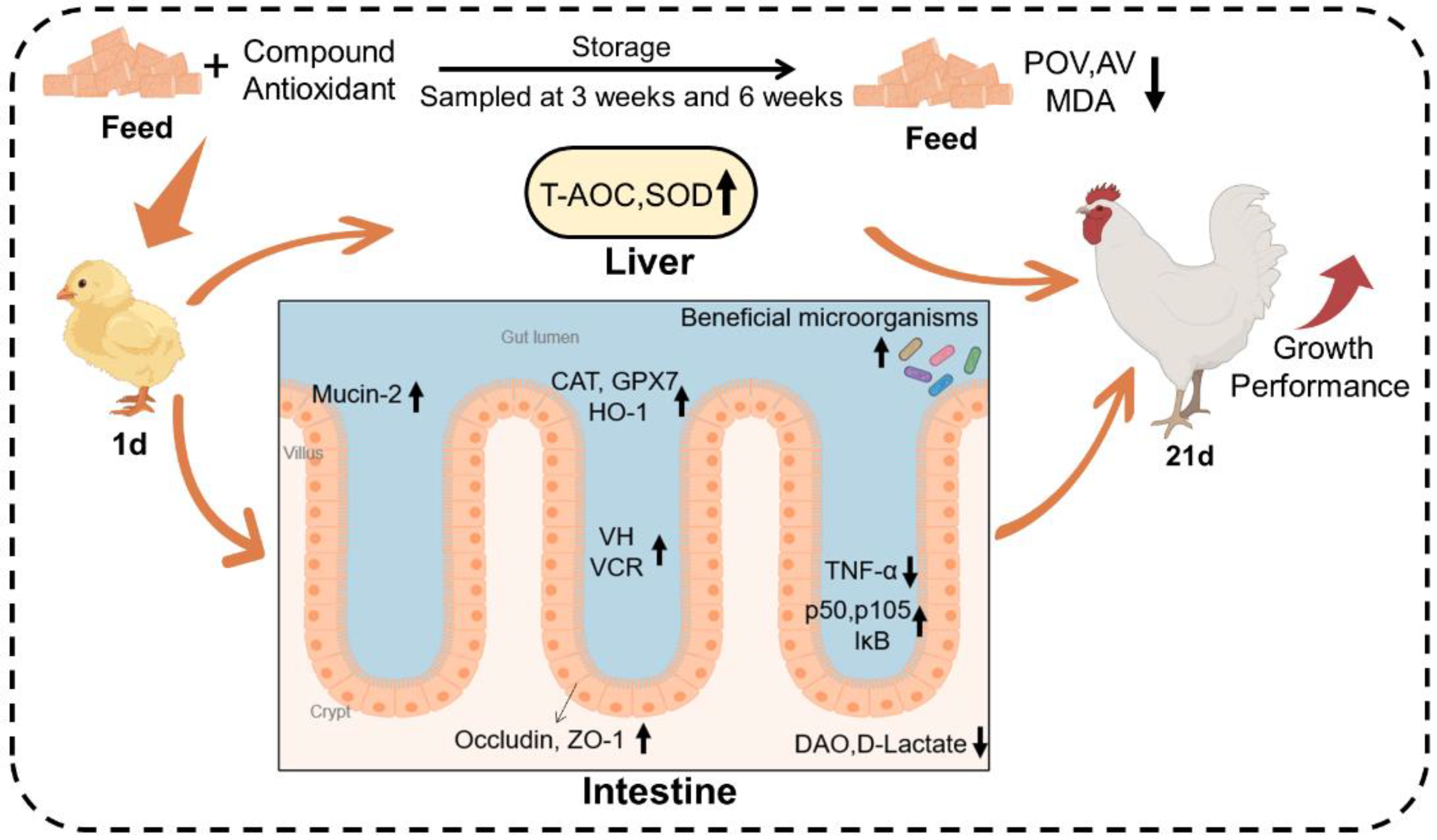

Table 1.
Ingredient and nutrient composition of the basal diet.
| Ingredients | % | Nutrient parameters | Nutrient level |
|---|---|---|---|
| Corn | 52.35 | Metabolic energy, Kcal/kg | 2950 |
| Soybean meal | 38.71 | Crude protein, % | 22.0 |
| Corn gluten meal | 1.31 | Lys, % | 1.31 |
| Soybean oil | 3.06 | Met, % | 0.48 |
| Dicalcium phosphate | 1.79 | Met + Cys, % | 0.96 |
| Limestone | 1.3 | Thr, % | 0.86 |
| Sodium chloride | 0.35 | Val, % | 0.89 |
| Choline chloride (50%) | 0.2 | Calcium, % | 1.02 |
| L-Lysine Hydrochloride (98.5%) | 0.26 | Non-phytate phosphorus, % | 0.39 |
| DL-Methionine (98%) | 0.31 | ||
| L-Threonine (98.5%) | 0.05 | ||
| L-Isoleucine (90%) | 0.03 | ||
| L-Arginine (98%) | 0.02 | ||
| Phytase (10000U/g) | 0.03 | ||
| Mineral premix1 | 0.2 | ||
| Vitamin premix2 | 0.03 | ||
| Total | 100 |
1 The mineral premix provided the following per kg of diets: Cu, 16 mg; Zn, 110 mg; Fe, 80 mg; Mn, 120 mg; Se, 0.30 mg; I, 1.50 mg. 2 The vitamin premix provided the following per kg of diets: vitamin A, 15,000 IU, vitamin D3, 3,600 IU; vitamin E, 30 IU; vitamin K3, 3.00 mg; vitamin B2, 9.60 mg; vitamin B12, 0.03 mg; biotin, 0.15 mg; folic acid, 1.50 mg; pantothenic acid, 13.80 mg; nicotinic acid, 45 mg.
Table 2.
Sequences of primers used for the quantitative real-time PCR analysis.
| Gene1 | Primer sequences (5’-3’)2 | Accession number |
|---|---|---|
| β-actin | F: TTGTTGACAATGGCTCCGGT | NM_205518.1 |
| R: TCTGGGCTTCATCACCAACG | ||
| IFN-γ | F: CTCGCAACCTTCACCTCACCATC | NM_205149.1 |
| R: CAGGAACCAGGCACGAGCTTG | ||
| IL-6 | F: GAACGTCGAGTCTCTGTGCTAC | NM_204628 |
| R: CACCATCTGCCGGATCGT | ||
| IL-1β | F: CAGCCTCAGCGAAGAGACCTT | NM_204524 |
| R: ACTGTGGTGTGCTCAGAATCC | ||
| TNF-α | F: CCCCTACCCTGTCCCACAA | NM_204267 |
| R: TGAGTACTGCGGAGGGTTCAT | ||
| IL-18 | F: GTGTGTGCAGTACGGCTTAG | NM_204608.1 |
| R: TCCACTGCCAGATTTCACCT |
1 Interferon γ (IFN-γ), interleukin 6 (IL-6), interleukin 1β (IL-1β), tumor necrosis factor-alpha (TNF-α), interleukin 18 (IL-18). 2 F, forward primer; R, reverse primer.
Table 3.
Intestinal morphology of broilers.
| Item1 | Duodenum | Jejunum | ||||
|---|---|---|---|---|---|---|
| VH | CD | VCR | VH | CD | VCR | |
| CON | 1413.64 | 214.71 | 6.28b | 965.15b | 175.31b | 5.57a |
| EQ | 1253.73 | 211.79 | 5.93b | 752.79c | 165.16b | 4.38b |
| CA | 1366.46 | 204.85 | 7.38a | 1112.82a | 203.73a | 5.07a |
| SEM | 29.265 | 4.838 | 0.159 | 37.593 | 5.567 | 0.147 |
| p-Value | 0.067 | 0.71 | < 0.001 | < 0.001 | 0.008 | 0.001 |
1 CON, basal diet; EQ, basal diet + 200 g/ton ethoxyquin; CA, basal diet + 200 g/ton compound antioxidant. N=9. Abbreviations: VH, villus height; CD, crypt depth; VCR, villus height-to-crypt depth ratio. ab Different superscripts letters indicate significant differences (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



