
Submitted:
13 September 2024
Posted:
16 September 2024
You are already at the latest version
Abstract
Chronic prostatitis has been found to be associated with the genesis of Benign Prostatic Hyper-plasia (BPH) and subsequent development of prostate cancer (PCa). It has been described that human papillomavirus (HPV) tends to infect the prostatic epithelium; however, correlation be-tween HPV infection and prostate pathology remains to be elucidated. Considering that HPV in-fection can affect cellular pathways, we investigated here the putative role of HPV in the expres-sion of selected miRNAs involved in inflammation, proliferation, and oncogenesis in BPH, BPH/prostatitis and PCa clinical samples. Our results demonstrated that 67.2 % of benign lesions and 93.4% of cancer samples were HPV-positive. Interestingly, we found downregulation of miR-34a, miR-143, and miR-145, as well as upregulation of miR-221 in tissue with BPH and BPH/prostatitis. In HPV-positive PCa samples, let-7c, miR-34a, miR-126, miR-221, miR-145, and miR-106a were found to be downregulated, suggesting their involvement in the formation of a pro-oncogenic microenvironment. Consistent with these results, we found that altered expression levels of miR-34a, miR-145, miR-106a, miR-21 and miR-221 showed a correlation with high-risk HPV-positive PCa samples. The differential expression of these miRNAs induced by HPV could be useful for the identification of potential therapeutic biomarkers, opening a new approach for di-agnosis, prognosis, and therapeutic targeting in prostate cancer.
Keywords:
HPV
; prostate cancer
; BPH
; prostatitis
; miRNAs
; inflammation
; proliferation
; apoptosis
; angiogenesis
; metastasis
1. Introduction
Prostatitis, the third most common urological disorder in males, affects 10-14 % of men of all ages and ethnic origins. Prostatitis has been found to be associated with the genesis of Benign Prostatic Hyperplasia (BPH), a condition that represents an important risk for the development and progression of prostate cancer (PCa) [1,2,3,4,5]. BPH prevalence increases after the age of 40 years, from 8% up to 60% at 90 years old [6]. According to the International Agency for Research on Cancer (IARC), in 2020, there were an estimated 1.4 million new cases of PCa worldwide, making it the second most diagnosed cancer in men [7].
Currently, it has been reported that in BPH, there is no direct relationship between the presence of HPV in the prostate and the development of PCa, as indicated by Bergh et al. [8]. Additionally, research by Adami et al. [9] and Effert et al. [10] suggests that there is no direct association between HPV types 16 or 18 and PCa, though a potential link between HPV type 33 and this disease has been observed.
Contrastingly, other research groups have detected HPV infection in prostate tissue samples, as exemplified by the Singh study [11]. They analyzed 95 PCa and 55 BPH samples. HPV infection was found in 41% of the prostate tumor biopsies and 20% in BPH samples. Specifically, 32% were infected with HPV type 16, and 6% with type 18, while only 5% of BPH controls had HPV 16. A significant proportion of HPV-infected cases were in advanced stages III and IV with high Gleason scores. This study suggests that HPV infection may be a cofactor in PCa progression.
Different studies have shown a strong correlation between high-risk human papillomavirus (HR-HPV) infection and the increased risk of PCa development. Despite this, the specific mechanisms through which these infections contribute to inflammatory lesions and the hyperproliferation in the prostate gland remain not fully elucidated [3,11,12]. Tumor development in the prostate is a complex process that involves successive changes in the genome of normal prostate cells until they transform into malignant cells. Chronic inflammation is regarded as the seventh hallmark of carcinogenesis and as a possible trigger for tumor initiation and progression at the cellular and molecular levels [13]. During pro-tumor inflammation, several inflammatory molecules are either activated or suppressed for a prolonged time, leading to undesirable consequences that encourage the growth and enrichment of aggressive tumor phenotypes in the tumor microenvironment [14]. This process stimulates the infiltration of immune cells and the production of chemokines, cytokines, and free radicals, which can damage DNA and activate the androgen receptor (AR) in prostatic cells [15,16,17].
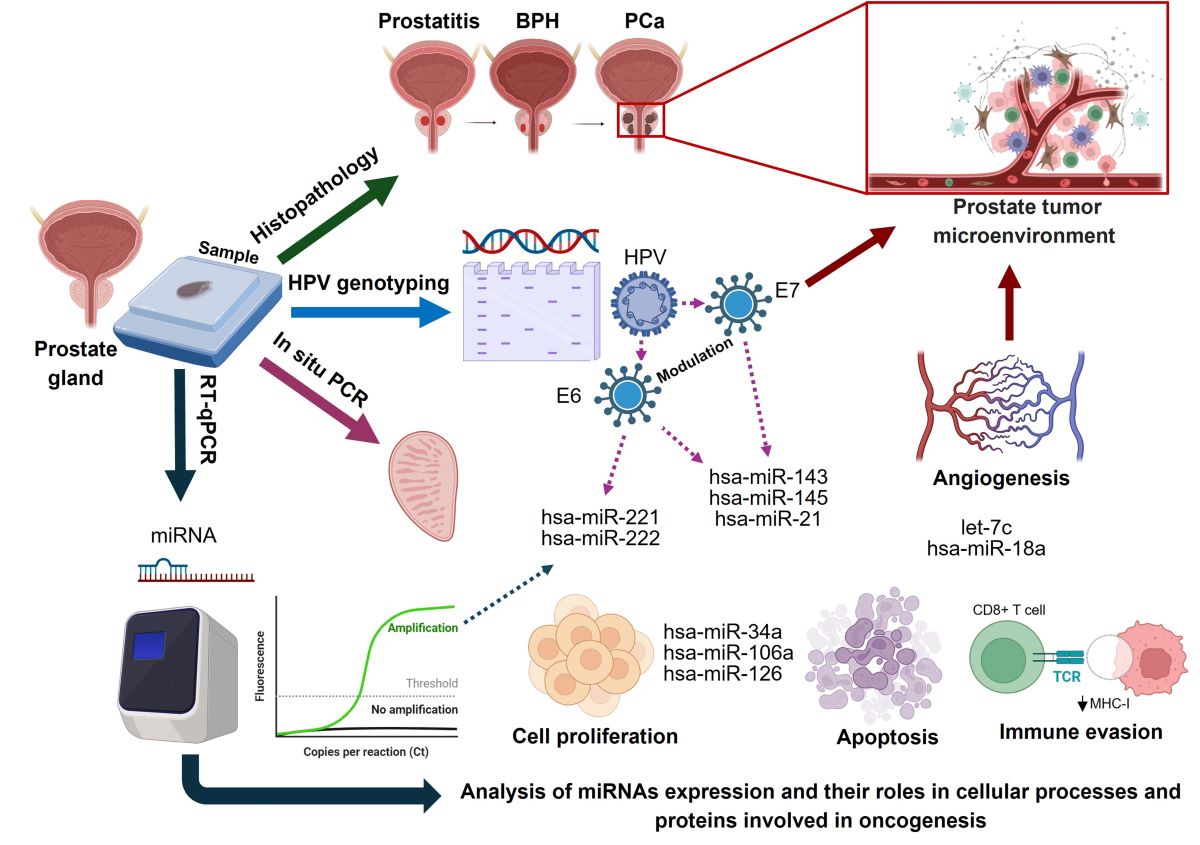
The expression of various micro-RNAs (miRNAs) has been associated with the progression and appearance of different types of cancer, including prostate, breast, lung, liver, gastric, pancreatic, and colorectal, among others [18,19]. Several studies have demonstrated the crucial role of miRNAs in the pathogenesis of cancer. Specifically, miR-34a, miR-106a, miR-143, miR-145, and miR-221 have garnered significant attention in cancer biology. These miRNAs have been implicated in regulating critical processes such as cell proliferation, apoptosis, differentiation, inflammatory response, angiogenesis, tumor suppression, adhesion, metastasis, and immune system evasion [18,19,20,21,22,23].
Particularly, the viral proteins E6 and E7 of HPV play a crucial role in altering genetic regulation in infected cells, specifically affecting miRNAs associated with PCa and tumorigenesis. E6 binds to the p53 protein, a key tumor regulator, promoting its degradation and affecting the expression of miRNAs regulated by p53 [24]. Both E6 and E7 also interfere with the cellular machinery for processing miRNAs, altering their stability and levels. These proteins impact cellular signaling pathways important for cell cycle control and apoptosis, which in turn influences the expression of miRNAs involved in these processes. Additionally, E6 and E7 can modify the transcription of genes, including those encoding miRNAs, either through changes in chromatin or direct interaction with genetic promoters [25,26].
Specifically, in PCa, it has been observed that the presence of HR-HPV leads to the aberrant expression of oncogenic and tumor-suppressive miRNAs. The viral proteins E6 and E7 of HPV play a crucial role in this process, interacting with key transcription factors such as c-Myc, p53, and E2F. This interaction modulates the expression of various miRNAs, including the miR-15/16 cluster, the miR-17-92 family, miR-21, miR-23b, miR-34a, miR-106b/93/25 cluster, miR-143/145 cluster, and let-7c [25,26]. Furthermore, Chiantore et al. [25] demonstrated that HPV E6 and E7 oncoproteins affect the expression of certain tumorigenesis-related miRNAs, including miR-18a, miR-19a, miR-34a, and miR-590-5p. They also identified miR-222, a critical miRNA that is often deregulated in various cancers, present in exosomes from cells infected with the HPV [27].
miRNAs, which originate from a common transcriptional precursor, are subject to the influence of various post-transcriptional factors that determine their final expression. These include precursor processing, efficiency in the generation of mature miRNAs and interactions with regulatory proteins, which are crucial factors for their expression levels. Notably, recent studies have highlighted the importance of transcriptional regulation, especially that mediated by nuclear receptors such as estrogen receptors (ER) and AR, in the modulation of miRNAs in oncological contexts. For instance, it has been observed that the ER interacts with the miR-221/222 gene to inhibit its expression in breast cancers, while in the progression of PCa, miRNAs such as miR-125b, miR-21, and miR-221/222 could be directly regulated by the AR. These findings suggest that in certain prostate pathologies, the predominant influence on the levels of miRNAs interacting with the AR could be of a transcriptional nature, as indicated by these studies [28].
In the Mexican population, there is a lack of documented studies analyzing the expression of miRNAs and their relationship with the development of prostate benign lesions (BPH and prostatitis) and its progression to PCa. Thus, the aim of this study was to analyze the expression profile of selected miRNAs in HR-HPV-positive patients with BPH/prostatitis and PCa. Our findings suggest that alterations in the expression levels of miR-34a, miR-145, miR-106a, miR-21, and miR-221 correlated with HPV infection, particularly in PCa. These results suggest that HPV infection can disrupt the expression of miRNAs, affecting the critical processes that maintain prostate cellular homeostasis.
2. Results
2.1. Histopathological Analysis of Pre-Malignant Lesions and Prostate Cancer
In this study, 14 control samples, 50 samples with premalignant lesions (13 with BPH, and 37 with BPH/prostatitis) and 33 PCa samples were included and stratified based on the Gleason scale, 21 samples were classified as high degree of malignancy (Gleason 8-10); 3 samples as intermediate degree of malignancy (Gleason 7) and 9 samples as Low degree of malignancy (Gleason ≤ 6) (Table 1 and Supplementary Table S1).
Representative histologic images of histopathologic samples are shown in Figure 1. The examination of the control group samples revealed prominent acinar lumens and well-organized epithelial acini, supported by stromal elements, showing a high degree of cellular differentiation and organization. BPH lesions were primarily located in the transitional and peripheral zone of the prostate gland; an increase in epithelial growth with enlarged nuclei restricted to the basal layer is observed; glandular sizes oscillate from medium to large acini, often showing luminal papillae. Furthermore, a decrease in the acinar lumen can be observed due to the hyperproliferation of luminal cells. This hyperproliferation leads to partial loss of cellular homogeneity and reduction of the stroma, although cells are still differentiated. In the BPH/prostatitis group, lesions were identified mainly in the transitional and peripheral zone of the prostate, characterized besides BPH alterations, by multifocal infiltrates of mononuclear cells (lymphocytes, monocytes, and plasma cells). Inflammation was found at the periglandular level, presenting a mononuclear leukocyte infiltrate adjacent to the prostatic acini with a decrease in the intraepithelial component. In some samples, stromal inflammation was found to coexist with the periglandular inflammation (Figure 1a). Important tissue structures such as acinar lumen, luminal cells, stroma, and cellular organization are affected due to cellular hyperproliferation and inflammation. Regarding PCa, low Gleason grade tumors were characterized by the formation of pseudoacinar structures in which the acinar lumen is reduced by hyperproliferation of luminal cells. Such proliferation leads to partial loss of cell homogeneity and stroma reduction, although the cells were still differentiated. Intermediate Gleason grade PCa adenocarcinomas showed the loss of the acinar lumen and the reduction of stroma due to increased cell proliferation. Finally, PCa high Gleason grade tumors completely lost their own structures, and hyperproliferation of luminal cells was observed. Basal cells, stroma, and cellular homogeneity were lost. The acinar lumen is almost null, and the cells are poorly differentiated (Figure 1b).
2.2. Histological Identification of Koilocytes and In Situ Molecular Detection of HPV Sequences
Histopathological analyzes were conducted to identify the presence of koilocytes or pseudokoilocyte-like structures, which are recognized as a pathognomonic morphological feature of HPV infection. A representative image of koilocyte-like morphological structures (pseudokoilocytes) is displayed in Figure 2a. In situ PCR for E6/E7 HPV DNA demonstrated specific detection of koilocytes in those BPH samples that were positive for any HPV genotype, while HPV-negative samples did not show such structures (Figure 2b). In situ semi-quantification of the viral presence, as illustrated in Figure 2c, revealed a heterogeneous positivity level.
2.3. Multiplex Amplification
PCR was performed to determine the genotype in each sample. The results shown that 67.2% of the samples with premalignant lesions were positive for any HPV genotype, in contrast to the 93.4 % positivity in the PCa samples (Table 2). In HPV-positive premalignant lesions, the predominant genotypes were the low-risk genotypes 6 and 11, either alone or in coinfection with intermediate-risk genotypes, while high-risk genotypes were poorly represented. Results also showed that 35.49 % of HPV-positive samples were diagnosed as high Gleason grade cancer (Gleason 8-10) and 48.38 % were intermediate Gleason grade PCa (≤7). Of the HPV-positive samples, 48.38 % were positive for one or more intermediate-risk HPV genotypes (IR-HPV), including 33, 31, 52, and 58 genotypes. Furthermore, HR-HPV 16 and 18 genotypes were identified, being the HPV 16 type the most frequent (Table 2). The analysis of BPH and BPH/prostatitis samples revealed a higher frequency of LR-HPVs instead of oncogenic genotypes. HR-HPVs were mainly present in high Gleason grade PCa specimens, but also in some low Gleason grade PCa specimens, in which genotypes 31 and 52 were the most common. Interestingly, HR-HPV genotypes were observed in all three PCa groups.
2.4. Expression Levels of miRNA in BPH and BPH/Prostatitis Samples
To identify specific molecular mechanisms that may be altered in preneoplastic lesions and CaP, we conducted a study to evaluate the expression levels of six miRNAs by RT-qPCR, including miR-34a, miR-106a, miR-143, miR-145, miR-221 and miR-222. These miRNAs were evaluated in HPV-negative or HPV-positive benign prostatic hyperplasia (BPH, Figure 3a) or in BPH/prostatitis samples (Figure 3b). Interestingly, our findings showed a significant increase in the expression levels of miR-221, as well as a decrease in the expression of miR-145 and miR-34a in HPV-positive samples, in both histopathological conditions. Furthermore, we observed down-regulation of miR-143 in HPV-positive samples from patients with BPH/prostatitis.
On the other hand, in HPV-positive PCa samples, we found a significant decrease in the expression levels of let-7c, miR-34a, miR-126, miR-221, miR-145, and miR-106a compared to control group prostate tissue. In contrast, no significant differences in the expression levels of miR-21 and miR-18a were observed (Figure 4). Due to the low frequency of HPV-negative PCa samples (6.1%), a comparative analysis of miRNA expression levels could not be performed in this patient group.
In addition, we evaluated the relative expression of miRNAs in HPV-positive PCa samples classified as high-risk, intermediate-risk and low risk, comparing them with the expression of HPV-negative PCa samples. Expression levels of miR-34a, miR-145 and miR-106a were significantly down-regulated in samples with high-risk genotypes (HR-HPV) compared to its expression in HPV-negative PCa samples. In contrast, miR-21 and miR-221 were up-regulated by the presence of HR-HPV, whereas miRNAs let7c, miR-126, and miR-18a, showed no significant changes in their expression levels (Figure 5). The statistical difference found in the expression of the 5 miRNAs in the HR-HPV group compared to HPV-negative supports the fact that the expression of viral oncoproteins may be involved in the dysregulation of the analyzed miRNAs expression, as well as in prostate carcinogenesis.
3. Discussion
Currently, it is understood that in the process of tumorigenesis of PCa, HPV modulates through its E6 and E7 oncoproteins to intricately modulate the expression of specific miRNAs. This modulation plays a crucial role in the molecular pathways associated with oncogenic processes and cancer development [27,28,29]. Thus, the aim of our investigation was to explore the potential correlation between specific miRNAs associated with cell proliferation, inflammation, cellular malignancy, and the HPV presence in BPH, BPH/prostatitis, and PCa. In this study, the molecular analysis of HPV in PCa samples revealed a significant relationship between HPV infection and prostatic carcinogenesis, particularly with intermediate- and high-risk genotypes. We found that samples with a Gleason score of ≥8 were highly correlated with the presence of the virus. Furthermore, the most frequent HR- and IR-HPV genotypes associated with PCa were 16, 31, and 52; interestingly, LR-HPV genotype 6 was also found in coinfection with IR-HPV or HR-HPV. Conversely, LR-HPV was more frequently detected in benign lesions (BPH alone or with prostatitis) compared to HR-HPV and IR-HPV genotypes. Our results suggest that premalignant and inflammatory lesions increase the risk of developing PCa.
According to Singh et al. [11] , HPV-16 is the most prevalent genotype found in PCa samples. However, they also identified genotypes 31 and 52. HR-HPV genotypes can lead to malignant tumors even in low viral concentrations. Nonetheless, HPV-16 can reach viral loads much higher than other high-risk genotypes, correlating with the severity of the disease in cervical cancer, as indicated by Swan et al. [30]. Thus, we believe this may explain why we found HPV-16 to be more common than HPV-18 in PCa samples. While it is well-known that LR-HPV has no oncogenic effect, its role as a promoter of persistent inflammation is reinforced by its presence in premalignant lesions such as BPH and BPH/prostatitis, especially in coinfection with IR-HPV and HR-HPV genotypes. This suggests that chronic inflammation may contribute to the initiation and progression of prostate cancer. Chronic inflammation is associated with higher rates of cellular mutations and genetic alterations, which could drive tumorigenesis [14]. Epidemiological data indicate that chronic infections and inflammation are linked to over 25% of all cancers [31], a finding consistent with the observations made by García-Lozano et al. [32] in cervical cancer.
In this study, we aimed to investigate the expression levels of specific miRNAs in premalignant inflammatory lesions and their association with the oncogenic process, particularly in relation to HPV presence. Our analysis revealed significant statistical differences (p<0.05) in the expression levels of let-7c, miR-34a, miR-221, miR-145, miR-106a, and miR-126 in comparison to control prostatic tissue. Among them, miR-221, miR-145, miR-126, and miR-106a showed the most significant statistical difference (p<0.001).
Of particular interest is the miRNA let-7c, which acts as a tumor suppressor in several cancers, including PCa [19]. let-7c targets genes such as NRAS, HMGA2, CCND2, c-Myc, HMGA1, AR, and IL6. Moreover, let-7c regulates other genes, such as TRAIL, Caspase 3, and Lin28b, which are associated with apoptosis, the Epithelial–mesenchymal transition (EMT) process, and angiogenesis. The inhibited expression of let-7c in PCa cells leads to overexpression of the androgen receptor (AR), c-Myc, and HMGA2, which are associated with cell proliferation, migration, invasion, and the EMT process [33,34,35,36,37,38]. It is noteworthy that let-7c and let-7a are considered particularly under-expressed in cervical cancer in comparison with other miRNAs, indicating their association with the presence of HPV in carcinogenic tissue [39,40,41,42].
We also focused on the role of miR-34a as a tumor suppressor that regulates cell differentiation, cell cycle, and apoptosis, as established by previous studies [43,44,45,46]. Our findings suggest that miRNA-34a expression is downregulated in PCa, resulting in the overexpression of several target genes associated with proliferation, motility, immune system evasion, and cellular senescence, including CDK6, NOTCH-1, MYCN, WNT proteins, BCL2, SIRT1, BIRC5, CD44, NANOG, SOX2, SNAI1, MET, AXIN2, PD-L1, DGKζ, and E2F3 [44]. Furthermore, our research indicates that miR-34a inhibits the transcription factor E2F3, which regulates survivin protein, thereby inhibiting apoptosis in samples with cervical cancer [19,43]. Although there is no scientific evidence that correlates E2F3 and survivin levels with HPV presence in PCa, our study suggests that the low levels of miR-34a and HPV presence in our samples could be associated with high levels of E2F3 and survivin, inhibiting apoptosis of cells and promoting their survival. According to previous studies, miR-34a can inhibit SIRT1 and CD44, which are associated with cell migration, cancer progression, and inflammatory cellular events [43]. Our findings suggest that low levels of miR-34a could keep the inflammatory events of the cell, promoting the development of gland transformation through the overexpression of targets like CD44. Additionally, the loss of expression of this miRNA has been strongly related to the overexpression of AR and Notch-1, as well as high-grade lesions [47]. It is worth noting that miR-34a expression depends on p53, and in cancer cells, p53 tends to be lower than in normal cells, as well as miR-34a. In addition, HR-HPV E6 and E7 oncoproteins tend to inhibit miR-34a expression, contributing to the overexpression of its target molecules [48,49]. Likewise, miR-21 plays a crucial role in the progression of PCa by regulating apoptosis, promoting cell proliferation, facilitating invasion and metastasis, potentially contributing to therapeutic resistance, and influencing key signaling pathways. Furthermore, miR-21 can alter the tumor microenvironment and the immune response by modulating the expression of cytokines and inflammatory factors [50]. Interestingly, in our study, we observed a decrease in miR-34a levels and an increase in miR-21 levels among high Gleason grade PCa patients. These findings align with the research conducted by Khatami et al. [39], who examined tissue samples from 112 PCa patients and 39 controls to detect HPV and evaluate miRNA expression profiles. Remarkably, they reported a significant decrease in miR-34a levels and an increase in miR-21 levels in PCa patients compared to those in control tissue. These observations suggest that miR-34a may play a role in the development and progression of prostate cancer.
Additionally, Stafford et al. [51] discovered a significant association between elevated miR-21 expression and advanced stages of PCa, as determined by the Gleason scale. This strongly suggests that elevated miR-21 expression could serve as a valuable biomarker for predicting prostate cancer prognosis.
The downregulation of miR-126 has been shown to be associated with various cellular processes such as apoptosis, proliferation, cell migration, adhesion, and metastasis by targeting SIRT1, CDK6, PIK3R2, E-cadherin, ADAM9, and integrins. Additionally, miR-126 targets the inflammatory molecule HMGB1, and its under-expression has been linked to increased inflammatory processes in tissues [52,53,54,55]. This indicates that the altered expression of miRNAs, such as miR-34a and miR-126, may play a role in the carcinogenic process of prostate cells by linking inflammatory processes to malignant lesions.
On the other hand, miR-221 has been found to inhibit the SOCS3 and IRF2 genes, which suppress the JAK/STAT pathway that is important in the IFN-II-mediated antiviral response [56,57]. Our investigation revealed a diminished expression of miR-221, suggesting that the suppression of the JAK/STAT pathway inhibits the antiviral HPV response [56,58]. Several studies have reported that the downregulation of miR-221 in PCa could activate the TGFβ pathway, which promotes the development and progression of cancer [59]. Moreover, the downregulation of miR-221 has been associated with the overexpression of AR, leading to the loss of the ability of the cells to regulate carcinogenic processes [59]. However, normal expression levels of miR-221 have been found to inhibit IRF2 and SOCS3, suppressing in turn the cell proliferation in androgen-independent PCa tumors, suggesting that miR-221 may be relevant in the control of cell growth [60].
According to Cui et al. [61], miR-145 is a tumor suppressor that plays a vital role in regulating various cellular processes such as cell cycle, proliferation, apoptosis, and cell invasion. Furthermore, under-expression of miR-145 is correlated with the transmembrane protease serine 2-ERG fusion (TMPRSS2-ERG) protein in PCa cells and has been shown to regulate the c-Myc gene that is involved in PCa progression [62]. Under normal conditions, miR-145 participates in anti-inflammatory events through its inhibitory effect on CD40, IL-6, CXCL8, and SMAD3 [63]. Moreover, the oncoproteins E6 and E7 of HR-HPVs facilitate the sub-expression of miR-145 and BRCA1 through the union between the E2F1 factor and Myc in the promoter region of BRCA. Furthermore, Gunasekharan and Laimins [64] reported that HPV-31 can control the expression of miR-145 to maintain its replicative cycle within the cell and suggested that the seed region of miR-145 has a strong affinity for HR-HPV genotypes, ensuring maintenance of infection in epithelial tissue.
In oropharyngeal carcinoma, it was reported that the subexpression of miR-199b, miR-143, miR-145, and miR-126 was due to the presence of HPV, identifying that miR-145 has a specific affinity with the ORF region of the E1 gene [42]. miR-145 is essential in the negative regulation of the replicative cycle of HPV, so the low expression levels of this miRNA are crucial in the tumor formation associated with HPV [42]. Furthermore, our results suggest that the normal expression of miR-145 has an anti-inflammatory effect in prostate cells. However, its subexpression in inflammatory lesions (BPH or BPH/prostatitis) could contribute to the transformation of cancer cells along with the deregulated expression of the miRNAs mentioned above.
Shen et al. [65] found that the under-expression of miR-106a is related to the overexpression of IL-8 in PCa cells. IL-8 is a molecule that promotes prostate tumorigenesis and is a direct target of miR-106a. IL-8 is also known as a primary inflammatory cytokine that facilitates cellular adhesion, neutrophil, and monocyte migration, as well as chronic inflammatory processes. The oncoprotein E6 of HPV positively regulates the expression of IL-8, which is associated with MMP-2 and MMP-9 metalloproteases [66]. If HR-HPVs increase IL-8, our findings about the HPV presence in PCa samples could explain the downregulation of miR-106a. In summary, the deregulation of miR-34a, miR-221, miR-145, and miR-106a in both chronic inflammatory lesions and PCa positive for HPV strongly suggests their participation in the development of the pro-oncogenic microenvironment.
In summary, in Figure 6 we propose a possible mechanism of miRNA expression and their role in viral protein modulation and malignant prostate transformation. According to the results, the deregulation of miR-34a, miR-221, miR-145, and miR-106a in chronic inflammatory lesions in samples of BPH/prostatitis and HPV-positive prostate cancer clearly suggests their participation in the pro-oncogenic microenvironment through the alteration of various molecular pathways.
Ongoing studies will elucidate whether these miRNAs are potential candidates as biomarkers for prognosis, diagnosis, or therapeutic targets in HPV-associated prostate cancer. These investigations are essential for establishing their clinical relevance and potentially paving the way for novel therapeutic strategies in the treatment of this disease.
4. Materials and Methods
4.1. Biological Samples
A retrospective analysis was conducted on prostatic tissue samples embedded in paraffin. The study included 50 samples with a positive histopathological diagnosis of benign prostatic lesions, 13 with BPH alone and 37 with BPH with prostatitis foci, and 33 samples diagnosed with PCa. In this study, 14 control samples were included: six samples obtained from autopsies of individuals who died in accidents, and eight BPH tissue samples. All these samples were negative for HPV.
4.2. Histopathological Analyses and DNA Extraction
For the processing of samples and hematoxylin-eosin staining, we made slight modifications to the protocols described by Pérez et al. [67] and Martínez et al. [68]. Formalin-fixed paraffin-embedded prostate tissue biopsies were processed using a mechanical tissue processor (Leica Instruments, Nussloch, BW, Germany). The samples were then incubated in xylene and subjected to a series of alcohol washes. The specimens were embedded in paraffin, sectioned at 5 micrometers (µm), and stained with hematoxylin-eosin. Two experienced pathologists examined the slides to confirm the diagnosis of each pathological entity and to determine the severity level of the PCa group using the Gleason scale.
DNA was extracted from the paraffin-embedded tissues using the deparaffinization technique of Johann et al. [69] and the DNeasy Blood & Tissue kit (QIAGEN, Germantown, MD, USA) according to the manufacturer’s recommendations. The extracted DNAs were quantified at 260 nm using a spectrophotometer (Epoch™, BioTek®) (BioTek Instruments, Winooski, VT, USA). Samples with DNA purity of 1.4-1.8 and DNA concentration of ≥100 ng/μl were used for HPV detection. The DNA quality was determined by amplifying the human β-globin gene (approximately 268 base pairs; bp) using the primers GH20 (5’-GAAGAGCCAAGGACAGGTAC-3’) and PC04 (5’-CAACTTCATCCACGTTCACC-3’). β-globin was used as an endogenous control to assess the integrity of DNA extraction from the samples. The amplification protocol was performed according to Zandnia, et al. and Williamson et al. [70,71], which consisted of an initial 5 min denaturation step at 95 °C, followed by 30 amplification cycles. Each cycle consisted of 30 s at 95 °C for denaturation, 30 s at 62 °C for annealing, and 30 s at 72 °C for extension. A final extension step of 5 min at 72 °C was applied to complete the amplification. The amplified products were analyzed by electrophoresis in 30% polyacrylamide gels. Only β-globin positive samples were selected.
4.3. HPV Detection by Multiplex PCR
HPV detection was carried out using the GP5+/6+ pair of degenerate oligonucleotides that amplify a 150 bp sequence within the L1 region of the HPV genome. The touchdown protocol described by Evans, et al. [72] was followed. To determine the genotype of HPV, the multiplex PCR (mPCR) Human Papilloma Virus Set 2 kit (Maxim Biotech, Rockville, MD, USA) was employed according to the manufacturer’s instructions. This kit was designed specifically for the simultaneous detection of the 8 most prevalent HPV genotypes in the population; 6, 11 (low-risk); 31, 33, 52, 58 (intermediate-risk); and 16, 18 (high-risk). PCR products were separated by electrophoresis using 12% polyacrylamide gels and analyzed after staining with ethidium bromide (Bio-Rad, Hercules, CA, USA).
4.4. In Situ PCR
To identify the High Risk-HPV (HR-HPV) E6/E7 viral genes, a direct in situ PCR was performed using specific primers, as previously described in the literature [73]. The procedure involved incubation of dried dewaxed sections on DNase/RNase-free electrocharged slides with Proteinase K, followed by washing with ultrapure water. PCR optimal solution containing digoxigenin-11-(2’-deoxy-uridine-5’)-triphosphate, DIG-11-dUTP, (Roche, Indianapolis, IN, USA) was used. In situ PCR was carried out using the system provided by Perkin Elmer, employing a hot start method and two consensus sequence primer pairs within E6 and E7 of HR-HPV (pU-1M and pU-2R primers) [74], and 5 U of Taq DNA polymerase. The cycling conditions included 2 min at 94 °C, followed by 18 cycles of 94 °C for 1 min, 60 °C for 1 min, and 72 °C for 1 min. Clips and AmpliCover discs were then removed, and slides were washed in PBS, followed by 5 min in 100% EtOH, before being air-dried.
4.5. Sequence Detection of In Situ PCR Products
To detect the PCR product, an indirect immunolabelling method using a primary anti-digoxigenin antibody (Roche, Indianapolis, IN, USA) conjugated to alkaline phosphatase was employed. Prior to labelling, a blocking step was carried out using 5% BSA (Sigma, St. Louis, MO, USA) in PBS for 30 min. The slides were then drained, and an anti-DIG antibody was added at a dilution of 1:200 in 100 mM Tris HCl pH 7.4 and 150 mM NaCl and incubated for 2 h at room temperature. Detection of alkaline phosphatase was achieved by using the NBT/BCIP kit (Roche, Indianapolis, IN, USA) for 10 min. After detection, the slides were rinsed in distilled water for 5 min and counterstained with Fast Green. Finally, the slides were air dried before being mounted in Permount histological mounting medium (Fisher Scientific, Waltham, MA, USA).
4.6. Digital Analysis and Relative Semi-Quantification of In Situ Positive Signal
A set of low-amplification (5x) images of up to 10 areas of each tissue sample were captured using a DFC290 HD digital camera (Leica Microsystems, Buffalo Grove, IL, USA). Subsequently, the experimental image files were digitally processed to obtain a uniform signal using PhotoImpact (Ulead PhotoImpact SE Version 3.02, Ulead Systems, Torrance, CA, USA). The relevant regions were digitally assessed using the Image-ProPlus Analysis Software (Version 4.5.0.19, Media Cybernetics, Inc., Rockville, MD, USA). All pixels exhibiting positive amplification signal for HPV E6/E7 oncogenes, and falling within the specified threshold parameters, were quantified to produce a graphical representation [73].
4.7. RT-qPCR for miRNAs
miRNAs were extracted from 20 µm thickness dewaxed tissue sections of biological samples using the standard TRIzol method [75] and conserved in DEPC-treated water to prevent degradation. The concentration of extracted RNA was quantified by spectrophotometer (Epoch™, BioTek®) (BioTek Instruments, Winooski, VT, USA) to ensure a purity of 1.7-1.9 (260/280 ratio) and a concentration of ≥50 ng/µl. The cDNA synthesis (RT) of each miRNA was performed using 50 ng of RNA and the TaqMan® Micro-RNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA) according to the manufacturer’s instructions. The RT-qPCR assays were carried out using the TaqMan Micro-RNA Assays with the Stratagene system MX 3000p (Thermo Fisher Scientific, Waltham, MA, USA) and included 40 cycles (denaturation at 95 °C, annealing and extension at 60 °C). We used commercial TaqMan probes (Applied Biosystems, Foster City, CA, USA) for the detection of hsa-miR-34a-5p, hsa-miR-106a-5p, hsa-miR-143-5p, hsa-miR-145-5p, hsa-miR-221-5p, hsa-miR-222-5p, and hsa-miR-34a-5p, hsa-miR-106a-5p, hsa-miR-143-5p, hsa-miR-145-5p, hsa-miR-221-5p, hsa-let-7c-5p, and hsa-miR126-5p. The internal control RNU48 was used to compare the relative expression of selected miRNAs in experimental tissue samples using the 2-ΔΔCt method [76]. All experiments were performed at least in triplicate.
4.8. Statistical Analysis
Graphics were generated using the GraphPad Prism version 8.0.1(GraphPad Software, San Diego, CA, USA) and IBM SPSS Statistics 25 software (IBM Corporation, Armonk, NY, USA), with a 95% confidence interval. The statistical analyses were conducted using one-way ANOVA and multiple comparison tests, including Tukey and Dunnett, as well as multiple T-tests per row. The levels of statistical significance were determined in accordance with the guidelines outlined by the American Psychological Association (APA), with *p≤0.003, **p≤0.002, and ***p≤0.001. The data presented represents the standard deviation obtained from at least three independent experiments. Finally, differences in intragroup, ∆Ct data were compared using boxplots in the JMP statistical software (SAS Institute, Cary, NC, USA).
5. Conclusions
Our study highlights the significance of miR-34a, miR-143, miR-145, and miR-221 in BPH and prostatitis, and let-7c, miR-34a, miR-126, miR-221, miR-145, and miR-106a in HPV-positive prostate cancer. These miRNAs modulate oncogenic pathways and contribute to the development of a pro-oncogenic environment, highlighting their potential as biomarkers for the prognosis, diagnosis, and treatment of HPV-driven prostate conditions.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org.
Author Contributions
Conceptualization, Sandra Salgado-Hernández, Lucero Martínez-Retamoza, María Gómez-García and David Pérez-Ishiwara; Data curation, Sandra Salgado-Hernández, Lucero Martínez-Retamoza, Gladys Cedeño-Arboleda and David Pérez-Ishiwara; Formal analysis, Sandra Salgado-Hernández, Lucero Martínez-Retamoza, Rodolfo Ocádiz-Delgado, Salvador Pérez-Mora, Gladys Cedeño-Arboleda, María Gómez-García and David Pérez-Ishiwara; Funding acquisition, María Gómez-García and David Pérez-Ishiwara; Investigation, Sandra Salgado-Hernández, Lucero Martínez-Retamoza, Rodolfo Ocádiz-Delgado, Salvador Pérez-Mora and David Pérez-Ishiwara; Methodology, Sandra Salgado-Hernández, Lucero Martínez-Retamoza, Rodolfo Ocádiz-Delgado, María Gómez-García, Patricio Gariglio and David Pérez-Ishiwara; Project administration, David Pérez-Ishiwara; Resources, María Gómez-García and David Pérez-Ishiwara; Software, Sandra Salgado-Hernández, Lucero Martínez-Retamoza, Rodolfo Ocádiz-Delgado and Patricio Gariglio; Supervision, Rodolfo Ocádiz-Delgado and David Pérez-Ishiwara; Validation, Sandra Salgado-Hernández, Rodolfo Ocádiz-Delgado, María Gómez-García and David Pérez-Ishiwara; Visualization, Sandra Salgado-Hernández, Rodolfo Ocádiz-Delgado, María Gómez-García and David Pérez-Ishiwara; Writing – original draft, Sandra Salgado-Hernández, Lucero Martínez-Retamoza and Salvador Pérez-Mora; Writing – review & editing, Sandra Salgado-Hernández, Rodolfo Ocádiz-Delgado, Salvador Pérez-Mora and David Pérez-Ishiwara.
Funding
This research was funded by National Polytechnic Institute; 20201289 project and 2023 Innovation Project given to D.G.P.-I.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
To CONAHCYT for their invaluable contribution through the postgraduate scholarship granted to S.V.S.-H., L.M.-R, S.P.-M. and G.C.-A. We would like to thank Biol. Elizabeth Alvarez-Rios and Mr. Lauro Macías (CINVESTAV-IPN, Mexico) for excellent technical support. Special thanks to the “Servicios Médicos Forenses” (SEMEFO), Mexico, for the samples obtained from autopsies.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dai, X.; Fang, X.; Ma, Y.; Xianyu, J. Benign Prostatic Hyperplasia and the Risk of Prostate Cancer and Bladder Cancer: A Meta-Analysis of Observational Studies. Medicine (Baltimore) 2016, 95, e3493. [CrossRef]
- Shen, P.; Sun, G.; Zhao, P.; Dai, J.; Zhang, X.; Zhao, J.; Zhu, S.; Chen, J.; Tao, R.; Yang, J.; et al. MicroRNA-106a Suppresses Prostate Cancer Proliferation, Migration and Invasion by Targeting Tumor-Derived IL-8. Transl. Cancer Res. 2020, 9, 3507–3517. [CrossRef]
- Shiau, M.-Y.; Fan, L.-C.; Yang, S.-C.; Tsao, C.-H.; Lee, H.; Cheng, Y.-W.; Lai, L.-C.; Chang, Y.-H. Human Papillomavirus Up-Regulates MMP-2 and MMP-9 Expression and Activity by Inducing Interleukin-8 in Lung Adenocarcinomas. PLoS ONE 2013, 8, e54423. [CrossRef]
- Johann, D.J.; Shin, I.J.; Roberge, A.; Laun, S.; Peterson, E.A.; Liu, M.; Steliga, M.A.; Muesse, J.; Emmert-Buck, M.R.; Tangrea, M.A. Effect of Antigen Retrieval on Genomic DNA From Immunodissected Samples. J. Histochem. Cytochem. 2022, 70, 643–658. [CrossRef]
- Zhang, L.; Wang, Y.; Qin, Z.; Gao, X.; Xing, Q.; Li, R.; Wang, W.; Song, N.; Zhang, W. Correlation between Prostatitis, Benign Prostatic Hyperplasia and Prostate Cancer: A Systematic Review and Meta-Analysis. J. Cancer 2020, 11, 177–189. [CrossRef]
- Lim, K.B. Epidemiology of Clinical Benign Prostatic Hyperplasia. Asian J. Urol. 2017, 4, 148–151. [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [CrossRef]
- Bergh, J.; Marklund, I.; Gustavsson, C.; Wiklund, F.; Grönberg, H.; Allard, A.; Alexeyev, O.; Elgh, F. No Link between Viral Findings in the Prostate and Subsequent Cancer Development. Br. J. Cancer 2007, 96, 137–139. [CrossRef]
- Adami, H.-O.; Kuper, H.; Andersson, S.-O.; Bergström, R.; Dillner, J. Prostate Cancer Risk and Serologic Evidence of Human Papilloma Virus Infection: A Population-Based Case-Control Study. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2003, 12, 872–875.
- Effert, P.J.; Frye, R.A.; Neubauer, A.; Liu, E.T.; Walther, P.J. Human Papillomavirus Types 16 and 18 Are Not Involved in Human Prostate Carcinogenesis: Analysis of Archival Human Prostate Cancer Specimens by Differential Polymerase Chain Reaction. J. Urol. 1992, 147, 192–196. [CrossRef]
- Singh, N.; Hussain, S.; Kakkar, N.; Singh, S.K.; Sobti, R.C.; Bharadwaj, M. Implication of High Risk Human Papillomavirus HR-HPV Infection in Prostate Cancer in Indian Population- A Pioneering Case-Control Analysis. Sci. Rep. 2015, 5, 7822. [CrossRef]
- Russo, G.I.; Calogero, A.E.; Condorelli, R.A.; Scalia, G.; Morgia, G.; La Vignera, S. Human Papillomavirus and Risk of Prostate Cancer: A Systematic Review and Meta-Analysis. Aging Male 2020, 23, 132–138. [CrossRef]
- Coussens, L.M.; Werb, Z. Inflammation and Cancer. Nature 2002, 420, 860–867. [CrossRef]
- Oseni, S.O.; Naar, C.; Pavlović, M.; Asghar, W.; Hartmann, J.X.; Fields, G.B.; Esiobu, N.; Kumi-Diaka, J. The Molecular Basis and Clinical Consequences of Chronic Inflammation in Prostatic Diseases: Prostatitis, Benign Prostatic Hyperplasia, and Prostate Cancer. Cancers 2023, 15, 3110. [CrossRef]
- Naiyila, X.; Li, J.; Huang, Y.; Chen, B.; Zhu, M.; Li, J.; Chen, Z.; Yang, L.; Ai, J.; Wei, Q.; et al. A Novel Insight into the Immune-Related Interaction of Inflammatory Cytokines in Benign Prostatic Hyperplasia. J. Clin. Med. 2023, 12, 1821. [CrossRef]
- Ullah, A.; Chen, Y.; Singla, R.K.; Cao, D.; Shen, B. Pro-Inflammatory Cytokines and CXC Chemokines as Game-Changer in Age-Associated Prostate Cancer and Ovarian Cancer: Insights from Preclinical and Clinical Studies’ Outcomes. Pharmacol. Res. 2024, 204, 107213. [CrossRef]
- Hatano, K.; Fujita, K.; Nonomura, N. Application of Anti-Inflammatory Agents in Prostate Cancer. J. Clin. Med. 2020, 9, 2680. [CrossRef]
- Coradduzza, D.; Solinas, T.; Balzano, F.; Culeddu, N.; Rossi, N.; Cruciani, S.; Azara, E.; Maioli, M.; Zinellu, A.; De Miglio, M.R.; et al. miRNAs as Molecular Biomarkers for Prostate Cancer. J. Mol. Diagn. 2022, 24, 1171–1180. [CrossRef]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Szyłło, K.; Hogendorf, P. miRNAs in Cancer (Review of Literature). Int. J. Mol. Sci. 2022, 23, 2805. [CrossRef]
- Pekarek, L.; Torres-Carranza, D.; Fraile-Martinez, O.; García-Montero, C.; Pekarek, T.; Saez, M.A.; Rueda-Correa, F.; Pimentel-Martinez, C.; Guijarro, L.G.; Diaz-Pedrero, R.; et al. An Overview of the Role of MicroRNAs on Carcinogenesis: A Focus on Cell Cycle, Angiogenesis and Metastasis. Int. J. Mol. Sci. 2023, 24, 7268. [CrossRef]
- Vanacore, D.; Boccellino, M.; Rossetti, S.; Cavaliere, C.; D’Aniello, C.; Di Franco, R.; Romano, F.J.; Montanari, M.; La Mantia, E.; Piscitelli, R.; et al. Micrornas in Prostate Cancer: An Overview. Oncotarget 2017, 8, 50240–50251. [CrossRef]
- Schitcu, V.H.; Raduly, L.; Nutu, A.; Zanoaga, O.; Ciocan, C.; Munteanu, V.C.; Cojocneanu, R.; Petrut, B.; Coman, I.; Braicu, C.; et al. MicroRNA Dysregulation in Prostate Cancer. Pharmacogenomics Pers. Med. 2022, 15, 177–193. [CrossRef]
- Abudoubari, S.; Bu, K.; Mei, Y.; Maimaitiyiming, A.; An, H.; Tao, N. Preliminary Study on miRNA in Prostate Cancer. World J. Surg. Oncol. 2023, 21, 270. [CrossRef]
- Tomaić, V. Functional Roles of E6 and E7 Oncoproteins in HPV-Induced Malignancies at Diverse Anatomical Sites. Cancers 2016, 8, 95. [CrossRef]
- Zheng, Z.-M.; Wang, X. Regulation of Cellular miRNA Expression by Human Papillomaviruses. Biochim. Biophys. Acta BBA - Gene Regul. Mech. 2011, 1809, 668–677. [CrossRef]
- Yim, E.-K.; Park, J.-S. The Role of HPV E6 and E7 Oncoproteins in HPV-Associated Cervical Carcinogenesis. Cancer Res. Treat. 2005, 37, 319. [CrossRef]
- Chiantore, M.V.; Mangino, G.; Iuliano, M.; Zangrillo, M.S.; De Lillis, I.; Vaccari, G.; Accardi, R.; Tommasino, M.; Columba Cabezas, S.; Federico, M.; et al. Human Papillomavirus E6 and E7 Oncoproteins Affect the Expression of Cancer-Related microRNAs: Additional Evidence in HPV-Induced Tumorigenesis. J. Cancer Res. Clin. Oncol. 2016, 142, 1751–1763. [CrossRef]
- Ali Syeda, Z.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory Mechanism of MicroRNA Expression in Cancer. Int. J. Mol. Sci. 2020, 21, 1723. [CrossRef]
- Boulet, G.; Horvath, C.; Broeck, D.V.; Sahebali, S.; Bogers, J. Human Papillomavirus: E6 and E7 Oncogenes. Int. J. Biochem. Cell Biol. 2007, 39, 2006–2011. [CrossRef]
- Swan, D.C.; Tucker, R.A.; Tortolero-Luna, G.; Mitchell, M.F.; Wideroff, L.; Unger, E.R.; Nisenbaum, R.A.; Reeves, W.C.; Icenogle, J.P. Human Papillomavirus (HPV) DNA Copy Number Is Dependent on Grade of Cervical Disease and HPV Type. J. Clin. Microbiol. 1999, 37, 1030–1034.
- Wu, Y.; Godoy, A.; Azzouni, F.; Wilton, J.H.; Ip, C.; Mohler, J.L. Prostate Cancer Cells Differ in Testosterone Accumulation, Dihydrotestosterone Conversion, and Androgen Receptor Signaling Response to Steroid 5α-Reductase Inhibitors. The Prostate 2013, 73, 1470–1482. [CrossRef]
- García Lozano, T.; García García, E.; González Monsalve, J.A.; Illueca Ballester, C.; Aznar Oroval, E.; San Juan Gadea, M.C.; Navarro Gallego, M.T.; Almenar Medina, S. Análisis de las coinfecciones mixtas por el virus del papiloma humano (VPH) de alto y bajo riesgo en lesiones de significado incierto. Clínica E Investig. En Ginecol. Obstet. 2015, 42, 18–24. [CrossRef]
- Wagner, S.; Ngezahayo, A.; Murua Escobar, H.; Nolte, I. Role of miRNA Let-7 and Its Major Targets in Prostate Cancer. BioMed Res. Int. 2014, 2014, 1–14. [CrossRef]
- Mulholland, E.J.; Green, W.P.; Buckley, N.E.; McCarthy, H.O. Exploring the Potential of MicroRNA Let-7c as a Therapeutic for Prostate Cancer. Mol. Ther. Nucleic Acids 2019, 18, 927–937. [CrossRef]
- Ma, Y.; Shen, N.; Wicha, M.S.; Luo, M. The Roles of the Let-7 Family of MicroRNAs in the Regulation of Cancer Stemness. Cells 2021, 10, 2415. [CrossRef]
- Zhang, W.-T.; Zhang, G.-X.; Gao, S.-S. The Potential Diagnostic Accuracy of Let-7 Family for Cancer: A Meta-Analysis. Technol. Cancer Res. Treat. 2021, 20, 15330338211033061. [CrossRef]
- Dong, Q.; Meng, P.; Wang, T.; Qin, W.; Qin, W.; Wang, F.; Yuan, J.; Chen, Z.; Yang, A.; Wang, H. MicroRNA Let-7a Inhibits Proliferation of Human Prostate Cancer Cells In Vitro and In Vivo by Targeting E2F2 and CCND2. PLoS ONE 2010, 5, e10147. [CrossRef]
- Alwhaibi, A.; Parvathagiri, V.; Verma, A.; Artham, S.; Adil, M.S.; Somanath, P.R. Regulation of Let-7a-5p and miR-199a-5p Expression by Akt1 Modulates Prostate Cancer Epithelial-to-Mesenchymal Transition via the Transforming Growth Factor-β Pathway. Cancers 2022, 14, 1625. [CrossRef]
- Khatami, A.; Nahand, J.S.; Kiani, S.J.; Khoshmirsafa, M.; Moghoofei, M.; Khanaliha, K.; Tavakoli, A.; Emtiazi, N.; Bokharaei-Salim, F. Human Papilloma Virus (HPV) and Prostate Cancer (PCa): The Potential Role of HPV Gene Expression and Selected Cellular MiRNAs in PCa Development. Microb. Pathog. 2022, 166, 105503. [CrossRef]
- Lajer, C.B.; Garnæs, E.; Friis-Hansen, L.; Norrild, B.; Therkildsen, M.H.; Glud, M.; Rossing, M.; Lajer, H.; Svane, D.; Skotte, L.; et al. The Role of miRNAs in Human Papilloma Virus (HPV)-Associated Cancers: Bridging between HPV-Related Head and Neck Cancer and Cervical Cancer. Br. J. Cancer 2012, 106, 1526–1534. [CrossRef]
- Bañuelos-Villegas, E.G.; Pérez-yPérez, M.F.; Alvarez-Salas, L.M. Cervical Cancer, Papillomavirus, and miRNA Dysfunction. Front. Mol. Biosci. 2021, 8, 758337. [CrossRef]
- Miller, D.L.; Davis, J.W.; Taylor, K.H.; Johnson, J.; Shi, Z.; Williams, R.; Atasoy, U.; Lewis, J.S.; Stack, M.S. Identification of a Human Papillomavirus–Associated Oncogenic miRNA Panel in Human Oropharyngeal Squamous Cell Carcinoma Validated by Bioinformatics Analysis of The Cancer Genome Atlas. Am. J. Pathol. 2015, 185, 679–692. [CrossRef]
- DUAN, K.; GE, Y.-C.; ZHANG, X.-P.; WU, S.-Y.; FENG, J.-S.; CHEN, S.-L.; ZHANG, L.; YUAN, Z.-H.; FU, C.-H. miR-34a Inhibits Cell Proliferation in Prostate Cancer by Downregulation of SIRT1 Expression. Oncol. Lett. 2015, 10, 3223–3227. [CrossRef]
- Slabáková, E.; Culig, Z.; Remšík, J.; Souček, K. Correction Alternative Mechanisms of miR-34a Regulation in Cancer. Cell Death Dis. 2018, 9, 783. [CrossRef]
- Slabáková, E.; Culig, Z.; Remšík, J.; Souček, K. Alternative Mechanisms of miR-34a Regulation in Cancer. Cell Death Dis. 2017, 8, e3100–e3100. [CrossRef]
- Li, W. (Jess); Liu, X.; Dougherty, E.M.; Tang, D.G. MicroRNA-34a, Prostate Cancer Stem Cells, and Therapeutic Development. Cancers 2022, 14, 4538. [CrossRef]
- Mekhail, S.M.; Yousef, P.G.; Jackinsky, S.W.; Pasic, M.; Yousef, G.M. miRNA in Prostate Cancer: New Prospects for Old Challenges. EJIFCC 2014, 25, 79–98.
- Li, B.; Guo, X.; Li, N.; Chen, Q.; Shen, J.; Huang, X.; Huang, G.; Wang, F. WNT1, a Target of miR-34a, Promotes Cervical Squamous Cell Carcinoma Proliferation and Invasion by Induction of an E-P Cadherin Switch via the WNT/β-Catenin Pathway. Cell. Oncol. Dordr. 2020, 43, 489–503. [CrossRef]
- Zhang, R.; Su, J.; Xue, S.-L.; Yang, H.; Ju, L.-L.; Ji, Y.; Wu, K.-H.; Zhang, Y.-W.; Zhang, Y.-X.; Hu, J.-F.; et al. HPV E6/P53 Mediated down-Regulation of miR-34a Inhibits Warburg Effect through Targeting LDHA in Cervical Cancer. Am. J. Cancer Res. 2016, 6, 312–320.
- Feng, Y.-H.; Tsao, C.-J. Emerging Role of microRNA-21 in Cancer. Biomed. Rep. 2016, 5, 395–402. [CrossRef]
- Stafford, M.Y.C.; Willoughby, C.E.; Walsh, C.P.; McKenna, D.J. Prognostic Value of miR-21 for Prostate Cancer: A Systematic Review and Meta-Analysis. Biosci. Rep. 2022, 42, BSR20211972. [CrossRef]
- Chen, Q.; Chen, S.; Zhao, J.; Zhou, Y.; Xu, L. MicroRNA-126: A New and Promising Player in Lung Cancer. Oncol. Lett. 2021, 21, 35. [CrossRef]
- Zhou, Y.; Feng, X.; Liu, Y.; Ye, S.; Wang, H.; Tan, W.; Tian, T.; Qiu, Y.; Luo, H. Down-Regulation of miR-126 Is Associated with Colorectal Cancer Cells Proliferation, Migration and Invasion by Targeting IRS-1 via the AKT and ERK1/2 Signaling Pathways. PLoS ONE 2013, 8, e81203. [CrossRef]
- Jalil, A.T.; Abdulhadi, M.A.; Al-Ameer, L.R.; Abbas, H.A.; Merza, Muna.S.; Zabibah, R.S.; Fadhil, A.A. The Emerging Role of microRNA-126 as a Potential Therapeutic Target in Cancer: A Comprehensive Review. Pathol. - Res. Pract. 2023, 248, 154631. [CrossRef]
- Miao, Y.; Lu, J.; Fan, B.; Sun, L. MicroRNA-126-5p Inhibits the Migration of Breast Cancer Cells by Directly Targeting CNOT7. Technol. Cancer Res. Treat. 2020, 19, 153303382097754. [CrossRef]
- Lu, H.; Gu, X. MicroRNA-221 Inhibits Human Papillomavirus 16 E1-E2 Mediated DNA Replication through Activating SOCS1/Type I IFN Signaling Pathway.
- Di Martino, M.T.; Arbitrio, M.; Caracciolo, D.; Cordua, A.; Cuomo, O.; Grillone, K.; Riillo, C.; Caridà, G.; Scionti, F.; Labanca, C.; et al. miR-221/222 as Biomarkers and Targets for Therapeutic Intervention on Cancer and Other Diseases: A Systematic Review. Mol. Ther. Nucleic Acids 2022, 27, 1191–1224. [CrossRef]
- Ye, T.; Zhong, L.; Ye, X.; Liu, J.; Li, L.; Yi, H. miR-221-3p and miR-222-3p Regulate the SOCS3/STAT3 Signaling Pathway to Downregulate the Expression of NIS and Reduce Radiosensitivity in Thyroid Cancer. Exp. Ther. Med. 2021, 21, 652. [CrossRef]
- Mishra, S.; Deng, J.J.; Gowda, P.S.; Rao, M.K.; Lin, C.-L.; Chen, C.L.; Huang, T.; Sun, L.-Z. Androgen Receptor and MicroRNA-21 Axis down-Regulates Transforming Growth Factor Beta Receptor II (TGFBR2) Expression in Prostate Cancer. Oncogene 2014, 33, 4097–4106. [CrossRef]
- Kneitz, B.; Krebs, M.; Kalogirou, C.; Schubert, M.; Joniau, S.; Van Poppel, H.; Lerut, E.; Kneitz, S.; Scholz, C.J.; Ströbel, P.; et al. Survival in Patients with High-Risk Prostate Cancer Is Predicted by miR-221, Which Regulates Proliferation, Apoptosis, and Invasion of Prostate Cancer Cells by Inhibiting IRF2 and SOCS3. Cancer Res. 2014, 74, 2591–2603. [CrossRef]
- Cui, S.-Y.; Wang, R.; Chen, L.-B. MicroRNA-145: A Potent Tumour Suppressor That Regulates Multiple Cellular Pathways. J. Cell. Mol. Med. 2014, 18, 1913–1926. [CrossRef]
- Hart, M.; Wach, S.; Nolte, E.; Szczyrba, J.; Menon, R.; Taubert, H.; Hartmann, A.; Stoehr, R.; Wieland, W.; Grässer, F.A.; et al. The Proto-oncogene ERG Is a Target of Micro RNA miR-145 in Prostate Cancer. FEBS J. 2013, 280, 2105–2116. [CrossRef]
- Tahamtan, A.; Teymoori-Rad, M.; Nakstad, B.; Salimi, V. Anti-Inflammatory MicroRNAs and Their Potential for Inflammatory Diseases Treatment. Front. Immunol. 2018, 9, 1377. [CrossRef]
- Gunasekharan, V.; Laimins, L.A. Human Papillomaviruses Modulate MicroRNA 145 Expression To Directly Control Genome Amplification. J. Virol. 2013, 87, 6037–6043. [CrossRef]
- Shen, P.; Sun, G.; Zhao, P.; Dai, J.; Zhang, X.; Zhao, J.; Zhu, S.; Chen, J.; Tao, R.; Yang, J.; et al. MicroRNA-106a Suppresses Prostate Cancer Proliferation, Migration and Invasion by Targeting Tumor-Derived IL-8. Transl. Cancer Res. 2020, 9. [CrossRef]
- Shiau, M.-Y.; Fan, L.-C.; Yang, S.-C.; Tsao, C.-H.; Lee, H.; Cheng, Y.-W.; Lai, L.-C.; Chang, Y.-H. Human Papillomavirus Up-Regulates MMP-2 and MMP-9 Expression and Activity by Inducing Interleukin-8 in Lung Adenocarcinomas. PLoS ONE 2013, 8, e54423. [CrossRef]
- Pérez-Mora, S.; Ocampo-López, J.; Gómez-García, M.D.C.; Pérez-Ishiwara, D.G. BFNB Enhances Hair Growth in C57BL/6 Mice through the Induction of EGF and FGF7 Factors and the PI3K-AKT-β-Catenin Pathway. Int. J. Mol. Sci. 2023, 24, 12110. [CrossRef]
- Martínez-Cuazitl, A.; Gómez-García, M.D.C.; Pérez-Mora, S.; Rojas-López, M.; Delgado-Macuil, R.J.; Ocampo-López, J.; Vázquez-Zapién, G.J.; Mata-Miranda, M.M.; Pérez-Ishiwara, D.G. Polyphenolic Compounds Nanostructurated with Gold Nanoparticles Enhance Wound Repair. Int. J. Mol. Sci. 2023, 24, 17138. [CrossRef]
- Johann, D.J.; Shin, I.J.; Roberge, A.; Laun, S.; Peterson, E.A.; Liu, M.; Steliga, M.A.; Muesse, J.; Emmert-Buck, M.R.; Tangrea, M.A. Effect of Antigen Retrieval on Genomic DNA From Immunodissected Samples. J. Histochem. Cytochem. 2022, 70, 643–658. [CrossRef]
- Zandnia, F.; Doosti, A.; Mokhtari-Farsani, A.; Kardi, M.T.; Movafagh, A. Application of Multiplex PCR for Rapid and Sensitive Detection of Human Papillomaviruses in Cervical Cancer. Pak. J. Med. Sci. 2016, 32, 444–447. [CrossRef]
- Williamson, A.; Rybicki, E.P. Detection of Genital Human Papillomaviruses by Polymerase Chain Reaction Amplification with Degenerate Nested Primers. J. Med. Virol. 1991, 33, 165–171. [CrossRef]
- Evans, M.F.; Adamson, C.S.; Simmons-Arnold, L.; Cooper, K. Touchdown General Primer (GP5+/GP6+) PCR and Optimized Sample DNA Concentration Support the Sensitive Detection of Human Papillomavirus. BMC Clin. Pathol. 2005, 5, 10. [CrossRef]
- Ocadiz-Delgado, R.; Castañeda-Saucedo, E.; Indra, A.K.; Hernandez-Pando, R.; Flores-Guizar, P.; Cruz-Colin, J.L.; Recillas-Targa, F.; Perez-Ishiwara, G.; Covarrubias, L.; Gariglio, P. RXRα Deletion and E6E7 Oncogene Expression Are Sufficient to Induce Cervical Malignant Lesions in Vivo. Cancer Lett. 2012, 317, 226–236. [CrossRef]
- Kato, I. Simultaneous Detection and Typing of Genital Human papiHomavirus DNA Using the Polymerase Chain Reaction.
- Ocadiz-Delgado, R.; Lizcano-Meneses, S.; Trejo-Vazquez, J.; Conde-Perezprina, J.; Garrido-Palmas, F.; Alvarez-Rios, E.; García-Villa, E.; Ruiz, G.; Illades-Aguiar, B.; Leyva-Vázquez, M.A.; et al. Circulating miR-15b, miR-34a and miR-218 as Promising Novel Early Low-invasive Biomarkers of Cervical Carcinogenesis. APMIS 2021, 129, 70–79. [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [CrossRef]
Figure 1.
Hematoxylin and Eosin in histopathological analysis of benign and PCa Samples. (a) Representative images of the tissue architecture of control samples. Benign Prostatic Hyperplasia (BPH); and Benign Prostate Hyperplasia plus prostatitis (BPH/prostatitis). The BPH and BPH/prostatitis tissues exhibit an increase in epithelial growth with enlarged nuclei restricted to the basal layer (indicated by black arrows). Some patients display a significant inflammatory infiltrate (indicated by red arrows); while in other samples, both conditions may be observed, with cells presenting prominent nuclei in the basal and suprabasal layers, along with an inflammatory infiltrate. (b) Representative images of prostate tissue architecture in prostatic carcinogenic tissue from low, intermediate, and high Gleason grade PCa. To facilitate visual comparison and analysis, the important structures in the tissue, such as acinar lumen (1), luminal cells (2), stroma (3), and cellular organization (4), have been labeled with corresponding numbers. The scale bar indicates 200 and 50 micrometers (µm).
Figure 1.
Hematoxylin and Eosin in histopathological analysis of benign and PCa Samples. (a) Representative images of the tissue architecture of control samples. Benign Prostatic Hyperplasia (BPH); and Benign Prostate Hyperplasia plus prostatitis (BPH/prostatitis). The BPH and BPH/prostatitis tissues exhibit an increase in epithelial growth with enlarged nuclei restricted to the basal layer (indicated by black arrows). Some patients display a significant inflammatory infiltrate (indicated by red arrows); while in other samples, both conditions may be observed, with cells presenting prominent nuclei in the basal and suprabasal layers, along with an inflammatory infiltrate. (b) Representative images of prostate tissue architecture in prostatic carcinogenic tissue from low, intermediate, and high Gleason grade PCa. To facilitate visual comparison and analysis, the important structures in the tissue, such as acinar lumen (1), luminal cells (2), stroma (3), and cellular organization (4), have been labeled with corresponding numbers. The scale bar indicates 200 and 50 micrometers (µm).
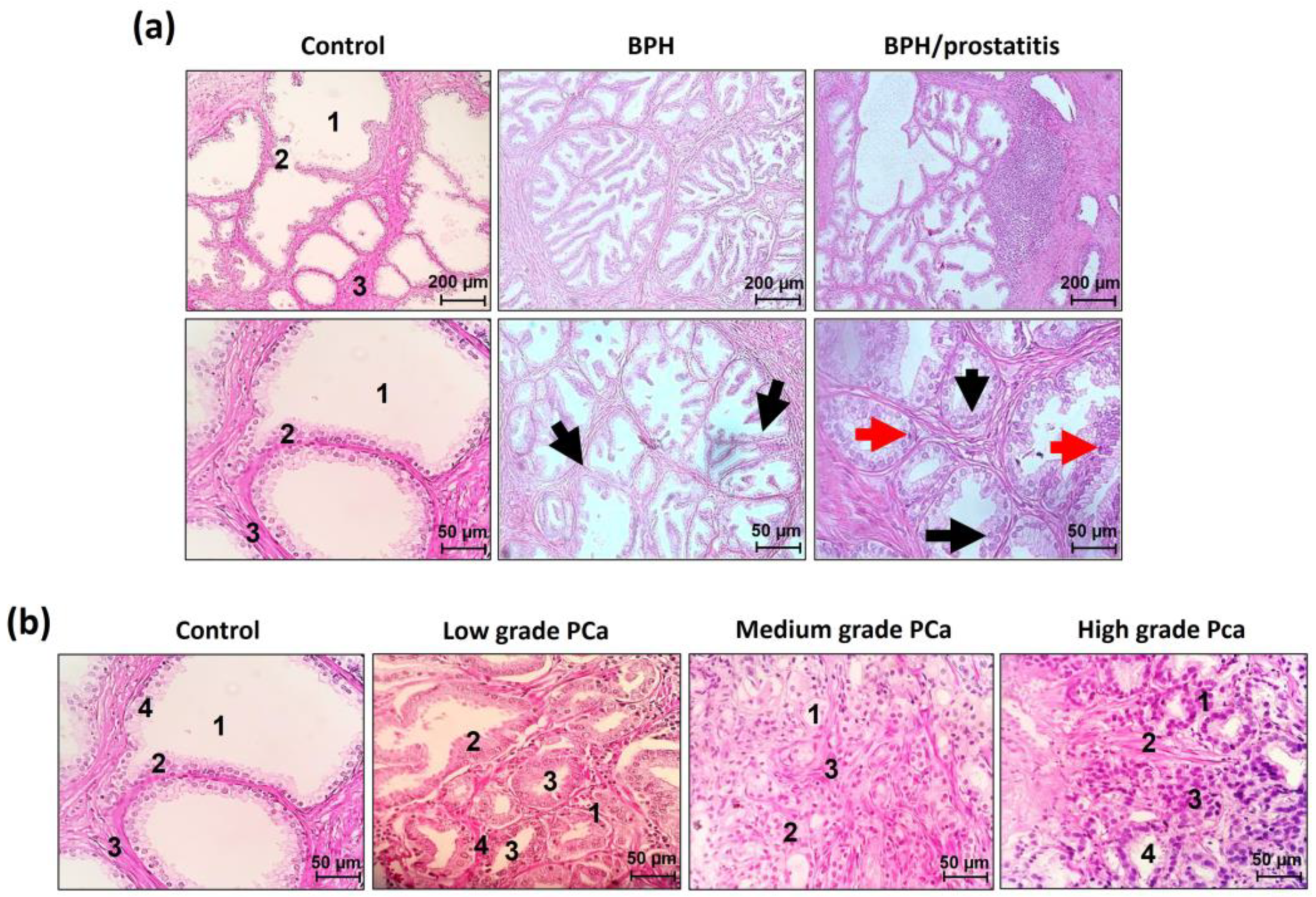
Figure 2.
Histological identification of koilocytes and in situ molecular detection of HPV sequences in BPH samples. Panel (a) displays the presence of koilocytes (black arrows), while panel (b) demonstrates the detection of HR-HPV E6/E7 DNA via in situ PCR. The signal was mainly localized in the cell nucleus (black arrows), with a positive signal of HPV DNA amplification detected in koilocytes (empty arrows with letter K). Patient control numbers are indicated. Samples 97 and 3680 were included as negative controls for HPV. Amplification: 40X and 63X. The obtained signal was digitally quantified and displayed in panel (c). Statistical analysis was performed using one-way ANOVA with Tukey’s multiple comparison test, and the results are presented as mean ± standard deviation. The asterisk symbol (*) indicates a significant difference between HPV-positive and HPV-negative samples, with a significance level of * p<0.033.
Figure 2.
Histological identification of koilocytes and in situ molecular detection of HPV sequences in BPH samples. Panel (a) displays the presence of koilocytes (black arrows), while panel (b) demonstrates the detection of HR-HPV E6/E7 DNA via in situ PCR. The signal was mainly localized in the cell nucleus (black arrows), with a positive signal of HPV DNA amplification detected in koilocytes (empty arrows with letter K). Patient control numbers are indicated. Samples 97 and 3680 were included as negative controls for HPV. Amplification: 40X and 63X. The obtained signal was digitally quantified and displayed in panel (c). Statistical analysis was performed using one-way ANOVA with Tukey’s multiple comparison test, and the results are presented as mean ± standard deviation. The asterisk symbol (*) indicates a significant difference between HPV-positive and HPV-negative samples, with a significance level of * p<0.033.
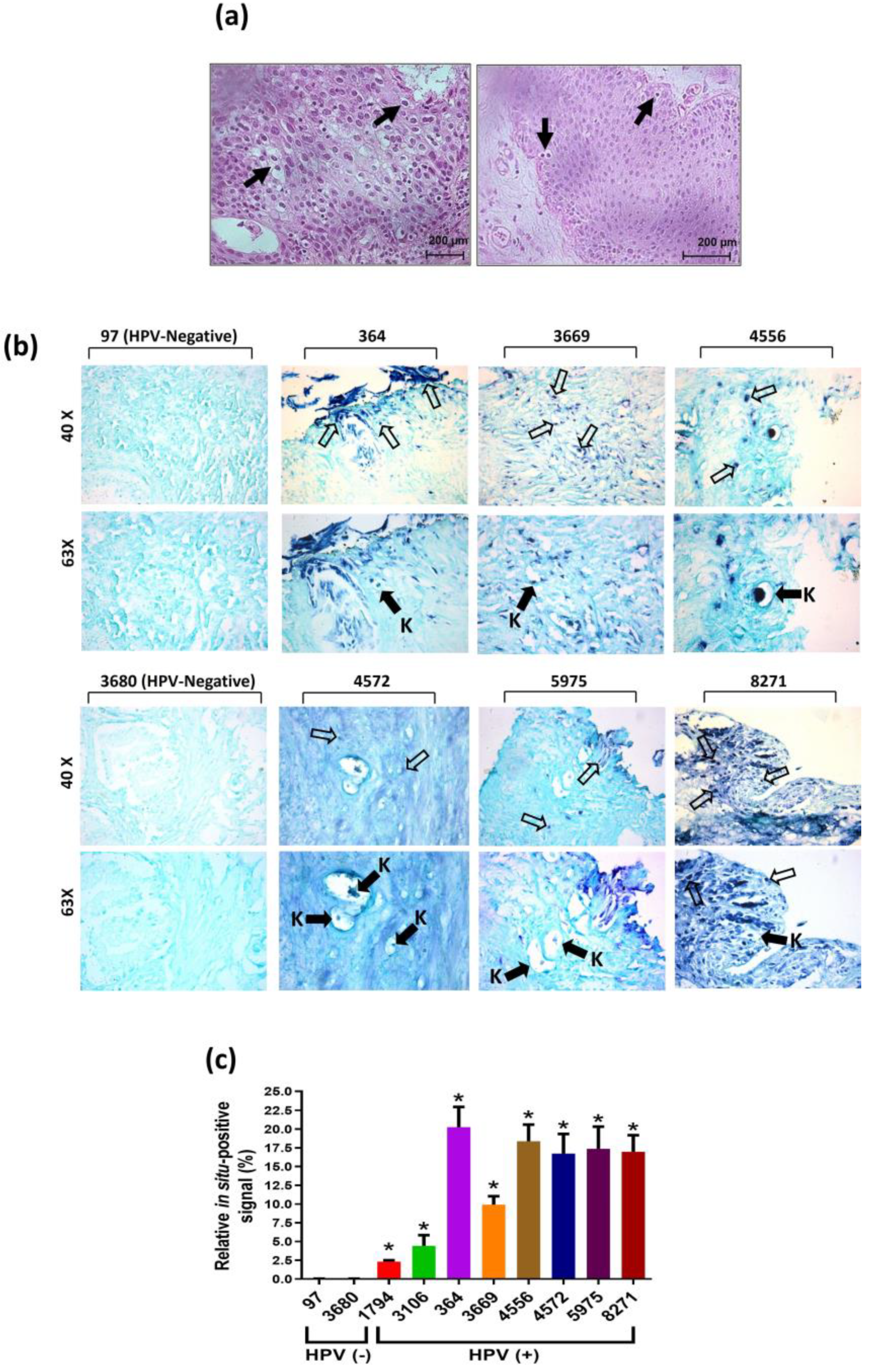
Figure 3.
miRNA expression levels in HPV-negative (−) or HPV-positive (+) benign prostatic samples. Expression of indicated miRNAs in BPH (a) or in BPH/prostatitis (b) is shown. The expression data were normalized using RNU48 as an internal control. Statistical analysis was performed using the multiple T-tests per row. The error bars represent the standard deviation. Three levels of significance were used for p-values: *p<0.033, **p<0.002, and ***p<0.001.
Figure 3.
miRNA expression levels in HPV-negative (−) or HPV-positive (+) benign prostatic samples. Expression of indicated miRNAs in BPH (a) or in BPH/prostatitis (b) is shown. The expression data were normalized using RNU48 as an internal control. Statistical analysis was performed using the multiple T-tests per row. The error bars represent the standard deviation. Three levels of significance were used for p-values: *p<0.033, **p<0.002, and ***p<0.001.
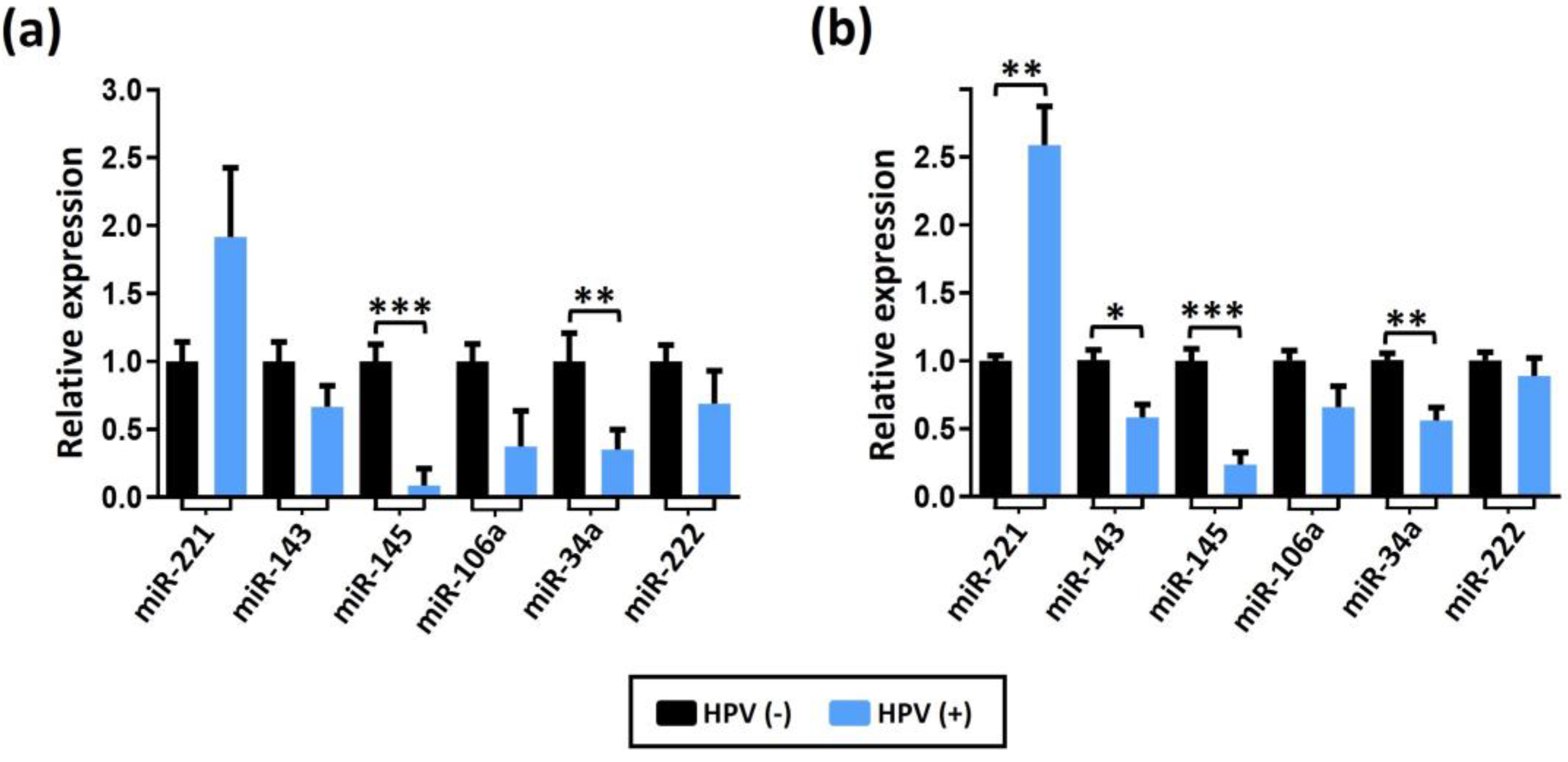
Figure 4.
miRNAs expression levels in HPV-positive PCa samples compared to their expression in control prostate tissue. Statistical analysis was performed using the multiple T-tests per row. The error bars represent the standard deviation. Three levels of significance were employed for p-values: *p<0.033, **p<0.002, and ***p<0.001.
Figure 4.
miRNAs expression levels in HPV-positive PCa samples compared to their expression in control prostate tissue. Statistical analysis was performed using the multiple T-tests per row. The error bars represent the standard deviation. Three levels of significance were employed for p-values: *p<0.033, **p<0.002, and ***p<0.001.
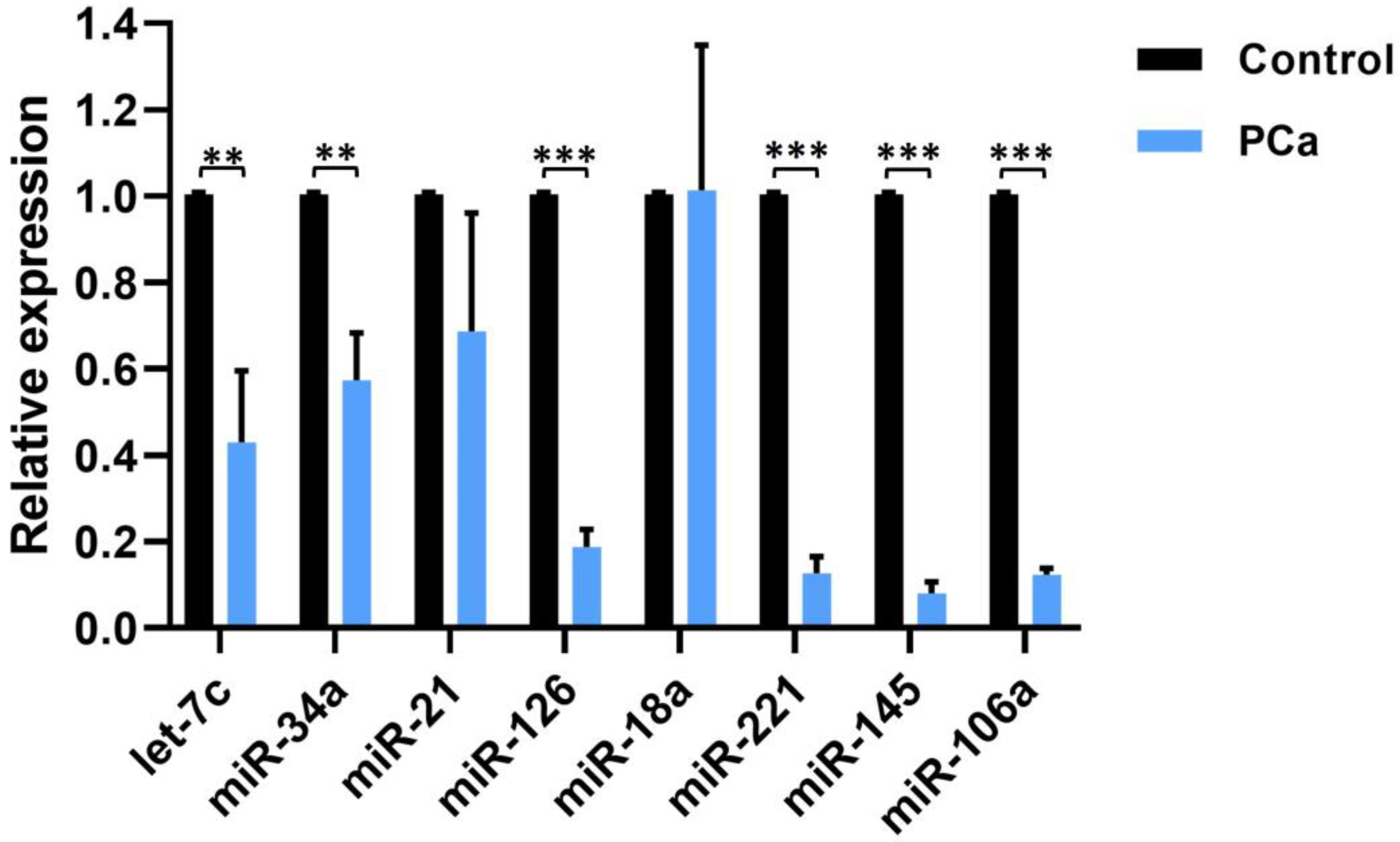
Figure 5.
Distribution of miRNA expression in high-risk (HR)-, low-risk (LR)- and intermediate-risk (IR)-HPV-PCa samples, compared to HPV-negative (NEG) PCa samples. Statistical analysis was performed using the Dunnett multiple comparison test. The p-values were reported as *p<0.033, **p<0.002, and ***p<0.001 for three levels of significance. Error bars represent the standard deviation.
Figure 5.
Distribution of miRNA expression in high-risk (HR)-, low-risk (LR)- and intermediate-risk (IR)-HPV-PCa samples, compared to HPV-negative (NEG) PCa samples. Statistical analysis was performed using the Dunnett multiple comparison test. The p-values were reported as *p<0.033, **p<0.002, and ***p<0.001 for three levels of significance. Error bars represent the standard deviation.
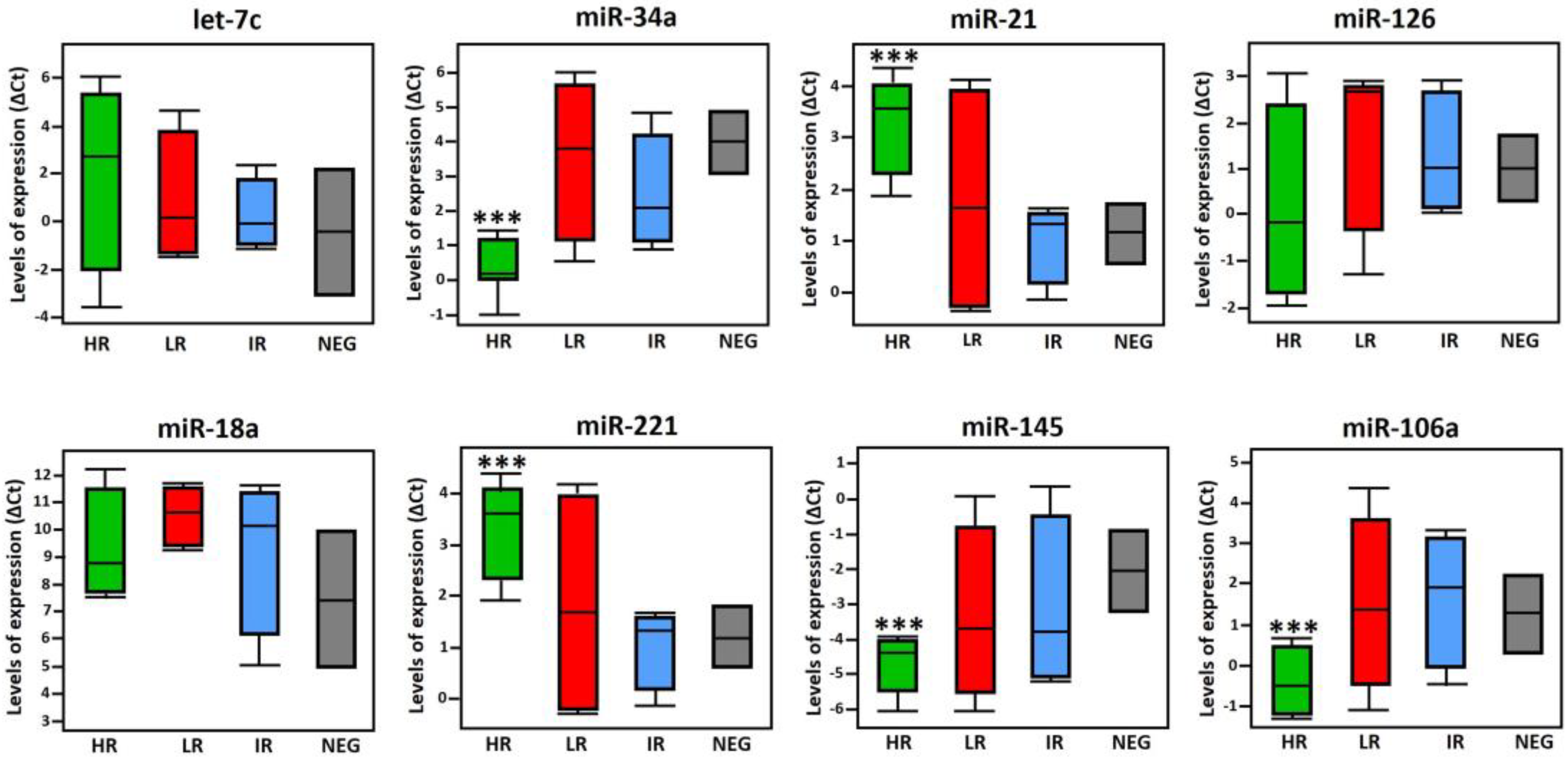
Figure 6.
Schematic overview of miRNAs potential role in HPV infection and prostate cancer progression. miRNAs such as miR-34a, miR-143, and miR-145 are frequently dysregulated in tissues affected by HPV, influencing key cellular processes like apoptosis and cell cycle arrest through their interactions with HPV oncoproteins E6 and E7. E6 leads to the degradation of p53, disrupting various miRNAs that emulate p53 functions, while E7 impedes the retinoblastoma (Rb) protein, altering E2F transcription factor activities. This miRNA dysregulation plays a crucial role in cancer progression by affecting multiple signaling pathways. Specifically, miR-34a and miR-145 suppress androgen receptor (AR) levels, and miR-21, miR-106a, and the miR-221/222 cluster target PTEN, leading to activation of the PI3K/AKT/mTOR pathway. These alterations enhance cellular proliferation, invasion, epithelial-mesenchymal transition (EMT), and metastasis. Importantly, let-7c and miR-143 downregulate the PI3K/AKT pathway, mitigating tumor growth. Moreover, miR-21 and the miR-221/222 cluster also interact with the JAK/STAT pathway, where they influence inflammatory and immune responses, further contributing to prostate cancer progression. Collectively, miRNAs are pivotal in mediating HPV’s oncogenic effects in prostate cancer by regulating essential signaling pathways, highlighting their potential as both biomarkers and therapeutic targets in the management of the disease.
Figure 6.
Schematic overview of miRNAs potential role in HPV infection and prostate cancer progression. miRNAs such as miR-34a, miR-143, and miR-145 are frequently dysregulated in tissues affected by HPV, influencing key cellular processes like apoptosis and cell cycle arrest through their interactions with HPV oncoproteins E6 and E7. E6 leads to the degradation of p53, disrupting various miRNAs that emulate p53 functions, while E7 impedes the retinoblastoma (Rb) protein, altering E2F transcription factor activities. This miRNA dysregulation plays a crucial role in cancer progression by affecting multiple signaling pathways. Specifically, miR-34a and miR-145 suppress androgen receptor (AR) levels, and miR-21, miR-106a, and the miR-221/222 cluster target PTEN, leading to activation of the PI3K/AKT/mTOR pathway. These alterations enhance cellular proliferation, invasion, epithelial-mesenchymal transition (EMT), and metastasis. Importantly, let-7c and miR-143 downregulate the PI3K/AKT pathway, mitigating tumor growth. Moreover, miR-21 and the miR-221/222 cluster also interact with the JAK/STAT pathway, where they influence inflammatory and immune responses, further contributing to prostate cancer progression. Collectively, miRNAs are pivotal in mediating HPV’s oncogenic effects in prostate cancer by regulating essential signaling pathways, highlighting their potential as both biomarkers and therapeutic targets in the management of the disease.
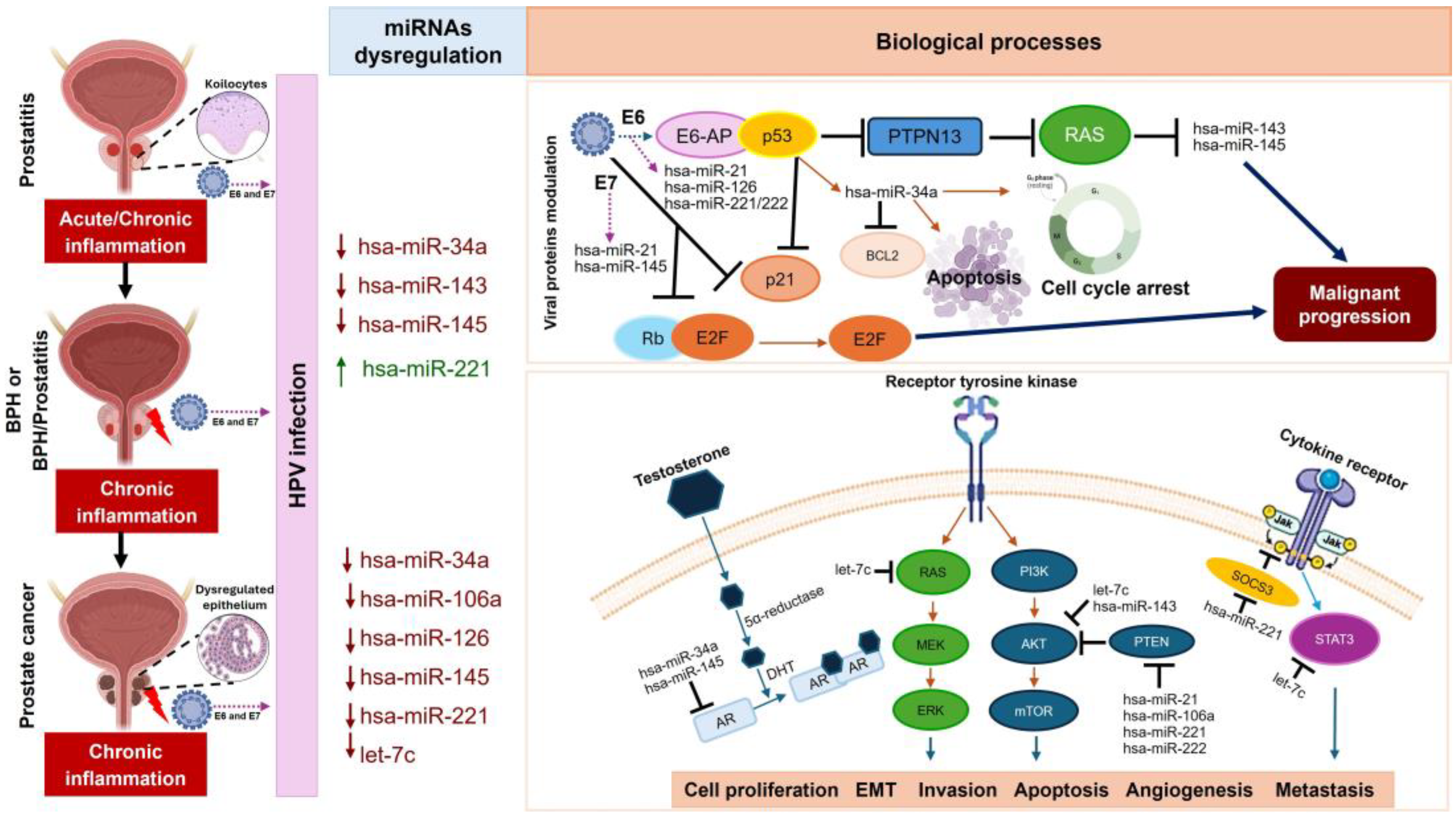

Table 1.
Histopathology stratification of selected prostate samples.
| Histopathological diagnosis | Number of samples | |
|---|---|---|
| Healthy tissues (HPV-negative tissue biopsy) |
14 | |
| BPH | 13 | |
| BPH / Prostatitis | 37 | |
| PCa | 33 | |
| Gleason (8-10) (High degree of malignancy) |
21 | |
| Gleason (7) (Intermediate degree of malignancy) |
3 | |
| Gleason <6 (Low degree of malignancy) |
9 | |
Table 2.
HPV Genotype frequency in benign and PCa lesions, according to the type and injury degree in the prostate gland.
Table 2.
HPV Genotype frequency in benign and PCa lesions, according to the type and injury degree in the prostate gland.
| Type of injury | HPV positivity frequency (%) | Predominant HPV genotypes | Coinfection [1] |
|---|---|---|---|
| Benign lesions | 67.2 | 6, 11 | |
| Malignant lesions | 93.4 | 16, 18, 31,33, 52, 58 | |
| HPV genotypes in benign lesions group | |||
| BPH | LR: 62.5 | 6,11 | ND |
| IR: 25.0 | 52,58 | 6,11 | |
| HR: 12.5 | 16,18 | 6, 11, 33 | |
| BPH/Prostatitis | LR:74 | 6, 11 | ND |
| IR: 21.7 | 52,58 | 6,11 | |
| HR: 4.3 | 16, 18 | 6,11,33 | |
| HPV genotypes in PCa group | |||
| Low risk cancer | 29.7 | 6, 11 | |
| Intermediate risk cancer | 9.1 | 31, 33, 52, 58 | |
| High risk cancer | 61.2 | 16, 18 | |
| HPV genotypes depending on PCa Gleason stratification | |||
| Grade Gleason score < 6 |
LR-HPV: 55.5 | 6, 11 | 58, 52[1] |
| IR-HPV: 11.1 | 52 | 16 | |
| HR-HPV: 33.3 | 16 | ND | |
| Grade Gleason 7 |
LR-HPV: ND | ND | ND |
| IR-HPV: 66.67 | 31,58 | 16 | |
| HR-HPV: 33.33 | 18 | ND | |
| Grade Gleason 8 - 9 |
LR-HPV: 10.6 | 6, 11 | 31, 52 |
| IR-HPV: 47.3 | 33, 31, 52 | 18 | |
| HR-HPV: 42.10 | 16, 18 | 6 | |
[1] In benign lesions BPH and BPH/prostatitis, coinfection of the low and intermediate genotype was found in samples with intermediate risk (IR) and high risk (HR) being observed in both groups of benign lesions. In contrast, in malignant PCa lesions, a higher frequency of coinfection of intermediate and high-risk genotypes is observed, in samples with a predominance of IR-HPV genotypes). ND; not detected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



