
Submitted:
15 September 2024
Posted:
16 September 2024
You are already at the latest version
Abstract
Fusarium wilt, caused by Fusarium oxysporum f.sp. cubense (Foc), is recognized as one of the most devastating diseases affecting banana cultivation worldwide. In India, Foc extensively affects the Malbhog banana (AAB genomic group) production. In this study, we isolated 25 Foc isolates from wilt-affected Malbhog plantations of India. Pathogenicity test confirmed the identity of these isolates as Foc, the primary causative agent of wilt in banana. Morpho-cultural characterization of the Foc isolates showed large variation in colony morphological features, intensity, and pattern of pigmentation, chlamydospores, and conidial size. The molecular identification of these isolates using Race-1 and Race-4 specific primers established their identity as Race- 1 of Foc, with absence of TR- 4. For a more comprehensive understanding of genetic diversity of Foc isolates, we em-ployed ISSR molecular typing which revealed five major clusters. Ahigh96% diversity within Foc population indicated the presence of polymorphic loci in individuals of a given population evi-dent from results of Nei’s genetic diversity, Shannon information index, and polymorphism in-formation content values, apart from analysis of molecular variance (AMOVA). The current findings provide significant insights towards detection of Foc variants, and consequently the deployment of effective management practices to keep the possible epidemic development of disease under control along Malbhog banana growing belts of northeast India.
Keywords:
Fusarium wilt
; Malbhog banana
; India
; Diversity analysis
; Molecular characterization
; Polymorphism
; Race
; AMOVA
1. Introduction
Bananas and plantains (Musaspp. Colla) belong to order Zingiberales [1,2], and are one of the most widely traded fruit crops worldwide. They are grown extensively in tropical and subtropical areas [3,4,5], and hold a significant position in the global market, ranking alongside other major staple crops such as rice, wheat, and maize in terms of gross value [6,7]. Banana holds a significant position as the fourth most important fruit crop globally after apples, grapes and citrus fruits [8]. The origin of bananas can be traced back to the Pleistocene period and their subsequent development is marked by gradual domestication of local and export cultivars, predominantly by the farmers of south-east Asia [9].Currently, the bananas are cultivated in more than 150 countries worldwide with the highest production (120 million tons) concentrated in Asia, Latin America, and Africa, whereas India and China are the largest producers in the domestic market [10].The Indian banana industry holds a significant position in the global market; it contributes 32% of the global basket of bananas, signifying the dominance of bananas in Indian horticulture. India’s annual export of bananas amounts to 1.35 million tons, worth 59.75million US dollars. These bananas are primarily exported to major banana destinations such as Oman, Iran, Saudi Arab, and the United Arab Emirates [11]. India is a significant player in the global banana market and holds a position as the second largest producer of bananas worldwide accounting 20.1% of global production. India cultivates bananas on an area of 0.80 million hectares producing 29.7 million tons with a productivity of 34 metric tons/ha [12]. The banana industry of northeast India contributes 2.4% of total Indian banana production [13], with Assam registering an annual production of 9.13 lakh tons of bananas from an area of 53.1 thousand hectares [14].
Northeast India, renowned globally for its banana diversity and being a national epicenter of banana production, is facing significant challenges due to prevalence of Fusarium wilt[15].The menace of this disease in this premier region assumes a more serious dimension, with favorable weather conditions for proliferation of Foc, including heavy rainfall allowing the spread of inoculums to newer areas [16], coupled with high humidity round the year, extended moisture availability in low pH coarse textured soils [17],and chemical- free low- input banana cultivation [13].
Improving our understanding about the progression of wilt disease in the field, adopting necessary cultural interventions, and eventually controlling the epidemic development of the pathogen have become increasingly formidable challenges for researchers due to susceptibility of banana cultivars to infectious diseases caused by pathogenic microorganism such as Fusarium oxysporum f. sp. cubense [18]. Malbhog banana (AAB genomic group) is a popular commercial cultivar, with the largest banana market located in Assam, India. The crop succumbs to several pests and diseases, amongst them, Fusarium wilt disease (also known as Panama wilt), poses an in surmount able risk to banana industry. ICAR-AICRP on Fruits (2020), reported incidence rates of this disease varying between 10 and 60% with an upsurge in epidemics following the replacement of susceptible Gros Michel banana variety with Cavendish cultivar[19].
Wilt disease is a vascular disease caused by soil-borne fungus, Fusarium oxysporum f. sp. cubense (E.F. Smith) and transmitted primarily through dissemination of infected plant material and long-lasting spores capable of surviving up to 20years in the soil [20,21].The identified causal agent attributed to Foc Race 1 has led to eradication of ‘Gros Michel’ banana cultivar [22,23]. Subsequently, the emergence of the novel Tropical race 4 (TR4) of Foc has proven susceptibility of Cavendish subgroup to this devastating disease [24]. In India, the Fusarium wilt disease was initially recorded in West Bengal, India. This reporting emphasized the need for implementing more stringent quarantine measures to prevent the unwanted spread of the disease [25]. Recently there has been a report of Foc TR4 in Indian states of Uttar Pradesh [26] and Bihar [27], highlighting the increasing economic losses and consequential decline in banana production in the affected regions.
Foc isolates have been classified into four evolutionary physiological races based on their pathogenicity [28]. These isolates have the ability to infect plants belonging to Musaceae (Banana plants) and Heliconiaceae (Related plants) families. In fact, there are more than 100 formae specialis of Fusarium wilt disease, each capable of affecting specific host plants in distinct ways [29,30]. Various methods viz., vegetative compatibility group test [31,32,33,34] volatile aldehyde production analysis [35], molecular markers such as restriction fragment length polymorphism (RFLP) [36], Random Amplified Polymorphic DNA (RAPD) [37,38], and amplified fragment length polymorphism (AFLP) [39] have been extensively investigated for assessing the variability of races of the highly variable Foc pathogen [35]. In contrast, molecular markers serve as a powerful tool to analyze the genetic variation present within the Foc isolates from around the world [40]. However, the present scenario in the India lacks sufficient data regarding the incidence and diversity of Foc in banana growing regions.
The present study was carried out focusing on Malbhog banana growing belts in India with the main objective of studying the genetic variation in banana wilt causing pathogen using patho-ecological and molecular variations to identify virulent strains for an effective management of Fusarium wilt disease.
2. Materials and Methods
2.1. Isolation, characterization, and maintenance of isolates
A detailed survey was undertaken to assess the distribution of Fusarium wilt in Malbhog banana growing belts of Assam, distributed over two major agro-climatic zones viz., lower Brahmaputra valley zone (Kokrajhar, Chirang, Barpeta, and Golpara) and upper Brahmaputra valley zone (Jorhat) (Table 1& Figure S1).
During the survey, the plantations were selected based on their typical visual symptoms associated with Fusarium wilt. These symptoms comprised of yellowing and browning of leaves, progressing from the older leaves coupled with upward progression, petiole breaking with a skirt like appearance, splitting of the pseudostem at the base, and internal discoloration of pseudostem. An additional screening was also done for oozing symptoms to rule out the possibility of bacterial wilt, and was thoroughly checked for the presence of pseudo-stem weevil. The collected samples were carefully packed in paper bags and labeled for further study at Department of Plant Pathology, Assam agricultural University, Jorhat, Assam (India).
Wilt infected vascular strands were carefully excised from diseased psuedostem, cleaned, and the lesions from the pseudostems were sliced into thin sections using sterilized blade. To avoid the contamination, blades were sterilized with 0.5% sodium hypochlorite (NaOCl) for 2-minutes, followed by two washings with sterile distilled water to eliminate all possible traces of NaOCl.
The sterilized samples were then dried by wrapping them in sterilized Whatmann No. 1 filter paper, thereafter 3-4 pre-sterilized sections were made from each sample andinoculated into sterilized Petri plates containing potato dextrose agar (PDA) media, supplemented with antibacterial agent in form of Streptomycin sulphate (1.2mL/240 mL PDA). Samples were labeled and incubated at 28±1°C in BOD-Incubator for 4 days until fungal mycelial growth was observed. The pathogens isolated were purified using hyphal-tip culture techniques and characterized. Subsequently, 25 cultures were maintained, and recorded for colony characteristics, mycelial texture, topography, margin, and pigmentations. They were also characterized for morphological characters viz., size, shape of macro-and microconidia along with chlamydospores and micro-photographed using trinocular light microscope (Carl Zeiss, Axiocam) at a magnification of 400x.
2.2. Molecular race identification of Foc isolates
The fungal genomic DNA was extracted using Himedia (HiPurATM Fungal DNA Purification kit) following the instructions provided in the manual. Molecular identity was validated through amplification using universal ITS primers (ITS1 5`-TCCGTAGGTGAACCTGCGG-3` and ITS4 (5`-TCCTCCGCTTATTGATATGC -3`), and sequence characterization by double pass sequencing from both forward and reverse directions. The data analysed by creating consensus using EMBOSS merger and further annotated using Basic Local Alignment Search Tool (BLAST) [53]. Furthermore, to rule out the possibility of non-pathogenic Fusarium oxysporum as well as race identification, the isolates were sequence characterized by sequencing PCR products amplified with Race 1 [41] and TR 4 specific primers [42].
2.3. Pathogenicity assay of Fusarium isolates
Pathogenicity tests were conducted for the isolates using tissue-culture (TC) banana plantlets (secondary hardened) of variety Malbhog collected from ICAR-CISH, Lucknow, India. The experiment was conducted in pot experiment using one-monthold plantlets with 3 replications for each isolate in protected shade net house. Another set of 3 plantlets was maintained as control for each of the isolate. To ensure a sterile environment, the soil with specific characteristics (pH: 5.8, organic carbon: 3.2 g/kg) was filled in polypropylene bags and autoclaved at 121°C for 15minutes under 15 lb psi pressure for 5consecutive days. The pots used for the experiment were sterilized in laminar airflow under UV light for 15-20 minutes for 3 consecutive days. Foc inoculum was prepared from 5day-old culture in Potato Dextrose broth (PDB) [43]. The cultures were incubated in a rotary shaker at 28±1°C for 5days.Subsequently, the broth was filtered through double-layered muslin cloth to obtain the spore suspension, which was then diluted 50timesto achieve a concentration of 106/mL.The number of spores was adjusted through hemocytometer before inoculation. For inoculation process, the root systems of Malbhog plantlets of uniform height were washed with running tap water and trimmed to one third of its original root mass and inoculated with 1000 mL of the conidial suspension (1x106 conidia /mL) for 30-minutes.After inoculation, the plants were closely observed for symptoms development after 30-days of inoculation. Another set of control plants were maintained by dipping the trimmed roots in sterilized water.
Pathogenicity test of the isolates was interpreted based on the following ratings for both leaf and vascular symptoms developed by International Network for the Improvement of Banana and Plantains (INIBAP) [44].
Leaf symptoms index: 1- No streaking/yellowing of older leaves, plants appear healthy; 3-Slight streaking or yellowing of older leaves; 5- streaking or yellowing on most of lower leaves; 7- extensive streaking or yellowing on all of the leaves; 9- dead/wilted plants. Ratings are mean of three replicates.
Vascular discoloration rating: 1- Corm completely clean, no vascular discoloration; 3- Isolated points of discoloration on vascular tissue; 5- discoloration of up to 1/3 of vascular tissue; 7- discoloration of between 1/3 and 2/3 of vascular tissue; 9- discoloration of greater than 2/3 of vascular tissue; 11- total discoloration of the vascular tissue. These ratings included the mean of three replicates.
2.4. Analysis of genetic diversity using ISSR markers
Genetic diversity of 25 Fusarium oxysporum isolates was evaluated using 12 highly polymorphic ISSR (Inter Simple Sequence Repeats) primers with di- or tri-nucleotide repeats [45] (Table 2). PCR amplification was carried out with 2720 Thermal Cycler in 0.2mL PCR tubes with a reaction volume of 10μL comprising the components (Emerald Amp® PCR Master Mix, Takara, Shiga, Japan) with 50 ng template DNA and 50 picomoles of primer. The standard annealing temperature was fixed in the range of48-52°C for 3 different sets of primers and PCR amplification was repeated at least twice to achieve consistency in a banding pattern. Initial denaturation at 94°C for 30 sec, annealing 40 sec for 48 or 52°C, extension for 1.30 min. at 72°C and final extension for 7 min. at 72°C.The amplified products of PCR were detected by staining with 1 µLof EtBr (0.5 µl/mL) and visualized under UV trans-illuminator (BIORAD, Molecular Imager Gel DOCTM XR).
2.5. Data analysis
The banding pattern observed in the agarose gel was scored as 1 or 0 for their occurrence or absence of bands and binary matrix was generated. The data was analyzed in NTSYS PC 2.0 software for generating dendrogram using unweighted pair group method of arithmetic mean (UPGMA) based on Jaccard’s similarity coefficients. The dataset was subsequently used for measuring genetic diversity among and within populations via parameters such as percentage of polymorphic loci, Nei’s gene diversity (h) [46] and Shannon’s information index (I) [47] using PopGene software version 1.32 software [48]. Average values for Nei's gene diversity index (h) and Shannon information index (I) were evaluated using GenAlex version 6.5 [49]. Additionally, the polymorphism information content (PIC) for dominant markers was calculated using the general equation
where f is the marker frequency in the data set. PIC assesses the discriminatory ability of a marker to detect polymorphism [50].
Genetic structure of Foc isolates was evaluated using Analysis of Molecular Variance (AMOVA), which estimates the variance components and their statistically significant levels of variation between and within the 25 isolates, with GenAlex version 6.5. Genetic differentiation was determined using PhiPT which calculates the level of genetic divergence in the samples. The test for statistical significance was done with 9999 permutations of random shuffling.
3. Results
3.1. Wilt symptomatology:
Malbhog banana is extensively cultivated as one of the commercial cultivars in the north-east India which is affected by Fusarium wilt and field diagnosis mainly relies on morphological symptoms. Typical symptoms of Fusarium wilt such as yellowing and gradual wilting of older leaves around the margins of leaf lamina, distortion of leaf blade, and petiole collapse to collectively produce a hanging skirt-like appearance were recorded (Figure 1). Eventually, all the infected plant leaves dried up coupled with pseudostem splitting visible at the base; while internal symptoms included discoloration of infected rhizome and pseudostem after examining either transverse or longitudinal section. As the wilt infection advanced, discoloration symptoms varied from pale-yellow to reddish-brown. These symptoms were recorded in more than 6monthsoldbanana plants. Many of the previous reports could be seen describing these distinguishing characteristic symptoms depending upon banana cultivars and growing conditions [51].
3.2. Isolation and morphological characterization of Foc isolates
The fungus was identified as Fusarium oxysporum using single spore isolation on PDA and based on the cultural (Table 3) and morphological characters (Table 4). Different Foc isolates were observed displaying significant diversity in morpho-cultural characters describing colony color as white, white with pink pigmentation at the centre. Among 25 isolates, white color was observed as most dominant color (44%), followed by white with pink centre (18%), pale white (12%), pinkish (8%), slightly purple (8%), pale yellow (4%) and purple (4%) in a pattern of decreasing order (Figure 2). The shape of the colonies also showed litte variability as 23 isolates had a circular smooth growth (92% isolates) and rest of the three displayed irregular shaped mycelial growth (8% isolates) (Table 3). Concerning the mycelial texture of Foc isolates, 11 isolates (44%) possessed abundant raised fluffy cottony mycelia, 6 isolates (24%) showing less fluffy hairy spaced mycelia, and rest of the 8 isolates (32%) showed a flat humid mycelia (Figure 3). These Foc isolates exhibited a great variability in their pigmentation on PDA media where white colored pigmentation was the most dominant color distributed in 11 isolates (44%), followed by light purple color in 5 isolates (20%), white purple centre in 3 isolates (12%), deep purple pigmentation in 2 isolates (8%), light purple with concentric rings in 2 isolates (8%), salmon red pigmentation in 1 isolate (4%), and pale yellow formed in only1 isolate (4%) (Figure 2).Previous studies reported varied colors and pigmentation pattern of Foc colonies from different banana belts, which comprise of hairy to cottony mycelia, spaced or abundant and variable from white to salmon to pale violet colour [52]. It was also indicated that some isolates tend to change rapidly from the pionnotal (with abundant greasy or brilliant conidia aggregates) to flat humid mycelia of white-pale yellowish to peach color on PDA culture [22,32].
A significant morphological variation amongst Foc isolates wasobserved based on conidial (macro- and micro-conidia) size (Table 4 & Figure 3). The micro-conidia were typically oval shaped cells with 0-2 septations and size (µm) ranging from 10-12 length × 2.2-3.0 width. The maximum micro-conidia size (12 × 3.0 µm) was recorded with the isolate 12 (I-12), while the lowest size (10 × 2.2 µm) was observed with the isolate 17 (I-17). Likewise, macro-conidia also showed a great variability, in size (27-30 × 3.2-3.6 µm) and septations (3-4) having falcate shape with attenuated tip and foot basal cell. Maximum macro-conidial size (30 ×3.6 µm) was observed with the isolate13 (I-13), while the lowest size (27 × 3.2 µm) was recorded with the isolate 7 (I-7). Presence and absence of chlamydospores was observed as another distinguishing feature in the different isolates which were typically round-oval-globose and developed singly or in chains, terminal or intercalary. The size of the chlamydospores ranged from 7.5 to 9.5 µm in diameter with maximum chlamydospore size (9.5 µm diameter) recorded with isolate 10 (I-10) and minimum size (7.5 µm diameter) with isolate 2 (I-2). None of the isolates were seen to producing sporodochia.
3.3. Molecular identification of Foc isolates
The high quality genomic DNA of 25 isolates was amplified with Foc Race 1 specific primer to produce ~354bp band in agarose gel characteristic to Foc. While TR4 specific primer did not produce any positive band at ~266 bp. Besides race identification, preliminarily, ITS based identification of Fusarium oxysporum f.sp. cubense using the PCR products generated by ITS1- ITS4primerfollowed by sequencing and annotation by BLAST analysis which resulted in identification till species level. The top hits of BLAST analysis was used for multiple sequence alignment and phylogenetic analysis using the CLUSTAL softwarewhich is shown as a radia tree in Figure 4. This phylogenetic tree is based on evolutionary relationship for various isolates following the maximum likelihood tree method with 500 bootstrap replicates showed Fusarium wilt fungus having highest homology with Fusarium oxysporum (>98%). Considering both the results, it is evident that all the isolates from Malbhog banana belt were identified as Foc race 1.
3.4. Pathogenicity of Foc isolates
The pathogenicity test of Foc isolates revealed the appearance of symptoms after 26 days to 33 days of post inoculation of isolates (Table 5). Initial symptoms included the appearance of slight vein clearing, yellowing on the outer portion of younger leaves followed by drooping of the leaves, stunting, yellowing of the leaves. Defoliation and necrosis of the younger leaves occur after 60 days of inoculation of pathogen and vascular discoloration of the corm was observed (Figure 5). Re-isolations were made from the inoculated Malbhog diseased plants on PDA- medium and isolated fungus was purified and characterized for morphological and cultural features to confirm as Fusarium oxysporum f.sp. cubenserace 1.
3.5. Genetic diversity analysis using ISSR markers
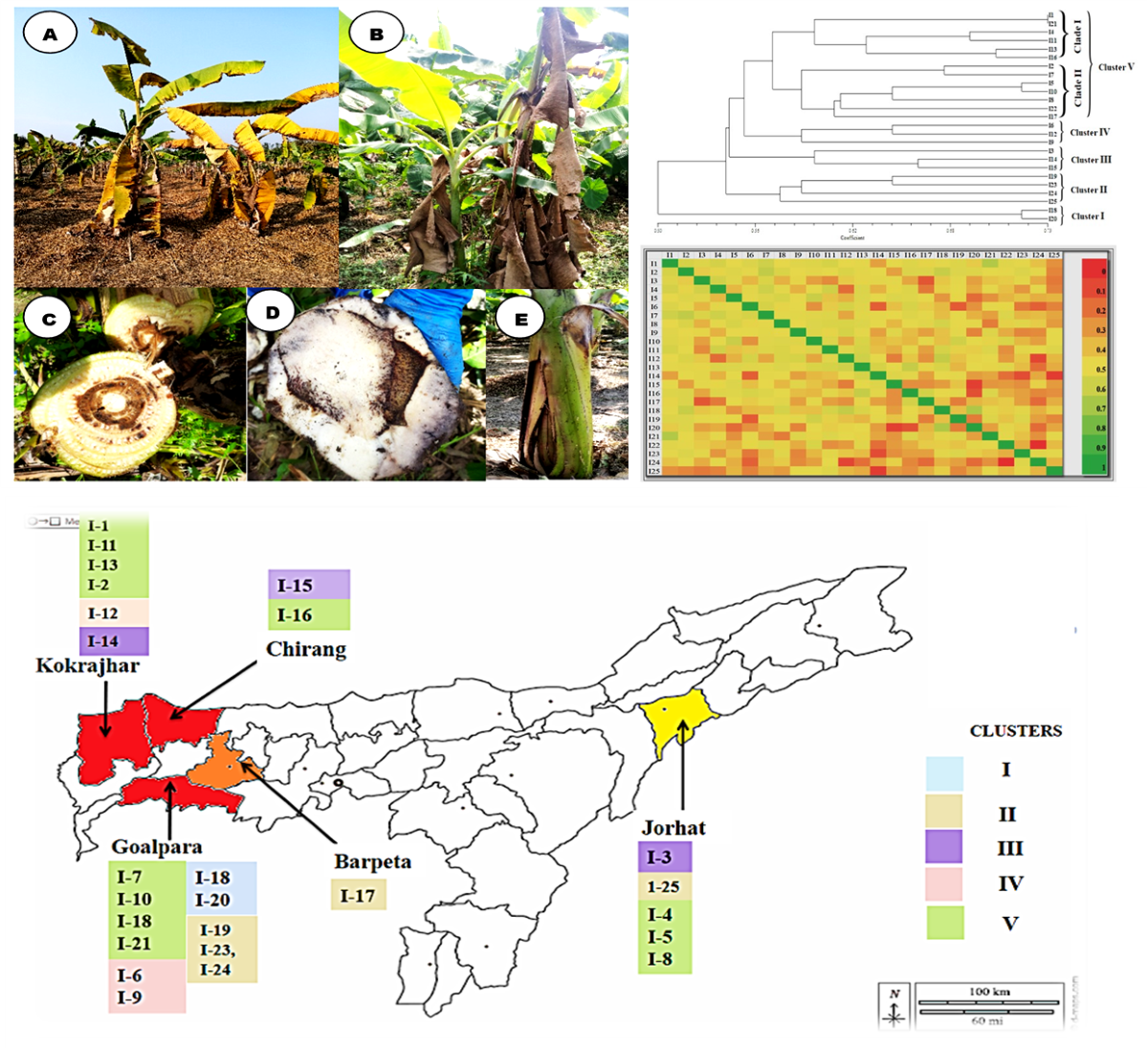
Foc isolates were subjected to genetic diversity analysis using 12 ISSR markers which produced amplicons of 200 bp to 1 Kbp (Table 6 & Figure 6). A total of 4-8 bands were obtained for each isolate and the presence of DNA bands was scored as 1 and 0 for presence or absence of the bands, respectively, followed by studies on genetic interactions. The genetic diversity and polyphyletic nature amongst 25 Foc isolates were observed using heat map analysis from distance matrix developed from finger prints of Foc isolates derived from Malbhog banana belt of India (Figure 7).The dendogram generated using UPGMA method grouped the 25 Foc isolates into five major clusters I, II, III, IV, and V. The distribution pattern of the isolates in the clusters were; 13 isolates in Cluster V (two sub- clades as clade I and clade II with a total of 13 isolates); 6 isolates cluster III and IV each with 3 isolates; 4 isolates in cluster II; and 2 isolates in Cluster I (Figure 8).The isolates from Kokrajhar (I-1, I-11, I-13, I-2), Goalpara (I-7, I-10, I-18, I-21) Jorhat (I-4, I-5, I-8) Barpeta (I-17), and Chirang (I-16)showed maximum similarity under Cluster V. Cluster I (1-18, 1-20) belonged to Goalpara and isolates from cluster II belonged to Jorhat (I-25) and Goalpara (I-19, I-23, I-24). Cluster III isolates belonged to Kokrajhar (I-14), chirang (I-15) and Jorhat (I-3). Similarly, isolates belonging to cluster IV belonged to Goalpara (I-6, I-9) and Kokrajhar (I-12) (Figure 8).
3.6. Population structure
A total of 115fragments were scored from agarose gel profiles of 12 ISSR markers, among which 97% were polymorphic. Individual primers generated 5-8 bands, with an average of 5.2bands. Number and percentages of polymorphic loci per primer (Table 6)showed highest polymorphism of 100% for six primers. Nei’s genetic diversity (h) varied from 0.2600 to 0.4735, while Shannon’s index ranged from 0.3604 to 0.6660. Evidently, both highest gene diversity (0.4735) and Shannon’s index (0.6660) were observed for primer (AC)8YA. Average values for Nei's genetic diversity index (h) and Shannon information index (I) were recorded as 0.34±0.010 and 0.50±0.014,respectively High polymorphism and Nei’s genetic diversity (h) of 0.34 denoted comparatively higher levels of genetic diversity. In the current study, average Shannon information index (I) was 0.50, denoting a moderate level of genetic diversity. The PIC value estimated to evaluate the informative capacity of the primers. In the current study ranged from 0.36 to 0.47 against the general maximum value of 0.5. High values of PIC accord for effectiveness of the ISSR primers used in assessing genetic diversity. On an average, high polymorphic information content was observed for all primers used. Primers (AC)8G, (AG)8C and (GA)8YG showed highest PIC values of 0.47, while (GA)8YT accounted for the lowest PIC of 0.36. AMOVA further confirmed the genetic variation of the Fusarium isolates. AMOVA results further showed the overall variation into two levels, among population and within populations (Table 7). A greater diversity to an extent of 96%was amongst isolates within populations, indicating polymorphic loci are prevalent in individuals of the given population. However, lower amount of variation (4%) between populations showed that the populations are not highly divergent. The molecular variation among populations accounted for meagre 4%, while the variation within population was 96%.
4. DISCUSSION
Our study delved into themorphological characters of 25 isolates from Focinfected Malbhog banana plants showing typical wilting and yellowing symptoms around the margins of the leaf lamina [55],distortion of leaf blade, and petiole collapse producing a hanging skirt-like appearance [43,56]with splitting of pseudostems at the base [57]. Internally, the infected rhizome and pseudostem showed discoloration [57,58,59]. These characteristics symptoms of Foc infection in banana plants have been documented in different studies [60,61]. Yellowing of leaves in Foc infection results from the release of fusaric acid by the pathogen and wilting and chlorosis symptoms reportedly due to the extensive formation of conidia in the xylem elements and blocking of vascular tissue [62,63]. Focinvades the root tissues of banana plants through natural openings, such as root hairs [41]. Upon invasion into the root, the pathogen colonizes and spreads through the xylem vessels, effectively blocking water and nutrient transport within the plant [64,65], leading to defoliation and necrosis of younger leaves as well as vascular discoloration in the corm. It has been reported that the production of cell wall-degrading enzymes such as cellulases and pectinases by the pathogen plays a critical role in the degradation of plant cell walls, allowing for successful penetration and colonization [66,67].
In order to understand the behavior and virulence of the pathogen, this article aims to explore the fascinating realm of morpho-cultural and molecular diversity in Foc. In our observations, we have found that 25Focisolates show different morphology including colony colour, and shape. Morpho-cultural variability amongst the 25 isolates, colour of the mycelia were predominantly white (44%), followed by white with pink centre (18%), pinkish (8%), pale white (12%), slightly purple (8%), pale yellow (4%) and purple (4%).Further, variations were observed on the margins, irregular or uniform circular shaped and smooth margins were observed amongst different isolates. Based on the topography; flat humid, less fluffy hairy spaced and abundant raised cottony fluffy characters were observed in the isolates. Most of the isolates had abundant aerial cotton mycelia (44%) as textural characters. The results of the present study are in consonance with the previous findings of earlier workers [32,35,68,69]. Das and his team also found that pigmentation varied from white to pinkish dark purple and light purplish [40].The variation in cultural and morphological characters may be due to the geographical locations of the isolates or it may be due sudden heritable changes of the isolates. Rapid mutation from the pionnotal (with abundant greasy or brilliant conidia aggregates) to flat humid mycelia of white-pale yellowish to peach colour on a PDA culture was earlier reported [22,32].
Morphological variation among the isolates through macroconidia, microconidia and chlamydospores in our study also indicates significant variation among isolates. The highest macro-conidial size (30 ×3.6 µm) was recorded in the isolateI-13 while the lowest size was recorded in the isolate I-7 with 27 × 3.2 µm. The results of morphological characterization of the present study are in agreement with those reported by several workers with reference to Foc [70,71] who also reported the size of the macroconidia, microconidia and chlamydospores as in range obtained in present investigation. Perez-Vincente in their findings described macroconidia (27-55×3.3-5.5 µm) as abundant, falcate to erect to almost straight of thin walls with 3-5 septa (usually 3) and microconidia (5-16 × 2.4-3.5µm) without septa, oval, elliptic to kidney shaped and develop abundantly in false heads in short monophialides [52]. The variation in macro and micro-conidial size observed in Foc isolates highlights the importance of understanding the genetic diversity within this pathogen. This diversity may have implications for the pathogenicity and virulence of Foc, as well as its ability to adapt and evolve [72].Previously several studies have been conducted to understand the pathogenicity of Foc and its effects on banana plants [73,74].
Pathogenicity test of our 25 the isolates of Foc showed symptom development at 26- 33 days after inoculation of all the isolates wherein isolates I-2, I-11, I-14, I-21 and I- 24 showed highest total vascular discoloration and highest leaf symptoms index. Earlier, Aguilar-Hawod observed that the isolates with high vascular rating produced more aggressive leaf symptoms however race diagnosis via morphological characters was inconsistent [75]. These five isolates belonging to Kokrajhar (LBVZ) and Goalpara (LBVZ) also showed higher disease severity in the fields which could be attributed to mono-cropping of Malbhog banana in these areas [15], that might have augmented virulence in these isolates besides congenial soil, climatic parameters and inocula load. It was also observed that Foc race 1 is commonly found in Assam due to various factors, including geographical location and environmental conditions[54]. Race 1 specifically was commonly found in tropical regions such as the Philippines and Indonesia, which share similar climatic conditions with Assam. Assam, located in North-eastern India, has a tropical climate characterized by high temperatures and humidity, providing favorable conditions for the survival and spread of Foc. These findings suggest that the virulence of Foc is not only influenced by genetic factors but also by environmental conditions specific to each geographical region [76].
The molecular identification of races within the Foc is crucial for their accurate classification and understanding of their genetic diversity.As a preliminary confirmation, ITS based sequence characterization identified all isolates to be Fusarium oxysporum isolates, and nearest top hits of BLAST analsyis based evolutionary phylogenetic tree constructed clustered the isolates into 3 broad clusters.In race identification of Foc isolates through race specific PCR analysis, the all the isolates invariably produced ~354bp amplicon characteristic to Foc Race 1 confirming them to be Foc race 1. Similar results have been reported with reference to race specific primers developed by [41] differentiated 11 Foc Race 1 isolates in China from Race 2.
Furthermore, studying genetic diversity is essential for understanding the evolutionary dynamics and population structure ofpathogens. Compared to Foc isolates of Indian banana belts distributed across seven Clusters identified through ISSR analysis [45], our diversity analysis in Malbhog banana belts revealed the presence of five different clusters I, II, III, IV and V of which the major cluster V represented by five districts viz., Kokrajhar, Barpeta, Chirang, Goalpara and Jorhat (I-1, I-2, I-4, I-5, I-7, I-8, I-10, I-11, I-13, I-16, I-17, I-21 & I-22 ) with two clades carried maximum isolates followed by Cluster IV, Cluster III, Cluster II and Cluster I in decreasing order of their dominance. Isolates (I-2) from Kokrajhar displayed maximum virulence causing more than 76.00 % percent disease incidence compared to other isolates. Maximum diversity was observed in Goalpara district with 4 nos. of clusters (Cluster I, II, IV &V) followed by Kokrajhar (Cluster III, IV & V) and Jorhat (Cluster II, III & V). Goalpara district has Asia’s largest Banana market (Darangiri Banana market) and the growers rely on suckers from various nearby districts as planting materials. So, it may be a reason Goalpara district showing highest morpho-cultural and genetic variability among the isolates.
Jaccards coefficient similarity matrix further indicated that the isolates collected from the same location/region belonged to the same cluster, however few of them belonged to other cluster that are partially related to geographical regions showing these 25 Foc isolates as a mixture of isolates from the populations in our study. Such possibility could be assigned to collection and use of infected planting material from other geographical locations or pathogen diversity of is influenced by the combination of agro-climate of the particular geographical location, relationship with the crop, and the pathogen [77]. Chittarath reported the isolates are uniquely associated with specific geographical regions [78]. However, there is a limited information available on the biogeography of this genus in relation to soil and climate. Environmental factors such as rainfall, temperature, humidity, soil conditions, and local vegetation play a crucial role in determining the severity and spread of Fusarium species [79]. Although, the foremost important factor is the relationship between pathogen growth rate and temperature, besides their survival ability at extreme temperatures [80]. Fusarium spp. thrives under warm and humid climates, with temperatures ranging between 24 and30℃ as optimal environmental conditions [81,82,83,84]. High rainfall or excessive irrigation can create favourable conditions for the pathogen's spores to germinate and infect the roots of susceptible Malbhog banana cultivar, due to development of more virulence and thereby, the elevated disease incidence. More specifically in India, virulent strain of Race 1 has been reported from TN, AP, Gujarat, and Assam, whereas, Foc TR4 from Bihar and U.P. [27,85]. Factors such as gene flow, spontaneous natural mutations, and genetic drift contribute towards variation in the Foc isolates [86].These reasons collectively attribute towards wide genetic diversity among the Foc isolates with their polyphyletic nature. The genetic variation within Foc isolates studied by various researchers using different markers such as RFLP, rDNA-ITS RFLP and DNA [36,87] indicated wide genetic variation among Foc isolates. The clustering pattern using ISSR marker data in the present study also established that the Foc isolates have clustered as per the eco-geographical adapatations or location specific clusters of Assam into five clusters. The present study has clearly indicated that ISSR could precisely measure the genetic diversity among the Foc isolates as reported by earlier workers [88,89].
5. CONCLUSION
Diversity analysis of Foc isolates from Malbhog banana belts studied through morphological, cultural, and molecular tools established the dominant presence of race 1, a big respite to banana belts of this region. Studies relating evolutionary relationship of Foc isolates would further help in developing future management strategies, apart from screening of Foc isolates from other commercial banana belts of India and introduction of defense-related genes from wild banana germplasm as banana hot spots, hence, become mandatory in order to develop a stringent strategy to contain the entry as well spread of Foc into newer banana belts.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, T.D. and PB.; methodology, A.B., B.S., and P.B.; software, A.K.B. and S.K.; validation, A.K.B., J.T. and S.S.A.; formal analysis, N.R., B.C.N.,M.M..; investigation, B.S.; resources, R.T., S.K..; data curation, S.Ki., P.B., T.D., M.M., A.K., B.K; writing—original draft preparation, P.B., A.S., S.S.A., P.P.; writing—review and editing, P.B and M.M.; visualization, U.D., J.T.; supervision, T.D and M.M.; project administration, P.B.; funding acquisition, T.D. All authors have read and agreed to the published version of the manuscript.
Funding
The authors acknowledge the funding by Department of Biotechnology (DBT), New Delhi, India funding for north east region (NER) project.
References
- Simmond, N.W.; Weatherup, S.T.C. Numerical taxonomy of the wild bananas (Musa), New Phytologist. 1990, 115, 567-571.
- Langhe, E. De.; Vrydaghs, L.; Maret, P.D.; Perrier, X.; Denham, T. Why Bananas Matter: An introduction to the history of banana domestication. Ethno. Res. and Appl. 2009, 7, 165–177.
- Zhang, P.; Whistler, R.L.; BeMiller, J.N.; Hamaker, B.R. Banana starch: production, physicochemical properties, and digestibility—a review. Carbo. poly. 2005, 59, 443-458.
- Sauco, V.; Robinson, J.C.; Tomer, E.; Daniells, J.W. Current situation and challenges of cultivating banana and other tropical fruits in the subtropics. Acta Hortic. 2012, 928, 19-30.
- Kamble, G.; Meenatai, A.; Singh, V.; Mishra, M.; Meghwal, K.; Prabhakar. Mass and surface modelling of green plantain banana fruit based on physical characteristics. Comput. and Electr. in Agric.2021, 186, 106194.
- Wardhan, H.; Das, S.; Gulati, A. Banana and Mango Value Chains. In: A. Gulati, K. Ganguly, H. Wardhan (eds) Agricultural Value Chains in India. India Studies in Business and Economics. Springer, Singapore. 2022.ISBN: 9789813342675,9789813342682.
- Singh, S.; Singh, N.; Kumar, V.; Datta, S.; Wani, A.B.; Singh, D.; Singh, J. Toxicity, monitoring and biodegradation of the fungicide carbendazim. Environ. Chemis. Lett.2016, 14, 317-329.
- Lassoudière, A. Le bananieretsa culture. editions Quae,2007,ISBN: 978-2-7592-0103-7.
- Langhe, E.; Vrydaghs, L.; Perrier, X.; Denham, T. Fahien reconsidered: Pleistocene exploitation of wild bananas and Holocene introduction of Musa cultivars to Sri Lanka. J. of Quarter. Sci. 2019, 1–5.
- FAOSTAT, www.faostat.fao.org,2021.
- https://oec.world.
- Thangavelu, R.; Saraswathi, M.S.; Uma, S.; Loganathan, M.; Backiyarani, S.; Durai, P.; Raj, E.E.; Marimuthu, N.; Kannan, G.; Swennen, R. Identification of sources resistant to a virulent Fusarium wilt strain (VCG 0124) infecting Cavendish bananas. Sci. reports. 2021, 11, 1-14.
- Hazarika, M.; Sarma, R.;Phukon, K.K. An analysis on area, production and productivity of banana in Assam. Agricul. Sci. digest –A Res. J. 2021, 41, 334-337.
- Anonymous (2018). Horticulture statistics at a glance. Govt. of India.
- Savani, A.K.; Bhattacharyya, A.; Baruah, A. Endophyte mediated activation of defense enzymes in banana plants pre-immunized with covert endophytes. Indian Phytopa.2020, 73, 433–441.
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti,E.S.G.; Staver, C.P. Fusarium wilt of banana: Current knowledge on epidemiology and research needs toward sustainable disease management. Front. Plant Sci.2018,9, 1468.
- Deltour, P.; França, S.C.; Pereira, O.L.; Cardoso, I.; Neve, S. D.; Debode, J.; Hofte, M. Disease suppressiveness to Fusarium wilt of banana in an agroforestry system: influence of soil characteristics and plant community. Agricul. Ecosy. Environ.2017, 239, 173-181.
- Ploetz, R.C. Panama disease: A classic and destructive disease of banana. Online. Plant Health Progress.2000, 1, 1.
- Stover, R.H.; Simmonds, N.W. Bananas. 3rd Edn. London: Longmans. 1987, pp. 468.
- Ploetz, R.C. Fusarium wilt of banana, Phytopath. 2015a, 105, 1512–1521.
- Ploetz, R. C. Management of Fusarium wilt of banana: a review with special reference to tropical race 4. Crop Prot. 2015b, 73, 7–15.
- Stover,R.Fusarial wilt (Panama disease) of Bananas and other Musa species. Kew: Commonwealth Mycol. Instit.1962, pp. 177.
- Ploetz,R.C. Panama disease: An old nemesis rears its ugly head part 2. The Cavendish era and beyond.Pl. Health Prog. 2005, 23, 1–17.
- Ploetz,R.C.Fusarium wilt of banana is caused by several pathogens referred to as Fusarium oxysporum f. sp. cubense. Phytopath. 2006, 96, 653- 656.
- Basu, S.K. Report on the banana disease of Chinsurah. Quart. J. Dep. Agric. Bengal. 2011, 4, 196-198.
- Damodaran, T.; Mishra, V.K.; Jha, S.K.; Gopal, R.; Rajan, S.; Ahmed,I. First report of Fusarium wilt in banana caused by Fusarium oxysporum f. sp. cubense tropical race 4 in India. Plant Disease. 2019, 103, 1022-1022.
- Thangavelu, R.; Mostert, D.; Gopi, M.; Devi, P.G.; Padmanaban, B.; Molina, A.B.; Viljoen, A. First detection of Fusarium oxysporum f.sp. cubense tropical race 4 (TR4) on Cavandish banana in India.Euro. J. of Plant Pathol. 2019, 154, 777-786.
- Moore, N.Y.; Bentley, S. Characterization of a unique population of Fusarium oxysporum f. sp. cubense causing Fusarium wilt in Cavendish bananas at Carnarvon, Western Australia. Austral. J. of Agricul. Res. 1995,46, 167-178.
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of soil fungi. Volume 1. Academic Press (London) Ltd.1980.
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium species: an illustrated manual for identification. USA News.1983, 10, 38-41.
- Ploetz, R.C.; Correll, J.C. Vegetative compatibility among races of Fusarium oxysporum f. sp. cubense. Plant Dis.1988, 72, 325–328.
- Ploetz, R.C. Population biology of Fusarium oxysporum f. sp. cubense. Fusarium wilt of banana. 1990, 63-76.
- Moore,N.Y.; Pegg, K.G.; Allen, R.N.; Irwin, J.A.C. Vegetative compatibility and distribution of Fusarium oxysporum f. sp. cubense in Australia, Austral. J. of Experi. Agricul. 1993, 33, 797- 802.
- Ploetz, R.C.; Pegg, K.G. Fusarium wilt. In Diseases of banana, abaca and enset. ed D. Jones (Wallingford, UK: CABI Publishing).2000, 143- 159.
- Stover,R.H. Studies on Fusarium wilt of bananas: VIII. Differentiation of clones by cultural interaction and volatile substances. Canadian J. of Bot. 1962,40, 1467-1471.
- Koenig, R.L.; Ploetz, R.C.; Kistler, H.C.;Fusarium oxysporum f. sp. Cubenseconsists of a small number of divergent and globally distributed clonal lineages. Phytopath.1997, 87, 915-923.
- Bentley,S.; Pegg,K.G.;Dale, J.L. Genetic variation among a worldwide collection of isolates of Fusarium oxysporum f. sp. Cubenseanalysed by RAPD-PCR fingerprinting. Mycol. Res. 1995, 99, 1378-1384.
- Bentley, S.; Pegg, K. G.; Moore, N.Y.; Davis, R.D.; Buddenhagen, I.W.; Genetic variation among vegetative compatibility groups of Fusarium oxysporum f. sp. cubense analyzed by DNA fingerprinting. Phytopath. 1998, 88, 1283-1293.
- Groenewald, S.; Van Den Berg N.; Marasas, W.F.O.; Viljoen, A. Application of AFLP in Genetic Analysis of Fusarium oxysporum f. sp. cubense. In Papers 1st. International, 2004.
- Das, A.; Venkataramana, M.; Chandranayaka, S.; Murali, H.S.; Batra,H.V. Molecular characterization of Fusarium oxysporum f. sp. cubense isolates from banana. Pest Mang. Horticul. Ecosys.2012, 18, 171-178.
- Li, M.H.; Yu, X.T.; Wang, H.F.; Zhou, J.N.; Xi, P.G.; Jiang, Z.D. Rapid detection and identification of Fusarium oxysporum f. sp. cubense race 1 and race 4. Scientia Agricult. Sinica. 2012, 45, 3971–3979.
- Dita, M.A.; Waalwijk, C.; Buddenhagen, I.W.; Souza, M.T.; Kema, G.H.I.; A molecular diagnostic for tropical race 4 of the banana fusarium wilt pathogen. Plant Pathology. 2010, 59, 348-357.
- Damodaran, T.; Rajan, S; Muthukumar, M.; Gopal, R.; Yadav, K.; Kumar, S.; Jha, S.K. Biological management of banana Fusarium wilt caused by Fusarium oxysporum f. sp. cubense tropical race 4 using antagonistic fungal isolate CSR-T-3 (Trichoderma reesei). Fronti.in micro.2020, 11, 595845.
- Carlier, J.; Waele, D.; Escalant, J. Global Evaluation of Musa Germplasm for Resistance to Fusarium wilt, Mycosphaerella Leaf Spot Diseases and Nematodes; INIBAP; Bioversity International: Rome, Italy, 2002.
- Thangavelu, R.; Muthu, K.K.; Devi,P.G.; Mustafa, M.M.; Genetic diversity of Fusarium oxysporum f.sp. cubense isolates (Foc) of India by inter simple sequence repeats (ISSR) analysis. Mol.Biotechnol. 2012, 51, 203–211.
- Nei,M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA. 1979, 76, 5269–5273.
- Lewontin, R.C. "The Apportionment of Human Diversity". Evolutionary Biol. SpringerUS.1972, 6, pp. 381–398. ISBN 978-1-4684-9065-7. S2CID 21095796.
- Yeh, F.C.; Yang, R.C.; Boyle, T. POPGENE—ver. 1.32 (32 bit): a microsoft windows-based freeware for population genetic analysis.1997. Available from http://www.ualberta.ca/~fcyeh/. Accessed 15 Dec 2011.
- Peakall, R.; Smouse, P.E.GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research—an update. Bioinfo.2012, 28, 2537–2539.
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphism. Am. J. Hum. Genet.1980, 32, 314–331.
- Ploetz, R.C. Fusarium Wilt of Banana. Phytopatho.2015, 105, 1512-1521.
- Perez Vicente, L.; Dita, M.A.;Parte, E.M.I. Technical Manual: Prevention and diagnostic of Fusarium wilt (Panama disease) of banana caused by Fusarium oxysporum f.sp. cubense Tropical Race 4 (TR4). Proceedings of Regional workshop on the Diagnosis of Fusarium wilt (Panama disease) caused by Fusarium oxysporum f.sp. cubense. 2014, pp. 74.
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. of mol. Bol.1990, 215, 403-410.
- Wu, K.L.; Chen, W.Z.; Shuai, Y.A.N.G.; Ya,W. E. N.; Zheng, Y.R.;Anjago, W.M.; Wang, Z.H. Isolation and identification of Fusarium oxysporum f. sp. cubense in Fujian Province, China. J of Integ. Agricul. 2019, 18, 1905-1913.
- Pant,B.; Bai,T.; Du,C. Baidya, S.; Magar,P.B.; Manandhar, S.; Shrestha, J.; Dita,M.;Rouard, M.; Fu,G.; Zheng, S.J. Molecular Diagnosis and Vegetative Compatibility Group Analysis of Fusarium Wilt of Banana in Nepal. J. of Fungi. 2023,9, 208.
- Jamil, F.N.; Hashim, A.M.; Yusof, M.T.; Saidi, N.B. Analysis of soil bacterial communities and physicochemical properties associated with Fusarium wilt disease of banana in Malaysia. Sci. Rep.2022, 12, 999.
- Yi, U.; Zaharah, S.S.; Ismail, S.I; Musa, M.H. Effect of Aqueous Neem Leaf Extracts in Controlling Fusarium Wilt, Soil Physicochemical Properties and Growth Performance of Banana (Musa spp.) 2021. 13, 22, 12335.
- Oyesigye, E.; Tinzara, W.; Karamura, G.; Cosmas, W. Distribution and farmers’ knowledge on Fusarium wilt (Race 1) in cropping systems of Uganda. Afr. J. of Pl. Sci. 2021, 15, 277-287.
- Thangavelu, R.; Edwinraj, E.; Gopi, M.; Pushpakanth, P.; Sharmila, K.; Prabaharan, M.; Loganathan, M.; Uma, S. Development of PCR-Based Race-Specific Markers for Differentiation of Indian Fusarium oxysporum f. sp. cubense, the Causal Agent of Fusarium Wilt in Banana, J. of Fungi (Basel), 2022,8, 53.
- Ulilalbab, A.R.; Widinugraheni, S.; Masanto, M.; Subandiyah, S.; Wibowo, A. Expression of SIX1b and SIX1c effector genes and banana resistance genes during Foc TR4 infection on banana cultivars. Biodiv. J. of Bol. Div. 2022, 23.
- Yuan, X.; Hong, S.; Xiong, W.; Raza, W.; Shen, Z.; Wang, B.; Li, R.; Ruan, Y.; Shen, Q.; Dini-Andreote, F. Development of fungal-mediated soil suppressiveness against Fusarium wilt disease via plant residue manipulation. Microbiome. 2021, 9,200.
- Dong, X.; Ling, N.; Wang, M.; Shen, Q.; Guo, S. Fusaric acid is a crucial factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants. Pl. Phy.and Bioche.2012, 60, 171-179.
- Dong, X.; Ling, N.; Wang, N.; Shen, Q.; Guo, S.; Fusaric Acid accelerates the senescence of leaf in banana when infected by Fusarium. World J. of Microbio. and Biotech.2014,30, 1399-1408.
- Effendi, Y.;Pambudi, A.; Pancoro, A. Metagenomic analysis of Fusarium oxysporum f.sp. cubense-infected soil in banana plantation, Sukabumi, Indonesia. Biodiversitas. 2019, 20, 1939-1945.
- Ding, Z.; Li, M.; Sun, F.; Xi, P.; Sun, L.; Zhang, L.; Jiang, Z. Mitogen-Activated Protein Kinases Are Associated with the Regulation of Physiological Traits and Virulence in Fusarium oxysporum f. sp. cubense. PLOS ONE. 2015, 10, e0122634.
- Roncero, M.I.;Di Pietro, A.; Ruiz-Roldán, M.C.; Huertas-González, M.D.; Garcia-Maceira, F.I.; Méglecz, E.; Jiménez, A.;Caracuel, Z.; Sancho-Zapatero, R.; Hera, C.; Gómez-Gómez, E.; Ruiz-Rubio, M.; González-Verdejo, C.I.; Páez, M.J. Role of cell wall-degrading enzymes in pathogenicity of Fusarium oxysporum. Rev Iberoam Micol. 2000, 17, 47-53.
- Zhang,L.; Yan,J.; Fu,Z.; Shi,W.;Ninkuu,V.; Li, G.; Yang, X.; Zeng, H. FoEG1, a secreted glycoside hydrolase family 12 protein from Fusarium oxysporum, triggers cell death and modulates plant immunity. Mol. plant Pathol. 2021, 22, 522-538.
- Perez-Vicente, L. Fusarium wilt (Panama disease) of bananas: an updating review of the current knowledge on the disease and its causal agent. In. Memorias de XV Reunion Internacional de ACORBAT (Oaxaca, MX). 2004, pp. 1-14.
- Nel, B.; Steinberg,C.; Labuschagne, N.; Viljoen, A. Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana. Crop Prot.2007, 26, 697-705.
- Nel, B.; Steinberg, C.; Labuschagne, N.; Viljoen, A. The potential of nonpathogenic Fusarium oxysporum and other biological control organisms for suppressing fusarium wilt of banana. Plant Pathol.2006, 55, 217-223.
- Mongkutkarn, U.; Soytong, K.Isolation, identification and pathogenicity test from Fusarium oxysporum f. sp. cubense causing banana wilt. Int. J. of Agri. Technol.2012, 12, 2181.
- Li, M.; Xie, L.; Wang, M.; Lin, Y.; Zhong, J.; Kong, G.; Xi, P.; Li, H.; Ma, L.; Jiang, Z.FoQDE2-dependent milRNA promotes Fusarium oxysporumf. sp. cubense virulence by silencing a glycosyl hydrolase coding gene expression. PLoSPathog. 2022, 18, e1010157.
- Meldrum,R.A.; Fraser-Smith, S.; Tran-Nguyen, L.T.T.; Daly, A.M.; Aitken, E.A.B. Presence of putative pathogenicity genes in isolates of Fusarium oxysporum f. sp.cubensefrom Australia. Australasian Plant Pathol.2012, 41, 551-557.
- San, E.J.; Su, H.J. Rapid method for determining differential pathogenicity of Fusarium oxysporum f. sp. cubense using banana plantlets. Trop. Agri.1984, 61.
- Aguilar-Hawod, K.G.I.; Cueva, F.M.D.L.;Cumagun, C.J.R. Genetic diversity of Fusarium oxysporum f sp. cubense causing panama wilt of banana in the philippines. Pathogens. 2020, 9, 32.
- Cruz-Martín, M; Leyva, L.; Acosta-Suárez, M.; Pichardo, T.; Bermúdez-Caraballoso, I.; Alvarado-Capó, Y. Antifungal activity of Bacillus amyloliquefaciens against Fusarium oxysporum f. sp. cubense race 1. Agronomía Mesoamericana, 2021, 32, ISSN: 2215-3608.
- Lonsdale, D.; Gibbs, J. Effects of climate change on fungal diseases of trees. British Mycological Society Symposium Series. Cambridge University Press. 1996, pp. 1–19.
- Chittarath, K.; Nguyen, C.H.; Bailey, W.C.; Zheng, S.J.; Mostert, D.; Viljoen, A.;Tazuba, A.F.; Ocimati, W.; Kearsley, E.; Chi, T.Y.; Tho, N.T.; Hung, N.T.; Dita, M.; Shah, T. Karanja, M.; Mahuku, G.; Blomme,G. Geographical Distribution and Genetic Diversity of the Banana Fusarium Wilt Fungus in Laos and Vietnam. J. of Fungi (Basel), 2022, 8, 46.
- Summerell, B.A.; Laurence, M.H.; Liew, E.C.; Leslie, J.F. Biogeography and phylogeography of Fusarium: a review. Fungal Diversity, 2010, 44, 3-13.
- Sangalang, A.E.; Backhouse, D.; Burgess, L.W. Survival and growth in culture of four Fusarium species in relation to occurrence in soils from hot climatic regions. Mycol. Res. 1995, 99, 529-533.
- Hibar, K.; Daami-Remadi, M.; Jabnoun-Khiareddine, H.; Mahjoub, El.M.; Temperature Effect on Mycelial Growth and on Disease Incidence of Fusarium oxysporumf.sp. radices lycopersici. Plant Pathol. J.2006, 5, 233-238.
- Nazari, L.; Pattori, E.; Terzi, V.;Morcia, C.; Rossi, V. Influence of temperature on infection, growth, and mycotoxin production by Fusarium langsethiaeand F. sporotrichioides in durum wheat. Food Microbiology. 2014, 39, 19-26.
- Kaur, S.; Barakat, R.; Kaur, J.; Epstein, L. The effect of temperature on disease severity and growth of Fusarium oxysporum f. sp. apii races 2 and 4 in celery. Phytopath. 2022, 112, 364–372.
- Cruz, D.R.; Leandro,L.F.S.; Munkvold,G.P. Effects of Temperature and pH on Fusarium oxysporum and Soybean Seedling Disease. Plant Dis.103, 2019, 3234-3243. [CrossRef]
- Damodaran, T.; Rajan, S.; Mishra,V.K; Jha, S.K.; Gopal,R.; Ahmad,I. First report of Fusarium wilt in banana caused by Fusarium oxysporum f. sp. cubense tropical race 4 in India. Plant Dis. 2018, 103,10-22.
- Abdel-Mawgood,A.L. DNA based techniques for studying genetic diversity. Gen. div. in micro. 2012,95-122.
- Leong, S.K.; Latiffah,Z.; Baharuddin,S. Genetic diversity of Fusarium oxysporum f. sp. cubenseisolates from Malaysia. Afr. J. of Microb. and Res.2010, 4, 1026-1037.
- Sudisha, J.; Kumar,S.A.; Thakur, R.P.; Rao,V.P.; Shetty,H.S.Molecular characterization of Sclerosporagraminicola, the incitant of pearl millet downy mildew using ISSR markers. J. of phytopath. 2009, 157, 748-755.
- Qi, Y.; Xie, Y.; Zhang; Pu,J.; Zhang, H.; Huang, S.; Zhang,H. Molecular and pathogenic variation identified among isolates of Corynesporacassiicola. Mol. biotechnol. 2009, 41, 145-151.
Figure 1.
Typical external and internal wilting symptoms observed in Malbhog banana(A) Yellowing and gradual wilting of older leaves, (B)Petiole collapse and hanging giving skirt-like appearance,(C-D)Dark brown discoloration of vascular tissue, (E) Splitting of pseudostem at the base.
Figure 1.
Typical external and internal wilting symptoms observed in Malbhog banana(A) Yellowing and gradual wilting of older leaves, (B)Petiole collapse and hanging giving skirt-like appearance,(C-D)Dark brown discoloration of vascular tissue, (E) Splitting of pseudostem at the base.

Figure 2.
Graphical representation of variation in cultural characteristics. (A): Colony colour, (B): Pigmentation, and (C): Topoghraphy of isolated Foc isolates.
Figure 2.
Graphical representation of variation in cultural characteristics. (A): Colony colour, (B): Pigmentation, and (C): Topoghraphy of isolated Foc isolates.
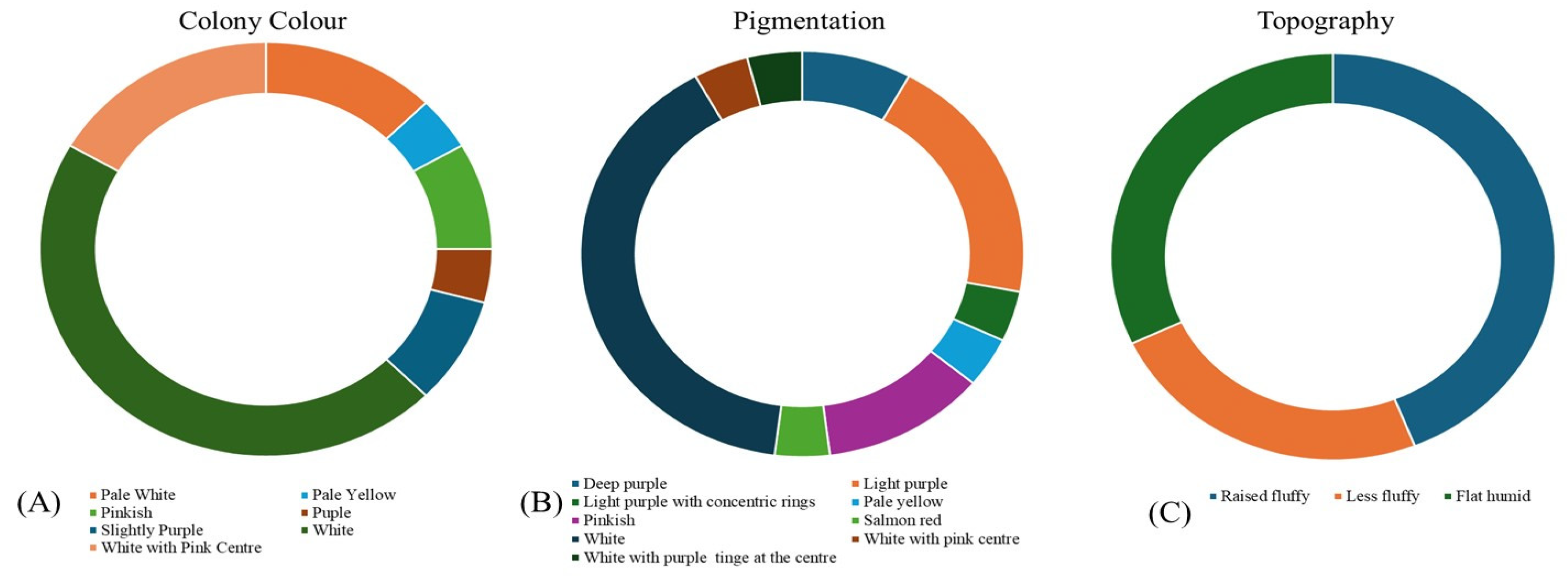
Figure 3.
Morpholo-cultural characteristics of five representative Foc isolates from five Malbhog banana belts of India.
Figure 3.
Morpholo-cultural characteristics of five representative Foc isolates from five Malbhog banana belts of India.
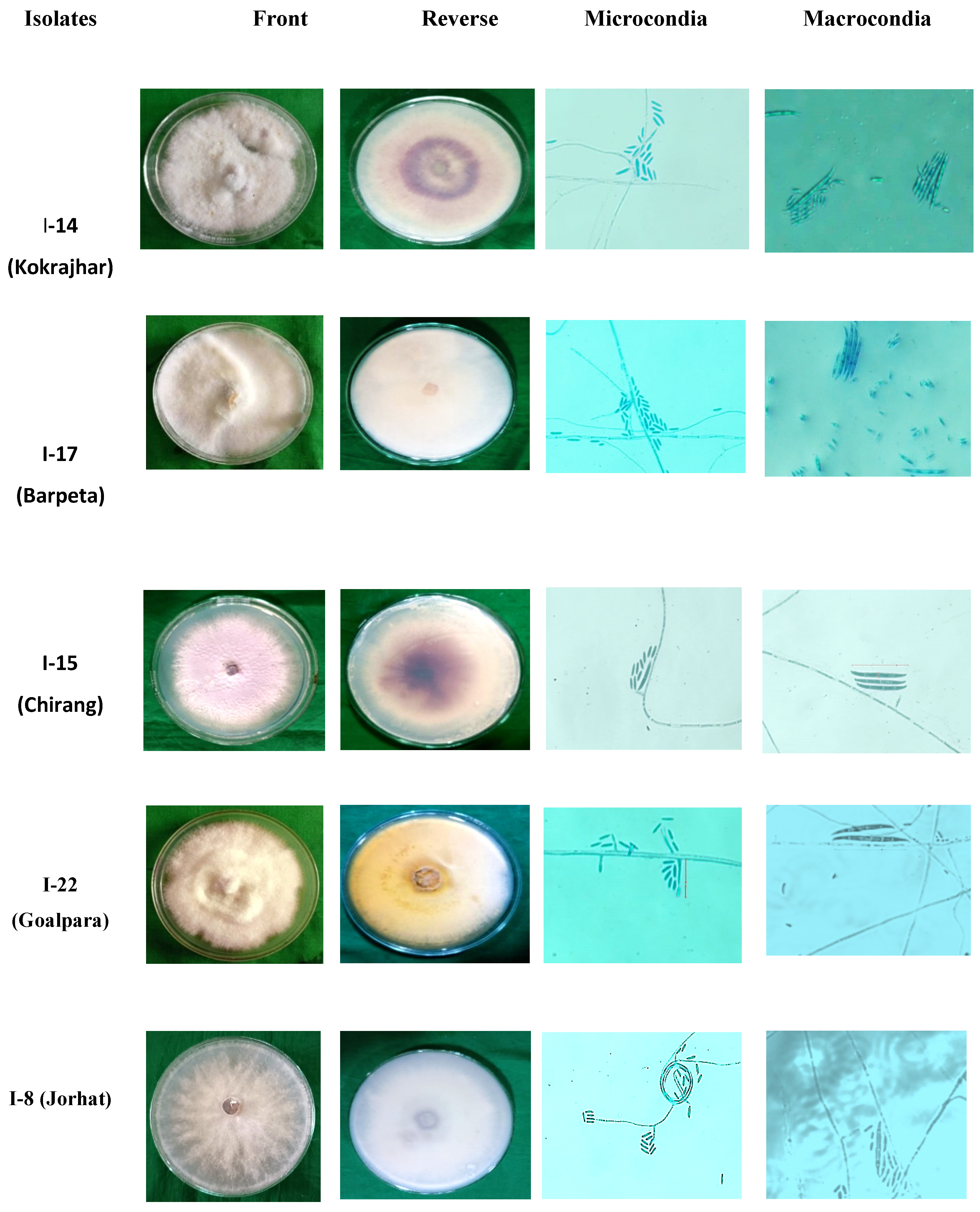
Figure 4.
Radial phylogenetic tree of Foc isolates from Malbhog banana plantation using maximum likelihood method.
Figure 4.
Radial phylogenetic tree of Foc isolates from Malbhog banana plantation using maximum likelihood method.

Figure 5.
A representative picture showing pathogenecity test of Foc isolates using TC Malbhog Banana plantlets.
Figure 5.
A representative picture showing pathogenecity test of Foc isolates using TC Malbhog Banana plantlets.

Figure 6.
Representative DNA fingerprinting profiles of Foc isolates generated by 12 different ISSR molecular markers.
Figure 6.
Representative DNA fingerprinting profiles of Foc isolates generated by 12 different ISSR molecular markers.
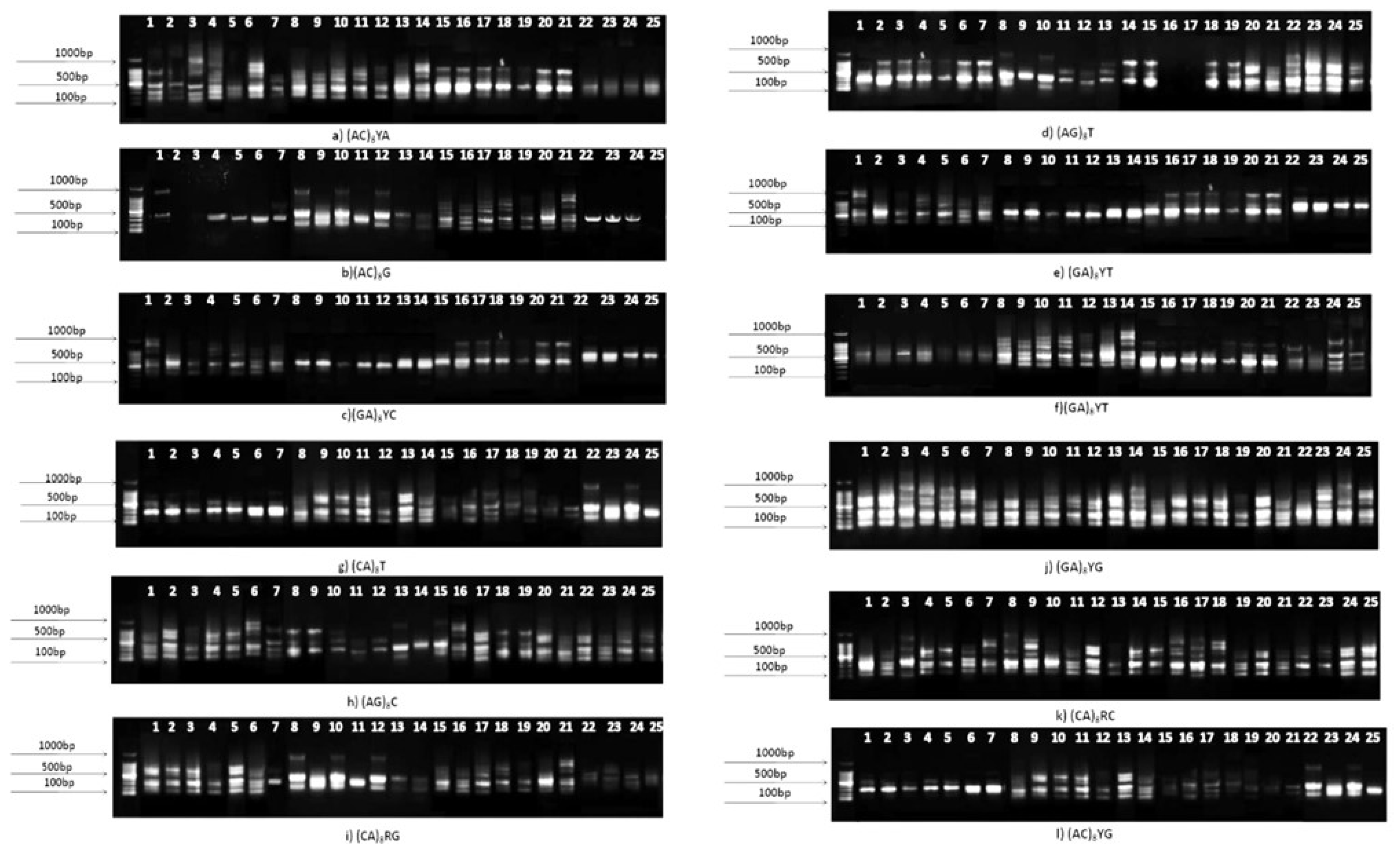
Figure 7.
Dendrogram and heatmap of 25 Foc isolates from Malbhog belts of India. (A): Dendrogram derived from UPGMA method using 12 ISSR markers showing the genetic relationships among the 25 Foc isolates from Malbhog belts of India. Five clusters were represented by Cluster I (I-18 & I-20); Cluster II ( I-19, I-23, I-24, I-25); Cluster III (I-3, I-14 & I-15); Cluster IV (I-6, I-12 & I-9) ) and ClusterV(I-1, I-2, I-4, I-5, I-7, I-8, I-10, I-11, I-13, I-16, I-17, I-21 & I-22), indicating Cluster V accommodating maximum isolates. (B): Heat map showing Jaccard’s similarity coefficient matrix.
Figure 7.
Dendrogram and heatmap of 25 Foc isolates from Malbhog belts of India. (A): Dendrogram derived from UPGMA method using 12 ISSR markers showing the genetic relationships among the 25 Foc isolates from Malbhog belts of India. Five clusters were represented by Cluster I (I-18 & I-20); Cluster II ( I-19, I-23, I-24, I-25); Cluster III (I-3, I-14 & I-15); Cluster IV (I-6, I-12 & I-9) ) and ClusterV(I-1, I-2, I-4, I-5, I-7, I-8, I-10, I-11, I-13, I-16, I-17, I-21 & I-22), indicating Cluster V accommodating maximum isolates. (B): Heat map showing Jaccard’s similarity coefficient matrix.
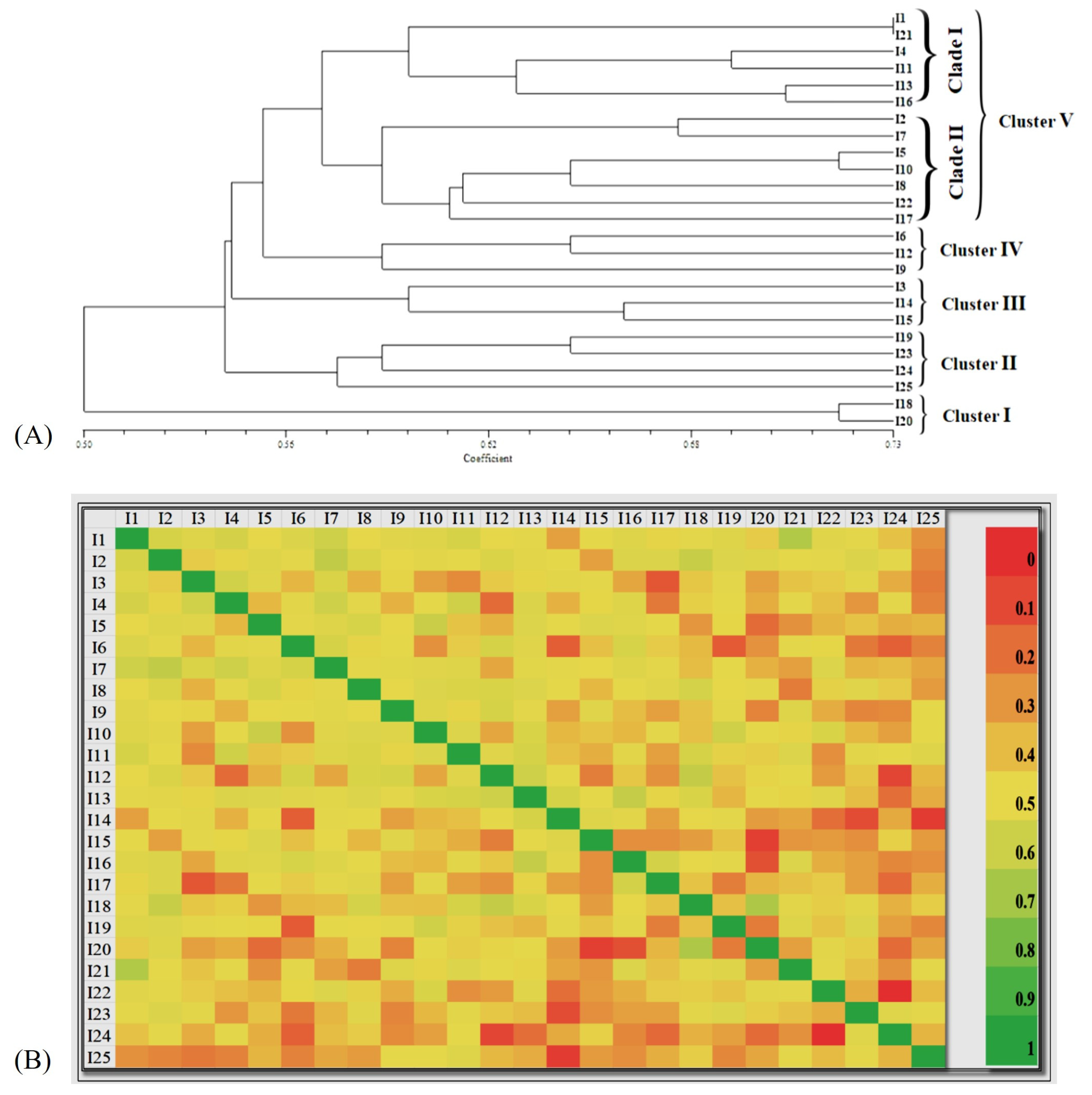
Figure 8.
Distribution of Foc isolates from major Malbhog belts of Assam, India using cluster-based analysis.
Figure 8.
Distribution of Foc isolates from major Malbhog belts of Assam, India using cluster-based analysis.
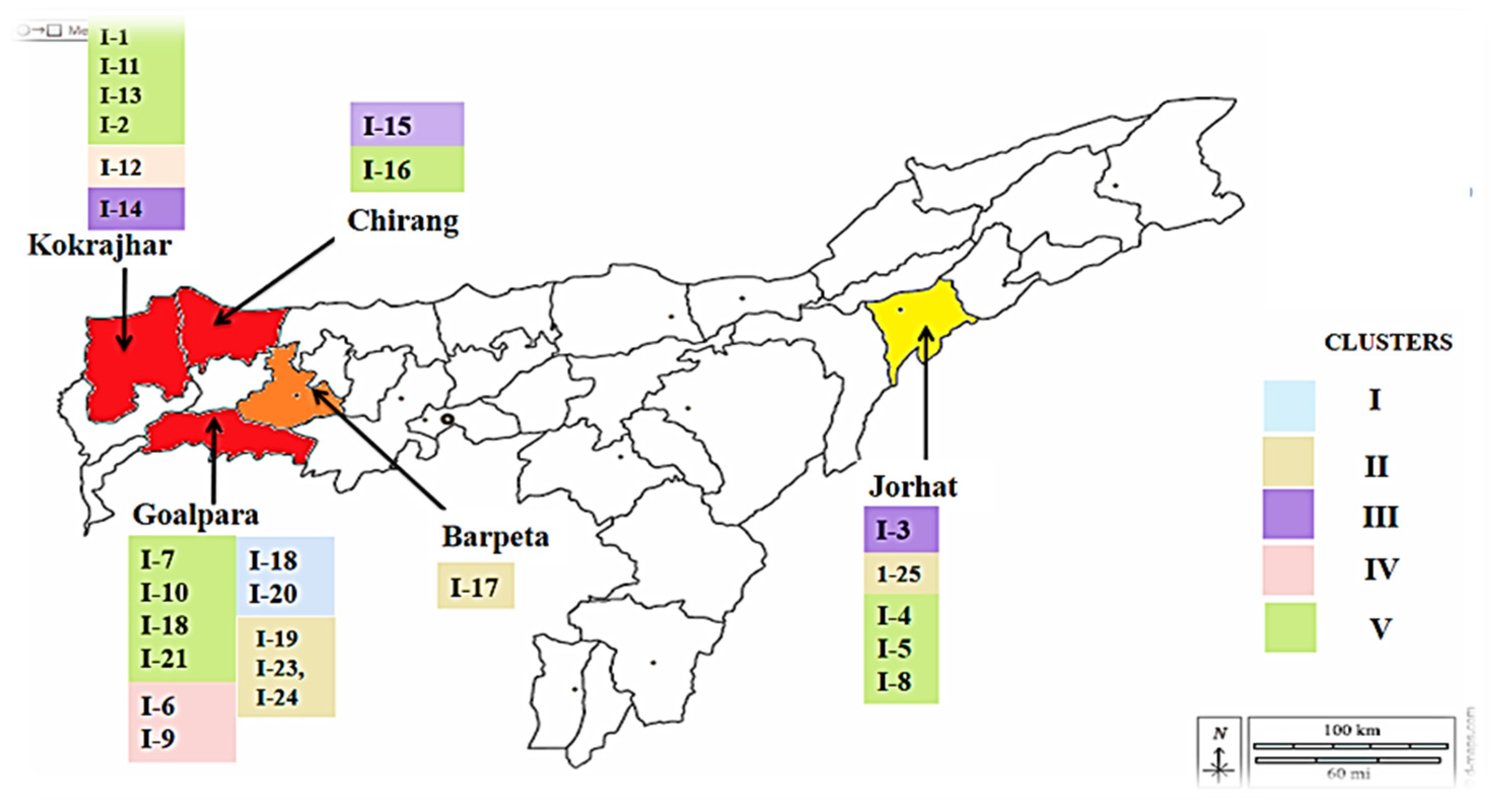

Table 1.
Details of geo-referenced sample collection sites representing Malbhog banana belts (Agro-climatically, cultivation is confined to lower and upper Brahmaputra valley zones) of India.
Table 1.
Details of geo-referenced sample collection sites representing Malbhog banana belts (Agro-climatically, cultivation is confined to lower and upper Brahmaputra valley zones) of India.
| Sl.No | Sample code | Locations | Latitude | Longitude | PDI (%) |
|---|---|---|---|---|---|
| 1. | I-1 (LBVZ) | Lakriguri, Gossaigaon, Assam, India | 26.597429˚N | 89.927368˚E | 46.00 |
| 2. | I-2 (LBVZ) | Khoksaguri-II, Gossaigaon, Asam, India | 26.50′76′′96˚N | 89.898279˚E | 76.00 |
| 3. | I-3 (UBVZ) | Horticulture experimental Farm, AAU, Jorhat, Assam,India | 26.726433˚N | 94.2046˚E | 18.00 |
| 4. | I-4 (UBVZ) | DakhinHatichungi, Jorhat, Assam, India | 26.67404˚N | 94.190131˚E | 12.00 |
| 5. | I-5 (UBVZ) | Assam Agricultural University, Jorhat, Assam, India | 26.726439˚N | 94.2021˚E | 20.00 |
| 6. | I-6 (LBVZ) | Dudhnoi, Assam,Assam, India | 25.975957˚N | 90.81605˚E | 31.00 |
| 7. | I-7 (LBVZ) | Fafal, Dudhnoi, Assam, India | 25.97548˚N | 90.815983˚E | 54.00 |
| 8. | I-8 (UBVZ) | Gohain Gaon, Jorhat, Assam, India | 26.481931°N | 94.162094°E | 17.00 |
| 9. | I-9 (LBVZ) | Dhanubhanga, Goalpara, Assam, India | 25.972059˚N | 90.980104˚E | 35.00 |
| 10. | I-10 (LBVZ) | Mandangpt-II, Assam, India | 25.940924˚N | 90.966416˚E | 29.00 |
| 11. | I-11(LBVZ) | BajugaonNo.1, Gossaigaon, Assam, India | 26.491766˚N | 89.899861˚E | 64.00 |
| 12. | I-12 (LBVZ) | Bajugaon, No1, Gossaigaon, Assam, India | 26.490674˚N | 89.899704˚E | 38.00 |
| 13. | I-13 (LBVZ) | BajugaonNo.2, Gossaigaon, Assam, India | 26.495045˚N | 89.903236˚E | 39.00 |
| 14. | I-14 (LBVZ) | BajugaonNo.3,Gossaigaon,Assam, India | 26.496353˚N | 89.902717˚E | 46.00 |
| 15. | I-15 (LBVZ) | Chirang, Assam, India | 26.545864˚N | 90.548317˚E | 29.00 |
| 16. | I-16 (LBVZ) | Chirang, Assam, India | 26.555215˚N | 90.527732˚E | 38.00 |
| 17. | I-17 (LBVZ) | Sulikata, Barpeta, Assam, India | 26.556428˚N | 90.848494˚E | 26.00 |
| 18. | I-18 (LBVZ) | Dahela, Goalpara, Assam, India | 26.030963˚N | 90.74946˚E | 22.00 |
| 19. | I-19 (LBVZ) | Karkashi, Goalpara, Assam, India | 26.029343˚N | 90.74946˚E | 31.00 |
| 20. | I-20 (LBVZ) | Dahela, Goalpara, Assam, India | 26.033461˚N | 90.747591˚E | 47.00 |
| 21. | I-21(LBVZ) | Karkashi, Goalpara, Assam, India | 26.034834˚N | 90.737234˚E | 19.00 |
| 22. | I-22 (LBVZ) | Kharamedhipara, Goalpara, Assam, India | 26.001823˚N | 90.794572˚E | 41.00 |
| 23. | I-23 (LBVZ) | Kharamedhipara, Assam, India | 26.001841˚N | 90.794601E | 30.00 |
| 24. | I-24 (LBVZ) | Mandang, Goalpara, Assam, India | 25.941226˚N | 90.966125˚E | 27.00 |
| 25. | I-25 (UBVZ) | AAU, Jorhat, Assam,India | 26.726433˚N | 94.2046˚E | 8.00 |
LBVZ and UBVZ represent lower Brahmaputra valley zone and upper Brahmaputra valley zone, respectively. PDI: Percent Diseases Incidence (%).
Table 2.
Details of ISSR primers used to differentiate Fusarium isolates from Malbhog banana belts of India.
Table 2.
Details of ISSR primers used to differentiate Fusarium isolates from Malbhog banana belts of India.
| Sl. No. | Primers | Sequence (5’-3’) | Primer length(bp) | Annealing temperature (°c) |
|---|---|---|---|---|
| 1. | (AC)8YA | ACACACACACACACACYA | 18 | 50 |
| 2. | (AC)8G | ACACACACACACACACG | 17 | 50 |
| 3. | (GA)8YC | GAGAGAGAGAGAGAGAYC | 18 | 50 |
| 4. | (AG)8T | AGAGAGAGAGAGAGAGT | 17 | 50 |
| 5. | (GA)8YT | GAGAGAGAGAGAGAGAYT | 18 | 52 |
| 6. | (GA)8YT | AGAGAGAGAGAGAGAGYT | 18 | 52 |
| 7. | (CA)8T | CACACACACACACACAT | 17 | 52 |
| 8. | (AG)8C | AGAGAGAGAGAGAGAGC | 18 | 52 |
| 9. | (CA)8RG | CACACACACACACACARG | 18 | 48 |
| 10. | (GA)8YG | GAGAGAGAGAGAGAGAYG | 18 | 48 |
| 11. | (CA)8RC | CACACACACACACACARC | 18 | 48 |
| 12. | (AC)8YG | ACACACACACACACACYG | 18 | 48 |
Table 3.
Cultural traits of 25 Fusarium isolates obtained from Malbhog banana belts of India.
| Sl.No. | Isolates | Colony colour | Topography/Texture of mycelia | Margin | Shape | Pigmentation |
|---|---|---|---|---|---|---|
| 1. | I-1 | White | Abundant Raised fluffy cottony | Smooth | Circular | White |
| 2. | I-2 | White with pink centre | Flat humid | Smooth | Circular | Light purple |
| 3. | I-3 | Light purple | Raised fluffy | Smooth | Circular | Deep purple |
| 4. | I-4 | White | Flat humid | Irregular | Circular | White |
| 5. | I-5 | White with pink centre | Raised fluffy | Smooth | Circular | Light purple |
| 6. | I-6 | Pinkish | Less fluffy | Smooth | Irregular | Light purple |
| 7. | I-7 | White | Flat humid | Smooth | Circular | White |
| 8. | I-8 | Pale white | Flat humid | Smooth | Circular | White |
| 9. | I-9 | Purple | Flat humid | Smooth | Irregular | Deep purple |
| 10. | I-10 | White with pink centre | Raised fluffy | Smooth | Circular | Light purple |
| 11. | I-11 | White | Flat humid | Smooth | Circular | White with purple centre |
| 12. | I-12 | White | Less fluffy | Irregular | Circular | White with purple centre |
| 13. | I-13 | Pinkish | Raised fluffy | Smooth | Irregular | Salmon red |
| 14. | I-14 | White | Raised fluffy | Smooth | Circular | Light purple with concentric rings |
| 15. | I-15 | Slightly purple | Less fluffy | Smooth | Circular | Light purple |
| 16. | I-16 | White | Less fluffy | Irregular | Circular | White |
| 17. | I-17 | Pale white | Less fluffy | Smooth | Circular | White |
| 18. | I-18 | White | Raised fluffy | Smooth | Circular | White with purple centre |
| 19. | I-19 | White with pink centre | Flat humid | Smooth | Irregular | Light purple with concentric rings |
| 20. | I-20 | White | Raised fluffy | Smooth | Circular | White |
| 21. | I-21 | Pale white | Flat humid | Smooth | Circular | White |
| 22. | I-22 | Pale yellow | Less fluffy | Smooth | Circular | Pale yellow |
| 23. | I-23 | White | Raised fluffy | Smooth | Circular | White |
| 24. | I-24 | White | Raised fluffy | Smooth | Circular | White |
| 25. | I-25 | White | Raised fluffy | Smooth | Circular | White |
Table 4.
Morphological characteristics of 25 Fusarium isolates obtained from Malbhog banana belts of India.
Table 4.
Morphological characteristics of 25 Fusarium isolates obtained from Malbhog banana belts of India.
| Foc isolates |
Microconidia* | Macroconidia* | Chlamydospores | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Size (µm) |
No. of septations | Shape | Size (µm) |
No. of septations | Shape | Size (µm) |
Shape | ||||
| I-1 | 10×2.5 | 1 | Oval | 28×3.4 | 3 | Falcate | 7.7 | Oval | |||
| I-2 | 10×2.4 | 0 | Oval | 28×3.4 | 3 | Falcate | 7.5 | Oval | |||
| I-3 | 11×2.5 | 0 | Oval | 30×3.5 | 3 | Falcate | 8.2 | Round | |||
| I-4 | 11×2.4 | 2 | Oval | 30×3.3 | 3 | Falcate | 8.3 | Oval | |||
| I-5 | 12×2.5 | 1 | Oval | 29×3.5 | 3 | Falcate | 9.0 | Round | |||
| I-6 | 11×2.6 | 1 | Oval | 28×3.6 | 3 | Falcate | 8.5 | Round | |||
| I-7 | 11×2.8 | 1 | Oval | 27×3.2 | 3 | Falcate | 8.0 | Round | |||
| I-8 | 11×3.0 | 1 | Oval | 30×3.5 | 3 | Falcate | 9.0 | Oval | |||
| I-9 | 11×2.6 | 2 | Oval | 29×3.5 | 3 | Falcate | 8.8 | Oval | |||
| I-10 | 11×3.0 | 0 | Oval | 28×3.2 | 3 | Falcate | 9.5 | Round | |||
| I-11 | 11×2.4 | 1 | Oval | 27×3.4 | 3 | Falcate | 8.1 | Oval | |||
| I-12 | 12 ×3.0 | 0 | Oval | 30×3.5 | 3 | Falcate | 8.5 | Oval | |||
| I-13 | 11×3.0 | 1 | Oval | 30×3.6 | 3 | Falcate | 9.0 | Oval | |||
| I-14 | 12×2.4 | 1 | Oval | 28×3.5 | 3 | Falcate | 8.7 | Round | |||
| I-15 | 12×2.5 | 2 | Oval | 28×3.5 | 3 | Falcate | 8.9 | Oval | |||
| I-16 | 11×2.5 | 1 | Oval | 29×3.4 | 3 | Falcate | 9.1 | Oval | |||
| I-17 | 10×2.2 | 0 | Oval | 29×3.5 | 3 | Falcate | 9.2 | Oval | |||
| I-18 | 12×2.5 | 1 | Oval | 27×3.6 | 3 | Falcate | 8.1 | Round | |||
| I-19 | 11×3.0 | 2 | Oval | 30×3.4 | 3 | Falcate | 8.6 | Round | |||
| I-20 | 11×2.6 | 1 | Oval | 28×3.5 | 4 | Falcate | 8.2 | Oval | |||
| I-21 | 10×2.5 | 0 | Oval | 30×3.6 | 3 | Falcate | 9.0 | Oval | |||
| I-22 | 10×2.4 | 0 | Oval | 30×3.5 | 3 | Falcate | 9.1 | Round | |||
| I-23 | 11×2.5 | 1 | Oval | 29×3.5 | 3 | Falcate | 8.3 | Oval | |||
| I-24 | 12×2.5 | 1 | Oval | 29×3.4 | 4 | Falcate | 8.7 | Round | |||
| I-25 | 11×3.0 | 1 | Oval | 28×3.5 | 3 | Falcate | 9.0 | Oval | |||
Table 5.
Pathogenicity of Foc isolates using tissue culture Malbhogbanana.
| Sl. No | Isolate Code | Days after first appearance of the symptoms | Leaf symptoms index | Vascular discoloration rating |
|---|---|---|---|---|
| 1. | I-1 | 26 | 7 | 7 |
| 2. | I-2 | 27 | 9 | 11 |
| 3. | I-3 | 30 | 7 | 9 |
| 4. | I-4 | 33 | 9 | 9 |
| 5. | I-5 | 27 | 7 | 7 |
| 6. | I-6 | 26 | 5 | 5 |
| 7. | I-7 | 32 | 9 | 9 |
| 8. | I-8 | 29 | 7 | 7 |
| 9. | I-9 | 30 | 3 | 3 |
| 10. | I-10 | 30 | 5 | 5 |
| 11. | I-11 | 28 | 9 | 11 |
| 12. | I-12 | 31 | 3 | 3 |
| 13. | I-13 | 26 | 9 | 9 |
| 14. | I-14 | 28 | 9 | 11 |
| 15. | I-15 | 32 | 5 | 5 |
| 16. | I-16 | 29 | 3 | 3 |
| 17. | I-17 | 33 | 7 | 7 |
| 18. | I-18 | 27 | 7 | 7 |
| 19. | I-19 | 29 | 3 | 3 |
| 20. | I-20 | 30 | 5 | 5 |
| 21. | I-21 | 32 | 9 | 11 |
| 22. | I-22 | 33 | 5 | 9 |
| 23. | I-23 | 29 | 5 | 5 |
| 24. | I-24 | 26 | 9 | 11 |
| 25. | I-25 | 27 | 5 | 5 |
Table 6.
Genetic diversity estimates of the 12 ISSR primers used for polymorphism.
| Primer | Number of PL* | Percentage of PL (%) | PIC† | h** | I†† |
|---|---|---|---|---|---|
| (AC)8YA | 25 | 100 | 0.41 | 0.4735 | 0.6660 |
| (AC)8G | 25 | 100 | 0.47 | 0.4672 | 0.6592 |
| (GA)8YC | 25 | 100 | 0.43 | 0.4480 | 0.6385 |
| (AG)8T | 25 | 100 | 0.43 | 0.4288 | 0.6178 |
| (GA)8YT | 24 | 96 | 0.36 | 0.3904 | 0.5701 |
| (AG)8YT | 13 | 52 | 0.44 | 0.2600 | 0.3604 |
| (CA)8T | 23 | 92 | 0.44 | 0.4160 | 0.5916 |
| (AG)8C | 24 | 96 | 0.47 | 0.4333 | 0.6166 |
| (CA)8RG | 22 | 88 | 0.40 | 0.3700 | 0.5367 |
| (GA)8YG | 25 | 100 | 0.47 | 0.4675 | 0.6596 |
| (CA)8RC | 25 | 100 | 0.39 | 0.4032 | 0.5902 |
| (AC)8YG | 24 | 96.00 | 0.42 | 0.4224 | 0.6047 |
*PL = polymorphic loci, †PIC = polymorphism information content, **h = Nei's gene diversity, ††I = Shannon's Information index.
Table 7.
Summary of Analysis of molecular variance (AMOVA) for 25 isolates of Fusarium oxysporum f.sp. cubense.
Table 7.
Summary of Analysis of molecular variance (AMOVA) for 25 isolates of Fusarium oxysporum f.sp. cubense.
| Source | Degrees of Freedom (df) | Sum of Squares | Mean Square | Estimated Variance | % variation | P value |
|---|---|---|---|---|---|---|
| Among Pops | 4 | 65.520 | 16.380 | 0.608 | 4% | <0.001 |
| Within Pops | 20 | 266.800 | 13.340 | 13.340 | 96% | <0.001 |
| Total | 24 | 332.320 | 29.72 | 13.948 | 100% | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



