Submitted:
15 September 2024
Posted:
16 September 2024
You are already at the latest version
Abstract
p97(also known as Cdc48 or VCP) is a conserved AAA+ ATPase that plays a key role in diverse cellular pathways and processes. p97 segregates damaged or misfolded substrates from cellular organelles or membranes in corporation with its cofactors. These cofactors regulate p97’s function by its directing to different cellular pathways and have diverse effect on p97´s conformation. Recently, several high-resolution structures of substrate-engaged p97 in complex with cofactors have been resolved, providing structural insights into the molecular mechanism of substrate processing. In this review, we present a comprehensive overview of how p97 in complex with substrate recruiting and substrate processing cofactors recognize, extract, translocate, unfold and release ubiquitinated substrates.
Keywords:
p97
; cofactors
; substrate processing
; Single particle cryo_electron microscopy
1. Introduction
Destabilizing mutations, metabolic challenges, and stress conditions are factors to form misfolded Destabilizing mutations, metabolic challenges, and stress conditions are factors to form misfolded state undergoes a twist rotation with respect to the D1 ring[9,17]. Despite the structural similarities between D1 and D2 domains, D1 binds nucleotides with stronger affinity than D2[20,21]. On the other hand, D2 domain was found as the main contributor of ATPase activity[22]. In addition, the nucleotide status of D1 changes the conformation of N domain (Figure 1A–D). In the UPS system, protein ubiquitination, which is a post translational modification, occurs to tag misfolded proteins using small proteins called ubiquitin molecules[23]. Ubiquitinated substrates then are recognized and extracted from their binding partners by p97 in complex with various cofactors[24]. In fact, cofactors control the substrate specificity and regulate p97 function by its directing to different cellular pathways [10]. There are two different types of cofactors including substrate-recruiting cofactors and substrate-processing cofactors that act as ubiquitin adaptors and regulators, respectively (Figure 1E). These cofactors contain binding domains which bind either the N-or C-terminal domain of p97. Most cofactors bind to N domain of p97 through either UBX or UBXL domains. Some major substrate-recruiting and substrate-processing cofactors interact with the N domain of p97 via UBXL domain such as UFD1/NPL4(UN), OTU1/YOD1, and VCIP135[25,26,27,28]. There are also cofactors that interact with p97 N domain through motifs which are including BS1(SHP box), VIM (VCP interacting motif), and VBM (VCP-binding motif)[28]. These motifs also divide into two groups some of them consist of α helices, while another group contain short stretches of unstructured hydrophobic sequences that are seen in BS1 and SHP motifs[29]. On the other hand, a small number of cofactors can bind the C-terminal disordered region of p97 via either PUB or PUL domains. Cofactors containing UBX domain can also be divided into two groups including UBA-UBX domain cofactors and UBX-domain cofactors[30]. The difference between these groups is that UBX-domain cofactors are the lack of the UBA domain , and thereby not able to bind the ubiquitinated substrates[30]. So far, more than 13 UBA-UBX proteins have been identified in human such as p47, FAF1, SAKS1, UBXD7, and UBXD8[30,31,32]. A variety of mass-spectrometry studies were conducted to understand what type of ubiquitin chains are recognized by UBA-UBX cofactors. The results showed that Lys48-linked chains are identified by all these cofactors to target polyubiquitinated substrates for proteasome degradation[33]. Intriguingly, in addition to Lys48, Lys11-linked chains have been identified by all UBA-UBX cofactors [30,34,35].
The hetero-dimer Ufd1/Npl4(UN) is involved in more than half of p97’s cellular functions. This cofactor as one of the most important ubiquitin adaptors recruits ubiquitinated substrates for p97 by interacting with the lys48-linked polyubiquitin chain[36]. Subsequently, p97 uses ATP hydrolysis by D2 domain to translocate the polypeptide through the central pore, resulting in unfolding the substrate[13,37]. In addition, substrate releasing occurs due to the ATP hydrolysis in D1 domain and also the cooperation of deubiquitinase cofactor(DUB)[13,38]. Therefore, the polyubiquitin chain is trimmed by DUB and the remaining oligoubiquitin is then pulled into the central pore. It has been suggested that at least two polypeptide chains might translocate through the central pore and then get unfolded, as this process has been reported for other hexametric AAA+ ATPases such as Cdc48, ClpXP [39,40]. In addition, these AAA+ structures indicate an asymmetric architecture when engaged with substrates, suggesting a hand-over-hand mechanism of substrate translocation[41,42,43,44]. In this review, we will focus on the main cofactors of p97 and their impacts on p97 conformation and function. We will then explain in detail the molecular mechanism of substrate processing by p97 or its fungal homologue (Cdc48) in complex with main cofactors based on the latest reports.
2. p97 Oligomerization
It was reported that almost all p97 structures determined from recombinant p97 produced in E. coli reveal a hexameric form consisting of six monomers. In addition, it is generally believed that the D1 domains of p97 recombinant are in ADP bound state even in the absence of nucleotide during sample purification[6,20,45]. The resolved p97 structure by single particle cryo-EM revealed that most D2 domains of p97 produced from E. coli are bound with ADP[17]. Furthermore, the nucleotide status of D1 changes the conformation of N domain, while that of D2 impacts the conformation of both the α helices number 9 located in C-terminus and disordered C-terminus, resulting in the oligomerization of p97. Yu et al determined a dodecamer structure of p97 in a tail-to-tail order in which both D1 and D2 domains are in apo state. They mentioned that since the D2 domain of p97 is often bound to a nucleotide, the dodecamer state is formed rarely. Therefore, it was reported that p97 dodecamer formation is due to the apo state of the D2 domain[46]. Hoq et al also reported a double-hexamer p97 structure resolved to 3.6 Å in which all the D2 domains were found to be in apo state[47].
It has been also suggested that the D2 domains of p97 hexamer hydrolyze the ATP sequentially[15,16]. On the other hand, since p97 dodecamer assembly needs all D2 domains to be empty, this protein is unlikely to be involved in the ATP hydrolysis cycle and UPS-mediated polyubiquitinated substrate degradation[46]. Therefore, dodecamerization process might keep p97 in hibernation mechanism to preserve ATP consumption[48,49]. In another study by Nandi et al, they reported the formation of dodecamer state around 30% of particles population from the p97R155H mutant. It was mentioned that the dodecamer seems to be unstable in the presence of nucleotide and thereby unable to form a complex with cofactor and substrate. However, in the absence of nucleotide, p97 mutant tends to form p97R155H dodecamer[50]. Thus, different type of nucleotides are the main elements to dissociate p97 dodecamer into two hexamer forms with an elevated ATPase activity. In addition, it was suggested that p97 dodecameric formation might be due to the shortage of ATP concentrations in the cell that leads to keeping p97 in the inactive state. In contrast to Yu et al, Gao et al reported that p97 in dodecamer state is an active ATPase and participate to process substrate unfolding[51]. Their resolved dodecamer structure showed that N domain in both sides is in up conformation when both D1 and D2 are bound to ATPγs. However, the other resolved structure of p97 double hexamer showed N domains are in down conformation when D1 is bound to ATPγs and D2 is in ADP bound state. In addition, it was found that there is an interaction between the C-terminal extension and a hydrophobic cleft in the D2 domain that is required for both ATPase activity and substrate unfolding. Moreover, the dimerization interface of p97 including α9 and conserved residues Arg745-Glu749, which forms a salt bridge with each other, are necessary for dimerization and substrate unfolding. In the other study by Liu et al, the p97 dodecamer structure was resolved in 3.0 A° resolution in complex with 12 non-ATP-competitive inhibitors of NMS-873, located in the D1/D2 interface[52]. In this structure, D1 ATPase sites were observed in the ADP-bound state, while all D2 sites were without nucleotide. In addition, in contrast to previous study, Liu et al claimed that the peptide region of Ser765 -Gly779 located in the unstructured C- terminus might induce the oligomerization of D2 domain. Meanwhile, in agreement with Gao et al, they reported that the dodecamer form is a real biological state of p97(Table 1).
3. Interaction between p97 and Ufd1/Npl4(UN)
The heterodimer UN adaptor is required for the activity of p97 in ubiquitin-proteasome system (UPS), ER-associated degradation (ERAD) pathways, mitochondria-associated degradation (MAD), and chromatin-associated degradation[32,53,54,55,56,57,58]. Intriguingly, the UPS misfolded proteins conjugated with lysine 48 (K48)-linked ubiquitin chains are recognized and processed by p97 in complex with UN cofactor[36,59]. UN plays a role as bridge to connect ubiquitinated substrate to the p97 N domain[36,60]. It has been reported that at least five ubiquitin proteins are required for recognition by the p97 in complex with UN[13], while the efficiency of substrate recognition increases in the presence of branched ubiquitin chains[37,61]. Ufd1 is composed of an N-terminal domain (UT3), and a C-terminal domain (UT6) which contains two SHP motifs. UT3 domain which is structurally similar to the p97 N domain, interacts with both mono- and poly-ubiquitinated substrate( Figure 2)[62,63]. Ufd1 binds to N-domain of p97 and Npl4 through two SHP motifs and UT6 domain, respectively[29,64]. Npl4 consists of an N-terminal UBXL domain, a central MPN domain and a C-terminal zinc finger motif. This cofactor interacts with p97 and ubiquitin protein through UBXL and zinc finger motif, respectively( Figure 2).[25,65] It has been showed that the interaction of heterodimeric UN and p97 is bipartite and both SHP1 motif of Ufd1 and the UBXL domain in NPL4 bind different p97 N domains[25,29,65].
Sato et al reported that NPL4 is the main cofactor for recognizing Lys48-linked polyubiquitylated misfolded substrate in the proteasomal degradation pathway in yeast[53]. They resolved a crystal structure of Npl4 complexed with Lys48-linked diubiquitin and revealed that both distal and proximal ubiquitin bind the Npl4 C-terminal domain (CTD). This C-terminal helix contains an N-terminal hydrophilic residue (Thr), which forms a hydrogen bonding with the NH groups of Ala46 and Gly47 of ubiquitin, and a central hydrophobic residue (Ile) that is involved in the interaction with the hydrophobic pocket formed by Leu8, Ile44 and Val70 of ubiquitin. In addition, the crystal structure of UN showed that Ufd1 interacts with a hydrophobic groove of the Mpr1/Pad1 N-terminal (MPN) domain of Npl4.
The polyubiquitinated substrate first binds to heterodimer UN and then moves into the central pore of Cdc48, which is an ATPase in yeast, through ATP hydrolysis by Cdc48- D2 domain[14]. The biochemical and structural analysis showed that different multisystem proteinopathy (MSP) mutations in p97 leads to increasing UN binding and substrate processing[66].The reason of tighter UN binding is a decoupling of p97’s nucleotide state and the preference for the up position of N domains in MSP mutant p97. Therefore, MSP mutant alters the N-domain movements, resulting in amplifying UN recruitment and substrate unfolding by p97[66]. In addition, affinity assays indicated that the amount of intercellular concentration of UN complexed with p97 in wild-type cells is less than that of UN-p97 in MSP-mutant cells, indicating the increase affinity between p97 and UN due to MSP-mutations. As a result of exceed binding to UN, less free p97 is involved to interact with other cofactors, causing a loss of cellular function in further p97-dependent pathways. Furthermore, since free UN is required for initial substrate binding, the absence of this free UN in mutant cells might impair p97-UN -dependent cellular functions[5,66,67].
Although the UN is a conserved cofactor among various species, there is an extra zinc finger in mammalian cells that is located at the C-terminus of Npl4[68]. It was proposed that since only one heterodimer UN binds to two adjacent p97 N-domains in a bipartite manner, other cofactors could potentially interact with vacant N domains and form a higher-order p97-cofactor complexes [10]. Moreover, immunoprecipitation of some UBA-UBX cofactors including UBXD7, UBXD8, SAKS1 and FAF1 contains not only p97, but also UN[30,69]. It is interesting to note that both cofactor groups are necessary for the export of misfolded proteins from the ER to the cytoplasm and recognize the K48-linked ubiquitin chain of misfolded substrate. A similar observation was also found in yeast where Ubxd2(UBA-UBX cofactor) participated in a higher order complex with Cdc48/UN[69]. Biochemical studies showed that both UBXD7 and FAF1 just bind to p97 when it is in complex with UN, but not free p97, indicating a hierarchy mechanism in p97-cofactor formation[70]. It was also observed that there is no interaction between FAF1 and UN in the absence of p97, while both cofactors bind to p97 with a stoichiometry of 1:1:6. Therefore, binding of UN to p97 might induce an asymmetry conformation in the p97, resulting in one of the vacant N domains bound tightly to FAF1. To confirm the hierarchical cofactor binding theory, depletion of UN in Hela cells led to abolishing the co-immunoprecipitation of FAF1 with p97[71].
In addition to some of the substrate recruiting UBA-UBX cofactors, UN was found in complex with p97 and a substrate processing cofactor named Otu1(YOD1)[26,72]. Isothermal titration calorimetry (ITC) assay determined that the stoichiometry between Otu1 and p97/ UN ternary complex is 1:1 binding. Meanwhile, biochemical studies suggesting a 1:1 stoichiometry of UN heterodimer and p97 hexamer[14,29,73]. Therefore, one OTU1 cofactor is involved in the ubiquitinated substrate processing through interaction with one p97 either in the absence or presence of UN[72].
It was reported that Npl4 plays a major role as a target for some anticancer medicines like the bis-diethyldithiocarbamate-copper complex (CuET). It has been shown that CuET induces aggregation and immobilization of NPL4 cofactor, resulting in impairing the cellular protein degradation machinery of p97/Npl4 in tumor cells[74,75]. In addition, both in vitro and cell-based assays revealed that under oxidative conditions, CuET inhibits the substrate unfolding process of p97 in complex with UN by releasing cupric ions, leading to disrupting the seesaw motion of Npl4 by destroying its zinc finger motifs, and locking the necessary conformational switch between Npl4 and p97[76].
UN also in complex with p97 and via NUB1L participate in the degradation of NEDD8 that is a ubiquitin like protein attached to its substrate. NUB1L is an interferon-inducible protein that interacts with p97 and joins to the p97-UN complex bound to NEDD8. The hydrolysis of ATP in D1 and D2 domains alters the p97 conformation and transfers the NEDD8 from Npl4 to NUB1L. Finally, NUB1L guides NEDD8 to the proteasome system for degradation via interacting with Rpn10 and Rpn1[77].
4. Interaction between p97 and p47
p47 plays a major role in Golgi reassembly during the cell cycle by recruiting p97 [79]. The UBA and UBX domains of p47 are the binding site for ubiquitinated substrate and p97 N-domain, respectively (Figure 2). The UBX domain contains conserved interacting residues that are involved in hydrophobic interactions with residues Phe52, Ile70, Leu72, Tyr110, and Tyr143 of p97N domain[70]. p47 can affect the ATPase activity of p97 and lower concentration of p47 avoids the ATP from binding to D1 domain. This prevents ATPase activity of both D1 and D2, because the binding of ATP to D1 is necessary for hydrolysis activity of D2 domain[80]. However, at higher concentrations of p47, this protein promotes more ATP binding to D1, resulting in hydrolysis ATP by D2 domain[80]. Moreover, the result of binding affinity between p47 and p97 in the absence of nucleotide showed that there are two different equilibrium affinity constants (KD values), suggesting there may be two different binding sites within p47[29]. In addition, it was found that p47 binds 10-fold more strongly to p97 in ATP-bound state compared to that of in ADP-bound state[81]. Therefore, the interactions between p47 and p97 residues are related to the bound nucleotide state of p97. P47 also contains intrinsically disordered regions including SHP motif and SEP-interacting motif (SIM) that are critical for regulating the interaction between p47 and p97[82]. Biophysical studies showed that p47 interacts with p97 N domain through both SHP motif (SHPN-terminus-SHPC-terminus) and UBX domain, indicating UBX domain and SHP motif have various binding sites on p97 N domain. However, two SHP motifs might seem to compete for the same interacting site on p97 N domain. In addition, SIM motif forms intramolecular contacts with SEP domain in either isolated p47 or when it is in complex with p97. This intramolecular interaction may facilitate delivery of ubiquitinated substrate from p47 UBA domain to p97. Previously, it was claimed that p47 forms a high-affinity trimer that is so stable[79,80,83]. However, a combination of analytical ultra-centrifugation(AUC), SAXS, and DLS analysis showed that p47 is completely monomer and not trimer[82]. A model of p97-p47 complex obtained from cryo-Em map showed that p47 forms a complex with p97 in both apo or ATP states through UBX and SHPC-terminus which interacts with p97 N-domain. In addition, p47 SEP domain seems to interact with p97 D1 domain and localized in the center of p97 ND1. Moreover, SHPN-terminus interacts with the adjacent p97 N-domain, indicating interactions are related to the bound nucleotide state of p97. Therefore, p47 can interact with p97 apo/ATP either bipartite or tripartite, while the interaction between p47 and p97ADP bound state is bipartite (Figure 3).
5. Interaction between p97 and p37
p37 is the other p97ʼs cofactor that is involved in Golgi and ER biogenesis like p47 and plays a pivotal role in the maintenance of Golgi and ER during interphase and reassembly of them at the end of mitosis[84]. This cofactor contains SEP and UBX domains which have a highly homology with those of p47(Figure 2). In addition, both proteins need VCIP135 deubiquitinase for Golgi biogenesis[30]. However, p47 is more active in reassembly of organelles at the end of mitosis, while p37 is specialized for organelle maintenance during interphase and reassembly of them during mitosis. The structural differences between p37 and p47 is the lack of UBA domain in the N-terminus of p37, which this domain in p47 binds to monoubiquitinated substrate in the presence of p97, and also the absence of the amino acid region 69–92 found in p47 [60,80]. Nevertheless, the biochemical binding assay showed that p37 does not bind the ubiquitin protein in either the presence or absence of p97. In addition, unlike p97/p47 complex, adding a ubiquitin mutant had no effect on p97/p37-dependent Golgi reassembly in vitro[84]. The other differences is that although both p47 and p37 regulate the p97 ATPase activity via binding to the N-terminus of p97 and induce conformational changes, which results in regulating ADP/ATP binding to the D1 domain, p47 inhibits the ATPase activity of p97, while p37 increases the p97 ATPase activity[80]. The mechanisms of regulation are not yet understood and require a comparison between p97-p47 and p97-p37 complex at high resolution.
There are some conserved residues in the UBX domain of both cofactors that form hydrophobic interactions with p97 N domain residues including PHE52, PRO106, ILE 70, ARG 53, LEU 72 and TYR 110[85,86]. Moreover, the SEP domain in both p37 and p47 directly bind protein phosphatase 1 inhibitor 3 (I3), a ubiquitin independent substrate of p97, and extract it from SDS22–PP1–I3 complex (SPI) [82,87,88,89,90]. According to immunoprecipitation from cells and in vitro unfolding assay, p37 in complex with p97 recognize and bind an internal recognition site (IRS) within I3 substrate of the SPI complex[89]. The IRS peptide loop is then translocated into the p97 rings and threaded through the central pore, resulting in disassembly of the SDS22–PP1–I3 complex (Figure 4). The cryo-EM of p97-p37-SPI substrate complex revealed that PP1, along with its associates SDS22 and inhibitor-3 (I3), is firmly attached to p97. Surprisingly, this attachment occurs through direct contact between SDS22 and the p97 N-domain. p37 cofactor, which serves as a bridge connecting two neighboring p97 N-domains beneath the substrate complex facilitates the loading process. Subsequently, a segment of I3 is inserted into the central pore of the hexameric structure of p97 arranged in a spiral shape, while additional components of I3 stay linked to PP1, suggesting a mechanism where p97 facilitates disassembly through a hold-and-extract process[91].
6. Substrate processing by p97 in complex with cofactor
Recently a variety of in vitro experiments have been conducted on purified Cdc48 in complex with UN cofactor and a polyubiquitinated model substrate to understand the mechanistic function of substrate processing by Cdc48[13,14,37,92]. Bodnar et al showed that UN cofactor can decrease the overall Cdc48 ATPase activity in the absence of substrate. In addition, substrate containing K48-linked polyubiquitin chain can bind to the p97 through UN cofactor. Substrate binding to the p97 in complex with UN leads to decreasing the ATPase activity in the D1 domain, altering the position of N domain to up conformation and enhancing ATP hydrolysis in the D2 domain[13]. Releasing the energy due to converting ATP to ADP generates the pulling force in aromatic pore loop residues of the D2 ring that causes movement of substrate through the central pore and its unfolding. During the translocation of substrate, most of the polyubiquitin remains bound to the UN, while a few of the ubiquitin chain enter the p97 central pore along with the substrate. Moreover, the ATP hydrolysis of D1 domain is necessary for releasing the substrate that happens through the cooperation between the Cdc48 and a deubiquitinase(DUB) processing cofactor. At this stage, the position of N-domain changes to down conformation due to the ATPase activity of D1 domain and thereby DUB cofactor has more access to the polyubiquitin chain for its trimming. Finally, the remaining oligoubiquitin chain is translocated through the central pore of Cdc48 and become unfolded (Figure 5)[13]. The research findings have indicated that at least two polypeptide chain can place in the central pore, which is consistent with other hexameric ATPease in different organisms[39].
In the other study, the structure of the Cdc48 ATPase with UN in complex with polyubiquitin substrate was determined using single-particle cryo-EM analysis in the presence of either ATPγs or ADP [14]. According to the resolved structure, four ubiquitin molecules are bound to the Cdc48 in complex with UN heterodimer cofactor. Since the interface between Ufd1 and Npl4 is very small, it might seem that Npl4 is separated after binding the ubiquitylated substrate. However, Ufd1 might be able to keep its connection with Cdc48 because of two SHP motifs. The UN cofactor needs at least five ubiquitin proteins to interact with substrate [92]. One of the ubiquitin molecules binds UT3 domain of Ufd1, and the other one is in the gap between MPN domain of Npl4 and the ATPase domain of p97, which this interaction might seem to be an initiation site for translocating into the central pore. Moreover, further ubiquitin protein binds the Npl4 domains apart from UBXL domain. Therefore, Npl4 binds directly to the ATPase ring of p97 and likely serves as a universal gatekeeper for all ubiquitin proteins which require translocation through the central pore of Cdc48 [14]. Cdc48 in complex with UN cofactor extracts polyubiquitilated and misfolded substrates from membrane and cellular organelles via a pulling force. The two ATPase domains (D1 and D2) play distinct roles, causing substrate unfolding and releasing via translocation into the central pore of Cdc48. Furthermore, the cooperation between Cdc48 and a deubiquitinase is required for the releasing substrate through the trimming the polyubiquitin chain to oligoubiquitin substrate[13]. A cryo–EM structure of the Cdc48 in complex with UN and polyubiquitinated substrate showed that substrate processing is started by unfolding a ubiquitin molecule. According to the structure, two ubiquitin proteins are at the top of Npl4, and surprisingly the further ubiquitin bound to a conserved groove of Npl4 is unfolded and inserted into the Cdc48 central pore[16]. Intriguingly, the unfolding process of this ubiquitin occurs through the interaction of its N-terminal domain with both Npl4 groove as well as Cdc48 and does not require ATP hydrolysis. The insertion of ubiquitin N-terminus into the central pore leads to engagement of the D2 pore loop residues and pulling on the N-terminal segment through ATP hydrolysis. The subunits of D2 ring from A to F form a staircase shape through engaging and disengaging with ATP and drag the polypeptide by interacting with every peptide attached to the substrate. In other words, the power stemmed from ATP binding and hydrolyzing in a rotary way in D2 subunits causes branch point movement through the central pore and unfolds the attached Ub(1). In addition, as a result of pulling on the proximal ubiquitin chain, Ub(-1) becomes unfolded. Finally, the substrate goes through the ATPase rings and become unfolded. It has also been mentioned releasing substrate from the Cdc48 complex needs cleavage of the distal ubiquitins Ub(2) -Ub(4) by a DUB cofactor.
Pan et al worked on human p97 to determine its interaction with UN cofactor and compare it with the resolved structure of Cdc48 complexed with UN[76]. Human Npl4 contains an additional C-terminal zinc finger motif in compared to that of fungal homologue[93]. The further differences is that fungal Npl4 binds to the D1 ring of Cdc48 by both zinc finger motifs, while human Npl4 binds to the D1 ring by just one zinc finger motif[76]. Three different conformational states of human p97 in complex with Npl4 were resolved in both the absence and presence of the polyubiquitinated substrate. The state I&II revealed that Npl4 interacts with the D1 ring of p97 through its zinc finger motif 1. However, the main difference between these two states is that the MPN and CTD domains of Npl4 rotate by ~ 8°and displace by ~13 A° with ZF1. In state III, Npl4 binds to the groove of the D1 domain through zinc finger 2. The zinc finger 3 was not observed to be involved in the interaction with D1 domain, but it was close to one of the p97 N domains. Meanwhile, another three major conformational states were resolved in the presence of polyubiquitinated substrate. Intriguingly, both state I and II of substrate-engaged p97 in complex with Npl4 were the same as those of structures without substrate. However, in substrate-engaged state III, the density of a single ubiquitin was observed on top of Npl4. The cryo-EM structure of substrate-engaged Cdc48 in complex with UN showed that a single unfolded ubiquitin binds in the groove of fungal Npl4 and then goes through the central pore of Cdc48, while none of these three substrate-engaged states of p97 showed the ubiquitin density in the groove of Npl4[14,16,76]. This might suggest that the resolved structures were in advance of ubiquitin unfolding and the substrate translocating. Therefore, since the unfolding of a single ubiquitin did not take place in the resolved structures of human p97 in complex with UN and polyubiquitin substrates, there might be a different mechanism between human 97 and fungal Cdc48. In addition, the structure of p97 complexes revealed a seesaw motion of Npl4 on top of the D1 ring which may initiate unfolding of the substrate. They also suggested that the reason of seesaw motion of Npl4 might be due to the interaction of one zinc finger motif with the p97 N domains. Therefore, the seesaw motion of Npl4 may occur when N-domain undergoes conformational changes upon ATP hydrolysis in the D1 and D2. In the further study by Pan et al, they worked on human p97 to determine its substrate processing in cooperation with UN cofactor. A series of cryo-EM structures of substrate-engaged p97 in complex with UN was reported in several working states. The obtained structures revealed that both the D1 and D2 domains undergo a specific conformational change upon substrate processing in their rings named power stroke motion[94]. This motion seems to be a unique feature for human p97, as it has not been reported for the other p97 homologues. In addition, it has been mentioned that the D2 ring has more ATPase activity compared to D1 ring and provides the force for substrate translocation through the backbone hydrogen bonds[22,94]. Therefore, the pore loops of D2 ring might seem to play a more significant role in interacting with the substrate. On the other hand, both structures of CDC48 and p97 showed fewer interactions between pore loops of D1 ring and translocating peptide[16]. Furthermore, it has been shown that Npl4 cofactor might play an important role in starting the translocation of substrate through unfolding the ubiquitin chain and guided it into the central pore by a seesaw motion. In the next step, the peptide, which is now in thread form, is captured by the pore loops in D1 and D2 rings. Following interaction with the substrate, both D1 and D2 rings undergo conformational change and get compressed to spiral forms. During the sequential ATP hydrolysis in D1 and D2 domains that provides the power stroke motions in their rings, the peptide is translocated downward into the central pore of p97 from the N-terminus to the C-terminus. The direction of substrate translocation is similar to that of Cdc48[15,16]. According to the solved structure, the density of the translocating substrate in D2 ring is much better than it is in D1 ring, which implies a stronger interaction with residues in pore loop-I of the D2 domain including Met550, Trp551, Phe552. These residues interact with the substrate like a pitching grip. Two hydrogen bonds are formed between one residue of translocating peptide, and both Met 550 and Phe 552 residues in each pore loop 1. Therefore, 12 residues of pore loop-I in D2 domain are involved in the hydrogen interaction with every other residue of translocating peptide. The pore loop-I residues in the D2 create a spiral in the open state in which chains F and E are at the bottom and up, respectively. However, in the closed state, chain F is separated from the translocating substrate. Moreover, the density of pore loop-II in D2 ring (Chains A to E) were resolved but at lower resolution. Meanwhile, pore loop-II residues are not engaged in hydrogen bonds with residues of translocating peptide, indicating their less important role in substrate translocation. The substrate unfolding assay indicated that mutation of pore loop-1 residues in D2 ring abolish p97’s unfoldase activity, while mutation of pore loop-II residues showed no effect. Pore loop-I residues in D1 ring bind to the translocating peptide in the same way as that of D2 ring. However, only four chains (B-E) are involved in the interaction with residues Lys277, Leu278 and Ala279(Figure 6). Moreover, two pore loop-II in D1 ring (Chains C and D) are engaged in the interaction with the translocating peptide by creating a hydrogen bond through His317.
The design of small molecules that interfere with the binding and assembly of cofactors is an attractive alternative to develop cofactor-specific inhibitors. The small molecule drugs can directly and uniquely bind to p97 and inhibit mutant p97’s activity. However, the lack of complete high-resolution structures of p97-cofactors impairs development of cofactor-specific inhibitors. Ji et al explained how the Cdc48 unfolds one ubiquitin, and translocates polypeptides, and how it releases its substrates[95]. According to in vivo, in vitro experiments and cryo-EM structure, the Cdc48 in complex with UN cooperatively recognize and bind the polyubiquitin chains with little or no specificity for the attached substrate. In agreement with the study of Twomey et al, they mentioned the interaction between Cdc48-UN complex and polyubiquitin chains initiate without the need of ATP hydrolysis, which leads to the unfolding of one of the ubiquitin molecules. The initiator ubiquitin needs to be succeeded into the polyubiquitin chain in which two folded ubiquitin are bound to top of Npl4 and two to the UT3 domain of Ufd1. It was also mentioned N-terminus of unfolded ubiquitin is separated from the rest of ubiquitin and captured by its insertion into the Cdc48 D1 ring[95,96]. The separated N-domain is engaged into the D2 pore loops and pulled through the central pore by ATP hydrolysis[16,95]. Dislodging the segment of ubiquitin which originally bound to the Npl4 groove leads to releasing folded Ub1 and Ub2 from Npl4. It was also seen that when the branch of initiator ubiquitin reaches the D1 pore, the C-terminus of initiator ubiquitin is translocated through the central pore but not the distal ubiquitin (Figure 7). This selectivity might be because Cdc48 uses less energy for translocation the unfolded C-terminal segment rather than the folded Ub1. Therefore, Ub1 and distal ubiquitins remain folded and place outside of the central pore of Cdc48. Translocating the initiator ubiquitin resulting in pulling the next branchpoint which is linked to the C-terminus of imitator ubiquitin. Since Cdc48 sequentially translocates all ubiquitin proteins located between the initiator and substrate, it might unfold both N- and C- terminus. There are two hypotheses about the number of polyubiquitin chain which insert into the central pore. The first one is that, as Cdc48 D2 pore loops surround one single polyubiquitin chain, the ATPase might translocate just this single branch and unfold the entire substrate in multiple rounds. However, according to the models of Vps4 and ClpB ATPases, two strands can be translocated at the same time[97,98]. Therefore, the second hypotheses might be that p97 or Cdc48 can pull several branches through their central pore stimulatingly. The entire unfolded polypeptide containing proximal ubiquitins and substrate is released from the central pore, while the distal ubiquitins seems to move outside of the ATPase. At the end, deubiquitinase cofactor (Otu1) might trim the unfolded ubiquitin chain to prevent the unfolded polyubiquitinated substrate re-binding to the ATPase complex. In addition, it was indicated that the substrate carrying an Otu1 is released from the complex of Cdc48 and UN which does not rebind. However, the previous study mentioned Otu1 may release the unfolded substrate from the complex by trimming all distal ubiquitins leading to substrate translocation through the central pore[13].
The side view of substrate engaged Cdc48 in complex with NPL4 based on resolved structure of Cdc48 in association with polyubiquitinated substrate ((PDB:1UBQ, PDB: 60A9)). The magenta sphere shows unfolded initiator ubiquitin. The proximal and distal ubiquitin proteins are indicated as orange and brown ubiquitin molecules, respectively. Translocated, unfolded ubiquitin proteins and substrate are illustrated as spirals.
The Cdc48 has a 6-fold rotational symmetry (C6) in the absence of substrates [7,14,17]. Cooney et al designed a co-immunoprecipitation (Co-IP) assay for purifying Cdc48 and native substrate through the substrate recruiting adaptor-shp1(homolog of human p47) to determine the different structural states of Cdc48 with and without substrate[15]. The Cdc48 complex structure in the absence of ADP⋅BeFx, which is a non-hydrolysable nucleotide analog, displayed a rotational symmetric with no density for substrate. However, the further complex in the presence of ADP⋅BeFx displayed an asymmetric configuration with a density for a substrate. According to the cryo-EM model of substrate engaged Cdc48 in complex with shp1,
several interactions between UBX domain of shp1 and Cdc48 N domain are observed, including UBX residues Lys 359, Val 417, Asn 415, Ser 416, Val 417, and Gly 358, and Cdc48 N residues Lys 119, Cly 64, Phe 62, Tyr 120, Lys 119, Asp 65, and Arg 63. Intriguingly all the Cdc48 N domain residues, which are involved in the interaction with residues of Shp1 UBX domain, are conserved in human p97 N domain and participate in the interaction with UBX residues. In addition, there are some interacting residues such as Val 417, Asn 415, and Gly 358 which are conserved in the UBX domain of both yeast Shp1 and human p47. The cryo-EM model also showed that the native substrate passes through the central pore of both the D1 and D2 rings (Figure 8A). In this model, the subunits A to E of both D1 and D2 domains are connected by 60o rotation that form a helical symmetry. However, the subunit F, which occupies two positions and form two conformations, is displaced from substrate. In addition, the interfaces of subunits AB, BC, CD and DE are stabilized through binding of ATP to the active site of A, B, C and D, respectively. This pattern provides a condition for substrate to interact in a β-strand conformation with Cdc48 pore. Hydrolysis of ATP at the D active site leads to decreasing the DE interface and allowing subunit E to move to the transitioning F state. The whole process is repeated by allowing the interface to exchange ADP to ATP and translocating A to the previous position. Therefore, the mechanism of hand over hand was suggested for substrate engagement and translocation through the central pore of Cdc48 in the presence of ATP analog and Shp1 adaptor[15]. According to both biochemical and resolved structure of Cdc48-Shp1-substrate, aromatic residues in D2 domain (W561and Y562) are more important for substrate binding and translocation compared to flexible or small D1 residues(M288 and A289) which bind substrate less tightly(Figure 8B)[13,15,37]. Furthermore, the less strong density of substrate has been observed in the D1 domain, indicating that the D2 domain is more significant for substrate translocation than D1 domain[13,37]. Moreover, the engaged residues of D2 domain are aromatic and seem fit for substrate binding. On the other hand, the D1 domain contains flexible and small residues that probably bind substrate less tightly and may cause slippage[99]. The hand over hand mechanism of substrate translocation was also observed in the other AAA+ ATPases including VAT, Vps4, proteasome, YME1, NSF, Hsp104 and ClpB, suggesting the same mechanism in these other AAA+ ATPases[15,42,43,97,100,101,102,103,104,105,106,107,108,109,110]. In addition to Cdc48-Shp1 complex associated with native substrate, the model of substrate-engaged p97 in complex with p47 showed an asymmetric reconstruction in which five of the six p97 monomers are in a helical arrangement. In contrast to subunit F, the five well-ordered and helical subunits (A-E) are involved in the subunit translocation (Figure 8C)[111]. Meanwhile in agreement with substrate/Cdc48 complex model, the substrate-engaged p97 subunits (A–E) are connected to each other by an ~60° rotation. In addition, both Pore loop 1 and 2 residues from D1 and D2 participate in substrate binding. These tight interaction with substrate formed by the pore loop 1 residues including Lys 277, Leu278 and Ala279 from D1 domain and Trp551-Phe552 from D2 domain (Figure 8D). The density of substrate in D1 is weaker than that of D2 domain, indicating a weaker interaction with the residues located in D1 pore loops which is consistent with the study of Cooney et al. On the other hand, the interaction between the aromatic residues in D2 and the side chain residues of substrate create stronger binding. Intriguingly, key residues in substrate binding and translocation are mainly conserved among both p97 and Cdc48. The K277, L278, and A279 pore loop 1 residues in the p97 D1, and K287, M288, as well as A289 in the D1 ring of Cdc48, are all important for substrate translocation. These residues interact with the substrate and help to move it through the central pore. Similarly, the aromatic residues M550, W551, and F552 in the p97 D2 Pore loop-I, and the conserved residues W561 and Y562 in the Cdc48 D2 Pore loop-I, form a hydrophobic pocket that can interact with hydrophobic residues on the substrate, allowing it to be pulled into the central pore.
The interaction of both pore loops I and II with the substrate residues explain how p97 and other AAA unfoldases could bind and translocate many various protein substrates[41,111].
Arie et al hypothesized that p97, in conjunction with its cofactor UN, undergoes specific structural transitions that regulate substrate processing and that these transitions are linked to the ATPase cycle of p97. The aim of the study was to resolve the structural changes involved in the early stages of UN binding and substrate processing. This study identified mutants that stabilize intermediate conformations without impairing p97’s activity, allowing the capture of discrete states involved in cofactor binding[112]. Specifically, the p97 F539A mutant was shown to stabilize a conformation where two D1 protomers are ATP-bound while the others remain in the ADP state. This intermediate conformation likely facilitates interaction with UN and represents a key state during the p97 catalytic cycle. In addition, the study demonstrated that the presence of UN induces a conformational change in p97’s N-terminal domain (NTD) to a configuration where five of the NTDs are in the ATP-bound "up" state. This conformational shift was shown to prevent nucleotide hydrolysis until substrate processing by the D2 ring is complete. This hierarchical control between the D1 and D2 rings enhances coordination in substrate handling, with D1 regulating substrate recruitment and D2 mediating translocation and unfolding. One limitation of this study was the absence of a specific p97 substrate to track the various stages of substrate processing and unfolding. The p97 F539A mutant, which behaves similarly to a "cold-sensitive" variant, becomes fully active only at higher temperatures. This characteristic could help capture distinct steps in the substrate unfolding process if analyzed through cryo-EM at lower temperatures using a substrate model. Another limitation was the inability to assign a specific density to Ufd1 within the p97-UN complex, despite attempts to do so.
Further study by Oppenheim, et al provided new structural insights into the interactions within the Cdc48-Npl4-Ufd1 (UN) ternary complex, which is critical for protein quality control. By combining crosslinking mass spectrometry (XL-MS) with integrative modelling, the researchers mapped interactions between Npl4, Ufd1, and Cdc48[113]. Their key hypothesis focused on identifying how specific residues stabilize this complex, particularly Cys115, a highly conserved cysteine at the Cdc48-Npl4 binding interface. Mutation of Cys115 to serine disrupts the interaction between Cdc48 and Npl4, leading to impaired complex stability, reduced cell growth, and altered protein degradation. Their findings highlight that Cdc48's N-terminal domain (NTD) serves as a multi-partner domain for binding various cofactors, including the UN complex. They demonstrate that Cdc48's NTD stabilizes the complex, primarily interacting with Npl4, while Ufd1 remains flexible. The study also suggests that Cdc48's preference for Npl4-binding may play a role in its diverse interactions with other cofactors. Mutations affecting the Cdc48-Npl4 interface have broader implications for protein degradation, cellular growth, and potential therapeutic targets for disrupting Cdc48 activity.
7. Conclusion and Future Directions
Human p97 is a critical therapeutic target for treatment of cancer and neurodegenerative diseases. The study of substrate engagement mechanism, translocation and unfolding by p97 in complex with cofactor clarify how uncontrolled cell division or missense mutations in human p97 affect substrate processing functions of the complex.
In recent years, many studies have carried out to unravel the molecular mechanism of substrate processing by ATPase complex. However, there are still some major questions that need to be considered. For instance, it is not clear whether p97 act upstream of shuttling factors and proteasome and deliver unfolded substrate to them, or p97 act both upstream and downstream of the proteasome. In addition, the impact of cofactors on p97’s conformation and function remains poorly understood. Meanwhile, the role of substrate-processing cofactors to release the unfolded protein after processing by p97 remains still unanswered. Although some structures of substrate-engaged p97 in complex with cofactor have been resolved, it is still unclear what the role of unresolved(flexible) domains of cofactors is in the substrate processing mechanism. It is still unclear how each specific cofactor recruits p97 in a distinct cellular function. To address these important questions, there is a need to combine structural biology methods with structural dynamics techniques. The hybrid structural studies provide clear insight into the atomic-level structure of substrate-engaged p97 in complex with cofactor, and conformational change of the complex upon protein-protein interaction. This structural information allows to understand function of both p97 and its cofactors in more details to develop drugs against disease-relevant functional property of p97 which target cofactor association.
Funding
This research was funded by the National Health and Medical Council (NHMRC) Ideas APP2000934 to I.R., and the start-up package of the University of Melbourne to I.R., Melbourne Research Scholarship to A.M.
Acknowledgments
We acknowledge the support of the ANR (ANR-19-CE11-0008-01 to SJ), The University of Melbourne (start-up fund to IR, Melbourne Research Scholarship to AM).
Declaration of Interests: The authors declare no competing interests.
References
- Chen, B., Retzlaff, M., Roos, T. & Frydman, J. (2011) Cellular strategies of protein quality control, Cold Spring Harbor perspectives in biology. 3, a004374. [CrossRef]
- Chou, T.-F., Brown, S. J., Minond, D., Nordin, B. E., Li, K., Jones, A. C., Chase, P., Porubsky, P. R., Stoltz, B. M. & Schoenen, F. J. (2011) Reversible inhibitor of p97, DBeQ, impairs both ubiquitin-dependent and autophagic protein clearance pathways, Proceedings of the National Academy of Sciences. 108, 4834-4839. [CrossRef]
- van den Boom, J. & Meyer, H. (2018) VCP/p97-mediated unfolding as a principle in protein homeostasis and signaling, Molecular cell. 69, 182-194. [CrossRef]
- Olszewski, M. M., Williams, C., Dong, K. C. & Martin, A. (2019) The Cdc48 unfoldase prepares well-folded protein substrates for degradation by the 26S proteasome, Communications biology. 2, 1-8. [CrossRef]
- Rouiller, I., DeLaBarre, B., May, A. P., Weis, W. I., Brunger, A. T., Milligan, R. A. & Wilson-Kubalek, E. M. (2002) Conformational changes of the multifunction p97 AAA ATPase during its ATPase cycle, Nature structural biology. 9, 950-957. [CrossRef]
- Brunger, A. T. & DeLaBarre, B. (2003) NSF and p97/VCP: similar at first, different at last, FEBS letters. 555, 126-133.
- Huyton, T., Pye, V. E., Briggs, L. C., Flynn, T. C., Beuron, F., Kondo, H., Ma, J., Zhang, X. & Freemont, P. S. (2003) The crystal structure of murine p97/VCP at 3.6 Å, Journal of structural biology. 144, 337-348.
- Wendler, P., Ciniawsky, S., Kock, M. & Kube, S. (2012) Structure and function of the AAA+ nucleotide binding pocket, Biochimica et Biophysica Acta (BBA)-Molecular Cell Research. 1823, 2-14. [CrossRef]
- Hänzelmann, P. & Schindelin, H. (2016) Structural basis of ATP hydrolysis and intersubunit signaling in the AAA+ ATPase p97, Structure. 24, 127-139. [CrossRef]
- Buchberger, A., Schindelin, H. & Hänzelmann, P. (2015) Control of p97 function by cofactor binding, FEBS letters. 589, 2578-2589. [CrossRef]
- DeLaBarre, B. & Brunger, A. T. (2005) Nucleotide dependent motion and mechanism of action of p97/VCP, Journal of molecular biology. 347, 437-452.
- Abaan, O. D., Hendriks, W., Üren, A., Toretsky, J. A. & Erkizan, H. V. (2013) Valosin containing protein (VCP/p97) is a novel substrate for the protein tyrosine phosphatase PTPL1, Experimental cell research. 319, 1-11.
- Bodnar, N. O. & Rapoport, T. A. (2017) Molecular mechanism of substrate processing by the Cdc48 ATPase complex, Cell. 169, 722-735. e9. [CrossRef]
- Bodnar, N. O., Kim, K. H., Ji, Z., Wales, T. E., Svetlov, V., Nudler, E., Engen, J. R., Walz, T. & Rapoport, T. A. (2018) Structure of the Cdc48 ATPase with its ubiquitin-binding cofactor Ufd1–Npl4, Nature structural & molecular biology. 25, 616-622.
- Cooney, I., Han, H., Stewart, M. G., Carson, R. H., Hansen, D. T., Iwasa, J. H., Price, J. C., Hill, C. P. & Shen, P. S. (2019) Structure of the Cdc48 segregase in the act of unfolding an authentic substrate, Science. 365, 502-505. [CrossRef]
- Twomey, E. C., Ji, Z., Wales, T. E., Bodnar, N. O., Ficarro, S. B., Marto, J. A., Engen, J. R. & Rapoport, T. A. (2019) Substrate processing by the Cdc48 ATPase complex is initiated by ubiquitin unfolding, Science. 365. [CrossRef]
- Banerjee, S., Bartesaghi, A., Merk, A., Rao, P., Bulfer, S. L., Yan, Y., Green, N., Mroczkowski, B., Neitz, R. J. & Wipf, P. (2016) 2.3 Å resolution cryo-EM structure of human p97 and mechanism of allosteric inhibition, Science. 351, 871-875. [CrossRef]
- Schuller, J. M., Beck, F., Lössl, P., Heck, A. J. & Förster, F. (2016) Nucleotide-dependent conformational changes of the AAA+ ATPase p97 revisited, FEBS letters. 590, 595-604.
- Tang, W. K., Li, D., Li, C. c., Esser, L., Dai, R., Guo, L. & Xia, D. (2010) A novel ATP-dependent conformation in p97 N–D1 fragment revealed by crystal structures of disease-related mutants, The EMBO journal. 29, 2217-2229.
- Briggs, L. C., Baldwin, G. S., Miyata, N., Kondo, H., Zhang, X. & Freemont, P. S. (2008) Analysis of nucleotide binding to P97 reveals the properties of a tandem AAA hexameric ATPase, Journal of Biological Chemistry. 283, 13745-13752. [CrossRef]
- Chou, T.-F., Bulfer, S. L., Weihl, C. C., Li, K., Lis, L. G., Walters, M. A., Schoenen, F. J., Lin, H. J., Deshaies, R. J. & Arkin, M. R. (2014) Specific inhibition of p97/VCP ATPase and kinetic analysis demonstrate interaction between D1 and D2 ATPase domains, J Mol Biol. 426, 2886-2899.
- Song, C., Wang, Q. & Li, C.-C. H. (2003) ATPase activity of p97-valosin-containing protein (VCP): D2 mediates the major enzyme activity, and D1 contributes to the heat-induced activity, Journal of Biological Chemistry. 278, 3648-3655.
- Ye, Y., Tang, W. K., Zhang, T. & Xia, D. (2017) A mighty “protein extractor” of the cell: structure and function of the p97/CDC48 ATPase, Frontiers in molecular biosciences. 4, 39.
- Erzberger, J. P. & Berger, J. M. (2006) Evolutionary relationships and structural mechanisms of AAA+ proteins, Annu Rev Biophys Biomol Struct. 35, 93-114.
- Bruderer, R. M., Brasseur, C. & Meyer, H. H. (2004) The AAA ATPase p97/VCP interacts with its alternative co-factors, Ufd1-Npl4 and p47, through a common bipartite binding mechanism, Journal of Biological Chemistry. 279, 49609-49616.
- Ernst, R., Mueller, B., Ploegh, H. L. & Schlieker, C. (2009) The otubain YOD1 is a deubiquitinating enzyme that associates with p97 to facilitate protein dislocation from the ER, Molecular cell. 36, 28-38. [CrossRef]
- Uchiyama, K., Jokitalo, E., Kano, F., Murata, M., Zhang, X., Canas, B., Newman, R., Rabouille, C., Pappin, D. & Freemont, P. (2002) VCIP135, a novel essential factor for p97/p47-mediated membrane fusion, is required for Golgi and ER assembly in vivo, The Journal of cell biology. 159, 855-866.
- Hänzelmann, P. & Schindelin, H. (2017) The interplay of cofactor interactions and post-translational modifications in the regulation of the AAA+ ATPase p97, Frontiers in molecular biosciences. 4, 21. [CrossRef]
- Hänzelmann, P. & Schindelin, H. (2016) Characterization of an additional binding surface on the p97 N-terminal domain involved in bipartite cofactor interactions, Structure. 24, 140-147. [CrossRef]
- Alexandru, G., Graumann, J., Smith, G. T., Kolawa, N. J., Fang, R. & Deshaies, R. J. (2008) UBXD7 binds multiple ubiquitin ligases and implicates p97 in HIF1α turnover, Cell. 134, 804-816. [CrossRef]
- Schuberth, C. & Buchberger, A. (2008) UBX domain proteins: major regulators of the AAA ATPase Cdc48/p97, Cellular and molecular life sciences. 65, 2360-2371.
- Kloppsteck, P., Ewens, C. A., Förster, A., Zhang, X. & Freemont, P. S. (2012) Regulation of p97 in the ubiquitin–proteasome system by the UBX protein-family, Biochimica et Biophysica Acta (BBA)-Molecular Cell Research. 1823, 125-129. [CrossRef]
- Liu, C., Liu, W., Ye, Y. & Li, W. (2017) Ufd2p synthesizes branched ubiquitin chains to promote the degradation of substrates modified with atypical chains, Nature communications. 8, 1-15.
- Parker, C. E., Mocanu, V., Warren, M. R., Greer, S. F. & Borchers, C. H. (2005) Mass spectrometric determination of protein ubiquitination in Ubiquitin-Proteasome Protocols pp. 153-173, Springer.
- Kirkpatrick, D. S., Hathaway, N. A., Hanna, J., Elsasser, S., Rush, J., Finley, D., King, R. W. & Gygi, S. P. (2006) Quantitative analysis of in vitro ubiquitinated cyclin B1 reveals complex chain topology, Nature cell biology. 8, 700-710. [CrossRef]
- Ye, Y., Meyer, H. H. & Rapoport, T. A. (2003) Function of the p97–Ufd1–Npl4 complex in retrotranslocation from the ER to the cytosol: dual recognition of nonubiquitinated polypeptide segments and polyubiquitin chains, The Journal of cell biology. 162, 71-84.
- Blythe, E. E., Olson, K. C., Chau, V. & Deshaies, R. J. (2017) Ubiquitin-and ATP-dependent unfoldase activity of P97/VCP• NPLOC4• UFD1L is enhanced by a mutation that causes multisystem proteinopathy, Proceedings of the National Academy of Sciences. 114, E4380-E4388.
- Bodnar, N. & Rapoport, T. (2017) Toward an understanding of the Cdc48/p97 ATPase, F1000Research. 6.
- Lee, C., Prakash, S. & Matouschek, A. (2002) Concurrent translocation of multiple polypeptide chains through the proteasomal degradation channel, Journal of Biological Chemistry. 277, 34760-34765. [CrossRef]
- Burton, R. E., Siddiqui, S. M., Kim, Y.-I., Baker, T. A. & Sauer, R. T. (2001) Effects of protein stability and structure on substrate processing by the ClpXP unfolding and degradation machine, The EMBO journal. 20, 3092-3100. [CrossRef]
- Han, H. & Hill, C. P. (2019) Structure and mechanism of the ESCRT pathway AAA+ ATPase Vps4, Biochemical Society Transactions. 47, 37-45. [CrossRef]
- Ripstein, Z. A., Huang, R., Augustyniak, R., Kay, L. E. & Rubinstein, J. L. (2017) Structure of a AAA+ unfoldase in the process of unfolding substrate, Elife. 6, e25754.
- Puchades, C., Rampello, A. J., Shin, M., Giuliano, C. J., Wiseman, R. L., Glynn, S. E. & Lander, G. C. (2017) Structure of the mitochondrial inner membrane AAA+ protease YME1 gives insight into substrate processing, Science. 358, eaao0464. [CrossRef]
- Zhao, M., Wu, S., Zhou, Q., Vivona, S., Cipriano, D. J., Cheng, Y. & Brunger, A. T. (2015) Mechanistic insights into the recycling machine of the SNARE complex, Nature. 518, 61-67. [CrossRef]
- Tang, W. K. & Xia, D. (2013) Altered intersubunit communication is the molecular basis for functional defects of pathogenic p97 mutants, Journal of Biological Chemistry. 288, 36624-36635. [CrossRef]
- Yu, G., Bai, Y., Li, K., Amarasinghe, O., Jiang, W. & Zhang, Z.-Y. (2021) Cryo-electron microscopy structures of VCP/p97 reveal a new mechanism of oligomerization regulation, Iscience. 24, 103310. [CrossRef]
- Hoq, M. R., Vago, F. S., Li, K., Kovaliov, M., Nicholas, R. J., Huryn, D. M., Wipf, P., Jiang, W. & Thompson, D. H. (2021) Affinity Capture of p97 with Small-Molecule Ligand Bait Reveals a 3.6 Å Double-Hexamer Cryoelectron Microscopy Structure, ACS nano. 15, 8376-8385.
- Mori-Konya, C., Kato, N., Maeda, R., Yasuda, K., Higashimae, N., Noguchi, M., Koike, M., Kimura, Y., Ohizumi, H. & Hori, S. (2009) p97/valosin-containing protein (VCP) is highly modulated by phosphorylation and acetylation, Genes to Cells. 14, 483-497.
- Tang, W. K., Odzorig, T., Jin, W. & Xia, D. (2019) Structural basis of p97 inhibition by the site-selective anticancer compound CB-5083, Molecular pharmacology. 95, 286-293. [CrossRef]
- Nandi, P., Li, S., Columbres, R. C. A., Wang, F., Williams, D. R., Poh, Y.-P., Chou, T.-F. & Chiu, P.-L. (2021) Structural and functional analysis of disease-linked p97 ATPase mutant complexes, International journal of molecular sciences. 22, 8079. [CrossRef]
- Gao, H., Li, F., Ji, Z., Shi, Z., Li, Y. & Yu, H. (2022) Cryo-EM structures of human p97 double hexamer capture potentiated ATPase-competent state, Cell Discovery. 8, 1-13.
- Liu, S., Ye, X., Liu, W., Liu, L., Li, D., Lin, Q. & Wang, T. (2022) Cryo-EM structure of dodecamer human p97 in complex with NMS-873 reveals S765-G779 peptide plays critical role for D2 ring oligomerization, Biochemical and Biophysical Research Communications.
- Sato, Y., Tsuchiya, H., Yamagata, A., Okatsu, K., Tanaka, K., Saeki, Y. & Fukai, S. (2019) Structural insights into ubiquitin recognition and Ufd1 interaction of Npl4, Nature communications. 10, 1-13.
- Heo, J.-M., Livnat-Levanon, N., Taylor, E. B., Jones, K. T., Dephoure, N., Ring, J., Xie, J., Brodsky, J. L., Madeo, F. & Gygi, S. P. (2010) A stress-responsive system for mitochondrial protein degradation, Molecular cell. 40, 465-480. [CrossRef]
- Wu, X., Li, L. & Jiang, H. (2016) Doa1 targets ubiquitinated substrates for mitochondria-associated degradation, Journal of Cell Biology. 213, 49-63.
- Moreno, S. P., Bailey, R., Campion, N., Herron, S. & Gambus, A. (2014) Polyubiquitylation drives replisome disassembly at the termination of DNA replication, Science. 346, 477-481. [CrossRef]
- Maric, M., Mukherjee, P., Tatham, M. H., Hay, R. & Labib, K. (2017) Ufd1-Npl4 recruit Cdc48 for disassembly of ubiquitylated CMG helicase at the end of chromosome replication, Cell reports. 18, 3033-3042. [CrossRef]
- Maric, M., Maculins, T., De Piccoli, G. & Labib, K. (2014) Cdc48 and a ubiquitin ligase drive disassembly of the CMG helicase at the end of DNA replication, Science. 346. [CrossRef]
- Ye, Y., Tang, W. K., Zhang, T. & Xia, D. (2017) A Mighty “Protein Extractor” of the Cell: Structure and Function of the p97/CDC48 ATPase, Frontiers in Molecular Biosciences. 4.
- Meyer, H. H., Wang, Y. & Warren, G. (2002) Direct binding of ubiquitin conjugates by the mammalian p97 adaptor complexes, p47 and Ufd1–Npl4, The EMBO Journal. 21, 5645-5652.
- Meyer, H.-J. & Rape, M. (2014) Enhanced protein degradation by branched ubiquitin chains, Cell. 157, 910-921.
- Park, S., Isaacson, R., Kim, H. T., Silver, P. A. & Wagner, G. (2005) Ufd1 exhibits the AAA-ATPase fold with two distinct ubiquitin interaction sites, Structure. 13, 995-1005. [CrossRef]
- Rumpf, S. & Jentsch, S. (2006) Functional division of substrate processing cofactors of the ubiquitin-selective Cdc48 chaperone, Molecular cell. 21, 261-269. [CrossRef]
- Hetzer, M., Meyer, H. H., Walther, T. C., Bilbao-Cortes, D., Warren, G. & Mattaj, I. W. (2001) Distinct AAA-ATPase p97 complexes function in discrete steps of nuclear assembly, Nature cell biology. 3, 1086-1091. [CrossRef]
- Isaacson, R. L., Pye, V. E., Simpson, P., Meyer, H. H., Zhang, X., Freemont, P. S. & Matthews, S. (2007) Detailed structural insights into the p97-Npl4-Ufd1 interface, Journal of Biological Chemistry. 282, 21361-21369. [CrossRef]
- Blythe, E. E., Gates, S. N., Deshaies, R. J. & Martin, A. (2019) Multisystem proteinopathy mutations in VCP/p97 increase NPLOC4· UFD1L binding and substrate processing, Structure. 27, 1820-1829. e4. [CrossRef]
- Kimura, Y., Fukushi, J., Hori, S., Matsuda, N., Okatsu, K., Kakiyama, Y., Kawawaki, J., Kakizuka, A. & Tanaka, K. (2013) Different dynamic movements of wild-type and pathogenic VCP s and their cofactors to damaged mitochondria in a P arkin-mediated mitochondrial quality control system, Genes to Cells. 18, 1131-1143.
- Pan, M., Zheng, Q., Yu, Y., Ai, H., Xie, Y., Zeng, X., Wang, C., Liu, L. & Zhao, M. (2020) Seesaw Conformations of Npl4 in the Human p97 Complex and the Inhibitory Mechanism of a Disulfiram Derivative, bioRxiv.
- Schuberth, C. & Buchberger, A. (2005) Membrane-bound Ubx2 recruits Cdc48 to ubiquitin ligases and their substrates to ensure efficient ER-associated protein degradation, Nature cell biology. 7, 999-1006. [CrossRef]
- Hänzelmann, P., Buchberger, A. & Schindelin, H. (2011) Hierarchical binding of cofactors to the AAA ATPase p97, Structure. 19, 833-843. [CrossRef]
- Lee, J.-J., Park, J. K., Jeong, J., Jeon, H., Yoon, J.-B., Kim, E. E. & Lee, K.-J. (2013) Complex of Fas-associated factor 1 (FAF1) with valosin-containing protein (VCP)-Npl4-Ufd1 and polyubiquitinated proteins promotes endoplasmic reticulum-associated degradation (ERAD), Journal of Biological Chemistry. 288, 6998-7011. [CrossRef]
- Kim, S. J., Cho, J., Song, E. J., Kim, S. J., Kim, H. M., Lee, K. E., Suh, S. W. & Kim, E. E. (2014) Structural basis for ovarian tumor domain-containing protein 1 (OTU1) binding to p97/valosin-containing protein (VCP), Journal of Biological Chemistry. 289, 12264-12274.
- Pye, V. E., Beuron, F., Keetch, C. A., McKeown, C., Robinson, C. V., Meyer, H. H., Zhang, X. & Freemont, P. S. (2007) Structural insights into the p97-Ufd1-Npl4 complex, Proceedings of the National Academy of Sciences. 104, 467-472.
- Ramadan, K. (2012) p97/VCP-and Lys48-linked polyubiquitination form a new signaling pathway in DNA damage response, Cell cycle. 11, 1062-1069.
- Majera, D., Skrott, Z., Chroma, K., Merchut-Maya, J. M., Mistrik, M. & Bartek, J. (2020) Targeting the NPL4 adaptor of p97/VCP segregase by disulfiram as an emerging cancer vulnerability evokes replication stress and DNA damage while silencing the ATR pathway, Cells. 9, 469.
- Pan, M., Zheng, Q., Yu, Y., Ai, H., Xie, Y., Zeng, X., Wang, C., Liu, L. & Zhao, M. (2021) Seesaw conformations of Npl4 in the human p97 complex and the inhibitory mechanism of a disulfiram derivative, Nature communications. 12, 1-12.
- Liu, S., Yang, H., Zhao, J., Zhang, Y.-H., Song, A.-X. & Hu, H.-Y. (2013) NEDD8 ultimate buster-1 long (NUB1L) protein promotes transfer of NEDD8 to proteasome for degradation through the P97UFD1/NPL4 complex, Journal of Biological Chemistry. 288, 31339-31349.
- Jumper, J., Evans, R., Pritzel, A., Green, T., Figurnov, M., Ronneberger, O., Tunyasuvunakool, K., Bates, R., Žídek, A. & Potapenko, A. (2021) Highly accurate protein structure prediction with AlphaFold, Nature. 596, 583-589.
- Kondo, H., Rabouille, C., Newman, R., Levine, T. P., Pappin, D., Freemont, P. & Warren, G. (1997) p47 is a cofactor for p97-mediated membrane fusion, Nature. 388, 75-78.
- Zhang, X., Gui, L., Zhang, X., Bulfer, S. L., Sanghez, V., Wong, D. E., Lee, Y., Lehmann, L., Lee, J. S. & Shih, P.-Y. (2015) Altered cofactor regulation with disease-associated p97/VCP mutations, Proceedings of the National Academy of Sciences. 112, E1705-E1714.
- Bulfer, S. L., Chou, T.-F. & Arkin, M. R. (2016) P97 disease mutations modulate nucleotide-induced conformation to alter protein–protein interactions, ACS chemical biology. 11, 2112-2116.
- Conicella, A. E., Huang, R., Ripstein, Z. A., Nguyen, A., Wang, E., Löhr, T., Schuck, P., Vendruscolo, M., Rubinstein, J. L. & Kay, L. E. (2020) An intrinsically disordered motif regulates the interaction between the p47 adaptor and the p97 AAA+ ATPase, Proceedings of the National Academy of Sciences. 117, 26226-26236.
- Beuron, F., Dreveny, I., Yuan, X., Pye, V. E., Mckeown, C., Briggs, L. C., Cliff, M. J., Kaneko, Y., Wallis, R. & Isaacson, R. L. (2006) Conformational changes in the AAA ATPase p97–p47 adaptor complex, The EMBO journal. 25, 1967-1976.
- Uchiyama, K., Totsukawa, G., Puhka, M., Kaneko, Y., Jokitalo, E., Dreveny, I., Beuron, F., Zhang, X., Freemont, P. & Kondo, H. (2006) p37 is a p97 adaptor required for Golgi and ER biogenesis in interphase and at the end of mitosis, Developmental cell. 11, 803-816.
- Mirzadeh, A., Kobakhidze, G., Vuillemot, R., Jonic, S. & Rouiller, I. (2022) In silico prediction, Characterization, Docking studies and Molecular dynamics simulation of human p97 in complex with p37 cofactor.
- Dreveny, I., Kondo, H., Uchiyama, K., Shaw, A., Zhang, X. & Freemont, P. S. (2004) Structural basis of the interaction between the AAA ATPase p97/VCP and its adaptor protein p47, The EMBO journal. 23, 1030-1039.
- Soukenik, M., Diehl, A., Leidert, M., Sievert, V., Büssow, K., Leitner, D., Labudde, D., Ball, L. J., Lechner, A. & Nägler, D. K. (2004) The SEP domain of p47 acts as a reversible competitive inhibitor of cathepsin L, FEBS letters. 576, 358-362.
- Weith, M., Seiler, J., van den Boom, J., Kracht, M., Hülsmann, J., Primorac, I., del Pino Garcia, J., Kaschani, F., Kaiser, M. & Musacchio, A. (2018) Ubiquitin-independent disassembly by a p97 AAA-ATPase complex drives PP1 holoenzyme formation, Molecular cell. 72, 766-777. e6.
- van den Boom, J., Kueck, A. F., Kravic, B., Müschenborn, H., Giesing, M., Pan, D., Kaschani, F., Kaiser, M., Musacchio, A. & Meyer, H. (2021) Targeted substrate loop insertion by VCP/p97 during PP1 complex disassembly, Nature Structural & Molecular Biology. 28, 964-971.
- Buchberger, A. (2022) Unfolding by Cdc48/p97: different strokes for different folks, Trends in Cell Biology.
- Van den Boom, J., Marini, G., Meyer, H. & Saibil, H. R. (2023) Structural basis of ubiquitin-independent PP1 complex disassembly by p97, The EMBO Journal. 42, e113110.
- Twomey, E. C., Ji, Z., Wales, T. E., Bodnar, N. O., Ficarro, S. B., Marto, J. A., Engen, J. R. & Rapoport, T. A. (2019) Substrate processing by the Cdc48 ATPase complex is initiated by ubiquitin unfolding, Science. 365, eaax1033.
- Alam, S. L., Sun, J., Payne, M., Welch, B. D., Blake, B. K., Davis, D. R., Meyer, H. H., Emr, S. D. & Sundquist, W. I. (2004) Ubiquitin interactions of NZF zinc fingers, The EMBO journal. 23, 1411-1421.
- Pan, M., Yu, Y., Ai, H., Zheng, Q., Xie, Y., Liu, L. & Zhao, M. (2021) Mechanistic insight into substrate processing and allosteric inhibition of human p97, bioRxiv.
- Ji, Z., Li, H., Peterle, D., Paulo, J. A., Ficarro, S. B., Wales, T. E., Marto, J. A., Gygi, S. P., Engen, J. R. & Rapoport, T. A. (2021) Translocation of polyubiquitinated protein substrates by the hexameric Cdc48 ATPase, Molecular Cell.
- Irbäck, A., Mitternacht, S. & Mohanty, S. (2005) Dissecting the mechanical unfolding of ubiquitin, Proceedings of the National Academy of Sciences. 102, 13427-13432.
- Han, H., Fulcher, J. M., Dandey, V. P., Iwasa, J. H., Sundquist, W. I., Kay, M. S., Shen, P. S. & Hill, C. P. (2019) Structure of Vps4 with circular peptides and implications for translocation of two polypeptide chains by AAA+ ATPases, Elife. 8, e44071.
- Avellaneda, M. J., Franke, K. B., Sunderlikova, V., Bukau, B., Mogk, A. & Tans, S. J. (2020) Processive extrusion of polypeptide loops by a Hsp100 disaggregase, Nature. 578, 317-320.
- Esaki, M., Islam, M. T., Tani, N. & Ogura, T. (2017) Deviation of the typical AAA substrate-threading pore prevents fatal protein degradation in yeast Cdc48, Scientific reports. 7, 1-11.
- Monroe, N., Han, H., Shen, P. S., Sundquist, W. I. & Hill, C. P. (2017) Structural basis of protein translocation by the Vps4-Vta1 AAA ATPase, Elife. 6, e24487.
- Puchades, C., Rampello, A. J., Shin, M., Giuliano, C. J., Wiseman, R. L., Glynn, S. E. & Lander, G. C. (2017) Structure of the mitochondrial inner membrane AAA+ protease YME1 gives insight into substrate processing, Science. 358.
- Andres, H., Goodall, E. A., Gates, S. N., Lander, G. C. & Martin, A. (2018) Substrate-engaged 26S proteasome structures reveal mechanisms for ATP-hydrolysis–driven translocation, Science. 362.
- Huang, R., Ripstein, Z. A., Rubinstein, J. L. & Kay, L. E. (2019) Cooperative subunit dynamics modulate p97 function, Proceedings of the National Academy of Sciences. 116, 158-167.
- Han, H., Monroe, N., Sundquist, W. I., Shen, P. S. & Hill, C. P. (2017) The AAA ATPase Vps4 binds ESCRT-III substrates through a repeating array of dipeptide-binding pockets, Elife. 6, e31324.
- de la Peña, A. H., Goodall, E. A., Gates, S. N., Lander, G. C. & Martin, A. (2018) Substrate-engaged 26 S proteasome structures reveal mechanisms for ATP-hydrolysis–driven translocation, Science. 362, eaav0725.
- Dong, Y., Zhang, S., Wu, Z., Li, X., Wang, W. L., Zhu, Y., Stoilova-McPhie, S., Lu, Y., Finley, D. & Mao, Y. (2019) Cryo-EM structures and dynamics of substrate-engaged human 26S proteasome, Nature. 565, 49-55.
- Gates, S. N., Yokom, A. L., Lin, J., Jackrel, M. E., Rizo, A. N., Kendsersky, N. M., Buell, C. E., Sweeny, E. A., Mack, K. L. & Chuang, E. (2017) Ratchet-like polypeptide translocation mechanism of the AAA+ disaggregase Hsp104, Science. 357, 273-279.
- Deville, C., Carroni, M., Franke, K. B., Topf, M., Bukau, B., Mogk, A. & Saibil, H. R. (2017) Structural pathway of regulated substrate transfer and threading through an Hsp100 disaggregase, Science advances. 3, e1701726.
- Yu, H., Lupoli, T. J., Kovach, A., Meng, X., Zhao, G., Nathan, C. F. & Li, H. (2018) ATP hydrolysis-coupled peptide translocation mechanism of Mycobacterium tuberculosis ClpB, Proceedings of the National Academy of Sciences. 115, E9560-E9569.
- White, K. I., Zhao, M., Choi, U. B., Pfuetzner, R. A. & Brunger, A. T. (2018) Structural principles of SNARE complex recognition by the AAA+ protein NSF, Elife. 7, e38888.
- Xu, Y., Han, H., Cooney, I., Guo, Y., Moran, N. G., Zuniga, N. R., Price, J. C., Hill, C. P. & Shen, P. S. (2022) Active conformation of the p97-p47 unfoldase complex, Nature communications. 13, 1-8.
- Arie, M., Matzov, D., Karmona, R., Szenkier, N., Stanhill, A. & Navon, A. (2024) A non-symmetrical p97 conformation initiates a multistep recruitment of Ufd1/Npl4, iScience. 27.
- Oppenheim, T., Radzinski, M., Braitbard, M., Brielle, E. S., Yogev, O., Goldberger, E., Yesharim, Y., Ravid, T., Schneidman-Duhovny, D. & Reichmann, D. (2023) The Cdc48 N-terminal domain has a molecular switch that mediates the Npl4-Ufd1-Cdc48 complex formation, Structure. 31, 764-779. e8. [CrossRef]
Figure 1.
p97 structure and different conformational states. A) Primary structure of p97. B-D) Various conformation of p97 in the presence of different nucleotide and substrate. E) Substrate recruiting and processing cofactors.
Figure 1.
p97 structure and different conformational states. A) Primary structure of p97. B-D) Various conformation of p97 in the presence of different nucleotide and substrate. E) Substrate recruiting and processing cofactors.
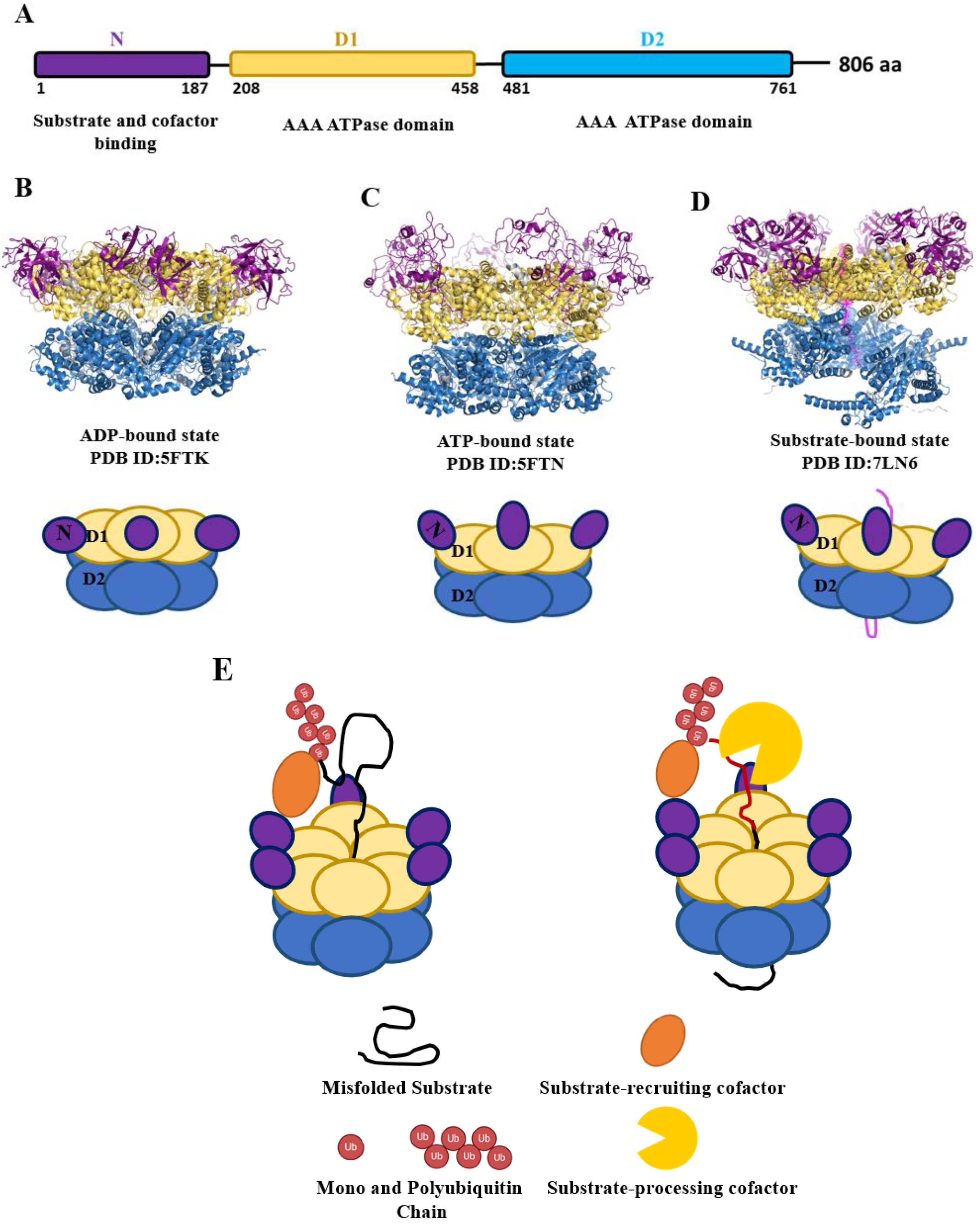
Figure 2.
Main p97’s cofactors. The domain architecture and predicted cofactors’ structures by alphafold[78].
Figure 2.
Main p97’s cofactors. The domain architecture and predicted cofactors’ structures by alphafold[78].
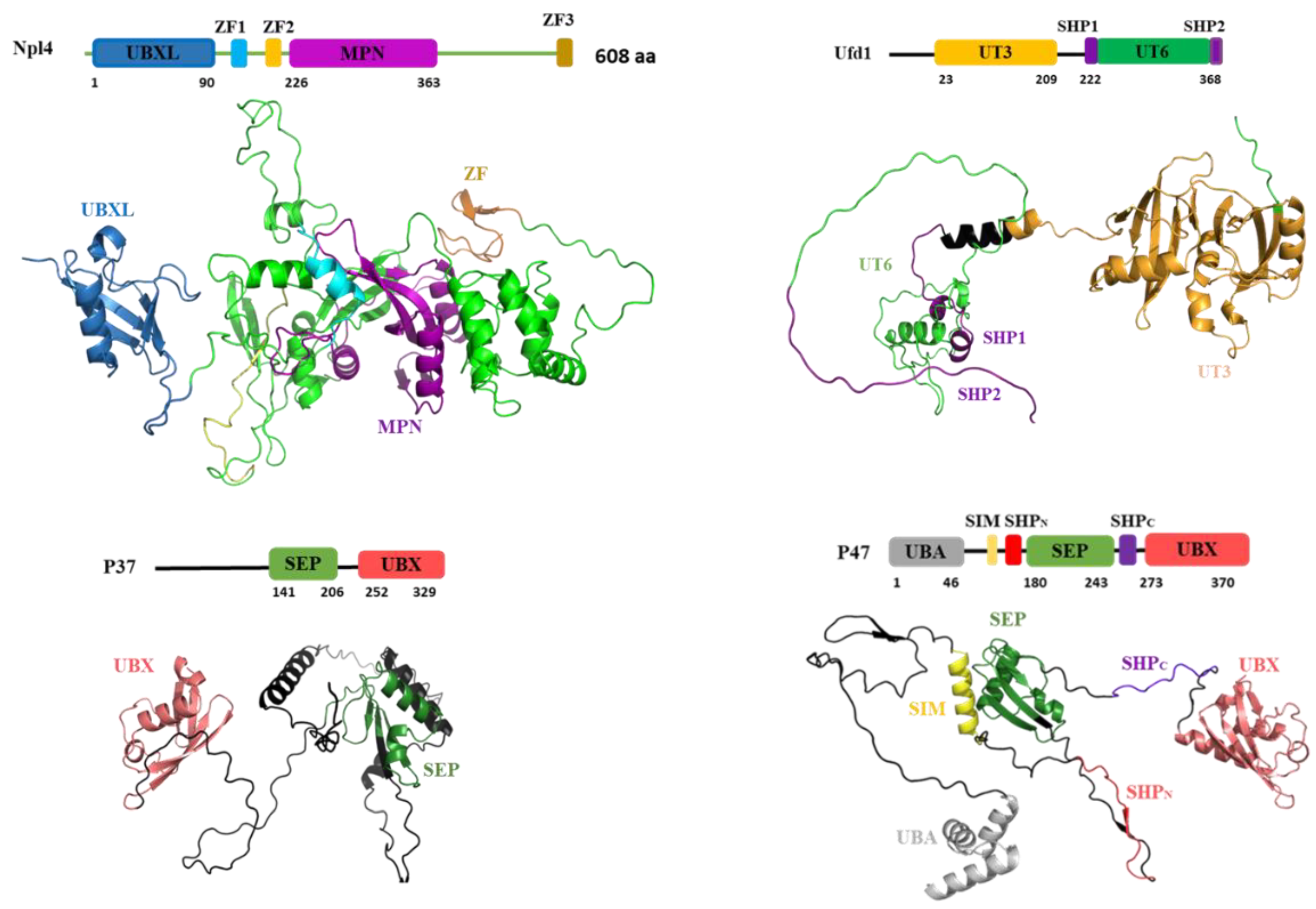
Figure 3.
A schematic model of p47-p97 complex in ATP and ADP bound state. A) p47–p97 complex in ATP-bound state (N domain in up conformation). B) p47–p97 complex in ADP-bound state (N domain coplanar with D1).
Figure 3.
A schematic model of p47-p97 complex in ATP and ADP bound state. A) p47–p97 complex in ATP-bound state (N domain in up conformation). B) p47–p97 complex in ADP-bound state (N domain coplanar with D1).
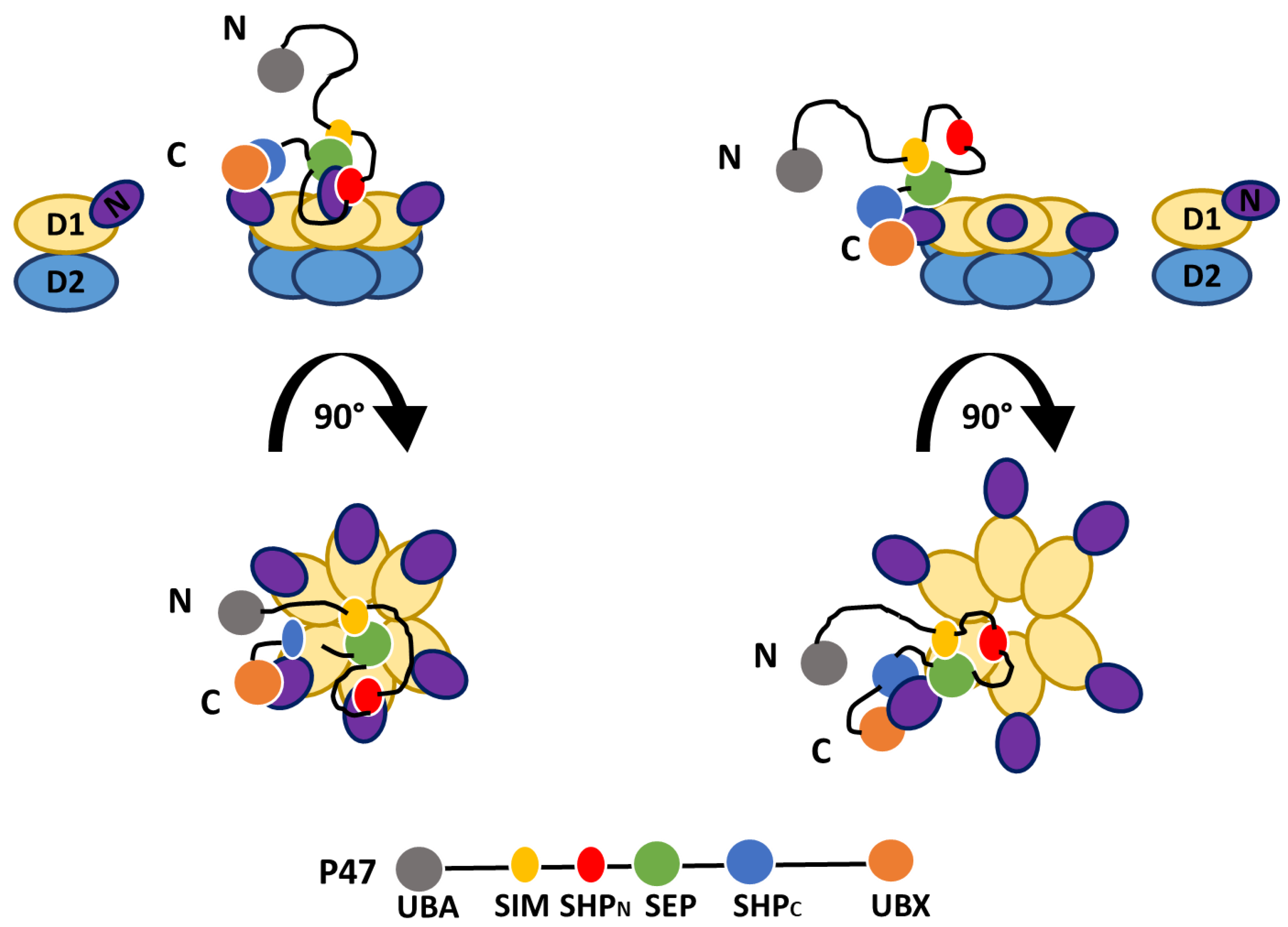
Figure 4.
A schematic model of I3 processing by p97 in complex with p37. The p97 interacts with SDS22-PP1-I3 complex through p37 cofactor. I3 is inserted into the p97 rings as peptide loop. The threading of I3 initiates the disassembly of the SDS22-PP1-I3 complex, allowing for the subsequent assembly of active Protein Phosphatase 1 (PP1) complexes.
Figure 4.
A schematic model of I3 processing by p97 in complex with p37. The p97 interacts with SDS22-PP1-I3 complex through p37 cofactor. I3 is inserted into the p97 rings as peptide loop. The threading of I3 initiates the disassembly of the SDS22-PP1-I3 complex, allowing for the subsequent assembly of active Protein Phosphatase 1 (PP1) complexes.
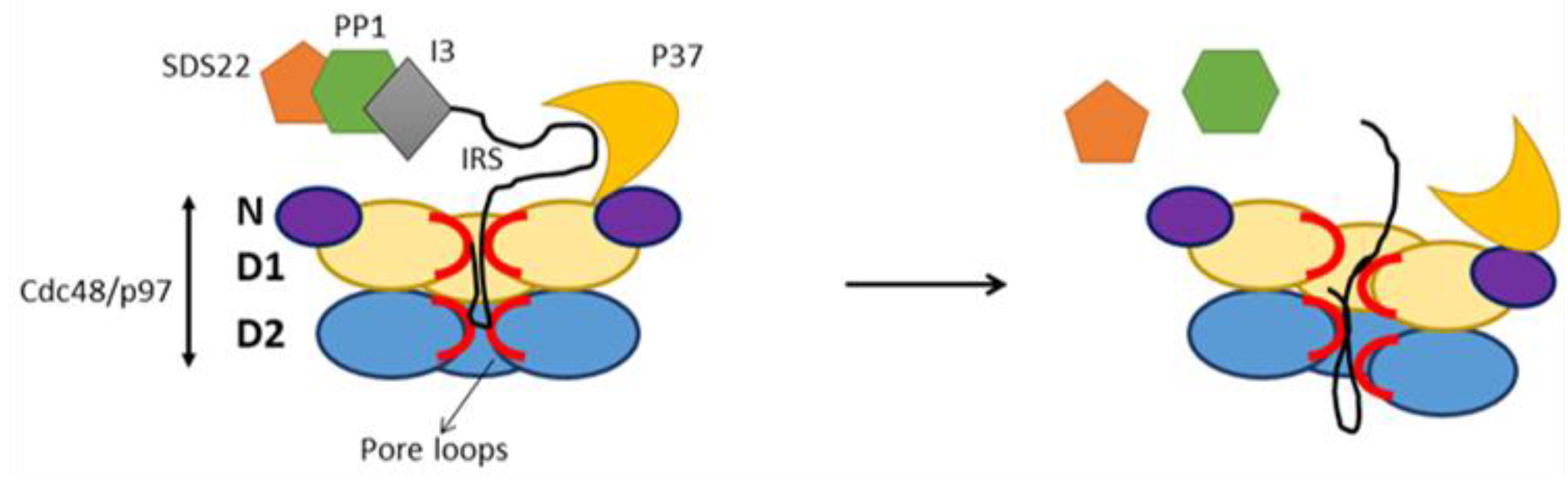
Figure 5.
Substrate processing by the Cdc48 in complex with UN and Otu1. A Schematic model of substrate translocation, unfolding and releasing pathway by the Cdc48 in complex with UN and Otu1.
Figure 5.
Substrate processing by the Cdc48 in complex with UN and Otu1. A Schematic model of substrate translocation, unfolding and releasing pathway by the Cdc48 in complex with UN and Otu1.
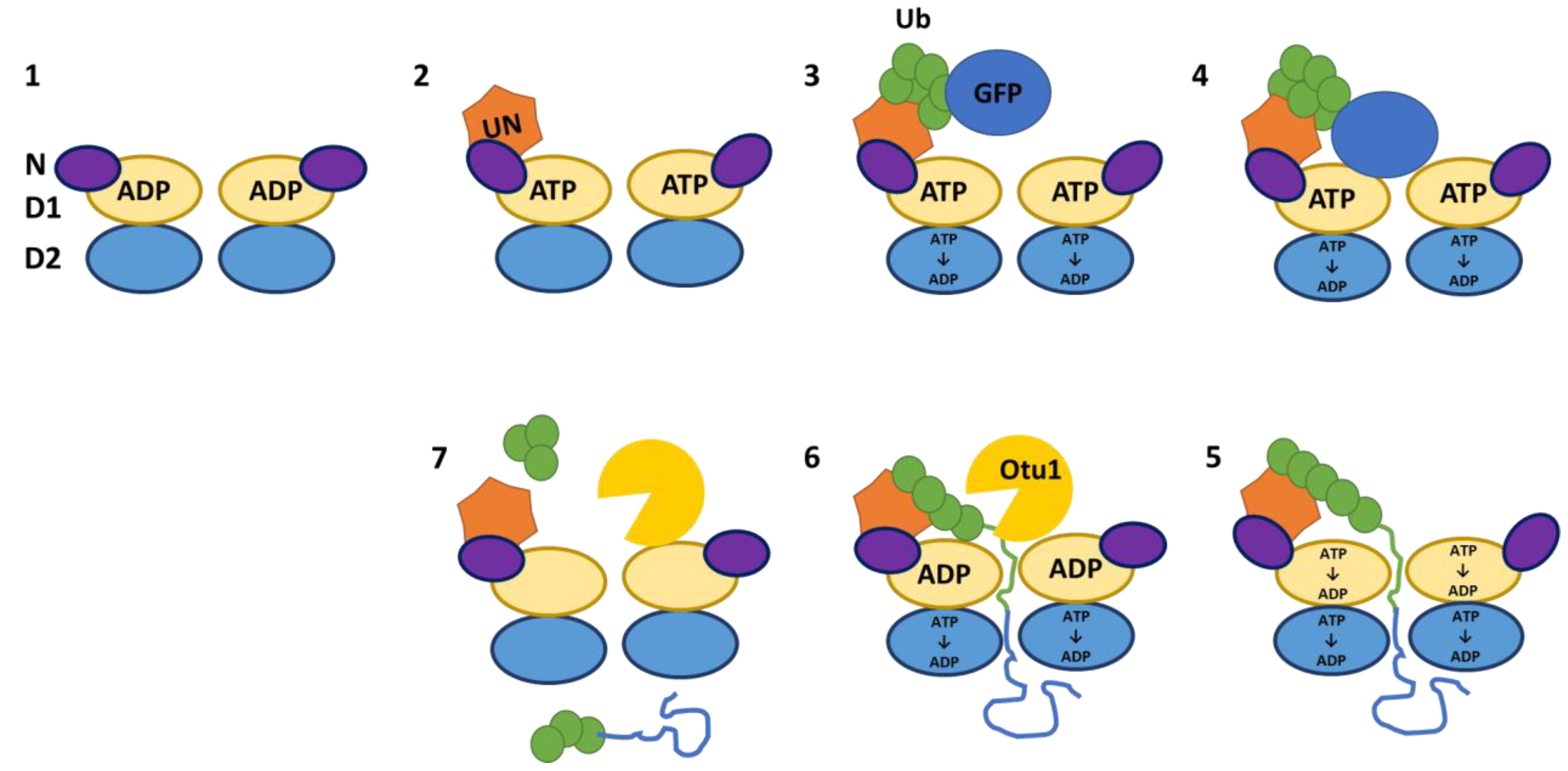
Figure 6.
Substrate processing by complex of human p97 and UN. A) The side-view of resolved structure of substrate-engaged p97 in association with Npl4(PDB: 7LN6). B) Interactions between the pore loops and translocating peptide. Interacting residues of pore loop I in D1 and D2 rings were colored in rainbow order from red to blue (residues K277, L278 and A279 from chain A are not involved in the interaction with translocating peptide). Interacting residues of pore loop II in D1 ring are colored in gray.
Figure 6.
Substrate processing by complex of human p97 and UN. A) The side-view of resolved structure of substrate-engaged p97 in association with Npl4(PDB: 7LN6). B) Interactions between the pore loops and translocating peptide. Interacting residues of pore loop I in D1 and D2 rings were colored in rainbow order from red to blue (residues K277, L278 and A279 from chain A are not involved in the interaction with translocating peptide). Interacting residues of pore loop II in D1 ring are colored in gray.
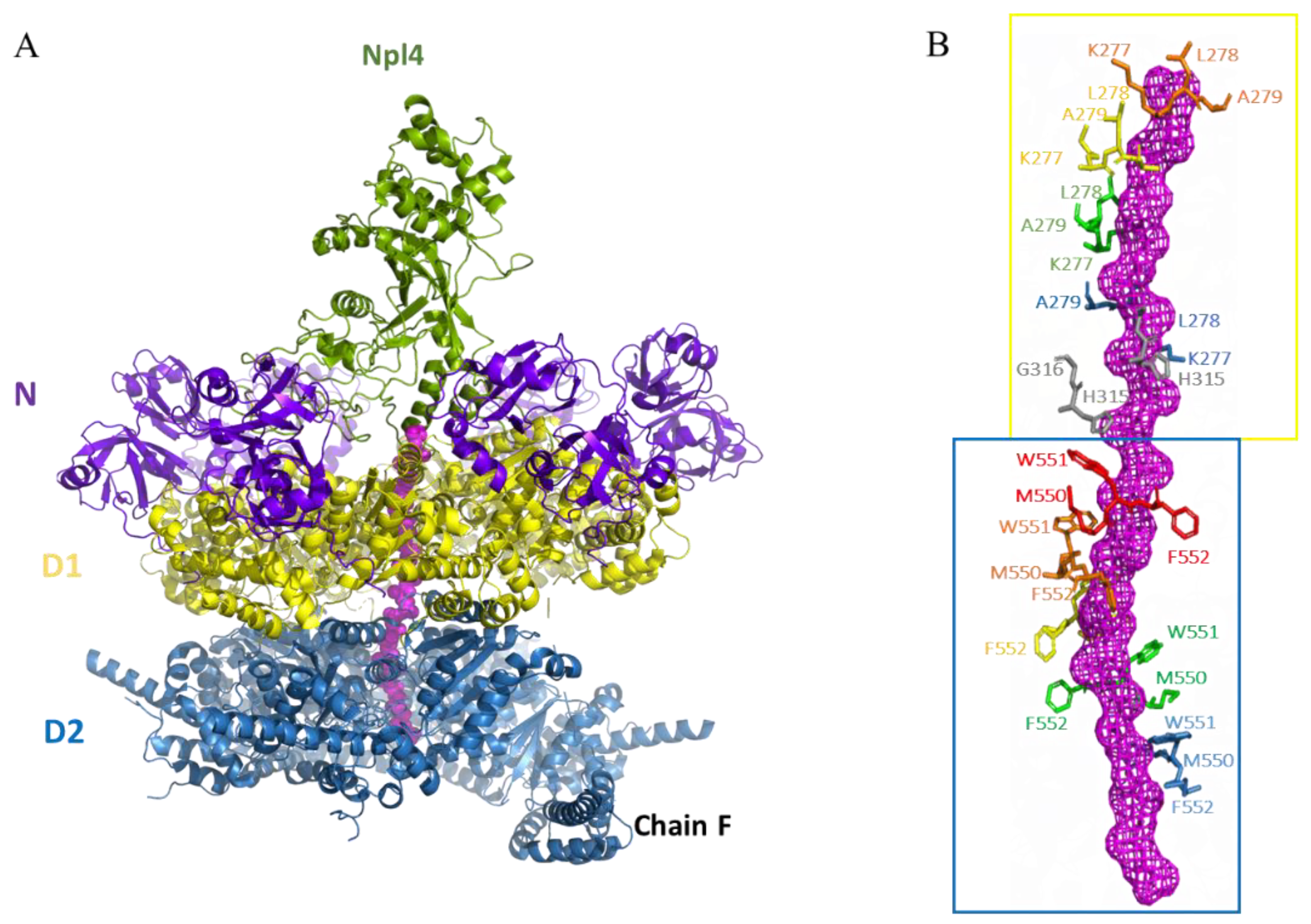
Figure 7.
Model of substrate translocation and unfolding steps by the complex of Cdc48 and UN.
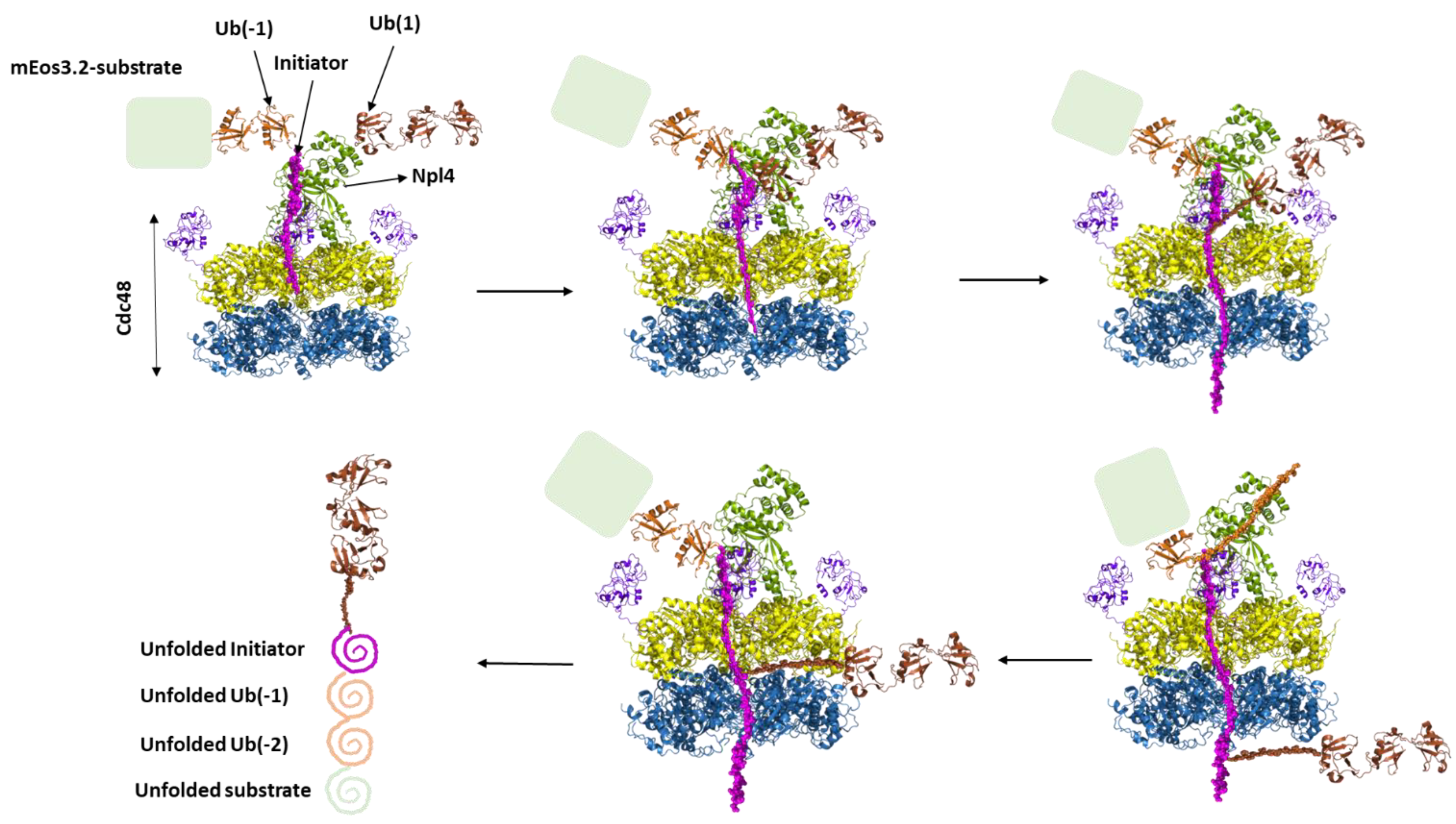
Figure 8.
Cryo-EM models of the Cdc48-Shp1-subtrate complex (PDB: 6OPC) and substrate-engaged p97 in complex with p47 UBX (PDB: 7MHS). A) side view of Cdc48 monomer complexed with Shp1-UBX and substrate in ribbon and spheres representations. B) Side view of translocating substrate (magenta spheres) and surrounding pore loop residues (Interacting residues were colored in rainbow order from chain A(red) to chain E (blue). C) Side view of a p97 protomer (subunit D shown) with the associated p47 UBX domain and engaged substrate(magenta). D) translocating substrate surrounded by interacting residues (Interacting residues were colored in rainbow order from chain A(red) to chain E (blue).
Figure 8.
Cryo-EM models of the Cdc48-Shp1-subtrate complex (PDB: 6OPC) and substrate-engaged p97 in complex with p47 UBX (PDB: 7MHS). A) side view of Cdc48 monomer complexed with Shp1-UBX and substrate in ribbon and spheres representations. B) Side view of translocating substrate (magenta spheres) and surrounding pore loop residues (Interacting residues were colored in rainbow order from chain A(red) to chain E (blue). C) Side view of a p97 protomer (subunit D shown) with the associated p47 UBX domain and engaged substrate(magenta). D) translocating substrate surrounded by interacting residues (Interacting residues were colored in rainbow order from chain A(red) to chain E (blue).
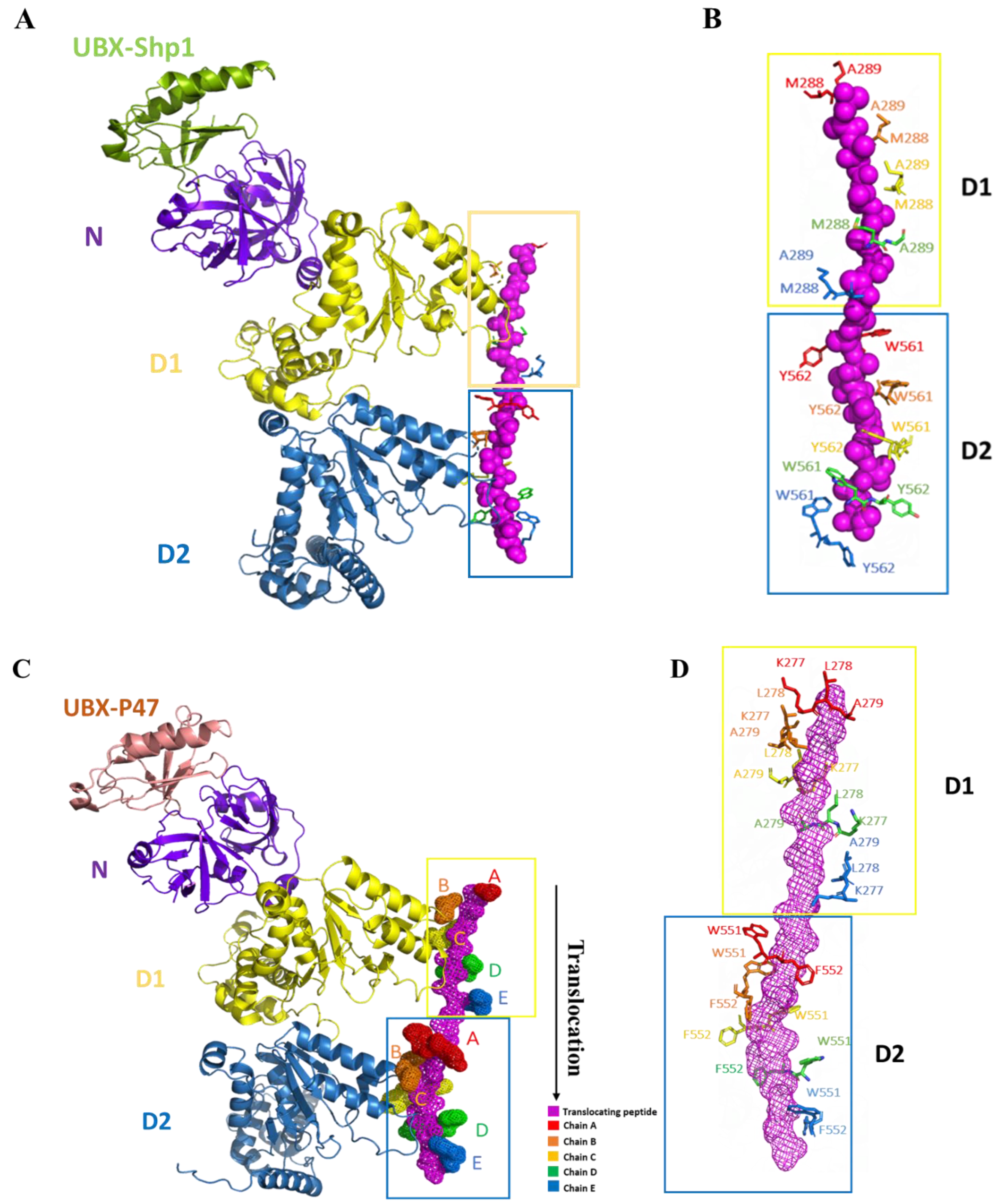

Table 1.
Details of the p97 dodecamer structures p97–main cofactor complexes.
| Reference | N-Domain in up conformation | N-Domain in coplanar conformation | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| D1 Domain Occupancy | D2 Domain Occupancy | Cryo-EM Map Resolution (A°) | EMDB ID | PDB ID | D1 Domain Occupancy | D2 Domain Occupancy | Cryo-EM Map Resolution (A°) | EMDB ID | PDB ID | |
| Gao et al. (2022) | ATP | ATP | 3.3 | EMD-31894 | 7VCS | ATP | ADP | 3.1 | EMD-31896 | 7VCU |
| G. Yu et al. (2021) | Empty | Empty | 3.9 | EMD-22675 | 7K56 | - | - | - | - | - |
| G. Yu et al. (2021) | Empty | Empty | 3.7 | EMD-22675 (at the presence of apyrase) | 7K57 (at the presence of apyrase) | - | - | - | - | - |
| P. Nandi (2021) | Empty | Empty | 3.3 | EMD-23191 | 7L5W | - | - | - | - | - |
| R. Hoq et al. (2021) | Empty | Empty | 3.6 | NA | NA | - | - | - | - | - |
| S. Liu, X. Ye, W. Liu et al. (2022) | - | - | - | - | - | ADP | NMS-873 | 3 | EMD-32827 | 7WUB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



