
Submitted:
14 September 2024
Posted:
17 September 2024
You are already at the latest version
Abstract
Primary themes in intergenerational justice are a healthy environment, the perpetuation of Earth’s biodiversity, and the sustainable management of the biosphere. However, the current rate of species declines globally, ecosystem collapses driven by accelerating and catastrophic global heating and a plethora of other threats, preclude the ability of habitat protection alone to prevent a cascade of amphibian and other species mass extinctions. Reproduction biotechnologies, biobanking, and conservation breeding programs (RBCs), enabling species restoration, offer a critical transformative change that can economically and reliably perpetuate species irrespective of environmental targets. These capacities extend indefinitely into the future and satisfy humanities needs as the biosphere expands into space. Currently applied RBCs include the hormonal stimulation of reproduction, the collection and refrigerated storage of sperm and oocytes, sperm cryopreservation, in vitro fertilisation, along biobanking. Advanced biotechnologies such as assisted evolution and cloning for species adaptation or restoration have yet to be fully realised. We broaden our discussion to include genetic management, broad political and cultural engagement, and future applications. The development and application of RBCs, and particularly assisted evolution and species restoration, raise intriguing ethical, theological, and philosophical issues. We address these themes with amphibian models to introduce the MDPI Special Issue, The Sixth Mass Extinction and Species Sustainability through Reproduction Biotechnologies, Biobanking, and Conservation Breeding Programs.

Keywords:
Mass extinction
; COP 15
; COP 28
; reproduction technologies
; intergenerational justice
; de-extinction
; climate catastrophe
; assisted evolution
; terraforming
; space colonisation
1. Introduction
The Earths biosphere is under threat from landscape modification, that along with catastrophic global heating and other threats, are driving profound changes in ecosystems globally and accelerating the sixth mass extinction [1]. COP 15, 2022, "Ecological Civilisation: Building a Shared Future for All Life on Earth", focused on direct habitat protection [2,3]. However, habitat modification even in protected habitats, driven by catastrophic global heating, exotic predators and competitors, emergent pathogens, and major losses in prey and other taxa [3,4], will inevitably result in the mass extinction of many amphibian species in the wild [5,6,7,8]. Amphibians are the most threatened vertebrate group, where ~65% of populations are decreasing with only ~40% stable; ~530 species are already Critically Endangered and 1,100 species Endangered, with these comprising ~20% of 8,450 described species [9,10,11]. Rapid declines are also occurring across all terrestrial vertebrates [12].
COP 28, 2023 [13], emphasised the urgency of addressing catastrophic global heating as a major factor in forcing the sixth mass extinction. Catastrophic global temperature increase from pre-industrial levels, based on Intergovernmental Panel on Climate Change (IPCC) estimates, are evidentially conservative. In 2023 the IPPC predicted a 1.10°C increase [1], whereas the actual was 1.46°C [14] and IPPC predictions for 2040 are 1.40°C [1], with alternative models predicting at least 1.85°C [14,15]. These IPPC underestimates could result from IPPC projection models including the reduction of CO2 emissions and increasing CO2 capture and disregarding increased heating through crossing global climate tipping points [14,15]. Unfortunately, meeting CO2 emission reduction targets have failed for most countries with some countries increasing emissions [16]. Amelioration projections for CO2 removal are also prohibitively costly and depend on speculative technologies and unlikely to be achieved within realistic timeframes [16,17,18].
Even more alarming are increasing emission of CO2 from disastrous forest fires rapidly increasing their role as a carbon source rather than a sink [19,20,21], increasing sea water temperatures and acidity reduced oceanic CO2 absorption [22,23], and predictions of the potential collapse by mid to late 21st Century of the Atlantic Meridional Overturning Circulation (AMOC) oceanic current affecting weather patterns globally, including possible season reversals in highly amphibian biodiverse major bioregions [24].
Plausibly, more than the current 1650 Endangered or Critically Endangered amphibian species will become extinct in the wild because of catastrophic global heating synergised by other threats [25,26]. Global heating will particularly affect amphibians subject to altitudinal habitat constraints [26,27,28], dependent on permanent stream flows or wetlands [29], or sensitive to forest destruction through fires [19,20,21] or elevated temperatures beneath tropical forest canopies [30]. Even under the current low IPPC global heating predictions [1], and with planned habitat protection [13], up to ~70% of remaining native vegetation structures will be significantly modified by 2070, with the loss of faunal suites, and faunal population loss is already occurring in otherwise supposedly pristine and protected ecosystems [4]. For example, bird populations have halved in large and otherwise considered pristine Ecuadorian rainforest reserves [31], and insects and other invertebrates have experiencing major populations declines globally [32]. Because biodiversity is irreplaceable, and its loss so functionally and ethically catastrophic, the precautionary principle directs that we should take the direst global heating predictions as our baseline for an immediate and emphatic response including the general adoption of RBCs for species perpetuation [33].
However, governments have not taken seriously the needs of intergenerational justice that entitles future generations to a healthy environment [34], sustainable biospheric management that includes the perpetuation of Earth’s biodiversity [3,35,36,37,38], and especially addressing catastrophic global heating [39,40]. Therefore, transformative change and supportive biotechnical, political, and cultural initiatives [41,42] are needed to reduce or prevent biodiversity loss and ameliorate the sixth mass extinction [40,43,44,45].
These transformative approaches must embrace biotechnological advances rather than relying on optimistic stopgap, adversarial, and traditionalist approaches [3,8,41,42,46,47]. Transformative approaches are slowly gaining traction through the International Geosphere Biosphere Programme [48], the United Nations Convention on Biodiversity [49], the IUCN One Plan Approach to Conservation [50,51] and the 2024 Amphibian Conservation Action Plan [52], Amphibian Ark [53,54], and private carers CBPs [3,55,56]. These transformative approaches include interventional RBC strategies [3] and the promotion of positive societal changes [57]. Futuristic approaches include species perpetuation through long term cell and tissue storage in biobanks that for security may even be eventually secured extra terrestrially [58]. However, the 2024 ACAP was not inclusive and disregarded both the enormous potential of private carers CBPs [56], and the perpetuation of species through biobanking and restoration [3].
Reproduction biotechnologies, biobanking, and conservation breeding programs (RBCs), enabling species restoration, offer a transformative change that irrespective of environmental targets can: 1) reliably and economically maintain genotypic variation within CBPs (Section 2. [10]); 2) produce genetically adaptable individuals for repopulation, augmentation, or supplementation programs [3,10]; 3) integrate ex situ and in situ conservation [59,60,61,62,63,64,65,66]; and 4) provide the potential for species perpetuation solely in biobanks [3,58,67,68,69,70,71,72,73]. These interventional activities can also benefit humanity through cultural and social inclusion, economic and technological advancement, and educational opportunities [3,57,74]. Overall, RBCs offer a significant advancement in the management of biospheric sustainability and the avoidance of mass extinctions by providing a safety net for the perpetuation of Endangered and Critically Endangered species [3,59,60,61,62,63,64,65,66,72,75,76,77]. Sustainability interventions, including amphibian RBCs, have mainly focused on anurans (frogs and toads) and salamanders. Caecilians (Gymnophiona) are an amphibian order of special concern. Besides a general disregard of caecilians in the public and scientific arena [78], other threats include many data deficient species of unknown conservation status, a lack of autecology knowledge because of their generally subterranean habitats [79], and little research supporting the development of caecilian RBCs [61,80,81,82].
Two decades have passed since the potentials of amphibian reproduction biotechnologies were first published in Australia in 1999 [60], fertilisation with cryopreserved amphibian sperm was developed at in Russia [83,84], its use for fertilisation in Australia [85,86], and heterocytoplasmic cloning in Japan in 1963 [87,88]. Despite these pioneering achievements, amphibian RBCs are only implemented for a few species, primarily in wealthy western polities [89,90] or through their satellite conservation programs [91].
This geopolitical limitation creates a critical gap in amphibian conservation, as very high biodiversity countries in the global south are not provided with the necessary independent resources to develop the full potentials of RBCs. The development of biobanks representative of all Critically Endangered and Endangered species globally is also particularly lacking [3]. Nevertheless, preventing the immediate extinction of some amphibian species through RBCs has already been achieved. Between 2007 and 2017, there was an increase in amphibian CBPs of ~60%, covering 2.9% of all amphibian species. Half of these CBPs involved repopulation or augmentation, and 70% of these having some success [10,52,92,93,94]. RBCs using sperm biotechnologies have also benefitted the linking of in situ and ex situ conservation programs [89,90]
Beyond the geopolitical and technical aspects of amphibian RBCs, cultural factors also play a role in their widespread adoption. Scientific publications and public media should utilize an international, clear, and concise nomenclature system that empowers public engagement and optimizes search engine visibility [3]. At an international level this approach, along with oversight beyond the scientific review process, avoids regionalized jargon, misnomers, and soft euphemisms that hinder the powerful and accurate scientifically based dissemination of information [3,95,96,97]. These initiatives are now even extending to addressing historical injustices in species nomenclature [98]. Pioneering achievements should be properly attributed to promote global cooperation in a multipolar world (Section 3.7,4.1.).
Intriguing, philosophical and ethical considerations concerning amphibian animal welfare in RBCs have been particularly prominent in respect to the collection of cells or tissues (Section 5.) the release of individuals into potentially hostile ecosystems for repopulation or assisted gene flow (Section 2.4., [93,94]), and advanced reproduction technologies including assisted evolution or species restoration (Section 4). Despite these ethical considerations (Section 5), intergenerational justice and looming sixth mass extinctions [25,26] demand the prioritisation of biobanking to enable the opportunist restoration of species through cloning or other advanced techniques (Section 4.). Furthermore, to ensure the perpetuation of amphibian species through RBCs, a concerted effort towards international democratic engagement within the emerging multipolar world is essential. This collaboration should broadly include local communities, environmental organizations, and private carers [3,55,99,100,101,102,103].
In summary, this review explores the potential of RBCs through exemplifying amphibians through: Section 2) management models for genotypic variation; Section 3) an overview of RBC protocols including their utility, potentials, limitations, and research; Section 4) advanced reproduction technologies including cloning and assisted evolution; and Section 5) the ethical and philosophy aspects of RBCs to foster public trust and support. Finally in Section 6) we summarise current and future applications of amphibian RBCs and particularly the fostering community engagement and international collaboration to secure a future rich in amphibian biodiversity for the foreseeable future [3]. Overall, this review presents a hopeful outlook for the perpetuation of amphibian biodiversity through embracing current and novel biotechnologies, along with fostering international cooperation through exemplary management, and social and cultural sensitivities.
2. Genetic Diversity and Species Management
Reduced genetic diversity can lead to genotypes with reduced potential for environmental adaptation, poor health, and lowered fecundity [104,105,106,107,108,109,110,111,112,113,114,115]. A goal of the IUCN's One Plan Approach to Conservation is the maintenance of sufficient genotypic variation to provide fitness and evolutionary adaptability. It includes all agencies operating in concert to perpetuate species as both wild and captive populations or biobanked genetic resources of individuals [50,51], in a single genetic management unit called a metapopulation [116]. Therefore, a structured approach using genetic databases of both living amphibians and biobanked material combined with genetic modelling is essential for a scientifically credible RBC program [71,72,89,107,109,110,111,112,117].
Capturing the maximum genotypic variation for ex situ genetic management [71,89,104,105,106,107,118,119] includes the number and proportions of living male, female [110] and biobanked founders [63,66,119]. The IUCN Amphibian Population Management Guidelines recommend 25 founder pairs to represent 99.5% of the target populations genotypic variation (Figure 1. Left Panel, [109,110]). An equal number of founder males and females is preferable; however, a bias toward live females would maximise progeny production for release [120].
The number of founders and initial allelic frequency can be used to calculate the likelihood that an allele will persist in future generations (Figure 1. Right Panel, [82]). However, a greater challenge is the quantification of an allele’s potential for future environmental adaptation. For example, a rare allele might be crucial for developing resistance to a new disease [121] or providing camouflage in a habitat with dramatic seasonality in the chromaticism of vegetation structure [122]. For example, the green and golden bell frog, Litoria aurea, a sun-basking species, inhabits wetlands subject to seasonal changes in vegetation colour from green to brown (Figure 2), with L. aurea varying genotypically throughout its life from pure gold to green, with most individuals a mixture (Figure 2.).
Other approaches have presented alternatives for the population type, ratio of males to females, the application, for genetic management through RBCs or assisted gene flow between populations (Table 1. ibid).
Genetic management of mating in CBPs is crucial for preventing inbreeding between closely related individuals [126]. Dependent on the species generation time and lifespan, with an initial population of 25 live males and females, to preserve 90% of genetic diversity in a CBP for 25 years requires the maintenance of a minimum number of 100 individuals with strict studbook pairing, and the 1,600 individuals with group management and random mating (Table 2., [110]). Unfortunately, despite careful studbook pairing to track the breeding history of individuals, unexpected deaths, domestic adaptations favouring survival in captive conditions, and epigenetic effects as changes in gene expression not caused by DNA alterations, can result in the loss of genetic diversity and allelic variation (Figure 1) and progeny poorly adapted to survival in the wild [106,113,114,127].
A key role of the biobanking of cells or tissues is to increase the effective population size beyond the limited number of breeding individuals typically available in CBPs (Figure 2.). Sperm storage and artificial fertilisation, and particularly utilising banked cryopreserved sperm, are the only current biotechnologies contributing to the prevention of the gradual loss of genotypic variation in CBPs [61,63,65,66,128]. Biobanked sperm pauses the genetic ageing process within a CBP and provides a reservoir of genetic variation including rare alleles that might be crucial for future adaptation to environmental changes such as disease resistance [121], modified ecosystems [4,6], or predators [122].
The benefits of biobanking sperm are exemplified by real-world examples. Case studies involving three frog species demonstrated how using biobanked sperm for breeding (back-crossing) reduces the required size of the CBPs population, minimize inbreeding, and lowers costs when compared to maintaining large numbers of individuals [129,130]. Biobanked sperm can restore genotypic variation to highly inbred colour morphs irrespective of their genetic diversity, in private carers collections thus avoiding costly intuitional CBPs altogether and encouraging private carers engagement in species perpetuation. For instance, with the popular and highly endangered Atelopus, cryopreserved sperm could potentially perpetuate many species potentially through private carers that have already reproduced many species (Figure 3, [55,131]).
Some publications have recommended the biobanking of all threatened amphibian species, Vulnerable, Endangered, and Critically Endangered [118,123]. However, the application of RBCs for species perpetuation should focus on the immediate needs of the current ~1,630 in total Endangered and particularly Critically Endangered species [9,10,11]. Vulnerable species are not in immediate danger of extinction [9,11,132] and their management should follow autecological recommendations for fieldwork, research and monitoring, and particularly habitat protection including encouraging community engagement [3,9,11,132].
2.3. Genetic Bias in Founder Sperm Collection
Minimising anthropocentric bias, terminal investment effects bias, and sperm yield bias in founder selection for sperm collection ensures the maximum representation the target population's genotypic variation [65,66]. Anthropogenic bias occurs through collectors favouring traits such as vocalisation [133], antagonism and larger size [134,135] and chromaticism [136]. However, these traits may not represent a species' adaptive genotypic variation [137,138,139] as they weakly associate with mating systems, habitat types, and life history [135]. Although age is associated with size and senescence theory predicts loss of sperm quality and therefore yield, no effect was found by [140].
Evolutionary trait constraint is possible through increased antagonistic inter-male competitive traits [134,135] corresponding with decreased female mate choice, along with ‘sneaker' males and delayed partial clutch fertilisation influencing fertilisation success [141]. Further research is needed to fully reveal the relationships between sperm genotypic variation [142,143], female sperm choice in fertilisation [144], and the fitness of resultant progeny [145].
Biases could also be due to genetically predisposed characteristics toward pathogens, including susceptibility to more overt parasite-mediated behaviour [146], greater size corresponding to the intensity of infection [147], or through terminal investment effects where sick or potentially dying individuals invest larger resources than usual to increase reproductive output [148]. Terminal investment effect was evident through increased spermatogenesis of males, from two anuran families, during hormonal stimulation when infected by the major threatening fungal pathogen of amphibians [148]. Furthermore, infections with parasites [147], from taxonomic groups with endangered species [149,150,151], also increased spermatogenesis in one species [148,150]. However, in another species male advertisement and mating outcomes were lower in fungal pathogen affected males [152]. Besides potential bias toward parasites, sperm collection through hormonal stimulation includes the possibility of lower sperm yield from less mature or more stress-prone males (See section 4.2. [153]).
2.4. Assisted Gene Flow
Assisted gene flow (AGF), where phenotypic variation is transferred between populations in the wild and in captivity, is an emerging RBC that could potentially increase population fitness but also entails considerable risks. The potential benefits of AGF are the transfer of desirable genotypes between CBPs, biobanks, and wild populations to reduce inbreeding depression [10,65,89,92,154,155,156,157]. Reduced inbreeding depression should increase reproductive capacity, evolutionary adaptability, and consequently species survivability [104,105,106,107,108,109,110,111,112,113,114,118,132,158,159,160]. These advantages are dependent on the target populations size, unique genotypic variation, whether it is a metapopulation including its fragmented sub-populations, or a genetically unique population isolated over geological periods [107,161,162]. AGF strategies toward fragmented subpopulations should utilise pooled genetic diversity from the core population [163] to avoid the diminished genetic diversity of the most divergent or fragmented populations [159,164]. In contrast, AGF should be within isolated population [165], except where historic bottlenecks have reduced genetic diversity to the extent of demonstrably reducing fitness and survivability [166,167]. Any advantages of AGF also depend on natural selective pressures toward genotypic variation that favours survival in the wild irrespective of any loss of genotypic variation [104,107,161,162].
Assisted gene flow toward wild populations can include any life stage, however, various beneficial genetic traits manifest throughout a species' life history [168]; for instance, fungal pathogen lethality varies between the tadpole and the adult stage [169]. Natural selection from early stages favours the retention of endemic genes or beneficial AGF genes along with the loss of detrimental genes [170,171,172,173,174,175]. Therefore, effective AGF strategies toward advantageous genotypes are through egg masses or early larvae. Oocytes could be sourced through on-site collection from wild gravid females, or females in CBPs, and fertilised with genetically diverse sperm [89].
Small, intermittently fragmented wild populations are targeted for AGF programs [176], with the potential benefits and risks dependent on the populations most recent fragmentation and any subsequent gene flow. However, the genetics of naturally fragmented populations are challenging to profile and are likely naturally trending toward inbreeding rather than outbreeding [177]. Therefore, AGF could reduce both fragmented and isolated population's survival through outbreeding depression and loss of alleles [177,178], the introduction of harmful genes [154,177], influencing male/female incompatibility [179,180], and through pathogen transmission [112,113,114,132].
Moreover, the impacts of inbreeding depression and the loss of genetic diversity on the extinction risk of amphibian populations, although widely theorised, are not evidenced on amphibians or other taxa’s declines or extinctions through reduced genetic diversity [181], and many species in thrive with very low genetic diversity [182,183]. These include island endemic species with effective population sizes of 500-1,000 individuals [183], a population size close to the theoretical minimum required to maintain genetic diversity [167]. An increasing number of species of amphibians [10,52], and other taxa [184], are also being repopulated in the wild from a few founders. These cases offer optimism for the survival of low populations of amphibians, if a supportive habitat remains for their survival, without the potential risks of AGF-supplemented genetic diversity [169,171,185].
Some large-scale repopulation programs are based on the hope that individuals develop unique genotypes that ameliorate or counteract new ecological realities such as lethal exotic pathogens [186]. However, these programs for toads [93,187] and frogs [94], have yet to result in unaugmented viable populations. Nevertheless, some initially very small natural populations persist despite the prevalence of lethal pathogens [94,169,188]; however, whether this is due to new genotypes, habitat preference or other behaviour at different life stages is uncertain [169,188]. Other programs simply bolster populations through releases with a recent emphasis toward increasing AGF [189].
The potentials for outbreeding depression [177] and pathogen transmission [190,191] should be evidenced before implementing an AGF, and there should be genetic and demographic monitoring both pre- and post-AGF [192,193]. Genetic and demographic monitoring will be a costly process over prolonged periods [167]. A major consideration before implementing costly and potentially risky AGF is that habitat loss may be the overwhelming cause of a species decline. In these cases, habitat protection, amelioration, or provision could maintain species for the time being without the risks and costs of AGF [171,183,185,194], with survival in the wild proving environmental adaptation and genetic fitness [184].
In summary, the benefits of AGF depend mainly on the genetic diversity of the target population, the translocated genotypes, the target population size, proportionate release numbers of individuals with highly beneficial genotypes, the life stage of release, and environmental selection toward the translocated genotypes. The repopulation of captive-bred individuals into the wild can also pose significant risks through outbreeding depression and pathogen transmission. Thorough planning of AGF is essential, including disease risk assessments, genetic management strategies, and post-release monitoring of demographics and population genetics including population viability analysis and computer modeling.
3. Reproduction Biotechnologies
Amphibian reproductive modes correlate with those of fish, and amphibian RBCs have reciprocal practical applications in fish reproductive management, biobanking, and CBPs [117,195,196,197,198,199,200]. Here, we provide an overview of amphibian RBCs including protocols, their application and future directions, along with the practicalities and ethics of sample collection, and address some recent historical and technical misrepresentations (also See section 5.). Amphibian sperm and oocyte collection, their refrigerated or cryopreserved storage, and artificial fertilisation only require basic laboratory facilities and animal handling procedures [201]. Whereas advanced techniques for cell culture and restoration technologies such as cloning, and assisted evolution, require sophisticated laboratory facilities and technical expertise [72]. Webinars describing details of amphibian reproduction biotechnologies (termed assisted reproductive technologies) only for the hormonal stimulation of sperm and oocytes, and in vitro fertilisation are available online [202].
3.1. Life Stages and Sample Collection
A generalised caution against using early life stages to provide adults for gamete collection was recently published [89]. However, the males of many anuran species mature in less than one year, and both males and females of most species in two years or less [203,204]. Nevertheless, salamanders have longer maturation periods than anurans of many years [61,205]. Average oocyte numbers show that many Endangered and Critically Endangered anuran species in the wild can provide surplus eggs, larvae, and early juveniles [26], and only a few eggs are needed for biobanking embryonic stem cells for species restoration. Different life stages of many Endangered and Critically Endangered species could also be sourced from zoos, CBPs, and private carers [52,55].
We accessed anuran oocyte numbers in Guirguis et al. [26] taken from IUCN-assessments of 4909 anurans, 575 salamanders and 84 caecilians, that included clutch size of 1611 species in total with 405 threatened species For anurans we added new data of 178 non-threatened species, then categorized the combined data, using IUCN Red List criteria, as species not-threatened, Vulnerable, Endangered, and Critically Endangered (Figure 4). However, further analysis showed that these differences were not consistent within clutch number ranges, particularly as the numbers in each category increase. The inclusion of some very large clutch sizes recorder for non-threatened species could also bias results (Table 3).
However, fecundity in respect to Redlist status is highly species specific. For example, Atelopus that include a very high proportion of Endangered and Critically Endangered species, with many species probably already extinct [206], spawn hundreds of oocytes [26]. In contrast, the 12 Oophaga species, with three Endangered, four Critically Endangered, and one Extinct, lay individual eggs (Figure 5., [206,207,208]). Both Atelopus and Oophaga species successfully reproduce in captivity and are held in private carer’s collections [55]. However, although 96% of Atelopus species habits are protected declines and extinctions are continuing [206], management plans do not emphasise the need for biobanking and disregard the potential of private carers collections [207,208]. Nevertheless, a RBC program for Atelopus is underway in Ecuador [131].
Vouchering, the process of preserving physical specimens, can provide valuable authenticated sources of biobanked sperm, and other cells or tissues, [209,210] through the taxonomic distinction of metapopulations [116], or genetically divergent isolated populations [194], as distinct management units [211,212,213]. Vouchering also contributes to amphibian conservation through the epidemiology of amphibian extinctions and declines [186], and by providing life history and ecological information required to facilitate CBPs and release programs [132]. Unfortunately, a decline in vouchering has led to impoverished museum collections that cannot even adequately address even the looming problem of global heating driven mass extinctions [212]. Fortunately, zoos are making significant contributions to biobanking and vouchering [213], but this activity could be more widely extended throughout RBCs.
3.2. Reproduction, Gamete Collection, Donor Stress, and Pathogens
The least stressful reproduction biotechnology is the simulation of their natural reproductive cues to promote reproduction [214,215]. Cues of temperature and humidity are generally intermittently circannual in temperate anurans and continuous in salamanders and tropical anurans [216,217] with precipitation generally triggering reproduction [218]. These are preferrable for animal welfare, and mostly used with most species to produce large numbers of progeny by private carers [55] and in some augmentation or repopulation programs [214]. Temperature regulated brumation can promote gonad maturation, mating and spawning responses to hormonal stimulation [64,214,219], with males and females of at least one salamander requiring different temperatures to promote reproductive maturity [220]. Sex differences in responses to environmental and social breeding cues have also been found in one anuran [221]. An alternative to mating to produce progeny is the collection of sperm to be used fresh, refrigerated for days to weeks, or cryopreserved (Section 3.7.), for the in vitro fertilisation of oocytes that may be stored for days to a week (Section 3.4,3.5.,3.6.)
Sperm from testicular tissues has made substantial contributions to both foundational [66,83,84,85,222] and recent amphibian RBC [76,179,223,224,225] and has the highest potential for producing large numbers of individuals for repopulation programs. An unsubstantiated bureaucratic impediment toward the use of this technique is found in the IUCN, Amphibian Conservation Action Plan 2024, “In cases where gamete recovery is part of a conservation strategy euthanasia is not recommended” [52] (p294).
However, sperm collection from testicular tissue should be the preferred method if males are available as it only requires euthanasia and is the least stressful technique [76,85,86,223,224] with the euthanasia of donors, dissection of testes, and the production of sperm suspensions achieved in a few minutes [179]. Testicular tissue also provides the highest sperm yield in highly concentrated sperm suspensions [61,64,66,76,85,86,222,223,224]. Sperm collection from testicular tissue is preferable with 1) males taken from the wild without threatening populations, 2) surplus males from CBPs [66,76,223,224], 3) vouchered males section 3.1), 4) the collection of large high-quality sperm yields from small and especially very small amphibians [61,95], 5) mortalities after strict pathogen screening [190,191], 6) the testing of multiple female/male compatibilities [179], 7) to produce quantities of concentrated sperm suspensions of consistent pH and osmolarity [179], and 8) essential for species recalcitrant to hormonal stimulation [10,226].
Hormonal stimulation, (misnamed hormone therapy is some amphibian RBC literature [227]), does not require euthanasia but generally requires injection, and is a more stressful and less efficient alternative to sperm collection from testicular tissue, and has been a popular research topic for more than 40 species [52,65]. In some species hormonal stimulation only yields low sperm numbers, and concentrations, of inferior sperm quality [10,226] further limiting its utility. An instance of hormone stimulation producing extremely low sperm concentrations of poor-quality sperm is found with the Endangered frog, Leiopelma hamiltoni, in the phylogenetically basal Anura family Leiopelmatidae, (Figure 6., [226]). A similar response is also likely with Le. archeyi, the highest priority amphibian species on the Zoological Society of London's Evolutionary Distinct and Globally Endangered (EDGE) list [196].
Unfortunately, many studies of hormonal stimulation to collect sperm do not present the sperm yields as the number of sperm making it difficult to assess the utility of this technique for the target species [52,65]. Nevertheless, even if not generally practicable for large scale progeny production with some species and particularly small amphibians, hormonal stimulation can yield sperm quantities, whether motile or immotile, that are suitable for intracytoplasmic sperm injection (ICSI) [228,229,230,231,232], and low sperm yields of fertile sperm can contribute to biobanks or the production of larvae that then can be raised to maturity to produce large numbers of progeny for release [3].
After hormonal stimulation some anuran species will spontaneously express sperm in urine [233,234,235] or a cannula or a pipette tip can be inserted into the urethra [10,236]. Salamander sperm can be collected after hormonal stimulation in pre-spermatophores or in gelatinous fluid [237,238], spermic urine [237,238] or semen collected by massaging the lower oviduct and cloaca [205,237,239,240], or through the natural deposition of spermatophores [66]. Very large quantities of hormonally stimulated giant salamanders (Cryptobranchidae, Cryptobranchus alleganiensis) hormonally stimulated semen is readily stripped in aquaculture and in the development of RBCs [205].
However, hormonal stimulation using injection is exceptionally stressful to small delicate salamanders [239] including the highly threatened plethodontids that comprise 65% of salamander species [61,239]. Alternative methods for hormonal stimulation of spermiation in small delicate salamanders or anurans include topical or nasal application [61,241,242]. The more robust body shape and consequently lower skin surface to body mass ratio could make small anurans less amenable to the topical or nasal application of hormones than salamanders of the same body weight. On the other hand, anurans have a patch of skin in the ventral pelvic region for absorbing aqueous solutions [243]. Spermiation was stimulated using hormonally injected crickets force fed to salamanders, termed non-invasive oral bioencapsulation [244]. This novel technique offers the potential to avoid the stress of hormone stimulation entirely and worked in one publication for both large and small salamanders [244].
Females of both anurans and salamanders are generally more recalcitrant to hormonal stimulation for oocyte collection, than males for spermiation, and often require priming doses [64,233,237,245,246,247]. Mature oocytes can then be collected ovarian excision [85], abdominal massage, or cannulation [61,62,63,64,237,248,249] Improved targeting of hormonal stimulation has been achieved through ultrasound identifying mature ovaries in both anurans [250] and salamanders [234,237]. Ovarian excision and spawning into physiological saline (Section 3.6.), are low stress techniques for the sampling of large numbers of oocytes [233,247]. Unseasonal hormonal stimulation of oocytes did not affect oocyte quality in one cool-temperate species [251].
An important factor in selecting gamete collection methods is pathogen transmission [190,191]. Testicular sperm may be contaminated by pathogens such as internal parasites and viruses [147,149], whereas hormonally stimulated sperm in various forms may additionally contaminated by bacteria, fungi and external parasites [252].
3.3. Sperm Qualities
Comparative evaluation of spermatological techniques through sperm quality metrics is confounded by inconsistencies in methods and terminology. Key sperm quality metrics are motility, membrane and DNA integrity [65,66,140,195,240,253,254,255,256,257]. Basic metrics of sperm motility are the percentages of immotile sperm, activated sperm as non-swimming motion, swimming sperm, and sperm speed. Sperm motility is subjectively assessed by observation [85], however, meaningful comparative studies need computer-aided sperm assessment (CASA) to give exact percentages and speeds of swimming sperm, along with other motility vectors for more precise analysis [66,258]. Damages to sperm morphology assessed by observation or vital stains are the integrity of acrosomes [176] and mitochondrial collars or sheaths [257,259,260] (misnamed in some in the amphibian RBC literature as ‘mitochondrial vesicles’ [259,261]). DNA integrity is assessed through comet assays and other tests [219,255,256], with other biochemical tests sometimes used for amphibians [253].
3.4. Refrigerated Storage of Sperm
Refrigerated storage at 0-4°C is a useful technique for the short-term storage of amphibian sperm. Refrigerated storage enables delayed in vitro fertilisation when there is asynchrony between sperm collection and oocyte availability [253], or for transport between breeding groups [89]. Refrigerated storage for ~10 min. can acclimate sperm to cryoprotectants [257] and be used to delay cryopreservation for days to weeks [66,86]. Refrigerated storage of anuran sperm is highly successful in cadavers [219,235],whole testes [86], macerated testicular tissue [86], and in spermic urine [65]. However, refrigerated salamander sperm only remains motile for a day to several days [61,66], except for weeks when held in spermatophores [262]. Oxygenation [263,264] as well as the addition of antibiotics to reduce bacterial concentrations, improves sperm storage periods while maintaining sperm concentrations and increasing biosecurity [65,263,264,265,266,267].
3.5. Sperm Cryopreservation and Freeze Drying
Cryopreservation is a freezing process that protects sperm using special solutions to create sperm cryosuspension and tailored freezing rates. The cryopreserved sperm can then be held indefinitely in biobanks [65,66,268]. Sperm cryopreservation uses aqueous formulations, termed cryodiluents, that when mixed with sperm to produce cryosuspensions that are then frozen [65,66]. Cryodiluents are formulated from sperm penetrating cryoprotectants such as dimethyl sulphoxide (DMSO, [83,84] or dimethylformamide (DMFA, [219,257]), and non-penetrating saccharides or salts, with supplements such as buffers, fetal bovine serum, and anti-biotics [65,66]. Fertilisation was first achieved with cryopreserved anuran testicular sperm in 1996 using DMSO [83], and with hormonally stimulated sperm in 2011 using DMFA t hen a novel cryoprotectant to amphibians [257]). Sperm of has also been freeze-dried and remain viable, then stored at room temperature [269]., but this technique has not been trailed amphibians either for fertilisation [61,66] or for ICSI [228,229,230,231]
A broad canvas of the RBC community in 2019 recommended 5-10% (v/v) DMSO or DMFA and 1-10% (w/v) saccharide as cryoprotectants with slow to moderate cooling rates [66]. Nevertheless, ranges of 12-15% DMSO have proved successful with anurans [83,84,86,219,223,224,257], with 12% DMFA proving superior to DMSO with urinal sperm [257] and consequently used in other studies [258,268,270]. Both fast and slow cooling rates have recently been successful [65,66,258]. Cryosuspension osmolarity on cryopreservation in some studies has shown a significant effect [223] while other studies have shown much less effect [270]. Freezing of sperm over a range of freezing rates can be achieved by constructing a simple and inexpensive device [271], with a comparative description of cooling rate impacts in [258].
The broad differences in successful cryopreservation protocols, even within the same species may be due to subtle variations in techniques for cryoprotectant penetration, freezing regimes, the thawing and the washing of cryoprotectants from cells [65,66], seasonal effects on sperm quality [272], in vitro fertilisation techniques lacking meaningful baselines (See 5.9.), unconcise sperm quality metrics (Section 3.3.,4.3.), and sperm genotypic and phenotypic characteristics [115,268].
Little difference was shown between the cryoresistance of motile urinal sperm or sperm from testicular tissues [270].
3.6. Oocyte Storage
The storage of oocytes enables delayed in vitro fertilisation if there is asynchrony between the availability of oocytes and sperm [65,66,273]. Because of their high yolk content and large size, the cryopreservation of anuran oocytes is not yet practicable [274]. The viability period of unfrozen oocytes during storage depends on the species' natural spawning temperature, the storage temperature, and the oocytes osmotic, ionic, and gaseous environment [64,86,222,275,276,277]. Early conventions for oocyte storage assumed that ionic formulations slowed oocyte gel hydration (described as hardening) that blocked sperm penetration [148,276,277], however, research over a broader range of species showed that hydrated oocytes of some species are fertilisable for an hour or more [222]. Unhydrated and refrigerated ovarian oocytes and post ovarian oocytes of cool temperate species have remained viable for many days [275] with a pressurised gaseous environment further extended storage life [278].
Oocyte storage is highly dependent on the target species natural spawning temperature, where the oocytes of species spawning in cold water may remain viable for many days [275] and tropical or subtropical species only for hours [86,276,277]. Studies with a single species showed that a percentage of post-ovarian oocytes undergo a very rapid loss of fertility [179]; possibly a female sperm choice mechanism for greater progeny heterozygosity with polyandrous species [279], or to maintain sub-population environmental adaptability where female/male genetic incompatibility lowered fertilisation rates in anurans with little dispersal ability [179,180]. In other species, some oocytes resist fertilisation during spawning, leaving fertilisable oocytes for later fertilisation [141]. However, the physiological or genetic mechanisms behind selective oocyte fertility loss over time, and any subsequent genetic biases toward progeny, are undetermined. Therefore, besides species specific temperature effects, the mechanisms for the loss of oocyte viability in aqueous solutions generally appears to be through the diffusion of ions or proteins from the oocyte gel needed for sperm motility or for oocyte metabolism [280,281,282,283] rather than gel hydration [276,277]. Specific genotypic mechanisms also influence the storage period and even extend to individual oocytes [179].
The cryopreservation of amphibian oocytes or early embryos has no parallel with techniques used for mammalian oocytes, which are very small, ~0.08-0.20 mm diam., and have a miniscule amount of egg yolk [284]. However, the cryopreservation of amphibian oocytes or embryos includes many parallels with fishes [197,198,274] due to both possessing highly fatty and structured egg yolk and oocytes sizes. Nevertheless, the success of cryopreservation techniques for fish embryos up to 0.8 mm diam. [197] are challenging to apply to the large size of more than 95% of amphibian oocytes that are over 0.9 mm diam. (Figure 7.). Therefore, the currently most the most promising techniques to perpetuate the amphibian female genome is using cryopreserved biomaterial for heterocytoplasmic cloning (Table 4, ibid) or stem cells to generate ovaries in surrogate species (Section 5. [274]).
3.7. Artificial Fertilisation; Sperm Concentrations, Fertilisation Periods, and Rates
The critical end point of artificial fertilisation comparing sperm concentrations and quality, and fertilisation rates, are subject to major limitations in techniques, terminologies, and in historic attribution. The term “artificial fertilisation" is generic and not only includes in vitro fertilisation where sperm is placed over oocytes as presented in [179,273,285], but also artificial insemination with sperm placed internally [286], and intracytoplasmic sperm injection (ICSI) with sperm placed into an oocyte [228,229,230,231]. In vitro fertilisation is the critical end point of much of amphibian RBC research and has been achieved with scores of anuran species, and tens of salamander species [52], but artificial insemination has been trialed with only one salamander species [286].
The first book devoted to amphibian RBCs published in 2022 in "Historical Perspectives on the Development of Amphibian Reproductive Technologies for Conservation” ([273], p1) stated that in vitro fertilisation in amphibian RBCs was ‘dry fertilisation’, and attributed “dry fertilisation” as a pioneering experimental achievement, to Rugh, 1961, in the USA [287]. Historically, dry fertilisation was pioneered in 1856 in Russia where inactivated fish sperm was mixed with oocytes then after a short period the sperm was activated with water to achieve fertilisation [288]. In contrast, Rugh first published the placement of activated sperm on anuran oocytes in 1934 [289]. This technique was then furthered in other early studies [290,291], first used in amphibian RBCs in Russia in 1996 [84] and subsequently generalised in amphibian RBCs [219,233,234,235,247].
Despite being subject to so many publications [52] our knowledge of in vitro fertilisation is limited by a lack in publications of comparable sperm quality metrics (Section 4.3.), sperm concentrations and fertilisation periods [85,222,257,292,293], and sperm activation and fertility in different osmotic environments [65,66,225,240,293,294]. The effect of osmolarity can even extend to osmotic environment of donor males [294]. Fertilisation curves are the end point of sperm quality; however, few publications have included amphibian fertilisation curves; first presented in the general scientific literature in 1975 [292] and the RBC literature in 1998 [86]. Furthermore, the fertilisation period in most amphibian studies is 10 min. and refers to concentrated sperm suspensions [86]. However, comprehensive fertilisation curves at different sperm concentrations show that fertilisation rates with Rhinella (Bufo) arenarum increased for at least 30 min. post application, with post application period more influential than linear sperm concentrations on fertilisation rates (Figure 8. [292]).
Other fertilisation curves show saturated fertilisation with sperm from testicular tissue at concentrations of 2.5 x 105 ml-1 after 60 min. [293], ~104 ml-1 after 60 min. [222], ~106 ml-1 after 10 min. [86]), and 105-7 ml-1 after 60 min. [295]. Fertilisation rate in general will also depend on the relatively short motility period, and lower quality, of cryopreserved sperm compared with fresh sperm [296]. For instance, after 15 min. application Rana temporaria spermic urine at ~106 ml-1 provided ~45% fertilisation while testicular sperm provided only ~2% fertilisation. Spermic urine achieved saturated fertilisation at ~107 ml-1 but testicular sperm only at ~108 ml-1 [257].
The need for robust fertilisation curves to compare fertilisation rates was exemplified through much higher fertilisation rates than expected with cryopreserved urinal Anaxyrus (Bufo) fowleri sperm. Control fresh swimming sperm provided ~85% saturated fertilisation at ~4.2 × 106 ml-1 concentration, whereas after cryopreservation swimming sperm at only ~2% concentration of the control provided ~20% fertilisation [297], even though the fertility of cryopreserved sperm is lower than fresh sperm [257,270]. A robust fertilisation curve could explain this apparent enigma, and in general provide baselines for meaningful comparisons between all studies.
4. Advanced Reproduction Biotechnologies (aARBs)
Advanced Reproduction Biotechnologies (aARBs) can perpetuate species through the cryopreservation of cultured or uncultured somatic cells [298] with subsequent cloning [89,198,274,284,299,300], or stem cells used for the generation of gametes in surrogate species [67,70,71,274]. The generation of gametes includes the implantation of cryopreserved immature ovarian follicles, primordial germ cells, or induced pluripotent stem cells to generate chimaeras [63,67,274]. These techniques were first conceptually introduced to the amphibian RBCs literature in 1999 [60].
4.1. Cloning
Heterocytoplasmic cloning enables species restoration solely from biobanked nuclei through surrogate species [89,198,274,284,299,300], and the critical need to develop heterocytoplasmic cloning has not received the attention it deserves in amphibian RBCs. To perpetuate amphibian biodiversity, heterocytoplasmic clones must develop to reproductively mature and fertile females to produce oocytes for fertilisation with cryopreserved sperm. Homocytoplasmic anuran clones from embryonic cells developed to late blastula as early as 1952 [301], with heterocytoplasmic cloning first accomplished with anurans in Japan and with salamanders in France (Table 5., ibid). Heterocytoplasmic clones then developed to the gastrula stage in 1957 [302], early neurula in 1958 [303], adults in 1961 [304], with naturally mating and spawning adults producing viable offspring in 1963 [87,88,305], second and third generations from 1963 to 1972 [306,307], and from as early as 1971/1972 adult heterocytoplasmic clones were produced from a wide range of other anurans [308,309], and also from salamanders (Pleurodeles sp.) (Table 4. ibid; [310,311]. In 1998 nuclei from cryopreserved totipotent cells developed to the gastrula stage [300] using the techniques of [299]. These foundational studies and the extraordinary rate of the development of de-extinction (species restoration) projects for other taxon relying on cloning [312], herald a new age in amphibian cloning that will lead species perpetuation solely from cryopreserved nuclei. This will greatly reduce the need for CBPs for species unable to survive in the wild, or CBPs for species of little cultural interest. This advance will not only relieve some of the financial burden through maintaining live amphibians but also have considerable animal welfare benefits.
The first book devoted to amphibian RBCs published in 2022 in "Historical Perspectives on the Development of Amphibian Reproductive Technologies for Conservation” ([273], p1) attributed “one of the most noteworthy and groundbreaking accomplishments toward amphibian reproductive technologies” to Gurdon’s, 1962, publication of amphibian homocytoplasmic cloning in the USA [313], with this accomplishment was also attributed to Gurdon receiving a Noble Prize in 2012 [314]. However, Gurdon’s homocytoplasmic cloning was not pioneering and extended Briggs and King's homocytoplasmic cloning of amphibians in 1952 [301], neither concerned heterocytoplasmic cloning a technique essential for amphibian species perpetuation [273]. Furthermore, the 2012 Nobel Prize for Physiology or Medicine was awarded jointly between John Gurdon and Shinya Yamanaka [314], not for cloning, but for the reprogramming of nuclei to pluripotency [314], as discussed by Gurdon in [315] his acceptance speech “The Egg and the Nucleus: A Battle for Supremacy” [316].
4.2. Assisted Evolution
Traditional methods of assisted evolution are the selection of desirable traits in domestic species [317] as found in amphibians with private carers coloured morphs [55]. Other assisted evolution techniques with amphibians include the natural selection of beneficial traits through mass releases [93,94], and approaches toward genetic engineering to provide pathogen immunity or adaptability to the climate crisis [52,119,318,319]. In contrast to most amphibians, the benefits of assisted evolution in mammals are challenged by their low reproductive rates [72]. Nevertheless, assisted evolution is a crucial strategy to select or increase the amphibian genotypic variation or for species restoration as amphibians are highly amenable to RBCs [3,52,67].
5. Ethics and Communication
By addressing ethical principles, communication challenges, and cultural normalization, we unlock the tremendous potential of RBCs to fulfil our obligations toward intergenerational justice [3,35,36,37]. However, there are few publications concerning the ethical standards of amphibian RBCs [52,320]. The ultimate standard for the ethical treatment of all animals is “The physical and psychological well-being of an animal (sic, in captivity). It is good or high if the individual is fit, healthy, free to express natural behavior, free from suffering and in a state of wellbeing.” [215].
Ethical principles dictate that RBC practitioners are moral agents responsible for the wellbeing of their moral subjects, the species of concern [321,322]. Practitioners’ ethical motivation towards amphibian RBCs is driven by a responsibility to perpetuate amphibian biodiversity while avoiding unnecessary harm [52]. Practitioners are guided by the precept, "How would I like it if I were them?" an ethical principal present in all cultures with ethical traditions [322]. RBC animal ethics standards include amphibians possessing sentience through the cognition of stimuli without association or interpretation [322,323,324], standard that can even extend to invertebrates [325].
In practice, the ethical use of RBCs depends on a balance between 1) the sentience of the target individuals [321,326], 2) the species' role as a biospheric entity thereby benefiting other moral subjects [327]; 3) support for intergenerational justice toward the environment [35,36,37], and 4) the principle of the greatest good for the greatest number as applied to the species perpetuation [328]. Ethical considerations depend on the reason for conducting the technique, its efficiency, general applicability, and associated stress to the targeted individuals (Table 6., ibid). Once an RBC protocol or program using animals is justified the three basic animal ethics principles of refinement, reduction, and replacement, in research or application can be applied [329,330,331].
Refinement can be achieved through procedures that minimize stress and pain during techniques to collect sperm and oocytes (Section 3.2, [244,320]), and avoiding hormonal stimulation by providing habitat simulations in CBP to promote mating and spawning [201,215]. Research regarding amphibian stress during confinement and handling will further guide the research and application of amphibian RBCs [153]. Reduction achieved through reducing the number of individuals in CBPs through using stored sperm, and by avoiding extensive hormonal stimulation trials to optimise hormonal sperm collection, when alternatively large yields of sperm could be obtained from testicular tissues (Section 3.2.). Replacement by programs maintaining genotypic variation through biobanking of sperm from any Endangered or Critically Endangered species for later use (Section 3.2., [3,65,66]). Replacement can also be achieved by genetic and demographic modelling to effectively reduce the required number of individuals to manage CBPs, biobanking, supplementation, repopulation, or assisted gene flow programs (Section 2.4.).
Biotechnologies for sperm collection have mainly focused on hormonal stimulation of sperm or oocytes, with research involving more than 40 species [52,320]. This high stress technique, and especially with small amphibians, generally only yields low sperm numbers and concentrations [226]. Furthermore dose-response curves for two hormones are recommended to optimise this technique [10], requiring at least 32 individuals being subject to hormonal stimulation, and the use of cannulation to collect sperm with most species. However, if mature males are available high sperm yields can be collected through the reliable low stress technique of testicular tissue to provide the sperm numbers for the fertilisation of large numbers of oocytes (Section 3.2). In either case even a few biobanked sperm can provide genotypic variation or produce mature adults to provide large numbers of progeny.
Nevertheless, the collection of spermic urine through cannulation is critical for the progress of the basic science of spermatology to provide sperm that are known to be equivalent to those expressed in natural spawning [257]. The comparative quality in amphibian RBCs is dependent on the basic science of the sperm structure, physiology and motility mechanisms particularly in respect to viscosities found in internally fertilising amphibians [61] and the physical structure of oocyte gel and female sperm choice mechanisms [279]. In many cases these advances can only be achieved using sperm from spermic urine, and cannulation is the best procedure to obtain this sperm in many species. Institutional animal ethics requirements should be sympathetic toward this critical need for the perpetuation of amphibian biodiversity.
Besides animal welfare, the ethics of amphibian RBCs extend to human/animal interactions through emotional and social engagement and the broad ecological impacts of repopulations or translocations [3,71,72,323,324,332]. Furthermore, speciesism, where one species is placed as morally more important than other species is shown where target species are selected on political or institutional rather than phylogenetic grounds [196]. There is considerable engagement of the theological community in the need to support biodiversity conservation [333] and avoiding language that may be theologically misinterpreted such as resurrection [75] for species restoration, and playing God helps to ensure broader public support [75,334,335,336]. Theologies can also contribute to biospheric sustainability through social and ethical models that respecting nature and consequently the activities of their members [337]. Although subject to ethical scrutiny the popularity of species restoration is shown by the global reach of the Colossus de-extinction project [311], with its potential for biospheric sustainability and profound community outreach [3,332].
6. Current and Future Application
The development of amphibian RBCs has progressed to the collection and storage of anuran and salamander sperm and oocytes and their use for in vitro fertilisation to produce sexually mature adults [61,76,77,224]. These RBCs can support either institutional or private CBPs, along with repopulation or translocation programs [3], and assisted gene flow (Section 2.4.) However, costly programs for wild populations should be highly targeted toward ecologically, phylogenetically, or culturally significant species that are likely to eventually independently survive in the wild for at least decades [52,116,196]. This targeting is particularly relevant in consideration of the rapid and accelerating anthropogenic modification of the biosphere (See Introduction). Species not expected to survive in the wild, irrespective of the environmental targets of COP 15 or COP 28, should be subject to CBPs supported by biobanked sperm or perpetuated solely in biobanks (See section 3.). Only through addressing these issues will the amphibian RBC community satisfy their ethical responsibilities toward providing reliable and cost-effective intergenerational justice. Caecilians offer an exceptional needs and opportunities for the development and application of RBCs, because of caecilians internal fertilisation, and in many species’ internal development, and maternal care in all terrestrial species [61,79,80,81].
The high costs of implementing recovery-based conservation are attributed to maintaining or providing specialized habitats, extensive research and monitoring, conservation breeding and reintroduction programs, and the need for professional management [3,338]. The financial burden of meeting these global biodiversity conservation targets through reducing extinction risk through interventions along with establishing and maintaining protected areas is substantial, with estimates of billions a year [339], with mammals and birds typically requiring the most resources [340]. To address these challenges, a globally inclusive approach is needed, focusing on developing highly cost-efficient RBC facilities in regions with high amphibian diversity, that welcome community engagement and international collaborations [3,341].
The short-term prevention of the extinction of amphibian species through RBCs to maintain, repopulate, or translocate amphibian populations in the wild [3], has already been reliably and economically demonstrated. Between 2007 and 2017, there was a ~60% increase in amphibian CBPs, including 80 species, or 2.9% of all species, with half of these CBPs involving repopulation or augmentation and 70% having some success [10,52]. However, these approaches must recognise a future of dramatic ecosystem collapses driven by global heating leading to mass extinctions [13,38,39,40,45]. The establishment of biobanks of amphibian cells and tissues will provide cost effect and reliable species perpetuation [3,58,67,68,69,70,71,72,73].
Exciting opportunities for amphibian RBC research and development include cloning and somatic cell techniques for species restoration (de-extinction) [312]), extraterrestrial biotechnologies [3,342,343], and terraforming for colonisation [3,342]. Terraforming conceptually includes the past, current, and future anthropogenic modification of Earth's biosphere as extended to extraterrestrial ecosystems along with their biodiversity [3,342].
Public education and outreach for amphibian conservation and include educational programs, community engagement, media campaigns, and interactive experiences [344]. These initiatives can foster empathy, promote citizen science, and encourage sustainable practices [3,52,55,345]. Perceptions of conservation success can vary but are generally perceived in terms of species and habitat improvements, effective program management, outreach initiatives, and the application of science-based conservation including RBCs [52,345]. The 2024 Amphibian Conservation Action Plan (ACAP) provides a guide for capturing outcomes, identifying gaps, and measuring progress in conservation efforts [52].
However, the 2024 ACAP [52] framework was limited by the mandate of supporting ‘Amphibians thriving in nature’ (in the wild) and largely disregards the transformational changes to species management when considering of the sixth mass extinction [3]. Chapter 12 ‘Amphibian assisted reproductive technologies and biobanking’, in the ‘Priorities and recommendations’ section, does not mention species perpetuation through biobanking, cloning, and species restoration, with similar viewpoints by the Amphibian Ark in neglecting species that cannot foreseeably be returned to the wild [346]. Furthermore, the ACAP ‘Chapter 11. Conservation breeding’ is not inclusive of private carers potential to contributions the perpetuation of amphibian biodiversity [55]. The disregard on many internet sources for the benefits of the biotechnical aspects of RBCs and biobanking [347], also shows the need and the challenges to the effective advocation and popularisation of the full potentials of RBCs.
The advocation and popularisation of amphibian RBCs includes engaging cultural discourses involving assisted evolution [348], cloning and species restoration [72,349], synthetic biology [342,350,351], and theological debates about humanities relationship to nature through synthetic biology, resurrecting species, and playing God [348,349,350,351]. Support for these initiatives includes standardised scientifically based terminologies popularised through the public media [3,95,96,97,259,352]. Furthermore, an emphatic effort must be made to remove anachronistic bureaucratic impediments including policy and media [52].
The development of solid RBC career streams will attract ambitious and talented researchers and activists to build RBC projects by competing for influence and resources [3,95]. Initiatives should focus on species perpetuation to ameliorate amphibian mass extinction and be inclusive of the broadest global community [3,333]. Community popularisation of advanced reproduction technologies is exemplified by the highly successful Colossal project, where their central theme of de-extinction (species restoration) is extended for their target species to community-based field conservation projects [312,332], advocating bioengineering as an initiative to help heal the Earth [353,354].
In summary, we have shown that the perpetuation of amphibian biodiversity, rather focusing on the limited goals of the 2024 ACAP, other institutions, and media sites of amphibians thriving in the wild should recognise the community’s responsibility to adapt to the environmental and cultural realities of the Anthropocene [355,356] including ecosystem modification’s, collapses and the sixth mass extinction [6,7,15,38,355]. Besides current initiatives, this transformation should harness the potential of biobanking and species restoration to perpetuate species and be inclusive of all global participants including private carers CBPs [3,55]. A greater focus is also needed to garner global support and international engagement toward developing RBCs in highly biodiverse regions, especially developing countries, and the provision of independent finance. An independent, democratic, and inclusive global organisation representing an increasingly assertive multipolar world is needed to advocate these needs [3].
Finally, will humanity over the coming millenniums look back to this century relegating plausibly thousands of amphibian species that will not continue to exist in the wild to extinction as promoted in [346,357]. Or alternatively will we face the challenges of the Anthropocene presented by inevitable biospheric modification and especially through global heating [39,40], and perpetuate species through realist innovative, cost effective, and reliable biotechnologies [3] supported by broader community inclusion in a multipolar world [3,44].
Author Contributions
All authors have read and agreed to the published version of the manuscript and have contributed substantially to the work reported.
Funding
This research received no external funding.
Data Availability Statement
Data concerning oocyte numbers and sizes are currently being used to support several research articles on the evolutionary ecology of amphibian sperm. Nevertheless, the data presented in this study are available, in confidence, on request to the corresponding author.
Acknowledgments
Special thanks to Jasmine Flores for collating spawn number and endangerment status data.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- IPCC, 2023: Summary for Policymakers. In: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (eds.)]. IPCC, Geneva, Switzerland, pp. 1-34, Available online:. [CrossRef]
- UNEP. UN Biodiversity Conference (COP 15). December 7-19, 2022. Available online: https://www.unep.org/un-biodiversity-conference-cop-15 (accessed on 23 September 2023).
- Browne, R.K.; Luo, Q.; Wang, P.; Mansour, N.; Kaurova, S.A.; Gakhova, E.N.; Shishova, N.V.; Uteshev, V.K.; Kramarova, L.I. .; Venu, G.; Vaissi, S.; Taheri-Khas, Z.; Heshmatzad, P.; Bagaturov, M.F.; Janzen, P.; Aguilar, R.E.N.; Swegen, A.; Strand, J.; McGinnity, D.; Dunce, I. Ecological Civilization COP 15, COP 28, and Amphibian Sustainability through Reproduction Biotechnologies, Biobanking, and Conservation Breeding Programs (RBCs). Animals 2024, 14, 1455. [Google Scholar] [CrossRef]
- Conradi, T.; Eggli, U.; Kreft, H.; Schweiger, A.H.; Weigelt, P.; Higgins, S.I. Reassessment of the Risks of Climate Change for Terrestrial Ecosystems. Nat Ecol. Evol. 2024, 8, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Grant, E.H.; Amburgey, S.M.; Gratwicke, B.; Chaves, V.A.; Belasen, A.M.; Bickford, D.; Bruhl, C.A.; Calatayud, N.E.; Clemann, N.; Clulow, S.; et al. Amphibian Conservation in the Anthropocene. Biol. Conserv. 2023, 236, 543–547. [Google Scholar] [CrossRef]
- Willcock, S.; Cooper, G.S.; Addy, J. Dearing, J.A. Earlier Collapse of Anthropocene Ecosystems Driven by Multiple Faster and Noisier Drivers. Nat. Sustain. 2023, s41893-023-01157-x. [CrossRef]
- Finn, C.; Gratarolla, F.; Pincheira-Donoso, D. More Losers than Winners: Investigating Anthropocene Defaunation through the Diversity of Population Trends. Biol. Rev. Camb. Philos. Soc. 2023, 98, 1732–1748. [Google Scholar] [CrossRef]
- Bradshaw, C.J.A.; Ehrlich, P.R.; Beattie, A.; Ceballos, G.; Crist, E.; Diamond, J.; Dirzo, R.; Ehrlich, A.H.; Harte, J.; Harte, M.E.; Graham, P.; et al. Underestimating the Challenges of Avoiding a Ghastly Future. Front. Conserv. Sci. 2021, 1, e615419. [Google Scholar] [CrossRef]
- The IUCN Red List of Threatened Species. Version 2022-2. Available online: https://www.iucnredlist.org (accessed on 27 September 2023).
- Silla, A.J.; Byrne, P.G. The Role of Reproductive Technologies in Amphibian Conservation Breeding Programs. Annu. Rev. Anim. Biosci. 2019, 15, 499–519. [Google Scholar] [CrossRef] [PubMed]
- AmphibiaWeb. Available online: https://amphibiaweb.org (accessed on 27 September 2023).
- 157. Smith, D.; Abeli, T.; Bruns, E.B.; Dalrymple, S.E.; Foster, J.; Gilbert, T.C.; Hogg, C.J.; Lloyd, N.A.; Meyer, A.; Moehrenschlager, A. et al. Extinct in the wild: The precarious state of Earth’s most threatened group of species. Science 2023, 379, eadd2889. [Google Scholar] [CrossRef]
- UNFCCC COP 28. November 30th -December 12th, 2023. UN Climate Change Conference. Available online: https://unfccc.int/cop28; (accessed on 29 December 2023).
- Hansen, J.E.; Sato, M.; Simons, L.; Nazarenko, L.S.; Sangha, I.; Kharecha, P.; Zachos, J.C.; von Schuckmann, K.; Loeb, N.G.; Osman, M.B.; et al. Global Warming in the Pipeline, Oxford Open Climate Change, 2023, 3, kgad008. [CrossRef]
- Global Tipping Points. Available online: https://global-tipping-points.org/ (accessed on 29 December 2023).
- CAT. Climate Action Tracker IMF. Available online: https://climateactiontracker.org/ (accessed on 6 August 2024).
- Smith, S M; Geden, O; Gidden, M J; Lamb, W F; Nemet, G F; Minx, J C; Buck, H; Burke, J; Cox, E; Browne, M.R. et al. The State of Carbon Dioxide Removal. 2nd Edition. TI - The State of Carbon Dioxide Removal. 2024. Available online: https://osf.io/f85qj/https://www.stateofcdr.org/ (accessed on 8 August 2024).
- GHR. Global Hydrogen Review 2023, IEA, Paris. Available online: https://www.iea.org/reports/global-hydrogen-review-2023 (accessed on 8 August 2024).
- Juárez-Orozco, S.M.; Siebe, C.; Fernández y Fernández, D. Causes and Effects of Forest Fires in Tropical Rainforests: A Bibliometric Approach. Trop. Conserv. Sci. 2017, 2017, 10. [Google Scholar] [CrossRef]
- Zheng, B.; Ciais, P.; Chevallier, F.; Yang, H.; Canadell, J.G.; Chen, Y.; van der Velde, I.R.; Aben, I.; Chuvielco, E.; et al. Record-high CO2 emissions from boreal fires in 2021. Science 2023, 379, 912–917. [Google Scholar] [CrossRef]
- Hong, X.; Liu, C.; Zhang, C.; Tian, Y.; Wu, H.; Yin, H.; Zhu, Y.; Yafang Cheng, Y. Vast ecosystem disturbance in a warming climate may jeopardize our climate goal of reducing CO2: a case study for megafires in the Australian ‘black summer’. Sci. Total Environ. 2023, 866, 161387. [Google Scholar] [CrossRef] [PubMed]
- Chikamoto, M.O.; DiNezio, P.; Lovenduski, N. Long-Term Slowdown of Ocean Carbon Uptake by Alkalinity Dynamics. Geophys. Res. Lett. 2023, 50, e2022GL101954. [Google Scholar] [CrossRef]
- World Ocean Review. The role of the ocean in the global carbon cycle. Available online: https://worldoceanreview.com/en/wor-8/the-role-of-the-ocean-in-the-global-carbon-cyclee/how-the-ocean-absorbs-carbon-dioxide/ (accessed on 2 August, 2024).
- Smolders, E.J.V.; Western, R.M.; Dijkstra, H.A. Probability Estimates of a 21st Century AMOC Collapse. Available online: https://arxiv.org/html/2406.11738v1 (accessed on 3 August 2024).
- Luedtke, J.A.; Chanson, J.; Neam, K.; Hobin, L.; Maciel, A.O.; Catenazzi, A.; Borzée, A.; Hamidy, A.; Aowphol, A.; Jean, A.; et al. Ongoing Declines for the World’s Amphibians in the Face of Emerging Threats. Nature 2023, 622, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Guirguis, J.; Goodyear, L.E.B.; Finn, C.; Johnson, J.V.; Pincheira-Donoso, D. Risk of Extinction Increases Towards Higher Elevations Across the World's Amphibians. Glob. Ecol. Biogeogr. 2023, 32, 1–12. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, D.; Wang, Liu, J. ; Du N. Altitudinal Patterns of Species Diversity and Phylogenetic Diversity across Temperate Mountain Forests of Northern China. PLOS ONE 2026, 1, e0159995. [Google Scholar] [CrossRef]
- Khatiwada, J.R.; Zhao, T.; Chen, Y.; Wang, B.; Xei, F.; Cannatella, D.C.; Jiang, J. Amphibian Community Structure along Elevation Gradients in Eastern Nepal Himalaya. BMC Ecol. 2019, 19, 1–11. [Google Scholar] [CrossRef]
- Rastegar-Pouyani, N; Gholamifard, A. ; Karamiani, R; Bahmani, Z.; Mobaraki, A.; Abtin E.; Faizi, H.; Heidari, N.; Takesh, M.; Sayyadi. F. et al. Browne RK. Sustainable Management of the Herpetofauna of the Iranian Plateau and coastal Iran. Amphib. Rept. Conser. 2015, 9, 1–15. [Google Scholar]
- Trew, B.T.; Edwards, D.P.; Klinges, D.H.; Early, R.; Svátel, M.; Plichta, R.; Mayula, R.; Okello, J.; Niessner, A.; Barthel, M.; et al. Novel Temperatures are Already Widespread Beneath the World’ Tropical Rainforests. Nat. Clim. Chang. 14, 753–759. [CrossRef]
- Blake, J.G.; Loiselle, B.A. Sharp Declines in Observation and Capture Rates of Amazon Birds in Absence of Human Disturbance. Glob. Ecol. Conserv. 2024, 51, e02902. [Google Scholar] [CrossRef]
- Campbell, L.G.; Anderson, K.A.; Marcec-Greaves, R.; Cardoso, P.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; et al. Scientists' Warning to Humanity on Insect Extinctions. Biol. Conserv. 2020, 242, 108426:1–108426:1. [Google Scholar] [CrossRef]
- Precautionary Principle. International Institute for Sustainable Development. Available online: https://www.iisd.org/articles/deep-dive/precautionary-principle 9acessed 13 August 2024).
- Gergis, J. An Intergenerational Crime Against Humanity What will it Take for Political Leaders to Start Taking Climate Change Seriously. Available online: https://theconversation.com/an-intergenerational-crime-against-humanity-what-will-it-take-for-political-leaders-to-start-taking-climate-change-seriously-231383. (accessed on 14 June 2024).
- Barry, B. Sustainability and Intergenerational Justice. Theoria 1997, 89, 43–65. [Google Scholar] [CrossRef]
- Spanning, R. Youth in the Anthropocene: Questions of Intergenerational Justice and Learning in a More-Than-Human World. In Youth Cultures in a Globalized World.; Knapp, G., Krall, H. Eds.; Springer, Cham, ZG, 2021; pp 113–133, ISBN 3030651762.
- Global Tipping Points. Section 4.6. Risks, Equity and Justice in the Governance of Positive Tipping Points. Pereira, L., Achieng, T., Castro, A., Sara M. Constantino, S.A., Ghadiali, A., Gifford, L., Newell, P., Smith, B., Smith, S.R., Villasante, S., Zimm, C. Eds.; Available online: https://global-tipping-points.org/ (accessed on 29 December 2023).
- Radchuk, V.; Reed, T.; Teplitsky, C.; van de Pol, M.; Charmantiet, A.; Hassall, C.; Adamik, P.; Adriaensen, F.; Ahola, M.P.; Arcese, P.; et al. Adaptive Responses of Animals to Climate Change are Most Likely Insufficient. Nat. Commun. 2019, 10, 3109. [Google Scholar] [CrossRef] [PubMed]
- Treves, A.; Artelle, K.A.; Darimont, C.T.; Lynn, W.S.; Paquet, P.; Santiago-Ávila, F.J.; Shaw, R.; Wood, M.C. Intergenerational Equity can Help to Prevent Climate Change and Extinction. Nat. Ecol. Evol. 2018, 2, 204–207. [Google Scholar] [CrossRef]
- Méjean, A.; Pottier, A.; Zuber, S.; Fleurbaey, M. Intergenerational Equity Under Catastrophic Climate Change. Global Priorities Institute Working Paper No. 5-2020. Availaable online: https://globalprioritiesinstitute.org/wp-content/uploads/M%C3%A9jean-et-al_Intergenerational-equity-under-catastrophic-climate-change.pdf (accesses 7 August 2024).
- Fedele, G.; Donatti, C.; I. , Harvey, C.A.; Hannah, L.; Hole, D.G. (2019). Transformative Adaptation to Climate Change for Sustainable Social-ecological Systems. Environ. Sci. Policy, 2019, 101, 116–125. [Google Scholar] [CrossRef]
- O’Brien, K. (2012). Global Environmental Change II: From Adaptation to Deliberate Transformation. Hum. Geogr. J. 2012, 36, 667–676. [Google Scholar] [CrossRef]
- Bolam, F.C.; Ahumada, J.; Akçakaya, H.R.; Brooks, T.M.; Elliott, W.; Hoban, S.; Mair, L.; Mallon, D.; McGowan, P.J.K.; Raimondo, D.; et al. Preventing Extinctions Post-2020 Requires Recovery Actions and Transformative Change. BioRx 2020. [Google Scholar] [CrossRef]
- Bowman, M. Chapter 1 Law, Legal Scholarship and the Conservation of Biological Diversity: 2020 Vision and Beyond. In Research Handbook on Biodiversity and Law; Bowman, M., Davies, P., Goodwin, E. Eds.; Edward Elgar Publishing, Cheltenham, United Kingdom, 2016. [CrossRef]
- Senior, R.A.; Bagwyn, R.; Leng, D.; Killion, A.K.; Jetz, W.; Wilcove, D.S. Global Shortfalls in Documented Actions to Conserve Biodiversity. Nature 2024, 630, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Bateman, I.; Balmford, A. Current Conservation Policies Risk Accelerating Biodiversity Loss. Nature 2023, 618, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Palacio, R.D.; Abarca, M.; Armenteras, D.; Balza, U.; Dollar, L.; Froese, G.Z.L.; Galligan, M.P.; Gula, J.; Giordano, A.J.; Jacobson, A.P.; et al. The Global Influence of the IUCN Red List can Hinder Species Conservation Efforts. Authorea 2023, preprint. [Google Scholar] [CrossRef]
- Crutzen, P.J.; Stoemer, E.F. The “Anthropocene”. The International Geosphere–Biosphere Programme (IGBP): A Study of Global Change of the International Council for Science (ICSU) Newsletter. Glob. Change Newsl. 2000, 41, 17–18. [Google Scholar]
- United Nations Convention on Biodiversity. Available online: https://www.un.org/en/observances/biological-diversity-day/convention (accessed on 27 September 2023).
- Byers, O.; Lees, C.; Wilcken, J.; Schwitzer, C. The One Plan Approach: The Philosophy and Implementation of CBSG’s Approach to Integrated Species Conservation Planning. WAZA Mag. 2013, 14, 2–5. [Google Scholar]
- Ziegler, T. The IUCN/SSC CPSG's One Plan Approach and the Role of Progressive Zoos in Conservation: Case Studies from Herpetology. In Proceedings of the 14th national congress of the Italian Society for Herpetology, Torino; 2023; pp. 195–222. [Google Scholar]
- IUCN SSC Amphibian Specialist Group (2024). Amphibian Conservation Action plan: A Status Review and Roadmap for Global Amphibian Conservation. Wren, S., Borzée. A., Marcec-Greaves, R. & Angulo, A. (Eds.). IUCN SSC Occasional Paper, No 57. Gland, Switzerland: IUCN. Available online: https://portals.iucn.org/library/sites/library/files/documents/SSC-OP-057-En.pdf (accessed on 6 August 2024).
- AArk. Amphibian Ark. Available online: www.amphibianark.org/ (accessed on 27th September 2023).
- Zippel, K.; Johnson, K.; Agliardo, R.G.; Gibson, R.; McFadden, M.; Browne, R.; Martinez, C.M.; Townsend, E. The Amphibian ARK: A Global Community for Ex situ Conservation of Amphibians. Herpetol. Conserv. Biol. 2011, 6, 340–352. [Google Scholar]
- Browne, R.K.; Janzen, P.; Bagaturov, M.F.; van Houte, D.K. Amphibian Keeper Conservation Breeding Programs. J. Zoo. Res. 2018, 2, 29–46. [Google Scholar] [CrossRef]
- Responsible Herpetological Project. conservation breeding programs. Available online: https://responsibleherpetoculture.foundation/ (accessed on 27 September 2023).
- Thomas-Walters, L.; McCallum, J.; Montgomery, R.; Petros, C.; Wan, A.K.Y.; Veríssimo, D. Systematic review of conservation interventions to promote voluntary behavior change. Conserv. Biol. 2023, 37, e14000. [Google Scholar] [CrossRef] [PubMed]
- Hagedorn, M.; Parenti, L.R.; Craddock, R.A.; Comizzoli, P.; Mabee, P.; Meinke, B.; Wolf, S.M.; Bischof, J.C.; Sandlin, R.D.; Tessier, S.N.; et al. Safeguarding Earth's biodiversity by creating a lunar biorepository. BioScience 2024, biae058. [Google Scholar] [CrossRef] [PubMed]
- Della-Tonga, G.; Howell, L.G.; Clulow, J.; Langhorne, C.J.; Marcec-Greaves, R.; Calatayud, N.E. Evaluating Amphibian Biobanking and Reproduction for Captive Breeding Programs According to the Amphibian Conservation Action Plan Objectives. Theriogenology 2020, 150, 412–431. [Google Scholar] [CrossRef]
- Clulow, J.; Mahony, M.; Browne, R.; Pomering, M.; Clark, A. Applications of Assisted Reproductive Technologies (ART) to Endangered Anuran Amphibians. In Declines and disappearances of Australian frogs; Campbell, A., Ed.; Environment Australia: Canberra, Australia, 1999; pp. 219–225. ISBN 0 642 54656 8. [Google Scholar]
- Browne, R.K.; Kaurova, S.A.; Vasudevan, K.; McGinnity, D.; Venu, G.; Gonzalez, M.; Uteshev, V.K.; Marcec-Greaves, R. Reproduction Technologies for the Sustainable Management of Caudata (salamander) and Gymnophiona (Caecilian) biodiversity. Reprod. Fert. Devel. 2022, 34, 479–497. [Google Scholar] [CrossRef]
- Ananjeva, N.B.; Uteshev, V.K.; Orlov, N.L.; Ryabov, S.A.; Gakhova, E.N.; Kaurova, S.A. , Kramarova L.I., Shishova N.V., Browne R.K. Comparison of the Modern Reproductive Technologies for Amphibians and Reptiles. Russ. J. Herpetol. 2017, 24, 275–290. [Google Scholar] [CrossRef]
- Clulow, J.; Upton, R.; Trudeau, V.L.; Clulow, S. Amphibian Assisted Reproductive Technologies: Moving from Technology to Application. Adv. Exp. Med. Biol. 2019, 1200, 413–463. [Google Scholar] [CrossRef] [PubMed]
- Uteshev, V.K.; Gakhova, E.N.; Kramarova, L.I.; Shishova, N.V.; Kaurova, S.A.; Kidova, E.A.; Kidov, A.A.; Browne, R.K. Russian Collaborative Development of Reproduction Technologies for the Sustainable Management of Amphibian Biodiversity. Asian Herpetol. Res. 2023, 14, 103–115. [Google Scholar] [CrossRef]
- Anastas, Z.M.; Byrne, P.G.; O’Brien, J.K.; Hobbs, R.J.; Upton, R.; Silla, A.J. The Increasing Role of Short-Term Sperm Storage and Cryopreservation in Conserving Threatened Amphibian Species. Animals 2023, 13, 2094:1–2094:40. [Google Scholar] [CrossRef]
- Browne, R.K.; Silla, A.J.; Upton, R.; Della-Togna, G.; Marcec-Greaves, R.; Shishova, N.V.; Uteshev, V.K.; Proano, B.; Pe-rez, O.D.; Mansour, N.; et al. Sperm Collection and Storage for the Sustainable Management of Amphibian Biodiversity. Theriogenology 2019, 133, 187–200. [Google Scholar] [CrossRef]
- Bolton, R.L.; Mooney, A.; Pettit, M.T.; Bolton, A.E.; Morgan, L.; Drake, G.J.; Appeltant, R.; Walker, S.L.; Gillis, J.D.; Hvilsom, C. Resurrecting Biodiversity: Advanced Assisted Reproductive Technologies and Biobanking. Reprod. Fertil. 2022, 3, R121–R146. [Google Scholar] [CrossRef] [PubMed]
- Seddon, P.J.; Griffiths, C.J.; Soorae, P.S.; Armstrong, D.P. Reversing Defaunation: Restoring Species in a Changing World. Science 2014, 345, 406–412. [Google Scholar] [CrossRef]
- Strand, J.; Thomsen, H.; Jensen, J.B.; Marcussen, C.; Nicolajsen, T.B.; Skriver, M.B.; Søgaard, I.M.; Ezaz, T.; Purup, S.; Callesen, H.; et al. Biobanking in Amphibian and Reptilian Conservation and Management: Opportunities and Challenges. Conserv. Gen. Res. 2020, 12, 709–725. [Google Scholar] [CrossRef]
- Strand, J.; Fraser, B.; Houck, M.L.; Clulow, S. Culturing and Biobanking of Amphibian Cell Lines for Conservation Applications. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CRC Press: London, UK, 2022; pp. 156–177. ISBN 9781486313334. [Google Scholar]
- Mooney, A.; Ryder, O.A.; Houck, M.L.; Staerk, J.; Conde, D.A.; Buckley, Y.M. Maximizing the Potential for Living Cell Banks to Contribute to Global Conservation Priorities. Zoo Biol. 2023, 42, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Cowl, V.B.; Comizzoli, P.; Appeltant, R.; Bolton, R.L.; Browne, R. Holt, W.V.; Penfold, L.M.; Swegen, A. et al. Cloning for the Twenty-First Century and Its Place in Endangered Species Conservation. Annu. Rev. Anim. Biosci. 2024, 12, 91–112. [Google Scholar] [CrossRef]
- Lermen, D.; Blömeke, B.; Browne, R.K.; Clarke, A.; Dyce, P.W.; Fixemer, T.; Fuhr, G.R.; Holt, W.V.; Jewgenow, K.; Lloyd, R.E.; et al. Meeting Review. Cryobanking of Viable Biomaterials: Implementation of New Strategies for Conservation Purposes. Molecular Ecology 2009, 18, 1030–1033. [Google Scholar] [CrossRef]
- Bowgen, K.M.; Kettel, E.F.; Butchart, S.H.M.; Carr, J.A.; Foden, W.B.; Magin, G.; Morecroft, M.D.; Smith, R.K.; Stein, B.A.; Sutherland, W.J.; et al. Conservation interventions can benefit species impacted by climate change, Biol. Conserv. 2022, 269, 109524. [Google Scholar] [CrossRef]
- Bolton, R.L.; Mooney, A.; Pettit, M.T.; Bolton, A.E.; Morgan, L.; Drake, G.J.; Appeltant, R.; Walker, S.L.; Gillis, J.D.; Hvilsom, C. Resurrecting Biodiversity: Advanced Assisted Reproductive Technologies and Biobanking. Reprod. Fertil. 2022, 3, R121–R146. [Google Scholar] [CrossRef]
- Upton, R.; Clulow, S.; Calatayud, N.E.; Colyvas, K.; Seeto, R.G.Y.; Wong, L.A.M.; Mahony, M.J.; Clulow, J. Generation of Reproductively Mature Offspring from the Endangered Green and Golden Bell Frog Litoria aurea using Cryopreserved Spermatozoa. Reprod. Fertil. Dev. 2021, 6. [Google Scholar] [CrossRef]
- Lampert, S.S.; Burger, I.J.; Julien, A.R.; Gillis, A.B.; Kouba, A.J.; Barber, D.; Kouba, C.K. Sperm Cryopreservation as a Tool for Amphibian Conservation: Production of F2 Generation Offspring from Cryo-Produced F1 Progeny. Animals 2023, 13, 53. [Google Scholar] [CrossRef]
- Holmes, B.; Ziermann, J.M.; Strzelecki, A.; Springer, S.; Zieger, M. Who notices Gymnophiona? Google Trends data reveal interesting trends for recent amphibian species. Ecol. Complex. 2024, 58, 101080. [Google Scholar] [CrossRef]
- Venu, G.; Raju, N.G.; Wilkinson, M.; Browne, R.K.; Varadh, K.; Balakrishna, G.N. , Ramakrishna, S.; Venkatachalaiah, G. First records of the Long-headed Caecilian, Ichthyophis longicephalus Pillai, 1986 (Gymnophiona: Ichthyophiidae) from the states of Karnataka and Tamil Nadu, India with comments on its conservation status. JAD 2020, 2, 5–10. Available online: https://jad.lu.ac.ir/article-1-93-en.html.
- Reinhard, S.; Kupfer, A. Maternal investment in the viviparous caecilian amphibian Typhlonectes natans (Gymnophiona: Typhlonectidae). Zool. Anz. 2022, 296, 33–36. [Google Scholar] [CrossRef]
- Kuehnel, S.; Kupfer, A. Sperm Storage in Caecilian Amphibians. Front Zool. 2012, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Parmley, D. General Husbandry of Terrestrial Caecilians. Available online: https://www.caudata.org/cc/articles/caecilian_care_Parmley.pdf, (accessed on 8 August 2024).
- Kaurova, S.A.; Chekurova, N.R.; Melnikova, E.V.; Uteshev, V.K.; Gakhova, E.N. 1996. Cryopreservation of Frog Rana temporaria Sperm Without Loss of Fertilizing Capacity. In Gakhova E. N., Karnaukhov V. N. Eds. Proceedings of the 14th Working Meeting, Pushchino, 13–15 May 1996, Pushchino: Pushchino Press, 106–108 (In Russian).
- Kaurova, S.A.; Uteshev, V.K.; Chekurova, N.R.; Gakhova, E.N. Cryopreservation of Testis of Frog Rana temporaria. Infusionsther Transfusionsmed 1997, 24, 379. [Google Scholar]
- Browne, R.K.; Clulow, J.; Mahony, M.; Clark, A. Successful Recovery of Motility and Fertility of Cryopreserved Cane Toad (Bufo marinus) sperm. Cryobiology 1998, 37, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Browne, R.K.; Clulow, J.; Mahony, M. Short-term Storage of Cane Toad (Bufo marinus) Gametes. Reproduction 2001, 121, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Karamura, T.; Nishioka, M. Reciprocal Diploid Nucleocytoplasmic Hybrids Between Two Species of Japanese Pond Frogs and their Offspring. J. Sci. Hiroshima Univ. 1963, 21, Ser.B. Div. 1., 65–84. [Google Scholar]
- Karamura, T.; Nishioka, M. Nucleo-cytoplasmic Hybrid Frog between Two Species of Japanese Brown Frogs and their Offspring. J. Sci. Hiroshima Univ. 1963, 21, Ser. B. Div. 1, 107–134. [Google Scholar]
- Kouba, C.K. ; Julien. A.R. Linking In situ and Ex situ Populations of Threatened Amphibian Species using Genome Resource Banks. In Reproductive Technologies and Biobanking for the Conservation of Amphibians, Silla, A.J., Kouba, A.J., H Heatwole, H., Eds.: CRC Press; London, 2022, pp. 188-203, ISBN 9781486313334.
- Taronga Conservation Society Australia. 2023, Available Online: https://taronga.org.au/conservation-and-science/current-research/frog-conservation-biobanking [accessed 20 September 2023).
- Panama Amphibian Conservation and Rescue Project. Available online: http://amphibianrescue.org/ (accessed on 13 August 2024).
- Burger, I.; Julien, A.R.; Kouba, A.J.; Councell, K.R.; Barber, B.; Pacheco, C.; Kouba, C.K. Linking In Situ and Ex Situ Populations of the Endangered Puerto Rican Crested Toad. Conserv. Sci. Pract. 2021, 3, e525. [Google Scholar] [CrossRef]
- Wyoming toad sees Recovery in the Southwest. Wyoming Game and Fish Department. Available online: https://wgfd.wyo.gov/News/Wyoming-toad-recovery-sees-success-in-southeast (accessed on 20 January 2024).
- Northern Corroboree Frog Recovery Program. Australia’s Nature Hub. Available online: https://www.australiasnaturehub.gov.au/action-inventory/northern-corroboree-frog-recovery-program (accessed on 20 January 2024).
- Browne, R.K. Book Review of ‘Reproductive Technologies and Biobanking for the Conservation of Amphibians’ by Silla, A. J., Kouba, A.J., Heatwole, H. (eds.); CSIRO Publishing: Victoria, Australia, 2022. 238 pp.; Herpetol. Rev. 2023, 54, 332–335. [Google Scholar]
- Martinez, A.; Mammola, S.S. Specialized Terminology Reduces the Number of Citations of Scientific Papers. Proc. R. Soc. B, Biol. Sci. 2021, 288. [Google Scholar] [CrossRef]
- Textor, M.; Rami, D. ; Proper Names: Philosophical and Linguistic Perspectives. Erkenn 2015, 80 (Suppl 2), 191–194. [Google Scholar] [CrossRef]
- Chala, D.; Endresen, D.; Demissew, S.; Slaughter, L. A.; Johnsen, E. B.; Stenseth, N. C. Address Social Injustices in Taxonomy: Implement Extended Revisions of Names with Ethical Issues and Persistent Identifiers for Tracing Name Changes. Preprints 2024, 2024071673. [Google Scholar] [CrossRef]
- Mc Cartney, A.M.; Head, M.A.; Tsosie, K.S.; Sterner, B.; Glass, J.R.; Paez, S.; Geary, J.; Hudson, M. Indigenous Peoples and Local Communities as Partners in the Sequencing of Global Eukaryotic Biodiversity. npj Biodivers. 2023, 2, 8. [Google Scholar] [CrossRef]
- ABS. Nagoya Protocol on Access and Benefit-sharing. 2014. Available online: https://www.cbd.int/abs/ (accessed on 6 August 2024).
- Khoday, K. Decolonising the Environment: Third World Approaches to the Planetary Crisis. Indones. J. Int. Law, 2022, 19, 189–211. [Google Scholar]
- Abiddin, N.Z.; Ibrahim, I.; Abdul Aziz, S.A. Non-Governmental Organisations (NGOs) and their Part towards Sustainable Community Development. Sustainability 2022, 14, 4386. [Google Scholar] [CrossRef]
- Adams, W.; Mulligan, M. Decolonising Nature; Strategies for Conservation in a Post-colonial Era. 2002. Routledge London. 2002, 320p, ISBN 9781853837494.
- Frankham, R. Genetics and Extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Williams, R.N.; Bos, D.H.; Gopurenko, D; Dewoody, J. A. Amphibian Malformations and Inbreeding. Biol. Lett. 2008, 4, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Brennan, R.S.; Garrett, A.D.; Huber, K.E.; Hargarten, H.; Pespeni, M.H. Rare Genetic Variation and Balanced Polymorphisms are Important for Survival in Global Change Conditions. Proc. R. Soc. B. 2019, 286, 20190943:1–20190943:9. [Google Scholar] [CrossRef] [PubMed]
- Ralls, K.; Ballou, J.D.; Dudash, M.R.; Eldridge, M.D.B.; Fenster, C.B.; Lacy, R.C.; Sunnucks, P.; Frankham, R. Call for a Paradigm Shift in the Genetic Management of Fragmented Populations. Conserv. Lett. 2018, 11, 1–6. [Google Scholar] [CrossRef]
- AArk Founder Numbers. Available online: https://www.amphibianark.org/conservation-programs/captive-programs/founder-animals/ (accessed on 2nd January 2023).
- Johnston, L.A.; Lacy, R.C. Genome Resource Banking for Species Conservation: Selection of Sperm Donors. Cryobiology 1995, 32, 68–77. [Google Scholar] [CrossRef]
- Amphibian Population Management Guidelines. Created at an Amphibian Population Management Workshop; 2007 December 10-11; San Diego, CA, USA. Schad, K., Ed.; Amphibian Ark, 2008, 31 p. Available online: https://www.amphibianark.org/wp-content/uploads/2018/08/AArk-Amphibian-Population-Management-Guidelines.pdf.
- Beebee, T.J.C. Conservation Genetics of Amphibians. Heredity 2005, 95, 423–427. [Google Scholar] [CrossRef]
- Johnson, W.E. Koepfli, K. The Role of Genomics in Conservation and Reproductive Sciences. In Reproductive Sciences in Animal Conservation. Holt, W., Brown, J., Comizzoli, P. Eds.; Springer, New York, NY. 2014, Vol 753, p 533.
- Farquharson, K.A.; Hogg, C.J.; Grueber, C.E. Offspring Survival Changes over Generations of Captive Breeding. Nat. Commun. 2021, 12, 3045. [Google Scholar] [CrossRef]
- Hinkson, K.M.; Poo, S. Inbreeding Depression in Sperm Quality in a Critically Endangered Amphibian. Zoo. Biol. 2020, 39, 197–204. [Google Scholar] [CrossRef]
- Hinkson, K.M.; Poo, S. Inbreeding Depression in Sperm Quality in a Critically Endangered Amphibian. Zoo Biol. 2020, 39, 197–204. [Google Scholar] [CrossRef]
- Conde, D.A.; Colchero, F.; Gusset, M.; Pearce-Kelly, P.; Byers, O.; Flesness, N.; Browne, R.K.; Jones, O.R. Zoos through the Lens of the IUCN Red List: A Global Metapopulation Approach to Support Conservation Breeding Pro-grams. PLOS One, 2013, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kardos. M.; Armstrong, E.E.;, Fitzpatrick, S.W.; Hauser, S.; Hedrick, P.W.; Miller, J.M.; Tallmon, D.A.; Funk, W.C. The Crucial Role of Genome-wide Genetic Variation in Conservation. Proc. Natl. Acad. Sci USA, 2021, 118, e2104642118. [Google Scholar] [CrossRef]
- Kouba, A.J. Genome Resource Banks as a Tool for Amphibian Conservation. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla A.J., Kouba A.J., Heatwole H., Eds.; CSIRO, Melbourne, Australia, 2022; pp. 204–220. ISBN 9781032372075.
- Onley, I.R.; Moseby, K.E.; Austin, J.J. Genomic Approaches for Conservation Management in Australia under Climate Change. Life 2021, 11, 653. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, V.; Barkha, S.; Venu, G.; Gowri, M.; Lisa, G. ; Kishor. G.B. Ex-situ Management of Amphibians in Indian Zoos. Central Zoo Authority, New Delhi, 2022; pp.102.
- Eskew, E.A.; Shock, B.C.; LaDouceur, E.E.B.; Keel, K.; Miller, M.R.; Foley, J.E.; Todd, B.D. Gene Expression Differs in Susceptible and Resistant Amphibians Exposed to Batrachochytrium dendrobatidis. R. Soc. Open. Sci. 2018, 5, 170910. [Google Scholar] [CrossRef] [PubMed]
- Hantak, M.M.; Kuchta, S.R. Predator Perception Across Space and Time: Relative Camouflage in a Colour Polymorphic Salamander, Biol. J. Linnean Soc. 2018, 123, 21–33. [Google Scholar] [CrossRef]
- Mahony, M.J.; Clulow, J. Appendix 2. Cryopreservation and Reconstitution Technologies: A Proposal to Establish A Genome Resource Bank For Threatened Australian Amphibians. In Guidelines for minimising disease risks associated with captive breeding, raising and restocking programs for Australian frogs; Murray, K., Skerratt, L.F., Marantelli, G., Berger, L., Hunter, D., Eds.; Canberra: Australian Government, Department of Sustainability, Environment, Water, Population and Communities, 2011, ID 1011-1151. [Google Scholar]
- FAO. 2012. Cryoconservation of Animal Genetic Resources. UN Food and Agriculture Organization. No. 12. Rome.
- Harnal, V.K.; Wildt, D.E.; Bird, D.M.; Monfort, S.L.; Ballou, J.D. Computer Simulations to Determine the Efficacy of Different Genome Resource Banking Strategies for Maintaining Genetic Diversity. Cryobiology, 2002, 44, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Witzenberger, K.A.; Hochkirch, A. Ex situ Conservation Genetics: a Review of Molecular Studies on the Genetic Consequences of Captive Breeding Programs for Endangered Animal Species. Biodivers. Conserv. 2011, 20, 1843–1861. [Google Scholar] [CrossRef]
- Crates, R.; Stojanovic, D.; Heinsohn, R. The Phenotypic Costs of Captivity. Biol. Rev. Camb. Philos. Soc. 2023, 98, 434–449. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Park, J.; Do, Y. Importance and Application of Amphibian Sperm Cryopreservation. J. Wetl. Res. 2023, 25, 257–266. [Google Scholar] [CrossRef]
- Howell, L.G.; Frankham, R.; Rodger, J.C.; Witt, R.R.; Clulow, S.; Upton, R.M.O.; Clulow, J. Integrating Biobanking Minimises Inbreeding and Produces Significant Cost Benefits for a Threatened Frog Captive Breeding Programs. Conserv. Lett. 2021, 14, e12776. [Google Scholar] [CrossRef]
- Howell, L.G.; Mawson, P.R.; Frankham, R.; Rodger, J.C.; Upton, R.M.O.; Witt, R.W.; Calatayud, N.E.; Clulow, S.; Clulow, J. Integrating Biobanking could Produce Significant Cost Benefits and Minimise Inbreeding for Australian Amphibian Captive Breeding Program. Reprod. Fert. Dev. 2021, 33, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, R.E.; Naydenova, E.; Proaño-Bolaños, C.; Vizuete, K.; Debut, A.; Arias, M.T.; Coloma, L.A. Development of Assisted Reproductive Technologies for the Conservation of Atelopus sp. (spumarius complex). Cryobiology 2022, 105, 20–31. [Google Scholar] [CrossRef]
- AArk Conservation Needs Assessments. 2023. Available online: https://www.conservationneeds.org/default.aspx (accessed on 23 September 2023).
- Rehberg-Besler, N.; Doucet, S.M.; Mennill, D.J. Vocal Behavior of the Explosively Breeding Neotropical Yellow Toad, Incilius luetkenii J. Herpetol. 2016, 50, 502–508. [Google Scholar] [CrossRef]
- Kupfer, A. Ch 5, Sexual size dimorphism in amphibians: an overview In Sex, Size and Gender Roles: Evolutionary Studies of Sexual Size Dimorphism Fairbairn, D.J., Wolf U. Blanckenhorn, W.U., Székely, T. Ed Oxford Academic Books, 2007, pp 50–59. [CrossRef]
- Liao, W.B.; Zeng, Y.; Yang, J.D. Sexual Size Dimorphism in Anurans: Roles of Mating System and Habitat Types. Front Zool. 2013, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.C.; Zamudio, K.R. Sexual Dichromatism in Frogs: Natural Selection, Sexual Selection and Unexpected Diversity. Proc. Biol. Sci. 2012, 279, 4687–4693. [Google Scholar] [CrossRef]
- Christe, P.; Keller, L.; Roulin, A. The Predation Cost of Being a Male: Implications for Sex-specific Rates of Ageing. Oikos 2006, 114, 381–384. [Google Scholar] [CrossRef]
- Han, X.; Fu, J. Does Life History Shape Sexual Size Dimorphism in Anurans? A Comparative Analysis. BMC Evol. Biol. 2013, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.J.; Tuttle, M.; Taft, LK. The Costs and Benefits of Frog Chorusing Behavior. Behav. Ecol. Sociobiol. 1981, 8, 273–278. [Google Scholar] [CrossRef]
- Watt, A.M.; Marcec-Greaves, R.; Hinkson, K.M.; Poo, S.; Roberts, B.; Pitcher, T.E. Effects of Age on Sperm Quality Metrics in Endangered Mississippi Gopher Frogs (Lithobates sevosus) from Captive Populations Used for Controlled Propagation and Reintroduction Efforts. Zoo Biol. 2021, 40, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Vieites, D.R.; Nieto-Román, S.; Barluenga, M.; Palanca, A.; Vences, M.; Meyer, A. Post-mating Clutch Piracy in an Amphibian. Nature 2004, 431, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Borowsky, R.; Luk, A.; He, X.; Kim, R.S. Unique Sperm Haplotypes are Associated with Phenotypically Different Sperm Subpopulations in Astyanax Fish. BMC Biol. 2018, 16, 72. [Google Scholar] [CrossRef]
- Scheltinga, D.M.; Jamieson, B.G.M. Spermatogenesis and the Mature Spermatozoon: Form, Function and Phylogenetic Implications, In Reproductive biology and phylogeny of Anura; Jamieson, G.M., Ed.; Science Publishers Inc.: NH, USA, 2013; Volume 2, pp. 119–252. ISBN 1-57808-288-9. [Google Scholar]
- Firman, R.C.; Gasparini, C.; Manier, M.K.; Pizzari, T. Postmating Female Control: 20 Years of Cryptic Female Choice. Trends Ecol. Evol. 2017, 32, 368–382. [Google Scholar] [CrossRef]
- Poo, S.; Bogisich, A.; Mack, M.; Lynn, B.K.; Devan-Song, A. Post-release Comparisons of Amphibian Growth reveal Challenges with Sperm Cryopreservation as a Conservation Tool. Conserv. Sci. Pract. 2022, 4, 5721–5729. [Google Scholar] [CrossRef]
- Hughes, DP.; Libersat, F. Parasite Manipulation of Host Behavior, Curr. Biol. 2019, 29, 45–47. [Google Scholar] [CrossRef]
- Browne, R.K.; Li, H.; Vaughan, M. Sexually Mediated Shedding of Myxobolus fallax Spores During Spermiation of Litoria fallax (anura). Dis. Aquat. Org. 2006, 72, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Brannelly, L.A.; Webb, R.; Skerratt, L.F.; Berger, L. Amphibians with Infectious Disease Increase their Reproductive Effort: Evidence for the Terminal Investment Hypothesis. Open Biol. 2016, 6, 150251. [Google Scholar] [CrossRef] [PubMed]
- Hartigan, A.; Phalen, D.N.; Slapeta, J. Myxosporean Parasites in Australian Frogs: Importance, Implications and Future Directions. Int. J. Parasitol Parasites Wildl. 2013, 2, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Eiras, J.C. An Overview on the Myxosporean Parasites in Amphibians and Reptiles. Acta Parasitol. 2005, 50, 267–275. [Google Scholar]
- Hartigan, A.; Phalen, D.N.; Šlapeta, J. Museum Material Reveals a Frog Parasite Emergence after the Invasion of the Cane Toad in Australia. Parasites Vectors, 2010, 3, 50. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, S.R.; Scheele, B.C.; Silla, A.J.; Keogh, S.; Hunter, D.A.; Endler, J.A.; Byrne, P.G. Disease influences male advertisement and mating outcomes in a critically endangered amphibian. Anim. Behav. 2021, 173, 145–157. [Google Scholar] [CrossRef]
- Narayan, E. 2022. Chapter 5. Non-invasive Monitoring of Stress Physiology During Management and Breeding of Amphibians in Captivity. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CRC Press: London, UK, 2022; pp. 49–68. ISBN 9781032372075. [Google Scholar]
- 94. Grummer, J.A.; Booker, T.R.; Matthey-Doret, R.; Nietlisbach, P.; Thomaz, A.T.; Whit-lock, M.C. The Immediate Costs and Long-term Benefits of Assisted Gene Flow in Large Populations. Conserv. Biol. 2022, 36, 13911. [Google Scholar] [CrossRef]
- Burger, I.J. The ART of Amphibian Conservation: Linking In-situ and ex-situ Populations of Endangered Species Through Genome Banking, 2021. MS Theses and Dissertations. 5379. https://scholarsjunction.msstate.edu/td/5379.
- Kelly, E.; Phillips, B.L. Targeted gene flow for conservation. Conserv. Biol. 2016, 30, 259–267. [Google Scholar] [CrossRef]
- Keller, L. F.; Waller, D. M. Inbreeding Effects in Wild Populations. Trends Ecol. Evol. 2002, 17, 230–241. [Google Scholar] [CrossRef]
- Brook, B.W.; Tonkyn, D.W.; O'Grady, J.J.; Frankham, R. Contribution of Inbreeding to Extinction Risk in Threatened Species. Conserv. Ecol. 2002, 6, 16. Available online: http://www.consecol.org/vol6/iss1/art16/138. [CrossRef]
- Stock, S.E.; Klop-Toker, K.; Wallace, S.; Kelly, O.; Callen, A.; Seeto, R.; Mahony, S.V.; Hayward, M.W.; Mahony, M.J. Uncovering Inbreeding, Small Populations, and Strong Genetic Isolation in an Australian Threatened Frog, Litoria littlejohni. Conserv. Genet. 2023, 24, 575–588. [Google Scholar] [CrossRef]
- Reed, D.H.; Frankham, R. Correlation Between Fitness and Genetic Diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Cheptou, P.O.; Hargreaves, A.L.; Bonte, D.; Jacquemyn, H. Adaptation to Fragmentation: Evolutionary Dynamics Driven by Human Influences. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160037. [Google Scholar] [CrossRef] [PubMed]
- Westram, A.M.; Stankowski, S.; Surendranadh, P.; Barton, N. What is Reproductive Isolation? J. Evol. Biol. 2022, 35, 1143–1164. [Google Scholar] [CrossRef] [PubMed]
- Nistelberger, H.M.; Roycroft, E.; Macdonald, A.J.; McArthur, S.; White, L.C.; Grady, P.G.S.; Pierson, J.; Sims, C.; Cowen, S.; Moseby, K.; et al. Genetic Mixing in Conservation Translocations Increases Diversity of a Keystone Threatened Species, Bettongia lesueur. Mol. Ecol. 2023, Epub ahead of print. [CrossRef]
- Albert, E.M.; Ferna´ndez-Beaskoetxea, S.; Godoy, J.A.; Tobler, U.; Schmidt, B.R.; Bosch, J. Genetic Management of an Amphibian Population after Chyridiomycosis Outbreak. Conserv. Genet. 2015, 16, 103–111. [Google Scholar] [CrossRef]
- Frankham, R.; Ballou, J.D.; Eldridge, M.D.; Lacy, R.C.; Ralls, K.; Dudash, M.R.; Fenster, C.B. Predicting the Probability of Outbreeding Depression. Conserv. Biol. 2011, 25, 465–475. [Google Scholar] [CrossRef]
- Lindsay, W.R.; Madsen, T.; Wapstra, E.; Lillie, M.; Loeb, L.; Ujvari, B.; Olsson, M. Long Term Effects of Outbreeding: Experimental Founding of Island Population Eliminates Malformations and Improves Hatching Success in Sand Lizards. Biol. Conserv. 2020, 249, 108710. [Google Scholar] [CrossRef]
- Frankham, R.; Bradshaw, C.J.A.; Brook, B.W. Genetics in Conservation Management: Revised Recommendations for the 50/500 rules, Red List Criteria and Population Viability Analysis. Biol. Conserv. 2014, 170, 56–63. [Google Scholar] [CrossRef]
- Silla, A.J.; Byrne, P.G. The importance of quantifying fitness-determining traits throughout life to assess the application of reproductive technologies for amphibian species recovery. Front. Conserv. Sci. 2024, 5. [Google Scholar] [CrossRef]
- Scheele, B.C.; Hunter, D.A.; Skerratt, L.F.; Brannelly, L.A.; Driscoll, D.A. Low Impact of Chytridiomycosis on Frog Recruitment Enables Persistence in Refuges Despite High Adult Mortality. Biol. Conserv. 2015, 182, 36–43. [Google Scholar] [CrossRef]
- Sgrò, C.M.; Lowe, A.J.; Hoffmann, A.A. Building Evolutionary Resilience for Conserving Biodiversity under Climate Change. Evol. Appl. 2011, 4, 326–337. [Google Scholar] [CrossRef]
- Carvalho, CS.; Lanes ECM. ; Silva AR.; Caldeira, CF.; Carvalho-Filho, N.; Gastauer, M.; Imperatriz-Fonsecaa, VL, Nascimento Jr, W.; Oliveeira, G.; Siqueira, J.O.; Viana, P.L. Jaffe, R. Habitat Loss Does Not Always Entail Negative Genetic Consequences. Front. Genet. 2019, 10, 1011. [Google Scholar] [CrossRef]
- Willi, Y.; Hoffmann, A.A. Demographic Factors and Genetic Variation Influence Population Persistence under Environmental Change. J. Evol. Biol. 2009, 22, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.; Phillips, B.L. Targeted Gene Flow for Conservation. Conserv. Biol. 2016, 30, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Tigano, A.; Friesen, V.L. ; Genomics of Local Adaptation with Gene Flow. Molec. Ecol. 2016, 25, 2144–2164. [Google Scholar] [CrossRef] [PubMed]
- Robert, A. Captive Breeding Genetics and Reintroduction Success. Biol. Conserv. 2009, 142, 2915–2922. [Google Scholar] [CrossRef]
- Calatayud, N.E.; Jacobs, L.; Della Togna, G.; Langhorne, C.J.; Mullen, A.C.; Upton, R. Hormonal Induction, Quality Assessments and the Influence of Seasonality on Male Reproductive Viability in a Long-term Managed Ex Situ Breeding Colony of Southern Rocky Mountain Boreal Toads, Anaxyrus boreas boreas. bioRxrv preprint, 2023. [CrossRef]
- Edmands, S. Between a Rock and a Hard Place: Evaluating the Relative Risks of Inbreeding and Outbreeding for Conservation and Management. Molec. Ecol. 2007, 16, 463–475. [Google Scholar] [CrossRef]
- Earnhardt, J.M. Reintroduction Programmes: Genetic Trade-offs for Populations. Anim. Conserv. 2006, 2, 279–286. [Google Scholar] [CrossRef]
- Byrne, P.G.; Silla, A.J. Genetic Management of Threatened Amphibians: Using Artificial Fertilisation Technologies to Facilitate Genetic Rescue and Assisted Gene Flow. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla A.J., Kouba A.J., Heatwole H., Eds.; CSIRO, Melbourne, Australia, 2022; pp. 124–146, ISBN 9781032372075.
- Byrne, P.G.; Keogh, J.S.; O'Brien, D.M.; Gaitan-Espitia, J.D.; Silla, A.J. Evidence that genetic compatibility underpins female mate choice in a monandrous amphibian. Evolution 2021, 75, 529–541. [Google Scholar] [CrossRef]
- Spielman, D.; Brook, B. W.; Frankham, R. Most Species are not Driven to Extinction before Genetic Factors Impact Them. PNAS, 2004, 101, 15261–15264. [Google Scholar] [CrossRef]
- Teixeira, J.C.; Huber, C.D. The Inflated Significance of Neutral Genetic Diversity in Conservation Genetics. Proc. Nat. Acad. Sci. 2021, 118, e2015096118. [Google Scholar] [CrossRef] [PubMed]
- Walter, H.S. The Mismeasure of Islands: Implications for Biogeographic Theory and the Conservation of Nature. J. Biogeog. 2004, 31, 177–197. [Google Scholar] [CrossRef]
- Revive and Restore. The Black-footed Ferret Project. Available online: https://reviverestore.org/projects/black-footed-ferret/ (accessed on 23 September 2023).
- Scheele, B.C.; Guarino, F.; Osborne, W.; Hunter, D.A.; Skerratt, L.F.; Driscoll, D.A. Decline and Re-expansion of an Amphibian with High Prevalence of Chytrid Fungus. Biol. Conserv. 2014, 170, 86–91. [Google Scholar] [CrossRef]
- O'Hanlon, S.J.; Rieux, A.; Farrer, R.A.; Rosa, G.M.; Waldman, B.; Bataille, A.; Kosch, T.A.; Murray, K.A.; Brankovics, B.; Fumagalli, M.; et al. Recent Asian Origin of Chytrid Fungi Causing Global Amphibian Declines. Science 2018, 360, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Dreitz, V.J. Issues in Species Recovery: An Example Based on the Wyoming Toad: Forum. BioScience 2006, 56, 765–771. [Google Scholar] [CrossRef]
- Scheele, B.C.; Hunter, D.A.; Skerratt, L.F.; Brannelly, L.A.; Driscoll, D.A. Low Impact of Chytridiomycosis on Frog Recruitment Enables Persistence in Refuges Despite High Adult Mortality. Biol. Conserv. 2015, 182, 36–43. [Google Scholar] [CrossRef]
- Burger, I.; Julien, A.R.; Kouba, A.J.; Councell, K.R.; Barber, B.; Pacheco, C.; Kouba, C.K. Linking In Situ and Ex Situ Populations of the Endangered Puerto Rican Crested Toad (Peltophryne lemur) through Genome Banking. Conserv. Sci. Pract. 2021, 3, e525. [Google Scholar] [CrossRef]
- Viggers, K.L.; Lindenmayer, D.B.; Spratt, D.M. The Importance of Disease in Reintroduction Programmes. Wildlife Research 1993, 20, 687–698. [Google Scholar] [CrossRef]
- Ballou, J.D. Assessing the risks of infectious diseases in captive breeding and reintroduction programs. J. Zoo. Wildl. Med. 1993, 24, 327–335. [Google Scholar]
- Chaudhary, V.; Oli, M.K. A Critical Appraisal of Population Viability Analysis. Conserv. Biol. 2019, 34, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Willi, Y.; Kristensen, T.N.; Sgrò, C.M.; Weeks, A.R.; Ørsted, M.; Hoffmann, A.A. Conservation genetics as a management tool: The Five Best-supported Paradigms to Assist the Management of Threatened Species. Proc Natl Acad Sci USA. 2022, 119, e2105076119. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R. Do Island Populations Have Less Genetic Variation than Mainland Populations? Heredity 1997, 78, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Fauvel, C.; Suquet, M.; Cosson, J. Evaluation and Determinism of Fish Sperm Quality. J. Appl. Ichthyol. 2010, 26, 636e43. [Google Scholar] [CrossRef]
- EDGE. Evolutionary Distinct and Globally Endangered. Zoological Society of London. Available online: https://www.edgeofexistence.org/species/ (accessed on 1 August 2023).
- Khosla, K.; Kangas, J.; Liu, Y.; Zhan, L.; Daly, J.; Hagedorn, M.; Bischof, J. Cryopreservation and Laser Nanowarming of Zebrafish Embryos Followed by Hatching and Spawning. Adv. Biosyst. 2020, 4, e2000138. [Google Scholar] [CrossRef]
- de Siqueira-Silva, D.H.; Saito, T.; dos Santos-Silva, A.P.; da Silva Costa, R.; Psenicka, M.; Yasui, G.S. Biotechnology Applied to Fish Reproduction: Tools for Conservation. Fish Physiol. Biochem. 2018, 44, 1469–1485. [Google Scholar] [CrossRef] [PubMed]
- Browne, R.K. Figiel, C. Chapter 2. Crypreservation in Amphibians In: Cryopreservation of Aquatic Species. Vol 8. Tiersch, T., Mazik, P. Eds.: World Aquaculture Society, Baton Rouge, La Hardcover, 2011, Vol 8, ~450 pages.
- Behera, B.J. Surrogacy Technology in Fisheries and Aquaculture. In: Current Trends in Fisheries Biotechnology. Behera, B.K. Ed.; Springer, 2024. [CrossRef]
- Browne, R.K.; Zippel, K.; Odum, A.R.; Herman, T. Physical Facilities and Associated Services. Use of Amphibians in Research, Laboratory, or Classroom Settings. Inst. Lab. Anim. Res. (ILAR), 2007, 48, 188–202. [Google Scholar] [CrossRef]
- AArk Reproduction Technologies videos. Available online: https://www.amphibianark.org/art-videos/ (accessed on 23 September 2023).
- Hamer, A.J.; Mahony, M.J. 2007. Life History of an Endangered Amphibian Challenges the Declining Species Paradigm. Australian J. Zool. 2007, 55, 79–88. [Google Scholar] [CrossRef]
- Richter, S.C.; Seigel, R.A. Annual Variation in the Population Ecology of the Endangered Gopher Frog, Rana sevosa Goin and Netting. Copeia 2002, 2002, 962–972. [Google Scholar] [CrossRef]
- McGinnity, D.; Reinsch, S.R.; Schwartz, H.; Trudeau, V.; Browne, R.K. Semen and Egg Collection, Sperm Cryopreservation, and in vitro Fertilisation with Threatened North American Giant Salamanders (Cryptobranchus alleganiensis). Reprod. Fertil. Dev. 2021, 34, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Lötters, S.; Plewnia, A.; Catenazzi, A.; Neam, K.; Acosta-Galvis, A.R.; Vela, Y.A.; Allen, J.P.; Segundo, J.O.A.; de Lourdes Almendáriz Cabezas, A. Barboza, G.A.; et al. Ongoing Harlequin Toad Declines Suggest the Amphibian Extinction Crisis is Still an Emergency. Commun. Earth Environ. 2023, 4, 412. [Google Scholar] [CrossRef]
- Valencia, L.M.; Fonte, L.F. The Fate of Harlequin Toads – Help Through a Synchronous Approach and the IUCN ‘Amphibian Conservation Action Plan? Oryx 2007, 39, 343–346. [Google Scholar] [CrossRef]
- Valencia, L.M.; Fonte, L.F. Atelopus Survival Initiative IUCN SSC ASG Atelopus Task Force. 2021. Available online: https://www.atelopus.org/_files/ugd/9db650_60f3e6095cbf4b1dabb7376a4fb88366.pdf (accessed on 7 August 2024).
- Buckner, J.C.; Sanders, R.C.; Faircloth, B.C.; Chakrabarty, P. The Critical Importance of Vouchers in Genomics. Elife 2021, 10, e68264. [Google Scholar] [CrossRef]
- Andreone, D.; Raselimanana, A.P. Crottini, A, Vouchering, Integrative Taxonomy and Natural History Collections: a Case Study with the Amphibians of Madagascar. Boll. Mus. Reg. Sci. Nat. Toronto 2023, 40(1–2), 41–57.
- Rocha, L.A.; Aleixo, A.; Allen, G.; Almeda, F.; Baldwin, C.C.; Barclay, M.V.L.; Bates, M.; Bauer, A.M.; Benzoni, F.; Berns, C.M.; et al. Specimen Collection: An Essential Tool. Science 2014, 344, 814–815. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hopkins, A.J.M.; Davis, R.A. Going, Going, Gone The Diminishing Capacity of Museum Specimen Collections to Address Global Change Research: A Case Study on Urban Reptiles. Animals 2023, 13, 1078. [Google Scholar] [CrossRef]
- Powell, D.M.; Meyer, T.G. , Duncan, M. By Bits and Pieces: The Contributions of Zoos and Aquariums to Science and Society via Biomaterials. J. Zool. Bot. Gard. 2023, 4, 277–287. [Google Scholar] [CrossRef]
- Goncharov, B.F.; Shubravy, O.I.; Serbinova, I.A.; Uteshev, V.K. The USSR programme for breeding amphibians, including rare and endangered species. Int. Zoo Yearbook, 1989, 28, 10–21. [Google Scholar] [CrossRef]
- World Animal Protection. Available online: https://www.worldanimalprotection.org/ (accessed on 27 July 2024).
- Rakesh, R.K. Seasonal Cycle in Anuran (Amphibia) Testis: The Endocrine and Environmental Controls. Italian J. Zool. 1976, 43(1-2), 151–172.
- Rastogi, R.K.; Tammaro, L.; Di Meglio, M.; Lela, L.; Di Matteo, l.; Giovanni Chieffi, G. Circannual Testicular Rhythm in the Green Frog, Rana esculenta. Italian J. Zool. 1981, 48, 97–105. [Google Scholar] [CrossRef]
- Ulloa, J.S.; Aubin, T.; Llusia, D.; Courtois, E.A.; Fouquet, A.; Gaucher, P.; Pavoine, S.; Sueur, J.; et al. Explosive Breeding in Tropical Anurans: Environmental Triggers, Community Composition and Acoustic Structure. BMC Ecol. 2019, 19, 28. [Google Scholar] [CrossRef]
- Kaurova, S.A.; Uteshev, V.K.; Gapeyev, A.B.; Shishova, N.V.; Gakhova, E.N.; Browne, R.K.; Kramarova, L.I. Cryopreservation of Spermatozoa Obtained Postmortem from the European Common Frog Rana temporaria. Reprod. Fert. Dev. 2021, 33. [Google Scholar] [CrossRef]
- Figiel Jr., C. R. Effects of Water Temperature on Gonads Growth in Ambystoma mexicanum Axolotl Salamanders. Animals 2023, 13, 874. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, D. M.; Silla, A. J.; Forsythe, P. S.; Byrne, P. G. Sex Differences in Response to Environmental and Social Breeding Cues in an Amphibian. Behaviour 2021, 158, 397–426. [Google Scholar] [CrossRef]
- Edwards, D.L.; Mahony, M.J.; Clulow, J. Effect of Sperm Concentration, Medium Osmolality and Oocyte Storage on Artificial Fertilisation Success in a Myobatrachid Frog (Limnodynastes tasmaniensis). Reprod. Fertil. Dev. 2004, 16, 347–354. [Google Scholar] [CrossRef]
- Upton, R.; Clulow, S.; Colyvas, K.; Mahony, M.; Clulow, J. Paradigm Shift in Frog Sperm Cryopreservation: Reduced Role for Non-penetrating Cryoprotectants. Reproduction 2023, 165, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Upton, R.; Clulow, S.; Mahony, M.J.; and Clulow, J. Generation of a Sexually Mature Individual of the Eastern Dwarf Tree Frog Litoria fallax, from Cryopreserved Testicular Macerates: Proof of Capacity of Cryopreserved Sperm Derived Offspring to Complete Development. Conserv. Physiol. 2018, 6, coy043. [Google Scholar] [CrossRef] [PubMed]
- Silla, A.J.; Keogh, L.M.; Byrne, P.G. Sperm Motility Activation in the Critically Endangered Booroolong Frog: the Effect of Medium Osmolality and Phosphodiesterase Inhibitors. Reprod. Fertil. Dev. 2017, 29, 2277–2283. [Google Scholar] [CrossRef] [PubMed]
- Germano, J.M.; Cree, A; Molinia, F. C; Arregui, L.; Bishop, P.J. Hormone Treatment does not Reliably Induce Spermiation or Mating in Hamilton’s frog from the Archaic Leiopelmatid Lineage. Reprod. Fert. Devel. 2021, 34, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Hormone therapy. Available online: https://en.wikipedia.org/wiki/Hormone_therapy (accessed on 17th July, 20240 (accessed on 7 August 2024).
- Miyamoto, K.; Simpson, D.; Gurdon, J.B. Manipulation and in vitro Maturation of Xenopus laevis Eggs, Followed by Intracytoplasmic Sperm Injection, to Study Embryonic Development. J. Vis. Exp. 2015, 96, 52496. [Google Scholar] [CrossRef]
- Herbert, D. Studies of Assisted Reproduction in the Spotted Grass Frog Limnodynastes tasmaniensis: Ovulation, Early Development and Microinjection (ICSI). Masters by Research thesis, 2004, The University of Newcastle, Australia.
- Ishibashi, S.; Kroll, K.L.; Amaya, E. Generation of Transgenic Xenopus laevis: III. Sperm Nuclear Transplantation. Cold Spring Harb. Protoc. 2007, 2007:pdb.prot4840. [CrossRef]
- Parmar, M.S.; Pant, C.; Karuppanasamy, K.; Mili, B.; Upadhyay, D.; Kant, V. Intracytoplasmic Sperm Injection (ICSI) and its Applications in Veterinary Sciences: An Overview. Sci. Int. 2013, 1, 266–270. [Google Scholar]
- Silla, A.J.; Byrne, P.G. Hormone-induced Ovulation and Artificial Fertilisation in Four Terrestrial-breeding Anurans. Reprod. Fertil. Dev. 2021, 33, 615–618. [Google Scholar] [CrossRef]
- Browne, R.K.; Seratt, J.; Vance, C.; Kouba, A. Hormonal Priming, Induction of Ovulation and In-vitro Fertilization of the Endangered Wyoming Toad (Bufo baxteri). Reprod. Biol. Endocrinol. 2006, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Marcec-Greaves, R.M.; Kouba, C.K.; Willard, S.T.; Andy, J. Kouba, A.J. Ovarian Ultrasound Analysis for Developing Temporal and Spatially Explicit Hormone Regimens for Induced Ovulation and Egg Deposition in the Tiger Salamander (Ambystoma tigrinum), Theriogenology Wild, 2023, 2,2023, 100038. [CrossRef]
- Shishova, N.V.; Uteshev, V. K.; Sirota, N.P.; Kuznetsova, E.A.; Kaurova, S.A.; Browne, R.K.; Gakhova, E.N. The Quality and Fertility of Sperm Collected from European Common Frog (Rana temporaria) Carcasses Refrigerated for up to 7 Days. Zoo Biol. 2013, 32, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Silla, A.J.; Roberts, J.D.; Byrne, P.G. The Effect of Injection and Topical Application of hCG and GnRH Agonist to Induce Sperm-release in the Roseate Frog, Geocrinia rosea. Conserv. Physiol. 2020, 8, coaa104. [Google Scholar] [CrossRef] [PubMed]
- Marcec, R.M. Development of Assisted Reproductive Technologies for Endangered North American Salamanders by Ruth Marie Marcec. Faculty of Mississippi State University, Degree of Doctor of Philosophy in Animal Physiology, 2016; Mississippi, MS, USA.
- Coxe, N.; Liu, Y.; Arregui, L.; Upton, R.; Bodenstein, S.; Voss, S.R.; Gutierrez-Wing, M.T.; Tiersch, T.R. Establishment of a Practical Sperm Cryopreservation Pathway for the Axolotl (Ambystoma mexicanum): A Community-Level Approach to Germplasm Repository Development. Animals 2024, 14, 206. [Google Scholar] [CrossRef] [PubMed]
- Guy, E.L.; Gillis, A.B.; Kouba, A.J.; Barber, D.; Poole, V.; Marcec-Greaves, R.M.; Kouba, C.K. Sperm Collection and Cryopreservation for Threatened Newt Species. Cryobiology 2020, 94, 80–88. [Google Scholar] [CrossRef]
- Chen, D.M.; Kouba, C.K.; Songsasen, N.; Roth, T.L.; Allen, P.J.; Kouba, A.J. Comparing novel sperm extenders for the internally-fertilizing tiger salamander (Ambystoma tigrinum). Front. Amphib. Reptile Sci. 2023, 1, 1320803. [Google Scholar] [CrossRef]
- Julien, A.R.; Kouba, A.J.; Kabelik, D.; Feugangm, J.M.; Willard, S.T.; Kouba, C.K. Nasal administration of gonadotropin releasing hormone (GnRH) elicits sperm production in Fowler’s toads (Anaxyrus fowleri). BMC Zool. 2019, 4, doi. [Google Scholar] [CrossRef]
- Campbell, L.G.; Anderson, K.A.; Marcec-Greaves, R.M. Topical Application of Hormone Gonadotropin-releasing Hormone (GnRH-A) Stimulates Reproduction in the Endangered Texas Blind Salamander (Eurycea rathbuni). Conserv. Sci. Pract. 2021, 4, e609. [Google Scholar] [CrossRef]
- Toledo RC, Jared C. Cutaneous adaptations to water balance in amphibians. Comp. Biochem. Physiol. B Biochem. Physiol. 1993, 105, 593–608. [Google Scholar] [CrossRef]
- Chen, D.M.; Chen, L.D.; Kouba, C.K.; Songsasen, N.; Roth, T.L.; Allen, P.J.; Kouba, A.J. Oral Administration of GnRH via a Cricket Vehicle Stimulates Spermiation in Tiger Salamanders (Ambystoma tigrinum). PLoS ONE, 2024, 19, e0289995. [Google Scholar] [CrossRef]
- John Clulow, Melissa Pomering, Danielle Herbert, Rose Upton, Natalie Calatayud, Simon Clulow, Michael J. Mahony, Vance L. Trudeau. Differential success in obtaining gametes between male and female Australian temperate frogs by hormonal induction: A review. Gen. Comp. Endocrinol. 2028, 265, 141–148. [CrossRef]
- Trudeau, V.L.; Raven, B.H.; Pahuja, H.K.; Narayan, E. J 2022. Chapter 4. Hormonal Control of Amphibian Reproduction. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CRC Press: London, UK, 2022; pp. 49–68. ISBN 9781032372075. [Google Scholar]
- Browne, R.K.; Li, H.; Seratt, J.; Kouba. A. Progesterone Improves the Number and Quality of Hormone Induced Fowler toad (Bufo fowleri) Oocytes. Reprod. Biol. Endocrinol. 2006, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mansour, N.; Lahnsteiner, F.; Patzner, R.A. Collection of Gametes from Live Axolotl, Ambystoma mexicanum, and Standardization of In Vitro Fertilization. Theriogenology 2011, 75, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Uteshev, V.K.; Kaurova, S.A.; Shishova, N.V.; Stolyarov, S.D.; Browne, R.K.; Gakhova, E.N. In Vitro Fertilization with Hormonally Induced Sperm and Eggs from Sharp-ribbed Newt Pleurodeles waltl. Russ. J. Herpetol. 2015, 22, 35–40. [Google Scholar] [CrossRef]
- Graham, K.M.; Langhorne, C.J.; Vance, C.K.; Willard, S,T. ; Kouba, A.J. Ultrasound Imaging Improves Hormone Therapy Strategies for Induction of Ovulation and In Vitro Fertilization in the Endangered Dusky Gopher Frog (Lithobates sevosa). Conserv. Physiol. 2018, 6, coy020. [Google Scholar] [CrossRef]
- Bogolyubova, I.O.; Bogolyubov, D.S. ; Effect of Hormonal Stimulation on the Oocyte Chromosomal Apparatus in the Common Frog. J. Evol. Biochem. Phys. 2023, 59, 1229–1236. [Google Scholar] [CrossRef]
- Browne, R.K.; Wang, Z.; Okada, S.; McGinnity, D.; Luo, Q.; Taguchi Y.; Kilpatrick, D.; Hardman, R.; Janzen, P.; Zhang, Z, Geng Y. The Sustainable Management of Giant Salamanders (Cryptobranchoidea). Report, Sustainability America: Belize, 2020.
- Silla, A.J.; Langhorne, C.J. Protocols for Hormonally Induced Spermiation, and the Cold Storage, Activation, and Assessment of Sperm. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla, A.J., Kouba, A.J., H Heatwole, H., Eds.: London: CRC Press; London, 2022; pp. 106-123.
- Burger, I.J.; Chen, L.; Lampert, S.S.; Kouba, C.K.; Barber, D.; Smith, D.; Cobos, C.; Kouba, A.J. Applying Sperm Collection and Cryopreservation Protocols Developed in a Model Amphibian to Three Threatened Anuran Species Targeted for Biobanking Management. Biol. Conserv. 2023, 277, 109850:1–109850:6. [Google Scholar] [CrossRef]
- Arregui, L.; Martinez-Pastor, F.; Arroyo, F.; Gosálvez, J. Determining the Effects of Sperm Activation in Anuran Cloaca on Motility and DNA Integrity in Epidalea calamita (Bufonidae). Reprod. Fertil. Dev. 2022, 34, 438–446. [Google Scholar] [CrossRef]
- Valencia, L.C.; García, A.; Ramírez-Pinilla, M.P.; Fuentes, J.L. Estimates of DNA Damage by the Comet Assay in the Direct-developing Frog Eleutherodactylus johnstonei (Anura, Eleutherodactylidae). Genet. Mol. Biol. 2011, 34, 681–688. [Google Scholar] [CrossRef]
- Shishova, N.V.; Uteshev, V.K.; Kaurova, S.A.; Browne, R.K.; Gakhova, E.N. Cryopreservation of Hormonally Induced Sperm for the Conservation of Threatened Amphibians with Rana temporaria as a Model Research Species. Theriogenology 2011, 75, 220–232. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Upton, R.; Calatayud, N.E.; Silla, A.J.; Daly, J.; McFadden, M.S.; O’Brien, J.K. Cryopreservation Cooling Rate Impacts Post-thaw Sperm Motility and Survival in Litoria booroolongensis. Animals (Basel) 2023, 13, 3014. [Google Scholar] [CrossRef] [PubMed]
- Browne, R.K.; Venu, G.; Kaurova, S.A. The Case for Considering the Term ‘Mitochondrial Vesicle’ as a Misnomer in Publications about Assisted Reproductive Technologies (ART) for Amphibians. Reprod. Fert. Develop. 2023. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.Y.; Jamieson, B.G.M. The Ultrastructure of the Spermatozoa of Bufonid and Hylid Frogs (Anure, Am-phibia): Implications for Phylogeny and Fertilisation Biology. Zool. Scripta 1993, 22, 309–323. [Google Scholar] [CrossRef]
- Chen, D.M.; Moore, M.G.; Willis, E.L.; Kouba, A.J.; Kouba, C.K. The Impact of Time and Environmental Factors on the Mitochondrial Vesicle and Subsequent Motility of Amphibian Sperm. Comp. Biochem. Physiol. A: Molec. Integ. Physiol. 2022, 111191. [Google Scholar] [CrossRef] [PubMed]
- Figiel Jr., C. R. Cold Storage of Sperm from the Axolotl, Ambystoma mexicanum. Herpetol. Conserv. Biol. 2022, 15, 367–371. [Google Scholar]
- Silla, A.J.; Keogh, L.M.; Byrne, P.G. Antibiotics and Oxygen Availability Affect the Short-term Storage of Spermatozoa from the Critically Endangered Booroolong Frog, Litoria booroolongensis. Reprod. Fertil. Dev. 2015, 27, 1147–1153. [Google Scholar] [CrossRef] [PubMed]
- Germano, J.M.; Arregui, L.; Kouba, A.J. Effects of Aeration and Antibiotics on Short-term Storage of Fowler’s toad (Bufo fowleri) Sperm. Aquaculture 2013, 396–399, 20–24. [Google Scholar] [CrossRef]
- Kaurova, S.A.; Browne, R.K.; Uteshev, V.K. Antibiotics for the Refrigerated Storage at 4°C of Hormonally Induced European Common Frog (Rana temporaria) Spermatozoa. Theriogenology Wild 2022, 1, 100009. [Google Scholar] [CrossRef]
- Kaurova, S.A.; Shishova, N.V.; Uteshev, V.K. The Effect of Gentamicin on the Motility of Hormonally Induced Spermatozoa of Toad Bufo bufo during Storage at 4°C. Bull. Exp. Biol. Med. 2023, 176, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Kaurova, S.A.; Uteshev, V.K. , Shishova, N.V. Effect of Antibiotics Metranidazole, Streptomycin, and Gentamicin on the Maintenance of Sperm Motility of the European Common Frog (Rana temporaria) During Refrigerated Storage. Russ. J. Herpetol. 2024, 31, 47–55. [Google Scholar] [CrossRef]
- Poo, S.; Hinkson, K.M. Applying Cryopreservation to Anuran Conservation Biology. Conserv. Sci. Pract. 2019, 1, e91. [Google Scholar] [CrossRef]
- Kaneko, T.; Ito, H.; Sakamoto, H.; Onuma, M.; Inoue-Murayama, M. Sperm Preservation by Freeze-drying for the Conservation of Wild Animals. PLoS One 2014, 9, e113381. [Google Scholar] [CrossRef] [PubMed]
- Shishova, N.V.; Kaurova, S.A.; Uteshev, V.K.; Gakhova, E.N. Comparison of Cryoresistance of Testicular and Urinal Spermatozoa of the Toad Bufo bufo (Amphibia, Anura, Bufonidae) during Slow Freezing. Inland Water Biol. 2023, 16, 788–792. [Google Scholar] [CrossRef]
- Liu, M.; Yuan, S.; Zhao, Z.; Liu, M.; Lin, A.; Gong, Q. Non-programmable Cryopreservation of Sperm from Industrial-farmed Murray Cod (Maccullochella peelii). Aquaculture 2021, 541, 736811. [Google Scholar] [CrossRef]
- Arregui, L.; Bóveda, P.; Gosálvez, J.; Kouba, A.J. Effect of Seasonality on Hormonally Induced Sperm in Epidalea calamita (Amphibia, Anura, Bufonidae) and its Refrigerated and Cryopreserved Storage. Aquaculture 2020, 529, e735677. [Google Scholar] [CrossRef]
- Silla, A.J.; Kouba, A.J. Chapter 1. Integrating Reproductive Technologies into the Conservation Toolbox for the Recovery of Amphibian Species. In Reproductive Technologies and Biobanking for the Conservation of Amphibians, Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CRC Press: London, UK, 2022; pp 1-9, ISBN 9781486313334.172.
- Clulow, J.; Upton, R.; Clulow, S. 2022. Cryopreservation of Amphibian Genomes: Targeting the Holy Grail, Cryopreservation of Maternal-haploid and Embryonic-Diploid Genomes. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla, A.J., Kouba, A.J., H Heatwole, H., Eds.: London: CRC Press; London, 2022; pp. 147-165.
- Uteshev, V.K.; Gakhova, E.N.; Kramarova, L.I.; Shishova, N.V.; Kaurova, S.A.; Browne, R.K. Refrigerated Storage of European Common Frog Rana temporaria Oocytes. Cryobiology 2018, 83, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Wolf, DP.; Hendrick, J.L. A Molecular Approach to Fertilisation. II. Viability and Artificial Fertilisation of Xenopus laevis Gametes. Dev. Biol. 1971, 25, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Hollinger, TG.; Corton, G.L. Artificial Fertilization of Gametes from the South African Clawed Frog, Xenopus laevis. Gamete Res. 1980, 3, 45–57. [Google Scholar] [CrossRef]
- Gagarinskiy, E.; Uteshev, V.; Fesenko, E.Jr. Prolonged hypothermic storage of oocytes of the European Common Frog Rana temporaria in a Gas Mixture of Oxygen and Carbon Monoxide. PLoS ONE 2023, 18, e0288370:1–e0288370:10. [Google Scholar] [CrossRef] [PubMed]
- Firman, R.C.; Gasparini, C.; Manier, M.K.; Pizzari, T. Postmating Female Control: 20 Years of Cryptic Female Choice. Trends Ecol. Evol. 2017, 32, 368–382. [Google Scholar] [CrossRef]
- Ukita, M.; Itoh, T.; Watanabe, T.; Watanabe, A.; Onitake, K. Substances for the Initiation of Sperm Motility in Egg-jelly of the Japanese Newt, Cynops pyrrhogaster. Zool. Sci. 1999, 16, 793–802. [Google Scholar] [CrossRef]
- Olson, J.H.; Chandler, D.E. Xenopus laevis Egg Jelly contains Small Proteins that are essential to fertilization. Dev. Biol. 1999, 210, 401–410. [Google Scholar] [CrossRef]
- Omata, S. Relative Roles of Jelly Layers in Successful Fertilization of Bufo japonicus. J. Exp. Zool. 1993, 265, 329–335. [Google Scholar] [CrossRef]
- Ukita, M.; Itoh, T.; Watanabe, T.; Watanabe, A.; Onitake, K. Substances for the Initiation of Sperm Motility in Egg-jelly of the Japanese Newt, Cynops pyrrhogaster. Zool. Sci. 1999, 16, 793–802. [Google Scholar] [CrossRef]
- Comizzoli, P. Biotechnologies for Wildlife Fertility Preservation, Anim. Front. 2015, 5, 73–78. [Google Scholar] [CrossRef]
- Kouba, A.; Vance, C.; Willis, E. Artificial Fertilization for Amphibian Conservation: Current Knowledge and Future Considerations. Theriogenology, 2009, 71, 214–227. [Google Scholar] [CrossRef]
- Taniguchi-Sugiura, Y.; Tanaka, E.M. Artificial Insemination in Axolotl. Methods Mol. Biol. 2023, 2562, 417–423. [Google Scholar] [CrossRef]
- Rugh 1962. Experimental Embryology: Techniques and Procedures. 3rd Edn. BurgessPublishing Company, Minneapolis MN, USA.
- Hoitsy, G.A. Woynarovich, T. Moth-Poulsen, T. Guide to the Small-scale Artificial Propagation of Trout. The FAO Regional Office for Europe and Central Asia, Budapest, Hungary. 2012; p 11, Para 3.
- Rugh, R. Induced Ovulation and Artificial Fertilisation in the Frog. Biol. Bull. 1934, 66, 22–29. [Google Scholar] [CrossRef]
- Wolf, D.P.; Hendrick, J.L. 1971. A molecular Approach to Fertilisation: II, Viability and Artificial Fertilisation of Xenopus laevis Gametes. Devel. Biol. 1971, 25, 348–359. [Google Scholar] [CrossRef]
- Hollinger, TG. , Corton GL. Artificial Fertilization of Gametes of the African Clawed Frog, Xenopus laevis. Gamete Res. 1980, 3, 45–57. [Google Scholar] [CrossRef]
- Cabada, M.O. Sperm Concentration and Fertilisation Rate in Bufo arenarum (Amphibia: Anura). J. Exp. Biol. 1975, 62, 481–486. [Google Scholar] [CrossRef]
- Silla, A.J. Artificial Fertilisation in a Terrestrial Toadlet (Pseudophryne guentheri): Effect of Medium Osmolarity, Sperm Concentration and Gamete Storage. Reprod. Fert. Devel. 2013, 25, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Byrne, P.G.; Anastas, Z.M.; Silla, A.J. A test for plasticity in sperm motility activation in response to osmotic environment in an anuran amphibian. Ecol Evol. 2022, 12, e9387. [Google Scholar] [CrossRef] [PubMed]
- Hollinger, T.G.; Corton, G.L. Artificial Fertilization of Gametes from the South African Clawed Frog, Xenopus laevis. Gamete Res. 1980, 3, 45–57. [Google Scholar] [CrossRef]
- Browne, R.K.; Kaurova, S.A.; Uteshev, V.K.; Shishova, N.V.; Kramarova, L.; McGinnity, D.; Figiel, C.R.; Mansour, N.; Agnew, D.; Wu, M.; et al. Sperm Motility of Externally Fertilizing Fish and Amphibians. Theriogenology 2015, 83, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Burger, I.J.; Lampert, S.S.; Kouba, C.K.; Morin, D.J.; Kouba, A.J. Development of an Amphibian Sperm Biobanking Protocol for Genetic Management and Population Sustainability. Conserv. Physiol. 2022, 10, coac032. [Google Scholar] [CrossRef]
- Strand, J.; Callesen, H.; Pertoldi, C.; Purupm, S. 2022. Amphibian cell lines: Usable tissue types and differences between individuals within a species. Amphib. Rept. Conserv. 2022, 16 [General Section]: 245–256.
- Nikitina, L.A. Nuclear Transplantation in Amphibians. Physiol. Gen. Biol. Rev. 1997, 13, 35–72. [Google Scholar]
- Kaurova, S.A.; Nikitina, L.A.; Uteshev, V.K.; Gakhova, E.N. Cryopreservation of Totipotent Embryo Cells and Their use in Reconstruction of Enucleated Eggs, In Proceedings of the 15th Working Meeting, Pushchino, 13–15, October 1998; Gakhova E., Karnaukhov V.N. Eds.; Pushchino Press, 1998; pp. 206–208 (In Russian).
- Briggs, R.; King, T.J. Transplantation of Living Nuclei from Blastula Cells into Enucleated Frogs’ Eggs. PNAS 1952, 38, 455–453. [Google Scholar] [CrossRef]
- Sambuichi, H. The Roles of the Nucleus and the Cytoplasm in Development. I. An Interspecific Hybrid Frog, Developed from a Combination of Rana nigromaculata nigromaculata Cytoplasm and a Diploid Nucleus of Rana nigromacula tabrevipoda. J. Sci. Hiroshima Univ. 1957, Ser. B. Div. 17, 33–41. [Google Scholar]
- Moore, J.A. Transplanation of Nuclei Between Rana pipiens and Rana sylvatica. Exp. Cell. Res. 1958, 14, 532–540. [Google Scholar] [CrossRef]
- Sambuichi, H. The Roles of the Nucleus and the Cytoplasm in Development. III. Diploid Nucleocytoplasmic Hybrids, Derived from Rana nigromaculata brevipoda Cytoplasm and Rana nigromaculata nigromaculata Nuclei. J. Sci. Hiroshima Univ. 1961, 20, 1–5.
- Hennan, S. Nucleocytoplasmic hybrids between Rana pipiens and Rana palustris. I. Analysis of the Developmental Properties of the Nuclei by Means of Nuclear Transplantation. Dev. Biol. 1963, 11, 243–267. [Google Scholar] [CrossRef] [PubMed]
- Toshijiro, K.; Nishioka, M. Viabiity and Abnormalities of the Offspring of Nucleo-cytoplasmic Hybrids between Rana japonica and Rana ornativentris. Sci. Rep. Lab. Amphib. Biol. Hiroshima Univ. 1972, 1, 95–209. [Google Scholar]
- Nishioka, M. Abnormalities of the Offspring of Nucleo-cytoplasmic Hybrids Between Rana nigromaculata and Rana brevipoda. Sci. Rel. Lab. Amphib. Biol. Hiroshima Univ.
- Nishioka, M. Nucleo-cytoplasmic Hybrids between Rana japonica and Rana temporaria temporaria. Sci. Rep. Lab. Amphibian Biol. Hiroshima Univ. 1972, 1, 211–243. [Google Scholar]
- Nishioka, M. Nucleo-cytoplasmic Hybrids between Rana brevipoda and Rana plancyichosenica. Rep. Lab. Amphibian Biol. Hiroshima Univ. 1972, 1, 245–257. [Google Scholar]
- Gallien, L.; Aimar, C. On a New Mode of Gemelarity, Produced by Nuclear Grafting in Amphibians (Urodeles) of the Genus Pleurodeles. Acad. Sci. 1972, 272, 3348–3351. [Google Scholar]
- Gallien, C.L.; Aimar, C.; Guillet, F. Nucleocytoplasmic Interactions During Ontogenesis in Individuals Obtained by Intra- and Interspecific Nuclear Transplantation in the Genus Pleurodeles (Urodele Amphibian). Develop. Biol. 1973, 33, 154–170. [Google Scholar] [CrossRef]
- De-extinction. Available online: https://colossal.com/de-extinction/ (accessed on 16 March 2024).
- Gurdon, J.B. Adult Frogs Derived from the Nuclei of Single Somatic Cells. Devel. Biol. 1962, 4, 250–273. [Google Scholar] [CrossRef]
- "The Nobel Prize in Physiology or Medicine 2012". Available online: https://www.nobelprize.org/prizes/medicine/2012/summary/ (accessed on 8 October 2012).
- Gurdon, J.B.; Byrne, J.A. The First Half-century of Nuclear Transplantation. Proc. Natl. Acad. Sci. USA. 2003, 100, 8048–8052. [Google Scholar] [CrossRef]
- Gurdon, J.B. The Egg and the Nucleus: A Battle for Supremacy. Development 2013, 140, 2449–2456. [Google Scholar] [CrossRef]
- Rangel, G. From Corgis to Corn a Brief Look at the Long History of Gmo-technology, 2015, Available online:. Available online: https://sitn.hms.harvard.edu/flash/2015/from-corgis-to-corn-a-brief-look-at-the-long-history-of-gmo-technology/ (accessed on 27 September 2023).
- Kosch, T.A.; Silva, C.N.S.; Brannelly, L.A.; Roberts, A.A.; Lau, Q. [ Marantelli, G.; Berger, L.F.L.; Skerratt L.F. Genetic Potential for Disease Resistance in Critically Endangered Amphibians Decimated by Chytridiomycosis. Anim. Conserv. 2019, 22, 238–250. [Google Scholar] [CrossRef]
- Eskew, E.A.; Shock, B.C.; LaDouceur, E.E.B.; Keel, K.; Miller, M.R.; Foley, J.E.; Todd, B.D. Gene Expression Differs in Susceptible and Resistant Amphibians Exposed to Batrachochytrium dendrobatidis. R. Soc. Open Sci. 2018, 5, 170910:1–170910:23. [Google Scholar] [CrossRef] [PubMed]
- Silla, A.J.; Calatayud, N.E.; Trudeau, V.L. Amphibian Reproductive Technologies: Approaches and Welfare Considerations. Conserv. Physiol. 2021, 9, coab011. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, M. Capturing Wild Animal Welfare: a Physiological Perspective. Biol. Rev. 2024, 99, 1–22. [Google Scholar] [CrossRef]
- Singer, P. 10th Anniversary Edition of The Life You Can Save. Available online: https://www.thelifeyoucansave.org/the-book/ (accessed on 22 July 2024).
- Irwin, L.N. Renewed Perspectives on the Deep Roots and Broad Distribution of Animal Consciousness. Front. Syst. Neurosci. 2020, 14, 57. [Google Scholar] [CrossRef]
- Lambert, H.; Elwin, A.; D’Cruze, N. Frog in the Well: A Review of the Scientific Literature for Evidence of Amphibian Sentience. Appl. Anim. Behav. Sci. 2022, 247, 105559. [Google Scholar] [CrossRef]
- Winlow, W.; Mather, J.; Cosmo, A.D. Postscript to Invertebrate Welfare: “We Have Met the Enemy and He Is Us”. Animals 2024, 14, 2082. [Google Scholar] [CrossRef] [PubMed]
- Lambert, H.; Elwin, A.; D’Cruze, N. Frog in the well: A Review of the Scientific Literature for Evidence of Amphibian Sentience. Appl. Anim. Behav. Sci. 2022, 247, 105559. [Google Scholar] [CrossRef]
- Offor, I. Second Wave Animal Ethics and (Global) Animal Law: a View from the Margins. J. Hum. Rights Environ. 2020, 11, 268–296. [Google Scholar] [CrossRef]
- Ikegbu, E.A.; Diana-Abasi, F.I. Utilitarianism as a Veritable Vehicle for the Promotion of a Just Society. LWATI: J. Contemp. Res. 2017, 14, 121–137. [Google Scholar]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Universities Federation for Animal Welfare: UFAW, Wheathampstead, UK, 1959; p. 238. ISBN 0-900767-78-2. [Google Scholar]
- Tannenbaum, J.; Bennett, B.T. Russell and Burch's 3Rs Then and Now: The Need for Clarity in Definition and Purpose. J. Am. Assoc. Lab. Anim. Science, 2015, 54, 120–132. [Google Scholar]
- NC3R. National Center for the Replacement, Reduction, and Refinement of Animals in Research. Available online: https://www.nc3rs.org.uk/who-we-are/3rs (accessed on 23 September 2023).
- Woolly Mammoth De-extinction Project & Process | Colossal. Available online: https://colossal.com/mammoth/ (accessed on 27 September 2023).
- van Urk-Costa, E. Introduction - Why (Reformed) Theology Needs Reflection on Biodiversity Loss and Extinction. J. Reform. Theol. 2023, 17, 121–123. [Google Scholar] [CrossRef]
- Link, H.J. Playing God and the Intrinsic Value of Life: Moral Problems for Synthetic Biology? Sci. Eng. Ethics 2013, 19, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Dabrock, P. Playing God? Synthetic Biology as a Theological and Ethical Challenge. Syst. Synth. Biol. 2009, 3, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Peters, T. Playing God” and Germline Intervention, Appl. Anim. Behav. Sci., 1995, 20, 365–386. [Google Scholar] [CrossRef]
- Negi, C.S. Religion and biodiversity conservation: not a mere analogy. Inter. J. Biodiv. Sci. Manag. 2005, 1, 85–96. [Google Scholar] [CrossRef]
- White, T.B.; Petrovan, S.O.; Christie, A.P.; Martin, P.A.; Sutherland, W.J. What is the Price of Conservation? A Review of the Status Quo and Recommendations for Improving Cost Reporting, BioScience 2022, 72, 461–471. [Google Scholar] [CrossRef]
- Rowling, M.; How much will it cost to save nature - and who will pay? World Economic Forum and the Thomas Reuters Foundation Trust.org. Available online: https://www.weforum.org/agenda/2021/05/biodiversity-deforestation-global-investment-inititive/ (accessed on 2 August 2024).
- Gordon, E.R.; Butt, N.; Rosner-Katz, H.; Binley, A.D.; Bennett, J.R. Relative Costs of Conserving Threatened Species Across Taxonomic Groups. Conserv. Biol. 2019, 34, 276–281. [Google Scholar] [CrossRef]
- Balmford, A.; Gaston, K.J.; Rodrigues, A.S.L.; James. A. Integrating Costs of Conservation into International Priority Setting. Conserv. Biol. 2013, 14, 597–605. [Google Scholar] [CrossRef]
- Conde-Pueyo, N.; Vidiella, B.; Sardanyés, J.; Berdugo, M.; Maestre, F.T.; De Lorenzo, V.; Solé, R. Synthetic Biology for Terraformation Lessons from Mars, Earth, and the Microbiome. Life 2020, 10, 14. [Google Scholar] [CrossRef]
- Gualandris-Parisot L, Husson D, Foulquier F, Kan P, Davet J, Aimar C, Dournon C, Duprat AM. Pleurodeles waltl, Amphibian, Urodele, is a Suitable Biological Model for Embryological and Physiological Space Experiments on a Vertebrate. Adv Space Res. 2001, 28, 569–578. [CrossRef]
- Jacobson, S.K.; McDuff, M.D.; Monroe, M.C. Conservation Education and Outreach Techniques. In 1st edn, Techniques in Ecology & Conservation; Oxford Academic, 2007, doi.org/10.1093/acprof:oso/9780198567714.001.0001, (accessed 19 July 2024).
- Meredith, H.M.R.; St. John, F.A.V.; Collen, B.; Black, S.A.; Griffiths R.A. Practitioner and scientist perceptions of successful amphibian conservation. Conserv. Biol. 2018, 32, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Bradfield, K.S.; Tapley, B.; Johnson, B. Amphibians and Conservation Breeding Pro-grammes: How do we Determine Who Should be on the Ark? Biodiver. Conserv. 2023, 32, 885–898. [Google Scholar] [CrossRef]
- The Nature Education Knowledge Project. Saving Endangered Species: A Case Study Using Global Amphibian Declines. Available online: https://www.nature.com/scitable/knowledge/library/saving-endangered-species-a-case-study-using-19445898/ (accessed on 8 August 2024).
- Rothschild, J. Ethical Considerations of Gene Editing and Genetic Selection. J. Gen. Fam. Med. 2020, 21, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Frieze, C. Cloning Wild Life - Zoos, Captivity, and the Future of Endangered Animals. 2013, NYU Press, USA, 258 p. 350.
- Dabrock, P. Playing God? Synthetic Biology as a Theological and Ethical Challenge. Syst. Synth. Biol. 2009, 3, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Link, H.J. Playing God and the Intrinsic Value of Life: Moral Problems for Synthetic Biology? Sci. Eng. Ethics 2013, 19, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Loy, I.; Worley, W. Your jargon-busting climate glossary. 2023, Available online:. Available online: https://www.thenewhumanitarian.org/news-feature/2023/12/01/oh-ffs-guide-climate-change-acronyms. (accessed on 8 August 2024).
- Charo, A. Charo: Bioengineering Can Help Heal the World. https://colossal.com/charo-bioengineering-can-help-heal-the-world/ (accessed 10 August 2024).
- Lann, B. Petition to the United Federation of Planets: Revert the Ban on Gene Editing for Ecological Applications. Available online: https://www.reddit.com/r/startrek/comments/1df045s/petition_to_the_united_federation_of_planets/?rdt=37953 (accessed on 10 August 2024).
- Schwägerl, C. The Anthropocene – The Human Era and How it Shapes the Planet. Synergetic Press, 2015, pp 236. ISBN: 9780907791546. [CrossRef]
- Malhi, Y. The Concept of the Anthropocene. Ann. Rev. Environ. Resour. 2017, 42, 77–104. [Google Scholar] [CrossRef]
- Tapley, B.; Bradfield, B.; Michaels, C.; Bungard, M. Amphibians and Conservation Breeding Programmes: do all Threatened Amphibians Belong on the Ark? Biodivers. Conserv. 2015, 24, 2625–2645. [Google Scholar] [CrossRef]
Figure 1.
(Left panel) illustrates the proportional heterozygosity/genetic diversity (y-axis) through random sampling of individuals (x-axis) for a founding population. Figure 1 (right panel) shows the probability that an allele will persist in the next generation (x-axis) dependent on the number of equally representative reproducing individuals in the captive population (x-axis).
Figure 1.
(Left panel) illustrates the proportional heterozygosity/genetic diversity (y-axis) through random sampling of individuals (x-axis) for a founding population. Figure 1 (right panel) shows the probability that an allele will persist in the next generation (x-axis) dependent on the number of equally representative reproducing individuals in the captive population (x-axis).
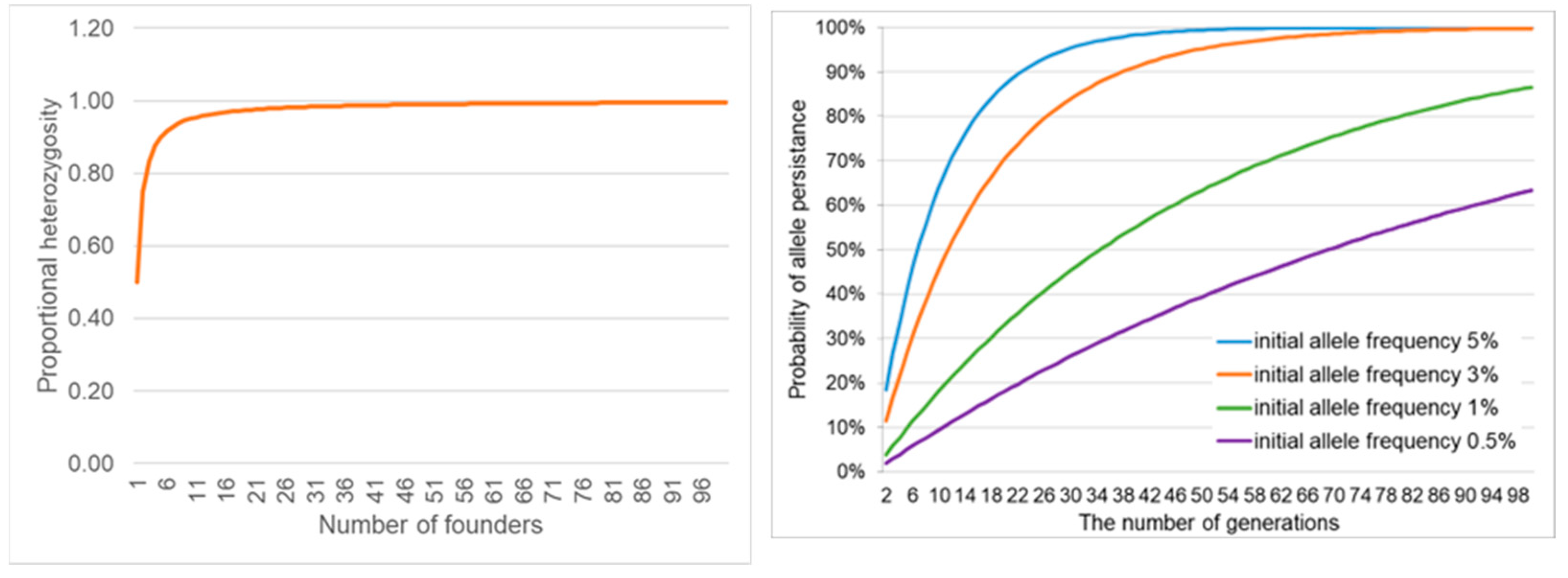
Figure 2.
The green and golden bell frog, Ranoidea (Litoria) aurea, varies in its green and gold colouration in natural populations. Left, most common morph (image-© 2008 Dr. Peter Janzen)); Right green morph (image- © 2004 Graeme R. Gillespie).
Figure 2.
The green and golden bell frog, Ranoidea (Litoria) aurea, varies in its green and gold colouration in natural populations. Left, most common morph (image-© 2008 Dr. Peter Janzen)); Right green morph (image- © 2004 Graeme R. Gillespie).

Figure 3.
Left, Atelopus bomolocus (Image – © University of Kansas); Middle, A. certus (Image –© 2019 Brian Gratwicke); Right, A. zeteki (Image – © Peter Janzen).
Figure 3.
Left, Atelopus bomolocus (Image – © University of Kansas); Middle, A. certus (Image –© 2019 Brian Gratwicke); Right, A. zeteki (Image – © Peter Janzen).

Figure 4.
The average oocyte numbers per anuran clutch by IUCN Redlist endangerment status.
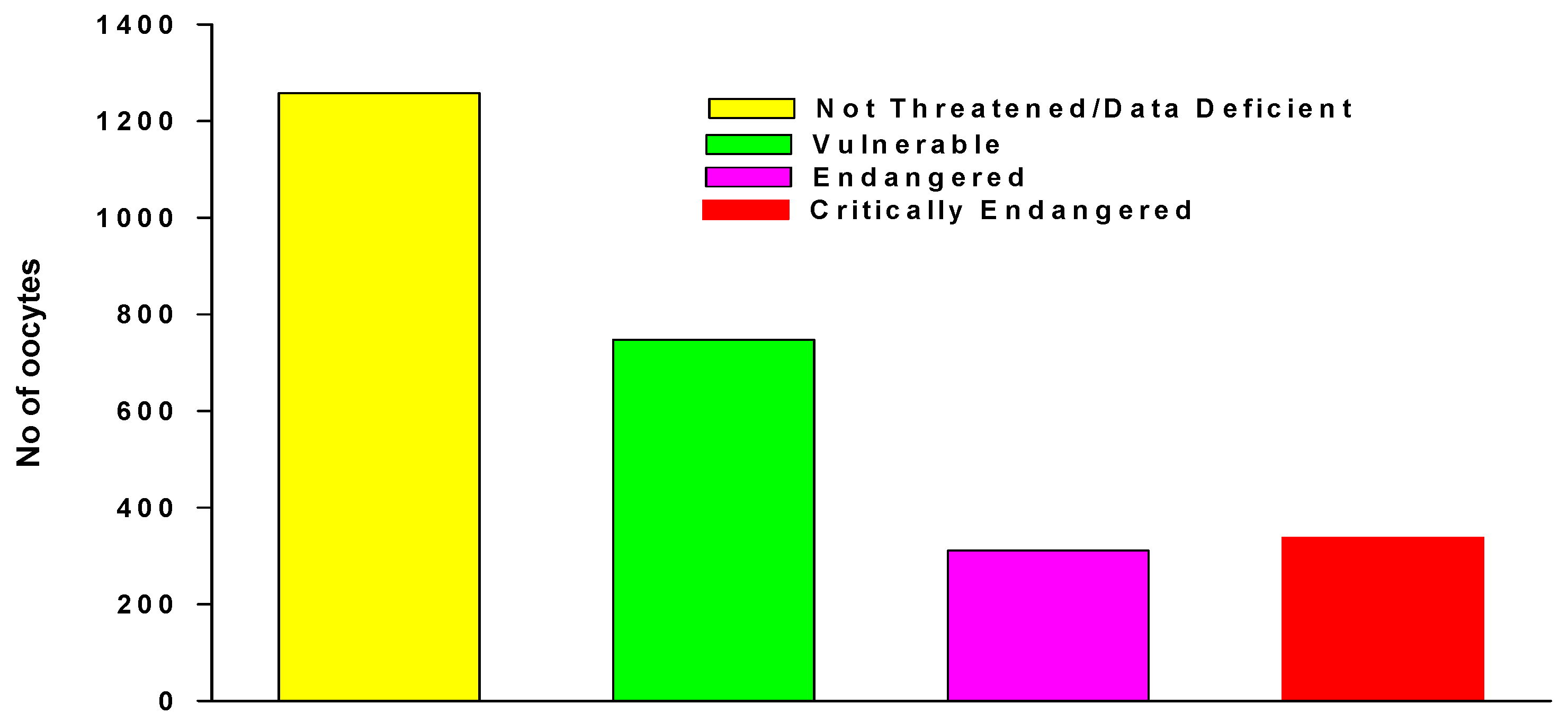
Figure 5.
Left, blue morph; Middle red morph; Right, yellow morph. Different colour morphs of the strawberry poison dart frog, Oophaga pumilio, and many other amphibian species are very popular with private carers and have a long history of domestication (Images Peter Janzen).
Figure 5.
Left, blue morph; Middle red morph; Right, yellow morph. Different colour morphs of the strawberry poison dart frog, Oophaga pumilio, and many other amphibian species are very popular with private carers and have a long history of domestication (Images Peter Janzen).

Figure 6.
Hamilton’s frog (Leiopelma hamiltoni). Image by Phil Bishop. Attribution ShareAlike 2.5.https://en.wikipedia.org/wiki/Hamilton%27s_frog#/media/File:Leiopelma_hamiltoni02.jpg Phil Bishop, CC BY-SA 2.5 <https://creativecommons.org/licenses/by-sa/2.5>, via Wikimedia Commons.
Figure 6.
Hamilton’s frog (Leiopelma hamiltoni). Image by Phil Bishop. Attribution ShareAlike 2.5.https://en.wikipedia.org/wiki/Hamilton%27s_frog#/media/File:Leiopelma_hamiltoni02.jpg Phil Bishop, CC BY-SA 2.5 <https://creativecommons.org/licenses/by-sa/2.5>, via Wikimedia Commons.

Figure 7.
The x-axis shows the number of anuran species, and the y-axis shows oocyte volumes (mm3). Left. 805 species with oocyte diameters to 4.9 mm and volumes to 31.3 mm3. (b) Right. ~430 of these 805 species from the first two bars in the left figure with oocyte diameters ≤ 1.8 mm and volumes ≤ 5.0 mm3.
Figure 7.
The x-axis shows the number of anuran species, and the y-axis shows oocyte volumes (mm3). Left. 805 species with oocyte diameters to 4.9 mm and volumes to 31.3 mm3. (b) Right. ~430 of these 805 species from the first two bars in the left figure with oocyte diameters ≤ 1.8 mm and volumes ≤ 5.0 mm3.
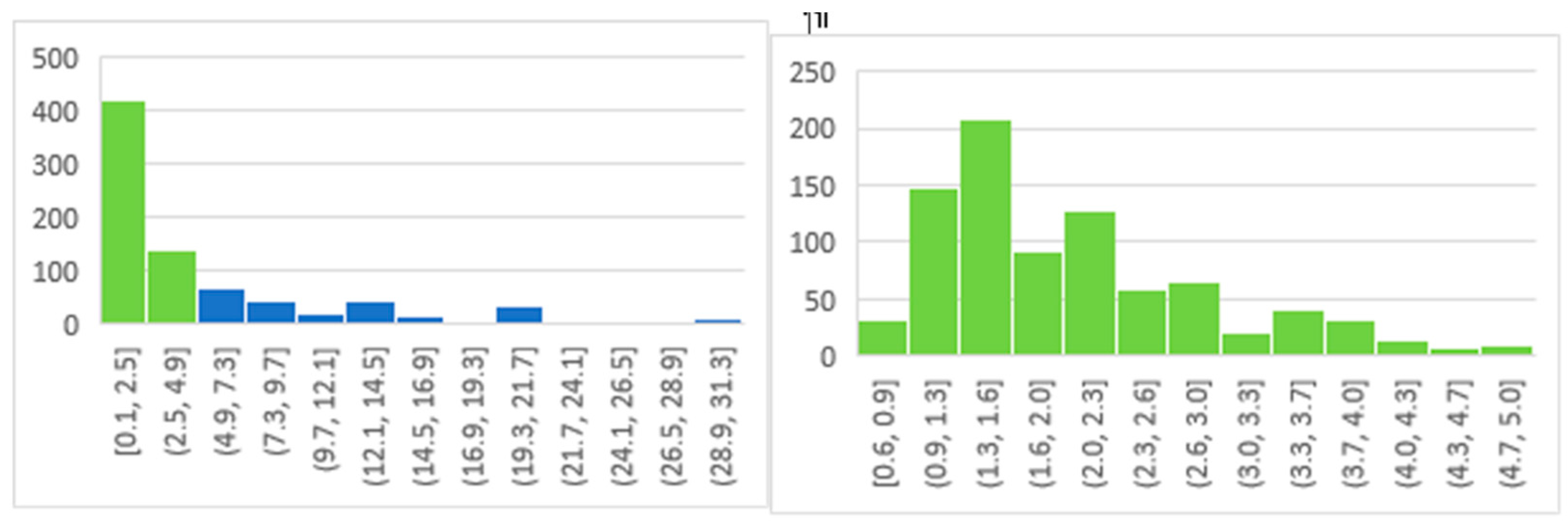
Figure 8.
Rhinella (Bufo) arenarum (Bufonidae) fresh testicular sperm concentrations, the percentage in vitro fertilisation, and minutes post-application before washing with fresh water [292].
Figure 8.
Rhinella (Bufo) arenarum (Bufonidae) fresh testicular sperm concentrations, the percentage in vitro fertilisation, and minutes post-application before washing with fresh water [292].
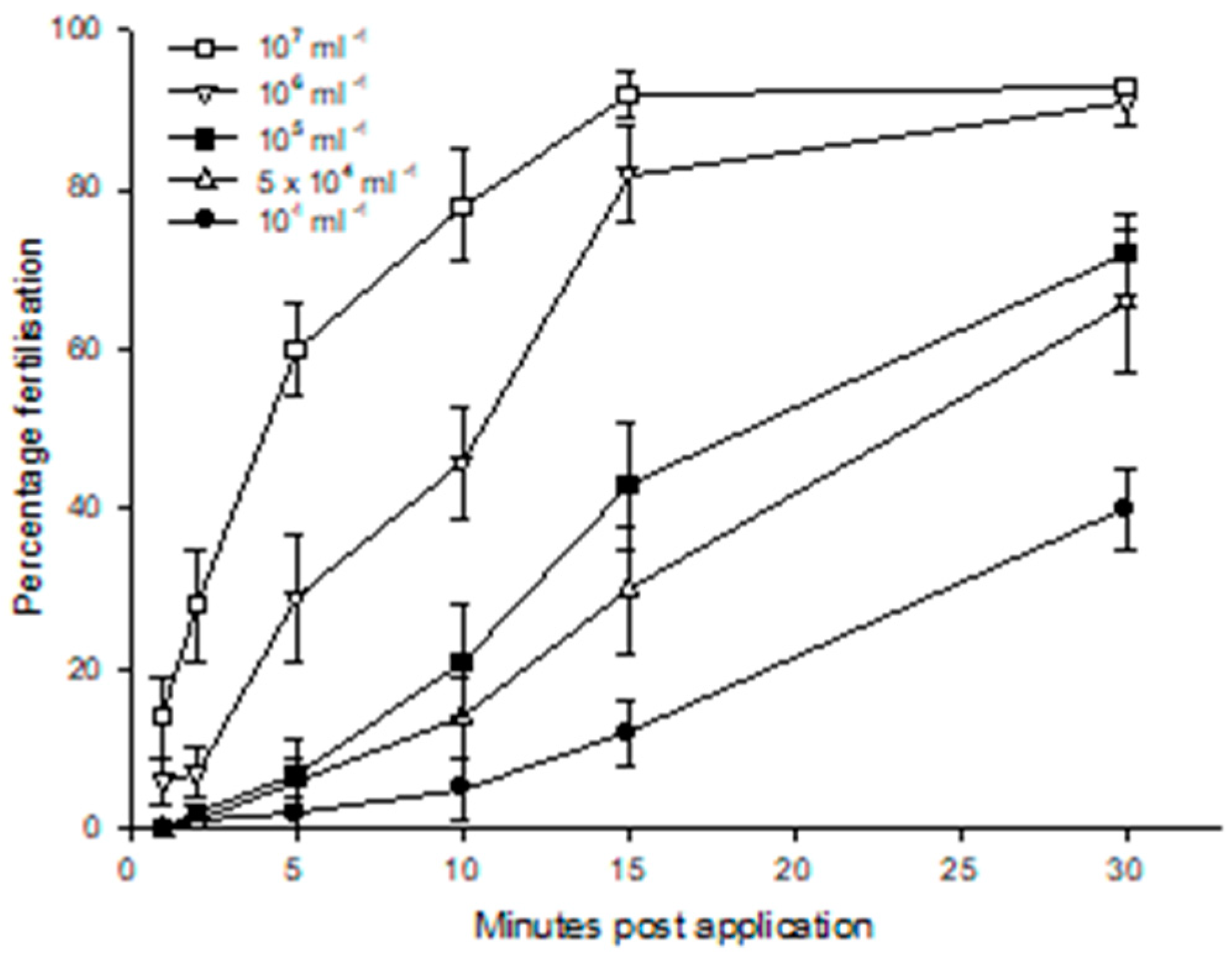

Table 1.
Other approaches to the numbers of biobanked founder individuals for maintaining ex-situ genetic diversity. M = male, F = female.
Table 1.
Other approaches to the numbers of biobanked founder individuals for maintaining ex-situ genetic diversity. M = male, F = female.
| Population | M | F | Application | Year | Reference |
|---|---|---|---|---|---|
| Metapopulation | 5 | 5 | 27 Threatened species. | 2005 | [123] |
| Metapopulation | 28 | 25 | CBP | 2007 | [110] |
| Metapopulation | 25 | - | CBP | 2012 | [124] |
| Fragmented/sub-pop. | 10 | - | AGF 4 years, depending on kinship | 2022 | [125] |
| Geographic clusters | 10 | 20 | Geographic clusters | 2022 | [120] |
| Metapopulation | 20+ | 20+ | CBP | 2024 | [108] |
Table 2.
The number of individuals to be housed in a CBP to maintain 90% of the original genetic diversity from 25 male and 25 female founders over programs of 25 years, based on age to maturity, longevity and whether individual studbook management (Individual) or group management (Group). * = Minimum population sizes of more than 100 are recommended to allow for unexpected loss of individuals [110]. # = Only group management because of short longevity.
Table 2.
The number of individuals to be housed in a CBP to maintain 90% of the original genetic diversity from 25 male and 25 female founders over programs of 25 years, based on age to maturity, longevity and whether individual studbook management (Individual) or group management (Group). * = Minimum population sizes of more than 100 are recommended to allow for unexpected loss of individuals [110]. # = Only group management because of short longevity.
| Examples of Genera | Age to maturity | Longevity | Individual | Group |
|---|---|---|---|---|
| Acris | < 1 year | 1 year | 1590 | # |
| Eleutherodactylus, Nectophrynoides, some Hyperoliidae | < 1 year | 2-5 year | 400 | 400 |
| Hylidae, some Hyperoliidae, Scaphiophryne | 1-5 year | < 5 years | 135 | 265 |
|
Dendrobatidae, Typhlonectes, Tylototriton/Echinotriton, Theloderma, Cynops, Leptodactylus, Ceratobatrachus, Mantella, Atelopus |
1-5 year | 5-15 years | 70* | 140 |
| Salamandra, some Ambystoma | 1-5 year | > 15 years | 60* | 80* |
| Cryptobranchus, Andrias | > 5 years | > 15 years | 45* | 80* |
Table 3.
The average clutch numbers of each IUCN Redlist status by range.
| Range | NoT | n | VU | n | EN | n | CE | n |
|---|---|---|---|---|---|---|---|---|
| 0-10 | 7 | 93 | 4 | 8 | 5 | 8 | 5 | 8 |
| 10-100 | 39 | 361 | 33 | 14 | 27 | 31 | 27 | 14 |
| 100-1000 | 410 | 571 | 483 | 8 | 394 | 4 | 272 | 14 |
| 1000 - 10,000 | 3013 | 328 | 5162 | 5 | 2466 | 5 | 4278 | 2 |
| 10,000 - 100,000 | 17980 | 28 | NA | 0 | NA | 0 | NA | 0 |
Table 4.
Heterocytoplasmic SCNT cloning in amphibians.
| Nucleus donors | Recipients | Results | Year | Ref |
|---|---|---|---|---|
| Rana pipiens | Aquarana (Rana) catesbeiana | Died late blastula | 1952 | [301] |
| R. n. brevipoda | R.n. nigromaculata | Metamorphs | 1957 | [302] |
| R. pipiens | R. sylvatica | Late blastula/early neurula | 1958 | [303] |
| R. n. nigromaculata | R. n. brevipoda | Adults | 1961 | [304] |
| R. pipiens | R. palustris | Post-neurula | 1963 | [305] |
| R. n nigromaculata | R. n. brevipoda | Adults –F1 reproduction | 1963 | [87,88] |
| R. japonica | R. ornativentris | F2 | 1963 | [306] |
|
R. nigromaculata R. temporaria |
R. brevipoda R. japonica |
F3. | 1972 | [307] |
|
R. japonica R. temporaria |
R. temporaria R. japonica |
Adults | 1972 | [308] |
|
R. brevipoda R. plancyi R. brevipoda R. esculenta |
R. plancyi R. brevipoda R. esculenta R. brevipoda |
Adults | 1972 | [309] |
|
Pleurodeles waltlii P. poireti |
P. poireti P. waltlii |
Adults | 1971 | [310] |
|
P. waltlii P. poireti |
P. poireti P. waltlii |
Adults | 1972 | [311] |
Table 5.
Ethical considerations with techniques for the natural or induced mating and spawning, and the collection of sperm or oocytes. .
Table 5.
Ethical considerations with techniques for the natural or induced mating and spawning, and the collection of sperm or oocytes. .
| Technique | Injection | Application stress | Collection technique | Collection stress | *Yield | Donor Size limitation | References |
|---|---|---|---|---|---|---|---|
| Mating and spawning | |||||||
| Natural mating and spawning | None | None | Fertilised eggs | None | Very high | None | [214,215] |
| Hormonally induced mating and spawning | Yes | Moderate | Fertilised eggs | None | Very high | None | [64,214] |
| Oocyte collection | |||||||
| Hormonal stimulation oocytes with spawning into solutions | Yes | Moderate | Spawning into physiological saline | Low | High to very high | None | [233,247] |
| Hormonally stimulated ovarian oocytes | Yes | Moderate | Abdominal massage or cannulation | High | Low | Avoid for small species | [64,233,237,245,246,247] |
| Excision of hormonally stimulated ovarian oocytes | Yes | Moderate | Direct sampling | None | Very high | None | [233,247]. |
| Sperm collection | |||||||
| Collection from testicular tissue | Yes | Moderate | Macerating testicular tissue | None | Very High | None | [61,64,66,76,85,86,222,223,224] |
| Hormonal stimulation sperm | Yes | Moderate | Urination | Moderate | High to none | Avoid for small species | [233,247] |
| Hormonal stimulation sperm | Yes | Moderate | Cannulation | High | Moderate - very low | Avoid for small species | [52,65] |
| Hormonal stimulation sperm - topical, nasal, oral, or food item | None | Low | Cannulation | High | Moderate - very low | Possibly for small species | [61,241,242] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



