
Submitted:
17 September 2024
Posted:
18 September 2024
You are already at the latest version
Abstract
In tropical and subtropical climate regions, heat stress is one of the main causes of production losses in laying quails, aggravated by the antinutritional effects of the phytate in diet ingredients, which negatively affect the bioavailability of minerals, especially calcium and phosphorus. This situation results in a reduction in production and quality of eggs from commercial laying hens. Several nutritional strategies are utilized to reduce the adverse effects of high temperatures and antinutritional factors such as phytate. Among these strategies, the use of exogenous enzymes, such as phytase, stands out as a viable alternative. Phytase breaks down phytate molecules, optimizing the absorption of essential minerals and improving productive performance and egg quality under unfavorable conditions. In this context, this bibliographic review sought to produce a document demonstrating the beneficial effects of the phytase enzyme on the hydrolysis of the phytate molecule and the availability of calcium for Japanese quails (Coturnix coturnix japonica), and its implications in thermal stress due to heat.
Keywords:
Antinutritional factors
; Calcium absorption
; Calbindin-D28K
; Exogenous enzymes
; TRPV6
1. Introduction
Japanese quail (Coturnix coturnix japonica) farming has aroused the interest of breeders due to rapid growth, early sexual maturity, high productivity, low feed intake, and long production period [1,2]. This increased interest also motivates researchers to conduct studies aimed at improving and strengthening quail farming as a highly profitable commercial activity [3].
Exposure of birds to heat stress (HS) environments in tropical and subtropical areas negatively affects their productive performance and causes substantial economic losses [4,5,6]. It is well reported that poor performance of heat stressed birds is mainly due to reduced feed intake in order to reduce the production of metabolic heat [6,7]. Furthermore, a state of imbalance occurs in the physiology and immunology of birds in response to exposure to HS [8]. In laying quails, exposure to heat of 34°C decreased egg production and feed conversion, besides impairing egg quality [9,10]. Furthermore, yolk and serum cholesterol levels and general stress biomarkers increased in laying quails in heat stress environments [11].
Therefore, many attempts have been made to minimize the negative effects of HS on the growth and physiological aspects of birds [12,13], including quails [5,6,9,11,14,15,16]. The use of the phytase enzyme stands out as an alternative, since its administration allows effective degradation of the phytate molecule present in diets, resulting in the subsequent release of minerals and other nutrients [17] to be used by animals subjected to heat stress.
Phytase (myo-inositol hexaphosphate phosphohydrolase - IP6) belongs to a class of exogenous enzymes whose function is to degrade the phytate molecule (inositol hexa-phosphate, IP6) present in diet ingredients, releasing phosphorus, calcium, and other nutrients to be used by the animals [18,19].
Studies have demonstrated the beneficial effects of using phytase on animal performance [17,20,21], on the quality of the eggs, especially the shells [17,22], and on bone characteristics [23]. It is worth noting that these studies were carried out with broiler chickens and laying hens. In this context, this bibliographic review sought to produce a document demonstrating the beneficial effects of the phytase enzyme on the breakdown of the phytate molecule and the availability of calcium for Japanese quails (Coturnix coturnix japonica), and the implications on heat stress.
2. Japanese Quail (Coturnix coturnix japonica)
Quails are birds originated in North Africa, Asia, and Europe. They are members of the Gallinacea order and the Phasianidae family, which also includes chicken and partridge; Pernicidae subfamily, Coturnix Genus [24,25]. They are animals that have been bred since ancient times as songbirds, decorative birds, and fighting birds [26]. The first records of domestication date back to the end of the 19th century and beginning of the 20th century [27], when the Japanese began to crossbreed European quails with wild species, resulting in the creation of the domesticated bird known as Coturnix coturnix japonica, thus marking the beginning of their exploitation [28,29].
In Brazil, the commercial exploitation of quails began around 1989, when a large poultry company decided to set up the first farm in South Brazil, and then began exporting frozen quail carcasses [30]. Since then, the activity has become very important in the Brazilian agricultural economy.
There are several attributes that have contributed to the increase in quail production in the country, such as: rapid growth of the birds, precocity in production and sexual maturity (35 to 42 days), high productivity (average of 300 eggs/year), longevity in high production (14 to 18 months), low investment, as well as rapid financial return [24,31] and the need for a small area for production (200-250 cm² bedding system and 150-200 cm² cage system) [32,33].
In recent years, quail egg production in Brazil has experienced significant growth, driven by increased consumer interest in this product. Eggs are recognized as a complete and balanced food source, rich in protein, and low in fat. Furthermore, their lipid portion contains high concentrations of unsaturated fatty acids [34], giving them a high biological value and making them affordable for consumers [35,36].
For a long time in Brazil, diets for quails were formulated based on the standards of the National Research Council – NRC [37] and of the Institut National de La Recherche Agronomique – INRA [38], which are based on the nutritional levels established for laying hens and/or broilers. However, this approach often resulted in diets with excess or deficiency of essential nutrients for quails. A rare exception to this practice is the Nutritional Recommendation Tables for Japanese and European Quails, developed by Silva and Costa [39] based on experiments carried out in Brazil.
The 4th and 5th editions of the Brazilian tables of nutritional requirements for poultry and swine, prepared by Rostagno et al. [40,41], introduced specific requirements for Japanese quails. However, the increasing research on the nutrition of laying quails is of utmost importance. This is due to recent increases in production costs, especially of ingredients used in poultry feed, making it vital to adopt nutritional strategies that maximize nutrient utilization, resulting in greater production efficiency.
3. Influence of Heat Stress on Japanese Quail Production
In thermoneutral conditions, birds require less energy to maintain a stable body temperature, which allows them to direct most of the energy from food to productive and reproductive processes. Considering the substantial influence of environmental conditions, such as temperature and relative humidity on the production and welfare of birds, it is crucial to monitor and manage these conditions appropriately in order to optimize production [42].
The poultry industry often faces adverse environmental conditions, with exposure to heat stress being one of the most common. Heat stress (HS) occurs when an animal’s heat production exceeds its ability to dissipate it into the environment [43]. In circumstances where the ambient temperature exceeds the thermoneutral range, birds tend to reduce their physical activity and food intake to control heat production. In addition, they increase panting and water consumption to facilitate heat dissipation through evaporation [44].
Panting is a phenomenon exhibited by birds when they open their beaks to increase the rate of respiration and evaporative cooling of the respiratory tract. During panting, CO2 excretion occurs faster than cellular CO2, production, which alters the standard bicarbonate buffer system in the blood. The reduction in CO2 leads to a decrease in the concentration of carbonic acids (H2CO3) and hydrogen ions (H+). In contrast, the concentration of bicarbonate ions (HCO3−) increases, thus raising the pH of the blood, i.e. the blood becomes alkaline. To manage this scenario and preserve blood pH balance, birds initiate a process of increased excretion of bicarbonate ions (HCO3−) and retention of hydrogen ions (H+) by the kidneys. Elevated H+ alters the acid-base balance, leading to respiratory alkalosis and metabolic acidosis, and is associated with a decline in productive performance of birds [45]. However, it is important to note that although the kidneys play a crucial role in compensating for alkalosis, prolonged exposure to high temperatures can result in the onset of a condition known as chronic alkalosis [46,47].
Furlan [48] conducted a study with five commercial strains of broiler chickens. Subjecting them to heat stress, he observed that, regardless of the strain, there was a decrease in CO2 content and an increase in blood pH as the temperature increased [47].
Ruzal et al. [49] state that respiratory alkalosis caused by panting has a deleterious effect on laying hens due to its effect on egg quality. Respiratory alkalosis evidently causes an increase in arterial blood pH associated with a reduction in the partial pressure of CO2. This leads to a decline in plasma bicarbonate level and an increase in the binding between organic acid and calcium ion content, thus causing declines in the availability of bicarbonate and calcium ions and weakening egg quality.
Laying quails may also suffer impacts on their acid-base balance due to periods of intense heat, resulting in electrolyte and mineral imbalances, potentially culminating in smaller eggs with thinner shells. This phenomenon is mainly attributed to alkalosis, which reduces the availability of free calcium in the blood and increases the proportion of calcium bound to proteins or complexed with organic acids. Consequently, during heat stress, an increase in blood pH is observed due to the loss of carbon dioxide, simultaneously accompanied by a reduction in available calcium [47].
Vercese et al. [10] evaluated the performance and quality of the eggs of Japanese quails subjected to cyclical heat stress and observed that Japanese quails exposed to a temperature of 27°C already showed signs of heat stress, such as reduced feed intake, egg weight, and egg mass. The cyclical increase in ambient temperature to 36°C negatively influenced the percentage of marketable eggs and egg production.
When evaluating the effect of heat stress on laying quails (Coturnix coturnix japonica), Akdemir et al. [16], stated that birds subjected to temperatures of 34°C have lower feed intake, lower egg production and weight, in addition to greater feed conversion.
In their research with laying quails, Moraes et al. [3] investigated the effects of heat stress compared to thermoneutral conditions. They highlighted that exposure to high temperatures can significantly impair bird performance, reflected in reduced feed intake, live weight gain, and efficiency. In addition, they observed a decrease in egg production, as well as in eggshell quality and thickness, due to the lower availability of calcium ions [50].
Lesson and Summers [51] state that, under conditions of respiratory alkalosis, there is a reduction in the calcification of eggs and bones, compromising the performance of birds and contributing to the increased incidence of leg problems in animals, as well as to the production of eggs with thin shells. Alkalosis limits the availability of anions necessary for the formation of calcium carbonate crystals in the eggshell [52] and, consequently leads to poor egg quality. According to Campos [53], this can result in a 12% reduction in eggshell thickness [10].
Another point is that in situations of stress due to high temperatures (30-33°C for quails) there is a decrease in the presence of the calcium transporter calbindin-D28k in the ileum, cecum, colon, and uterus of the birds [3], which results in deterioration of eggshell quality under these conditions [54]. his occurrence can be explained by heat stress, which reduces the conversion of vitamin D3 into its metabolically active form, 1,25 (OH)2D3, essential for the absorption and utilization of calcium [3]. Furthermore, stress also decreases antioxidant patterns, leading to deficiency of or increased requirements for vitamins and minerals, especially zinc [55], considered a cofactor ion of the enzyme carbonic anhydrase responsible for calcium deposition in the eggshell [19,56].
4. Phytase Enzyme and the Hydrolysis of the Phytate Molecule
Phytic acid (myoinositol 1, 2, 3, 4, 5, 6-hexakis dihydrogen phospodium) also known as Inositol-6-phosphate (Figure 1), phytate in its salt form [57], phytin, or phytic phosphorus is the main phosphorus storage system in plant seeds, such as cereal grains, nuts, oilseeds, and legumes [57,58]. Table 1 highlights the phytate levels in some foods used in animal feed.
Approximately 60–80% of P in most cereal grains and oilseeds is bound to phytate [69], which is a chemical form of low availability for poultry and swine [70]. Phytate traps phosphorus and other cationic substances, preventing them from being fully utilized by birds [71]. According to Woyengo and Nyachoti. [72] phytic acid contains 12 ionizable protons with pKa values ranging from 1.5 to about 10 [73,74]. Therefore, at all pH values normally observed throughout the gastrointestinal tract, phytate is capable of binding to cations, causing mineral unavailability.
In addition to making phosphorus unavailable, phytate also negatively influences the digestion and absorption of other minerals, amino acids, and the energy utilization of diets [75]. Phytate can create a wide variety of insoluble salts with divalent cations, such as calcium (Ca2+), magnesium (Mg2+), iron (Fe2+), zinc (Zn2+), copper (Cu2+) and manganese (Mn2+) [76], minerals which are considered nutritionally important, especially for laying hens. Thus, blood concentrations of phosphorus and calcium can be reduced, negatively affecting production processes [77]. Furthermore, phytic acid can also bind with proteins and inhibit the activity of some digestive enzymes, including trypsin, pepsin, and alpha-amylase [78].
Phytase or myo-inositol hexaphosphate phosphohydrolase (IP6) is a phosphatase commonly used in the diets of non-ruminant animals as a way to increase the bioavailability of phosphorus phytate and other nutrients [18]. This enzyme hydrolyzes phytic acid and its salts (phytate) producing inositol, inositol monophosphate, and inorganic phosphorus [79], releasing phosphorus, and improving the availability of other nutrients bound to the phytate molecule [80]. The activity of this enzyme is expressed in a Phytase Activity Unit (FTU), which means the amount of enzyme required to release 1 μmol of inorganic phosphorus per minute of reaction from sodium phytate, at a temperature of 37°C and pH of 5.5 [81].
Phytase catalyzes the release of phosphate present in phytate, and then produces myo-inositol pentakis-, tetrakis-, tris-, bis-, and monophosphates, as well as inorganic phosphate [82,83], (Figure 2). Enzymes can be classified into different classes according to some criteria, including the stereospecificity of phytate hydrolysis (carbon number in the myo-inositol ring of phytate at which dephosphorylation is initiated), pH optimum (alkaline or acid phytases) and their catalytic mechanism [84].
Based on stereospecificity, according to the International Union of Biochemistry (UIB), three groups of phytases can currently be distinguished, depending on the position in the Inositol ring where dephosphorylation is initiated, namely: 3-phytases (EC 3.1.3.8), 6-phytases (EC 3.1.3.26) and 5-phytases (EC 3.1.3.72) [85,86,87], as shown in Figure 3.
Description of the three phytases:
a) 3-phytase (EC 3.1.3.8): Myo-inositol hexakisphosphate-3-phosphohydrolase, initiates the hydrolysis of the ester bond at the third position of myo-inositolhexakisphosphate into myo-inositol-1,2,4,5,6-pentakisphosphate and orthophosphate, isolated from Aspergillus niger, Neurospora crassa, Pseudomonas and Klebsiella sp. ASR1.
b) 4-phytase, also known as 6-phytase (EC 3.1.3.26): myo-inositol hexakisphosphate-6-phosphohydrolase, hydrolyzes the ester bond at the sixth position of myo-inositolhexakisphosphate to myo-inositol-1,2,3,4,5-pentakisphosphate and orthophosphate, isolated from plants, E. coli, Paramecium.
c) 5-phytase (EC 3.1.3.72): Myo-inositol-hexakisphosphate-5-phosphohydrolase, initiates the hydrolysis of the ester bond at the fifth position of myo-inositolhexakisphosphate to myo-inositol-1,2,3,4,6-pentakisphosphate and orthophosphate, isolated from Medicago sativa, Phaseolus vulgaris and Pisum sativum.
Depending on their optimal pH, phytases can be further classified as acidic or alkaline, acidic when their pH is between 2.5 and 6.0, and alkaline when their pH is close to 8.0 [88,89,90]. Based on their structural differences and their catalytic mechanism, phytases can be classified into four groups, which are histidine acid phosphatases (HAPs), ß-helical phytases (BPPs), cysteine phytases (CPs), or purple acid phosphatases (PAP) [84,91] (Figure 3).
5. Phytase Overdose
Phytase overdose refers to the addition of phytase to diets at levels higher than those recommended (500 FTUs/kg) [92], and may even be double or triple these recommendations. Such dosages have been used to effectively combat the antinutritional effects of phytate, improving animal performance [20,80].
According to Cowieson et al. [93], there are three main mechanisms for which the use of high doses of phytase can have beneficial effects:
1. Greater quantity of phosphate made available by the enzyme or greater proportion in the release of calcium and phosphorus;
2. Less phytate excreted, i.e. destruction of the antinutritional effect and increased generation of more soluble esters;
3. Generation of myo-inositol with vitamin/lipotropic effects
There are several studies demonstrating the relevant effects of the use of phytase overdoses in broiler chicken diets:
Nelson et al. [94] used 1-8 g of a phytase from Aspergillus ficum with 950 FTUs/kg in the diet of broilers (950 and 7,600 FTUs/kg of feed). These authors evaluated that apparently 38.9% of the phytate disappeared when using the concentration of 950 FTUs/kg of the enzyme and 94.4% when using 7,600 FTUs/kg, with greater responses for weight gain and bone ash in 21-day-old chickens when using 7,600 FTUs/kg of phytase in the diets [80,93].
When evaluating the extra-phosphoric effects of overdoses of a microbial phytase produced from E. coli, Walk et al. [95] observed better feed conversion in broilers fed with 1,500 FTUs/kg of phytase in the diet with a reduction of Ca by 0.16%, and P by 0.15%. According to these authors, these findings could be related to the reduction of the antinutritional factors of phytate when using the enzyme in the diet.
When evaluating the effects of phytase overdose (SUNPHASE 5000 G) on the digestibility and bone development of broilers from 1 to 21 days of age, Fernandes et al. [96] observed a quadratic effect of phytase inclusion for the diameter and percentage of Ca and P of the femur, in which the inclusion of 1,494 and 1,220 FTU, respectively, presented the best results. For the evaluation of the tibia, the diameter, length, and weight increased linearly with the inclusion of phytase, and for resistance a quadratic effect was observed, with 1265 FTU being the best level.
In a study with four commercially available phytase sources supplemented at regular levels (250 and 500 FTUs/kg) and overdoses (100; 1500 and 2000 FTUs/kg) on live performance, bone mineralization, and apparent ileal digestible energy in broilers from 7 to 24 days of age, Leyva-Jimenez et al. [97] stated that on days 14 and 22 all phytase sources improved (P < 0.05) body weight, weight gain, and bone mineralization when compared to birds on the negative control diet. Overall, phytase supplementation at the S level improved apparent ileal digestibility by 17% at 24 d. Throughout the growing period, the phytase overdose yielded (P < 0.05) better performance, bone characteristics, and energy digestibility than at the regular dietary level. In conclusion, all phytase sources were able to compensate for phosphorus deficiency and promote performance and bone mineralization. Higher phytase levels showed a greater response when compared to lower supplementation levels.
Given the satisfactory results with broilers, there is growing interest in the results with laying hens as well, taking into account that the use of Ca and P in laying hens is probably more important than in broilers [98] due to the greater requirement of these minerals in the formation of eggshells [80].
When evaluating the effects of overdose (450 FTUs or 900 FTUs) of two phytases (bacterial phytase or fungal phytase) on the performance, egg quality, biometry of the digestive organs, and bone quality of light laying hens in the first (58 weeks) and second (87 weeks) production cycles, Farias et al. [17], concluded that bacterial phytase produced from Escherichia coli, at a dosage of 450 FTUs, improved the egg production of light laying hens. According to these authors, the use of phytase in the diet of laying hens implies lower feed costs.
When evaluating the effects of an overdose (0; 500; 1000; 1500 and 3000FTU) of phytase on bone parameters and the concentration of the epithelial calcium carrier Calbindin-D28k in Japanese quails, Ribeiro et al. [99] had results that showed that phytase overdose not only increased the efficiency of calcium absorption, but also stimulated a greater expression of calbindin-D28K in the duodenum and jejunum of the birds. This resulted in improved calcium mobilization to the tibiae and increased egg production. Additionally, supplementation with 1500 FTU of phytase reduced the negative effects of heat stress at 36°C by increasing eggshell thickness. These studies highlight the importance of phytase overdosing as an effective strategy to improve bird health and productivity under heat stress conditions.
6. Role of Phytase in Reducing Heat Stress
As already well documented, heat stress influences the performance of laying hens, as well as in any production system, constituting one of the main causes of production losses in tropical and subtropical regions [100]. Factors such as time of day, genotype, breeding system, and animal density interfere in thermoregulatory responses, leading to changes in the natural behavior of birds [19].
Nutritional strategies are widely used to reduce the negative effects of heat stress, such as adjusting protein and/or energy levels in diets, including fats or oils, supplementing with synthetic amino acids, using vitamins, and offering drinking water with mineral additives [101]. The use of exogenous enzymes such as phytase in the diet of birds stands out as one of these nutritional strategies implemented to overcome the effects caused by heat stress, resulting in better animal welfare [19]. Since birds do not produce enough phytase enzymes to hydrolyze the phytate molecule present in diet ingredients, the introduction of this enzyme in their diets is necessary, as phytase catalyzes the hydrolysis of phytic acid, eliminating its antinutritional properties [78], thus promoting the release of phytic phosphorus present in plant foods, in addition to making other minerals available, such as calcium and vitamins, and increasing the digestibility of the nutrients in diets [102].
Phytase also has extra phosphoric effects, such as the release of proteins, amino acids, carbohydrates, minerals, and vitamins that are complexed to the phytate molecule [103], besides improving the utilization of the dietary energy by the animals [97]. The phytase enzyme has also been recognized for facilitating the availability of zinc [104]. According to Borges [105], This mineral is incorporated into some dietary formulations to reduce the effects of heat stress in birds, promoting better nutrient absorption during periods of stress, which can reduce losses, especially energy losses.
According to Hu et al. [106], at elevated temperatures, enzyme activity is greatly affected in birds. There is a significant change in the activity of metabolic enzymes in the body, thereby increasing the metabolic rate and therefore increasing the production of free radicals. Excess free radicals disrupt the oxidative and antioxidant balance of the body, leading to lipid peroxidation, DNA and protein damage, and the generation of oxidative stress.
Oxidative damage to the cell membrane by free radicals occurs during zinc deficiency, thus altering the status of enzymes and antioxidant substances [107]. Zinc (Zn) is a vital mineral element found in birds, predominantly in the bones and liver. It plays an essential role in growth, and in the proper functioning of the immune system of birds. In addition, zinc contributes to the development of birds, improves feed conversion efficiency, strengthens immunity, and helps prevent diseases [106].
Zinc is required for the activity of over 300 enzymes and participates in many enzymatic and metabolic functions in the body [108]. One of the most important functions of Zn is its participation in the antioxidant defense system. Zinc deficiency increases oxidative damage to cell membranes caused by free radicals [55,108]. Zn exerts its antioxidant action by increasing the synthesis of metallothionein, a protein rich in cysteine, which acts as a free radical scavenger [109]. Another proposed mode of action for Zn as an antioxidant is its interaction with vitamin E, because vitamin E status is impaired in zinc-deficient animals [55]. Furthermore, zinc can occupy iron and copper binding sites in lipids, proteins, and DNA, and thus exert a direct antioxidant action [107]. Furthermore, zinc increases antioxidant capacity in birds by increasing the activity of copper-zinc superoxide dismutase and zinc metalloenzymes and altering DNA and chromatin structure to influence gene expression [106].
Another point related to the importance of the phytase enzyme in the availability of Zn in the diets of birds subjected to heat stress is that this mineral is a cofactor of the enzyme carbonic anhydrase, which plays a vital role in the formation of the eggshell. In addition, Zn can be incorporated in the growth stage during the formation of the calcite crystal [110]. Carbonic anhydrase is an enzyme that catalyzes the hydration of metabolic CO2 to HCO3 −, the precursor of eggshell carbonate. Several studies have reported that partial or complete inhibition of carbonic anhydrase can result in thin-shelled or shell-free eggs [110,111]. It has also been reported that eggshell quality is reduced by inhibition of carbonic anhydrase activity or anhydrase mRNA expression in laying hens [110,112,113].
Studies have demonstrated the positive effects of using phytase in the feed of poultry under heat stress: Mohebbifar et al. [114] investigated the effects of phytase supplementation (0 and 150 FTU) in diets with different levels of rice bran and non-phytic phosphorus on productive performance, egg quality, leukocyte profile, and serum lipids in laying hens raised indoors under high environmental temperatures. They found that diets with high phytate content (such as those rich in rice bran) and/or low non-phytic phosphorus (2.5 g/kg), when supplemented with phytase, resulted in significant improvements in the performance and egg quality of birds subjected to heat stress.
When evaluating the simple and combined effects of phytase and citric acid on performance, nutrient digestibility, bone characteristics, intestinal morphology and blood parameters of Japanese quails (aged 1 to 35 days) and fed with low-phosphorus diets, Hezaveh et al. [115], found that phytase supplementation resulted in significant improvements. There was an increase in bird performance, protein, ash and phosphorus digestibility coefficients, metabolizable energy, bone mineralization, and intestinal morphology, even with a 0.12% reduction in non-phytic phosphorus in the diet.
When evaluating phytase levels (0; 200; 400 and 600 FTU/kg) in the diet on the performance and quality of eggs of Japanese quails subjected to heat stress, Lima et al. [116] concluded that the use of phytase improved the performance and quality of the eggs of the birds. The best phytase level for better efficiency in the use of phosphorus by the birds and for better egg mass was 463 FTU/kg.
When evaluating the effects of an overdose of phytase on bone parameters and the concentration of the epithelial calcium carrier Calbindin-D28k in Japanese quails under heat stress, Ribeiro et al. [99] demonstrated that phytase supplementation brought benefits to the birds, especially those kept at temperatures of 30°C, given the greater demand for vitamins and minerals, due to changes in their metabolism. Phytase improved Ca absorption efficiency and positively influenced the increased expression of anti-calbindin-D28K in the duodenum and jejunum of the birds. This provided greater Ca mobilization to the tibiae and higher total egg production for the birds kept at this temperature. Furthermore, these results demonstrate that dietary phytase supplementation at a level of 1500 FTU was able to reduce the deleterious effects of heat stress (36°C), and positively influenced the increase of quail eggshell thickness.
In this context, it can be seen that the use of phytase can positively influence the reduction of the effects of heat stress and antinutritional factors such as phytate in Japanese quails.
7. Calcium and Phosphorus in the Diet of Laying Quails (Coturnix coturnix japônica)
According to current practice, diets for commercial laying quails are formulated based on the recommendations of tables of nutritional requirements specific to each lineage, such as editions 4 and 5 of the Brazilian Tables of Nutritional Requirements for Poultry and Swine, prepared by Rostagno et al. [40,41]. Reducing the density of nutrients such as phosphorus in the feed and using phytase supplementation can reduce the total cost of the diet, thus contributing to the profitability of egg production [117].
Calcium (Ca) and phosphorus (P) are essential for laying quails, and their availability is most crucial during the laying period [118]. Among all of the minerals, calcium and phosphorus play the most fundamental role in the construction of the skeleton, from 80 to 85% of its structure. These minerals are essential in the formation of eggshells and in muscle development. Thus, they are indispensable for the proper functioning of the animal’s body [119].
Calcium is the most abundant mineral in the animal organism, representing a body proportion of 1:75 [120]. Most calcium (99%) is present in bones [121,122] in the form of hydroxyapatite, an inorganic crystalline structure composed of calcium and phosphorus [Ca10(PO4)6(OH)2] [120]. During the processes of absorption, metabolism, and excretion, calcium and phosphorus interact, maintaining a ratio of approximately 2:1 [123]. In addition to composing the bone structure, calcium is also present in soft tissues (1%), where it performs several functions, such as activating enzymes, transmitting nerve stimuli, and participating in the blood coagulation process [120].
The use of calcium by the bird’s body varies mainly with age. During the growth period of laying hens, most of the dietary calcium is directed towards the formation of bones [120]. In contrast, in the adult phase, calcium is predominantly used for egg production, specifically for shell formation [119]. The Ca²⁺ necessary for the formation of the shell, which comes from bone resorption and intestinal absorption, is transported by the blood to the calcigenous chamber [120,124]. This transport is mediated by the activity of carbonic anhydrase, a crucial enzyme in eggshell formation [110].
Approximately 30% of the calcium required for eggshell formation is mobilized from the bones. This is because in the uterus, where calcium carbonate is deposited, there is no calcium storage. The organic fraction of the shell is synthesized by glands, and calcium is mobilized directly from the blood, and the transfer of plasma to the uterus occurs very quickly [120,125].
The most obvious symptoms of calcium deficiency in young birds include delayed growth, reduced feed intake, and bone fragility [120]. In adult birds, the deficiency manifests itself in reduced egg production, thin-shelled eggs, and reduced ash and calcium content in bones [126,127].
Phosphorus, in turn, plays a crucial role in both the absorption and metabolism of calcium [128]. More than 80% of phosphorus is associated with calcium in bone formation, while the remainder is present in soft tissues [129,130]. Its functions include participation in bone structure (hydroxyapatite), in cell membranes in the form of phospholipids (lecithin), as phosphate in deoxyribonucleic acid (DNA) and ribonucleic acid (RNA) molecules, and in the form of energy molecules, such as adenosine diphosphate (ADP) and triphosphate (ATP) [128,129].
The skeleton functions as a reservoir of calcium and phosphorus in the body, and this function is especially crucial in laying hens, since the eggshell contains approximately 10% of the total calcium in the bird’s body [120]. Therefore, there is a high demand for calcium to maintain blood homeostasis while the shell is forming [131]. Physiological mechanisms regulate skeletal integrity in response to different needs during the productive life of laying hens. The relative importance of the intestine and bones as sources of calcium depends on the concentration of this mineral in the diet [120,132].
In general, there are intestinal barriers, such as physical-chemical conditions, pH, and viscosity, which make it difficult to absorb most minerals [133]. Therefore, dietary levels of calcium and phosphorus are often higher than actual requirements, resulting in a lower utilization rate and environmental impact due to excess excretion [134].
To ensure eggshell quality, calcium and phosphorus levels in the diet must meet the nutritional requirements of the birds [135]. It is important to note that these requirements may vary depending on the type of quail (Japanese or European), sex, age, and purpose of the farm, whether for the production of eggs for consumption, incubation, or for meat [136].
Several studies are being conducted with to establish precise information on the nutritional needs of Japanese quails, aiming to achieve maximum productive performance and maintain health. Table 2 presents the recommended values of calcium and available phosphorus, proposed by different authors for Japanese quails in the laying phase.
Variations in calcium (Ca) and available phosphorus (P) recommendations for Japanese quails derive from several reasons, such as the source and bioavailability of the minerals, the age of the birds, the efficiency of absorption, and the energy levels of the diets.
8. Calcium (Ca) Absorption
Active absorption and reabsorption of Ca is of great importance in maintaining the homeostasis of this mineral for many physiological functions [148,149]. It is also essential for the formation of skeletal hydroxyapatite and for the mineralization of bones, as well as for satisfying the high Ca requirement in the uterus (where the eggshell is formed) for egg calcification in laying hens [150,151].
In modern commercial strains of laying hens, about 2-3 g of Ca (equivalent to 10% of total body calcium) is transferred daily for deposition in the eggshell [118,152]. The largest source of Ca comes from dietary absorption in the intestine, renal reabsorption, and bone stores [148].
Plasma Ca concentrations are precisely controlled within a narrow range of variation, both intracellular and extracellular, through interactive mechanisms between parathyroid hormone (PTH), calcitonin (CT), 1,25 dihydroxycholecalciferol (1,25(OH)2D3) and estrogen and their respective receptors located in the intestine, bones, and kidneys [120]. In response to low blood Ca2+ levels, parathyroid cells rapidly release PTH, which stimulates bone resorption by releasing calcium from the bone matrix into the circulation and also by promoting reabsorption in the kidneys. These actions help restore serum Ca2+ concentrations. In chronic situations, PTH increases the production of 1,25(OH)2D3 to facilitate intestinal Ca2+ absorption Ca2+ [153].
Among the various mechanisms aimed at reducing circulating Ca, phytate can be highlighted as one of these, as it has chelating power over several divalent cations, mainly Ca, making it less available [77,154].
There are two mechanisms for the mobilization of Ca that is deposited in the bones. The first is represented by the transfer of ions from the hydroxyapatite crystals to the interstitial fluid, from which the calcium passes into the blood. This mechanism is considered purely physical and takes place mainly in the spongy bones. The second mechanism, which is characterized by having a slower action on the bone tissue, uses the parathyroid hormone (PTH) produced in the parathyroid gland [155]. PTH increases the number of osteoclasts and the resorption of bone matrix [156], releasing calcium phosphate and increasing calcemia [157] nd acts on the kidneys, decreasing phosphorus excretion and stimulating the synthesis of active vitamin D [158]. Parathyroid hormone secretion activity is regulated in response to fluctuations in calcium concentration [120,159].
Calcitonin is another hormone that acts on Ca metabolism to maintain normal levels in blood plasma. It is produced by the parafollicular cells of the thyroid and acts by inhibiting the reabsorption of bone matrix and, consequently, the mobilization of bone Ca. Its secretion is stimulated when Ca levels are high in the blood [160,161].
Modern commercial strains of layers can produce about 300 eggs per year, which require an amount of Ca equivalent to more than 20x their total body weight annually [162,163]. Therefore, layers must absorb a large amount of calcium to support the demand for this production. Epithelial calcium absorption involves two distinct pathways: active transcellular transport of calcium and diffusion via the paracellular pathway [148].
In the mammalian intestine, transcellular calcium transport plays an important role in calcium homeostasis, which is a three-step process comprising the passive entry of calcium into the enterocyte via TRPV6; the cytoplasmic transfer of calcium bound to the protein calbindin-D9k; and in birds the analogous protein is calbindin-D28k; and calcium extrusion across the basolateral membrane via Ca2⁺ –ATPase (PMCA 1b) and/or Na – Ca exchanger [164,165]. A similar mechanism exists in tissues such as the kidney, uterus, and placenta [148,166,167,168,169,170,171].
The kidney also plays an essential role in calcium balance by regulating the excretion of calcium from the body. To meet the requirement for eggshell calcification, calcium filtered in the glomerulus is extensively reabsorbed as it passes through the nephrons [168,172].
Egg formation in laying hens is a complex process, which depends on the maturation of the ovum in the ovary, passage of the mature oocyte through the oviduct via the infundibulum, addition of albumen in the magnum and synthesis of the membrane in the isthmus, and deposition of the shell in the uterus. The mineral components of the shell in laying hens are composed mainly of calcium carbonate and, to a lesser extent, of magnesium carbonate and tricalcium phosphate. The initial deposition of minerals occurs in the isthmus where the shell membrane is formed. The calcification of the shell in the uterus takes approximately 20 hours, during which time 2g of Ca is deposited [173].
The function of the different cell types of the uterus and tubular glands is not clearly defined, but they are implicated in the process of Ca secretion. High concentration of calcium-activated ATPase is located mainly in the microvilli of the uterus tubular cell glands [174]. Although the pathway of calcium absorption and reabsorption is physiologically known, there are no studies in quails that aim to elucidate the absorption, reabsorption, and deposition of calcium in the uterus, including its sites and expression, and they are also scarce in poultry farming in general.
9. Transepithelial Calcium Transport Mediated by TRPV6 and Calbindin-D28K
Epithelia play a fundamental role as linings in various biological compartments, where they create specialized surfaces for essential functions such as protection, secretion, absorption, and reabsorption of substances [175]. For example, the ability of epithelial cells to regulate the absorption and secretion of vital ions, such as calcium, plays a crucial role in maintaining electrolyte balance, thus ensuring the proper processes of several vital functions in the body [168,176,177].
The absorption and reabsorption of calcium in the body occurs through the epithelium of various organs, such as the intestines, kidneys, mammary glands and placenta. This process is mediated by a complex sequence of events regulated by different factors, such as pH, extracellular calcium concentrations and hormones [168,178]. There are two main pathways described for calcium transport across epithelia: the paracellular pathway, which allows the movement of ions through the spaces between cells, and the transcellular pathway, in which ions are transported through the cytoplasm of epithelial cells. In the transcellular pathway, calcium transport occurs through a process that involves several steps, including the entry of calcium into the cell through the apical membrane, the translocation of calcium through the cytoplasm to the basolateral membrane, and, finally, the release of calcium into the blood [168], (Figure 4).
Calcium entry into epithelial cells occurs through channels located in the apical membrane, known as TRPV5 and TRPV6 (Transient Receptor Potential Vanilloid), previously called ECaC1 and ECaC2, respectively. These proteins belong to the family of channels known as Transient Receptor Potential Vanilloids, specifically grouped in the Vanilloid subfamily [168], which includes six components, with only TRPV subtypes 5 and 6 having calcium transport capacity [179].
The ion channel TRPV6 (Transient Receptor Potential Vanilloid Channel Type 6) (Figure 5) functions as an epithelial Ca channel in the intestine, kidney, bone, skin, placenta, and exocrine glands of mammals, tissues that are characterized by a high demand for calcium transport [168,180].
TRPV6 is described as having a facilitating effect on Ca entry into epithelial cells, with a significant correlation between TRPV6 expression and Ca transport [150,181], and this ion channel is vitamin D dependent [182]. Observations have led to the suggestion that TRPV6 is an important limiter of calcium entry and homeostasis [150,183]. This ion channel is expressed in the absorption and reabsorption epithelia, that is, in the intestine and kidney, however, there is little information about the expression pattern of TRPV6 in laying hens [148] and none for laying quails.
Immunohistochemical studies indicated anti-TRPV6 positivity in the apical border of the cells of the duodenum, jejunum, ileum, cecum, and rectum. The positivity was highest in the duodenum and weakest in the rectum, and was absent in the crypts and goblet cells. PCR and Western blot also demonstrated the presence of TRPV6 in the aforementioned segments, mainly in the duodenum and jejunum of laying hens [148]. This finding is consistent, since the main sites of calcium absorption in birds are the duodenum and jejunum [184]. In humans, TRPV6 (mRNA) is also most abundant in the duodenum and jejunum, with low levels in the ascending colon, and none in the ileum or distal segments of the large intestine [185].
The significant presence of TRPV6 (mRNA and protein) in the ileum of laying hens [148], contrasts with that found in rats, humans, and sheep [180,185,186], suggesting that the ileum may also have a significant role in intestinal absorption in laying hens. In the kidney of laying hens, TRPV6 (protein) is positive at the apical border of the proximal convoluted tubules, loops of Henle, and distal convoluted tubules, however, weaker than in the digestive system.
Despite the cited study [148], for other authors [187,188,189], the presence of TRPV6 in bird tissues, including laying hens or quails, is still uncertain. Considering that TRPV6 is found in the intestinal segments of laying hens, calcium absorption should play a significant role in egg production during laying peak, including shell production, in the uterus. These observations are in line with Yang et al. [148], who describe areas for future studies covering the mechanisms of calcium transfer in laying hens. These studies would include the expression pattern of TRPV6 between laying and non-laying hens, co-expression with other vitamin-dependent proteins such as calbindin-D28k, and expression in other calcium-transporting tissues such as the avian uterus and bone marrow.
Thus, Yang et al. [148] propose that the epithelial channel TRPV6 has a crucial function in the absorption and reabsorption of calcium in laying hens. However, further studies are needed to elucidate its action, especially in the uterus and kidney, and there are still no studies of such calcium transporters in quails. Thus, the increase in the expression and positivity of TRPV6 may physiologically prove the better use of calcium after environmental or nutritional changes.
The Calbindin-D28K protein (Figure 5) plays a crucial role in calcium transport: once in the cytoplasm, calcium ions bind with high affinity to calbindin D28K, which acts as a carrier facilitating the diffusion of calcium between the apical and basolateral cytoplasm of cells [3].
Passive calcium absorption involves diffusion of calcium from the intestinal lumen into the enterocyte, while active absorption is the process in which calcium passes through channels found in the apical membrane of the microvilli of the enterocytes. These then bind to a protein that binds to calcium, Calbindin (vitamin D-induced calcium-binding protein), and diffuses it into the cytoplasm, which is finally extruded by CA2⁺-ATPase in the basolateral membrane, reaching the vascular system through vessels of the lamina propria [152].
Thus, the potential for active calcium absorption depends on the presence of calbindin in the cytoplasm [190], with its biosynthesis being dependent on circulating levels of vitamin D [191], and on the activity of Ca2⁺-ATPase in the basolateral membrane [192], with the majority of dietary calcium being absorbed by the intestinal segments via the active pathway [193].
Calbindin exists in 2 main forms, with low molecular weight, one 9kDa protein (Calbindin-D9k) present in mammalian intestines, and another with high molecular weight with 28kDa (Calbindin-D28k), present in the kidney, brain, intestine, and uterus of birds, and in the kidneys of mammals [194].
The localization of calbindin-D28k in birds was initially described in the intestine of chickens [187] and later in the uterus of laying hens [195] and in the distal portion of the isthmus [196], the latter locations being sites of calcium deposition in the avian oviduct. The synthesis of calbindin-D28k is dependent on vitamin D [195]. However, during the eggshell production cycle, the concentration of calbindin-D28k in the uterus is relatively insensitive to changes in plasma vitamin D levels [197,198].
In the intestine of laying hens, there is positivity (protein) of calbindin-D28k in all segments. It is higher in the duodenum and jejunum, mainly in the apical portions of the villi [152,191], and lower in the ileum. Positivity also occurs in the cecum and colon, to a lesser extent, which indicates that laying hens absorb calcium in both the small and large intestines [152]. Intestinal calbindin-D28k mRNA expression in laying hens is twice as high in laying hens (25 weeks) than in immature hens (11-17 weeks). However, these immature hens do not show positivity (protein) for anti-calbindin-D28k [191]. Despite the mode of action of calbindin-D28k demonstrated in the aforementioned study in laying hens, it is still not known how it behaves in other laying species such as quails, and the raising of quails has been growing in the Northeast region of Brazil [28].
It is well established that there is an increased requirement for dietary calcium after sexual maturation in laying hens. This increase is reflected in the elevated expression of calbindin-D28k in the enterocytes of laying hens when there is egg production [191]. The concentration of calbindin-D28k would also be related to the amount of calcium transported to the eggshell in the uterus of laying hens [199], and these levels would be induced by the increase in estrogen. Therefore, calbindin-D28k would modulate the intestinal absorption capacity of calcium [3] and its deposition in the uterus [99,199], influencing the production and quality of the eggshell.
It is known that stress due to high temperatures (30-33°C) decreases the presence of calbindin-D28k in the ileum, cecum, colon, and uterus of birds, causing deterioration of eggshell quality under this condition [54]. This can be explained by the fact that heat stress reduces the conversion of vitamin D3 into its metabolically active form, 1,25 (OH)2D3, which is essential for the absorption and utilization of calcium [99]. In fact, the calcium requirement for laying hens increases under conditions of high environmental temperatures [3]. Therefore, the increase in expression of and positivity to Calbindin-D28k can also physiologically prove the better use of calcium after a specific diet or environmental treatment.
5. Conclusions
The incorporation of phytase into Japanese quail diets is a promising strategy to optimize poultry production. The ability of the phytase enzyme to release essential nutrients is crucial to improving performance and egg production, allowing quails to reach their maximum potential. In addition, phytases help to reduce the negative effects of heat stress on the functions of epithelial calcium transporters, preserving the digestive and reproductive integrity of laying quails. This ability to counteract the impacts of heat stress further highlights the benefits of phytases in poultry production, contributing to the sustainability and quality of production, in addition to promoting bird welfare. In summary, the addition of phytase to quail diets is an effective and comprehensive approach to increase production efficiency.
Author Contributions
Conceptualisation, A.G.R.; R.S.S.; D.A.S.; J.C.S.N.; L.F.A.S.; E.G.S.; J.E.S.R.; D.B.C.; C.V.B.V.A.; E.P.S.; F.G.P.C.; and R.R.G. writing—original draft preparation, A.G.R.; writing—review and editing, A.G.R. and R.R.G.; funding acquisition, R.R.G. All authors have read and agreed to the published version of the manuscript.
Funding
The Funding for this research was performed by National Council for Scientific and Technological Development (CNPq) Announcement nº 09/2020 - Research Productivity Grants, project no. 309318/2020-3 and Announcement 03/2020 - Research productivity PROPESQ-PRPG/UFPB Project PVN13248-2020 by through of the Cooperation Agreement by Universidade Federal da Paraíba (UFPB).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author.
Acknowledgments
The authors would like to thank the Center for Agricultural Sciences (CCA) of the Federal University of Paraíba (UFPB), the National Council for Scientific and Technological Development (CNPq) and the Coordination for the Improvement of Higher Education Personnel (CAPES).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Santos, T.C.; Gates, R.S.; Tinôco, I.F.F.; Zolnier, S.; Baêta, F.C. Behavior of Japanese quail in different air velocities and air temperatures. Pesquisa Agropecuária Brasileira 2017, 52, 344-354. [CrossRef]
- Truong, L.; Miller, M.R.; Sainz, R.D.; King, A.J. Changes in Japanese quail (Coturnix coturnix japonica) blood gases and electrolytes in response to multigenerational heat stress. PlosClimate 2023, 2, e0000144. [CrossRef]
- Moraes, L.R.; Delicato, M.E.A.; Cruz, A.S.; Silva, H.T.F.N.P.; Alves, C.V.B.V.; Campos, D.B.; Saraiva, E.P.; Costa, F.G.P.; Guerra, R.R. Methionine supplementing effects on intestine, liver and uterus morphology, and on positivity and expression of Calbindin-D28k and TRPV6 epithelial calcium carriers in laying quail in thermoneutral conditions and under thermal stress. PlosOne 2021, 16, e0245615. [CrossRef]
- Niu, Z.Y.; Liu, F.Z.; Yan, Q.L.; Li, W.C. Effects of different levels of vitamin E on growth performance and immune responses of broilers under heat stress. Poultry Science 2009, 88: 2101–2107. [CrossRef]
- Mehaisen, G.M.K.; Ibrahim, R.M.; Desoky, A.A.; Safaa, H.M.; El-Sayed, O.A.; Abass, A.O. The importance of propolis in alleviating the negative physiological effects of heat stress in quail chicks. PlosOne 2017, 12, 1–17. [CrossRef]
- Mehaisen, G.M.K.; Desoky, A.A.; Sakr, O.G.; Sallam, W.; Abass, A.O. Propolis alleviates the negative effects of heat stress on egg production, egg quality, physiological and immunological aspects of laying Japanese quail. PlosOne 2019, 14, e0214839. [CrossRef]
- Miller, D.B.; O’Callaghan, J.P. Neuroendocrine aspects of the response to stress. Metabolism 2002, 51, 5–10. [CrossRef]
- Mashaly, M.M.; Hendricks, G.L.; Kalama, M.A.; Gehad, A.E.; Abbas, A.O.; Patterson, P.H. Effect of Heat Stress on Production Parameters and Immune Responses of Commercial Laying Hens. Poultry Science 2004, 83, 889–894. [CrossRef]
- Sahin, K.; Sahin, N.; Onderci, M. Vitamin E supplementation can alleviate negative effects of heat stress on egg production, egg quality, digestibility of nutrients and egg yolk mineral concentrations of Japanese quails. Research in Veterinary Science 2002, 73, 307–312. [CrossRef]
- Vercese, F.; Garcia, E.A.; Sartori, J.R.; Silva, A.P.; Faitarone, A.B.G.; Berto, D.A.; Molino, A.B.; Pelícia, K. Performance and egg quality of Japanese quails submitted to cyclic heat stress. Brazilian Journal of Poultry Science 2012, 14, 37-41. [CrossRef]
- Akdemir, F.; Sahin, N.; Orhan, C.; Tuzcu, M.; Sahin, K.; Hayirli, A. Chromium-histidinate ameliorates productivity in heat-stressed Japanese quails through reducing oxidative stress and inhibiting heat-shock protein expression. British Poultry Science 2015, 56: 247–254. [CrossRef]
- Patra, T.; Pati, P.K.; Mohapatra, A.K. Study on carcass quality of coloured broiler chicks supplemented with vitamin E and C during summer stress. SAARC Agricultural Information Centre (SAIC) 2011, 9, 123–132. Available at: https://www.cabdirect.org/cabdirect/abstract/20123050083.
- Deng, W.; Dong, X.F.; Tong, J.M.; Zhang, Q. The probiotic Bacillus licheniformis ameliorates heat stress-induced impairment of egg production, gut morphology, and intestinal mucosal immunity in laying hens. Poultry Science 2012, 91: 575–582. [CrossRef]
- Sandikci, M.; Eren, U.; Onol, A.G.; Kum, S. The effect of heat stress and the use of Saccharomyces cerevisiae or (and) bacitracin zinc against heat stress on the intestinal mucosa in quails. Revue de Medecine Veterinaire 2004, 11, 552–556. Available at: https://www.researchgate.net/publication/289759420_The_effect_of_heat_stress_and_the_use_of_Saccharomyces_cerevisiae_or_and_bacitracin_zinc_against_heat_stress_on_the_intestinal_mucosa_in_quails.
- Sahin, N.; Orhan, C.; Tuzcu, M.; Sahin, K.; Kucuk, O. The Effects of Tomato Powder Supplementation on Performance and Lipid Peroxidation in Quail. Poultry Science 2008, 87, 276–283. [CrossRef]
- 16. Akdemir F, Köseman A; Şeker I. Alchemilla vulgaris effects on egg production and quality expressed by heat-stressed quail during the late laying period. South African Journal of Animal Science 2019, 49, 1-12. [CrossRef]
- Farias, M.R.S.; Leite, S.C.B.; Silva, H.P.; Pacheco, D.B.; Alves, G.C.; Abreu, C.G.; Freitas, E.R. Superdosing Phytases in the Diets of Light Laying Hens: Productive Performance and Bone Quality. Brazilian Journal of Poultry Science 2021, 23, 001-010. [CrossRef]
- Hirvonen, J.; Liljavirta, J.; Saarinen, M.T.; Lehtinen, M.J.; Ahonen, I.; Nurminen, P.I. Effect of Phytase on in Vitro Hydrolysis of Phytate and the Formation of myo-Inositol Phosphate Esters in Various Feed Materials. Journal of Agricultural and Food Chemistry 2019, 67, 11396−11402. [CrossRef]
- Sena, T.L.; Leite, S.C.B.; Vasconcelos, A.M.; Bezerra, M.M.R.; Abreu, C.G.; Farias, M.R.S.; Silveira, R.M.F. Does dietary supplementation with phytases affect the thermoregulatory and behavioral responses of pullets in a tropical environment? Journal of Thermal Biology 2020a, 88: 102499. [CrossRef]
- Pirgozliev, V.; Bedford, M.R.; Acamovic, T.; Allymehr, M. The effects of supplementary bacterial phytase on dietary energy and total tract amino acid digestibility when fed to young chickens. British Poultry Science 2011, 52, 245-254. [CrossRef]
- Martínez-Vallespín, B.; Männer, K.; Ader, P.; Zentek, J. Evaluation of high doses of phytase in a low-phosphorus diet in comparison to a phytate-free diet on performance, apparent ileal digestibility of nutrients, bone mineralization, intestinal morphology, and immune traits in 21-day-old broiler chickens. Animals 2022, 12, 1955. [CrossRef]
- Rojas, I.Y.M.; González, E.A.; Menocal, J.A.; Santos, T.T.; Arguello, J.R.; Coello, C.L. Assessment of a phytase included with lactic acid on productive parameters and on deposition of phosphorus, calcium, and zinc in laying hens fed with sorghum-soybean-meal-based diets. Journal of Applied Animal Research 2017, 46, 314-321. [CrossRef]
- Manobhavan, M.; Elangovan, A.V.; Sridhar, M.; Shet, D.; Ajith, S.; Pal, D.T.; Gowda, N.K.S. Effect of super dosing of phytase on growth performance, ileal digestibility and bone characteristics in broilers fed corn-soya-based diets. Journal of Animal Physiology and Animal Nutrition 2016, 100, 93-100. [CrossRef]
- Pinto, R.; Ferreira, A.S.; Albino, L.F.T.; Gomes, P.C.; Vargas Júnior, J.G. Protein and Energy Levels for Laying Japanese Quails. Revista Brasileira de Zootecnia 2002, 31, 1761-1770. [CrossRef]
- Vogado, G.M.S.; Silva, L.P. Características anatômicas e fatores genéticos ligados ao desenvolvimento reprodutivo de codornas de corte. In: I CONGEB. Editora Agron Science. 2023. p.119-133. [CrossRef]
- Lukanov, H.; Pavlova, I. Domestication changes in Japanese quail (Coturnix japonica): a review. World’s Poultry Science Journal 2020, 76, 787–801. [CrossRef]
- Lukanov, H. Domestic quail (Coturnix japonica domestica), is there such farm animal? World’s Poultry Science Journal 2019, 75, 547–558. [CrossRef]
- Pastore, S.M.; Oliveira, W.P.; Muniz, J.C.L. Panorama Da Coturnicultura No Brasil. Revista Eletrônica Nutritime 2012, 9, 2041–2049. Available at: https://portalidea.com.br/cursos/25e421f08de4aab6d494d4a76b957d11.pdf.
- Almeida, T.J.O.; Araújo, V.V.; Silva, A.V.; Silva, R.F.; Santos, N.A.; Santana, M.D.; Oliveira, V.P. Evolução Da Produção De Codornas Para Abate E Postura No Brasil. XIII Jornada de Ensino, Pesquisa e Extensão – JEPEX - UFRPE: Recife, Anais 2013. Available at: https://portalidea.com.br/cursos/6409d8df3ed7101311dc5da38592c83c.pdf.
- Silva, J.H.V.; Jordão Filho, J.; Costa, F.G.P.; Lacerda, P.B.; Vargas, D.G.V.; Lima, M.R. Exigências nutricionais de codornas. Revista Brasileira de Saúde e Produção Animal 2012, 13, 775–790. Available at: https://www.scielo.br/j/rbspa/a/kJDrRVLb6cMr7p6hskmZKzj/?format=pdf&lang=pt.
- Vieira, S.S. Desempenho e qualidade dos ovos de codornas japonesas (coturnix coturnix japônica) alimentadas com dietas contendo diferentes níveis óleo de palma. Diessertação (Mestre em Produção Animal), Universidade Federal Rural da Amazônia, Belém, Amazônia, Brasil, 2014. Available at: http://repositorio.ufra.edu.br/jspui/handle/123456789/754.
- Nasar, A.; Rahman, A.; Hoque, N.; Kumar Talukder, A.; Das, Z.C. A survey of Japanese quail (Coturnix coturnix japonica) farming in selected areas of Bangladesh. Veterinary World 2016, 9(9): 940-947. [CrossRef]
- Matos Júnior, J.J.L.; Furtado, D.A.; Ribeiro, N.L.; Marques, J.I.; Leite, P.G.; Nascimento, J.W.B.; Rodrigues, V.P.; Lopes Neto, J.P.; Rodrigues, L.R.; Santos, S.G.C.G.; Oliveira, A.G.; Silva, R.S. Productive performance, egg quality and the morphometry of the organs of Japanese quails (Cotournix cotournix japônica) kept at different temperatures. Food Science and Technology 2023, 43, 117822. [CrossRef]
- Sarcinelli, M.F.; Venturini, K.S.; Silva, L.C. Características dos ovos. Boletim Técnico, Universidade Federal do Espirito Santo. 2007. Available at: https://www.agais.com/telomc/b00707_caracteristicas_ovos.pdf.
- Carvalho, F.B.; Stringhini, J.H.; Jardim Filho, R.M.; Leandro, N.S.M.; Café, M.B.; Deus, H.A.S.B. Qualidade interna e de casca para ovos de poedeiras comerciais de diferentes linhagens e idades. Ciência Animal Brasileira 2007, 8(1): 25-30. 2007. https://www.revistas.ufg.br/vet/article/view/1155.
- Mota, A.S.B.; Lima, P.M.S.; Silva, D.S.; Abreu, V.K.G.; Freitas, E.R.; Pereira, A.L.F. Internal quality of eggs coated with cassava and yam starches. Revista Brasileira de Ciências Agrárias 2017, 12, 47-50. [CrossRef]
- NRC. Nutrient Requirements of Poultry. 9th ed. National Academics Press, Washington, DC, 1994. Available at: https://books.google.com.br/books?hl=pt-BR&lr=&id=bbV1FUqRcM0C&oi=fnd&pg=PT13&dq=NRC+1994+Nutrient+Requirements+of+Poultry.+9th+ed.+National+Academics+Press,+Washington&ots=IleM4AkqTs&sig=2Ll0iW3NGbzHsQc19UoHD17wtxM#v=onepage&q&f=false.
- INRA - Institut National de la Recherche Agronomique. Alimentação dos animais monogástricos: Suínos, Coelhos e Aves. 2.ed. São Paulo: Roca, 1999. 245p.
- Silva, J.H.V.; Costa, F.G.P. Tabelas para codornas japonesas e europeias. 2.ed. Jaboticabal: FUNEP, 2009. 107p.
- Rostagno, H.S.; Albino, L.F.T.; Hannas, M.I.; Donzele, J.L.; Sakomura, N.K.; Perazzo, F.G.; Brito, C.O. Brazilian tables for poultry and swine : composition of feedstuffs and nutritional requirements, 4 ed. Viçosa, MG. 2017. 488p.
- Rostagno, H.S.; Albino, L.F.T.; Calderano, A.A.; et al. Tabelas Brasileiras para aves e suínos. 5th edn. (Universidade Federal de Viçosa: Viçosa). 2024. 576p.
- Silva, R.C. Trocas de calor e desempenho de codornas japonesas confinadas em ambiente termoneutro e sob estresse térmico. Tese (Doutor em Engenharia Agrícola), Programa de Pós- Graduação em Engenharia Agrícola do Centro de Tecnologia e Recursos Naturais da Universidade Federal de Campina Grande, Campina Grande - Paraíba, Brasil. 2017. 108p. Available at: http://dspace.sti.ufcg.edu.br:8080/jspui/handle/riufcg/28065.
- Kim, D.H.; Kim, Y.B.; Lee, S.H.; Lee, Y.K.; Lee, S.D.; Lee, K.W. Identical thermal stress coupled with different temperature and humidity combinations affects nutrient digestibility and gut metabolites of laying hens. Brazilian Journal of Animal Science 2023, 52, e20220067. [CrossRef]
- Teyssier, J.R.; Brugaletta, G.; Sirri, F.; Dridi, S.; Samuel, J.; Rochell, S.J. A review of heat stress in chickens. Part II: Insights into protein and energy utilization and feeding. Frontiers in Physiology 2022, 13. [CrossRef]
- Wasti, S.; Sah, N.; Mishra, B. Impact of Heat Stress on Poultry Health and Performances, and Potential Mitigation Strategies. Animals 2020, 10, 1266. [CrossRef]
- Linsley, J.G.; Burger, R.E. Respiratory and cardiovascular responses in the hyperthermic domestic cock. Poultry Science 1964, 43, 291-305. [CrossRef]
- Vercese, F. Efeito da temperatura sobre o desempenho e a qualidade dos ovos de codornas japonesas. Dissertação (Mestre em Zootecnia), Programa de Pós-Graduação em Zootecnia da universidade Estadual Paulista - Faculdade de Medicina Veterinária e Zotecnia, Botucatu - São Paulo. 2010. 70p. Available at: https://www.fmvz.unesp.br/Home/ensino/pos-graduacao768/zootecnia/dissertacoeseteses/francine-vercese.pdf.
- Furlan, R.L.; Macari, M.; Moraes, V.M.B. Malheiros, R.D.; Malheiros, E.B.; Secato, E.R. Alterações hematológicas e gasométricas em diferentes linhagens de frangos de corte submetidos ao estresse calórico agudo. Revista Brasileira de Ciência Avícola 1999, 1, 77-84.
- Ruzal, M.; Shinder, D.; Malka, I.; Yahav, S. Ventilation plays an important role in hens’ egg production at high ambient temperature. Poultry Science 2011, 90, 856-862. [CrossRef]
- Abdulkadir, A.; Reddy, D. A scoping review of the impact of heat stress on the organs of the Japanese quail (Coturnix japonica). The Journal of Basic and Applied Zoology 2023, 84. [CrossRef]
- Lesson, S.; Summers, J. D. Commercial poultry nutrition, Nottingham. Guelph: University Books, 1991. 283 p.
- Mongin, P. Role of acid-base balance in the physiology of egg-shell formation. World’s Poultry Science Journal 1968, 24, 200–230. [CrossRef]
- Campos, E.J. Avicultura: razões fatos e divergências. Belo Horizonte: Editora FEPMVZ; 2000. 311p.
- Ebeid, T.A.; Suzuki, T.; Sugiyama, T. High ambient temperature influences eggshell quality and calbindin-D28k localization of eggshell gland and all intestinal segments of laying hens. Poultry Science 2012, 91, 2282–2287. [CrossRef]
- Sahin, K.; Sahin, N.; Kucuk, O.; Hayirli, A.; Prasad, A.S. Role of dietary zinc in heat-stressed poultry: A review. Poultry Science 2009, 88, 2176-2183. [CrossRef]
- Melo, A.S.; Fernandes, R.T.V.; Marinho, J.B.M.; Arruda, A.M.V.; Figueirêdo, L.C.; Fernandes, R.T.V. Relação temperatura e nutrição sobre o desempenho de galinhas poedeiras. PUBVET 2016, 10, 855-860. [CrossRef]
- Kumar, S.; Anand, R. Effect of Germination and Temperature on Phytic Acid Content of Cereals. International Journal of Research in Agricultural Sciences 2021, 8, 1–13. Available at: https://ijras.org/administrator/components/com_jresearch/files/publications/IJRAS_932_FINAL.pdf.
- Kim, D.H.; Lee, Y.K.; Lee, S.D.; Kim, S.H.; Lee, S.R.; Lee, H.G.; Lee, K.W. Changes in Production Parameters, Egg Qualities, Fecal Volatile Fatty Acids, Nutrient Digestibility, and Plasma Parameters in Laying Hens Exposed to Ambient Temperature. Frontiers in Veterinary Science 2020, 7, 412. [CrossRef]
- Lehrfeld, J. High-performance Liquid Chromatography Analysis of Phytic Acid on a pH-stable, Macroporous Polymer Column. Cereal Chemistry 1989, 66, 510–515. Available at: https://www.cerealsgrains.org/publications/cc/backissues/1989/Documents/66_510.pdf.
- Bloot, A.P.M.; Kalschne, D.L.; Amaral, J.A.S.; Baraldi, I.J.; Canan, C. A Review of Phytic Acid Sources, Obtention, and Applications. Food Reviews International 2021, 39, 73-92. [CrossRef]
- Lolas, G.; Palamidis, N.; Markakis, P. Phytic Acid Total Phosphorus Relationship Relationship in Barley, Oats, Soybeans and Wheat. Cereal Chemistry 1976, 53, 867–871. Available at: https://www.cerealsgrains.org/publications/cc/backissues/1976/Documents/chem53_867.pdf.
- Stein, H.H. Analyzed values for P and phytate in feed ingredients. Monogastric Nutrition Laboratory, 2023. Available at: https://nutrition.ansci.illinois.edu/node/1753.
- Ravindran, S.; Ravindran, V.; Sivalogan, G. Total and Phytate Phosphorus Contents of Various Foods and Feedstuffs of Plant Origin. Food Chemistry 1994, 50, 133–136. [CrossRef]
- Frossard, E.; Bucher, M.; Mächler, F.; Mozafar, A.; Hurrell, R. Potential for Increasing the Content and Bioavailability of Fe, Zn and Ca in Plants for Human Nutrition. Journal of the Science of Food and Agriculture 2000, 80, 861–879. [CrossRef]
- Banaszkiewicz, T. Nutritional Value of Soybean Meal. Soybean and Nutrition. InTech. 2011. [CrossRef]
- Kasim, B.; Edwards,; H.M. The Analysis of Inositol Phosphate Forms in Feed Ingredients. Journal of the Science of Food and Agriculture 1998, 76, 1–9. [CrossRef]
- Canan, C.; Cruz, F.T.L.; Delaroza, F.; Casagrande, R.; Sarmento, C.P.M.; Shimokomaki, M.; Ida, E.I. Studies on the Extraction and Purification of Phytic Acid from Rice Bran. Journal of Food Composition and Analysis 2011, 24, 1057–1063. [CrossRef]
- Garcı́a-Estepa, R.; García-Estepa, R. M.; Guerra-Hernández, E.; García-Villanova, B. Phytic Acid Content in Milled Cereal Products and Breads. Food Research International 1999, 32, 217–221. [CrossRef]
- Hu, Y.X.; Van Harn, J.; Hendriks, W.H.; Van Baal, J.; Dijkslag, M.A.; Van Krimpen, M.M.; Bikker, P. Low-calcium diets increase duodenal mRNA expression of calcium and phosphorus transporters and claudins but compromise growth performance irrespective of microbial phytase inclusion in broilers. Poultry Science 2021, 100, 101488. [CrossRef]
- Figueirêdo, A.V.; Fialho, E.T.; Vitti, D.M.S.S.; Lopes, J.B.; Silva Filho, J.C.; Teixeira, A.S.; Lima, J.A.F. Ação da Fitase sobre a Disponibilidade Biológica do Fósforo, por Intermédio da Técnica de Diluição Isotópica, em Dietas com Farelo de Arroz Integral para Suínos. Revista Brasileira de Zootecnia 2000, 29, 177–182. [CrossRef]
- Payne, R.L.; Lavergne, T.K.; Southern, L.L. A comparison of two sources of phytase in liquid and dry forms in broilers. Poultry Science 2005, 84, 265–272. [CrossRef]
- Woyengo, T.A.; Nyachoti, C.M. Review: Supplementation of phytase and carbohydrases to diets for poultry. Canadian Journal of Animal Science 2011, 91, 177–192. [CrossRef]
- Costello, A.J.R.; Glonek, T.; Myers, T.C. 31P Nuclear Magnetic Resonance-pH Titrations of Myo-Inositol Hexaphosphate. CarbohyrIrate Research 1976, 46, 159-171. [CrossRef]
- Vasconcelos, D.M. Diferentes níveis nutricionais e de fitase nas dietas para codornas japonesas. Dissertação (Mestre em Zootecnia) Centro de Ciências Agrárias da Universidade Federal da Paraíba-UFPB, Areia, Paraíba – Brasil, 2018. Available at: https://repositorio.ufpb.br/jspui/bitstream/123456789/14997/1/DZ329.pdf.
- Gautier, A.E.; Walk, C.L.; Dilger, R.N. Effects of a high level of phytase on broiler performance, bone ash, phosphorus utilization, and phytate dephosphorylation to inositol. Poultry Science 2017, 97, 211–218. [CrossRef]
- Alves, N.M.; Guimarães, L.H.S.; Piccoli, R.H.; Cardoso, P.G. Production and Partial Characterizationof an Extracellular Phytase Produced by Muscodor sp. under SubmergedFermentation. Advances in Microbiology 2016, 6, 23-32. [CrossRef]
- Rezaeipour, V.; Barsalani, A.; Abdullahpour, R. Effects of phytase supplementation on growth performance, jejunum morphology, liver health, and serum metabolites of Japanese quails fed sesame (Sesamum indicum) meal-based diets containing graded levels of protein. Tropical Animal Health and Production 2016, 48, 1141–1146. [CrossRef]
- Lelis, G.R.; Albino, L.F.T.; Silva, C.R.; Rostagno, H.S.; Gomes, P.G.; Borsatto, CG. Suplementação dietética de fitase sobre o metabolismo de nutrientes de frangos de corte. Revista Brasileira de Zootecnia 2010. 39, 1768-1773. [CrossRef]
- Jatuwong, K.; Suwannarach, N.; Kumla, J.; Penkhrue, W.; Kakumyan, P.; Lumyong, S. Bioprocess for Production, Characteristics, and Biotechnological Applications of Fungal Phytases. Frontiers in Microbiology 2020, 11, 188. [CrossRef]
- Sena, T.L.; Leite, S.C.B.; Farias, M.R.S.; Abreu, C.G.; Freitas, E.R.; Costa, A.C. Phytase Superdosing in the Diet of Lightweight Replacement Pullets: Performance, Organ Biometry and Bone Characteristics. Brazilian Journal of Poultry Science 2020b, 22, 001-008. [CrossRef]
- Kriseldi, R.; Walk, C.L.; Bedford, M.R.; Dozier, W.A. Inositol and gradient phytase supplementation in broiler diets during a 6-week production period: 2. Effects on phytate degradation and inositol liberation in gizzard and ileal digesta contentes. Poultry Science 2021, 100, 100899. [CrossRef]
- Vats, P.; Banerjee, U.C. Production studies and catalytic properties of phytases (myo-inositolhexakisphosphate phosphohydrolases): an overview. Enzyme and Microbial Technology 2004, 35, 3-14. [CrossRef]
- Sato, V.S.; Jorge, J.A.; Oliveira, W.P.; Souza, C.R.F.; Guimarães, L.H.S. Phytase production by Rhizopus microsporus var. microsporus biofilm: characterization of enzymatic activity after spray drying in presence of carbohydrates and nonconventional adjuvants. Journal of Microbiology and Biotechnology 2014, 24, 177-87. [CrossRef]
- Dailin, D.J.; Hanapi, S.Z.; Elsayed, E.A.; Sukmawati, D.; Azelee, N.I.W.; Eyahmalay, J.; Siwapiragam, V.; El Enshasy, H. Fungal Phytases: Biotechnological Applications in Food and Feed Industries. Recent Advancement in White Biotechnology Through Fungi 2019, 65-99. [CrossRef]
- Greiner, R.; Konietzny, U. Phytases: Biochemistry, enzymology and characteristics relevant to animal feed use. In: Enzymes in farm animal nutrition, Bedford MR, Partridge GG. (eds). CAB Intl. Publishing, Oxfordshire, UK: 2010; pp.96–128. https://www.researchgate.net/publication/286044581_Phytases_Biochemistry_Enzymology_and_Characteristics_Relevant_to_Animal_Feed_Use.
- Bhavsar, K.; Khire, J.M. Current research and future perspectives of phytase bioprocessing. Royal Society of Chemistry advances 2014, 4, 26677–26691. [CrossRef]
- Santos, K.O.; Costa-Filho, J.; Riet, J.; Spagnol, K.L.; Nornberg, B.F.; Kütter, M.T.; Tesser, M.B.; Marins, L.F. Probiotic expressing heterologous phytase improves the immune system and attenuates inflammatory response in zebrafish fed with a diet rich in soybean meal. Fish Shellfish Immunol 2019, 93, 652-658. [CrossRef]
- Greiner, R.; Alminger, M.L. Carlsson, N.G. Stereospecificity of myo-Inositol Hexakisphosphate Dephosphorylation by a Phytate-Degrading Enzyme of Baker’s Yeast. Journal of Agricultural and Food Chemistry 2001, 49, 2228-2233. [CrossRef]
- Naves, L.P.; Corrêa, A.D.; Bertechini, A.G.; Gomide, E.M.; Santos, C.D. Effect of ph and Temperature on the Activity of Phytase Products Used in Broiler Nutrition. Brazilian Journal of Poultry Science 2012, 14, 159-232. [CrossRef]
- Delmaschio, I.B. Produção de fitases por fermentação em estado sólido e imobilização das enzimas por spray drying. Dissertação (Mestre em Ciências da Microbiologia) Programa de Pós-Graduação em Microbiologia, Área de Concentração – Microbiologia Industrial e Aplicada, do Instituto de Biociências, Letras e Ciências Exatas da Universidade Estadual Paulista (UNESP) “Júlio de Mesquita Filho”, Campus de São José do Rio Preto – Brasil. 2014. 115p. https://bdtd.ibict.br/vufind/Record/UNSP_5ea0a9a9549bebefeae1fbf3090e0ce6.
- Nezhad, N.G.; Rahman, R.N.Z.R.A.; Normi, Y.M.; Oslan, S.N.; Shari, F.M.; Leow, T.C. Integrative Structural and Computational Biology of Phytases for the Animal Feed Industry. Catalysts 2020, 10, 844. [CrossRef]
- Dallmann, H.M.; Avila, V.S.; Krabbe, E.L.; Surek, D.; Bedendo, G.C.; Toledo, T.S.; Dallmann, P.R.; Roll, A.A.P.; Roll, V.F.B.; Rutz, F. Different phytase levels and energy densities in broiler diets on performance, nutrient digestibility, and bone integrity from 28 to 35 days of age. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 2023, 75, 280-292. [CrossRef]
- Cowieson, A.J.; Wilcock, P.; Bedford, M.R. Super-dosing effects of phytase in poultry and other monogastrics. World’s Poultry Science Journal 2011, 67, 225 – 236. [CrossRef]
- Nelson, T.S.; Shieh, T.R.; Wodzinski, R.J.; Ware, J.H. Effect of supplemental phytase on the utilization of phytate phosphorus by chicks. The Journal of Nutrition 1971, 101, 1289-1294. [CrossRef]
- Walk, C.L.; Bedford, M.R.; Santos, T.S.; Paiva, D.; Bradley, J.R.; Wladecki, H.; Honaker, C.; McElroy, A.P. Extra-phosphoric effects of superdoses of a novel microbial phytase. Poultry Science 2013, 92, 719–725. [CrossRef]
- Fernandes, J.I.M.; Horn, D.; Ronconi, E.J.; Buzim, R.; Lima, F.K.; Pazdiora, D.A. Effects of Phytase Superdosing on Digestibility and Bone Integrity of Broilers. Journal of Applied Poultry Research 2019, 28, 390–398. [CrossRef]
- Leyva-Jimenez, H.; Alsadwi, A.M.; Gardner, K.; Voltura, E.; Bailey, C.A. Evaluation of high dietary phytase supplementation on performance, bone mineralization, and apparent ileal digestible energy of growing broilers. Poultry Science 2019. 98, 811-819. [CrossRef]
- Kim, J.H.; Pitargue, F.M.; Jung, H.; Han, G.P.; Choi, H.S.; Kil, D.Y. Effect of superdosing phytase on productive performance and egg quality in laying hens. Asian-Australasian Journal of Animal Sciences 2017, 30, 994-998. [CrossRef]
- Ribeiro, A.G.; Silva, R.S.; Costa, F.S.; Silva, E.G.; Santos Ribeiro, J.E.; Saraiva, E.P.; Costa, F.G.P.; Guerra, R.R. (2024) Phytase super-dosing modulates bone parameters and the concentration of the calcium epithelial carrier Calbindin-D28k in quails (Coturnix coturnix japonica) under thermal stress. Animal Production Science 2024, AN24057. [CrossRef]
- Saeed, M.; Abbas, G.; Alagawany, M.; Ali Kamboh, A.; Abd El-Hack, M.E.; Khafaga, A.F.; Chao, S. Heat stress management in poultry farms: A comprehensive overview. Journal of Thermal Biology 2019, 84, 414-425. [CrossRef]
- Farag, M.R.; Alagawany, M. Physiological alterations of poultry to the high environmental temperature. Journal of Thermal Biology 2018, 76, 101-106. [CrossRef]
- Farias, M.R.S.; Leite, S.C.B.; Vasconcelos, A.M.; Silva, T.A.G.; Leitão, A.M.F.; Sena, T.L.; Pacheco, D.B.; Abreu, C.G.; Silveira, R.M.F. Thermoregulatory, behavioral and productive responses of laying hens supplemented with different types and dosages of phytases raised in a hot environment: An integrative approach. Journal of Thermal Biology 2020, 94, 102773. [CrossRef]
- Dersjant-Li, Y.; Awati, A.; Schulze, H.; Partridge, G. Phytase in non ruminant animal nutrition: a critical review on phytase activities in the gastrointestinal tract and influencing factors. Journal of the Science of Food and Agriculture 2014, 95, 878-896. [CrossRef]
- Freeland-Graves, J.H.; Sanjeevi, N.; Lee, j.j. Global perspectives on trace element requirements. Journal of Trace Elements in Medicine and Biology 2015, 31, 135-141. [CrossRef]
- Borges, G.C.S. Peroxidação lipídica, desempenho e características de carcaça de frangos de corte estressados pelo calor e suplementados com zinco e selênio. Dissertação (Mestrado em Ciências Veterinárias). Faculdade de Medicina Veterinária, Uberlândia, Minas Gerais, Brasil. 2008. 61p. Available at: https://repositorio.ufu.br/handle/123456789/12987.
- Hu, P.; Li, K.; Peng, X.; Yao, T.; Zhu, C.; Gu, H.; Liu, H-Y.; Sun, M.; Hu, Y.; Ennab, W.; Luo, X.; Cai, D. Zinc intake ameliorates intestinal morphology and oxidative stress of broiler chickens under heat stress. Frontiers in Immunology 2023, 14, 1308907. [CrossRef]
- Kazim, S.; Omer, K. Zinc Supplementation Alleviates Heat Stress in Laying Japanese Quail. The Journal of Nutrition 2003, 133, 2808-2811. [CrossRef]
- Prasad, A.S.; Kucuk, O. Zinc in cancer prevention. Cancer and Metastasis Reviews 2002, 21, 291-295. [CrossRef]
- Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. The Role of Metallothionein in Oxidative Stress. International Journal of Molecular Science 2013, 14 6044–6066. [CrossRef]
- Zhang, Y.N.; Zhang, H.J.; Wang, J.; Yue, H.Y.; Qi, X.L.; Wu, S.G.; Qi, G.H. Effect of dietary supplementation of organic or inorganic zinc on carbonic anhydrase activity in eggshell formation and quality of aged laying hens. Poultry Science 2017, 96, 2176-2183. [CrossRef]
- Benesch, R.; Barron, N.S.; Mawson, C.A. Carbonic anhydrase, sulphonamides and shell formation in the domestic fowl, Nature 1944, 153, 138-139. [CrossRef]
- Nys, Y.; Hincke, M.; Arias, J.; Garcia-Ruiz, J.; Solomon, S. Avian eggshell mineralization. Poultry and Avian Biology Reviews 1999, 10, 143-166. Available at: https://www.researchgate.net/profile/Yves-Nys/publication/279562431_Avian_Eggshell_Mineralization/links/5630e38208ae0530378cf614/Avian-Eggshell-Mineralization.pdf.
- Holm, L.; Blomqvist, A.; Brandt, I.; Brunström, B.; Ridderstråle, Y.; Berg, C. Embryonic exposure to o, p’-DDT causes eggshell thinning and altered shell gland carbonic anhydrase expression in the domestic hen. Environmental Toxicology and Chemistry 2006, 25, 2787-2793. [CrossRef]
- Mohebbifar, A.; Torki, M.; Ghasemi, H.A. Effect of phytase supplementation of diets with different levels of rice bran and non-phytate phosphorus on productive performance, egg quality traits, leukocytes profile and serum lipids of laying hens reared indoor under high environmental temperatures. Animal Feed Science and Technology 2015, 207, 222-233. [CrossRef]
- Hezaveh, M.S.S.; Ghasemi, H.A.; Hajkhodadadi, I.; Moradi, M.H. Single and combined effects of phytase and citric acid on growth performance, nutrient digestibility, bone characteristics, intestinal morphology, and blood components in meat-type quails fed low-phosphorous diets. Animal Feed Science and Technology 2020, 269, 114677. [CrossRef]
- Lima, H.J.D.A.; Barreto, S.L.T.; Donzele, J.L.; Valeriano, M.H.; Vieira, P.A.F.; Costa, C.H.R. Dietary phytase levels on performance and egg quality of Japanese quails. Revista Brasileira de Zootecnia 2011, 40, 129-134. [CrossRef]
- Gouveia, A.B.V.S.; Paulo, L.M.; Dias, A.G.F.; Batista, J.M.; Brasileiro, J.C.L.; Minafra, C.S.; Stringhini, J.H. Fitase na nutrição de aves de corte e postura: revisão de literatura. II Congresso Brasileiro de Produção Animal e Vegetal: “Produção Animal e Vegetal: Inovações e Atualidades 2022. 2, 984. [CrossRef]
- Ribeiro, C.L.N.; Barreto, S.L.T.; Reis, R.S.; Muniz, J.C.L.; Viana, G.S.; Ribeiro Junior, V.; Mendonça, M.O.; Ferreira, R.C.; DeGroot, A.A. The Effect of Calcium and Available Phosphorus Levels on Performance, Egg Quality and Bone Characteristics of Japanese Quails at End of the Egg-Production Phase. Brazilian Journal of Poultry Science 2016, 18, 033-040. [CrossRef]
- Stanquevis, C.E.; Furlan, A.C.; Marcato, S.M.; Oliveira-Bruxel, T.M.; Perine, T.P.; Finco, E.M.; Grecco, E.T.; Benites, M.I.; Zancanela, V.T. Calcium and available phosphorus requirements of Japanese quails in early egg-laying stage. Poultry Science 2021, 100, 147-158. [CrossRef]
- Souza, C.S.; Barreto, S.L.T.; Vieites, F.M.; Calderano, A.A.; Moraes, G.H.K.; Oliveira, M.G.A. Cálcio e fósforo na nutrição de codornas japonesas em postura. Science and Animal Health 2017, 5, 260-281. Available at: https://periodicos.ufpel.edu.br/index.php/veterinaria/article/view/9166/8350.
- Lautrou, M.; Pomar, C.; Dourmad, J.-Y.; Narcy, A.; Schmidely, P.; Létourneau-Montminy, M.P. Phosphorus and calcium requirements for bone mineralisation of growing pigs predicted by mechanistic modelling. Animal 2020, 14, s313–s322. [CrossRef]
- David, L.S.; Anwar, M.N.; Abdollahi, M.R.; Bedford, M.R.; Ravindran, V. Calcium Nutrition of Broilers: Current Perspectives and Challenges. Animals 2023, 13, 1590. [CrossRef]
- Pelicia, K.; Garcia, E.A.; Faitarone, A.B.G.; Silva, A.P.; Berto, D.A.; Molino, A.B.; Vercese, F. Calcium and Available Phosphorus Levels for Laying Hens in Second Production Cycle. Brazilian Journal of Poultry Science 2009, 11, 39 - 49. Available at: https://www.scielo.br/j/rbca/a/VgmDHbKHdXPv4vKMXvb9qhk/?format=pdf&lang=en.
- Carvalho, L.S.S.; Fernandes, E.A. Formation and eggshell quality laying and breeding hens. Medicina Veterinária, Recife 2013, 7, 35-44. Available at: https://www.journals.ufrpe.br/index.php/medicinaveterinaria/article/view/604.
- Mello, J. F. Influência dos níveis de cálcio e fósforo na dieta de matrizes de codornas japonesas, no desempenho produtivo e no desenvolvimento ósseo embrionário da progênie. Dissertação (Mestrado em Zootecnia), Programa de Pós-Graduação em Zootecnia, Universidade Estadual de Maringá, Paraná. 2015. 82p. Available at: http://repositorio.uem.br:8080/jspui/bitstream/1/1764/1/000220522.pdf.
- Chaves Hernández, A.J. Poultry and Avian Diseases. Encyclopedia of Agriculture and Food Systems. 2014; pp.504–20. [CrossRef]
- Zhao, S.C.; Teng, X.Q.; Xu, D.L.; Chi, X.; Ge, M.; Xu, S.W. Influences of low level of dietary calcium on bone characters in laying hens. Poultry Science 2020, 99, 7084-7091. [CrossRef]
- Serna, J.; Bergwitz, C. Importance of Dietary Phosphorus for Bone Metabolism and Healthy Aging. Nutrients 2020, 12, 3001. [CrossRef]
- Adedokun, S.A.; Adeola, O. Calcium and phosphorus digestibility: metabolic limits. The Journal of Applied Poultry Research 2013, 22, 600-608. [CrossRef]
- IFP - Inorganic Feed phosphates. Phosphorus: a vital source of animal nutrition. 2024. Available at: https://www.feedphosphates.org/index.php/guides/11-guides/11-phosphorus-a-vital-source-of-animal-nutrition.
- Sinclair-Black, M.; Garcia-Mejia, R.A.; Blair, L.R.; Angel, R.; Arbe, X.; Cavero, D.; Ellestad, L.E. Circadian regulation of calcium and phosphorus homeostasis during the oviposition cycle in laying hens. Poultry Science 2024, 103, 103209. [CrossRef]
- Mazzuco, H. Osteoporose em poedeiras comerciais: uma doença metabólica multifatorial. Circular Técnica, n. 43, Concórdia: Embrapa Aves e Suínos, 2005; pp.1-8. Available at: https://ainfo.cnptia.embrapa.br/digital/bitstream/item/57891/1/CUsersPiazzonDocumentsCIT-43.pdf.
- Miranda, C.C. Formas inorgânicas e orgânicas de minerais e Temperatura ambiente sobre o desempenho, Imunidade e parâmetros sanguíneos em frangos de Corte. Dissertação (Mestre em Zootecnia), Programa de Pós-Graduação em Zootecnia da Universidade Estadual Paulista, Faculdade de Medicina Veterinária e Zootecnia, Campus de Botucatu, Botucatu - SP, 2010. 59p. Available at: https://www.fmvz.unesp.br/Home/ensino/pos-graduacao768/zootecnia/dissertacoeseteses/carolina-carvalho-de-miranda.pdf.
- Bertechini, A. G. Metabolismo dos minerais. In: Bertechini, A. G. Nutrição de monogástricos. Lavras: Editora UFLA, 2012. p. 207-255.
- Silva, A.P. Níveis de cálcio e fósforo na dieta de codornas Japonesas (coturnix coturnix japonica) em diferentes fases do ciclo de produção e seus efeitos sobre desempenho Produtivo e qualidade dos ovos. Dissertação (Mestre em Zootecnia), Programa de Pós-Graduação em Zootecnia da Universidade Estadual Paulista, Faculdade de Medicina Veterinária e Zootecnia, Campus de Botucatu, Botucatu - SP, 2011. 58p. Available at: https://repositorio.unesp.br/server/api/core/bitstreams/d866ecc9-3f1e-4417-a647-48ead5a8c2f5/content.
- Albino, L.F.T.; Barreto, S.L.T. Criação de codornas para produção de ovos e carne. Viçosa: Aprenda Fácil, 2003. 268p.
- Pedroso, A.A.; Moraes, V.M.B.; Ariki, J.; Kronka, S.N. Níveis de cálcio e fósforo na ração sobre o desempenho e qualidade dos ovos de codornas japonesas. Ars Veterinaria 1999, 15, 135 –139.
- Masukawa, Y.; Fernandes, E.; Moraes, V.; Ariki, J.; Bruno, L. Níveis de cálcio da dieta sobre o desempenho e a qualidade da casca de ovos de codornas japonesas. Ars Veterinária 2001, 17, 144-148.
- Brandão, P.A.; Costa, F.G.P.; Silva, J.H.V.; Brandão, J.S.; Nobre, J.G.S.; Goulart, C.C. Exigência de cálcio para codornas japonesas (coturnix coturnix japonica) em postura. Acta Scientiarum. Animal Sciences 2007, 29, 17-21. Available at: https://www.redalyc.org/pdf/3031/303126486001.pdf.
- Costa, C.H.R.; Barreto, S.L.T.; Moura, W.C.O.; Reis, R.S.; Leite, C.D.S.; Maia, G.V.C. Níveis de fósforo e cálcio em dietas para codornas japonesas em postura. Revista Brasileira de Zootecnia 2007, 36, 2037-2046. [CrossRef]
- Costa, C.H.R.; Barreto, S.L.T.; Umigi, R.T.; Lima, H.J.D.A.; Araujo, M.S.; Medina, P. Balanço de cálcio e fósforo e estudo dos níveis desses minerais em dietas para codornas japonesas (45 a 57 semanas de idade). Revista Brasileira de Zootecnia 2010a, 39, 1748-1755. [CrossRef]
- Costa, C.H.R.; Barreto, S.L.T.; Gomes, P.C.; Maia, G.V.C.; Lipari, C.A.; Hosoda, L.H. Teores de cálcio em dietas para codornas japonesas no terço final de postura (45 a 57 semanas de idade). Arquivo Brasileiro de Medicina Veterinária e Zootecnia 2010b, 62, 1225-1231. [CrossRef]
- Amoah, J.K.; Martin, E.A.; Barroga, A.J.; Garillo, E.P.; Domingo, I. Calcium and phosphorus requirements of Japanese quail layers. Journal of Applied Biosciences 2012, 54, 3892-3900. Available at: https://m.elewa.org/JABS/2012/54/5.pdf.
- Vieira, D.V.G.; Barreto, S.L.T.; Valeriano, M.H.; Jesus, L.F.D.; Silva, L.F.F.; Mencalha, R.; Barbosa, K.S.; Mendes, R.K.V.; Cassuce, M.R.; Melo, T.S. Exigências de cálcio e de fósforo disponível para codornas japonesas de 26 a 38 semanas de idade. Revista Brasileira de Saúde e Produção Animal 2012, 13, 204-213. Available at: https://www.scielo.br/j/rbspa/a/NvXwgKTH5RX5gJQmx4DkHsp/?format=pdf&lang=pt.
- Nascimento, M.Q. Níveis de cálcio e de fósforo em dietas para codornas japonesas utilizando fosfato bicálcico com duas granulometrias. Dissertação (Mestrado em Ciências Veterinárias), Universidade Federal do Espírito Santo, Alegre-ES, 2013. 84p. Available at: https://repositorio.ufes.br/server/api/core/bitstreams/52bfca24-9ea6-4806-87f3-a81e761c8309/content.
- Aguda, A.Y.; Sekoni, A.A.; Omage, J.J. Requirement of calcium and available phosphorus for laying Japanese quail birds (Coturnix coturnix japonica) in Nigeria. Journal of Animal and Poultry Sciences 2015, 4, 31-38. Available at: https://www.researchgate.net/publication/313387866_Requirement_of_Calcium_and_available_Phosphorus_for_Laying_Japanese_quail_birds_Coturnix_coturnix_japonica_in_Nigeria.
- Nascimento, M.Q.; Vargas Junior, J.G.; Pinto, C.E.L.; Demuner, L.F.; Petrucci, F.B.; Vieites, F.M. Optimal Available Phosphorus Levels in Diets Containing Different Dicalcium Phosphate Particle Sizes for Japanese Quails. Brazilian Journal of Poultry Science 2019, 21, 001-008. [CrossRef]
- Yang, J.H.; Hou, J.F.; Farquharson, C.; Zhou, Z.L.; Deng, Y.F.; Wang, L.; Yu, Y. Localisation and expression of TRPV6 in all intestinal segments and kidney of laying hens. British Poultry Science 2011, 52, 507–516. [CrossRef]
- Yu, E.; Sharma, S. Physiology, Calcium. StatPearls, 2023. Available at: https://www.ncbi.nlm.nih.gov/books/NBK482128/.
- Bianco, S.D.C.; Peng, J.-B.; Takanaga, H.; Suzuki, Y.; Crescenzi, A.; Kos, C.H.; Zhuang, L.; Freeman, M.R.; Gouveia, C.H.A.; Wu, J.; Luo, H.; Mauro, T.; Brown, E.M.; Hediger, M.A. Marked disturbance of calcium homeostasis in mice with targeted disruption of the Trpv6 calcium channel gene. Journal of Bone and Mineral Research 2007, 22, 274–285, . [CrossRef]
- Hwang, C.-S.; Shemorry, A.; Varshavsky, A. Two proteolytic pathways regulate DNA repair by cotargeting the Mgt1 alkylguanine transferase. Proceedings of the National Academy of Sciences 2009, 106, 2142–2147. [CrossRef]
- Sugiyama, T.; Kikuchi, H.; Hiyama, S.; Nishizawa, K.; Kusuhara, S. Expression and localisation of calbindin D28k in all intestinal segments of the laying hen. British Poultry Science 2007, 48, 233–238. [CrossRef]
- Chang, W.; Tu, C.; Chen, T-H.; Bikle, D.; Shoback, D. The extracellular calcium-sensing receptor (CaSR) is a critical modulator of skeletal development. Science Signaling 2008, 1, 1-23. [CrossRef]
- Schoulten, N.A.; Teixeira, A.S.; Freitas, R.T.F.; Bertechini, A.G.; Conte, A.J.; Silva, H.O. Níveis de cálcio em rações de Frangos de Corte na Fase Inicial Suplementadas com Fitase. Revista Brasileira de Zootecnia 2003, 32, 1190-1197. https://www.scielo.br/j/rbz/a/3YD6bYvWGrv8n6mW5YpyQxM/?format=pdf&lang=pt.
- Gobesso, A.A.O.; Wajnsztejn, H.; Ribeiro, R.M.; Bastos, F.L.; Etchichury, M.; Araújo Júnior, A.M.C. Comparison between different sources of minerals in horses with nutritional secondary hyperparathyroidism. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 2021, 73, 73 – 81. [CrossRef]
- Silva, B.C.; Bilezikian, J.P. Parathyroid hormone: anabolic and catabolic actions on the skeleton. Current Opinion in Pharmacology 2015, 22, 41–50. [CrossRef]
- Moe, S.M. Disorders Involving Calcium, Phosphorus, and Magnesium. Primary Care: Clinics in Office Practice 2008, 35, 215-237. [CrossRef]
- McDowell, L.R. Calcium and Phosphorus. In: Minerals in Animal and Human Nutrition. 2nd Edition, Elsevier Science BV, Amsterdam, 2003; 144p.
- Pinheiro, S.R.F. Niveis De Fósforo, De Cálcio E De Cloreto De Sódio Para Aves De Linhagens De Crescimento Lento Criadas Em Sistema Semi-Confinado. Tese (Doutor em Zootecnia) Universidade Estadual Paulista – UNESP, “Júlio de Mesquita Filho”, Jaboticabal – São Paulo – Brasil. 2009. https://repositorio.unesp.br/server/api/core/bitstreams/d5d73a2d-e13f-472a-8597-0e30913775cb/content.
- Underwood, E. J., & Suttle, N.F. The mineral nutrition of livestock 3rd edition. 3rd edn. CAB International, Wallingford, UK. 1999. Available at: https://www.cabidigitallibrary.org/doi/book/10.1079/9780851991283.0000.
- Naot, D.; Musson, D.S.; Cornish, J. The Activity of Peptides of the Calcitonin Family in Bone. Physiological Reviews 2019, 99, 781-805. [CrossRef]
- Cross, H.S.; Peterlik, M. Calcium and inorganic phosphate transport in embryonic chick intestine: Triiodothyronine enhances the genomic action of 1,25-dihydroxycholecalciferol. Journal of Nutrition 1988, 118, 1529–1534. [CrossRef]
- Sugiyama, T.; Kusuhara, S. Avian calcium metabolismo and bone function. Asian-Australasian Journal of Animal Sciences 2001, 14, 82–90. https://hero.epa.gov/hero/index.cfm/reference/details/reference_id/7351228.
- Bronner, F. Mechanisms of intestinal calcium absorption. Journal of Cellular Biochemistry 2003, 88, 387–393. [CrossRef]
- Van Abel, M.; Hoenderop, J.G.J.; Bindels, R.J.M. The epithelial calcium channels TRPV5 and TRPV6: Regulation and implications for disease. Naunyn-Schmiedeberg’s Archives of Pharmacology 2005, 371, 295–306. [CrossRef]
- Hirnet, D.; Olausson, J.; Fecher-Trost, C.; Bödding, M.; Nastainczyk, W.; Wissenbach, U.; Flockerzi, V.; Freichel, M. The TRPV6 gene, cDNA and protein. Cell Calcium 2003, 33, 509–518. [CrossRef]
- Belkacemi, L.; Bédard, I.; Simoneau, L.; Lafond, J. Calcium channels, transporters and exchangers in placenta: A review. Cell Calcium 2005, 37, 1–8. [CrossRef]
- Hoenderop, J.G.J.; Nilius, B.; Bindels, R.J.M. Calcium Absorption Across Epithelia. Physiological Reviews 2005, 85, 373-422. [CrossRef]
- Kim, H.J.; Lee, G.S.; Ji, Y.K.; Choi, K.C.; Jeung, E.B. Differential expression of uterine calcium transporter 1 and plasma membrane Ca 2+ ATPase 1b during rat estrous cycle. American Journal of Physiology-Endocrinology and Metabolism 2006, 291, E234–E241. [CrossRef]
- Suzuki, Y.; Kovacs, C.S.; Takanaga, H.; Peng, J.B.; Landowski, C.P.; Hediger, M.A. Calcium channel TRPV6 is involved in murine maternal-fetal calcium transport. Journal of Bone and Mineral Research 2008, 23, 1249–1256. [CrossRef]
- Lee, G.S.; Jeung, E.B. Uterine TRPV6 expression during the estrous cycle and pregnancy in a mouse model. American Journal of Physiology - Endocrinology and Metabolism 2007, 293, 132–139. [CrossRef]
- Lambers, T.T.; Bindels, R.J.M.; Hoenderop, J.G.J. Coordinated control of renal Ca2+ handling. Kidney International 2006, 69, 650–654. [CrossRef]
- Burley, R.W.; Vadhera, D.V. The Avian Egg. Chemistry and Biology. John Wiley & Sons Co. New York. 1989; 472p.
- Yamamoto, K.R. Steroid receptor regulated transcription of specific genes and gene networks. Annual review of genetics 1985, 19, 209–252. [CrossRef]
- Ali, A.; Tan, H.Y.; Kaiko, G.E. Role of the Intestinal Epithelium and Its Interaction With the Microbiota in Food Allergy. Frontiers Immunology 2020, 11, 604054. [CrossRef]
- Tang, V.W.; Goodenough, D.A. Paracellular ion channel at the tight junction. Biophysical Journal 2003, 84, 1660 –1673. [CrossRef]
- Clemente-Suárez, V.J.; Martín-Rodríguez, A.; Redondo-Flórez, L.; Villanueva-Tobaldo, C.V.; Yáñez-Sepúlveda, R.; Tornero-Aguilera, J.F. Epithelial Transport in Disease: An Overview of Pathophysiology and Treatment. Cells 2023, 12, 2455. [CrossRef]
- Friedman, P.A.; Gesek, F.A. Cellular calcium transport in renal epithelia: measurement, mechanisms, and regulation. Physiological Reviews 1995, 75, 429 – 471. [CrossRef]
- Peng, J.B.; Suzuki, Y.; Gyimesi, G.; Hediger, M.A. TRPV5 and TRPV6 Calcium-Selective Channels. In: Kozak JA, Putney JW Jr., editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 13. Available at: https://www.ncbi.nlm.nih.gov/books/NBK531440/.
- Nijenhuis, T.; Hoenderop, J.G.J.; Van der Kemp, A.W.C.M.; Bindels, R.J.M. Localization and Regulation of the Epithelial Ca 2+ Channel TRPV6 in the Kidney. Journal of the American Society of Nephrology 2003, 14, 2731–2740. [CrossRef]
- Brown, A.J.; Krits, I.; Armbrecht, H.J. Effect of age, vitamin D, and calcium on the regulation of rat intestinal epithelial calcium channels. Archives of Biochemistry and Biophysics 2005, 437, 51–58. [CrossRef]
- Bar, A. Calcium transport in strongly calcifying laying birds: Mechanisms and regulation. Comparative Biochemistry and Physiology - A Molecular and Integrative Physiology 2009, 152, 447–469. [CrossRef]
- Bianco, S.D.; Peng, J.B.; Takanaga, H.; Kos, C.H.; Crescenzi, A.; Brown, E.M.; Hediger, M.A. Mice lacking the epithelial calcium channel CaT1 (TRPV6) show a deficiency in intestinal calcium absorption depite high plasma levels of 1,25-dihydroxy vitamin D. FASEB Journal 2004, 18, A706. https://eurekamag.com/research/035/305/035305455.php.
- Hurwitz, S.; Harrison, H.C.; Harrison, H.E. Effect of vitamin D3 on the in vitro transport of calcium by the chick intestine. The Journal of nutrition 1967, 91, 319–323. [CrossRef]
- Peng, J.B.; Chen, X.Z.; Berger, U.V.; Weremowicz, S.; Morton, C.C.; Vassilev, P.M.; Brown, E.M.; Hediger, M.A. Human calcium transport protein cat1. Biochemical and Biophysical Research Communications 2000, 278, 326–332. [CrossRef]
- Wilkens, M.R.; Kunert-Keil, C.; Brinkmeier, H.; Schröder, B. Expression of calcium channel TRPV6 in ovine epithelial tissue. The Veterinary Journal 2009, 182, 294–300. [CrossRef]
- Wasserman, R.H.; Taylor, A.N. Vitamin D3-induced calcium-binding protein in chick intestinal mucosa. Science 1966, 152, 794–796. [CrossRef]
- Qin, X.; Klandorf, H.; Porter, D.W.; Holt, S.B.; Martin, W.G. Estrogen Enhancement of Ca-, Mg-, and Ca-Mg-Stimulated Adenosine Triphosphatase Activity in the Chick Shell Gland. General and Comparative Endocrinology 1993a, 89, 4–10. [CrossRef]
- Qin, X.; Klandorf, H. Effect of Estrogen in Relation to Dietary Vitamin D3 and Calcium on Activity of Intestinal Alkaline Phosphatase and Ca-ATPase in Immature Chicks. General and Comparative Endocrinology 1993b, 90, 318–327. [CrossRef]
- Morrissey, R.; Wasserman, R. Calcium absorption and calcium-binding protein in chicks on differing calcium and phosphorus intakes. American Journal of Physiology-Legacy Content 1971, 220, 1509–1515. [CrossRef]
- Wu, J.C.Y.; Smith, M.W.; Mitchell, M.A.; Peacock, M.A.; Turvey, A.; Keable, S.J. Enterocyte expression of calbindin, calbindin mRNA and calcium transport increases in jejunal tissue during onset of egg production in the fowl (Gallus domesticus). Comparative Biochemistry and Physiology -- Part A: Physiology 1993, 106, 263–269. [CrossRef]
- Cai, Q.; Chandler, J.S.; Wasserman, R.H.; Kumar, R.; Penniston, J.T. Vitamin D and adaptation to dietary calcium and phosphate deficiencies increase intestinal plasma membrane calcium pump gene expression. Proceedings of the National Academy of Sciences of the United States of America 1993, 90, 1345–1349. [CrossRef]
- McCormick, C.C. Passive diffusion does not play a major role in the absorption of dietary calcium in normal adults. Journal of Nutrition 2002, 132, 3428–3430. [CrossRef]
- Fleet, J.C.; Schoch, R.D. Molecular mechanisms for regulation of intestinal calcium absorption by vitamin D and other factors. Critical Reviews in Clinical Laboratory Sciences 2010, 47, 181–195. [CrossRef]
- Corradino, R.A.; Wasserman, R.H.; Pubols, M.H.; Chang, S.I. Vitamin D3 induction of a calcium-binding protein in the uterus of the laying hen. Archives of Biochemistry and Biophysics 1968, 125, 378–380. [CrossRef]
- Wasserman, R.H.; Smith, C.A.; Smith, C.M.; Brindak, M.E.; Fullmer, C.S.; Krook, L.; Penniston, J.T.; Kumar, R. Immunohistochemical localization of a calcium pump and calbindin-D28k in the oviduct of the laying hen. Histochemistry 1991, 96, 413-8. [CrossRef]
- Bar, A.; Cohen, A.; Eisner, U.; Risenfeld, G.; Hurwitz, S. Differential Response of Calcium Transport Systems in Laying Hens to Exogenous and Endogenous Changes in Vitamin D Status. The Journal of Nutrition 1978, 108, 1322–1328. [CrossRef]
- Nys, Y.; Parkes, C.O.; Thomasset, M. Effects of suppression and resumption of shell formation and parathyroid hormone on uterine calcium-binding protein, carbonic anhydrase activity, and intestinal calcium absorption in hens. General and Comparative Endocrinology 1986, 64, 293–299. [CrossRef]
- Corradino, R.A.; Smith, C.A.; Krook, L.P.; Fullmer, C.S. Tissue-specific regulation of shell gland calbindin D28K biosynthesis by estradiol in precociously matured, vitamin D-depleted chicks. Endocrinology 1993, 132, 193–198. [CrossRef]
Figure 1.
Myo-inositol (a) and myo-inositol 1, 2, 3, 4, 5, 6-hexakis dihydrogen phospodium (InsP6) (b).
Figure 1.
Myo-inositol (a) and myo-inositol 1, 2, 3, 4, 5, 6-hexakis dihydrogen phospodium (InsP6) (b).
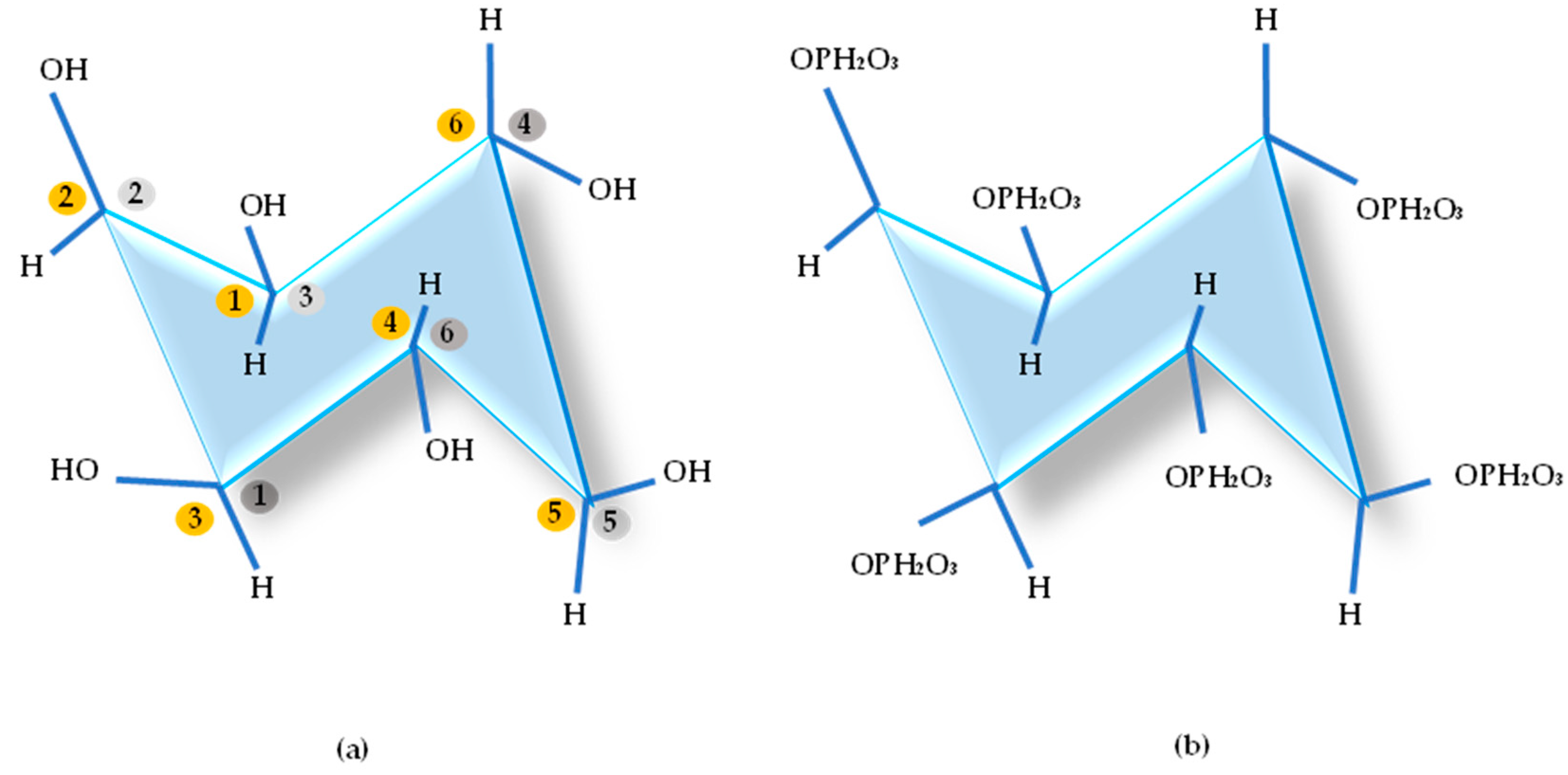
Figure 2.
Hydrolysis of the phytate molecule through the action of the phytase enzyme.
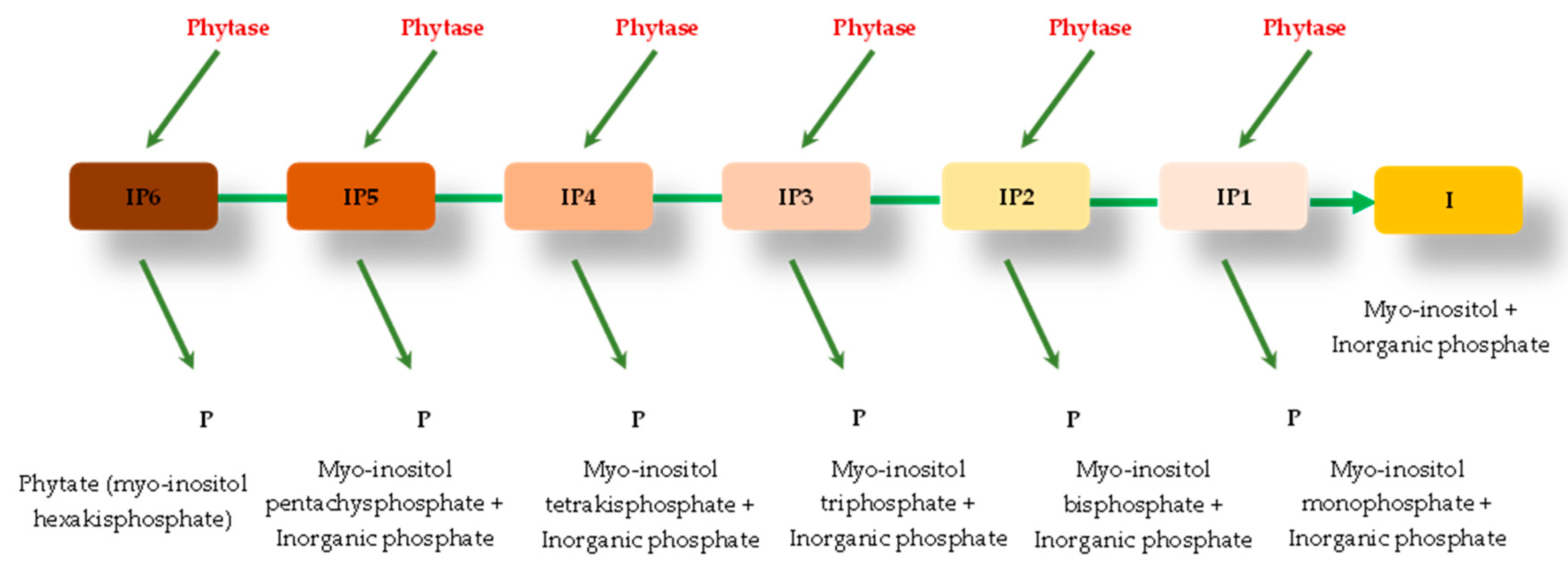
Figure 3.
Classification of phytases.
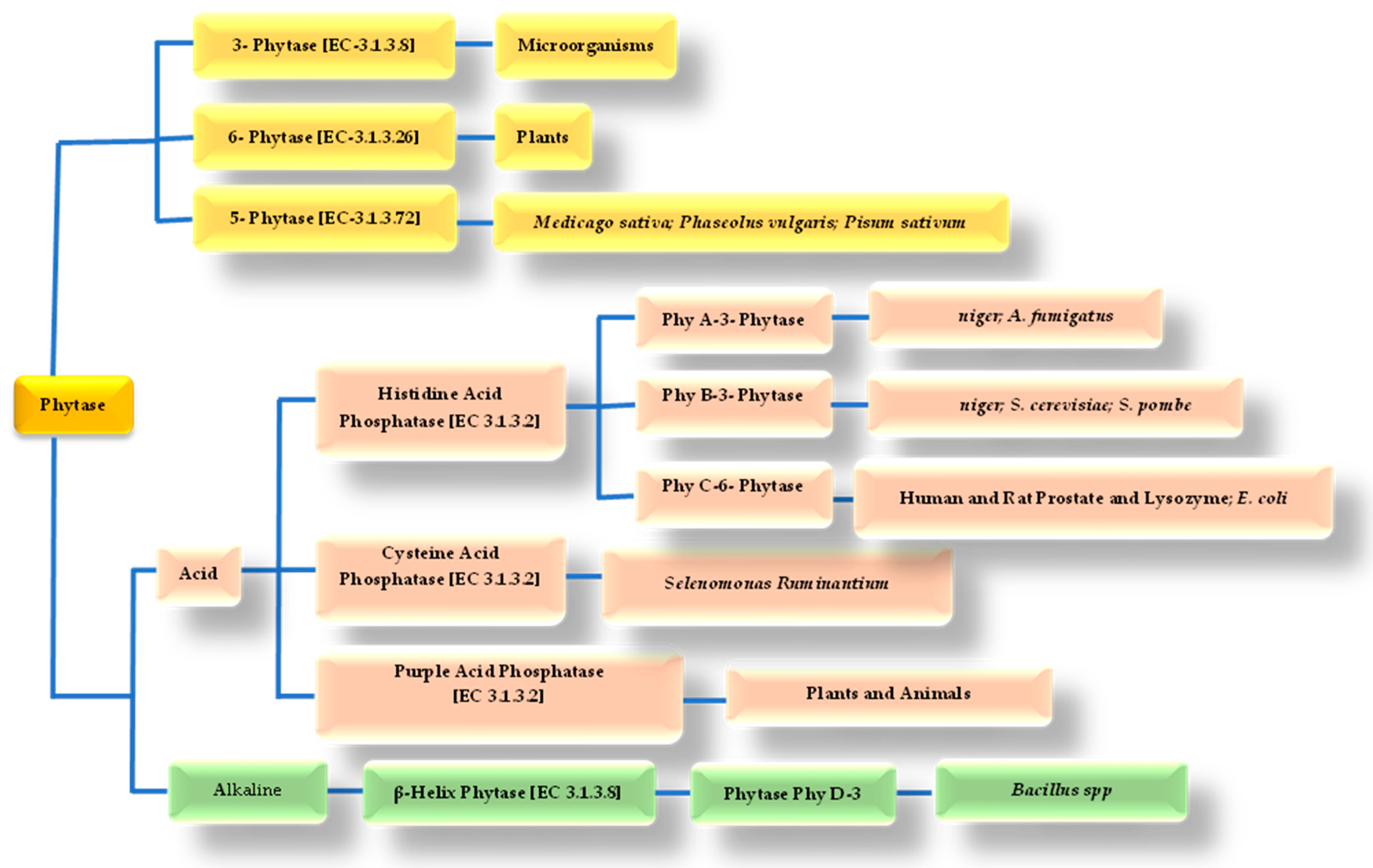
Figure 4.
Schematic drawing of transepithelial calcium transport.
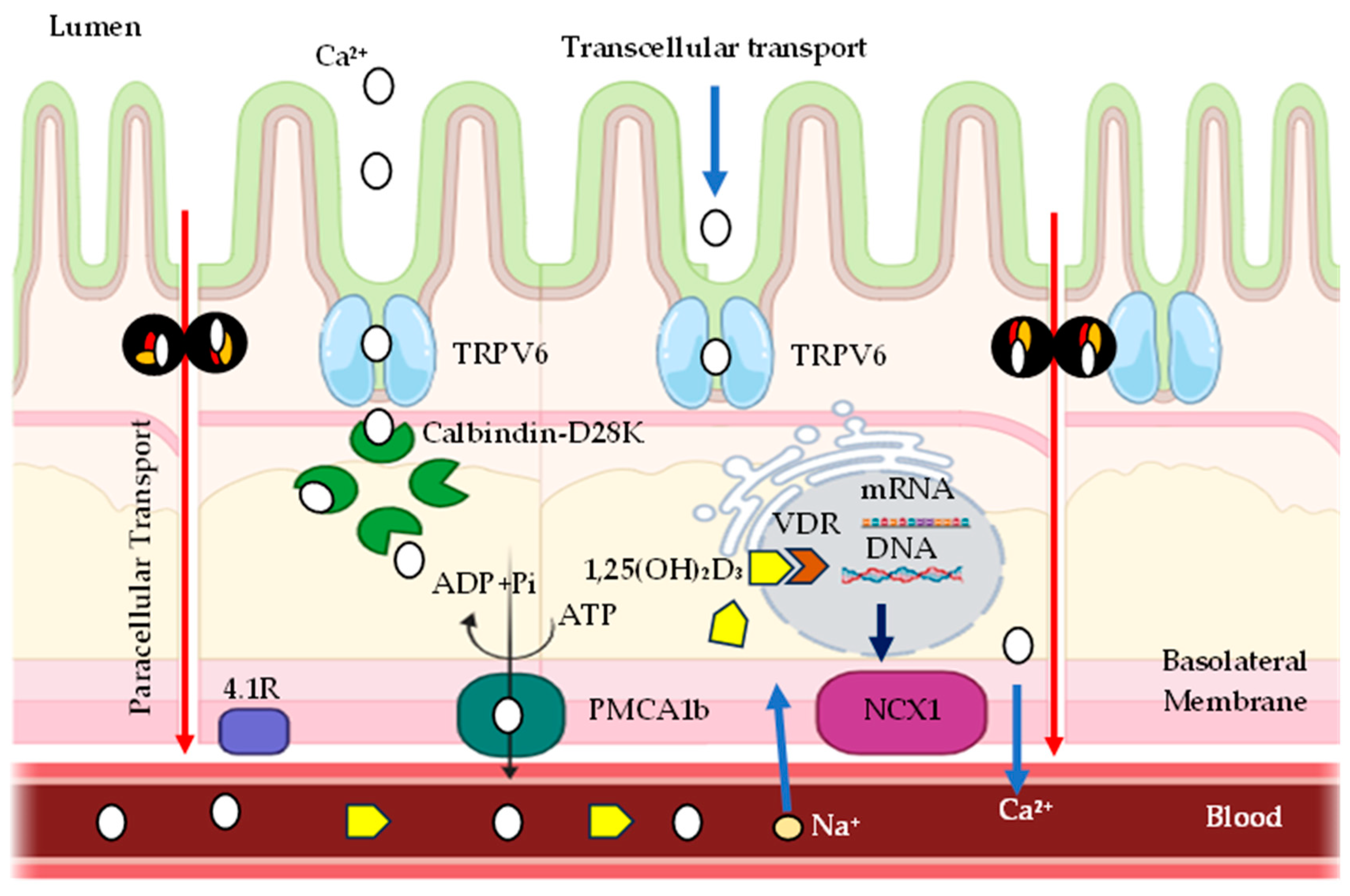
Figure 5.
Calcium Selective Channel TRPV6 and Calbindin-D28K.
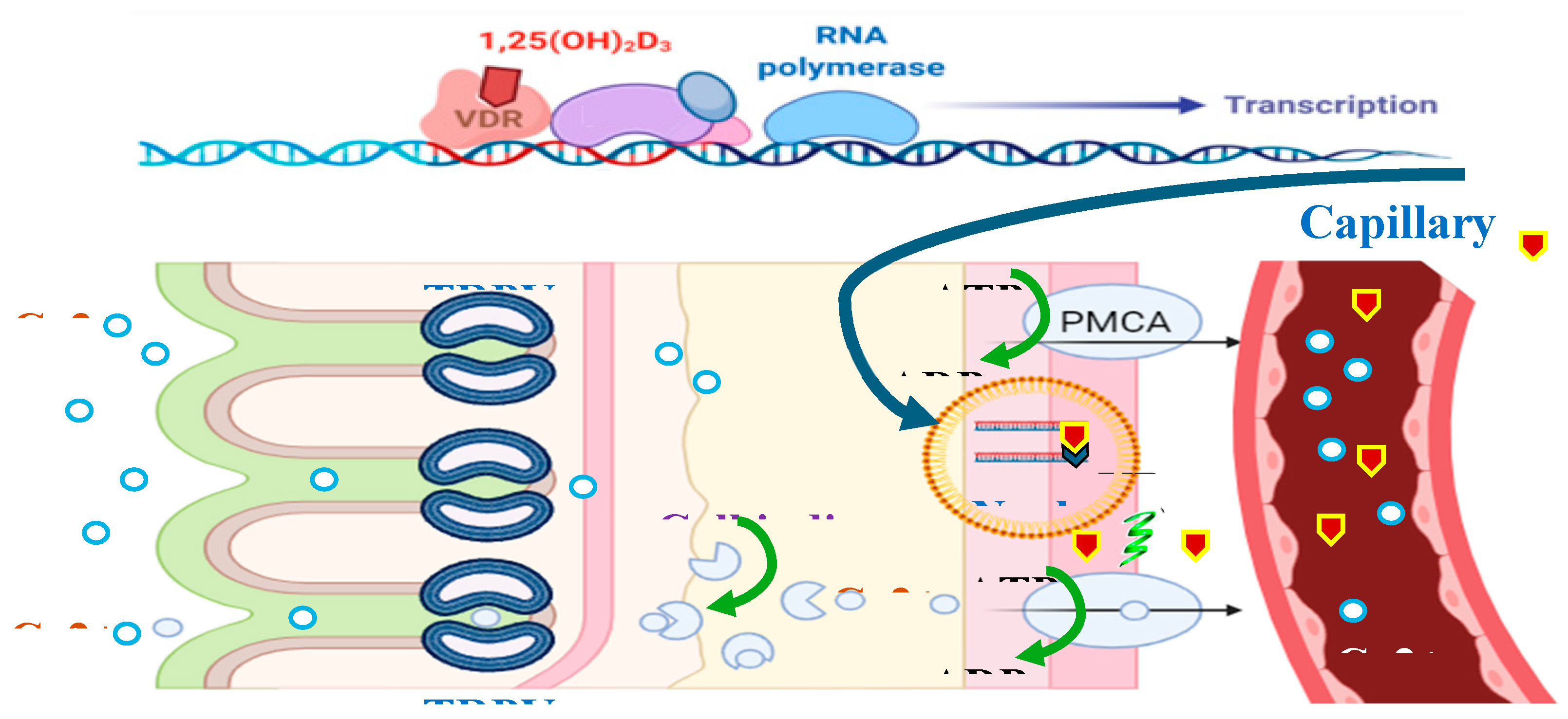

Table 1.
Phytate content in foods (based on dry matter).
| Products | % Phytate | Reference |
| Corn | 0.78–1.05 | [59,60] |
| Soybean | 1.01–1.47 | [60,61] |
| Sorghum | 0.80 | [62] |
| Cottonseed meal | 2.65 | [62] |
| Corn germ | 2.97 | [62] |
| Polished rice | 0.60 | [60,63] |
| Oats | 0.79–1.01 | [60,61] |
| Wheat | 0.39–1.35 | [60,64] |
| Soybean bran | 1.0-1.5 | [65] |
| Rice bran | 5.90–6.48 | [66,67] |
| Wheat bran | 5.38 | [59,60] |
| Whole wheat flour | 2.22 | [60,68] |
| White wheat flour | 0.404 | [60,68] |
Table 2.
Recommendations for available calcium (Ca) and phosphorus (P) for Japanese quails.
| Age - weeks | Ca (%) | P (%) | Literature |
| 6-16 | 3.50 | 0.45 | [137] |
| 6-29 | 2.00-3.05 | - | [138] |
| 6-19 | 3.51 | - | [139] |
| 8-21 | 2.50 | 0.31 | [140] |
| 45-57 | 3.50 | 0.15 | [141] |
| 45-57 | 3.80 | - | [142] |
| 7-54 | 2.50 | 0.25 | [135] |
| 12-42 | 2.50 | 0.35 | [143] |
| 26-38 | 2.00 | 0.31 | [144] |
| 21-36 | 3.10 | 0.32 | [145] |
| 7-20 | 2.50 | 0.35 | [146] |
| 8-56 | 2.99 | 0.31 | [40] |
| 6-57 | 2.0-3.8 | 0.15-0.45 | [120] |
| 20-32 | - | 0.39-0.44 | [147] |
| 9-24 | 2.68 | 0.38 | [119] |
| 8-56 | 3.01-3.04 | 0.31-0.80 | [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



