
Submitted:
16 September 2024
Posted:
20 September 2024
You are already at the latest version
Abstract
A polyphasic taxonomic study was carried out on the rod-shaped, orange pigmented strain C11T, isolated from gold mine tailings. Sequencing of the 16S rRNA gene showed relatedness to Brevundimonas, with 98.4 % and 98.2 % similarity to Brevundimonas bacteroides and Brevundimonas variabilis, respectively. Average nucleotide identity and digital DNA–DNA hybridization with the closest phylogenetic neighbor of C11T indicates distinction at the species level, further confirmed by differences in physiology. C18:1 ω7c is the dominant cellular fatty acid. DNA G + C content is 68.3 mol %. The predominant ubiquinone is Q-10; 1,2-Di-O-acyl-3-O-α-D-glucopyranuronosyl glycerol, phosphatidylglycerol, 1,2-di-O-acyl-3-O-α-D-glucopyranosyl glycerol and 1,2-di-O-acyl-3-O-[D-glucopyranosyl-(1->4)-α-D-glucopyranuronosyl] glycerol are the major polar lipid constituents. This bacterium produces bacteriochlorophyll a and tolerates high concentrations of (μg/mL): tellurium (>1500), selenium (1000 to >5000), and vanadium (>5000) oxyanions. Data supports the inclusion of C11T into the genus Brevundimonas as a new species with the proposed name Brevundimonas aurifodinae sp. nov., (C11T = NRRL B-61758T; = DSM 118059T).
Keywords:
Aerbic anoxygenic phototroph
; Brevundimonas
; Brevundimonas aurifodinae
; Bacteriochlorophyll a
; Mine tailings
; Metalloid oxide resistance
; Tellurium
; Selenium
; Vanadium
1. Introduction
Brevundimonas is a genus, belonging to the family Caulobacteraceae, that was proposed in 1994 to re-classify two species, which had a distinct taxonomic position among the Pseudomonas [1]. It is also closely related to Caulobacter, with several current Brevundimonas members formerly residing in the genus [2]. Currently, there are 37 validly recognized members with B. diminuta as the type species [3]. Members are gram-negative, rod shaped, aerobic, oligotrophic, contain Q-10 as the major isoprenoid quinone and have relatively high DNA G+C content [4]. However, they vary in traits such as motility, pigmentation, and prosthecae formation [4]. Species have been isolated from a wide range of habitats: soils, marine/freshwater environments, sand, and activated sludge [5,6]. Brevundimonas spp. are becoming of greater interest as human pathogens, with B. diminuta and B. vesicularis being found in clinical specimens of patients with underlying conditions [6]. One interesting feature is some members produce bacteriochlorophyll a (Bchl a), indicating they can be classified as aerobic anoxygenic phototrophs (AAP) [7]. Currently, three species are known to have this ability: B. bacteroides [7], B. subvibrioides [8], and B. variabilis [9]. AAP are a diverse group found in many different environments and various genera of the Proteobacteria [10]. Their core characteristics include requiring oxygen for growth and survival, being incapable of autotrophy and utilizing energy derived from photosynthesis as a supplemental source to cellular respiration [11]. A standout characteristic of AAP is their ability to withstand extremely high levels of metalloid oxyanions [10]. Resistance to toxic metals is a known attribute of many Brevundimonas spp. For example, B. vancanneytii SMA3 mitigates heavy metals cadmium, lead, and mercury from soils [12], B. diminuta can survive increased levels of arsenic, cadmium, and zinc [13,14], while B. vesicularis remediates copper, lead, and nickel [15,16]. Previously, the tailings of Central Gold Mine in Nopiming Provincial Park, Manitoba, Canada were investigated to explore the diversity of AAP in this extreme environment and their resistance to metalloid oxides [17]. From here, strain C11T, was isolated and found to produce Bchl a, resist high levels of Te, Se, and V oxyanions as well as have close relations to Brevundimonas [17]. Since the genus has potential for bioremediation, the discovery of more species which tolerate and remove toxic metals could lead to future applications. As such, we set forth to taxonomically classify C11T as a new species with the proposed name Brevundimonas aurifodinae.
2. Materials and Methods
2.1. Strains and Cultivation
Strain C11T was isolated from gold mine tailings at Nopiming Provincial Park, Manitoba, Canada using Rich Organic (RO) solid medium [17]. Cells in all experiments were grown aerobically on Caulobacter medium [18] at 30 °C and pH 7.0 in the dark unless otherwise noted. Long term storage was at -80 °C in 30 % glycerol and modified RO with 10% (w/v) organics. The following strains were obtained for comparison: From the Korean Agricultural Culture Collection: Brevundimonas alba KACC 12015T [2], Brevundimonas bacteroides KACC 12013T [2], Brevundimonas basaltis KACC 17487T [19], Brevundimonas subvibrioides KACC 12014T [2], and Brevundimonas variabilis KACC 12016T [2]; From the USDA-ARS Culture Collection (NRRL): Brevundimonas diminuta NRRL B-1496T [1].
2.2. Physiological and Biochemical Experiments
Taxonomical markers such as the production of specific enzymes and utilization of carbon sources was investigated using API ZYM and API 20NE test strips (Biomeriux) and Biolog GEN III Microplate (Biolog Inc.). Motility was determined by the hanging drop method [20]. In addition to the antibiotic susceptibility tested with the Biolog GEN III Microplate, disk diffusion assays were carried out using BD BBL™ Sensi-Disc™ [21] with the following: penicillin G (10 IU), ampicillin (10 µg), polymyxin B (300 IU), tetracycline (30 µg), erythromycin (15 µg), imipenem (10 µg), streptomycin (10 µg), chloramphenicol (30 µg), bacitracin (2 IU), kanamycin (30 µg), and rifampin (5 µg). Temperature range for growth was evaluated from 0 to 50 °C at 5 °C intervals while pH tolerance was assessed from 4.0 to 11.0 with 0.5 increments and NaCl from 0.0 % to 6.0 % at 0.5 % intervals. Gram stain, spore formation, anaerobic growth, catalase, oxidase, methyl red, Voges-Proskauer, indole and other carbon source utilization were completed as described [22].
Anoxygenic photosynthetic complex formation and Bchl a synthesis was assessed in absorption spectra taken of whole cells and pigment extracts using standard methods [11]. Resistance and reduction of metal(loid) oxides was evaluated previously [17].
For fatty acid profiling, C11T was grown in Caulobacter medium at 30 °C for 72 h. Biomass was collected, lipids were extracted with the Folch method [23] and analyzed via gas chromatography [24]. Polar lipids were discerned using two-dimensional TLC with the appropriate detection reagents [25]. Cellular quinones obtained from 100 mg of freeze-dried cells were separated by TLC and identified [25].
2.3. Microscopy
The cell size and shape of a 48 h culture was observed with a phase contrast light microscope (Zeiss Axioskop 2).
2.4. Phylogenetic Analysis
DNA was extracted as in protocol [26] and sent to Azenta (South Plainfield, NJ, USA) for 16S rRNA gene Sanger sequencing using universal primers 27F (5’-AGAGTTTGATCCTGGCTCAG-3’) and 1492R (5’-GGTTACCTTGTTACGACTT-3’). A 1404 bp fragment (GenBank accession number: PP885399) was produced. Closest relatives were identified with a NCBI standard nucleotide BLAST search. A Maximum Likelihood phylogenetic tree was created in MEGA 11 and pairwise aligned 16S rRNA fragments of other Brevundimomas spp. collected from NCBI GenBank [27]. Genome sequence of C11T was obtained as done previously [28]. Briefly, the sequencing library of the genomic DNA was prepared using the Illumina DNA Library Prep kit. The genome was sequenced with the Illumina MiniSeq platform using 500 μl of a 1.8pM library. Paired-end (2x150 bp) sequencing generated 2,690,380 reads and 406.3 Mbps. Quality control of the reads was performed using FASTQC (v1.0.0), using a k-mer size of 5 and contamination filtering for overrepresented sequences against the default contamination list. Genome assembly with the Illumina sequencing was performed using Unicycler (v0.5.0) [29] within BV-BRC [30]. This resulted in a 3.3 Mbp genome consisting of 26 contigs (116x coverage). It was then annotated using the NCBI prokaryotic genome annotation pipeline [31]. The Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank under the accession JBEGDD000000000. The version described in this paper is version JBEGDD010000000. Average nucleotide identity (ANI) was assessed with the OrthoANI algorithm through ChunLab’s online ANI calculator [32]. Formula d4 of the Genome-Genome Distance Calculator from DSMZ [33] was used to define digital DNA:DNA hybridization (dDDH) values. A Genome Blast Distance Phylogeny tree was created with the Type Strain Genome Server (TYGS) from DSMZ [34]. Intergenomic distances were applied to generate a balanced minimum evolution tree via FASTME 2.1.6.1 including SPR post-processing [35]. Branch support was inferred from 100 pseudo-bootstrap replicates each. The genome-based tree was rooted at the midpoint [36] and visualized with PhyD3 [37].
3. Results and Discussion
3.1. Physiology and Morphology of Cells
API ZYM, API 20NE test strips and Biolog Gen III Microplate revealed C11T used the following carbon sources for growth: gluconate, dextrin, D-cellobiose, gentibiose, sucrose, D-turanose, stachyose, α-D-lactose, D-melibiose, N-acetyl-D-glucosanmine, N-acetyl-beta-D-mannosamine, N-acetyl-D-galactosamine, N-acetyl-nuraminic acid, α-D-glucose, D-mannose, D-galactose, 3-methyl-glucose, D-fucose, L-fucose, L-rhamnose, D-mannitol, D-arabitol, myo-inositol, D-glucose-6-PO4, D-fructose-6-PO4, D-aspartic acid, Glycyl-L-proline, L-alanine, L- aspartic acid, L-glutamic acid, L-serine, D-galacuronic acid, L-galacturonic acid, D-gluconic acid, D-glucuronic acid, glucuronamide, quinic acid, D-saccharic acid, D-lactic acid methyl ester, citric acid, α-keto-glutaric acid, D-malic acid, L-malic acid, Bromo-succinate acid, α-hydroxy-butyric acid, β-hydroxy-D, L-butyric acid, α-keto-butyric acid, acetoacetic acid, acetic acid, casamino acids, yeast extract, and bactopeptone. Alkaline phosphatase, esterase (C4), esterase lipase (C8), leucine arylamidase, valine arylamidase, trypsin, α-chymotrypsin, acid phosphatase, naphthol-AS-Bi-phosphohydrolase, and α-glucosidase activity is present. Esculin and gelatin are hydrolyzed. Carbon sources and enzyme activities not cited here which are included in these test kits, produced negative results. C11T is susceptible to chloramphenicol, kanamycin, polymyxin B, streptomycin, imipenem, vancomycin, tetrazolium violet, tetrazolium blue, aztreonam, macrolide, rifamycin, minocycline, lincomycin, guanidine HCL, niaproof 4, fusidic acid, D-serine, and sodium bromate, but resistant to nalidixic acid, lithium chloride, penicillin, and ampicillin. C11T does not require vitamin supplements. Growth occurs between 5 and 40 °C, from pH 6.0 to 10.5 and up to 2.0 % NaCl with optimums at 30 °C, pH 8.0 and 0 %, respectively (Table 1).
C11T was assessed for its ability to resist and potentially reduce metal(loid) oxyanions [17]. It had the broadest and greatest tolerance among other isolates from the gold mine tailings in Nopiming Provincial Park. This included surviving high levels of (µg/ml): tellurite (>1500), tellurate (>1500), selenite (1000), selenate (>5000), metavanadate (>5000), and orthovanadate (>5000). Furthermore, C11T has potential bioremediation and biometallurgy applications as it reduced tellurite to elemental tellurium [17].
In vivo, C11T has an anoxygenic photosynthesis complex containing a light-harvesting I complex (870 nm) and reaction center (802 nm) (Figure 1A, light orange line). Bchl a is detected in the pigment extract absorbance spectrum (770 nm, Figure 1A, dashed dark orange line). As such, C11T is classified as an aerobic anoxygenic phototroph. Carotenoids are also synthesized (Figure 1A orange line: 458, 515 nm; Figure 1A, dashed dark orange line: 423, 453, 481, 531 nm).
Strain C11T is gram-negative, oxidase- and catalase-positive, non-spore forming and obligately aerobic. It produces circular (1-2 mm), raised, orange colonies with entire margins and a mucoid consistency on Caulobacter media plates after 72 h. Morphologically, cells are rod shaped, 1.5-2.0 µm in length and 0.75-1.0 µm in width and do not form prosthecae after 48 h of growth (Figure 1B).
3.2. Chemotaxonomic Characterization
Whole cell fatty acid analysis revealed C11T contains predominately C18:1 ω7c. The major polar lipids are 1,2-Di-O-acyl-3-O-α-D-glucopyranuronosyl glycerol (MGDOx), phosphatidylglycerol (PG), 1,2-di-O-acyl-3-O-α-D-glucopyranosyl glycerol (MGD) and 1,2-di-O-acyl-3-O-[D-glucopyranosyl-(1→4)-α-D-glucopyranuronosyl]glycerol (DGL). MGDOx production differentiates Brevundimonas from Caulobacter and as such, its presence in C11T supports the conclusions [4]. The predominant ubiquinone in the cells is Q-10. Fatty acid, polar lipid and quinone profiles are representative of Brevundimonas spp. [4], solidifying its genus placement.
3.3. Phylogenetic and Genomic Analysis
A nearly complete 16S rRNA fragment (1404 bp) of C11T was produced via Sanger sequencing with the full gene (1461 bp) identified in the genome on contig 7. Pairwise comparisons to type species revealed 16S rRNA relatedness of strain C11T was 98.4% to B. bacteroides KACC 12013T [2] and 98.2% to B. variabilis KACC 12016T [2]. Using these sequences and those from validated Brevundimonas members, a Maximum-Likelihood 16S rRNA gene phylogenetic tree was created (Figure 2).
The final product positions C11T within the genus (Figure 2).
The genome of C11T is 3.3 Mb and has a G + C content of 68.3 mol %, falling within range of other genus members (Table 2) [4]. It contains 26 contigs an L50 of 5, and 3186 protein-coding genes. OrthoANI between C11T and B. bacteroides is 83.5 % and dDDH is 26.6 %. For B. variabilis it is 77.7 % and 21.8 %, respectively. All fall below the accepted cutoffs for species differentiation [38]. The Genome Blast Distance Phylogeny tree created shows a distinct lineage for C11T among other Brevundimonas spp. that supports its classification as a new species (Figure 3).
While cells were non-motile under test conditions, C11T possesses the genes for flagellar production. As shown via spectrophotometry, C11T synthesizes Bchl a and as such, has bhcB, bchC, bchF, bchL, bchM, bchY, and bchZ sequences within its genome. Besides being resistant to very high levels of tellurite, tellurate, selenite, selenate, metavanadate, and orthovanadate [17], copper and silver resistance via a resistance-nodulation-division efflux transporter is also encoded, suggesting C11T may be able to tolerate high levels of Cu and Ag. These genes alongside those encoding for conjugative transfer, polygalacturonase as well as additional hypothetical proteins, comprise the 404 PATRIC cross-genus families found in C11T but not in closest relatives B. bacteroides, B. subvibrioides and B. variabilis.
4. Conclusion
Strain C11T, isolated from gold mine tailings in Nopiming Provincial Park, possesses significant differences in both phenotype (carbon metabolism and enzyme activities) and genotype (ANI and dDDH), which sufficiently differentiate it from the closest relatives. As such, this bacterium represents a new species in the genus and the name Brevundimonas aurifodinae is proposed.
4.1. Description of Brevundimonas aurifodinae sp. nov.
Brevundimonas aurifodinae (au.ri.fo.di’nae. L. neut. n. aurum, gold; L. fem. n. fodina, mine; N.L. gen. n. aurifodinae, indicates discovery at a gold mine).
Gram-negative, non-motile, non-spore forming and obligately aerobic. Circular (1-2 mm), raised, orange colonies with entire margins and a mucoid consistency form on Caulobacter medium plates after 72 h. Cells are rod-shaped, 1.5-2.0 µm in length and 0.8-1.0 µm in width, non-prosthecate. Catalase- and oxidase-positive. Growth occurs in the following conditions (optimum): between 5 and 40 °C (30 °C), from pH 6.0 to 10.5 (8.0) and up to 2.0 % NaCl (0 %). Carbon sources utilized include: Gluconate, dextrin, D-cellobiose, gentibiose, sucrose, D-turanose, stachyose, α-D-lactose, D-melibiose, N-acetyl-D-glucosanmine, N-acetyl-beta-D-mannosamine, N-acetyl-D-galactosamine, N-acetyl-nuraminic acid, α-D-glucose, D-mannose, D-galactose, 3-methyl-glucose, D-fucose, L-fucose, L-rhamnose, D-mannitol, D-arabitol, myo-inositol, D-glucose-6-PO4, D-fructose-6-PO4, D-aspartic acid, Glycyl-L-proline, L-alanine, L- aspartic acid, L-glutamic acid, L-serine, D-galacuronic acid, L-galacturonic acid, D-gluconic acid, D-glucuronic acid, glucuronamide, quinic acid, D-saccharic acid, D-lactic acid methyl ester, citric acid, α-keto-glutaric acid, D-malic acid, L-malic acid, Bromo-succinate acid, α-hydroxy-butyric acid, β-hydroxy-D, L-butyric acid, α-keto-butyric acid, acetoacetic acid, acetic acid, casamino acids, yeast extract, and bactopeptone. Alternatively, capric acid, adipic acid, phenylacetic acid, D-maltose, D-trehalose, D-raffinose, β-methyl-D-glucoside, D-salicin, D-fructose, inosine, D-sorbitol, glycerol, D-serine, L-arginine, L-histidine, L-pyroglutamic acid, pectin, mucic acid, p-hydroxy-phenylacetic acid, methyl pyruvate, L-lactic acid, ɣ-amino-butyric acid, propionic acid, ethanol, methanol, and formic acid were not. Can grow without vitamins supplemented. Alkaline phosphatase, esterase (C4), esterase lipase (C8), leucine arylamidase, valine arylamidase, trypsin, α-chymotrypsin, acid phosphatase, naphthol-AS-Bi-phosphohydrolase, α-glucosidase, and amylase activity were present, while arginine dihydrolase, lipase, cystine arylamidase, α-galactosidase, β-galactosidase, β-glucuronidase, β-glucosidase, N-acetyl-β-glucosaminidase, α-mannosidase, α-fucosidase, urease, and nitrate reductase were not. Esculin and gelatin were hydrolyzed. Indole, methyl red, and Voges-Proskauer were negative. The primary fatty acid was C18:1 ω7c. Ubiquinone Q-10 was the dominant isoprenoid quinone. MGDOx, PG, MGD, and DGL were the major polar lipids. Produces bacteriochlorophyll a and is resistant to high levels of (µg/ml): tellurite (>1500), tellurate (>1500), selenite (1000), selenate (>5000), metavanadate (>5000), and orthovanadate (>5000). Can reduce tellurite to elemental tellurium. The DNA G + C content is 68.3 mol %.
The type strain C11T (= NRRL B-61758T = DSM 118059T) was isolated from gold mine tailings at Nopiming Provincial Park, Manitoba, Canada. Strain C11T ribosomal 16S rRNA gene sequence under GenBank accession number: PP885399. This Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank under the accession JBEGDD000000000, which is used in this work.
Author Contributions
Conceptualization, Chris Maltman and Vladimir Yurkov; Formal analysis, Chris Maltman, Katia Messner, John Kyndt and Vladimir Yurkov; Funding acquisition, Chris Maltman, John Kyndt and Vladimir Yurkov; Investigation, Chris Maltman, Katia Messner, John Kyndt and Vladimir Yurkov; Methodology, Chris Maltman, Katia Messner, John Kyndt and Vladimir Yurkov; Project administration, Chris Maltman and Vladimir Yurkov; Resources, Chris Maltman, John Kyndt and Vladimir Yurkov; Supervision, John Kyndt and Vladimir Yurkov; Writing – original draft, Chris Maltman; Writing – review & editing, Chris Maltman, Katia Messner, John Kyndt and Vladimir Yurkov. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Department of Biology, Slippery Rock University, held by C. Maltman; Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant 1501 and a University of Manitoba GETS grant, both held by V. Yurkov; and the Wilson Enhancement Fund for Applied Research in Science at Bellevue University, given to J. Kyndt.
Data Availability Statement
Strain C11T ribosomal 16S rRNA gene sequence under GenBank accession number: PP885399. This Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank under the accession JBEGDD000000000, which is used in this work.
Acknowledgments
We would like to thank Dr. George Lengyel for assistance with quinone and polar lipid analysis as well as Dr. Aharon Oren for his assistance with checking Latin grammar and spelling of the new name.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Segers, P.; Vancanneyt, M.; Pot, B.; Torck, U.; Hoste, B.; Dewettinck, D.; Falsen, E.; Kersters, K.; De Vos, P. Classification of Pseudomonas diminuta Leifson and Hugh 1954 and Pseudomonas vesicularis Büsing, Döll, and Freytag 1953 in Brevundimonas gen. nov. as Brevundimonas diminuta comb. Nov. and Brevundimonas vesicularis comb. Nov., Respectively. Int. J. Syst. Evol. Microbiol. 1994, 44, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Abraham, W.-R.; Strompl, C.; Meyer, H.; Lindholst, S.; Moore, E.; Christ, R.; Vancanneyt, M.; Tindall, B.; Bennasar, A.; Smit, J.; Tesar, M. Phylogeny and Polyphasic Taxonomy of Caulobacter species. Proposal of Maricaulis gen. nov. with Maricaulis maris (Poindexter) comb. nov. as the Type Species, and Emended Description of the Genera Brevundimonas and Caulobacter. Int. J. Syst. Evol. Microbiol. 1999, 49, 1053–1073. [Google Scholar] [CrossRef] [PubMed]
- Parte, A.; Sardà Carbasse, J.; Meier-Kolthoff, J.; Reimer, L.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Bacteriol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Vancanneyt, M.; Segers, P.; Abraham, W.-R.; Vos, P. Brevundimonas. In Bergey’s Manual of Systematics of Archaea and Bacteria, 2nd ed.; Trujillo, M.E., Dedysh, S., DeVos, P., Hedlund, B., Kämpfer, P., Rainey, F.A., Whitman, W.B., Eds.; Springer Science + Business Media, Inc.: New York, NY, USA, 2005; Volume 2, pp. 308–315. [Google Scholar]
- Friedrich, I.; Klassen, A.; Neubauer, H.; Schneider, D.; Hertel, R.; Daniel, R. Living in a Puddle of Mud: Isolation and Characterization of Two Novel Caulobacteraceae Strains Brevundimonas ondensis sp. nov. and Brevundimonas gottingensis sp. nov. Appl. Microbiol. 2021, 1, 38–59. [Google Scholar] [CrossRef]
- Ryan, M.; Pembroke, J. Brevundimonas spp: Emerging Global Opportunistic Pathogens. Virulence 2018, 9, 480–493. [Google Scholar] [CrossRef]
- Tanabe, Y.; Yamaguchi, H.; Yoshida, M.; Kai, A.; Okazaki, Y. Characterization of a Bloom-Associated alphaproteobacterial Lineage, ‘Candidatus Phycosocius’: Insights into Freshwater Algal-Bacterial Interaction. ISME Comm. 2023, 3, 20. [Google Scholar] [CrossRef]
- Imhoff, J.; Than, T.; Kunzel, S.; Neulinger, S. Phylogeny of Anoxygenic Photosynthesis Based on Sequences of Photosynthetic Reaction Center Proteins and a Key Enzyme in Bacteriochlorophyll Biosynthesis, the Chlorophyllide Reductase. Microorganisms 2019, 7, 576. [Google Scholar] [CrossRef]
- Tahon, G.; Willems, A. Isolation and Characterization of Aerobic Anoxygenic Phototrophs from Exposed Soils for the Sor Rondane Mountains, East Antarctica. Syst. Appl. Microbiol. 2017, 40, 357–369. [Google Scholar] [CrossRef]
- Hughes, E.; Yurkov, V. Aerobic Anoxygenic Phototrophs: Four Decades of Mystery. In Modern Topics in the Phototrophic Prokaryotes: Environmental and Applied Aspects; Hallenbeck, P., Ed.; Springer: Cham, Switzerland, 2017; pp. 193–214. [Google Scholar]
- Yurkov, V.; Beatty, T. Aerobic Anoxygenic Phototrophic Bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 695–724. [Google Scholar] [CrossRef]
- Ghosh, A.; Sah, D.; Chakraborty, M.; Rai, J. Bio-mediated Detoxification of Heavy Metal Contaminated Soils and Phytotoxicity Reduction Using Novel Strain of Brevundimonas vancanneytii SMA3. Heliyon 2023, 9, e22344. [Google Scholar] [CrossRef]
- Singh, N.; Marwa, N.; Mishra, S.; Mishra, J.; Verma, P.; Rathaur, S.; Singh, N. Brevundimonas diminuta Mediated Alleviation of Arsenic Toxicity and Plant Growth Promotion in Oryza sativa L. Ecotoxicol. Environ. Saf. 2016, 125, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Li, M.; Su, J.; Li, Y.; Wang, Z.; Bai, Y.; Ali, E.; Shaheen, S. Brevundimonas diminuta Isolated from Mines Polluted Soil Immobilized Cadmium (Cd2+) and Zinc (Zn2+) Through Calcium Carbonate Precipitation: Microscopic and Spectroscopic Investigations. Sci. Total Environ. 2022, 813, 152668. [Google Scholar] [CrossRef] [PubMed]
- Resmi, G.; Thampi, S.; Chandrakaran, S. Brevundimonas vesicularis: A Novel Bio-sorbent for Removal of Lead from Wastewater. Int. J. Environ. Res. 2010, 4, 281–288. [Google Scholar]
- Singh, N.; Gadi, R. Bioremediation of Ni(II) and Cu(II) from Wastewater by the Nonliving Biomass of Brevundimonas vesicularis. J. Environ. Chem. Ecotoxicol. 2012, 4, 137–142. [Google Scholar]
- Hughes, E.; Head, B.; Maltman, C.; Piercey-Normore, M.; Yurkov, V. Aerobic Anoxygenic Phototrophs in Gold Mine Tailings in Nopiming Provincial Park, Manitoba, Canada. Can. J. Microbiol. 2017, 63, 212–218. [Google Scholar] [CrossRef]
- Poindexter, J. Biological Properties and Classification of the Caulobacter Group. Bacteriol. Rev. 1964, 28, 231–295. [Google Scholar] [CrossRef]
- Choi, J.-H.; Kim, M.-S.; Roh, S.; Bae, J.-W. Brevundimonas basaltis sp. nov., Isolated from Black Sand. Int. J. Syst. Evol. Microbiol. 2010, 60, 1488–1492. [Google Scholar]
- Jordan, E.; Caldwell, M.; Reiter, D. Bacterial Motility. J. Bacteriol. 1934, 27, 165–174. [Google Scholar] [CrossRef]
- Bauer, A.; Kirby, W.; Sherry, J.; Turck, M. Antibiotic Susceptibility Testing by Using a Standardized Single Disc Method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Holt, J.; Krieg, N.; Sneath, P. Bergey’s Manual of Determinative Bacterology; The Williams and Wilkins Co: Baltimore, MD, USA, 1994. [Google Scholar]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A Simple Method for the Isolation and Purification of Total Lipids from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Estrela, A.; Abraham, W. Brevundimonas vancanneytii sp. nov., Isolated from Blood of a Patient with Endocarditis. Int. J. Syst. Evol. Microbiol. 2010, 60, 2129–2134. [Google Scholar] [CrossRef] [PubMed]
- Minnikin, D.E.; O’Donnell, A.G.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.; Parlett, J.H. An Integrated Procedure for the Extraction of Bacterial Isoprenoid Quinones and Polar Lipids. J. Microbiol. Methods 1984, 2, 233–241. [Google Scholar] [CrossRef]
- Chen, W.; Kuo, T. A Simple and Rapid Method for the Preparation of Gram-negative Bacterial Genomic DNA. Nucleic Acids Res. 1993, 21, 2260. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Molec. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Maltman, C.; Kuzyk, S.B.; Kyndt, J.; Lengyel, G.; Yurkov, V. Shewanella metallivivens sp. nov., a Deep-sea Hydrothermal Vent Tube Worm Endobiont Capable of Dissimilatory Anaerobic Metalloid Oxyanion Reduction. Int. J. Sys. Evol. Microbiol. 2023, 73, 005980. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; Gerdes, S.; Henry, C.S.; Kenyon, R.W.; Machi, D.; Mao, C.; Nordberg, E.K.; Olsen, G.J.; Murphy-Olson, D.E.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; Shukla, M.; Vonstein, V.; Warren, A.; Xia, F.; Yoo, H.; Stevens, R.L. Improvements to PATRIC, the All-Bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI Prokaryotic Genome Annotation Pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Lim, J.M.; Kwon, S.J.; Chun, J. A Large-scale Evaluation of Algorithms to Calculate Average Nucleotide Identity. Antonie van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Sardà Carbasse, J.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A Database Tandem for Fast and Reliable Genome-based Classification and Nomenclature of Prokaryotes. Nucleic Acid Res. 2021, 50, D801–D807. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.; Goker, M. TYGS is an Automated High-throughput Platform for State-of-the-art Genome-based Taxonomy. Nat. Comm. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A Comprehensive, Accurate, and Fast Distance-based Phylogeny Inference program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [PubMed]
- Farris, J.S. Estimating Phylogenetic Trees from Distance Matrices. Am. Nat. 1972, 106, 645–667. [Google Scholar] [CrossRef]
- Kreft, L.; Botzki, A.; Coppens, F.; Vandepoele, K.; Van Bel, M. PhyD3: A Phylogenetic Tree Viewer with Extended phyloXML Support for Functional Genomics Data Visualization. Bioinformatics 2017, 33, 2946–2947. [Google Scholar] [CrossRef]
- Richter, M.; Rossello-Mora, R. Shifting the Genomic Gold Standard for the Prokaryotic Species Definition. PNAS 2009, 106, 19126–19131. [Google Scholar] [CrossRef]
Figure 1.
Photosynthetic complex and cell morphology of C11T. (A) Whole cell (light orange line) and pigment extract (dark orange, dashed line) absorbance spectra. Peaks and shoulders of importance are indicated. (B) Phase contrast micrograph of cells. Bar is 5 μm.
Figure 1.
Photosynthetic complex and cell morphology of C11T. (A) Whole cell (light orange line) and pigment extract (dark orange, dashed line) absorbance spectra. Peaks and shoulders of importance are indicated. (B) Phase contrast micrograph of cells. Bar is 5 μm.
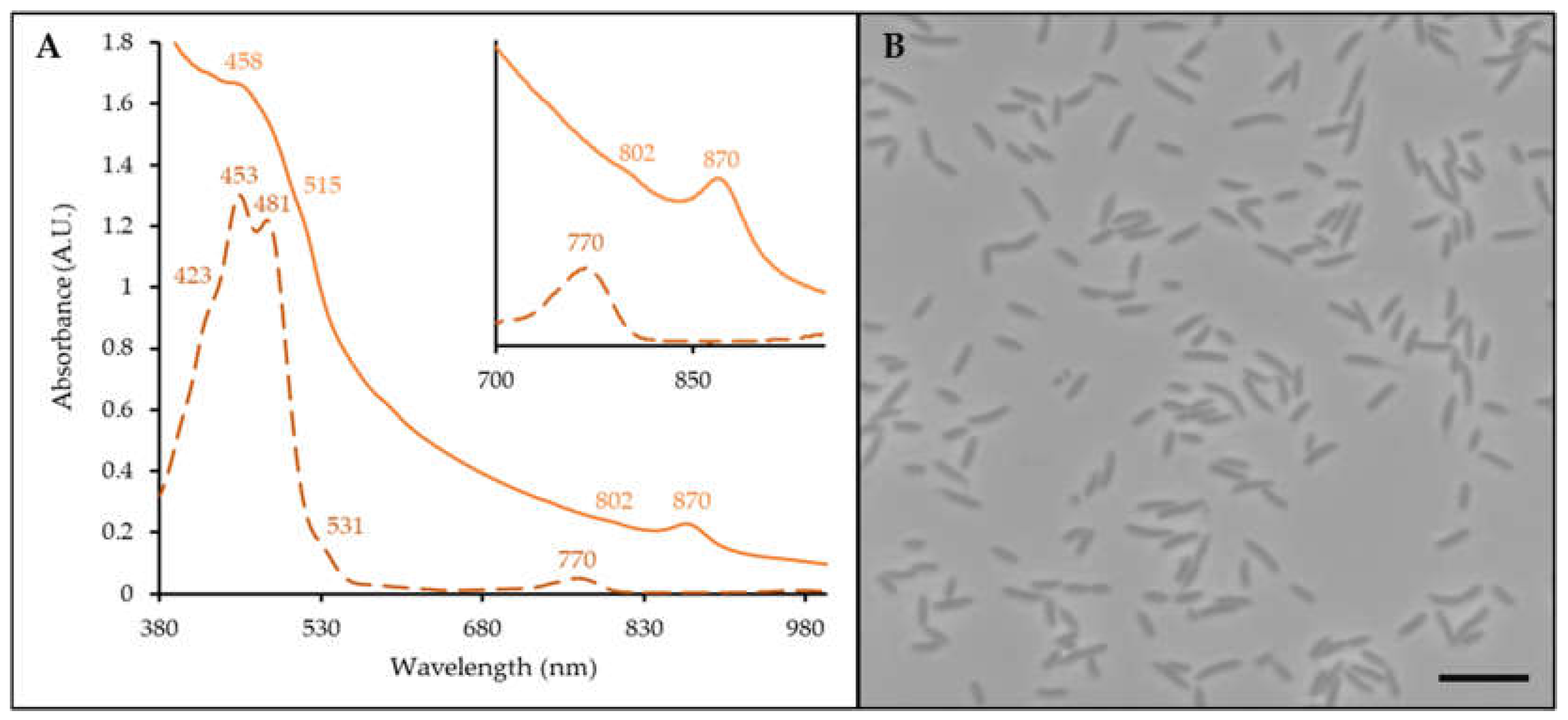
Figure 2.
Maximum Likelihood 16S rRNA phylogenetic tree of strain C11T and validated species in Brevundimonas. The scale bar represents the amount of substitutions per nucleotide. Accession numbers are in parenthesis. Value at nodes indicate the bootstrap support calculated with neighbor joining analysis of 1000 resampled datasets; only values >50 % are shown. Aquidulcibacter paucihalophilus LMG 28362T was used as the outgroup.
Figure 2.
Maximum Likelihood 16S rRNA phylogenetic tree of strain C11T and validated species in Brevundimonas. The scale bar represents the amount of substitutions per nucleotide. Accession numbers are in parenthesis. Value at nodes indicate the bootstrap support calculated with neighbor joining analysis of 1000 resampled datasets; only values >50 % are shown. Aquidulcibacter paucihalophilus LMG 28362T was used as the outgroup.
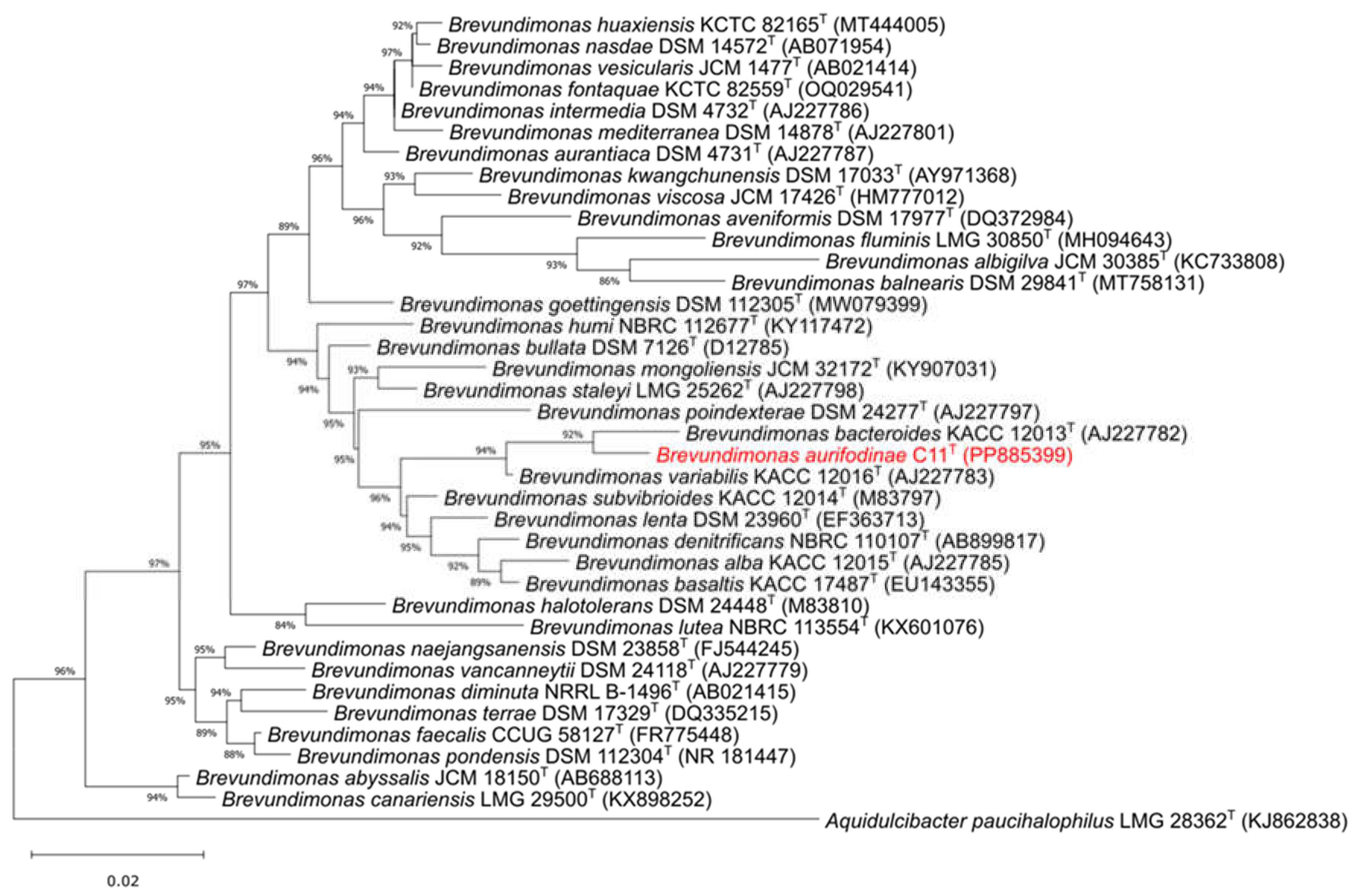
Figure 3.
Genome Blast Distance Phylogeny tree of C11T among species of the genus Brevundimonas. The numbers above branches are pseudo-bootstrap support values > 60 % from 100 replications, with an average branch support of 80.1 %. The tree was rooted at the midpoint.
Figure 3.
Genome Blast Distance Phylogeny tree of C11T among species of the genus Brevundimonas. The numbers above branches are pseudo-bootstrap support values > 60 % from 100 replications, with an average branch support of 80.1 %. The tree was rooted at the midpoint.
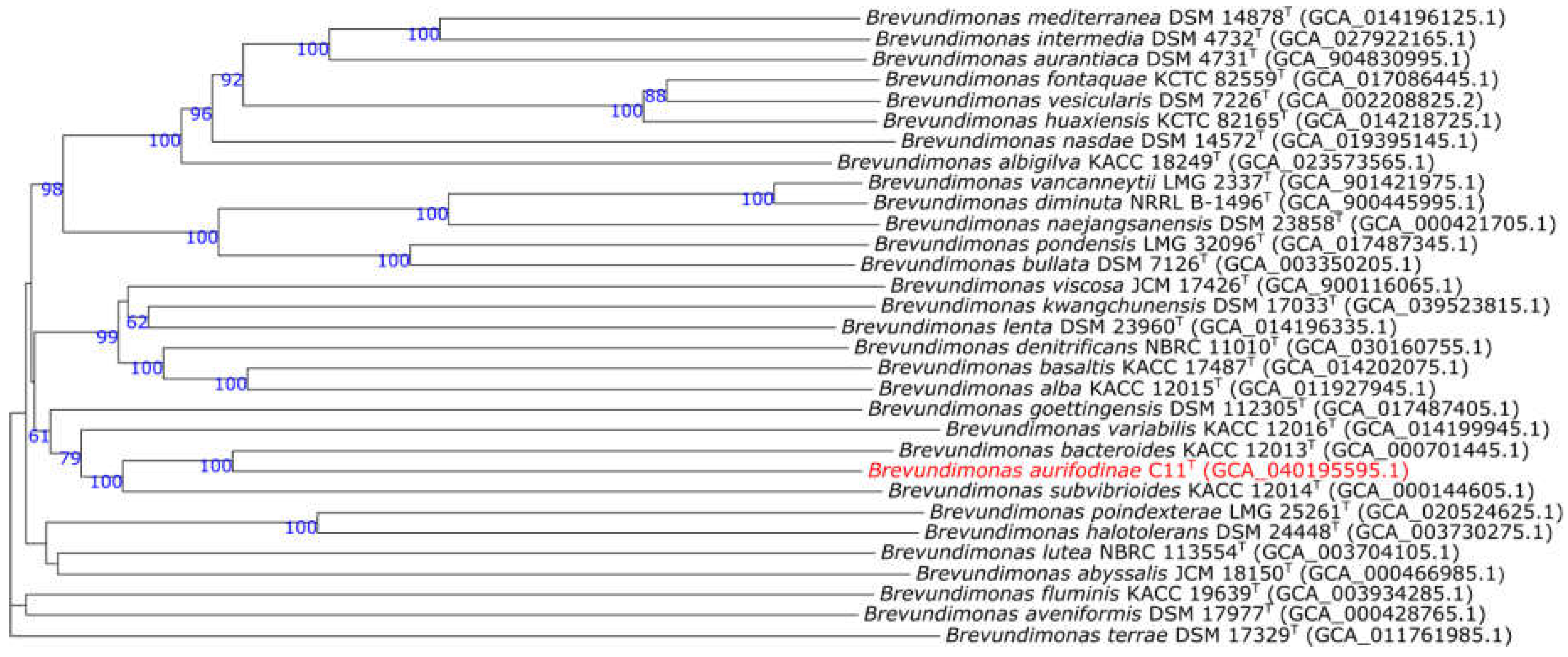

Table 1.
Physiological and biochemical features of C11T compared to related members and the type species of the genus Brevundimonas. 1,2.
Table 1.
Physiological and biochemical features of C11T compared to related members and the type species of the genus Brevundimonas. 1,2.
| Species | B. aurifodinae | B. bacteroides | B. variabilis | B. basaltis | B. alba | B. subvibrioides | B. diminuta |
| Strain | C11T | KACC 12013T | KACC 12016T | KACC 17487T | KACC 12015T | KACC 12014T | NRRL B-1496T |
| Temperature (°C) | 5-40 | 25-40 | 10-40 | 10-40 | 10-40 | 25-40 | 10-40 |
| Optimum | 30 | 30 | 30 | 30 | 30 | 20 | 30 |
| pH | 6.0-10.5 | 6.0-8.0 | 6.0-8.0 | 5.5-10.0 | 6.0-8.0 | 6.0-8.0 | 6.0-8.0 |
| Optimum | 8.0 | 7.0 | 7.0 | 7.5 | 7.0 | 7.0 | 7.0 |
| NaCl Tolerance (%) | 2 | 4 | 4 | 4 | 2 | 2 | 6 |
| Utilization of: | |||||||
| D-maltose | - | - | + | - | - | - | + |
| D-cellobiose | + | - | + | - | - | + | + |
| Gentibiose | + | + | + | + | - | - | + |
| Sucrose | + | - | + | - | - | - | + |
| Stachyose | + | + | + | - | - | - | + |
| D-raffinose | - | - | + | - | - | + | + |
| β-methyl-D-glucoside | - | + | - | + | - | + | + |
| D-salicin | - | + | + | - | - | + | + |
| α-D-glucose | + | - | + | + | + | + | + |
| D-mannose | + | + | - | - | + | + | + |
| D-fructose | - | - | - | + | - | - | + |
| Gelatin | + | + | + | - | + | - | + |
| L-arginine | - | + | + | + | - | - | + |
| Pectin | - | - | + | + | - | + | + |
| Mucic acid | - | + | + | + | - | + | + |
| D-malic acid | + | + | - | + | - | + | + |
| α-hydroxy-butyric acid | + | + | - | - | - | - | + |
| α-keto-butyric acid | + | - | - | - | - | - | + |
| Propionic acid | - | + | - | + | - | - | - |
| Enzyme Activities | |||||||
| Amylase | + | + | + | - | - | + | - |
| Gelatinase | + | + | + | - | + | - | - |
| Leucine arylamidase | + | - | + | + | + | + | + |
| Valine arylamidase | + | - | + | + | + | + | + |
| Cysteine arylamidase | - | - | - | + | - | + | - |
| α-chymotrypsin | + | - | + | + | + | + | + |
| α-glucosidase | + | + | + | + | + | + | - |
| Oxidase | + | + | - | - | + | + | + |
| Esculin Hydrolysis | + | + | + | + | + | + | - |
1 +, Growth occurs, enzyme activity detected; -, No growth, no enzyme activity 2 Physiological results not included here produced identical results among C11T and Brevundimonas spp. tested.
Table 2.
General genome features of C11T compared to related species, and the type strain, of the genus Brevundimonas.
Table 2.
General genome features of C11T compared to related species, and the type strain, of the genus Brevundimonas.
| Species | B. aurifodinae | B. bacteroides | B. variabilis | B. basaltis | B. alba | B. subvibrioides | B. diminuta |
| Strain1 | C11T | KACC 12013T | KACC 12016T | KACC 17487T | KACC 12015T | KACC 12014T | NRRL B-1496T |
| 16s rRNA Gene Similarity (%)2 | 100 | 98.4 | 98.2 | 97.5 | 97.4 | 97.2 | 94.8 |
| Genome Size (Mb) | 3.3 | 3.2 | 3.4 | 2.6 | 3.1 | 3.4 | 3.4 |
| G+C Content | 68.3 | 68.2 | 65.3 | 68.5 | 68.6 | 68.4 | 67.3 |
| Genes | 3258 | 3225 | 3320 | 2715 | 3056 | 3385 | 3438 |
| Protein-coding genes | 3186 | 3169 | 3246 | 2649 | 3000 | 3325 | 3358 |
| No. of contigs | 26 | 17 | 8 | 8 | 3 | 1 | 2 |
| No. of tRNA operons | 43 | 44 | 46 | 43 | 46 | 47 | 53 |
| L50 | 5 | 4 | 2 | 2 | 1 | 1 | 1 |
| OrthoANI (%) 2 | 100 | 83.5 | 77.7 | 77.7 | 77.4 | 80.9 | 77.3 |
| dDDH (%) 2 | 100 | 26.6 | 21.8 | 21.4 | 21.3 | 23.5 | 20.8 |
1 GenBank Assembly Accession numbers of strains from left to right: GCA_040195595.1, GCA_000701445.1, GCA_014199945.1, GCA_014202075.1, GCA_011927945.1, GCA_000144605.1, GCA_900445995.1. 2 Compared to C11T.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



