
Submitted:
19 September 2024
Posted:
19 September 2024
You are already at the latest version
Abstract
Fucoidans, known for their diverse biological properties such as anti-inflammatory, antiviral, antitumor, and immune-stimulatory effects, have recently gained attention for their potential benefits in exercise endurance, muscle mass, and anti-fatigue. However, the mechanisms by which fucoidans enhance exercise performance are still unclear. To investigate these effects, we administered 400 mg/kg/day of fucoidan extract derived from Undaria pinnatifida to sixty-four C57BL/6J mice over ten weeks. We evaluated changes in running activity, mitochondrial-related gene expression in skeletal muscle, and alterations in the intestinal microbiome. Our results showed that fucoidan supplementation significantly increased daily running distance and muscle mass by 25.5% and 10.4%, respectively, in mice on a standard chow diet, and with more modest effects observed in those on a high-fat diet (HFD). Additionally, fucoidan supplementation led to a significant increase in beneficial gut bacteria, including Bacteroides/Prevotella, Akkermansia muciniphila, and Lactobacillus, along with a notable reduction in the Firmicutes/Bacteroidetes ratio, indicating improved gut microbiome health. Mechanistically, fucoidan supplementation upregulated the mRNA expression of key genes related to mitochondrial biogenesis and oxidative capacity, such as COX4, MYH1, PGC-1α, PPAR-γ, and IGF1, in both standard chow and HFD-fed mice. Our findings suggest that fucoidan supplementation enhances exercise performance, improves muscle function, and positively modulates the gut microbiome in mice, regardless of diet. These effects may be attributed to fucoidans' potential prebiotic role, promoting the abundance of beneficial gut bacteria and contributing to enhanced exercise performance, increased muscle strength, and improved recovery.
Keywords:
Fucoidans
; Undaria pinnatifida
; exercise performance
; gut microbiome
; Firmicutes/Bacteroidetes ratio
; Mitochondrial biogenesis
; IGF-1
; COX4
; MYH1
1. Introduction
Fucoidans are complex polysaccharides primarily extracted from various species of marine brown algae, containing substantial amounts of L-fructose and sulphate ester groups [1,2]. Over the past decades, fucoidans have been extensively studied for their wide range of biological properties including anti-inflammatory, antiviral, antitumor as well as immune stimulatory activities [3,4,5,6,7,8].
In the field of exercise performance, recent interest has focused on the biological role of fucoidan which demonstrated beneficial effects on exercise endurance, muscle mass and function, and anti-fatigue actions in mice [9,10,11]. Three weeks of Laminaria japonica fucoidan supplementation (310 and 620 mg/kg/day) increased grip strength and endurance swimming time in a dose-dependent manner, suggesting that fucoidan possesses a wide spectrum of bioactivities that can improve exercise performance and exhibit anti-fatigue effects [9]. In a later study, eight weeks of Undaria pinnatifida fucoidan (UPF) supplementation (0.25% of diet weight) enhanced mitochondrial biogenesis, increased oxidative muscle fibre, and promoted angiogenesis in skeletal muscles, resulting in increased treadmill distance and skeletal muscle mass [11]. Similarly, four weeks of oral administration of a UPF and a Fucus vesiculosus fucoidan (FVF) blend (400 mg/kg/day) increased muscle size and strength in both exercised and non-exercised mice, suggesting an important influence of fucoidan on skeletal muscle physiology [10].
Clinical trials in athletes consuming fucoidan demonstrated that the bioactive compound may influence various aspects of human physiology. For instance, in a double blind randomised controlled clinical trial, two weeks of UPF supplementation (1 g/day) had no effects on exercise performance, but it promoted a modest effect on inflammatory cytokines [12]. In another study, a blend of UPF and FVF increased the concentration of faecal lysozyme [6], a protein known for its antimicrobial and anti-inflammatory properties suggesting that fucoidan plays a potential role in protecting mucosal barrier integrity [13].
Additionally, in recent years, researchers have produced a significant amount of literature dedicated to understanding how fucoidans influence the gut microbiome’s composition and function. Several studies suggest that fucoidans induce favourable microbiota alterations exerting prebiotic effects [2,14] and scientific evidence suggests a link between fucoidan, diet, exercise performance and composition of the gut microbiome [6,15,16,17]. In mice fed a high fat diet (HFD), 8 weeks of UPF supplementation alleviated dyslipidaemia decreasing the total serum cholesterol, LDL cholesterol (LDL-C), and liver cholesterol levels as well as modulating gut microbiota [18]. Similarly, 5 weeks of fucoidan administration reduced obesity and improved gut microbiota in mice [19].
Regular exercise can enrich the microflora diversity improving the development of beneficial bacteria, including Bifidobacterium and Akkermansia which are linked to gut health for their anti-inflammatory properties and their positive impact on gut barrier integrity [20,21]. Exercise can also modify the ratio of two dominant phyla, Firmicutes and Bacteroidetes which has been associated with several diseases including diabetes, gut and brain health, and some cancers [22,23]. In obese mice, twelve-week free running wheel exercise intervention ameliorated the metabolic consequences caused by HFD consumption and changed the gut microbiome composition, increasing the abundance of Bacteroides while decreasing Firmicutes [24].
Based on the effects of UPF on gut microbiome and skeletal muscle, in the current study we aimed to examine whether UPF would mitigate the negative impact of HFD consumption on exercise performance while increasing the abundance of beneficial gut bacteria. For this purpose, UPF was administered orally to both male and female mice for ten weeks. The daily voluntary running activity and changes in intestinal microbiome were assessed. To further investigate the mechanisms involved, the expression of genes involved in mitochondrial activity and energy utilization in skeletal muscle that influence contraction speed, endurance, and metabolic adaptation to exercise were measured [11,25]. Specifically, markers such as mitochondrial cytochrome c oxidase subunit 2 and 4 (COX2 and COX4) as well as muscle myosin heavy chain 1 (MYH1) were measured.
2. Results
2.1. Effects of UPF on Running Activity and Muscle Mass
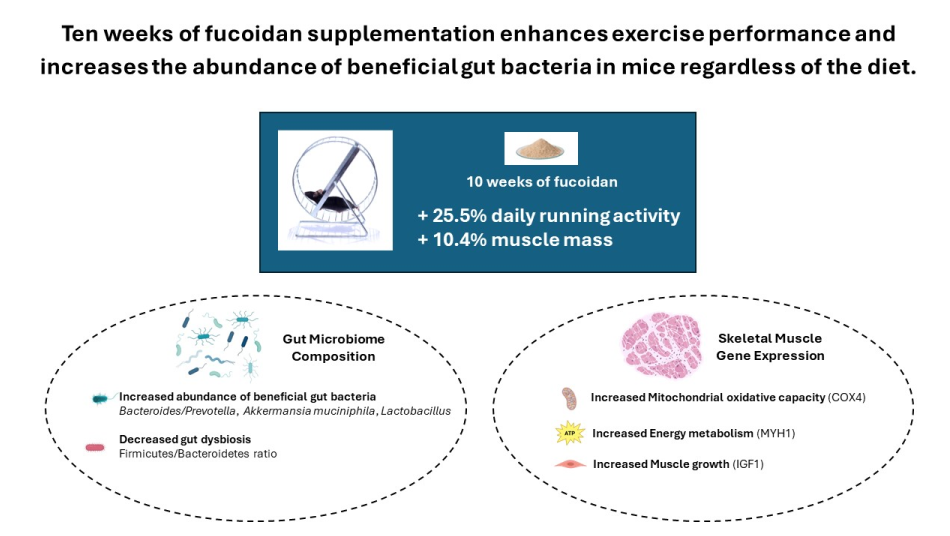
In mice consuming a standard chow diet, weekly running distance difference reached statistical significance after two weeks of UPF treatment and was maintained until the end of the experiment (p < 0.05, Figure 1A).
In mice consuming HFD, weekly running distance difference reached statistical significance after 1 week of UPF treatment and was maintained for the following 4 weeks. (p < 0.05, Figure 1B).
Mice consuming a chow diet ran significantly greater distances daily than those consuming HFD (Figure 1C). Specifically, in mice consuming a standard chow diet, UPF significantly increased daily running activity compared to the control group (25.5 %; p < 0.05; Figure 1C). UPF did not improve running activity in mice consuming HFD.
Mice consuming a chow diet together with UPF showed significantly higher muscle mass than mice consuming a chow diet only (10.4%; p < 0.05; Figure 1D). This difference was not observed between mice on HFD with or without fucoidan consumption.
2.2. Effects of UPF on Total DNA Content in the Faeces
Overall, mice consuming chow diet had higher DNA content in the faeces (20.6%) compared to mice consuming HFD (Figure 2A). In mice consuming HFD, UPF increased the total DNA content (14.8%), (Figure 2A, p < 0.05), while the compound had no effect on mice consuming chow diet.
2.2.1. Effects of Diet and UPF Administration on Gut Microbial Profiling
Overall, Bacteroides/Prevotella, Akkermansia muciniphila, and Lactobacillus group (Figure 2B-D) had higher DNA abundance in mice fed HFD. Conversely, in the phylum Bacteroidetes (Figure 2E), HFD had significantly lower DNA abundance compared to CHOW groups.
In mice eating standard chow diet, UPF supplementation significantly increased the DNA levels of Bacteroides/Prevotella (p < 0.05, Figure 2B) and the phylum Bacteroidetes (p < 0.05, Figure 2E).
In mice consuming HFD, animals receiving UPF showed a significant increase in two bacterial species, including Bacteroides/Prevotella (p < 0.05, Figure 2B) and Akkermansia muciniphila (p < 0.05, Figure 2C). Likewise, the administration of UPF induced a significant increase in Lactobacillus group (p < 0.05, Figure 2D) and Bacteroidetes phylum (p < 0.05, Figure 2E). However, consuming UPF was not associated with a change in abundance of bacterial Clostridial coccoides (Figure 2F) and Firmicutes (Figure 2G).
In contrast, in mice receiving UPF the ratio of Firmicutes to Bacteroidetes phylum in the faeces of mice consuming HFD was significantly reduced (p < 0.05, Figure 2H), but no UPF effect was observed in the chow group.
2.3. Correlation of Gut Microbial Abundance with Mouse Body Weight, Visceral Fat, and Running Activity
When the abundance of specific bacterial groups/species in mouse faecal contents were plotted against body weight, visceral fat (gonadal and retroperitoneal fat pads), and daily running distance, Bacteroidetes phylum (r = -0.708; p = 0.0001; n = 10) and Akkermansia muciniphila (r = -0.399; p = 0.0354; n = 7) were found to be significantly negatively correlated with body weight. In addition, visceral fat was negatively correlated with the phylum Bacteroidetes (r = -0.779; p = 0.0001; n = 10). Conversely, running distance showed significantly positive correlations with Bacteroidetes phylum (r = 0.598; p = 0.0001; n = 10) and Akkermansia muciniphila (r = 0.420; p = 0.0259; n = 7); however, a negative correlation with running distance was found in Clostridial coccoides (r = -0.430; p = 0.0089; n = 9). From the perspective of F/B (Firmicutes to Bacteroidetes) ratio, body weight (r = 0.568; p = 0.0001; n = 10) and visceral fat (r = 0.593; p = 0.0001; n = 10) showed significantly positive correlations; conversely, the running distance was negatively correlated with F/B ratio (r = -0.559; p = 0.0002; n = 10). The correlation matrix is presented in Table 1.
2.4. Effects of Diet and UPF on Muscle Gene Expression
Overall, HFD increased the muscle mRNA expression of cytochrome c oxidase subunit 2 and 4 (COX2 and COX4, Figure 3A, B), myosin heavy chain 1 (MYH1, Figure 3C), peroxisome proliferator-activated receptor gamma coactivator 1α (PGC-1α, Figure 3D), peroxisome proliferator-activated receptor gamma (PPAR-γ, Figure 3E), and Insulin-like growth factor 1 (IGF1, Figure 3F) compared with standard chow diet.
3. Discussion
The current study identified potential mechanisms by which UPF may enhance exercise performance, focusing on changes in the abundance of beneficial gut bacteria and alterations in skeletal muscle gene expression.
Over the ten-week experimental period, UPF supplementation (400 mg/kg/day) increased voluntary running activity in mice on both a standard chow diet and a high-fat diet (HFD). In mice on the standard chow diet, two weeks of UPF treatment enhanced running distance, with this effect sustained throughout the experiment. Additionally, mice that ingested UPF had an increase in muscle mass of 10.4% compared to the untreated group. In the HFD group, mice that ingested UPF also significantly increased running distance, but this effect lasted only five weeks and there was no impact on muscle mass. The results also show that UPF upregulated the key genes involved in mitochondrial function and energy homeostasis, including COX4, MYH1, PGC-1α, PPAR-γ, and IGF1 and these effects were associated with changes in the abundance of some beneficial gut bacteria belonging to the Firmicutes and Bacteroides phyla.
To build upon previous studies investigating the exercise performance-enhancing properties of UFP [6,9,10], we aimed to elucidate its potential mechanism of action. Our results suggest that UPF may improve exercise performance by modifying the abundance of beneficial gut bacteria which are known to influence muscle strength and energy utilization [17,26,27].
According to several studies, fucoidans promote beneficial effects on exercise endurance, muscle mass and function, and anti-fatigue actions [9,10,11]. Consistent with these findings, we confirmed that UPF increased weekly running activities in mice on both chow and high-fat diets, with a more pronounced effect in those on chow diet. This effect emerged after just two weeks of treatment and persisted throughout the experiment, whereas in the high-fat diet group, the effect lasted four weeks. We also noted that the increase in running activity was accompanied by an increase in muscle mass, but this was observed only in mice consuming chow diet. Our results are in line with previous results, where three weeks of fucoidan supplementation derived from Laminaria japonica (620 mg/kg/day) improved the exercise performance of male mice in a dose-dependent manner [9]. Results of the study showed that fucoidan significantly increased grip strength and endurance swimming time and that this treatment led to dose-dependent reductions of exercise-induced fatigue-related parameters including serum lactate and ammonia levels, along with an increase in glucose levels following a 15-minute swimming test [9]. Similarly, four weeks of UPF and Fucus vesiculosus fucoidan (400 mg/kg/day) increased muscle size and strength in both exercised and no-exercised mice, suggesting an important influence of fucoidan on skeletal muscle physiology [10]. In humans, several physiological effects of fucoidan consumption have been observed in athletes. For example, in a clinical trial, two weeks of fucoidan supplementation (1 g/day) had no effects on exercise performance, but it promoted a modest effect on inflammatory cytokines [12]. In another study, fucoidan increased the concentration of faecal lysozyme [6], a protein known for its antimicrobial and anti-inflammatory properties, suggesting that fucoidan plays a potential role in protecting mucosal barrier integrity [13]. Collectively, our findings and previous research suggest that long-term fucoidan supplementation may offer broad health benefits, including performance enhancement and anti-fatigue effects.
On the basis of these results, we tested the effects of UPF on key genes involved in mitochondrial function and energy homeostasis, including COX4, MYH1, PGC-1α, PPAR-γ, and IGF1 in skeletal muscle. In mice consuming a standard chow diet or HFD, UPF significantly upregulated the mRNA expression of COX4, MYH1, PGC-1α, PPAR-γ, and IGF1, indicating improved mitochondrial biogenesis and oxidative capacity [28,29].
COX2 and COX4 are integral subunits of respiratory chain complex IV in the mitochondrial electron transport chain; COX2, encoded by mitochondrial DNA, forms the catalytic core essential for electron transfer and oxygen reduction, while COX4, encoded by nuclear DNA, modulates the enzyme’s activity and efficiency [30]. The synergism between COX2 and COX4 ensures effective oxidative phosphorylation (OXPHOS) and ATP synthesis, which are vital for cellular energy production [31]. The upregulation of COX2 and COX4 in this study supported the hypothesis that UPF enhances mitochondrial function and oxidative metabolism [25], crucial for maintaining muscle efficiency under metabolic stress [31,32]. Additionally, we report that UPF increases MYH1 expression, which is central for maintaining muscle fibre composition and structural integrity during endurance activities in physiological conditions [33,34]. Furthermore, increased PPAR-γ expression suggests a potential role in regulating lipid metabolism within muscle tissues, which may contribute to improved muscle performance and adaptation [35]. These effects likely explain the observed improvements in voluntary running activity and muscle mass in the UPF-treated chow group, suggesting that UPF supports enhanced muscle function and endurance [11,36]. Collectively, our findings suggest that UPF may enhance muscle energy metabolism and improve fatigue resistance by modulating genes involved in mitochondrial biogenesis, oxidative metabolism, and muscle growth, across both standard chow and high-fat diet contexts. Further research is needed to elucidate the molecular mechanisms and assess the long-term benefits of UPF supplementation on muscle function and metabolic health.
In addition to previous studies investigating the impacts of UPF on exercise performance and muscle function [9,10], the present study aimed to elucidate how ten weeks of UPF supplementation would influence the changes in the gut microbiome, and specifically on the abundance of Firmicutes and Bacteroides phyla.
It is interesting to note that in our study, mice consuming HFD supplemented with UPF exhibited a 14.9% increase in total DNA content compared to their control group. The change in DNA content that we observed could be explained, at the least in part, by the evidence that exercise training alters the composition and functional capacity of the gut microbiota independently of the diet consumed [15,16,22,37,38,39]. Additionally, other studies demonstrated that the magnitude of the changes in bacterial DNA may be related to the obesity status as well as exercise intensity and modality [17,27,38,40].
In our study, we investigated the abundance of Akkermansia muciniphila, a bacterium widely accepted as biomarker of intestinal health because of its numerous beneficial properties, such as improved glucose metabolism, enhanced insulin sensitivity, inflammation [41,42,43]. Here, we report that while a chow diet did not affect the abundance of Akkermansia muciniphila, mice on a high-fat diet showed reduced levels of this bacterium. However, UPF supplementation significantly increased Akkermansia muciniphila abundance, suggesting a prebiotic role in promoting the growth of beneficial gut bacteria [14,44,45].
We also measured the abundance of Lactobacillus group, a bacterium that plays a major role in human health reducing the release of cortisol, decreasing inflammatory cytokines, and improving the immune system [46,47]. Lactobacillus group is associated to a probiotic agent itself because it helps to create a favourable gut environment [46] lowering the gut pH and inhibiting the growth of harmful intestinal bacteria [46,47]. In mice fed a standard chow diet and treated with UPF, the levels of Lactobacillus remained unchanged. However, in those consuming HFD, the abundance of Lactobacillus increased, and the treatment with UPF further augmented Lactobacillus levels within this group. Additionally, UPF increased the abundance of Bacteroides/Prevotella, bacterial species recognized for their prebiotic effects [48] suggesting and reinforcing a potential prebiotic role of UPF.
A novel finding of our study is that UPF positively altered the Firmicutes-to-Bacteroidetes (F/B) ratio. Firmicutes and Bacteroidetes represent the predominant gastrointestinal bacterial phyla, collectively constituting over 90% of the gut microbiota [49]. The F/B ratio has been linked to the maintenance of healthy intestinal homeostasis, and disruption of this balance can result in several serious complications in both humans and animals [50,51,52]. For instance, obese individuals typically exhibit an elevated F/B ratio compared to healthy counterparts [40,53]. Conversely, patients with breast cancer tend to have an F/B ratio three times lower than that of healthy controls [52]. Inflammatory bowel disease is also associated with a low F/B ratio, while benign prostatic hyperplasia is linked to a high F/B ratio [50,51].
In our study, and in line with previous research in which HFD consumption increased the phylum Firmicutes and decreased Bacteroidetes [54], mice consuming HFD had a significant higher F/B ratio. However, ten weeks of oral UPF supplementation significantly lowered this ratio compared to their control group, suggesting a potential benefit of the supplement in modulating beneficial gut bacteria. We also found that the F/B ratio positively correlated with body weight, and negatively with running activity as demonstrated in previous studies where exercise reduced the F/B ratio in obese mice [39]. Overall, our findings substantiated the effects of UPF on beneficial gut bacteria including Akkermansia muciniphila and Lactobacillus and on the F/B ratio suggesting a prebiotic role of the supplement especially in concomitance of HFD consumption.
While our targeted approach to analyse specific genes of interest has shed light on the potential impacts on mitochondrial and fibre function in muscle, global “omics” approaches such as mRNA sequencing and proteomics would lead to more comprehensive insight into the activity of UPF. Furthermore, detailed characterization of UPF constituents would facilitate investigation into the molecular mechanism involved in any therapeutic effects of the supplement. There is a consensus that it is important to consider the source of fucoidan when comparing outcomes, as its properties can vary significantly based on factors such as species, molecular weight fractions, and component percentages [55].
In conclusion, based on the data presented in this study, we propose a descriptive mechanism by which ten weeks of UPF supplementation enhanced exercise performance, upregulated muscle function-related genes, and modulated the gut microbiome abundance of beneficial bacteria in mice consuming a standard chow diet as well as HFD. Our findings suggest that UPF may play a prebiotic role in the gut contributing to an increase in the abundance of beneficial gut bacteria which may result in enhanced exercise performance, increased muscle strength, or improved recovery times. Further research is needed to elucidate the underlying molecular mechanisms by which UPF exerts its beneficial therapeutic effects.
4. Materials and Methods
4.1. Ethics Statement
This investigation was authorised by the Animal Ethics Committee of the University of Tasmania (A0027164). Animal work and procedures in this study were executed in strict adherence to the provisions delineated in the Tasmanian Animal Welfare Act (1993/63) and the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes 8 edition 2013 [56].
4.2. Animals and Diet
Four-week-old male and female C57BL/6J mice (n=32 males and n=32 females, Animal Services, University of Tasmania) were housed under controlled conditions at a temperature of 20 ± 2°C and maintained on a standard 12:12h light/dark cycle. Following a one-week acclimatization period, 32 mice were provided with ad libitum access to a commercial high-fat pelleted diet (HFD) (19.4 MJ/kg, 23.5% fat, 23% protein, 5.4% crude fibre, SF16-059, Specialty Feeds, Perth, Australia), while the control group (32 mice) received a standard chow diet (12.8 MJ/kg, 6% fat, 20% protein, 3.2% crude fibre, product code 102108, Ridly Agri-Products). Mice were singularly housed with unrestricted access to food and drinking water ad libitum. Each cage was equipped with a free-running wheel, shown in Supplementary Information (Figures S1 and S2), to assess the mice’s exercise performance and mitigate social stress associated with isolation. Throughout the study, running distance was recorded daily, while body weight and food intake were recorded weekly. Mice were euthanized after ten weeks of UPF administration.
4.3. Experimental Groups
64 mice were randomly divided into four groups (n=16): CHOW, CHOW+UPF, HFD, and HFD+UPF.
4.4. UPF Jelly Preparation
A fucoidan extract (≥85% fucoidan) derived from Undaria pinnatifida (UPF, Batch No.: UPF2022532) was provided by Marinova Pty Ltd. (Tasmania, Australia) and stored at ambient temperature (21-29°C). The chemical composition of fucoidan used in this study is described in Supplementary Information (Tables S1 and S2). Fucoidan was incorporated into artificially flavoured and sweetened jelly for voluntary oral administration to mice by using 6% w/v gelatine (Ward McKenzie, Australia) as the gelling agent, 9% v/v strawberry essence (Roberts Edible Craft, Australia) for flavour, and 2% w/v sucralose (Cat# 69293, Sigma-Aldrich, Australia) as a non-caloric sweetener.
In CHOW+UPF and HFD+UPF groups, each serve of jelly was prepared with a total volume of 200 µL, incorporating 12 mg of UPF (equivalent 400 mg/kg/day). In CHOW and HFD groups, each serve of jelly was prepared in the same manner but without the addition of fucoidan. Both UPF and vehicle jellies were served daily in small dishes.
4.5. Training and Jelly Administration
Mice were trained to consume the jelly using a previously reported method [57] to ensure mice were accustomed to its taste and texture. The jelly was prepared weekly and stored at 4°C to maintain freshness. Jelly was consistently served to mice at 9:00 am each day, alongside their regular food (standard chow diet or HFD) to ensure continuous food intake and prevent potential nutritional deficiencies. All animals consumed the full 200 µL jelly provided daily (Supplementary Video S1).
4.6. Sample Collection
After ten weeks of drug administration, mouse faecal samples were collected in the morning and stored at −80°C freezer for gut microbial profiling study. Mice were then fasted overnight (12 h) and euthanized via carbon dioxide inhalation. Blood was collected by cardiac puncture and centrifuged (12,000 rpm/10 min). Plasma was separated and stored at −80°C for analysis of metabolic markers. Visceral fat pads (gonadal and retroperitoneal fat) and skeletal muscle (gastrocnemius) were dissected, weighed, snap-frozen in liquid nitrogen, and stored at −80°C freezer for further analysis.
4.7. Gut Microbial Profile
Total bacterial DNA was extracted from approximately 100 mg of mice faecal samples after ten weeks of UPF administration using QIAamp PowerFecal Pro DNA Kits (Cat# 51804, Qiagen, Japan), according to the manufacturer’s instructions and stored at −80°C for further processing. DNA concentration and purity ratios (A260/280 and A260/230) were measured by Nanodrop™ 8000 Spectrophotometer (NanoDrop Technologies Inc., Japan) and only samples with absorbance ratio of ~1.8 were used for quantitative polymerase chain reaction (qPCR) assays.
Specific primers for Firmicutes phylum, Bacteroidetes phylum, Enterobacteriales order, Bifidobacterium spp., Lactobacillus group, Bacteroides/Prevotella, Akkermansia muciniphila, Clostridial coccoides, and Total Bacteria were obtained from Sigma-Aldrich (Supplementary Table S3). The abundance of specific bacterial groups and species was determined by measuring DNA expression of the 16S rRNA gene sequences by qPCR with the CFX Connect TM Real-Time PCR Detection system (Bio-Rad), following previously described protocols [58,59].
The reactions were conducted with 2.5 μL DNA (2 ng/μL), 1.5 μL nucleus-free distilled water, 0.5 μL forward primer (10 μM), 0.5 μL reverse primer (10 μM), and 5 μL PowerUp SYBR Green Master Mix (Cat# A25918, Applied Biosystems) in a total volume of 10 μL. A total of 2.5 μL of nucleus-free distilled water (Cat# 10977015, Thermo Fisher Scientific) was used as a negative control in each run. Target bacterial DNA expression was normalized against Total Bacteria (housekeeping gene) using the average of the control group as a calibrator, as previously described [58]. Analysis was performed using the ΔΔCt method [60].
4.8. Real-Time Quantitative PCR (RT-qPCR) Assay
Frozen skeletal muscle tissue was used to extract and purify total RNA using RNeasy Mini Kit (Cat# 74106, Qiagen, Japan) according to the manufacturer’s recommendations. A NanodropTM 8000 Spectrophotometer (NanoDrop Technologies Inc., Japan) was used to measure the RNA concentration and purity ratios (A260/280 and A260/230) and only samples with absorbance ratio of ~2.0 were used for complementary DNA (cDNA) synthesis. One microgram of RNA template from each sample was reverse transcribed into cDNA using High-Capacity cDNA Reverse Transcription Kit (Cat# 4368814, Applied Biosystem, Waltham, MA, USA). All primers were purchased from Kicqstart SYBR Green Primers (Sigma-Aldrich, St. Louis, MO, USA). The sequence of the primers used for quantification of gene expression of COX2, COX4, MYH1, IGF1, PGC-1α, PPAR-γ, b-actin, and GAPDH can be found in Supplementary Table S4.
RT-qPCR reactions were performed using PowerUp SYBR green master mix (Cat# A25918, Applied Biosystems, Waltham, MA, USA) in the CFX Connect TM Real-Time PCR Detection system (Bio-Rad) following the manufacturer’s instructions. The amplification program included an initial denaturation step at 95 °C for 5 min, followed by 40 cycles of 10 s at 95 °C, 10 s at 60 °C, and 20 s at 72 °C. After each amplification, a melting curve study was conducted to validate the reliability of the results and product specificity. Target gene expression was normalized against β-actin and GAPDH housekeeping genes using the average of control group as a calibrator. Analysis was performed using the comparative ΔΔCt method [60].
4.9. Statistical Analysis
Statistical analysis was performed using GraphPad Prism version 8.3.0 for Windows (GraphPad Software, San Diego, California USA, www.graphpad.com) and results were expressed as mean ± SEM. Body weight change and weekly running distance were analysed by Two-way repeated measures ANOVA with fucoidan effect and time points as factors. The effects of fucoidan on final body weight, energy daily intake, visceral adipose fat, skeletal muscle, and gut microbial DNA expression levels were performed by One-way ANOVA. Data were then assessed by a post hoc analysis using Fisher’s least significant difference test (LSD) as appropriate. Results were considered statistically significant when p < 0.05.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Additional information is available in “Supplementary File”. Figure S1: Running Wheel; Figure S2: Running Distance analyser; Table S1: Absolute mass percentages of Undaria pinnatifida fucoidan (UPF) extract; Video S1: Effective Voluntary Oral Drug Administration in Mice: Drug-Containing/Vehicle Jelly; Table S2: Carbohydrate breakdown (mass %) of neutral carbohydrates in Undaria pinnatifida fucoidan (UPF) extract; Table S3: Sequence of primers used for RT-qPCR assay in mouse muscle samples; Table S4: Sequence of primers used for bacterial profiling by quantitative PCR; Table S5: Effects UPF, diet and exercise on energy intake and anthropometric parameters.
Author Contributions
Conceptualization, C.Y. and V.C.; methodology, C.Y. and V.C.; software, C.Y. and V.C.; validation, C.Y., C.D., B.W. and V.C.; formal analysis, C.Y., C.D., B.W., R.W., L.J. and V.C.; investigation, C.Y., C.D., B.W. and V.C.; resources, C.D., B.W. and V.C.; data curation, C.Y., C.D., B.W. and V.C.; writing original draft preparation, C.Y. and V.C.; writing review and editing, C.Y., C.D., B.W., R.W., L.J. and V.C.; visualization, C.Y. and V.C.; supervision, R.W, L.J. and V.C.; project administration, C.D., B.W. and V.C.; funding acquisition, V.C., C.D. and B.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Marinova Pty. Ltd.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) of Washington Biotechnology IACUC (Approval number A0027164 (A-94788) on 19 April 2023).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Conflicts of Interest
The current study was sponsored by Marinova Pty. Ltd. (Cambridge, TAS, Australia) C.D. and B.W. are employees of Marinova Pty. Ltd.
References
- Li B, Lu F, Wei X, Zhao R. Fucoidan: Structure and Bioactivity. Molecules. 2008;13(8):1671-95. [CrossRef]
- Fitton HJ, Stringer DS, Park AY, Karpiniec SN. Therapies from Fucoidan: New Developments. Mar Drugs. 2019;17(10). [CrossRef]
- Ahmad T, Eapen MS, Ishaq M, Park AY, Karpiniec SS, Stringer DN, et al. Anti-Inflammatory Activity of Fucoidan Extracts In Vitro. Mar Drugs. 2021;19(12). [CrossRef]
- Mathew L, Burney M, Gaikwad A, Nyshadham P, Nugent EK, Gonzalez A, et al. Preclinical Evaluation of Safety of Fucoidan Extracts From Undaria pinnatifida and Fucus vesiculosus for Use in Cancer Treatment. Integr Cancer Ther. 2017;16(4):572-84. [CrossRef]
- Fitton J, Dell’Acqua G, Gardiner V-A, Karpiniec S, Stringer D, Davis E. Topical Benefits of Two Fucoidan-Rich Extracts from Marine Macroalgae. Cosmetics. 2015;2(2):66-81. [CrossRef]
- Cox AJ, Cripps AW, Taylor PA, Fitton JH, West NP. Fucoidan Supplementation Restores Fecal Lysozyme Concentrations in High-Performance Athletes: A Pilot Study. Mar Drugs. 2020;18(8). [CrossRef]
- Irhimeh MR, Fitton JH, Lowenthal RM. Pilot clinical study to evaluate the anticoagulant activity of fucoidan. Blood Coagulation & Fibrinolysis. 2009;20(7):607-10. [CrossRef]
- Ahmad T, Ishaq M, Karpiniec S, Park A, Stringer D, Singh N, et al. Oral Macrocystis pyrifera Fucoidan Administration Exhibits Anti-Inflammatory and Antioxidant Properties and Improves DSS-Induced Colitis in C57BL/6J Mice. Pharmaceutics. 2022;14(11):2383. [CrossRef]
- Chen YM, Tsai YH, Tsai TY, Chiu YS, Wei L, Chen WC, et al. Fucoidan supplementation improves exercise performance and exhibits anti-fatigue action in mice. Nutrients. 2014;7(1):239-52. [CrossRef]
- McBean SE, Church JE, Thompson BK, Taylor CJ, Fitton JH, Stringer DN, et al. Oral fucoidan improves muscle size and strength in mice. Physiol Rep. 2021;9(3):e14730. [CrossRef]
- Ahn J, Ha TY, Ahn J, Jung CH, Seo HD, Kim MJ, et al. Undaria pinnatifida extract feeding increases exercise endurance and skeletal muscle mass by promoting oxidative muscle remodeling in mice. The FASEB Journal. 2020;34(6):8068-81. [CrossRef]
- McFadden BA, Vincenty CS, Chandler AJ, Cintineo HP, Lints BS, Mastrofini GF, et al. Effects of fucoidan supplementation on inflammatory and immune response after high-intensity exercise. J Int Soc Sports Nutr. 2023;20(1):2224751. [CrossRef]
- Antoni L, Nuding S, Weller D, Gersemann M, Ott G, Wehkamp J, et al. Human colonic mucus is a reservoir for antimicrobial peptides. J Crohns Colitis. 2013;7(12):e652-64. [CrossRef]
- Lopez-Santamarina A, Miranda JM, Mondragon ADC, Lamas A, Cardelle-Cobas A, Franco CM, et al. Potential Use of Marine Seaweeds as Prebiotics: A Review. Molecules. 2020;25(4). [CrossRef]
- Mailing LJ, Allen JM, Buford TW, Fields CJ, Woods JA. Exercise and the Gut Microbiome: A Review of the Evidence, Potential Mechanisms, and Implications for Human Health. Exerc Sport Sci Rev. 2019;47(2):75-85. [CrossRef]
- Clauss M, Gérard P, Mosca A, Leclerc M. Interplay Between Exercise and Gut Microbiome in the Context of Human Health and Performance. Front Nutr. 2021;8:637010. [CrossRef]
- Allen JM, Mailing LJ, Niemiro GM, Moore R, Cook MD, White BA, et al. Exercise Alters Gut Microbiota Composition and Function in Lean and Obese Humans. Med Sci Sports Exerc. 2018;50(4):747-57. [CrossRef]
- Liu M, Ma L, Chen Q, Zhang P, Chen C, Jia L, et al. Fucoidan alleviates dyslipidemia and modulates gut microbiota in high-fat diet-induced mice. Journal of Functional Foods. 2018;48:220-7. [CrossRef]
- Huang J, Huang J, Li Y, Lv H, Yin T, Fan S, et al. Fucoidan Protects Against High-Fat Diet-Induced Obesity and Modulates Gut Microbiota in Institute of Cancer Research Mice. J Med Food. 2021;24(10):1058-67. [CrossRef]
- Monda V, Villano I, Messina A, Valenzano A, Esposito T, Moscatelli F, et al. Exercise Modifies the Gut Microbiota with Positive Health Effects. Oxid Med Cell Longev. 2017;2017:3831972. [CrossRef]
- McLoughlin RF, Berthon BS, Jensen ME, Baines KJ, Wood LG. Short-chain fatty acids, prebiotics, synbiotics, and systemic inflammation: a systematic review and meta-analysis. The American Journal of Clinical Nutrition. 2017;106(3):930-45. [CrossRef]
- Meng D, Ai S, Spanos M, Shi X, Li G, Cretoiu D, et al. Exercise and microbiome: From big data to therapy. Comput Struct Biotechnol J. 2023;21:5434-45. [CrossRef]
- Magne F, Gotteland M, Gauthier L, Zazueta A, Pesoa S, Navarrete P, et al. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients. 2020;12(5). [CrossRef]
- Evans CC, LePard KJ, Kwak JW, Stancukas MC, Laskowski S, Dougherty J, et al. Exercise Prevents Weight Gain and Alters the Gut Microbiota in a Mouse Model of High Fat Diet-Induced Obesity. PLOS ONE. 2014;9(3):e92193. [CrossRef]
- Watson SA, McStay GP. Functions of Cytochrome c Oxidase Assembly Factors. International Journal of Molecular Sciences. 2020;21(19):7254. [CrossRef]
- Allen JM, Miller MEB, Pence BD, Whitlock K, Nehra V, Gaskins HR, et al. Voluntary and forced exercise differentially alters the gut microbiome in C57BL/6J mice. Journal of Applied Physiology. 2015;118(8):1059-66. [CrossRef]
- Aykut MN, Erdoğan EN, Çelik MN, Gürbüz M. An Updated View of the Effect of Probiotic Supplement on Sports Performance: A Detailed Review. Current Nutrition Reports. 2024. [CrossRef]
- Alvehus M, Boman N, Söderlund K, Svensson MB, Burén J. Metabolic adaptations in skeletal muscle, adipose tissue, and whole-body oxidative capacity in response to resistance training. European Journal of Applied Physiology. 2014;114(7):1463-71. [CrossRef]
- Gómez-Pérez Y, Capllonch-Amer G, Gianotti M, Lladó I, Proenza AM. Long-term high-fat-diet feeding induces skeletal muscle mitochondrial biogenesis in rats in a sex-dependent and muscle-type specific manner. Nutrition & Metabolism. 2012;9(1):15. [CrossRef]
- Čunátová K, Reguera DP, Houštěk J, Mráček T, Pecina P. Role of cytochrome c oxidase nuclear-encoded subunits in health and disease. Physiol Res. 2020;69(6):947-65. [CrossRef]
- Kadenbach, B. Complex IV—The regulatory center of mitochondrial oxidative phosphorylation. Mitochondrion. 2021;58:296-302. [CrossRef]
- Pajuelo Reguera D, Čunátová K, Vrbacký M, Pecinová A, Houštěk J, Mráček T, et al. Cytochrome c Oxidase Subunit 4 Isoform Exchange Results in Modulation of Oxygen Affinity. Cells. 2020;9(2):443. [CrossRef]
- Zhang L, Zhou Y, Wu W, Hou L, Chen H, Zuo B, et al. Skeletal Muscle-Specific Overexpression of PGC-1α Induces Fiber-Type Conversion through Enhanced Mitochondrial Respiration and Fatty Acid Oxidation in Mice and Pigs. International Journal of Biological Sciences. 2017;13(9):1152-62. [CrossRef]
- Sellers RS, Mahmood SR, Perumal GS, Macaluso FP, Kurland IJ. Phenotypic Modulation of Skeletal Muscle Fibers in LPIN1-Deficient Lipodystrophic (fld) Mice. Veterinary Pathology. 2019;56(2):322-31. [CrossRef]
- Hancock CR, Han DH, Chen M, Terada S, Yasuda T, Wright DC, et al. High-fat diets cause insulin resistance despite an increase in muscle mitochondria. Proc Natl Acad Sci U S A. 2008;105(22):7815-20. [CrossRef]
- Yoshida T, Delafontaine P. Mechanisms of IGF-1-Mediated Regulation of Skeletal Muscle Hypertrophy and Atrophy. Cells. 2020;9(9):1970. [CrossRef]
- Wegierska AE, Charitos IA, Topi S, Potenza MA, Montagnani M, Santacroce L. The Connection Between Physical Exercise and Gut Microbiota: Implications for Competitive Sports Athletes. Sports Med. 2022;52(10):2355-69. [CrossRef]
- Cerdá B, Pérez M, Pérez-Santiago JD, Tornero-Aguilera JF, González-Soltero R, Larrosa M. Gut Microbiota Modification: Another Piece in the Puzzle of the Benefits of Physical Exercise in Health? Frontiers in Physiology. 2016;7. [CrossRef]
- Koutouratsas T, Philippou A, Kolios G, Koutsilieris M, Gazouli M. Role of exercise in preventing and restoring gut dysbiosis in patients with inflammatory bowel diseases: A review. World J Gastroenterol. 2021;27(30):5037-46. [CrossRef]
- Abenavoli L, Scarpellini E, Colica C, Boccuto L, Salehi B, Sharifi-Rad J, et al. Gut Microbiota and Obesity: A Role for Probiotics. Nutrients. 2019;11(11). [CrossRef]
- Belzer C, de Vos WM. Microbes inside--from diversity to function: the case of Akkermansia. Isme j. 2012;6(8):1449-58. [CrossRef]
- Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci U S A. 2013;110(22):9066-71. [CrossRef]
- Shang Q, Song G, Zhang M, Shi J, Xu C, Hao J, et al. Dietary fucoidan improves metabolic syndrome in association with increased Akkermansia population in the gut microbiota of high-fat diet-fed mice. Journal of Functional Foods. 2017;28:138-46. [CrossRef]
- Kim NY, Kim JM, Son JY, Ra CH. Synbiotic Fermentation of Undaria pinnatifida and Lactobacillus brevis to Produce Prebiotics and Probiotics. Appl Biochem Biotechnol. 2023;195(10):6321-33. [CrossRef]
- Li H, Dong T, Tao M, Zhao H, Lan T, Yan S, et al. Fucoidan enhances the anti-tumor effect of anti-PD-1 immunotherapy by regulating gut microbiota. Food & Function. 2024;15(7):3463-78. [CrossRef]
- Baenas I, Camacho-Barcia L, Miranda-Olivos R, Solé-Morata N, Misiolek A, Jiménez-Murcia S, et al. Probiotic and prebiotic interventions in eating disorders: A narrative review. European Eating Disorders Review. 2024. [CrossRef]
- Dempsey E, Corr SC. Lactobacillus spp. for Gastrointestinal Health: Current and Future Perspectives. Front Immunol. 2022;13:840245. [CrossRef]
- Sonnenburg ED, Zheng H, Joglekar P, Higginbottom SK, Firbank SJ, Bolam DN, et al. Specificity of polysaccharide use in intestinal bacteroides species determines diet-induced microbiota alterations. Cell. 2010;141(7):1241-52. [CrossRef]
- Rinninella E, Raoul P, Cintoni M, Franceschi F, Miggiano G, Gasbarrini A, et al. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms. 2019;7(1):14. [CrossRef]
- Takezawa K, Fujita K, Matsushita M, Motooka D, Hatano K, Banno E, et al. The Firmicutes/Bacteroidetes ratio of the human gut microbiota is associated with prostate enlargement. The Prostate. 2021;81(16):1287-93. [CrossRef]
- Stojanov S, Berlec A, Štrukelj B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms. 2020;8(11). [CrossRef]
- An J, Kwon H, Kim YJ. The Firmicutes/Bacteroidetes Ratio as a Risk Factor of Breast Cancer. J Clin Med. 2023;12(6). [CrossRef]
- Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. Obesity alters gut microbial ecology. Proc Natl Acad Sci U S A. 2005;102(31):11070-5. [CrossRef]
- Mujico JR, Baccan GC, Gheorghe A, Díaz LE, Marcos A. Changes in gut microbiota due to supplemented fatty acids in diet-induced obese mice. Br J Nutr. 2013;110(4):711-20. [CrossRef]
- Fitton JH, Stringer DN, Karpiniec SS. Therapies from Fucoidan: An Update. Mar Drugs. 2015;13(9):5920-46. [CrossRef]
- Health N, Council MR. Australian code of practice for the care and use of animals for scientific purposes: Australian Government Publishing Service; 1997. [CrossRef]
- Zhang, L. Method for voluntary oral administration of drugs in mice. STAR Protoc. 2021;2(1):100330. [CrossRef]
- Chaplin A, Parra P, Serra F, Palou A. Conjugated Linoleic Acid Supplementation under a High-Fat Diet Modulates Stomach Protein Expression and Intestinal Microbiota in Adult Mice. PLoS One. 2015;10(4):e0125091. [CrossRef]
- Mujico JR, Baccan GC, Gheorghe A, Díaz LE, Marcos A. Changes in gut microbiota due to supplemented fatty acids in diet-induced obese mice. British Journal of Nutrition. 2013;110(4):711-20. [CrossRef]
- Pfaffl, MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29(9):e45. [CrossRef]
Figure 1.
Effects of UPF on running activity and muscle mass. (A) Changes in running activity of mice consuming standard chow diet (CHOW) (black round, n=10), and those treated with UPF (open round, n=10). (B) Changes in running activity of mice consuming HFD (black square, n=10), and those treated with UPF (open square, n=10). (C) Daily running activity (n=10). (D) Muscle mass of mice at killing calculated as percentage of wet weight to body weight (n=15-16). Results are expressed as mean ± SEM. In graphs (A, B) data were analysed by Repeated Measures two-way ANOVA with UPF treatment and weeks of treatment as factors. In graphs (C, D) data were analysed by one-way ANOVA followed by a post hoc assessment using Fisher’s least significant difference test (LSD). #, significant difference for overall diet effect (CHOW and CHOW + UPF) vs. (HFD and HFD + UPF) (p < 0.05); *, significant differ-ence for UPF effect (p < 0.05).
Figure 1.
Effects of UPF on running activity and muscle mass. (A) Changes in running activity of mice consuming standard chow diet (CHOW) (black round, n=10), and those treated with UPF (open round, n=10). (B) Changes in running activity of mice consuming HFD (black square, n=10), and those treated with UPF (open square, n=10). (C) Daily running activity (n=10). (D) Muscle mass of mice at killing calculated as percentage of wet weight to body weight (n=15-16). Results are expressed as mean ± SEM. In graphs (A, B) data were analysed by Repeated Measures two-way ANOVA with UPF treatment and weeks of treatment as factors. In graphs (C, D) data were analysed by one-way ANOVA followed by a post hoc assessment using Fisher’s least significant difference test (LSD). #, significant difference for overall diet effect (CHOW and CHOW + UPF) vs. (HFD and HFD + UPF) (p < 0.05); *, significant differ-ence for UPF effect (p < 0.05).
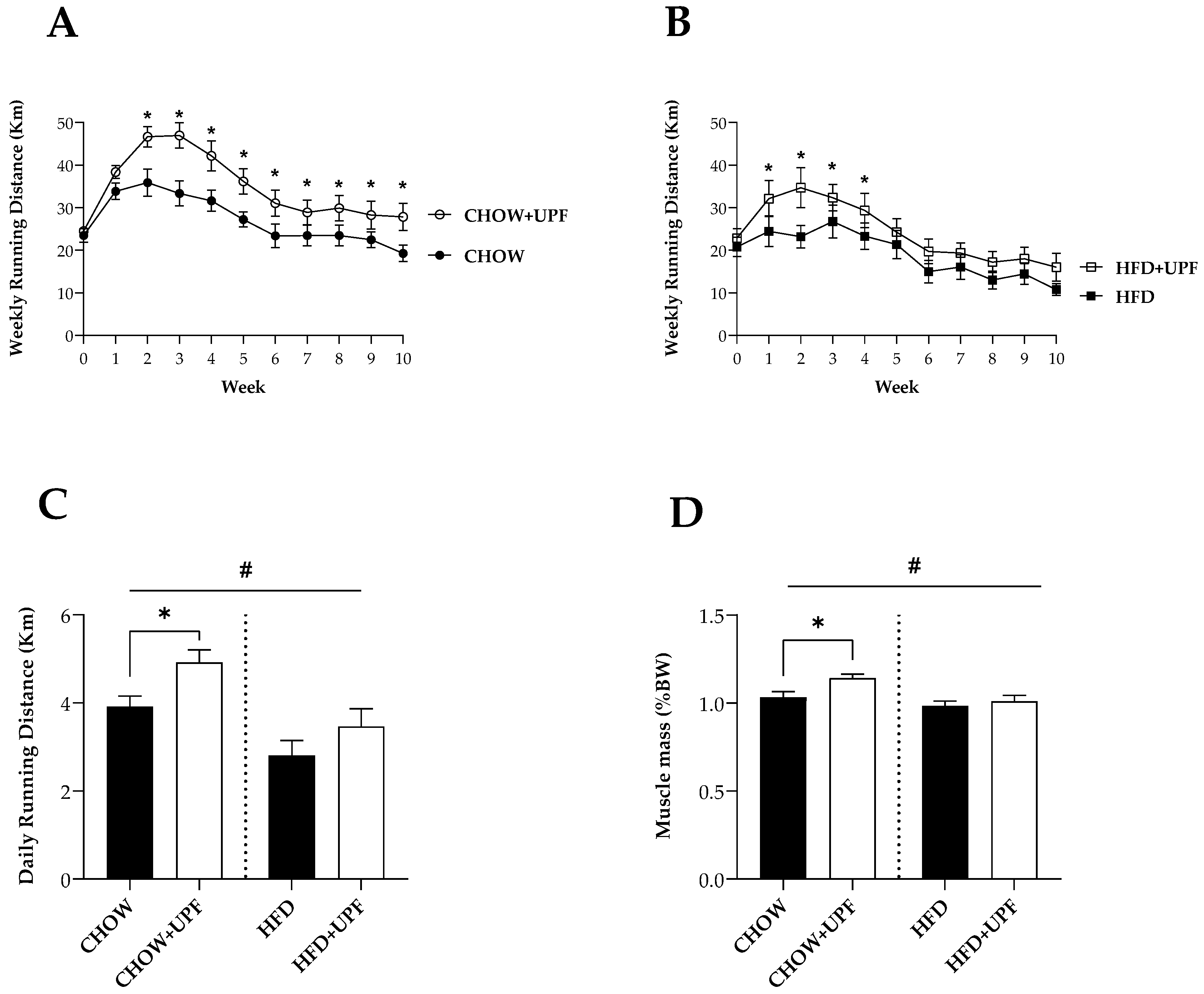
Figure 2.
Total DNA content and abundance of representative bacterial groups and species in mice faeces. Total DNA content is expressed as ng DNA per mg of faeces. (A) Total DNA content in the faeces (n=15-16) after ten weeks of UPF administration. DNA abundance of 16S rRNA gene of representative bacterial groups/species (B) Bacteroides/Prevotella, (C) Akkermansia muciniphila, (D) Lactobacillus group, (E) Bacteroidetes phylum, (F) Clostridial coccoides, and (G) Firmicutes phylum were analysed in mouse faeces and normalised with the average of the CHOW group (2−ΔΔCt). Bacterial quantities are expressed as relative units. And (H) the ratio of Firmicutes to Bacteroidetes was calculated by dividing the relative DNA expression value of Firmicutes by the relative DNA expression value of Bacteroidetes. n = 7-10. Results are presented as mean ± SEM. In the graphs (A-H) data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between CHOW vs CHOW+UPF and HFD vs HFD+UPF.
Figure 2.
Total DNA content and abundance of representative bacterial groups and species in mice faeces. Total DNA content is expressed as ng DNA per mg of faeces. (A) Total DNA content in the faeces (n=15-16) after ten weeks of UPF administration. DNA abundance of 16S rRNA gene of representative bacterial groups/species (B) Bacteroides/Prevotella, (C) Akkermansia muciniphila, (D) Lactobacillus group, (E) Bacteroidetes phylum, (F) Clostridial coccoides, and (G) Firmicutes phylum were analysed in mouse faeces and normalised with the average of the CHOW group (2−ΔΔCt). Bacterial quantities are expressed as relative units. And (H) the ratio of Firmicutes to Bacteroidetes was calculated by dividing the relative DNA expression value of Firmicutes by the relative DNA expression value of Bacteroidetes. n = 7-10. Results are presented as mean ± SEM. In the graphs (A-H) data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between CHOW vs CHOW+UPF and HFD vs HFD+UPF.
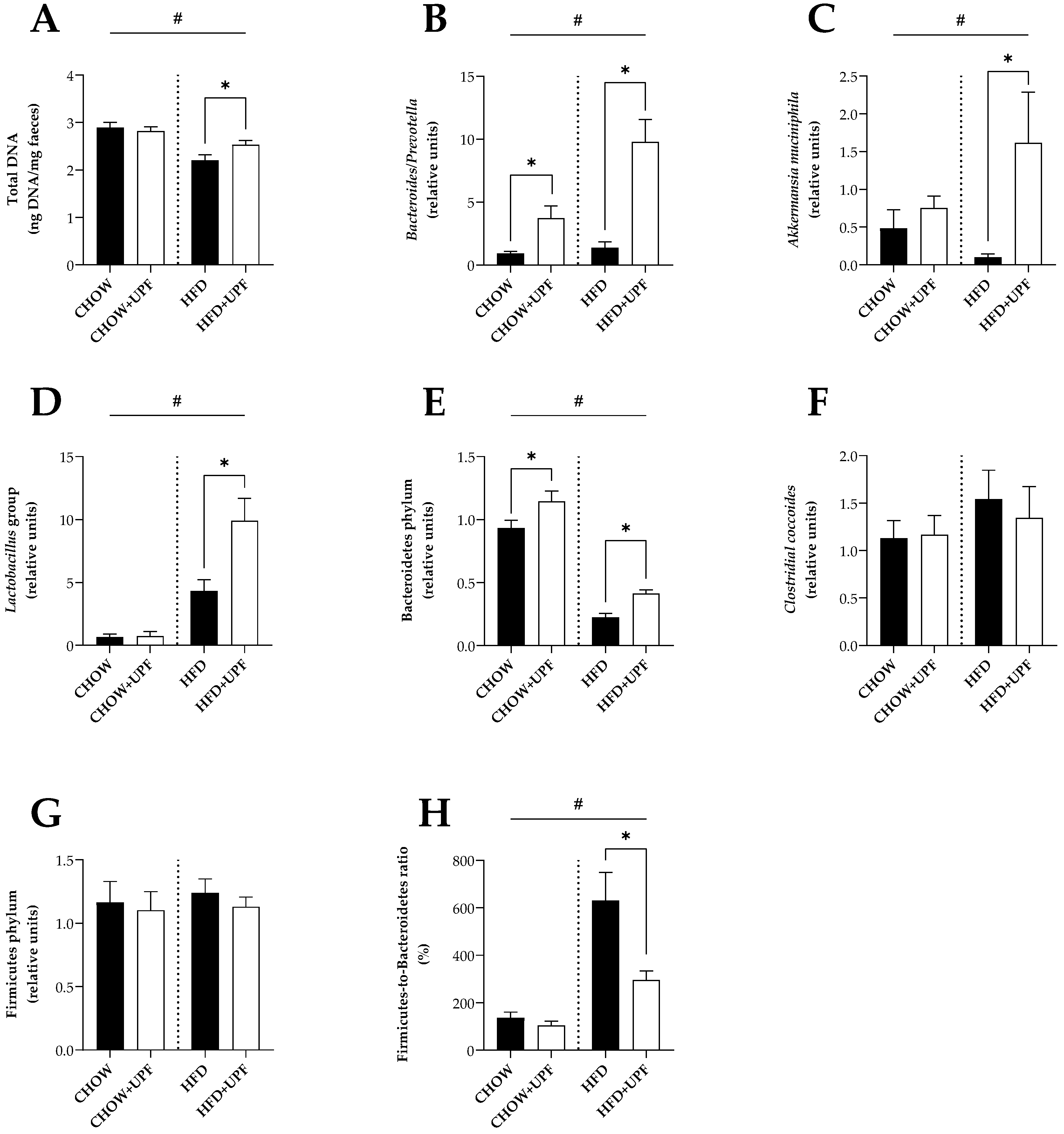
Figure 3.
Effects of UPF on muscle gene expression. mRNA levels of (A) COX2, (B) COX4, (C) MYH1, (D) PGC-1α, (E) PPAR-γ, and (F) IGF1. n = 13-16. Results are expressed as mean ± SEM. Data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between CHOW vs CHOW+UPF and HFD vs HFD+UPF.
Figure 3.
Effects of UPF on muscle gene expression. mRNA levels of (A) COX2, (B) COX4, (C) MYH1, (D) PGC-1α, (E) PPAR-γ, and (F) IGF1. n = 13-16. Results are expressed as mean ± SEM. Data were analysed by one-way ANOVA, followed by post hoc LSD tests. #, p < 0.05, overall difference between (CHOW and CHOW+UPF) vs (HFD and HFD+UPF). *, p < 0.05, difference between CHOW vs CHOW+UPF and HFD vs HFD+UPF.
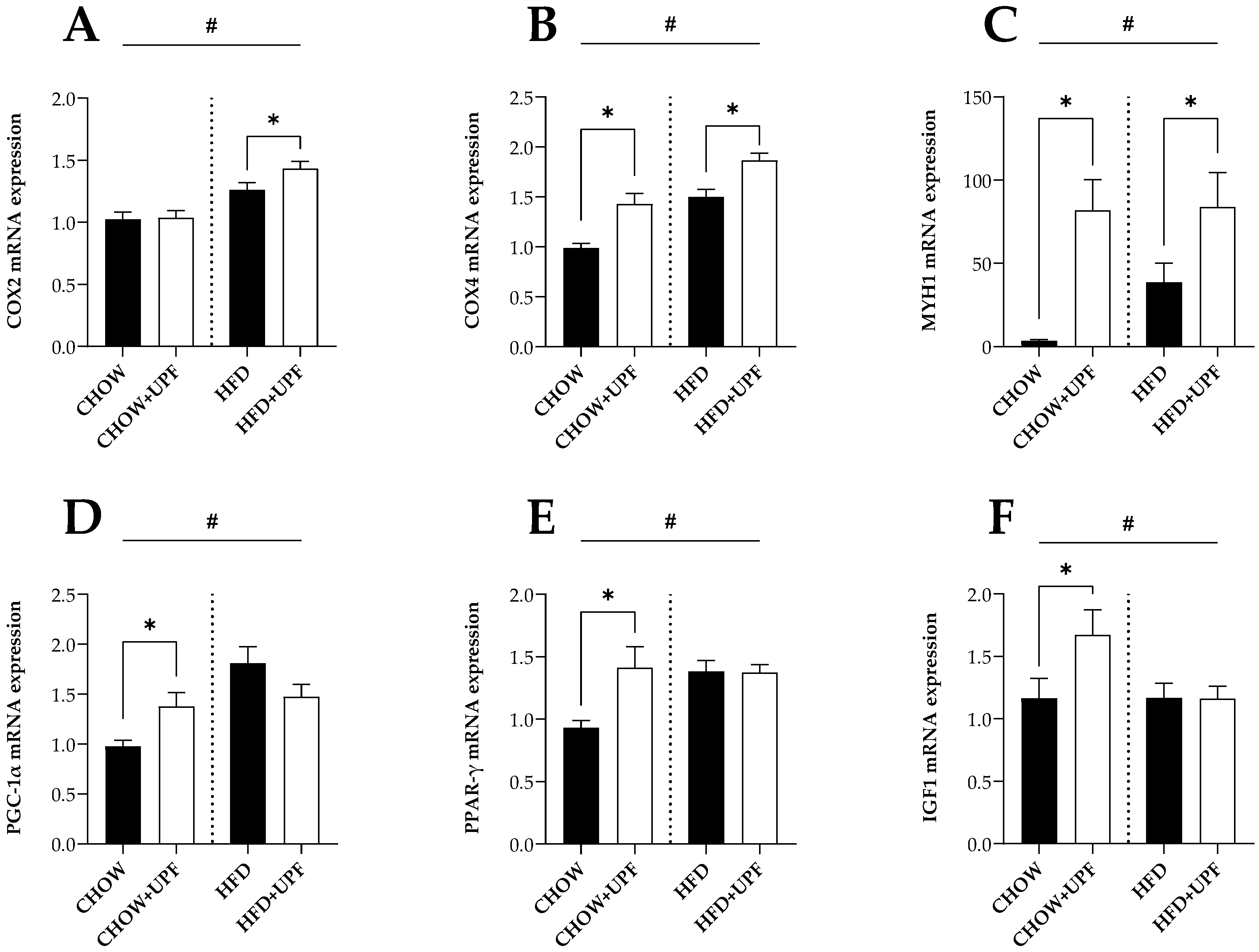

Table 1.
Correlations between the abundance of bacterial groups/species in faeces with mouse body weight, visceral fat, and running distance.
Table 1.
Correlations between the abundance of bacterial groups/species in faeces with mouse body weight, visceral fat, and running distance.
| Bacterial Groups/species | Body Weight | Visceral Fat | Running Distance | ||||||
|---|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | ||||
| Bacteroidetes phylum | - | 0.708 ** | 0.0001 | - | 0.779 ** | 0.0001 | 0.598 ** | 0.0001 | |
| Firmicutes phylum | 0.136 | 0.4022 | 0.111 | 0.4950 | - | 0.261 | 0.1035 | ||
| Clostridial coccoides | 0.107 | 0.5362 | 0.217 | 0.2039 | - | 0.430 ** | 0.0089 | ||
| Akkermansia muciniphila | - | 0.399 * | 0.0354 | - | 0.140 | 0.4756 | 0.420 * | 0.0259 | |
| F/B ratio | 0.568 ** | 0.0001 | 0.593 ** | 0.0001 | - | 0.559 ** | 0.0002 | ||
Linear relationships were tested using Pearson’s correlation coefficients (r). Significant correlations are marked as fellow: *, p < 0.05, **, p < 0.01. F/B ratio: Firmicutes to Bacteroidetes ratio. Visceral fat: gonadal and retroperitoneal fat pads.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



