
Submitted:
20 September 2024
Posted:
20 September 2024
You are already at the latest version
Abstract
Antimicrobial resistance (AMR) is a worsening global health crisis that has arisen due to antibiotic overuse and misuse. Antibiotic resistance renders standard treatments less effective, making it difficult to control some infections, thereby increasing morbidity and mortality. Medicinal plants are sustainable treatment options that are becoming increasingly important as antibiotics lose their efficacies. The present study evaluates the antibacterial activity of aqueous, methanolic and ethyl acetate extracts prepared using Terminalia bellirica and Terminalia chebula fruit against six bacterial pathogens using disc diffusion and broth microdilution assays. The aqueous and methanol extracts of T. bellirica and T. chebula showed substantial zones of inhibition (ZOIs) against S. aureus and MRSA. The activity against those bacteria was determined to be strong, with minimum inhibitory concentrations (MIC) ranging from 94 µg/mL to 392 µg/mL. Additionally, the T. bellirica methanolic extract showed noteworthy antibacterial activity against E. coli and an ESBL E. coli strain (MIC values of 755 µg/mL for both). The aqueous T. bellirica and T. chebula extracts also inhibited K. pneumoniae growth (MIC values of 784 µg/mL and 556 µg/mL respectively), whilst the methanolic extracts of these plants also inhibited ESBL K. pneumoniae growth (MIC values of 755 µg/mL and 1509 µg/mL respectively). Eighteen additive interactions were observed when extracts were combined with reference antibiotics. Notably, strong antagonistic interactions occurred when any of the extracts were mixed with polymyxin B. All extracts were subjected to LC-MS analysis, revealing several interesting flavonoids and tannins, including 6-galloylglucose, 1,2,6-trigalloyl-β-D-glucopyranose, 6-O-[(2E)-3-phenyl-2-propenoyl]-1-O-(3,4,5-trihydroxybenzoyl)-β-D-glucopyranose, propyl gallate, methyl gallate, sanguiin H4, hamamelitannin, pyrogallol, gallic acid, ellagic acid, chebulic acid, and chebuloside II. All extracts were nontoxic when tested in brine shrimp assays. This lack of toxicity, combined with their antibacterial activities suggests that these plant species may be promising sources of antibacterial compound(s) that warrant further study.
Keywords:
MRSA
; ESBL
; plant extracts
; natural therapies
; combinational therapies
; metabolomics
1. Introduction
Antimicrobial resistance (AMR) is one of the most concerning public health issues of the 21st century. The development of AMR mechanisms in bacterial pathogens leads to lower efficacies for standard antimicrobial therapies, which in turn promotes the transmission of these infections. The World Health Organisation (WHO) has highlighted antimicrobial resistance (AMR) as a major danger to global health, food security, and future development, with far-reaching effects on human health and economic stability [1]. Antibiotic resistant bacterial strains have rapidly arisen due to antibiotic misuse and overuse in humans and animals, as well as in agriculture. Antibiotic misuse includes excessive antibiotic prescriptions, poor treatment compliance, and the widespread use of antibiotics in livestock for growth promotion and disease prevention. This results in longer illness durations, higher mortality rates, and elevated healthcare costs [1]. AMR mortality and morbidity are concerning. AMR caused 1.27 million deaths worldwide in 2019, and current trends indicate that this number will climb dramatically [1]. Infections caused by antibiotic-resistant bacteria, such as methicillin-resistant Staphylococcus aureus (MRSA) and multidrug-resistant Escherichia coli, are particularly concerning due to their high resistance rates and severe clinical outcomes [2]. The costs associated with AMR are multifaceted, encompassing direct medical costs such as longer hospital stays and more intensive care, as well as indirect costs such as lost productivity and long-term disability. According to the World Bank, by 2050 AMR is likely to cause a loss of global economic output amounting to USD 1 trillion annually, with healthcare costs also projected to increase significantly [3].
The 2022 Global Antimicrobial Resistance and Use Surveillance System (GLASS) report underscores the significant concern regarding the alarming resistance rates of third-generation cephalosporin-resistant E. coli and MRSA in 76 countries [1,4]. That report also found a significant rise in E. coli infections resistant to ampicillin, co-trimoxazole, and fluoroquinolones. The common intestinal bacterium Klebsiella pneumoniae has also shown increased resistance to penicillins, cephalosporins and fluoroquinolones. These elevated levels of antibiotic resistance may lead to an increase in the use of last-resort medications, such as carbapenems [1,4]. The rise of antibiotic-resistant bacteria has outpaced drug development. The high expenditures and limited financial returns of antibiotic research and development have caused a sharp fall in the number of novel antibiotics approved for clinical use [5]. According to the WHO, there is a concerning lack of new antibacterials in the pipeline due to the lengthy research and development processes, and the high probability of failure during clinical testing [6]. Thirteen new antibiotics have received therapeutic authorisation since July 2017. However, only two of these antibiotics constitute a new chemical class and are considered innovative [6]. This highlights the significant scientific and technical difficulties in finding new antibacterials that are both effective against bacteria and safe for human use.
Exploring the potential of medicinal plants in the development of antimicrobial drugs presents numerous benefits compared to conventional antibiotics. Traditional medicinal plants offer an accessible and cost-effective alternative, requiring fewer resources and less time for production [7]. They often interact with the body in a safe manner, causing minimal side effects, and often have complementary effects that enhance overall well-being. However, medicinal plants can also cause toxicity if misused. Therefore, careful dosing, accurate identification, and monitoring are essential to avoid adverse effects. Phytochemicals in medicinal plants may work together to boost antibacterial efficacy, potentiating the treatment’s efficacy [7,8]. Medicinal plants possess a variety of mechanisms of action, which holds great promise in combating antibiotic-resistant bacteria [8]. The combination of plant extracts/compounds with conventional antibiotics is a strategy that can be utilised in the development of new antimicrobial therapies [9]. Phytochemicals may enhance antibiotic efficacies through synergistic and additive interactions, potentially overcoming resistance issues. This approach offers a promising solution for developing effective treatments against antibiotic-resistant bacterial strains. For example, propolis ethanolic extract potentiates the efficacy of ampicillin and vancomycin against the methicillin resistant S. aureus (MRSA) [10].
The current study focuses on the antibacterial activity, phytochemistry, and toxicity of fruit extracts (aqueous, methanol, and ethyl acetate) derived from Terminalia bellirica (Gaertn.) Roxb. and Terminalia chebula Retz. Antibiotic sensitive and resistant bacterial pairs of S. aureus, E. coli, and K. pneumoniae were examined for their growth in the presence of the extracts, as well as a panel of reference antibiotics. Our previous work revealed enhancements of E. coli antibacterial activity when T. bellirica and T. chebula fruit extracts were combined [11]. However, antibiotic-resistant bacterial species were not tested in that study, nor were combinational interactions between pure antibiotics and the plant extracts examined. Thakur et al. (2024) recently identified good antibacterial activities of the T. bellirica aqueous fruit extract against S. aureus, E. coli, and E. coli 385 (an antibiotic-resistant strain), with minimum inhibitory concentrations (MICs) of 312 µg/mL, 1250 µg/mL and 625 µg/mL against these species, respectively [12]. Phytochemical analysis was conducted by liquid chromatography–mass spectrometry (LC-MS), which identified a single polyphenol compound, bergenin [12]. Interestingly, flavonoids, tannins and terpenoids were not identified in the T. bellirica fruit extracts in that study. Furthermore, combinational interactions between reference antibiotics and the extracts were not explored. In addition, prior studies have demonstrated that fruit extracts of T. bellirica exhibit antibacterial activity against S. aureus, E. coli, and K. pneumoniae [13,14,15]. However, those investigations did not include combinational studies of extracts with antibiotics and testing of antibiotic-resistant bacterial strains. Other investigations reported inconsistent results for T. chebula fruit extracts (aqueous, ethanol, and dimethylformamide) against S. aureus and K. pneumoniae, with ZOIs of 15-18 mm on agar, using 500 µg extract per disc [16]. Furthermore, low to inactive antibacterial activities of T. chebula fruit extracts were observed against antibiotic sensitive and resistant strains of S. aureus and E. coli [17,18].
In order to address the discrepancies in the previous studies regarding the activities of T. bellirica and T. chebula, and the absence of thorough phytochemical analyses of the extracts, we investigated the antibacterial properties of aqueous, methanolic, and ethyl acetate fruit extracts obtained from T. bellirica and T. chebula. Our study focused on selected bacterial pathogens, but included resistant strains of S. aureus, E. coli, and K. pneumoniae. We conducted disc diffusion and liquid microdilution assays to assess their effectiveness. Additionally, LC-MS was utilised to examine the phytochemical composition of T. bellirica and T. chebula and to identify specific flavonoid, phenolic acid, terpenoid, and tannin molecules. In addition, our study investigated the interactions between active plant extracts and a panel of reference antibiotics when tested as extract-antibiotic combinations in MIC assays. Finally, the toxicities of the extracts were assessed using Artemia franciscana Kellogg nauplii lethality assays to provide initial insights into the safety of the extracts for possible clinical use.
2. Results
2.1. Antimicrobial Susceptibility Assays
Individual 1 g masses of T. bellirica and T. chebula fruit powder was extracted separately using sterile deionised water, methanol, or ethyl acetate. The extracts were subsequently dried and resuspended in 10 mL of 1% DMSO. The extract concentrations for the two plants were generally similar, where the highest yields were obtained using methanol as a solvent, whilst ethyl acetate produced very low yields for each plant. Specifically, T. bellirica extract yields for water, methanol and ethyl acetate solvent extractants were 50.2, 48.3, and 3.6 mg/mL respectively, whilst for T. chebula they were 35.6, 48.3, and 4.9 mg/mL.
The antibacterial efficacies of all extracts and reference antibiotics were evaluated using agar disc diffusion assays and broth microdilution assays. The aqueous and methanolic extracts of both plants exhibited activities varying from low to high against the six bacterial pathogens tested in the disc diffusion and broth dilution assays (Figure 1 and Table 1, respectively). The broth microdilution assays were used to quantify MIC values of the extracts and reference antibiotics (Table 1). The differences between the antibiotic-sensitive and resistant strains of the pathogens can be readily observed by the generally higher MIC values for many of the antibiotics against the resistant bacteria compared to their sensitive counterparts, as well as the lower ZOIs resulting from agar disc diffusion assays. This demonstrates that the antibiotic-resistant strains are much less sensitive to many of the antibiotics tested in this study.
When aqueous and methanol extracts of T. bellirica were evaluated against S. aureus and MRSA, they exhibited MIC values ranging from 94 and 392 µg/mL, respectively. The T. bellirica ethyl acetate extract also inhibited S. aureus and MRSA growth, albeit with a higher MIC value of 900 µg/mL. In contrast, all T. chebula extracts exhibited good antibacterial activity against S. aureus and MRSA, with MIC values ranging from 139 µg/mL to 306 µg/mL. Concordant findings were obtained for both plant extracts against S. aureus and MRSA in agar disc diffusion assays, where only small but detectable ZOIs were observed for the T. bellirica ethyl acetate extracts against the two Staphylococcus species, although much larger ZOIs of 9-15 mm were produced by all other extracts against S. aureus and MRSA (Figure 1).
Similarly, the extracts were also screened against E. coli and K. pneumoniae, as well as their antibiotic-resistant ESBL counterparts. The agar disc diffusion experiments showed limited growth inhibition for most of the extracts when tested against E. coli, ESBL E. coli, K. pneumoniae, and ESBL K. pneumoniae. However, these results cannot be considered as zones of inhibition (ZOIs) because complete inhibition surrounding the disc is necessary. Interestingly, the T. chebula ethyl acetate extract produced a small but distinct ZOI against E. coli and K. pneumoniae on agar (7-9 mm) in the disc diffusion assay (Figure 1). Furthermore, relatively high MIC values were determined for the T. bellirica aqueous extracts and the T. chebula aqueous and methanol extracts against E. coli and ESBL E. coli (MIC = 1509 µg/mL to 3138 µg/mL). Notably, an MIC value of 755 µg/mL was obtained for the T. bellirica methanolic extract against these two species. The aqueous T. bellirica and T. chebula extracts also exhibited noteworthy antibacterial activity against K. pneumoniae, with MIC values of 784 µg/mL and 556 µg/mL respectively. Moderate activities towards this bacterium were found when the T. bellirica and T. chebula methanolic extracts were tested (MIC = 1509 µg/mL), as well as the ethyl acetate T. chebula extract (MIC = 1225 µg/mL). Additionally, the ESBL K. pneumoniae was inhibited in broth assays by the methanolic T. bellirica and T. chebula extracts (MIC values of 755 µg/mL and 1509 µg/mL, respectively). However, the aqueous extracts yielded low activities (relatively high MIC values) against ESBL K. pneumoniae (MIC values of 3138 µg/mL and 2225 µg/mL respectively).
2.2. Combination Assays: Sum of Fractional Inhibitory Concentration (ƩFIC) Determinations
Combinations of the T. bellirica and T. chebula extracts with conventional antibiotics were tested to detect any possible interactions between the two components against the antibiotic-sensitive and resistant bacterial strains (Table 2). For these assays to be conducted, both the extract and the antibiotic must demonstrate efficacy against the bacterial species under examination, as MIC values for both components are required to determine ƩFIC values. No synergistic interactions were observed for any of the combinations. Eighteen combinations caused additive effects, and eighty combinations were indifferent. Additionally, nineteen combinations had antagonistic effects.
2.3. Compound Identification by LC-MS Metabolomics Fingerprinting
LC-MS fingerprinting analysis was employed to investigate the metabolomic fingerprints of all extracts, with particular focus on the flavonoid, tannin, and terpenoid compounds. Most of the extract compounds eluted in the gradient stage of the chromatogram 30% to 90% acetonitrile (Supplementary Materials Figures S1 and S2), suggesting that most of the extract components were of relatively high polarity [19]. Organic acids and amines are more likely to elute in polar environments and therefore do so early in the chromatogram. Conversely, lipophilic substances and hydrocarbons, which exhibit a stronger interaction with the non-polar stationary phase, elute later as the gradient persists. Only compounds that were completely matched in any of the databases (and were not present in the blank controls) were chosen to create a potential inventory of all the identified compounds (Supplementary Materials: Table S1 and S2). In this study, we have focussed on the flavonoid, tannin, terpenoid and phenolic acid compounds (Table 3).
2.4. Toxicity Quantification
The plant extracts were evaluated for toxicity in triplicate assays conducted on 48-well plates using Artemia franciscana Kellogg nauplii lethality assays (ALA). The toxicity level was determined, and plant extracts were classified as toxic if their LC50 values were less than 1000 µg/mL following 24-hour exposure [11]. More than half of the nauplii evaluated in this study remained alive for all extracts after 24-hour duration of incubation. Consequently, all plant extracts tested were classified as nontoxic. Indeed, the outcomes of all extracts were comparable to those of the negative control (artificial seawater).
3. Discussion
Our study tested T. bellirica and T. chebula fruit extracts against a group of bacterial pathogens that are significant for human health, including strains that are resistant to antibiotics. Significantly, the aqueous and methanolic extracts effectively suppressed the growth of the six bacterial pathogens examined in this study, underscoring their potential as targets for plant-based antibiotics development. The methanolic extracts exhibited the highest antibacterial efficacy in both the disc diffusion and liquid microdilution experiments against all pathogens. The ethyl acetate T. bellirica and T. chebula extracts also exhibited good antibacterial activities against S. aureus and MRSA in the disc diffusion and broth microdilution assays. In addition, the T. chebula ethyl acetate extract showed moderate antibacterial efficacy against K. pneumoniae. The differences in net yields and the populations of phytochemicals extracted by different solvents may account for these variances in potency. Methanol and water have a higher polarity compared to ethyl acetate, which results in the extraction of a larger quantity of phytochemicals with high to mid-polarity. In contrast, ethyl acetate extracts a smaller number of compounds with mid to lower polarity [20]. The differences in phytochemical contents among the extracts may potentially contribute to the varied antibacterial growth inhibition effects observed in the disc diffusion and liquid dilution investigations. Phytochemicals that have lower polar or are larger in size diffuse less rapidly through solid agar, resulting in a decrease in their apparent antibacterial effectiveness in disc diffusion assays [21]. The solubility of these phytochemicals in broth is also influenced by their polarity [22], which in turn impacts their ability to dissolve and may lead to inaccurate MIC values. Prior research has demonstrated that the depth of agar in petri dishes, as well as the regularity of the agar, may impact the extent of zones of inhibition (ZOIs) in agar diffusion studies [23]. Whilst we followed manufacturers’ instructions to create agar of uniform consistency and poured it at a consistent depth of 4 mm, repeating the disc diffusion experiments at varying agar depths may be useful to confirm the ZOIs of all the extracts and antibiotics. In this investigation, we noticed that the borders of the zone of inhibition (ZOI) were clearly visible for S. aureus and MRSA, although they were less clear (regarded as no ZOI) for the other bacterial pathogens studied. It is important to note that in future research, the use of methylene blue or crystal violet to stain the plates may enhance the visibility of ZOIs if there is low clarity [24].
Notably, the MRSA strain tested herein showed resistance to many commonly used antibiotics, such as β-lactams (penicillin G, oxacillin, amoxicillin), and macrolides (erythromycin). Nevertheless, the emergence of extend spectrum β-lactamase enzymes has made these medications less effective against some bacterial strains. Likewise, macrolide antibiotics such as erythromycin are commonly employed because they have the ability to impede protein synthesis in bacteria [25]. The existence of MRSA resistance to macrolides complicates the availability of treatment options, highlighting the need for new therapeutic drugs and procedures. Hence, it is imperative to identify innovative compounds that can elude or surmount these resistance mechanisms. The antibacterial activities observed with T. bellirica and T. chebula extracts against S. aureus and MRSA produced similar MIC values ranging from 94 µg/mL to 900 µg/mL. These findings indicate that the resistance mechanisms observed in the MRSA strain have little influence on the active compounds of the extracts. Thus, the extract compounds either function through different mechanisms, or they may inhibit the bacterial antibiotic-resistance pathways.
The mecA gene plays a crucial role in the resistance of MRSA [26]. This gene codes for a new penicillin-binding protein (PBP2a) that has a reduced ability to bind to β-lactam antibiotics. This protein provides the bacteria with resistance against many β-lactam antibiotics by enabling them to produce cell walls, even in the presence of these medications. Hence, the mechanisms by which the phytochemicals in the extract function may vary from those of β-lactam antibiotics, even in bacteria that are resistant to β-lactam antibiotics. Alternatively, these extracts may contain phytochemicals that disrupt the bacterial strains’ defence mechanisms against these medicines, thereby allowing them to function at higher potency [26]. This outcome is promising because, when compared to the susceptible strain, the MRSA strain in our study exhibited significantly diminished susceptibilities/ increased resistance to a range of antibiotics from the β-lactams, macrolides and fluoroquinolones classes.
Aqueous and methanol T. bellirica and T. chebula extracts exhibited antibacterial activity against E. coli and its antibiotic resistant counterpart, ESBL E. coli. The methanolic T. bellirica extract showed noteworthy antibacterial activity against both bacterial pathogens, with identical MIC values. Similarly, the methanolic T. bellirica and T. chebula extracts also inhibited the growth of K. pneumoniae and ESBL K. pneumoniae, with similar MIC values. These results suggest that the methanolic extracts of both plants may contain compounds that have a broad efficacy against strains of E. coli and K. pneumoniae, including those that produce ESBL enzymes. Their efficacy might be linked to a distinct mechanism of action that contrasts those of β-lactam antibiotics. For example, the plant extracts may affect how bacteria form their cell walls, how well their membrane’s function, or they may affect other essential processes independent of the β-lactam antibiotic mechanism [27]. In addition, it is possible that the methanolic extracts of both plants are exerting antibacterial effects through mechanisms that do not directly interfere with or inhibit the ESBL enzymes. Additional research is necessary to ascertain whether the plant extracts specifically hinder the resistance mechanisms of ESBL bacterial pathogens, or if they function via independent antibiotic pathways. This may require assessment of the effects of the extract/isolated components on β-lactamase inhibition, or by investigating the impact of the extract on the production of resistance genes in ESBL strains.
We also examined the use of T. bellirica and T. chebula extracts in combinations with conventional antibiotics. This method holds great promise for developing novel antibiotic chemotherapies, as many bacteria have developed resistance to conventional antibiotics, and plant compounds may provide ways of inhibiting/blocking these resistance mechanisms [4]. Our goal was to enhance the efficacy of antibiotics and possibly negate the bacterial resistance mechanisms by mixing them with extracts from plants. Augmentin®, a combination of amoxicillin and clavulanic acid, is a well-known example of how combining medications can improve treatment outcomes [28] since clavulanic acid inhibits β-lactamase enzymes present in resistant bacteria. This enables amoxicillin to specifically target and eliminate the bacteria with greater efficiency, even in β-lactam resistant bacterial strains. Clavulanic acid acts as a β-lactamase inhibitor by binding permanently to the active site of the enzyme, therefore inhibiting the breakdown of the antibiotic.
Our investigation found that penicillin G, amoxicillin, in combination with the plant extracts, have additive effects against S. aureus, which may be due to the presence of phytochemicals that possess anti-β-lactamase activities [29]. These phytochemicals can hinder the activity of β-lactamase enzymes, thereby contributing to antibiotic resistance by breaking down the β-lactam ring found in β-lactam antibiotics, such as penicillin and amoxicillin [30]. In addition, phytochemicals found in the plant extracts may interact with β-lactamase enzymes in a way that is comparable to clavulanic acid, although this requires confirmation. Regardless of the mechanism, this interaction helps protect the antibiotics from being broken down by bacterial enzymes and improves their ability to fight against bacterial pathogens. Plant extracts may possess the capacity to function as an alternative treatment method and could be a safe and less detrimental option for addressing antibiotic resistance [9]. In comparison to synthetic antibiotics, natural compounds found in plants frequently exhibit fewer adverse effects and a lower likelihood of generating resistance [8]. Moreover, the likelihood of further antibiotic-resistance development may be reduced by the variety of phytochemicals present in plant extracts, which may act on multiple bacterial pathways.
The ethyl acetate T. chebula extract exhibited additive effects against S. aureus in combination with erythromycin. It is possible that the extract and erythromycin may target distinct bacterial processes. Erythromycin inhibits protein synthesis by binding to the 50S ribosomal subunit [31], whereas the plant extract may impact other bacterial targets, including cell wall synthesis, membrane integrity, or metabolic processes [30]. Further studies are required to determine which of these mechanisms are affected by the extracts. Notably, this complementary targeting has the potential to improve the overall antibacterial effect. Furthermore, the plant extract may also influence other mechanisms associated with bacterial resistance, such as the disruption of efflux pumps or the modification of cell wall structures [30], which could render bacteria more susceptible to erythromycin. The extract may also disrupt bacterial protective barriers, which could facilitate the improved penetration of erythromycin into the cell. Additionally, the plant extract may contain a variety of bioactive compounds [8] that work in conjunction to enhance the efficacy of erythromycin. Alternatively, the extract may exhibit broad-spectrum antimicrobial activity that complements the specific effects of erythromycin. Similarly, the methanolic and ethyl acetate extracts of T. chebula exhibited additional antibacterial effects against K. pneumoniae when combined with chloramphenicol. This antibiotic binds to the 23S rRNA component of the 50S ribosomal subunit of the bacterial ribosome [32]. This binding suppresses the peptidyl transferase activity, a crucial process for the creation of peptide bonds during protein synthesis. Chloramphenicol inhibits bacterial protein production by blocking the formation of peptide bonds.
Our research determined that when tetracycline is combined with T. chebula extracts, the combination exhibited additive interactions against S. aureus, MRSA, E. coli and K. pneumoniae, suggesting enhanced antibacterial efficacy. However, the aqueous T. bellirica extract only showed an additive interaction against MRSA. Tetracycline resistance is mostly commonly attributed to tetracycline-specific efflux pumps [33]. Hence, the observed additive effect indicates that the T. bellirica and T. chebula extracts might have hindered the functioning of these efflux pumps. By inhibiting efflux pumps, tetracycline is allowed to remain inside cells for a longer period, which increases its effectiveness. Whilst ribosomal changes may also contribute to tetracycline resistance, this pathway is substantially less common [33]. Our research findings suggest that a broad range of phytochemicals found in the extracts of T. bellirica and T. chebula may have antibacterial action against several different bacteria. Similarly, plant extracts prepared from Phyllanthus niruri L., Berberis vulgaris L., and Piper nigrum L. exhibit additive interactions in combination with tetracycline, and can inhibit tetracycline efflux pumps [34,35].
Interestingly, the T. bellirica and T. chebula ethyl acetate extracts exhibited additive interactions in combination with ciprofloxacin against MRSA. This indicates that ethyl acetate extracts contain phytochemicals that may target different bacterial processes such as cell membrane disruption, inhibition of cell wall synthesis, or interference with metabolic pathways [30]. The plant extract’s unique mechanisms may enhance the effectiveness of ciprofloxacin’s DNA-targeting action, resulting in a more comprehensive treatment [8]. The plant extract may also enhance the cellular absorption of ciprofloxacin (thereby increasing its intracellular concentration) or may block/inhibit MRSA’s antibiotic-resistance mechanisms [35].
Notably, polymyxin B and T. bellirica, and T. chebula extracts combinations exhibited substantial antagonistic interactions. The antagonistic interaction between polymyxin B and the plant extracts may be affected by variations in pH levels in the broth. Polymyxin B, which breaks bacterial cell membranes by binding to lipopolysaccharides, is pH sensitive, with substantially reduced potency under acidic or alkaline circumstances [36]. The inclusion of plant extracts may cause a change in the pH of the broth, which could affect the efficacy of both polymyxin B and the plant chemicals, potentially resulting in a decrease in overall antibacterial activity, although this remains to be confirmed in future studies. Changes in pH may also impact bacterial physiology, thereby reducing the vulnerability of bacteria to the combined actions of the two substances. Polymyxin B and the components of plant extracts may also have chemical interactions that are regulated by pH [36], which may further contribute to the antagonistic effects of the combination. Moreover, the antagonistic interactions between the extracts and polymyxin B can be ascribed to the binding of bioactive phytochemicals with polymyxin B, which impedes its absorption and efficacy in targeting bacterial cells [37]. Hence, understanding these dynamics is critical for optimising combination medicines and assuring antimicrobial efficacy. Future studies are planned to examine these effects.
LC-MS metabolomics analysis of the T. bellirica and T. chebula fruit extracts highlighted the presence of flavonoids, tannins, terpenoids and phenolic acid compounds (Table 3). Complete, comprehensive lists of phytochemicals present in the individual plant extracts are available as the supplementary files (Supplementary Materials: Tables S1 and S2). Notable phytochemicals identified in both plant extracts include quinic acid (Figure 2A), shikimic acid (Figure 2B), aureusidin 6-glucuronide (Figure 2C), madecassic acid (Figure 2D), pedunculoside (Figure 2E), 6-galloylglucose (Figure 2F), 1,2,6-trigalloyl-β-D-glucopyranose (Figure 2G), propyl gallate (Figure 2H), methyl gallate (Figure 2I), theogallin (Figure 2J), gallic acid (Figure 2K), ellagic acid (Figure 2L), sanguiin H4 (Figure 2M), hamamelitannin (Figure 2N), pyrogallol (Figure 2O), chebulic acid (Figure 2P), chebuloside II (Figure 2Q), and 1,6-bis-O-(3,4,5-trihydroxybenzoyl) hexopyranose (Figure 2R). The scientific literature has documented the presence of gallic acid, ellagic acid, chebulic acid, chebuloside II, methyl gallate, propyl gallate, ethyl gallate, phloroglucinol, pyrogallol, quercetin, kaempferol, and various others compound in the fruit extracts of T. bellirica and T. chebula [38,39,40]. In our previous study, we conducted qualitative GC-MS headspace analysis on aqueous, methanol and ethyl acetate fruit extracts of T. bellirica and T. chebula. Our analysis revealed the presence of several notable volatile terpenoids, including eucalyptol, linalool, methoxycitronellal, terpinene-4-ol, camphor, pinocarveol, carvone, endo borneol, L-fenchone, hyscylene, patchoulane, p-cumic aldehyde, and phenylbutanal [11].
Notably, Embaby et al. (2019) showed synergistic antibacterial activity of quinic acid-rich acetone bark extract of Ficus macrocarpa var. nitida. with tetracycline against E. coli and S. aureus [41]. Furthermore, an in-silico molecular docking study revealed that 5-caffeol quinic acid (and other phenolic compounds) may have antibacterial activity due to their efflux pump inhibitory effect. However, the study did not investigate the interactions between the plant extracts and conventional antibiotics. Another study reported that the antibacterial activity of quinic acid against E. coli and S. aureus, with MIC of 500 µg/mL to 1000 µg/mL, respectively [42]. Additionally, Zhang et al. (2024) reported synergistic antibacterial effects for shikimic acid (625 µg/mL) in combination with penicillin, ampicillin, amoxicillin, and ceftiofur against MRSA, with and significantly reduced MIC values reported for the combinations (4 to 16-fold decreases) [43]. Similarly, other investigations have shown that phloroglucinol derivatives exhibited synergistic antibacterial activity against MRSA in combination with vancomycin, penicillin and doxycycline [44,45,46], despite phloroglucinol itself lacking antibacterial activity against MRSA, yielding a high MIC of over 10,000 µg/mL [45].
We identified the flavonoid glucuronide, aureusidin 6-glucuronide in the ethyl acetate extracts of T. bellirica and T. chebula. Notably, studies evaluating the antibacterial activity of this compound are lacking in the literature, as are studies evaluating its effects in combination with conventional antibiotics. In addition, madecassic acid, a pentacyclic triterpenoid has been identified in the T. chebula ethyl acetate extracts in our study. Previous studies have reported noteworthy antibacterial activity of madecassic acid against S. aureus, MRSA, and E. coli, with MIC values of 31.25 (61.9 μM), 62.5 (124 μM), 250 µg/mL (495 μM), respectively [47]. These findings indicate that the madecassic acid exhibits more efficacy against Gram-positive bacteria, including S. aureus and MRSA, compared to Gram-negative bacteria such as E. coli. This discrepancy in effectiveness may be attributed to variations in cell wall structures and methods of action. These values aid in determining the most effective dosage levels and emphasise the potential of madecassic acid as a specific treatment for antibiotic-resistant bacteria, such as MRSA. Moreover, these findings establish a basis for future investigations, such as doing combination studies with different antibiotics to improve effectiveness. Antibacterial mechanism-focused experiments showed that madecassic acid destroys cell wall integrity, inhibiting the synthesis of soluble proteins and DNA topoisomerase I and II [47]. Pedunculoside, a triterpene saponin has been identified in the methanol and ethyl acetate T. chebula extracts. Interestingly, an older study reported the antibacterial effects of pedunculoside against S. aureus and E. coli, with MIC values of 200 µg/mL [48]. However, there is a shortage of literature on the combined effects of pedunculoside and antibiotics.
Previous work has documented the antibacterial activity of penta-galloyl-glucose against S. aureus and E. coli, with a MIC of 250 µg/mL (266 µM) against both bacteria [49]. The postulated mechanism of antibacterial activity is related to the suppression of the bacterial type II fatty acid production pathway. Another study reported broad-spectrum antibacterial activity of penta-galloyl-glucose against both methicillin-resistant S. aureus and quinolone-resistant S. aureus and E. coli, with MIC values ranging from 64–128 µg/mL (68 µM and 136 µM, respectively) [50]. The antibacterial activities of four derivatives of galloyl-β-D-glucose have also been investigated against multidrug-resistant E. coli and K. pneumoniae strains, with MIC values from 32 µg/mL to 128 µg/mL respectively (26-136 µM), [51]. However, combinational interactions of galloyl-β-D-glucose derivatives with reference antibiotics have not been conducted to date.
Our LC/MS experiments also identified the galloyl-glucose derivatives 1,2,6-trigalloyl-β-D-glucopyranose, 1,6-bis-O-(3,4,5-trihydroxybenzoyl) hexopyranose, and 6-O-[(2E)-3-phenyl-2-propenoyl]-1-O-(3,4,5-trihydroxybenzoyl)-β-D-glucopyranose in both T. bellirica and T. chebula extracts. Previous studies have reported that relatively high levels of gallotannins are present in T. bellirica and T. chebula extracts [40,52,53,54]. Notably, efflux pump inhibitory activity has previously been demonstrated for 1,2,6-tri-O-galloyl-β-D-glucopyranose against multidrug-resistant (MDR) uropathogenic E. coli [55]. Additionally, 1,2,6-tri-O-galloyl-β-D-glucopyranose exhibits synergistic antibacterial activity in combination with gentamicin and trimethoprim against E. coli [56]. Synthetic gallotannins also exhibit antibiofilm and antimicrobial activity against S. aureus and MRSA strains [57].
Synergistic interactions between orbifloxacin and propyl gallate against the E. coli resistant strain KVCC 1423 have previously been reported, with MIC values of the combination being reduced from 125 µg/mL (316 µM, orbifloxacin only) to 7.8 µg/mL (19.7 µM), and 312.5 µg/mL (1472 µM, propyl gallate only) to 78 µg/mL (367 µM), respectively [58]. In the present study, LC-MS analysis, propyl gallate was identified only in the T. chebula extracts, although methyl gallate was identified in both Terminalia species. Tamang et al. (2022) investigated the antibacterial activities of erythromycin, ampicillin, gentamicin, kanamycin, and ciprofloxacin in combination with gallic acid, methyl gallate, ethyl gallate, propyl gallate, butyl gallate, octyl gallate, dodecyl gallate, and stearyl gallate against MRSA [59]. It was noted in that study that octyl gallate (4 µg/mL) exhibited significant antimicrobial synergy against MRSA in combination with penicillin, ampicillin, cephalothin, gentamicin, tetracycline, erythromycin and lincomycin, reducing their MIC values from 64 µg/mL to 0.25–16 µg/mL.
A recent study reviewed the antimicrobial characteristics of sanguiins, highlighting their capacity to inhibit bacterial growth and the production of biofilms [60]. The antibacterial activity of sanguiin H6, a closely related compound to sanguiin H4, was demonstrated against S. aureus and MRSA, with a MIC value of 250 µg/mL [60,61]. Sanguiin H4 has been identified in the polyphenolic extract of Sanguisorba officinalis L., which exhibited antibacterial activity S. aureus and E. coli [62]. However, data is not available for the combinatorial interactions of sanguiins with antibiotics to combat AMR. The combination of gallic acid and thiamphenicol, and the combination of hamamelitannin with thiamphenicol or erythromycin, exhibited synergistic antibacterial effects against E. coli. (ATCC 25922) [63]. Furthermore, study detected additional antibacterial interactions against E. coli when combining gallic acid with ampicillin, or cefotaxime, and/or marbofloxacin, as well as when combining hamamelitannin with amoxicillin or marbofloxacin. Interestingly, gallic acid and hamamelitannin each have moderate antibacterial activity against E. coli, with MIC values of 1024 µg/mL (6024 µM) and 2048 µg/mL (4230 µM), respectively [63]. Bassyouni et al. (2015) noted that hamamelitannin (20 µg/mL) reduces the MIC of vancomycin (4 µg/mL), and clindamycin (32 µg/mL) to 0.25 µg/mL against MRSA strains [64]. Furthermore, hamamelitannin in combination with vancomycin and clindamycin effectively inhibit the biofilm formation in MRSA strains.
Our present study identified chebulic acid, a gallotannin compound in the aqueous extract of T. bellirica and all extracts of T. chebula. Chebulic acid has demonstrated numerous biological activities including anti-tumor activity, anti-atherogenic, anti-fibrotic, anti-ulcer, and antioxidant effects [65]. Yang et al. (2020) identified and isolated twenty chebulic acid and brevifolincarboxylic acid derivatives from an ethanolic extract prepared from the arial parts of Euphorbia hirta L. [66]. All of those compounds exhibited significant free radical scavenging activities, although antibacterial activity was not investigated in that study. Both ethyl gallate and tri-n-butyl chebulate have been isolated from the aqueous T. chebula fruit extract [67]. Both compounds inhibit K. pneumoniae growth, with MIC values of 156 µg/mL (787 µM) and 1250 µg/mL, respectively. However, combinational interactions with antibiotics were not investigated in that study. In the present study, chebuloside II was only identified in the methanolic T. chebula fruit extract. In contrast, several earlier studies reported the presence of chebuloside II in both T. bellirica and T. chebula extracts [40,53,54,68]. Chebuloside II rich T. chebula extracts exhibit hepatoprotective effects and prevent liver toxicity in animal models [68], although there is a lack of studies examining the antibacterial activity and combinational interactions of chebuloside II with conventional antibiotics.
In the present study, the extracts were found to be rich in gallic acid, ellagic acid and pyrogallol. Previous studies showed that gallic acid acts synergistically in combination with norfloxacin against S. aureus [69]. This combination decreases the MIC of norfloxacin from 156 μg/mL to 49 μg/mL. In addition, gallic acid lowered the MIC of gentamicin against S. aureus from 49 μg/mL to 2.5 μg/mL. Similarly, ellagic acid lowered the MIC of tetracycline, chloramphenicol, and tobramycin against a multidrug resistant isolate of E. coli [70]. Quave et al. (2012) showed that ellagic acid and its derivatives isolated from the roots of Rubus ulmifolius Schott. inhibited S. aureus growth by inhibiting biofilm formation [71]. Pyrogallol exhibited antibacterial activity against S. aureus and reduced the MIC of norfloxacin from 156 μg/mL to 78 μg/mL, and gentamicin from 49 μg/mL to 2.5 μg/mL [69]. In addition, pyrogallol exhibited broad-spectrum antibacterial activities against methicillin-susceptible S. aureus, MRSA, E. coli (ATCC 25922), colistin-resistant E. coli, and colistin-resistant K. pneumoniae [72].
We were unable to detect synergistic enhancement of antibacterial activity between T. bellirica and T. chebula extracts and any of the antibiotics we selected against the bacterial pathogens investigated. However, additive interactions of extracts were noted in some combinations containing penicillin G, amoxicillin, erythromycin, chloramphenicol, tetracycline and ciprofloxacin against S. aureus, MRSA, E. coli, and ESBL K. pneumoniae. This indicates that the phytochemicals present in the T. bellirica and T. chebula extracts may possess β-lactamase and/or efflux pump inhibitory properties. Further research is required to investigate the effects of these extracts against those resistance mechanisms.
The toxicity assays using Artemia nauplii revealed that all T. bellirica and T. chebula extracts are nontoxic, thereby indicating their safety as an antimicrobial agent. To determine whether these extracts are suitable for use in medicine, additional testing should be conducted utilising a panel of mammalian cell lines. Taken together, the findings of our study indicate that T. bellirica and T. chebula fruit extracts may be a valuable source of antimicrobial compounds for future research and development in the fight against bacterial infections.
4. Materials and Methods
4.1. Plant Origins
Terminalia bellirica fruit powder (batch no: BNFP/01) was produced by Organic Prime and was purchased online from Navafresh Australia. Terminalia chebula fruit powder (batch no: HRP1020) developed by Aarshaveda was obtained online from Sattvic Australia. The traditional Ayurvedic names of T. bellirica (Bhitaki, Baheda) and T. chebula (Haritaki, Harad) were used to search for the plant herb on the supplier’s website. The provider verified the authenticity and quality of the plant materials. The plant samples were labelled, and voucher specimens NBG-TB0220GU and NBG-TC0220GU for T. bellirica and T. chebula, respectively, were kept at Griffith University’s Gold Coast campus in the School of Pharmacy and Medical Sciences.
4.2. Extract Preparation
After weighing individual 1 g masses of T. bellirica and T. chebula fruit powders into three separate 50 mL tubes, sterile deionized water, methanol (AR grade), or ethyl acetate (AR grade) were added individually to achieve a total volume of 50 mL [11,34]. The organic solvents (methanol and ethyl acetate) were provided by ChemSupply (Gillman, Australia). Samples were mixed at room temperature for 24 hours by continuous oscillation at 30 rpm. The samples were then filtered through Whatman No. 54 filter paper (Sigma-Aldrich, Melbourne, Australia) into pre-weighed 50 mL tubes under vacuum pressure. The aqueous samples were lyophilized in an Alpha 1-4 LSC plus benchtop freeze dryer (Martin Christ, Osterode am Harz, Germany) for three days to dry the aqueous extracts. The organic solvent samples were subjected to evaporation at a temperature of 40°C for a duration of two days, or until the evaporation process was fully completed. To calculate the final yields, all dried extracts were weighed, and the mass of extract was determined. Subsequently, the extracts were resuspended in 10 mL of 1% dimethyl sulfoxide (DMSO; Merck, Macquarie Park, Australia) and sonicated three times using 20 s pulses of a probe sonicator set at 1kHz, with a 30 s rest period between pulses. All extracts were subsequently sterilised by passing through 0.22 µm syringe-driven filters (Sarstedt, Mawson Lakes, Australia) and stored at −20 °C until use.
4.3. Antibiotics and Bacterial Strains
Powdered antibiotics were purchased from Sigma-Aldrich (Melbourne, Australia), which included penicillin G (potency of 1440–1680 µg/mg), erythromycin (potency ≥850 µg/mg), tetracycline (≥95% purity by HPLC), chloramphenicol (≥98% purity by HPLC), ciprofloxacin (≥98% purity by HPLC), polymyxin B (purity >90%), oxacillin (≥95% purity by TLC), amoxycillin (potency of 900 µg/mg), gentamicin (≥98% purity by HPLC), and vancomycin (potency of ≥900 μg per mg). Antibiotic stock solutions (1 mg/mL) were prepared for broth microdilution assays and stored at −20 °C until required. Oxoid Ltd. (Thebarton, Australia) supplied preloaded standard discs that contained penicillin G (10 IU), erythromycin (10 µg), tetracycline (30 µg), chloramphenicol (30 µg), ciprofloxacin (1 µg), polymyxin B (300IU), oxacillin (1 µg), gentamicin (10 µg), vancomycin (30 µg), Augmentin® (15 µg) and cefoxitin (30 µg). A 10 µL volume of amoxicillin stock solution (0.01 mg/mL) was infused into sterile filter paper discs, which were subsequently placed on Mueller–Hinton (MH) agar and used immediately for disc diffusion assays. Broth microdilution assays were conducted with all reference antibiotics, except for Augmentin® and cefoxitin.
The American Type Culture Collection (ATCC, Manassas, VA, USA) provided the reference strains of Escherichia coli (ATCC 25922), Staphylococcus aureus (ATCC 25923), MRSA (ATCC 43300), Klebsiella pneumoniae (ATCC 13883), and ESBL Klebsiella pneumoniae (ATCC 700603). A clinical isolate strain of ESBL Escherichia coli was obtained from the Gold Coast University Hospital in Southport, QLD, Australia and its resistance profile has been previously confirmed by our group [73]. Mueller–Hinton (MH) agar and broth (Oxoid Ltd., Australia) were utilised to cultivate all bacterial strains. The resistance phenotype of the MRSA strain was preserved by culture at 35 °C [74], whilst all other bacterial strains were cultured at 37 °C for 18–24 hours. All MH agar plate was prepared in accordance with the instructions provided by the manufacturer and with an agar depth of 4 millimetres.
4.4. Antibacterial Susceptibility Screening
The antibacterial activity of all plant extracts in MH agar was investigated using a modified Kirby–Bauer disc diffusion method [34]. In summary, individual colonies isolated from MH agar plates were inoculated in 40 mL MH broth and grown at 37 °C for 18–24 h, except for MRSA, which was grown at 35 °C. The individual bacterial cultures were used to make 0.5 McFarland standards for each strain. Subsequently, 100 µL of the 0.5 McFarland standards was spread on fresh MH agar plates. With sterile forceps, Whatman sterile filter paper discs (6 mm in diameter) were affixed to MH agar, and 10 µL of all extracts resuspended in 1% DMSO was infused into them. Each bacterial strain was inoculated on MH agar, and all reference antibiotic discs were inserted. The extract and reference antibiotic plates were incubated at 37 °C for 18–24 hours, except for MRSA, which was incubated at 35 °C. All samples were examined in triplicate.
The zones of inhibition (ZOI) were reported as the diameter of the inhibition zones around each disc, which was measured to the nearest whole millimetre (mm), to assess the bacterial growth inhibition. Samples with no visible inhibition were reported to have ZOIs of 6 mm (the diameter of the discs). The ZOI data are presented in the form of bar graphs as the average ± SEM (standard error of mean) of a minimum of three independent studies. One-way analysis of variance (ANOVA) was employed to analyse the differences between the treatment groups and the negative controls. The p-values of less than 0.01 and 0.001 were deemed to be statistically highly significant and very highly significant, respectively.
4.5. Minimum Inhibitory Concentration Determinations
The minimum inhibitory concentration (MIC) values for all extracts and reference antibiotics were determined using a standard 96-well microtitre plate broth microdilution assay [11,34]. A 100 µL volume of the individual extracts and reference antibiotics were added to the top row of the plates. The dilutions were then made down each column of the plates using doubling dilutions. After adding a volume of 100 µL of a 1:100 dilution of 0.5 McFarland cell suspension to every well (except from the sterile controls), the wells were incubated for 20–24 hours at 37 °C. After the initial incubation period, a solution (0.4 mg/mL) of p-iodonitrotetrazolium violet (INTZ; Sigma Aldrich, Australia) dye was added to each well on the plate. The plate was then incubated for an additional 2-4 hours at room temperature. For determining the MIC values, a visual inspection method was employed. This involved identifying the lowest concentration of plant extracts or antibiotics that effectively prevented bacterial growth, which was indicated by the absence of a red-pink colour change. The experiments were conducted in duplicate. MIC values greater than 10,000 μg/mL were classified as inactive, while values between 2000 and 10,000 μg/mL were considered to have low activity. Moderate activity was assigned to MIC values between 1000 and 2000 μg/mL, noteworthy activity to values between 400 and 1000 μg/mL, and good activity to values between 100 and 400 μg/mL. MIC values below 100 μg/mL were regarded as having high activity.
4.6. Fractional Inhibitory Concentration Evaluation
Plant extracts and antibiotics that showed antibacterial activity at minimum inhibitory concentration (MIC) ≤ 3000 µg/mL and ≤2.5 µg/mL, respectively were selected for combination studies. These were combined in equal proportions (50:50) to study their interactions with susceptible bacterial pathogens. The study examined the interactions between the extracts and antibiotics by calculating the sum of the fractional inhibitory concentrations (ΣFIC) for each combination using the following equations (a = extracts; b = antibiotics):
FIC(a) = MIC (a in combination with b)/MIC (a independently)
FIC(b) = MIC (b in combination with a)/MIC (b independently).
The sum of the fractional inhibitory concentrations (∑FIC) was determined by applying the formula ∑FIC = FIC(a) + FIC(b).
The interactions were classified as synergistic (∑FIC ≤ 0.5), additive (∑FIC > 0.5–≤ 1.0), indifferent (∑FIC > 1.0–≤ 4.0), or antagonistic (∑FIC > 4.0) [75].
4.7. Toxicity Assays
The toxicity of the plant extracts and controls were assessed using standard Artemia franciscana Kellogg nauplii lethality assays (ALA) [76]. In summary, 400 µL of plant extracts (2mg/mL concentration, diluted in artificial seawater) and 400 µL of artificial seawater with newly hatched (within 1 day) A. franciscana nauplii were added separately in each well of a 48-well plate. On all plates, a 400 µL volume of negative control (32 g/L artificial seawater; Red Sea) and positive control (1 mg/mL sodium azide) was added. The plates were then incubated at 25 °C ± 1 °C for 24 hours, and the number of live shrimps was determined. Probit analysis was employed to determine the LC50 values, which represent the concentration of extract or control required to cause mortality in 50% of A. franciscana nauplii in separate wells. The LC50 values were calculated graphically using the mean percentage of three repeated experiments, representing the concentration that resulted in 50% mortality.
4.8. Non-Targeted Headspace LC-MS for Quantitative Analysis
A Vanquish Ultra High-Performance Liquid Chromatography (UHPLC) system (Thermo Fisher Scientific, Waltham, MA, USA) was utilised to conduct a non-targeted headspace metabolic profile analysis on all samples [34]. The separation of compounds was achieved using an Accucore ™ RP-MS column (100 mm × 2.1 mm) with a particle size of 2.6 μm, which was linked to an Orbitrap Exploris 120 mass spectrometer (Thermo Fisher Scientific). The UHPLC system was fitted with a quaternary pump at a flow rate of 0.6 mL/min. The separation used the following mobile phases: (A) A solution of 0.1% v/v formic acid in ultrapure water and (B) acetonitrile (MeCN) solution containing 0.1% v/v formic acid. The Xcalibur acquisition software (version 2.0) was utilised to create a gradient flow for compound separation. The flow consisted of the following steps, with a total duration of 24 minutes: 5% B for 5 minutes, a linear increase from 5% to 30% B over 5 minutes, 30% B for 3 minutes, a linear increase from 30% to 90% B over 4 minutes, isocratic elution at 90% B for 4 minutes to flush the column, and a linear decrease from 90% to 5% B over 1 minute. The column was re-equilibrated by running an isocratic separation at 5% B of the mobile phase for a duration of 2 minutes between each separation. To get chromatogram peaks of high quality, plant extract samples were analysed at three distinct concentrations (0.25 mg/mL, 0.5 mg/mL, and 1 mg/mL) using the Vanquish HPLC system (Thermo Fisher Scientific, Waltham, MA, USA) linked to the same column as mentioned above. Plant samples exhibited enhanced chromatogram peaks of superior quality when used at a concentration of 1 mg/mL.
The mass spectra of the eluted compounds were examined using the Orbitrap Exploris 120 mass spectrometer in information-dependent acquisition (IDA) mode. The Orbitrap system employed electrospray ionisation (ESI) in negative ionisation mode, utilising specified settings including a vaporiser temperature of 350 °C, sheath gas pressure of 60 psi, auxiliary gas pressure of 15 psi, sweep gas pressure of 2 psi, and a spray voltage ranging from 2.5 KV to 5 KV. The data was analysed using Compound DiscovererTM software 3.3. The identification of the unknown natural products utilised local database searches and online statistical analysis. To remove the background and differentiate individual components of the extract, the data files for each extract were analysed individually and compared to the blank file. In the Compound Discoverer function, result filters were employed to utilise databases such as Mz cloud, ChemSpider, Predicted Compositions, and MassList to achieve either a partial or complete match of the compounds that were discovered. The Compound Discoverer software 3.3 was utilised to generate Excel files containing the discovered compounds from each extract. The files were subsequently analysed to generate a catalogue containing potential putative compounds. Following the consolidation of duplicates by pivot table analysis in Microsoft Excel, the relative abundance was calculated as a percentage of the total area.
5. Conclusions
Due to increasing levels of antibiotic-resistant bacterial strains, there is a pressing demand for new antibacterial medications, which has generated interest in natural products as potential sources. The results of our study demonstrate that the active extracts derived from Terminalia bellirica and Terminalia chebula can effectively hinder the growth of both resistant and susceptible strains of bacteria. This suggests that the extracts possess unique antibacterial mechanisms that require further examination. In addition, all extracts enhanced the effectiveness of various conventional antibiotics, specifically β-lactam antibiotics and tetracycline. This may allow the reactivation of these antibiotics, even in bacterial strains that are otherwise resistant to their actions. The method of potentiation has not been discovered yet, although it is probable that the components of the extract may deactivate bacterial extended spectrum β-lactamase enzymes and efflux pumps, leading to an increase in the concentration of antibiotics inside the cells. Future studies are required to confirm these mechanisms, as well as to test other possible potentiation pathways. Several phytochemicals that were identified in the extracts may contribute to these actions, suggesting that they could be valuable targets for new antibacterial agent development, although further confirmation is required. Subsequent studies should prioritise the isolation of these compounds and examine their capacity to augment the effectiveness of existing antibiotics. Also, nuclear magnetic resonance (NMR) is essential for the conclusive structural identification of the components present in the extract.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: LC-MS total compound chromatograms of (A) TB-Aq (Terminalia bellirica aqueous), (B) TB-MeOH (Terminalia bellirica methanol), and (C) TB-EtOAc (Terminalia bellirica ethyl acetate). Figure S2: LC-MS total compound chromatograms of (A) TCh-Aq (Terminalia chebula aqueous), (B) TB-MeOH (Terminalia chebula methanol), and (C) TB-EtOAc (Terminalia chebula ethyl acetate). Table S1: LC-MS putative identification and % relative abundance of phytochemicals identified in the fruit extracts of Terminalia bellirica. Compounds less than 0.01% of the total area were considered as trace amounts and denoted as T. Table S2: LC-MS putative identification and % relative abundance of phytochemicals identified in the fruit extracts of Terminalia chebula. Compounds less than 0.01% of the total area were considered as trace amounts and denoted as T.
Author Contributions
Conceptualization, investigation, resources, M.J.C. and I.E.C.; methodology, G.T. and M.J.C.; validation, formal analysis, data curation, G.T.; writing—original draft preparation, G.T..; writing—review and editing, supervision, M.J.C. and I.E.C.; project administration and funding acquisition, I.E.C. and M.J.C. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support for this work was provided by the Environmental Futures Research Institute, Griffith University, Australia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are either presented within the manuscript or are available from the corresponding author upon reasonable request.
Acknowledgments
The authors wish to thank Muhammad Jawad Zai for assistance with compound identification.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- WHO. Antimicrobial resistance. World Health Organization. 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 26 July 2024).
- Centers for Disease Control and Prevention (CDC). 2019 Antibiotic Resistance Threats Report. Available online: https://www.cdc.gov/antimicrobial-resistance/data-research/threats/ (accessed on 26 July 2024).
- World Bank. Drug-Resistant Infections: A Threat to Our Economic Future. Available online: https://www.worldbank.org/en/topic/health/publication/drug-resistant-infections-a-threat-to-our-economic-future (accessed on 6 June 2024).
- Ajulo, S.; Awosile, B. Global antimicrobial resistance and use surveillance system (GLASS 2022): Investigating the relationship between antimicrobial resistance and antimicrobial consumption data across the participating countries. PLoS ONE 2024, 19, e0297921. [Google Scholar] [CrossRef] [PubMed]
- Renwick, M.J.; Brogan, D.M.; Mossialos, E. A systematic review and critical assessment of incentive strategies for discovery and development of novel antibiotics. J. Antibiot. (Tokyo) 2016, 69, 73–88. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO releases report on state of development of antibacterials. 2024. Available online: https://www.who.int/news/item/14-06-2024-who-releases-report-on-state-of-development-of-antibacterials (accessed on 18 June 2024).
- Abdallah, E.M.; Alhatlani, B.Y.; de Paula Menezes, R.; Martins, C.H.G. Back to nature: medicinal plants as promising sources for antibacterial drugs in the post-antibiotic era. Plants 2023, 12, 3077. [Google Scholar] [CrossRef] [PubMed]
- Tiwana, G.; Cock, I.E.; Cheesman, M.J. A Review of Ayurvedic principles and the use of Ayurvedic plants to control diarrhoea and gastrointestinal infections. Pharmacogn. Commun. 2023, 13, 152–162. [Google Scholar] [CrossRef]
- Cheesman, M.; Ilanko, A.; Blonk, B.; Cock, I. Developing new antimicrobial therapies: Are synergistic combinations of plant extracts/compounds with conventional antibiotics the solution? Pharmacogn. Rev. 2017, 11, 57. [Google Scholar] [CrossRef]
- Belmehdi, O.; Bouyahya, A.; Jekő, J.; et al. Synergistic interaction between propolis extract, essential oils, and antibiotics against Staphylococcus epidermidis and methicillin-resistant Staphylococcus aureus. Int. J. Sec. Metab. 2021, 8, 195–213. [Google Scholar] [CrossRef]
- Tiwana, G.; Cock, I.E.; White, A.; Cheesman, M.J. Use of specific combinations of the triphala plant component extracts to potentiate the inhibition of gastrointestinal bacterial growth. J. Ethnopharmacol. 2020, 260, 112937. [Google Scholar] [CrossRef]
- Thakur, M.; Khushboo; Yadav, A. ; et al. Antimicrobial activity against antibiotic-resistant pathogens and antioxidant activity and lcms/ms phytochemical content analysis of selected medicinal plants. J. Pure Appl. Microbiol. 2024, 18, 722–738. [Google Scholar] [CrossRef]
- Badoni, H.; Sharma, P.; Mohsin Waheed, S.; Singh, S. Phytochemical analyses and evaluation of antioxidant, antibacterial, and toxic properties of Emblica officinalis and terminalia bellirica fruit extracts. Asian J. Pharm. Clin. Res. 2016, 9. [Google Scholar] [CrossRef]
- Mani, P.; Devi, P.N.; Kaleeswari, S.; Poonkothai, M. Antimicrobial activity and phytochemical analysis of fruit extracts of Terminalia bellirica. Int. J. Pharm. Pharm. Sci. 2014, 43, 364. [Google Scholar]
- Sharma, R.; Raizada, S.; Gautam, A.; Bhatia, A.K. Phytochemical and antibacterial analysis of Terminalia chebula and Terminalia bellirica. In Green Chemistry in Environmental Sustainability and Chemical Education; Springer, 2018; pp. 131–137. [CrossRef]
- Parekh, J.; Chanda, S. Evaluation of antimicrobial activity of Terminalia chebula Retz. fruit in different solvents. J. Herbs Spices Med. Plants 2008, 13, 107–116. [Google Scholar] [CrossRef]
- Bag, A.; Kumar Bhattacharyya, S.; Bharati, P.; et al. Evaluation of antibacterial properties of Chebulic myrobalan (fruit of Terminalia chebula Retz.) extracts against methicillin-resistant Staphylococcus aureus and trimethoprim-sulphamethoxazole resistant uropathogenic Escherichia coli. Afr. J. Plant Sci. 2009, 3, 25–29. [Google Scholar]
- Zearah, S.A. Antifungal and antibacterial activity of flavonoid extract from Terminalia chebula retz. fruits. Journal of Basrah Researches (Sciences) 2014, 40, 122–131. [Google Scholar]
- Cajka, T.; Hricko, J.; Rudl Kulhava, L.; et al. Optimization of mobile phase modifiers for fast LC-MS-based untargeted metabolomics and lipidomics. Int. J. Mol. Sci. 2023, 24, 1987. [Google Scholar] [CrossRef]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; et al. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef]
- Bonev, B.; Hooper, J.; Parisot, J. Principles of assessing bacterial susceptibility to antibiotics using the agar diffusion method. J. Antimicrob. Chemother. 2008, 61, 1295–1301. [Google Scholar] [CrossRef]
- Jain, S.; Patel, N.; Lin, S. Solubility and dissolution enhancement strategies: Current understanding and recent trends. Drug Dev. Ind. Pharm. 2015, 41, 875–887. [Google Scholar] [CrossRef]
- Flanagan, J.N.; Steck, T.R. The relationship between agar thickness and antimicrobial susceptibility testing. Indian J. Microbiol. 2017, 57, 503–506. [Google Scholar] [CrossRef]
- Bhattacharjee, M.K. Better visualization and photodocumentation of zone of inhibition by staining cells and background agar differently. J. Antibiot. 2015, 68, 657–659. [Google Scholar] [CrossRef]
- Livermore, D.M. Antibiotic resistance in staphylococci. Int. J. Antimicrob. Agents 2000, 16, 3–10. [Google Scholar] [CrossRef]
- Kim, C.; Mwangi, M.; Chung, M.; et al. The Mechanism of heterogeneous beta-lactam resistance in MRSA: Key role of the stringent stress response. PLoS ONE 2013, 8, e82814. [Google Scholar] [CrossRef] [PubMed]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; et al. Towards advances in medicinal plant antimicrobial activity: A review study on challenges and future perspectives. Microorganisms 2021, 9, 2041. [Google Scholar] [CrossRef] [PubMed]
- Huttner, A.; Bielicki, J.; Clements, M.N.; et al. Oral amoxicillin and amoxicillin–clavulanic acid: properties, indications, and usage. Clin. Microbiol. Infect. 2020, 26, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Oelschlaeger, P. β-Lactamases: Sequence, structure, function, and inhibition. Biomolecules 2021, 11, 986. [Google Scholar] [CrossRef] [PubMed]
- Khameneh, B.; Eskin, N.A.M.; Iranshahy, M.; Fazly Bazzaz, B.S. Phytochemicals: A promising weapon in the arsenal against antibiotic-resistant bacteria. Antibiotics 2021, 10, 1044. [Google Scholar] [CrossRef]
- Platon, V.M.; Dragoi, B.; Marin, L. Erythromycin formulations—A journey to advanced drug delivery. Pharmaceutics 2022, 14, 2180. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Schwarz, S.; Jacobsen, L.; et al. A new mechanism for chloramphenicol, florfenicol, and clindamycin resistance: Methylation of 23S ribosomal RNA at A2503. Mol. Microbiol. 2005, 57, 1064–1073. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, V.K.; Pathania, R. Efflux pump inhibitors for bacterial pathogens: From bench to bedside. Indian J. Med. Res. 2019, 149, 129. [Google Scholar] [CrossRef]
- Tiwana, G.; Cock, I.E.; Cheesman, M.J. Phyllanthus niruri Linn.: Antibacterial activity, phytochemistry, and enhanced antibiotic combinatorial strategies. Antibiotics 2024, 13, 654. [Google Scholar] [CrossRef]
- Gibbons, S. Phytochemicals for bacterial resistance--strengths, weaknesses, and opportunities. Planta Med. 2008, 74, 594–602. [Google Scholar] [CrossRef]
- Cai, X.; Javor, S.; Gan, B.H.; et al. The antibacterial activity of peptide dendrimers and polymyxin B increases sharply above pH 7.4. Chem. Commun. 2021, 57, 5654–5657. [Google Scholar] [CrossRef] [PubMed]
- Zavascki, A.P.; Goldani, L.Z.; Li, J.; Nation, R.L. Polymyxin B for the treatment of multidrug-resistant pathogens: A critical review. J. Antimicrob. Chemother. 2007, 60, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Cock, I.E. The medicinal properties and phytochemistry of plants of the genus Terminalia (Combretaceae). Inflammopharmacology 2015, 23, 203–229. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; J Krishana, M.; Joshi, A.B.; et al. A pharmacognostic, phytochemical, and pharmacological review of Terminalia bellirica. J. Pharmacogn. Phytochem. 2017, 6, 368–376. [Google Scholar]
- Bag, A.; Bhattacharyya, S.K.; Chattopadhyay, R.R. The development of Terminalia chebula Retz. (Combretaceae) in clinical research. Asian Pac. J. Trop. Biomed. 2013, 3, 244. [Google Scholar] [CrossRef]
- Embaby, M.A.; El-Raey, M.A.; Zaineldain, M.; et al. Synergistic effect and efflux pump inhibitory activity of Ficus nitida phenolic extract with tetracycline against some pathogenic bacteria. Toxin Rev. 2021, 40, 1187–1197. [Google Scholar] [CrossRef]
- Adamczak, A.; Ożarowski, M.; Karpiński, T.M. Antibacterial activity of some flavonoids and organic acids widely distributed in plants. J. Clin. Med. 2020, 9, 109. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, Q.; Wang, Y.; et al. Exploiting the synergistic antibacterial activity of shikimic acid and ceftiofur against methicillin-resistant Staphylococcus aureus. World J. Microbiol. Biotechnol. 2024, 40, 1–11. [Google Scholar] [CrossRef]
- Yang, X.; Gao, X.; Ou, J.; et al. Antimicrobial activity and mechanism of anti-MRSA of phloroglucinol derivatives. DARU J. Pharm. Sci. 2024, 32, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Duan, S.; Hu, Y.; et al. Antibacterial activity and synergistic antibiotic mechanism of trialdehyde phloroglucinol against methicillin-resistant Staphylococcus aureus. Phytother. Res. 2023, 37, 490–504. [Google Scholar] [CrossRef]
- Mittal, N.; Tesfu, H.H.; Hogan, A.M.; et al. Synthesis and antibiotic activity of novel acylated phloroglucinol compounds against methicillin-resistant Staphylococcus aureus. J. Antibiot. 2019, 72, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Cui, P.; Liu, X. Antibacterial activity and mechanism of madecassic acid against Staphylococcus aureus. Molecules 2023, 28, 1895. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, H.; Kataoka, S.; Okamoto, S.; et al. Antimicrobial triterpenes from Ilex integra and the mechanism of antifungal action. Phytother. Res. 1999, 13, 151–160. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Kim, S.H.; et al. Anti-cancer, anti-diabetic and other pharmacologic and biological activities of penta-galloyl-glucose. Pharm. Res. 2009, 26, 2066–2080. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Sohn, M.J.; Lee, J.; Kim, W.G. Isolation and identification of pentagalloylglucose with broad-spectrum antibacterial activity from Rhus trichocarpa Miquel. Food Chem. 2010, 123, 501–506. [Google Scholar] [CrossRef]
- Masota, N.E.; Ohlsen, K.; Schollmayer, C.; et al. Isolation and Characterization of Galloylglucoses Effective against Multidrug-Resistant Strains of Escherichia coli and Klebsiella pneumoniae. Molecules 2022, 27, 5045. [Google Scholar] [CrossRef]
- Pfundstein, B.; El Desouky, S.K.; Hull, W.E.; et al. Polyphenolic compounds in the fruits of Egyptian medicinal plants (Terminalia bellerica, Terminalia chebula and Terminalia horrida): Characterization, quantitation and determination of antioxidant capacities. Phytochemistry 2010, 71, 1132–1148. [Google Scholar] [CrossRef]
- Avula, B.; Wang, Y.H.; Wang, M.; et al. Simultaneous determination and characterization of tannins and triterpene saponins from the fruits of various species of Terminalia and Phyllantus emblica using a UHPLC-UV-MS Method: Application to Triphala. Planta Med. 2013, 29, 181–188. [Google Scholar] [CrossRef]
- Manosroi, A.; Jantrawut, P.; Ogihara, E.; et al. Biological activities of phenolic compounds and triterpenoids from the Galls of Terminalia chebula. Chem. Biodivers. 2013, 10, 1448–1463. [Google Scholar] [CrossRef]
- Bag, A.; Chattopadhyay, R.R. Efflux-pump inhibitory activity of a gallotannin from Terminalia chebula fruit against multidrug-resistant uropathogenic Escherichia coli. Nat. Prod. Res. 2014, 28, 1280–1283. [Google Scholar] [CrossRef]
- Bag, A.; Chattopadhyay, R.R. Synergistic antibiofilm efficacy of a gallotannin 1,2,6-tri-O-galloyl-β-D-glucopyranose from Terminalia chebula fruit in combination with gentamicin and trimethoprim against multidrug resistant uropathogenic Escherichia coli biofilms. PLoS ONE 2017, 12, e0178712. [Google Scholar] [CrossRef] [PubMed]
- Hricovíniová, Z.; Mascaretti, Š.; Hricovíniová, J.; et al. New unnatural gallotannins: A way toward green antioxidants, antimicrobials and antibiofilm agents. Antioxidants 2021, 10, 1288. [Google Scholar] [CrossRef]
- Abbas, M.A.; Lee, E.B.; Boby, N.; et al. A pharmacodynamic investigation to assess the synergism of orbifloxacin and propyl gallate against Escherichia coli. Front. Pharmacol. 2022, 13, 989395. [Google Scholar] [CrossRef] [PubMed]
- Tamang, M.D.; Bae, J.; Park, M.; Jeon, B. Potentiation of β-Lactams against Methicillin-Resistant Staphylococcus aureus (MRSA) Using octyl gallate, a food-grade antioxidant. Antibiotics 2022, 11, 266. [Google Scholar] [CrossRef] [PubMed]
- Gesek, J.; Jakimiuk, K.; Atanasov, A.G.; Tomczyk, M. Sanguiins—Promising molecules with broad biological potential. Int. J. Mol. Sci. 2021, 22, 12972. [Google Scholar] [CrossRef]
- Aguilera-Correa, J.J.; Fernández-López, S.; Cuñas-Figueroa, I.D.; et al. Sanguiin H-6 fractionated from cloudberry (Rubus chamaemorus) seeds can prevent the methicillin-resistant Staphylococcus aureus biofilm development during wound infection. Antibiotics 2021, 10, 1481. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.L.; Chen, G.; Chen, S.N.; et al. Characterization of polyphenolic constituents from Sanguisorba officinalis L. and its antibacterial activity. Eur. Food Res. Technol. 2019, 245, 1487–1498. [Google Scholar] [CrossRef]
- Hossain, M.A.; Park, H.C.; Park, S.W.; et al. Synergism of the combination of traditional antibiotics and novel phenolic compounds against Escherichia coli. Pathogens 2020, 9, 811. [Google Scholar] [CrossRef]
- Bassyouni, R.H.; Dwedar, R.A.; Farahat, M.G.; et al. Protective effect of hamamelitannin against biofilm production by methicillin-resistant Staphylococci Isolated from blood of patients at intensive Care Units. Microbiol. Res. J. Int. 2015, 10, 1–8. [Google Scholar] [CrossRef]
- Dhingra, A.K.; Chopra, B.; Grewal, A.S.; Guarve, K. Pharmacological properties of chebulinic acid and related ellagitannins from nature: An emerging contemporary bioactive entity. Pharmacol. Res. Mod. Chin. Med. 2022, 5, 100163. [Google Scholar] [CrossRef]
- Yang, Z.N.; Su, B.J.; Wang, Y.Q.; et al. Isolation, absolute configuration, and biological activities of chebulic acid and brevifolincarboxylic acid Derivatives from Euphorbia hirta. J. Nat. Prod. 2020, 83, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Lin, Y.; Li, B.; et al. Antibacterial constituents of Fructus Chebulae Immaturus and their mechanisms of action. BMC Complement. Altern. Med. 2016, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tasduq, S.A.; Singh, K.; Satti, N.K.; et al. Terminalia chebula (fruit) prevents liver toxicity caused by sub-chronic administration of rifampicin, isoniazid and pyrazinamide in combination. Hum. Exp. Toxicol. 2006, 25, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Lima, V.N.; Oliveira-Tintino, C.D.M.; Santos, E.S.; et al. Antimicrobial and enhancement of the antibiotic activity by phenolic compounds: Gallic acid, caffeic acid and pyrogallol. Microb. Pathog. 2016, 99, 56–61. [Google Scholar] [CrossRef]
- Jenic, D.; Waller, H.; Collins, H.; Erridge, C. Reversal of tetracycline resistance by cepharanthine, cinchonidine, ellagic Acid and propyl gallate in a multi-drug resistant Escherichia coli. Nat. Prod. Bioprospect. 2021, 11, 345–355. [Google Scholar] [CrossRef]
- Quave, C.L.; Estévez-Carmona, M.; Compadre, C.M.; et al. Ellagic acid derivatives from Rubus ulmifolius inhibit Staphylococcus aureus biofilm formation and improve response to antibiotics. PLoS ONE 2012, 7, e28737. [Google Scholar] [CrossRef]
- Anek, P.; Kumpangcum, S.; Roytrakul, S.; et al. Antibacterial activities of phenolic compounds in Miang extract: Growth inhibition and change in protein expression of extensively drug-Resistant Klebsiella pneumoniae. Antibiotics 2024, 13, 536. [Google Scholar] [CrossRef]
- Cheesman, M.J.; Alcorn, S.R.; White, A.; Cock, I.E. Hamamelis virginiana L. leaf extracts inhibit the growth of antibiotic-Resistant Gram-positive and Gram-negative bacteria. Antibiotics 2023, 12, 1195. [Google Scholar] [CrossRef]
- Vitko, N.P.; Richardson, A.R. Laboratory maintenance of methicillin-resistant Staphylococcus aureus (MRSA). Curr. Protoc. Microbiol. 2013, 28, 9C.2.1–9C.2.14. [Google Scholar] [CrossRef]
- Doern, C.D. When does 2 plus 2 equal 5? A review of antimicrobial synergy testing. J. Clin. Microbiol. 2014, 52, 4124–4128. [Google Scholar] [CrossRef]
- Ruebhart, D.R.; Wickramasinghe, W.; Cock, I.E. Protective efficacy of the antioxidant’s vitamin E and Trolox against Microcystis aeruginosa and microcystin-LR in Artemia franciscana Nauplii. J. Toxicol. Environ. Health A 2009, 72, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Antimicrobial activities of T. bellirica and T. chebula fruit extracts in agar disc diffusion assays against (A) S. aureus, (B) MRSA, (C) E. coli, (D) ESBL E. coli, (E) K. pneumoniae, (F) ESBL K. pneumoniae. TB-AQ = T. bellirica aqueous; TB-MeOH = T. bellirica methanol; TB-EtOAc = T. bellirica ethyl acetate. TCh-AQ = T. chebula aqueous; TCh-MeOH = T. chebula methanol; TCh-EtOAc = T. chebula ethyl acetate. Negative controls = 1% DMSO and Blank = sterile water. Reference antibiotics: PEN G = penicillin G (10 IU), ERY = erythromycin (10 µg), TET = tetracycline (30 µg), CHL = chloramphenicol (30 µg), CIP = ciprofloxacin (1 µg), POL B = polymyxin B (300IU), OXA = oxacillin (1 µg), AMX = amoxycillin (10 µL of 0.01mg/mL stock solution), GEN = gentamicin (10 µg), VAN = vancomycin (30 µg), AUG = Augmentin® (15 µg), CEF = cefoxitin (30 µg). Horizontal red lines on the y-axis at 6 mm indicates the disc diameter used in the assay. Mean values (±SEM) are reported from three independent studies. p-values < 0.01 are represented with a single asterisk symbol (*), while p-values < 0.001 are represented with a double asterisk symbol (**).
Figure 1.
Antimicrobial activities of T. bellirica and T. chebula fruit extracts in agar disc diffusion assays against (A) S. aureus, (B) MRSA, (C) E. coli, (D) ESBL E. coli, (E) K. pneumoniae, (F) ESBL K. pneumoniae. TB-AQ = T. bellirica aqueous; TB-MeOH = T. bellirica methanol; TB-EtOAc = T. bellirica ethyl acetate. TCh-AQ = T. chebula aqueous; TCh-MeOH = T. chebula methanol; TCh-EtOAc = T. chebula ethyl acetate. Negative controls = 1% DMSO and Blank = sterile water. Reference antibiotics: PEN G = penicillin G (10 IU), ERY = erythromycin (10 µg), TET = tetracycline (30 µg), CHL = chloramphenicol (30 µg), CIP = ciprofloxacin (1 µg), POL B = polymyxin B (300IU), OXA = oxacillin (1 µg), AMX = amoxycillin (10 µL of 0.01mg/mL stock solution), GEN = gentamicin (10 µg), VAN = vancomycin (30 µg), AUG = Augmentin® (15 µg), CEF = cefoxitin (30 µg). Horizontal red lines on the y-axis at 6 mm indicates the disc diameter used in the assay. Mean values (±SEM) are reported from three independent studies. p-values < 0.01 are represented with a single asterisk symbol (*), while p-values < 0.001 are represented with a double asterisk symbol (**).
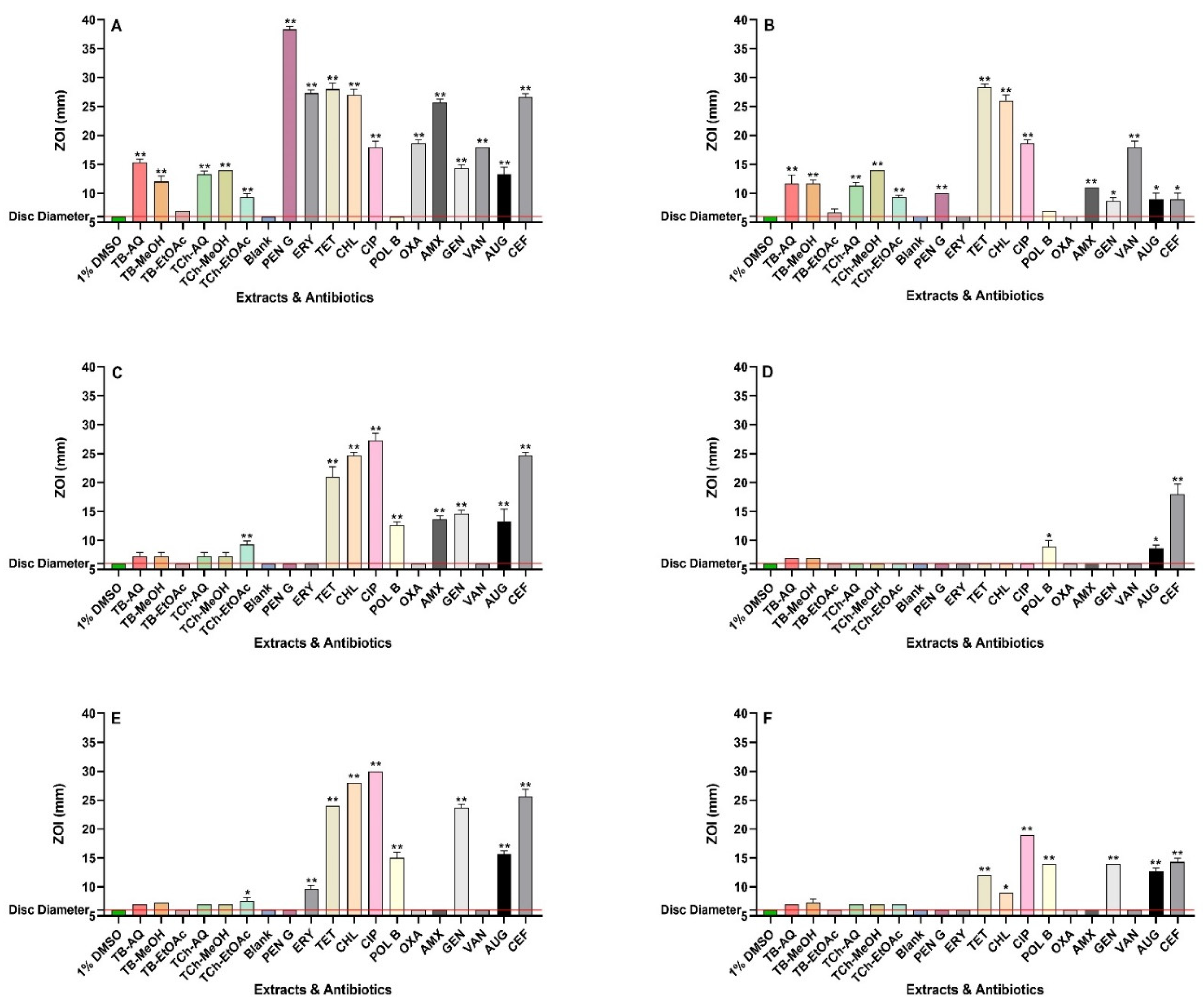
Figure 2.
Structures of noteworthy compounds identified in the fruit extracts of T. bellirica and T. chebula. quinic acid (A), shikimic acid (B), aureusidin 6-glucuronide (C), madecassic acid (D), pedunculoside (E), 6-galloylglucose (F), 1,2,6-trigalloyl-β-D-glucopyranose (G), propyl gallate (H), methyl gallate (I), theogallin (J), gallic acid (K), ellagic acid (L), sanguiin H4 (M), hamamelitannin (N), pyrogallol (O), chebulic acid (P), chebuloside II (Q), and 1,6-bis-O-(3,4,5-trihydroxybenzoyl) hexopyranose (R).
Figure 2.
Structures of noteworthy compounds identified in the fruit extracts of T. bellirica and T. chebula. quinic acid (A), shikimic acid (B), aureusidin 6-glucuronide (C), madecassic acid (D), pedunculoside (E), 6-galloylglucose (F), 1,2,6-trigalloyl-β-D-glucopyranose (G), propyl gallate (H), methyl gallate (I), theogallin (J), gallic acid (K), ellagic acid (L), sanguiin H4 (M), hamamelitannin (N), pyrogallol (O), chebulic acid (P), chebuloside II (Q), and 1,6-bis-O-(3,4,5-trihydroxybenzoyl) hexopyranose (R).
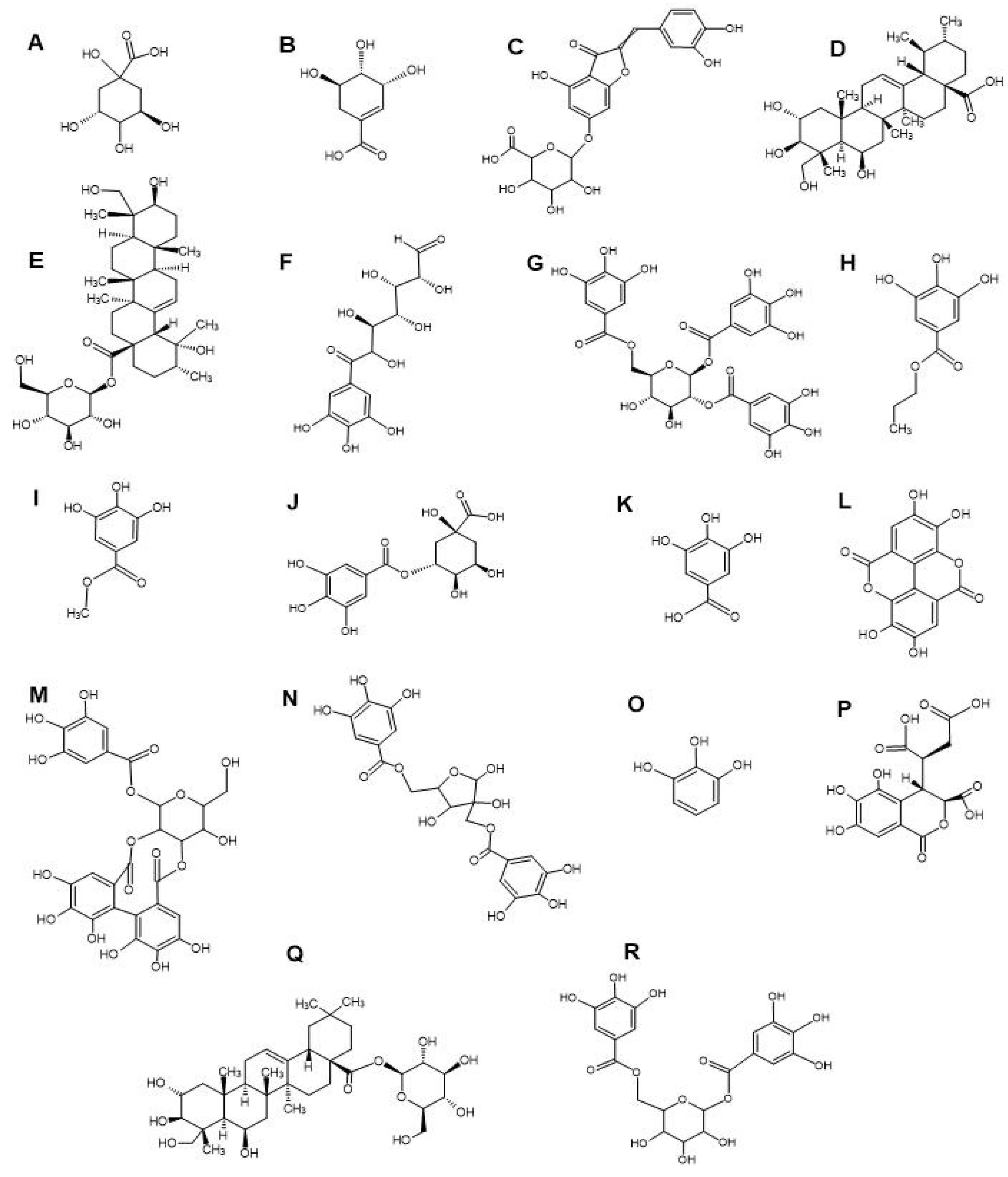

Table 1.
MIC values (µg/mL) for aqueous, methanol, and ethyl acetate of T. bellirica and T. chebula extracts, and the reference (positive control) antibiotics against the six bacterial strains.
Table 1.
MIC values (µg/mL) for aqueous, methanol, and ethyl acetate of T. bellirica and T. chebula extracts, and the reference (positive control) antibiotics against the six bacterial strains.
| Extract Type or Antibiotic | Bacterial species & MIC (µg/mL) | |||||
|---|---|---|---|---|---|---|
| S. aureus | MRSA | E. coli | ESBL E. coli | K. pneumoniae | ESBL K. pneumoniae | |
| TB-AQ | 392 | 196 | 3138 | 1569 | 784 | 3138 |
| TB-MeOH | 94 | 189 | 755 | 755 | 1509 | 755 |
| TB-EtOAc | 900 | 900 | Inactive | Inactive | Inactive | Inactive |
| TCh-AQ | 139 | 139 | 2225 | 2225 | 556 | 2225 |
| TCh-MeOH | 189 | 189 | 3019 | 1509 | 1509 | 1509 |
| TCh-EtOAc | 306 | 306 | Inactive | Inactive | 1225 | Inactive |
| PENG | 1.25 | >2.5 | >2.5 | >2.5 | >2.5 | >2.5 |
| ERY | 0.31 | >2.5 | >2.5 | >2.5 | >2.5 | >2.5 |
| TET | 0.16 | 0.04 | 0.31 | >2.5 | 0.625 | >2.5 |
| CHL | >2.5 | >2.5 | 2.5 | >2.5 | 2.5 | >2.5 |
| CIP | 0.16 | 0.625 | 0.02 | >2.5 | 0.02 | 0.16 |
| POLB | >2.5 | >2.5 | 0.02 | 0.02 | 0.02 | 0.04 |
| OXA | 0.16 | >2.5 | >2.5 | >2.5 | >2.5 | >2.5 |
| AMX | 0.625 | >2.5 | >2.5 | >2.5 | >2.5 | >2.5 |
| GEN | >2.5 | >2.5 | 0.625 | >2.5 | 0.625 | >2.5 |
MIC values for extracts with no antibacterial activity (Inactive; MIC > 10,000 µg/mL), low activity (2000–5000 µg/mL; shown in bold), moderate activity (1000–2000 µg/mL; shown in blue), noteworthy activity (400–1000 µg/mL; shown in red), or good activity (100–400 µg/mL; shown in green), or potent activity (<100 µg/mL; highlighted in yellow). Reference antibiotics: PEN G = penicillin G, ERY = erythromycin, TET = tetracycline, CHL = chloramphenicol, CIP = ciprofloxacin, POLB = polymyxin B, OXA = oxacillin, AMX = amoxycillin, GEN= gentamicin, VAN= vancomycin. Active antibiotics; shown in bold, and no antibiotic activity (MIC >2.5 µg/mL).
Table 2.
∑FIC values calculated for combinations of T. bellirica and T. chebula extracts and conventional antibiotics.
Table 2.
∑FIC values calculated for combinations of T. bellirica and T. chebula extracts and conventional antibiotics.
| Bacteria | Extracts | PENG | ERY | TET | CHL | CIP | POLB | OXA | AMX | GEN | VAN |
|---|---|---|---|---|---|---|---|---|---|---|---|
| S. aureus | TB-AQ | 0.53 | 1.26 | 1.50 | - | 1.50 | - | 1.25 | 0.56 | - | 1.06 |
| TB-MeOH | 1.02 | 2.13 | 1.13 | - | 2.25 | - | 1.25 | 1.03 | - | 2.03 | |
| TB-EtOAc | 0.75 | 1.13 | 1.06 | - | 2.06 | - | 2.13 | 1.25 | - | 1.50 | |
| TChAQ | 1.06 | 1.25 | 0.75 | - | 1.50 | - | 1.50 | 1.13 | - | 1.06 | |
| TCh-MeOH | 1.03 | 1.13 | 1.25 | - | 1.25 | - | 1.25 | 1.06 | - | 1.03 | |
| TCh-EtOH | 0.75 | 0.75 | 1.25 | - | 2.50 | - | 2.50 | 1.00 | - | 1.50 | |
| MRSA | TB-AQ | - | - | 1.00 | - | 1.06 | - | - | - | - | 1.03 |
| TB-MeOH | - | - | 1.50 | - | 1.03 | - | - | - | - | 1.02 | |
| TB-EtOAc | - | - | 1.02 | - | 0.63 | - | - | - | - | 1.50 | |
| TCh-AQ | - | - | 1.00 | - | 2.13 | - | - | - | - | 2.06 | |
| TCh-MeOH | - | - | 1.00 | - | 1.06 | - | - | - | - | 1.03 | |
| TCh-EtOAc | - | - | 1.06 | - | 1.00 | - | - | - | - | 1.50 | |
| E. coli | TB-AQ | - | - | 1.50 | 1.25 | 2.25 | 64.50 | - | - | 2.00 | - |
| TB-MeOH | - | - | 1.52 | 1.06 | 4.00 | 17.50 | - | - | 1.25 | - | |
| TCh-AQ | - | - | 1.50 | 1.25 | 2.25 | 64.50 | - | - | 2.00 | - | |
| TCh-MeOH | - | - | 0.76 | 1.25 | 2.25 | 32.25 | - | - | 2.00 | - | |
| ESBL E. coli | TB-AQ | - | - | - | - | - | 16.50 | - | - | - | - |
| TB-MeOH | - | - | - | - | - | 5.00 | - | - | - | - | |
| TCh-AQ | - | - | - | - | - | 32.50 | - | - | - | - | |
| TCh-MeOH | - | - | - | - | - | 16.50 | - | - | - | - | |
| K. pneumoniae | TB-AQ | - | - | 1.25 | 1.06 | 2.00 | 9.00 | - | - | 1.25 | - |
| TB-MeOH | - | - | 1.50 | 1.12 | 3.00 | 16.50 | - | - | 1.50 | - | |
| TB-EtOAc | - | - | - | - | - | - | - | - | - | - | |
| TCh-AQ | - | - | 1.25 | 1.06 | 4.00 | 9.00 | - | - | 1.25 | - | |
| TCh-MeOH | - | - | 0.75 | 0.56 | 3.00 | 16.50 | - | - | 1.50 | - | |
| TCh-EtOAc | - | - | 0.63 | 1.00 | 2.12 | 63.00 | - | - | 4.00 | - | |
| ESBL K. pneumoniae | TB-AQ | - | - | - | - | 1.25 | 8.25 | - | - | - | - |
| TB-MeOH | - | - | - | - | 1.00 | 2.50 | - | - | - | - | |
| TCh-AQ | - | - | - | - | 2.50 | 16.60 | - | - | - | - | |
| TCh-MeOH | - | - | - | - | 1.50 | 8.75 | - | - | - | - |
∑ FIC values of aqueous (TB-AQ, TCh-AQ), methanolic (TB-MeOH, TCh-MeOH) and ethyl acetate (TB-EtOAc, TCh-EtOAc) extracts of T. bellirica and T. chebula, respectively, in combination with reference antibiotics against antibiotic-sensitive and resistant strains of S. aureus, MRSA, E. coli, ESBL E. coli, K. pneumoniae, and ESBL K. pneumoniae. Additive interaction: >0.5≤1.00, Indifferent interaction: >1.01–≤4.00, Antagonistic interaction: > 4.0.—indicates the extract and/or the antibiotic was inactive against the bacteria being tested. Reference antibiotics: PEN G = penicillin G, ERY = erythromycin, TET = tetracycline, CHL = chloramphenicol, CIP = ciprofloxacin, POL B = polymyxin B, OXA = oxacillin, AMX = amoxycillin, GEN = gentamicin, and VAN = vancomycin.
Table 3.
LC-MS putative identification and % relative abundance of phytochemicals identified in the fruit extracts of T. bellirica (TB) and T. chebula (TCh).
Table 3.
LC-MS putative identification and % relative abundance of phytochemicals identified in the fruit extracts of T. bellirica (TB) and T. chebula (TCh).
| Retention Time (min) | Molecular Weight | Empirical Formula | Putative Compounds | % Relative Abundance (TB) | % Relative Abundance (TCh) | ||||
|---|---|---|---|---|---|---|---|---|---|
| AQ | MeOH | EtOAc | AQ | MeOH | EtOAc | ||||
| 1.496 | 192.06305 | C7H12O6 | Quinic acid | 3.13% | 0.33% | 0.19% | 0.39% | - | 1.53% |
| 1.573 | 344.07409 | C14H16O10 | Theogallin | - | 3.47% | - | - | 0.03% | 0.02% |
| 1.784 | 302.06328 | C12H14O9 | Pyrogallol-2-O-glucuronide | - | 0.21% | - | 0.07% | - | - |
| 1.793 | 244.0581 | C10H12O7 | 1-O-Galloylglycerol | - | - | 0.02% | - | - | - |
| 1.905 | 236.0684 | C12H12O5 | Dillapional | 0.01% | - | - | - | T | 0.01% |
| 1.923 | 316.0793 | C13H16O9 | Ginnalin B | 0.01% | - | - | - | - | - |
| 1.975 | 149.08397 | C9H11NO | Venoterpine | - | - | 0.05% | 0.04% | - | - |
| 2.045 | 294.03749 | C13H10O8 | Banksiamarin B | - | - | 0.15% | 0.17% | 0.05% | 0.13% |
| 2.058 | 174.05268 | C7H10O5 | Shikimic acid | - | 0.41% | 0.67% | 2.64% | 3.14% | 4.80% |
| 2.237 | 154.02646 | C7H6O4 | Gentisic acid | 0.02% | 0.05% | - | 0.01% | - | 0.09% |
| 2.296 | 448.15769 | C19H28O12 | 8-O-Acetyl shanzhiside methyl ester | - | - | - | 0.19% | 0.18% | - |
| 2.356 | 332.07411 | C13H16O10 | 6-Galloylglucose | 6.06% | 3.97% | 3.39% | 3.74% | 0.68% | - |
| 2.444 | 448.06386 | C20H16O12 | Ellagic acid 2-rhamnoside | 0.11% | 1.44% | 0.69% | 0.01% | 0.21% | 0.33% |
| 2.51 | 292.02177 | C13H8O8 | Brevifolincarboxylic acid | 0.09% | - | - | 0.11% | 0.03% | 0.61% |
| 2.56 | 288.0844 | C12H16O8 | Phlorin | - | 0.02% | - | 0.03% | 0.02% | - |
| 3.11 | 484.0848 | C20H20O14 | Gallic acid 3-O-(6-galloylglucoside) | 0.74% | - | - | - | - | - |
| 3.158 | 212.06824 | C10H12O5 | Propyl gallate | - | - | - | 0.30% | 0.35% | 0.12% |
| 4.4 | 277.05821 | C13H11NO6 | Salfredin C1 | - | - | - | 0.14% | - | 0.03% |
| 4.847 | 184.03703 | C8H8O5 | Methyl gallate | 0.25% | 11.39% | - | 0.04% | 1.05% | - |
| 5.638 | 444.19969 | C21H32O10 | Cynaroside A | - | - | - | 0.02% | 0.02% | - |
| 8.848 | 636.09627 | C27H24O18 | 1,3,4-Trigalloyl-β-D-glucopyranose | - | - | - | - | 0.07% | 0.25% |
| 8.44 | 484.0855 | C20H20O14 | 1,6-Bis-O-(3,4,5-trihydroxybenzoyl) hexopyranose | 5.13% | 5.19% | 2.52% | 6.52% | 2.16% | 3.78% |
| 8.961 | 478.07436 | C21H18O13 | Quercetin 7-glucuronide | - | - | - | - | - | 0.49% |
| 8.985 | 478.0743 | C21H18O13 | Quercetin 3-O-glucuronide | - | 0.08% | - | - | - | - |
| 9.316 | 470.01176 | C21H10O13 | Sanguisorbic acid dilactone | - | 0.08% | - | 0.56% | 0.61% | 0.19% |
| 9.567 | 634.08082 | C27H22O18 | Sanguiin H4 | 0.84% | - | 0.07% | - | 5.63% | 0.54% |
| 9.811 | 478.07452 | C21H18O13 | Miquelianin | 0.04% | 0.42% | - | 0.37% | 0.60% | 0.39% |
| 9.902 | 152.04703 | C8H8O3 | Vanillin | - | - | - | 0.16% | 0.17% | 0.17% |
| 9.921 | 126.03146 | C6H6O3 | Phloroglucinol | 0.10% | 1.93% | - | 0.07% | 0.24% | 0.27% |
| 10.043 | 636.09614 | C27H24O18 | 1,2,6-Trigalloyl-β-D-glucopyranose | 0.09% | 5.44% | 2.85% | 0.11% | 1.42% | 2.22% |
| 10.158 | 634.08037 | C27H22O18 | Corilagin | - | - | - | - | 0.15% | - |
| 10.357 | 524.1533 | C24H28O13 | Barbatoflavan | 0.01% | - | - | - | - | - |
| 10.419 | 484.08522 | C20H20O14 | Hamamelitannin | 2.10% | 0.02% | 0.62% | 0.02% | 0.11% | - |
| 10.421 | 126.0316 | C6H6O3 | Pyrogallol | 6.28% | 5.28% | 2.19% | 6.29% | 3.82% | 4.87% |
| 10.422 | 296.05248 | C13H12O8 | cis-Coutaric acid | 0.33% | 0.86% | 0.19% | 0.31% | 0.93% | 1.05% |
| 10.476 | 636.09612 | C27H24O18 | 1,4,6-Trigalloyl-β-D-glucopyranose | - | - | - | - | - | 0.63% |
| 10.55 | 601.99658 | C28H10O16 | Diellagilactone | - | - | - | 1.13% | 0.60% | - |
| 10.703 | 372.1055 | C16H20O10 | Veranisatin C | 0.02% | 0.04% | - | - | - | - |
| 10.754 | 636.09608 | C27H24O18 | 1,3,6-Tri-O-galloyl-β-D-glucose | - | 0.25% | 0.93% | 0.64% | - | - |
| 11.305 | 170.02138 | C7H6O5 | Phloroglucinic acid | 0.98% | 2.80% | 4.33% | 0.98% | 0.83% | 1.24% |
| 11.091 | 610.1535 | C27H30O16 | Rutin | - | 0.14% | 0.10% | - | 0.03% | 0.08% |
| 11.176 | 432.10539 | C21H20O10 | Vitexin | - | - | - | - | 0.05% | - |
| 11.257 | 304.0579 | C15H12O7 | Nigrescin | T | 0.02% | - | - | - | - |
| 11.272 | 610.1527 | C27H30O16 | Quercetin 3-O-rhamnoside-7-O-glucoside | 0.05% | - | - | - | - | - |
| 11.448 | 302.04213 | C15H10O7 | Quercetin | 0.11% | 0.33% | 0.17% | 0.02% | - | - |
| 11.473 | 464.0951 | C21H20O12 | Myricitrin | 0.05% | 0.13% | 0.19% | - | - | - |
| 11.529 | 422.0846 | C19H18O11 | 1,5,8-Trihydroxy-9-oxo-9H-xanthen-3-yl β-D-glucopyranoside | 0.02% | 0.05% | - | - | - | - |
| 11.589 | 170.02147 | C7H6O5 | Gallic acid | 26.23% | 8.64% | 2.60% | 23.90% | 1.90% | 1.56% |
| 11.756 | 216.0994 | C10H16O5 | (4S,5S,8S,10R)-4,5,8-trihydroxy-10-methyl-3,4,5,8,9,10-hexahydro-2H-oxecin-2-one | 0.05% | - | - | 0.06% | - | 1.23% |
| 11.822 | 462.07961 | C21H18O12 | Aureusidin 6-glucuronide | - | - | 2.52% | - | - | 0.04% |
| 11.905 | 286.04729 | C15H10O6 | Maritimetin | - | - | - | T | 0.01% | 0.01% |
| 11.906 | 594.15829 | C27H30O15 | Palasitrin | - | - | - | 0.01% | - | 0.03% |
| 11.938 | 176.04708 | C10H8O3 | 4-Methylumbelliferone hydrate | - | - | - | 0.03% | 0.03% | - |
| 11.953 | 310.10477 | C15H18O7 | (E)-1-O-Cinnamoyl-β-D-glucose | - | - | - | - | - | 1.67% |
| 12.141 | 584.1169 | C28H24O14 | 2”-O-Galloylisovitexin | - | - | - | - | - | 0.01% |
| 12.183 | 448.0996 | C21H20O11 | Maritimein | - | 0.01% | 0.02% | - | - | - |
| 12.185 | 302.00627 | C14H6O8 | Ellagic acid | 2.45% | 10.74% | 33.49% | 8.39% | 10.46% | 11.96% |
| 12.201 | 220.07336 | C12H12O4 | Eugenitin | - | - | - | - | - | 0.04% |
| 12.233 | 262.0476 | C13H10O6 | Maclurin | - | 0.02% | - | 0.04% | - | - |
| 12.254 | 148.05223 | C9H8O2 | trans-Cinnamic acid | - | - | - | 0.76% | 0.96% | 0.87% |
| 12.38 | 334.0325 | C15H10O9 | 3,5,6,7,2′,3′,4′-Heptahydroxyflavone | - | 0.20% | - | - | - | - |
| 12.503 | 192.07847 | C11H12O3 | (R)-Shinanolone | - | - | - | 0.04% | 0.03% | 0.05% |
| 12.504 | 310.10483 | C15H18O7 | (2S,3R,4S,5S,6R)-3,4,5-trihydroxy-6-(hydroxymethyl) oxan-2-yl (2E)-3-phenylprop-2-enoate | - | - | - | 2.25% | - | - |
| 12.509 | 610.1894 | C28H34O15 | Neohesperidin | - | - | 0.16% | - | - | - |
| 12.584 | 498.1741 | C23H30O12 | Eucaglobulin | - | 0.02% | - | - | - | - |
| 12.593 | 436.13688 | C21H24O10 | Nothofagin | - | - | - | - | - | 0.12% |
| 12.721 | 680.37692 | C36H56O12 | Tenuifolin | - | - | - | - | 0.11% | 0.37% |
| 12.8 | 486.33397 | C30H46O5 | Bassic acid | - | - | - | - | - | 0.05% |
| 12.844 | 518.1785 | C26H30O11 | Phellodensin E | - | - | 0.05% | - | - | - |
| 12.889 | 346.1052 | C18H18O7 | Hamilcone | - | - | - | 0.02% | - | 0.01% |
| 12.914 | 190.13544 | C13H18O | β-Damascenone | - | - | - | - | 0.02% | 0.06% |
| 12.915 | 666.39739 | C36H58O11 | Chebuloside II | - | - | - | - | 3.85% | - |
| 12.924 | 302.1152 | C17H18O5 | Lusianin | 0.01% | - | - | - | - | - |
| 12.929 | 356.03763 | C14H12O11 | (+)-Chebulic acid | 0.54% | - | - | 3.98% | 1.40% | 2.88% |
| 12.967 | 330.14651 | C19H22O5 | Hericenone A | - | - | - | - | 0.03% | - |
| 12.969 | 504.34417 | C30H48O6 | Madecassic acid | - | - | - | - | - | 1.42% |
| 12.971 | 202.13522 | C14H18O | (±)-Anisoxide | - | - | - | - | - | 0.05% |
| 12.973 | 244.14583 | C16H20O2 | Lahorenoic acid C | - | - | - | - | - | 0.02% |
| 13.208 | 288.06318 | C15H12O6 | Eriodictyol | - | - | - | - | - | 0.27% |
| 13.354 | 314.1154 | C18H18O5 | Crotaoprostrin | - | - | - | - | - | 0.01% |
| 13.428 | 442.1992 | C25H30O7 | Exiguaflavanone M | - | - | 0.13% | - | - | - |
| 13.443 | 460.13716 | C23H24O10 | 7-Hydroxy-5,6-dimethoxyflavone 7-glucoside | - | - | - | - | 0.01% | - |
| 13.451 | 274.08371 | C15H14O5 | Phloretin | - | - | - | 0.01% | 0.02% | 0.01% |
| 13.536 | 462.11641 | C22H22O11 | Leptosin | - | 0.01% | - | 0.01% | 0.06% | - |
| 13.59 | 302.04255 | C15H10O7 | Bracteatin | - | - | - | 0.01% | 0.01% | - |
| 13.592 | 450.07961 | C20H18O12 | Quercetin 4′-galactoside | - | - | - | 0.01% | - | - |
| 13.593 | 502.1836 | C26H30O10 | Flavaprin | - | - | 0.07% | - | - | - |
| 13.642 | 514.18403 | C27H30O10 | Baohuoside 1 | - | - | - | T | 0.01% | - |
| 13.678 | 568.12178 | C28H24O13 | Isoorientin 2”-p-hydroxybenzoate | - | - | - | - | - | 0.04% |
| 13.703 | 280.13067 | C15H20O5 | Artabsinolide A | - | - | - | - | - | 0.02% |
| 13.725 | 330.0374 | C16H10O8 | Blighinone | 0.12% | - | - | - | - | - |
| 13.805 | 444.10499 | C22H20O10 | 3′-O-Methylderhamnosylmaysin | - | - | - | 0.04% | 0.12% | 0.08% |
| 13.805 | 292.09447 | C15H16O6 | (S)-Angelicain | - | - | - | 0.05% | - | - |
| 13.806 | 462.11629 | C22H22O11 | 6-O-[(2E)-3-Phenyl-2-propenoyl]-1-O-(3,4,5-trihydroxybenzoyl)-β-D-glucopyranose | - | - | - | 0.08% | 1.02% | 0.18% |
| 14.024 | 500.1674 | C26H28O10 | Ikarisoside A | - | - | 0.03% | - | - | - |
| 14.078 | 534.28285 | C29H42O9 | Corchoroside A | - | - | - | - | - | 0.02% |
| 14.358 | 272.06832 | C15H12O5 | Naringenin | - | - | - | - | - | 0.18% |
| 14.367 | 470.33907 | C30H46O4 | Glycyrrhetinic acid | - | - | - | - | 0.08% | 0.01% |
| 14.561 | 226.1203 | C12H18O4 | Allixin | - | - | 0.03% | - | - | - |
| 14.783 | 234.16167 | C15H22O2 | Valerenic acid | - | - | - | - | - | 0.13% |
| 14.928 | 202.17175 | C15H22 | Rulepidadiene B | - | - | - | - | - | 0.08% |
| 14.93 | 222.16153 | C14H22O2 | Rishitin | - | - | - | - | - | 0.04% |
| 15.046 | 426.09477 | C22H18O9 | Epiafzelechin 3-O-gallate | - | - | - | - | - | 0.01% |
| 15.454 | 650.4026 | C36H58O10 | Pedunculoside | - | - | - | - | 1.18% | 2.61% |
| 16.257 | 738.41926 | C39H62O13 | Isonuatigenin 3-[rhamnosyl-(1->2)-glucoside] | - | - | - | - | 0.04% | 0.12% |
| 16.301 | 470.1941 | C26H30O8 | Limonin | - | - | 0.14% | - | - | - |
| 16.478 | 540.1663 | C25H32O11S | Sumalarin B | - | - | 0.02% | - | - | - |
| 16.532 | 316.1308 | C18H20O5 | Methylodoratol | - | - | 0.02% | - | - | - |
| 16.787 | 472.2097 | C26H32O8 | Kushenol H | - | - | 0.07% | - | - | - |
| 16.844 | 344.0532 | C17H12O8 | 3,4,3′-Tri-O-methylellagic acid | - | - | 0.01% | - | - | - |
| 16.876 | 252.20856 | C16H28O2 | Isoambrettolide | - | - | - | - | - | 0.32% |
| 16.889 | 300.1361 | C18H20O4 | Angoletin | - | - | T | - | - | - |
| 17.064 | 440.1828 | C25H28O7 | Lonchocarpol E | - | - | 0.15% | - | - | - |
| 17.209 | 544.2668 | C30H40O9 | Physagulin F | - | - | 0.10% | - | - | - |
| 17.275 | 342.14638 | C20H22O5 | Brosimacutin C | - | - | - | - | T | - |
| 17.276 | 180.11471 | C11H16O2 | Jasmolone | - | 0.61% | 1.48% | 0.05% | - | - |
| 17.31 | 504.34409 | C30H48O6 | Protobassic acid | - | - | - | 1.26% | 0.02% | 0.01% |
| 17.33 | 468.32326 | C30H44O4 | Glabrolide | - | - | - | 0.87% | - | 3.19% |
| 17.33 | 502.32964 | C30H46O6 | Medicagenic acid | - | - | - | - | T | 0.07% |
| 17.331 | 200.15605 | C15H20 | (S)-gamma-Calacorene | - | - | - | 0.02% | - | 0.29% |
| 17.363 | 696.40877 | C37H60O12 | Momordicoside E | - | - | - | - | 0.01% | 0.04% |
| 17.626 | 286.08357 | C16H14O5 | Homobutein | - | T | 0.01% | - | - | 0.01% |
| 17.635 | 282.12541 | C18H18O3 | Ohobanin | 0.01% | T | 0.09% | T | - | 0.06% |
| 17.671 | 652.27302 | C32H44O14 | Dicrocin | - | - | - | - | - | 0.03% |
| 17.902 | 424.1881 | C25H28O6 | Paratocarpin G | T | - | - | - | - | - |
| 17.949 | 456.2144 | C26H32O7 | Antiarone J | - | 0.54% | - | - | - | - |
| 18.124 | 372.2509 | C20H36O6 | Sterebin Q4 | - | - | 0.01% | - | - | - |
| 18.164 | 328.1309 | C19H20O5 | 2′,3′,4′,6′-Tetrameth oxychalcone | - | - | 0.05% | - | - | - |
| 18.237 | 428.1831 | C24H28O7 | Heteroflavanone B | - | - | 0.05% | - | - | - |
| 18.237 | 312.13615 | C19H20O4 | Desmosdumotin C | T | - | 0.02% | T | - | 0.02% |
| 18.292 | 546.35535 | C32H50O7 | Hovenidulcigenin B | - | - | - | 0.04% | - | - |
| 18.339 | 236.1776 | C15H24O2 | Capsidiol | - | - | - | - | - | 0.02% |
| 18.346 | 268.13069 | C14H20O5 | Kamahine C | 0.02% | 0.01% | - | - | 0.01% | 0.15% |
| 18.363 | 336.0994 | C20H16O5 | Ciliatin A | - | - | - | - | 0.01% | - |
| 18.47 | 466.1989 | C27H30O7 | Eriotriochin | - | 0.50% | - | - | - | - |
| 18.498 | 340.09447 | C19H16O6 | Ambanol | - | - | - | - | 0.01% | - |
| 18.507 | 484.31803 | C30H44O5 | Liquoric acid | - | - | - | T | 0.01% | 0.04% |
| 18.623 | 488.35017 | C30H48O5 | Pitheduloside I | - | - | - | T | 0.06% | 0.11% |
| 19.641 | 226.09903 | C15H14O2 | 7-Hydroxyflavan | - | 0.84% | - | 0.03% | - | 0.05% |
| 18.748 | 208.1096 | C12H16O3 | Isoelemicin | 0.01% | - | 0.19% | - | - | - |
| 18.767 | 526.2565 | C30H38O8 | Kosamol A | - | 0.52% | - | - | - | - |
| 18.824 | 300.0997 | C17H16O5 | 2′,4′-Dihydroxy-3,4-dimethoxychalcone | - | - | 0.07% | - | - | - |
| 18.987 | 372.1208 | C20H20O7 | Tangeretin | - | - | 0.03% | - | - | - |
Compounds identified in both plant extracts are in black; compounds identified only in T. bellirica (TB) extracts are in red; compounds identified only in T. chebula (TCh) extracts are in green; compounds less than 0.01% of the total area were considered as trace amounts and denoted as T.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



