
Submitted:
20 September 2024
Posted:
20 September 2024
You are already at the latest version
Abstract
Cardiovascular disease (CVD) is a major global health concern. The number of people with CVD is expected to rise due to aging populations and increasing risk factors such as obesity and diabetes. Identifying new molecular markers is crucial for early diagnosis and treatment. Among these, plasma levels of some miRNAs specifically expressed in cardiac and skeletal muscle known as myomiRs, has gained attention for their roles in cardiovascular health. This study analyzed the plasma levels of myomiR-133a, -133b, and -206 in the pathogenesis of cardiovascular diseases. Exploiting of a case-control study design with patients recruited from several nursing homes form Calabria (Southern Italy) characterized by different types of CVD compared to non-CVD controls, we found downregulation of miR-133a in heart failure and miR-133b in stroke, along with the overall decreased expression of miR-133b and miR-206 in CVD patients. In silico functional characterization of their targets and signaling pathways revealed their involvement in critical cardiovascular processes. Although further research is necessary to fully elucidate their mechanisms and clinical utility, the results here reported support the idea that myomiRs could serve not only as biomarkers but also as therapeutic targets, offering new avenues for managing and potentially preventing CVD progression.
Keywords:
CVD
; cardiovascular disease
; heart failure
; stroke
; miRNA
; myomiRs
; miRNA-133a
; miRNA-133b
; miRNA-206
1. Introduction
Cardiovascular disease (CVD), which refers to a wide range of diseases that affect the heart and blood vessels, including ischemic heart disease, stroke, and heart failure, is one of the leading causes of morbidity and mortality worldwide, accounting for more than 17 million deaths each year [1,2]. This condition appears to be highly prevalent in both developed and developing countries, and the rising prevalence of risk factors such as obesity and diabetes, along with population aging, is expected to increase the global burden of CVDs in the coming decades, bringing a massive burden on public health worldwide.
The pathogenic mechanisms of CVDs are complex and multifactorial, involving a combination of genetic, environmental, and lifestyle factors, and although recent research has been successful in uncovering novel risk factors and the underlying genetic and molecular mechanisms, seeking novel molecular players involved in the development of CVD is crucial to improve early diagnosis in high-risk patients and develop new potential therapeutic targets.
MicroRNAs (miRNAs) are endogenous, conserved, single-stranded non-coding RNAs of 21-25 nucleotides in length that regulate gene expression post-transcriptionally by base pairing to complementary sequences usually located to the 3′-untranslated region (UTR) of target messenger RNAs (mRNAs), leading to their degradation or translational repression [3]. They are important regulatory elements able to fine-tune almost all biological pathways and are essential for maintaining cellular homoeostasis. Consequently, their dysregulation plays a relevant role in the onset and progression of diverse diseases [4,5].
In the cardiovascular system, miRNAs regulate a wide range of biological processes relevant to cardiovascular health, such as inflammation, oxidative stress, fibrosis, angiogenesis, cardiac cell contractility and growth [6,7], thus acting as promoters or suppressors of disease processes. Altered expression of several miRNAs has indeed been associated with several CVDs, holding promise as biomarkers for early disease detection [8,9,10,11,12,13].
Recently, a small group of miRNAs selectively enriched or expressed in cardiac and/or skeletal muscle (also called myomiRs), which includes miRs-1, -133a, -133b, -206, -208a, -208b, -499a, and -499b, has been proved to play important activities in the functional maintenance of the cardiovascular system as regulators of several processes such as proliferation, differentiation, regeneration, and metabolism [14,15,16,17,18]. Evidence for altered myomiRs expression patterns in both human and animal models of cardiovascular related diseases has been reported and summarized in some recent reviews [19,20] and references therein]. To name a few, decreased expression of both miR-1 and miR-133 was found in mouse and human models of cardiac hypertrophy [21] and reported in patients with myocardial infarction [22]. In the same study by Boštjančič and colleagues [22] miR-208 was found upregulated, in agreement with high expression levels in acute myocardial infarction patients found by Han and colleagues [23]. Likewise, it has been reported that circulating miR-499 was substantially elevated in patients with acute myocardial infarction as compared with healthy control groups [24]. In one report, Kumar and colleagues [25] showed that miR-133b was downregulated in plasma samples of coronary artery disease patients.
In the present study the plasma expression patterns of miRNA-1, miRNA-133a, miRNA-133b, miRNA-206, miRNA-208b, and miRNA-499 were investigated in patients with different types of CVD and non-CVD controls, with the aim to further highlight the importance of these miRNAs as contributors to CVD pathogenesis and to explore whether they could be used as biomarkers for discrimination among different forms of CVD.
2. Results
As stated in materials and methods, subjects were classified as having cardiovascular disease (CVD) whether they had one or more of the following physician-confirmed diagnoses: ischemic cardiomyopathy, atrial fibrillation, heart failure and stroke. The demographic and clinical characteristics of the study cohort stratified according to absence/presence of CVD (CVD−/CVD+) are shown in Table 1. Compared to CVD− controls, CVD+ patients were older (p = 0.001), while the ratio of males to females was not significantly different between the two groups (p > 0.05).
Risk factors for CVD such as diastolic pressure, serum total cholesterol, low-density lipoprotein (LDL) cholesterol levels were significantly lower in CVD patients than in control subjects (P<0.01); this is probably due to the use by patients of pharmacological treatments for blood pressure and lipid control. Conversely, levels of uric acid, an emergent biomarker of CVD, as well as marker of kidney dysfunction (urea and creatinine), resulted to be higher in CVD patients compared to controls. Finally, as expected, CVD patients were characterized by a higher mortality rate during the study follow-up (49 % vs 20%).
qRT–PCR was conducted to quantify the levels of miR-1, miR-133a, miR-133b, miR-206, miR-208a, miR-208b, and miR-499 in the plasma of healthy controls and CVD patients. For data analysis, we eliminated miR-1, miR-208b, and miR-499 since the expression levels were below the threshold of detection be quantified reliably and analyzed for differential expression.
First, we explored the association between these miRNAs and the variables in Table 1 in the overall study population. A slightly positive correlation between and age (r = 0.14, p = 0.042) and a very weak negative correlation with the level of albumin and HDL cholesterol (r = -0.15, p = 0.037 and r = -0.17, p = 0.02, respectively) were observed for miR-133a. No other significant correlations between miRNA expression and laboratory data were observed.
We then examined whether the levels of circulating miR-133a, miR-133b, and miR-206 were associated with the diagnosis of CVD. Logistic regression analysis adjusted for age, sex and for statin exposure (Table 2) showed that unitary increase in the expression of miR-133b and miR-206 was associated with a decreased risk of CVD (OR= 0.66; 95% CI= 0.44-0.97; p=0.038 and OR= 0.57; 95% CI= 0.36-0.85; p=0.009 for miR-133b and miR-206 respectively).
Since all subjects were followed-up for a median duration of approximately three years for their overall survival [28 healthy controls (27%) and 40 CVD patients (49%) died during this period], we also evaluate if the detected effects of the analyzed miRNAs on CVD risk might affect survival of the subjects. MiRNAs levels were divided by quartiles and the highest quartile was compared to others. The Kaplan-Meier survival curves are plotted and shown in Figure S1. For all analyzed miRNAs, p-values for both log-rank test and Cox proportional hazards regression analysis adjusted for age and sex indicated no statistically significant differences in all-cause death between “high-expression” and “low/middle-expression” groups.
Next, we performed a sub-analysis to evaluate whether a characteristic miRNA expression profile is linked to a specific cardiovascular condition. The results, reported in Table 3, indicated that, compared with the control group, unitary increase in plasma levels of miR-133a were significantly associated with decreased odds of heart failure by 53% (OR = 0.47, C.I. 0.24-0.95; p=0.031); a similar trend, although not significant at nominal level, was observed for miR-206 (OR= 0.62, C.I.0.37-1.06; p=0.074). Furthermore, increase in serum levels of miR-133b was significantly associated with decreased risk of stroke of about 37% (OR = 0.63, C.I. 0.39-0.99; p=0.048).
2.1. In Silico Functional Characterization
We performed miRNA-target enrichment analysis by using MIENTURNET tool setting the three miRNAs as the inputs and using experimentally validated miRNA-target interactions (miRTarBase). We retrieved one hundred thirty-nine genes significantly associated with these miRNA markers, see Supplementary Table S1. As this table reports, a total of fifty-seven mRNAs were predicted to be targeted by miR-133a, forty-seven by miR-133b, and thirty-five by miR-206. Fifty-six mRNAs were in common between miR-133a and miR-133b, while four targets were shared between miR-133a and miR-206, and two between miR-133b and miR-206. Three mRNAs were targeted by all three miRNAs. The top-ranked targets of these miRNAs and the number of interactions is indicated in Figure 1a, while the significant interaction network is shown in Figure 1b. Functional enrichment analysis using the database REACTOME by MIENTURNET revealed the main pathways involved with the three miRNAs (Figure 1c). Among these pathways, target genes of miR-133a and miR133b were involved in potassium channels, EGFR signaling, and Jak-stat signaling pathways. The latter pathway was also targeted by miR-206; other targets of this miRNA are involved in the dissolution of fibrin clot and mainly in signaling by the receptor tyrosine kinase MET.
3. Discussion
In recent years, interest has grown in determining the role of miRNAs in cardiovascular biology and in identifying circulating miRNA signatures related to the onset and progression of CVD, a leading cause of worldwide morbidity and mortality.
In the present study, we focused our attention on a subset of miRNAs, known as myomiRs, that are highly enriched in both skeletal and cardiac muscles and that have recently garnered attention for their involvement in most of the CVD-related processes, such as cardiac hypertrophy, cardiac fibrosis, apoptosis, inflammation (see references in the Introduction section). Six myomiRs were selected, but only three of them were detectable and analyzed in the plasma samples, namely miR-133a, miR-133b, and miR-206. The study showed that in the whole sample of patients with CVD (which included ischemic cardiomyopathy, atrial fibrillation, heart failure or stroke) the expression levels of both miR-133b and miR-206 were downregulated compared to those in healthy controls, suggesting that low levels of these miRNAs can have a detrimental effect on cardiovascular risk. When analyzing the levels of these miRNAs in subjects categorized according to specific CVDs, we observed a significant downregulation of miR-133b in patients with stroke and a significant downregulation of miR-133a in patients with heart failure, with a trend in the same direction for miR-206.
Taken all together, these findings suggest that specific molecular targets and cellular mechanisms involved in the action of these miRNAs likely act in response to different cardiac damages to obtain distinct expression patterns in distinct types of CVDs. This view is supported by findings of several studies from different cardiovascular conditions, which highlight the complex and context-dependent roles of these miRNAs in CVD development. In line with our findings, studies have found that patients with stroke have lower levels of miR-133b in their blood compared to healthy individuals, and that increasing miR-133b levels could promote recovery after a stroke [26,27]. Also, it has been reported that downregulation of miR-133b might contribute to the development of ventricular fibrillation in patients with myocardial infarction (MI) [28], as well as to the risk of coronary artery disease (CAD) [25]. By contrast, its upregulation has been reported in patients with ST-segment elevation MI and in a mouse model of MI [29]. Also, association between increased concentration of miR-133b and increased odds of lipid-rich coronary plaque has been reported in patients with stable CAD [30]. Furthermore, consistent with our observations, de Gonzalo-Calvo et al [31] found that patients with heart failure have lower levels of miR-133a compared to healthy individuals. Consistently, it has been reported that synthetic miR-133a has the potential to prevent heart failure in a model of pressure overload [32], and that a miR-133a mimic and miR-133a overexpression significantly decreased heart fibrosis in rats with chronic heart failure [33]. On the other hand, the expression of this miRNA, which plays a role in the pathogenesis of most cardiovascular disease, is either upregulated or downregulated due to the pathological changes in particular cell types or specific CVD conditions. For instance, it is downregulated in patients with MI but, by contrast, its expression level is elevated significantly in patients with acute myocardial infarction (AMI) (for a review see [34]). Regarding miR-206, while it was found to be downregulated when the whole group of patients with CVD was considered, only a slight decrease in its expression levels in the group of patients with heart failure was observed. Although not statistically significant, this fits well with the broad role of this miRNA in regulating heart muscle growth (hypertrophy), which is known to contribute to heart failure [35,36]. Several studies in model organisms evaluated miR-206 expression levels in different cardiac lesions. In rat heart with acute MI, it has been found that miR-206 expression is significantly decreased in infarcted myocardial areas and hypoxia-induced cardiomyocytes compared to non-infarcted areas; further investigation revealed that loss-of-function of miR-206 increased cardiomyocyte apoptosis [37]. On the other hand, Shan et al [38] found miR-206 expression significantly increased in a rat model of MI. Also, studies have shown that miR-206 expression is upregulated in cardiac hypertrophy [35], and that its overexpression in adult cardiomyocytes causes arrhythmia [39].
From all the above discussion, it appears that miRs-133a, -133b, and -206 are likely to exhibit differential expression patterns depending on the type of CVD and the clinical and physiological context. This raises the possibility that they can serve as diagnostic and/or prognostic markers not only for overall CVD but also for specific CVD types, pathophysiological processes and/or clinical outcomes. This possibility only comes through a better understanding of the regulation of these miRNAs, as well as the elucidation of their relevant targets, downstream pathways and biological functions in the context of the cardiovascular system, that presently have not been fully clarified.
The miR-133a forms a cluster with miR-1-2 on chromosome 18 (18q11.2), which is transcribed as a bicistronic transcript regulated by many myocytes’ differentiation factors, including myocardin, a nuclear protein that promotes activation of cardiac gene expression by associating with the transcription factor SRF (serum response factor) [40]. MiR-133b and miR-206 are clustered on chromosome 6 (6p12.2) and transcribed as either a single bicistronic lncRNA, or under certain conditions, from separate promoters to yield separate primary transcripts [41]. The primary transcript encoding miR-133b/miR-206 is induced by Wnt/β-catenin signals [42]. This signaling pathway regulates the stability and nuclear localization of β-catenin and plays an important role cardiac function and cardiac tissue homeostasis [43,44]. Its activation is related to pathological stages after MI, including inflammation, angiogenesis, and fibrosis [45,46].
The target enrichment analysis showed that mRNAs of eleven genes were the most significant interacting targets, some of which deserve to be mentioned for their possible association with the cardiac function and pathophysiological state. Among the mRNAs targeted by both miR-133a and miR-133b were PRDM16, a zing-finger transcription factor whose cardiac-specific deletion results in cardiac hypertrophy, fibrosis, mitochondrial dysfunction, and heart failure [47], the hyperpolarization-activated cyclic nucleotide–gated 4 (HCN4) channel, a transmembrane protein expressed in the cardiovascular system reported to play an important role in heart rate regulation [48], and the glutathione transferase P1 (GSTP1), an enzyme involved in detoxification processes, that has gained attention as a biomarker for myocardial stress, particularly in the context of heart failure [49,50]. Among the mRNAs targeted by miR-133a and miR-206 were Annexin A2 (AnxA2), a Ca2+-dependent phospholipid-binding protein important for heart health that has been linked to mechanisms involved in CVD, such as angiogenesis, thrombosis and lipid metabolism [51,52], and the nuclear receptors NR4A2 that has been reported as associated to impairment of cardiac function and heart failure [53,54]. The results of the functional enrichment analysis showed that validated targets of both miR-133a and miR133b were enriched in biological processes related to regulation of cardiovascular function such as the potassium channels, membrane-spanning proteins that regulate cardiac repolarization and the shape and duration of the cardiac action potential and that are actively involved in the mechanism of several heart diseases [55,56,57] and processes mediated by the EGFR signaling, an important signaling hub associated with vascular physiology whose relevance in cardiovascular health and disease has been shown [58,59,60]. Target genes of all the three miRNAs were enriched in the Janus kinase/signal transducer and activator of transcription (JAK/STAT) pathway. This pathway, regulated by cytokines, triggers multiple signals involved in development, homeostasis and inflammation, participates plays a central role in transducing stress and growth signals in various myocardial injuries such as myocardial apoptosis, hypertrophy, fibrosis and ischemia [46,61,62]. For miR-206 there was also a significant enrichment for gene sets related to the signaling pathways activated by the receptor tyrosine kinase MET, the HGF (hepatocyte growth factor) receptor. MET-dependent signals control heart homeostasis and prevents oxidative stress in normal cardiomyocytes and plays also important cardioprotective roles after heart injuries, such as ischemic injury, by promoting pro-survival effect such as protection from cardiomyocytes apoptosis and autophagy [63,64]. Furthermore, one of the enriched pathways was associated to dissolution of fibrin clot, the final step of the coagulation cascade. Impaired plasma fibrin clot properties characterize patients with acute and chronic manifestations of CVD [65].
This study has many strengths: first, it provides valuable insights into the role of myomiRs in the development of cardiovascular diseases and highlights their potential as biomarkers, contributing to a growing body of evidence that could lead to improved diagnostic and therapeutic strategies for CVD. Second, in silico functional enrichment analysis add robustness to study results, highlighting targets and pathways that are likely involved in the pathophysiological mechanisms in CVD risk; third, this study directly addressed the clinical relevance of circulating miRNA in a real-world population of older nursing home residents, making study findings potentially highly relevant to clinical practice.
The study has also some limitations: the small sample size of the whole cohort and of subgroups of patients with different types of CVDs limit the generalizability of study findings; furthermore, the study cohort consisted of older nursing home residents enrolled in the Calabria region of Southern Italy; while this is relevant for studying cardiovascular disease in the aging population, the results may not be applicable to other age groups and geographic populations, as genetic and environmental factors may influence myomiRs expression.
4. Materials and Methods
4.1. Study Population
The cohort of this retrospective study included 209 individuals, which consisted of 90 patients (31% were males) clinically diagnosed with CVD including ischemic cardiomyopathy (52%), atrial fibrillation (29%), heart failure (18%) and stroke (31%) and a control group of 119 healthy controls (28% were males). The mean age of the study population was 85 ± 6.3 years for patients with CVD and 82 ± 7.8 years for the healthy controls.
All participants were recruited from nursing homes located in the provinces of Crotone and Cosenza, Calabria region (southern Italy), as part of a larger study aimed at assessing and monitoring the quality of aging in the entire region. Subjects were eligible to participate in the study if they were of Calabrian ancestry.
4.2. Clinical and Laboratory Assessments
Each participant underwent a complete clinical and geriatric assessment designed to evaluate cognitive status, functional abilities, physical health, and social aspects. A structured questionnaire administered by a trained interviewer was used to collect data. Demographic (age, sex) and anthropometric (height, weight, BMI) variables were available for all participants; clinical variables included systolic and diastolic blood pressure (SBP and DBP, respectively), history of hypertension and diabetes, and presence of CVD; the diagnosis of CVD was based on patients’ detailed medical histories, clinical symptoms, imaging studies and physical or laboratory findings as determined by a board-certified cardiologist in accordance with the international clinical guidelines. Additionally, a peripheral blood sample was drawn from each participant for laboratory examinations, including traditional biomarkers and myomiR, assessed via standardized protocols. Traditional biomarkers included fasting lipid levels (total cholesterol, triglycerides, and high- and low-density lipoprotein cholesterol (HDL-c, and LDL-c, respectively), fasting plasma glucose, HbA1c, urea, creatinine, electrolytes, uric acid, total proteins, albumin, C-reactive protein (CRP), iron, ferritin, and total bilirubin.
4.3. Plasma RNA Extraction
The peripheral blood specimens from all participants were collected in an EDTA-containing tube and centrifuged at 12,000 rpm for 15 min. Afterward, the plasma was stored in a freezer at −80°C until analysis. The MiRNeasy Serum/Plasma Kit (Qiagen, Germany) was used to extract the total RNA from 200 µL of the plasma. In brief, QIAzol lysis buffer (1000µl) were added to the sample to facilitate lysis. Upon addition of 200 µL of chloroform and 3.5 μL of Arabidopsis thaliana miR-159a (assay ID 000338) as a spike-in control, the lysate was separated into an organic and aqueous phase by centrifugation, with RNA partitioning to the upper aqueous phase which was separated and mixed with 1.5 volumes of 100% ethanol and purified on the RNeasy MinElute spin column provided in the kit. RNA was eluted in 14 µL of RNase-free water. The RNA was quantified on a Qubit 2.0 Fluorometer (Life Technologies, Milan, Italy). The yield was around 30–50 ng/mL each sample.
4.4. Quantitative Real-Time Polymerase Chain Reaction
5 μL of RNA was reverse transcribed to cDNA using TaqMan® microRNA Reverse Transcription Kit (Life Technologies) and stem–loop specific RT primers for each of the miRNA examined: hsa-miR-1 (ID 002222); hsa-miR-133a (ID 002246); hsa-miR133b (ID 002247); hsa-miR-206 (ID 000510); hsa-miR-208b (ID 002290); hsa-miR-499 (ID 001045). RNU6B (U6) (ID 001973) was used as an endogenous control to normalize the miRNA qPCR data.
After incubation, quantitative real-time PCR reactions were performed using the TaqMan MicroRNA Assay kit (Applied Biosystems). Each PCR reaction contained 1.33 μl of the RT reaction product, 10 μl of TaqMan 2x Universal PCR Master Mix without uracil-N-glycoslyase (UNG), and 1 μl of 20x TaqMan MicroRNA Assay reagent in a total volume of 20 μl. Real-time reaction was carried out at 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s and 60 °C for 60 s., using the QuantStudio3™ Real-Time PCR System (Applied Biosystems, Milan, Italy) with automatic baseline setting. All reactions, including the no-template controls, were run in triplicate. The relative expression levels of each miRNA in comparison with the normalizer were calculated using the comparative threshold (Ct) method 2−ΔCt [66]. Expression levels (2−ΔCt) were log-transformed to better fit a normal distribution.
4.5. Pathway Enrichment Analysis of the Predicted Targets
The Mienturnet (MicroRNA ENrichment TURned NETwork) web tool (http://userver.bio.uniroma1.it/apps/mienturnet/) [67] was used to access the miRTarBase 9.0 database to search for experimentally validated target genes of the myomiRs analyzed in our study, and to generate a graphical representation of the network of miRNA-target interactions identified by the enrichment analysis. Networks were constructed considering miRNA-mRNA interactions with strong functional level of evidence (such as Western blot and luciferase assay), a threshold of 1 for the minimum number of miR–target interactions and FDR < 0.05. The Reactome database was used for pathway enrichment analysis to search pathways related to miRNA targets with strong empirical evidence and FDR < 0.01. Mienturnet tool also allows the graph of the enriched plot, where each dot indicates one miRNA whose targets are participate in the corresponding pathway. The dots are colored according to the p-adjusted value and sized according to the ratio of genes to pathways.
4.6. Statistical Analysis
The descriptive data were presented as means (SD) for continuous variables and percentages for categorical variables. The Shapiro–Wilk test was used to evaluate the distribution of the variables. Demographic and clinical variables were compared between groups using independent sample t-test for continuous variables and Fisher's exact test for categorical variables. Correlation analyses were performed using Spearman’s correlation coefficient to determine the magnitude of association between plasma miRNA levels and biochemical markers. A binary logistic regression analysis was used to evaluate the association between presence of CVD and the variability of the assessed plasma miRNA levels. Multivariate analysis was performed after adjustment for age, sex, and assumption of statin to exclude their effect. Survival data for all-cause of mortality were available for patients, gathered during an average of three years. For each miRNA, Kaplan-Meier survival curves of subjects in the highest quartile were compared to those of remaining quartiles by using the log-rank test. Cox proportional hazard regression analyses were also performed considering age and gender as confounder variables. P < 0.05 was considered statistically significant. All statistical data were analyzed with R (v4.3.3).
5. Conclusions
This study provides further evidence that myomiRs, particularly miR-133a, miR-133b, and miR-206, may play crucial roles in the pathogenesis of various cardiovascular diseases. The downregulation of miR-133a in heart failure and miR-133b in stroke, along with the overall decreased expression of miR-133b and miR-206 in CVD patients, underscores the potential of these miRNAs as diagnostic biomarkers. While the in silico functional characterization of their targets and pathways revealed their involvement in critical cardiovascular processes, further research is necessary to fully elucidate their mechanisms and clinical utility. These results support the idea that myomiRs could serve not only as biomarkers but also as therapeutic targets, offering new avenues for managing and potentially preventing CVD progression.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1 Kaplan-Meier survival curves of the three miRNAs in CVD patients. The p-value refers to the comparison between the highest quartile (Q1) versus the others (Q2–Q4), as assessed using the log-rank test. Table S1: Mienturnet target enrichment results as obtained from miRTarBase. The threshold for the minimum number of miRNA-target interactions was set up to 1.
Author Contributions
Conceptualization, G.R. and P.C.; methodology, P.C., S.D., R.L.G., and E.P; formal analysis, G.R., and A.M.; writing—original draft preparation, G.R.; writing—review and editing, G.R., P.C., A.M., R.L.G., E.P., L.S., S.D., and G.P.; visualization, G.R.; funding acquisition, G.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was co-funded by Italian Ministry of Health (Ministero della Salute) through the program POS Trajectory line 3.1 project FaPerME, by the Next Generation EU ([DM 1557 11.10.2022] and by project Tech4You—Technologies for climate change adaptation and quality of life improvement, n. ECS0000009.
Institutional Review Board Statement
The study was conducted in accordance with the Declara-tion of Helsinki, and the protocol was approved by the local Ethical Committee (Comitato Etico Regione Calabria-Sezione Area Nord) on 2017–10-31 (code n. 25/2017).
Informed Consent Statement
Informed consent was obtained from the study participants or, where appropriate, a relative or legal representative.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon request.
Acknowledgments
We acknowledge co-funding from the Italian Ministry of Health (Ministero della Salute) through the program POS Trajectory line 3.1 project FaPerME, and from the Next Generation EU, in the context of the National Recovery and Resilience Plan, Investment PE8 – Project Age-It: “Ageing Well in an Ageing Society”. This resource was co-financed by the Next Generation EU [DM 1557 11.10.2022] and, Mission 4, Component 2, Investment 1.5, call for the creation and strengthening of ‘Innovation Ecosystems’, building ‘Territorial R&D Leaders’ (Directorial Decree n. 2021/3277)—project Tech4You—Technologies for climate change adaptation and quality of life improvement, n. ECS0000009. The work has been made possible by the collaboration with Gruppo Baffa (Sadel Spa, Sadel San Teodoro srl, Sadel CSsrl, Casa di Cura Madonna dello Scoglio, AGI srl, Casa di Cura Villa del Rosario srl, Savelli Hospital srl,) in the frame of the agreement TECNOLOGIE AVANZATE PER IL MIGLIORAMENTO DEI SERVIZI ALLA PERSONA.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z,; Benjamin, E.J.; Benziger, C.P.; et al. GBD-NHLBI-JACC Global Burden of Cardiovascular Diseases Writing Group. Global Burden of Cardiovascular Diseases and Risk Factors, 1990-2019: Update From the GBD 2019 Study. J Am Coll Cardiol. 2020, 76, 2982-3021.
- Vaduganathan, M.; Mensah, G.; Turco, J.V; Fuster, V.; Roth, G.A. The Global Burden of Cardiovascular Diseases and Risk: A Compass for Future Health. J Am Coll Cardiol. 2022, 80, 2361–2371.
- Filipowicz, W.; Bhattacharyya, S.N., Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nature reviews genetics, 2008; 9, 102-114.
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D., Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as biomarkers in disease: latest findings regarding their role in diagnosis and prognosis. Cells, 2020; 9, 276. [CrossRef]
- Venneri, M.; Passantino, A. MiRNA: what clinicians need to know. European Journal of Internal Medicine, 2023, 113, 6-9.
- Wojciechowska, A.; Braniewska, A.; Kozar-Kamińska, K. MicroRNA in cardiovascular biology and disease. Advances in Clinical & Experimental Medicine 2017, 26, 5. [CrossRef]
- Varghese, L.N.; Schwenke, D.O.; Katare, R. Role of noncoding RNAs in cardiac ageing, Frontiers in Cardiovascular Medicine, 2023, 10, 1142575. [CrossRef]
- Poller, W.; Dimmeler, S.; Heymans, S., Zeller, T.; Haas, J.; Karakas, M.; Leistner, D.M.; Jakob, P.; Nakagawa, S.; Blankenberg, S.; et al. Non-coding RNAs in cardiovascular diseases: diagnostic and therapeutic perspectives. European heart journal, 2018, 39, 2704-2716. [CrossRef]
- Zhou; S.S.; Jin, J.P.; Wang, J.Q.; Zhang, Z.G.; Freedman, J.H.; Zheng, Y., Cai, L. miRNAS in cardiovascular diseases: potential biomarkers, therapeutic targets and challenges. Acta Pharmacol Sin. 2018, 39, 1073–1084. [CrossRef]
- Çakmak, H.A., Demir, M. MicroRNA and Cardiovascular Diseases. Balkan Med J. 2020, 37, 60-71.
- Sessa, F.; Salerno, M.; Esposito, M.; Cocimano, G.; Pomara, C. miRNA dysregulation in cardiovascular diseases: current opinion and future perspectives. International Journal of Molecular Sciences, 2023, 24, 5192. [CrossRef]
- Searles, C.D. MicroRNAs and Cardiovascular Disease Risk. Curr Cardiol Rep 2024, 26, 51–60. [CrossRef]
- Lozano-Velasco, E.; Inácio, J.M.; Sousa, I.; Guimarães, A.R.; Franco, D.; Moura, G.; Belo, J.A. miRNAs in Heart Development and Disease. International Journal of Molecular Sciences, 2024, 25, 1673. [CrossRef]
- Li, X.; Wang, J.; Jia, Z.; Cui, Q.; Zhang, C.; Wang, W.; Chen, P.; Ma, K.; Zhou, C. MiR499 regulates cell proliferation and apoptosis during late-stage cardiac differentiation via Sox6 and cyclin D1. PLoS ONE. 2013, 8, e74504. [CrossRef]
- Zhao, X.; Wang, Y.; Sun, X. The functions of microRNA-208 in the heart. Diabetes Res Clin Pract. 2020;160: 108004. [CrossRef]
- Huang; X.H., Li, J.L., Li, X.Y., Wang, S.X., Jiao, Z.H., Li, S.Q., Liu, J; Ding, J. miR-208a in cardiac hypertrophy and remodeling. Frontiers in Cardiovascular Medicine, 2021, 8, 773314. [CrossRef]
- Zhu, Y.F.; Wang, R.; Chen, W.; Cao, Y.D.; Li, L.P.; Chen, X. miR-133a attenuates cardiomyocyte hypertrophy through inhibiting pyroptosis activation by targeting IKKε. Acta Histochemica, 2021, 123, 151653. [CrossRef]
- Ouyang, Z; Ke, W. miRNA in cardiac development and regeneration. Cell Regeneration 2021, 10.1, 14. [CrossRef]
- Song, Z.; Gao, R.; Yan, B. Potential roles of microRNA-1 and microRNA-133 in cardiovascular disease. Reviews in Cardiovascular Medicine 2020, 21(1), 57-64. [CrossRef]
- Abkhooie, L.; Sarabi, M.M.; Kahroba, H.; Eyvazi, S.; Montazersaheb, S.; Tarhriz, V.; Hejazi, M.S. Potential Roles of MyomiRs in Cardiac Development and Related Diseases. Curr Cardiol Rev. 2021, 17, e010621188335. [CrossRef]
- Care, A.; Catalucci, D.; Felicetti, F.; Bonci, D.; Addario, A.; Gallo, P.; Bang, M.L.; Segnalini, P.; Gu,Y.; Dalton, N.D.; et al. MicroRNA-133 controls cardiac hypertrophy. Nature medicine, 2007, 13, 613-618. [CrossRef]
- Boštjančič, E.; Zidar, N.; Štajer, D.; Glavač, D. MicroRNAs miR-1, miR-133a, miR-133b and miR-208 are dysregulated in human myocardial infarction. Cardiology, 2010, 115, 163-169. [CrossRef]
- Han, Z., Zhang, L.; Yuan, L.; Liu, X.; Chen, X.; Ye, X.; Yang, C., Yan, Z. Change of plasma microRNA-208 level in acute myocardial infarction patients and its clinical significance. Ann Transl Med. 2015, 3, 307. [CrossRef]
- Zhang, L.; Chen, X.; Su, T.; Li, H.; Huang, Q.; Wu, D.; Yang, C.; Han, Z. Circulating miR-499 are novel and sensitive biomarker of acute myocardial infarction. J Thorac Dis 2015, 7:303-308. [CrossRef]
- Kumar, D.; Narang, R.; Sreenivas, V.; Rastogi, V.; Bhatia, J.; Saluja, D.; Srivastava, K. Circulatory miR-133b and miR-21 as Novel Biomarkers in Early Prediction and Diagnosis of Coronary Artery Disease. Genes (Basel). 2020, 11, 164. [CrossRef]
- Yu, Y.; Zheng, Y.; Yu, T.; Bai, J.; Jiang, T. The relationship between serum rhoa and mir-133b in acute ischemic stroke Int. J. of Adv. Res. 2020, 8, 358-363.
- Xin, H.; Li, Y.; Liu, Z.; Wang, X.; Shang, X.; Cui, Y.; Zhang, Z.G.; Chopp, M. MiR-133b promotes neural plasticity and functional recovery after treatment of stroke with multipotent mesenchymal stromal cells in rats via transfer of exosome-enriched extracellular particles. Stem Cells. 2013, 31, 2737–46. [CrossRef]
- Boštjančič, E.; Brandner, T.; Zidar, N.; Glavač, D.; Štajer, D. Down-regulation of miR-133a/b in patients with myocardial infarction correlates with the presence of ventricular fibrillation. Biomedicine & Pharmacotherapy. 2018, 99, 65-71. [CrossRef]
- D’Alessandra, Y.; Devanna, P.; Limana, F; Straino, S.; Di Carlo, A.; Brambilla, P.G.; Rubino, M.; Carena, M.C.; Spazzafumo, L.; De Simone, M.; et al. Circulating microRNAs are new and sensitive biomarkers of myocardial infarction. Eur Heart J, 2010; 31, 2765-2773. [CrossRef]
- Sæther, J.C.; Vesterbekkmo, E.K.; Taraldsen, M.D.; Gigante, B-; Follestad, T.; Røsjø, H.R.; Omland, T.; Wiseth, R.; Madssen, E.; Bye, A. Associations between circulating microRNAs and lipid-rich coronary plaques measured with near-infrared spectroscopy. Sci Rep 2023, 13, 7580. [CrossRef]
- de Gonzalo-Calvo, D.; Martinez-Camblor, P.; Belmonte, T.; Barbé, F.; Duarte, K.; Cowie, M.R.; Angermann, C.E.; Korte, A.; Riedel, I.; Labus J; et al. Circulating miR-133a defines a low-risk subphenotype in patients with heart failure and central sleep apnea: a decision tree machine learning approach. Journal of Translational Medicine, 2023, 21, 742. [CrossRef]
- Modica, J.; Di Mauro, V.; Barandalla-Sobrados, M.; Chavez, S.E.P.; Carullo, P.; Nemska, S.; Anselmo, A.; Condorelli, G.; Iafisco, M.; Miragoli, M.; Catalucci D. Nano-miR-133a replacement therapy blunts pressure overload–induced heart failure. Circulation. 2021; 144, 1973-1976. [CrossRef]
- Sang, H.Q.; Jiang, Z.M.; Zhao, Q.P.; Xin, F. MicroRNA-133a improves the cardiac function and fibrosis through inhibiting Akt in heart failure rats. Biomed. Pharmacother. 2015, 71, 185–189. [CrossRef]
- Xiao, Y.; Zhao, J.; Tuazon, J.P.; Borlongan, C.V.; Yu, G. MicroRNA-133a and myocardial infarction. Cell transplantation 2019, 28, 831-838. [CrossRef]
- Yang, Y.; Del Re, D.P.; Nakano, N.; Sciarretta, S.; Zhai, P.; Park, J.; Sayed, D., Shirakabe, A.; Matsushima, S.; Park, Y:, et al. miR-206 Mediates YAP-Induced Cardiac Hypertrophy and Survival. Circ Res. 2015, 117, 891-904. [CrossRef]
- Caturano, A.; Vetrano, E.; Galiero, R.; Salvatore, T.; Docimo, G.; Epifani, R.; Alfano, M.; Sardu, C.; Marfella, R.; Rinaldi L, Sasso, F.C. Cardiac hypertrophy: from pathophysiological mechanisms to heart failure development. Reviews in Cardiovascular Medicine. 2022, 23, 165. [CrossRef]
- Yan, Y.; Dang, H.; Zhang, X.; Wang, X.; Liu, X. The protective role of MiR-206 in regulating cardiomyocytes apoptosis induced by ischemic injury by targeting PTP1B. Biosci Rep. 2020, 40, BSR20191000. [CrossRef]
- Shan, Z.X.; Lin, Q.X.; Fu, Y.H.; Deng, C.Y.; Zhou, Z.L.; Zhu, J.N.; Liu, X.Y.; Zhang, Y.Y.; Li, Y.; Lin, S.G.; Yu, X.Y. Upregulated expression of miR-1/miR-206 in a rat model of myocardial infarction. Biochemical and biophysical research communications. 2009, 381, 597-601. [CrossRef]
- Jin, Y.; Zhou, T.Y.; Cao, J.N.; Feng, Q.T.; Fu, Y.J.; Xu, X.; Yang, C.J. MicroRNA-206 Downregulates Connexin43 in Cardiomyocytes to Induce Cardiac Arrhythmias in a Transgenic Mouse Model. Heart Lung Circ. 2019, 28, 1755–1761. [CrossRef]
- Wang, D.Z.; Chang, P.S.; Wang, Z.; Sutherland, L.; Richardson, J.A.; Small, E.; Krieg, P.A.; Olson, E.N. Activation of cardiac gene expression by myocardin, a transcriptional cofactor for serum response factor. Cell. 2001. 105, 851-862. [CrossRef]
- Cesana, M.; Cacchiarelli, D.; Legnini, I.; Santini, T.; Sthandier, O.; Chinappi, M.; Tramontano, A.; Bozzoni, I. A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell. 2011, 147, 358-369. [CrossRef]
- Cui, S.; Li, L.; Mubarokah, S.N.; Meech, R. Wnt/β-catenin signaling induces the myomiRs miR-133b and miR-206 to suppress Pax7 and induce the myogenic differentiation program. Journal of cellular biochemistry. 2019, 120, 12740-12751. [CrossRef]
- Ni, B.; Sun, M.; Zhao, J.; Wang, J.; Cao, Z. The role of β-catenin in cardiac diseases. Frontiers in Pharmacology, 2023, 14, 1157043. [CrossRef]
- Balatskyi, V.V.; Sowka, A.; Dobrzyn, P.; Piven, O.O. WNT/β-catenin pathway is a key regulator of cardiac function and energetic metabolism. Acta Physiol (Oxf). 2023, 237, e13912. [CrossRef]
- Fu, W.B.; Wang, W.E.; Zeng, C.Y. Wnt signaling pathways in myocardial infarction and the therapeutic effects of Wnt pathway inhibitors. Acta Pharm. Sin. 2019, 40, 9-12. [CrossRef]
- Zhang, Q.; Wang, L.; Wang, S.; Cheng, H.; Xu, L.; Pei, G.; Wang, Y.; Fu, C.; Jiang, Y.; He, C.; Wei, Q. Signaling pathways and targeted therapy for myocardial infarction. Sig Transduct Target Ther 2022, 7, 78 (2022). [CrossRef]
- Cibi, D.M.; Bi-Lin, K.W.; Shekeran, S.G.; Sandireddy, R.; Tee, N.; Singh, A.; Wu, Y, Srinivasan, D.K.; Kovalik, J.P.; Ghosh, S.; Seale, P.; Singh, M.K. Prdm16 deficiency leads to age-dependent cardiac hypertrophy, adverse remodeling, mitochondrial dysfunction, and heart failure. Cell reports. 2020, 33, 108288. [CrossRef]
- Hennis, K.; Biel, M.; Fenske, S.; Wahl-Schott, C. Paradigm shift: new concepts for HCN4 function in cardiac pacemaking. Pflugers Arch. 2022, 474, 649-663. [CrossRef]
- Andrukhova, O.; Salama, M.; Rosenhek, R.; Gmeiner, M.; Perkmann, T.; Steindl, J.; Aharinejad, S. Serum Glutathione S-Transferase P1 1 in Prediction of Cardiac Function. J. Card. Fail. 2012, 18, 253-261. [CrossRef]
- Simeunovic, D.; Odanovic, N.; Pljesa-Ercegovac, M.; Radic, T.; Radovanovic, S.; Coric, V.; Milinkovic, I.; Matic, M.; Djukic, T.; Ristic, A.; et al. Glutathione Transferase P1 Polymorphism Might Be a Risk Determinant in Heart Failure. Dis. Markers 2019, 2019, 6984845. [CrossRef]
- Méndez-Barbero, N.; San Sebastian-Jaraba, I.; Blázquez-Serra, R., Martín-Ventura, J.L.; Blanco-Colio, L.M. Annexins and cardiovascular diseases: Beyond membrane trafficking and repair. Frontiers in cell and developmental biology, 2002; 10, 1000760. [CrossRef]
- Kayejo, V. G.; Fellner, H.; Thapa, R.; Keyel, P.A. Translational implications of targeting annexin A2: From membrane repair to muscular dystrophy, cardiovascular disease and cancer. Clinical and Translational Discovery, 2023, 3, e240. [CrossRef]
- Liu, H., Liu, P., Shi, X., Yin, D., Zhao, J. NR4A2 protects cardiomyocytes against myocardial infarction injury by promoting autophagy. Cell Death Discov. 2018, 4, 27. [CrossRef]
- Ashraf, S.; Taegtmeyer, H.; Harmancey, R. Prolonged cardiac NR4A2 activation causes dilated cardiomyopathy in mice. Basic Res Cardiol. 2022, 117, 33. [CrossRef]
- Ye, P.; Zhu, Y.R.; Gu, Y.; Zhang, D.M.; Chen, S.L. Functional protection against cardiac diseases depends on ATP-sensitive potassium channels. Journal of cellular and molecular medicine. 2018, 22, 5801-5806. [CrossRef]
- Weisbrod, D. Small and intermediate calcium activated potassium channels in the heart: role and strategies in the treatment of cardiovascular diseases. Frontiers in Physiology 2020, 11, 590534. [CrossRef]
- Lei, M.; Salvage, S.C.; Jackson, A.P.; Huang, C.L.H. Cardiac arrhythmogenesis: roles of ion channels and their functional modification. Frontiers in Physiology, 2024, 15, 1342761. [CrossRef]
- Forrester, S.J.; Kawai, T.; O'Brien, S.; Thomas, W.; Harris, R.C.; Eguchi, S. Epidermal growth factor receptor transactivation: Mechanisms, pathophysiology, and potential therapies in the cardiovascular system. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 627-653. [CrossRef]
- AlShatnawi, M.N.; Shawashreh, R.A.; Sunoqrot, MA.; Yaghi, A.R. A systematic review of epidermal growth factor receptor tyrosine kinase inhibitor-induced heart failure and its management. The Egyptian Journal of Internal Medicine, 2022, 34, 85. [CrossRef]
- Algül, S.; Schuldt, M.; Manders, E.; Jansen, V.; Schlossarek, S.; de Goeij-de Haas, R.; Henneman, A.A.; Piersma, S.R.; Jimenez, C.R.; Michels, M; Carrier, L., et al.EGFR/IGF1R signaling modulates relaxation in hypertrophic cardiomyopathy. Circulation Research, 2023, 133, 387-399. [CrossRef]
- Bolli, R.; Dawn, B.; Xuan, Y.T. Role of the JAK–STAT pathway in protection against myocardial ischemia/reperfusion injury. Trends Cardiovasc. Med. 2003; 13:72–79. [CrossRef]
- Barry, S.P.; Townsend, P.A.; Latchman, D.S.; Stephanou, A. Role of the JAK–STAT pathway in myocardial injury. Trends Mol. Med. 2007; 13:82–89. [CrossRef]
- Gallo, S.; Sala, V.; Gatti, S.; Crepaldi, T. HGF/Met Axis in Heart Function and Cardioprotection. Biomedicines. 2014, 2, 247-262. [CrossRef]
- Gallo, S.; Sala, V.; Gatti, S.; Crepaldi, T. Cellular and molecular mechanisms of HGF/Met in the cardiovascular system. Clin Sci (Lond). 2015, 129, 1173-1193. [CrossRef]
- Ząbczyk, M.; Ariëns, R.A.; Undas, A. Fibrin clot properties in cardiovascular disease: from basic mechanisms to clinical practice. Cardiovascular research, 2023, 119, 94-111. [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3,1101–1108. [CrossRef]
- Licursi, V.; Conte, F.; Fiscon, G.; Paci, P. MIENTURNET: An interactive web tool for microNA-target enrichment and network-based analysis. BMC Bioinform. 2019; 20, 545. [CrossRef]
Figure 1.
miRNA-target enrichment analysis by MIENTURNET. (a) Bar plot of the results of the enrichment analysis obtained as obtained from miTarBASE. In the Y-axis are reported the top ranked target genes of the submitted miRNAs, while the X-axis represents the number of miRNAs targeting them. The color code reflects the adjusted p-values. The threshold for the minimum number of miRNA-target interactions was set up to 1; (b) Graphical representation of the network of miRNA-target interactions identified by the enrichment analysis; (c) Functional Enrichments for miRNA target genes according to REACTOME. Dots are colored by p-adjusted value (FDR) and sized by gene ratio per pathway (i.e., the number of miRNA targets found enriched in each category over the number of total genes associated with that category).
Figure 1.
miRNA-target enrichment analysis by MIENTURNET. (a) Bar plot of the results of the enrichment analysis obtained as obtained from miTarBASE. In the Y-axis are reported the top ranked target genes of the submitted miRNAs, while the X-axis represents the number of miRNAs targeting them. The color code reflects the adjusted p-values. The threshold for the minimum number of miRNA-target interactions was set up to 1; (b) Graphical representation of the network of miRNA-target interactions identified by the enrichment analysis; (c) Functional Enrichments for miRNA target genes according to REACTOME. Dots are colored by p-adjusted value (FDR) and sized by gene ratio per pathway (i.e., the number of miRNA targets found enriched in each category over the number of total genes associated with that category).
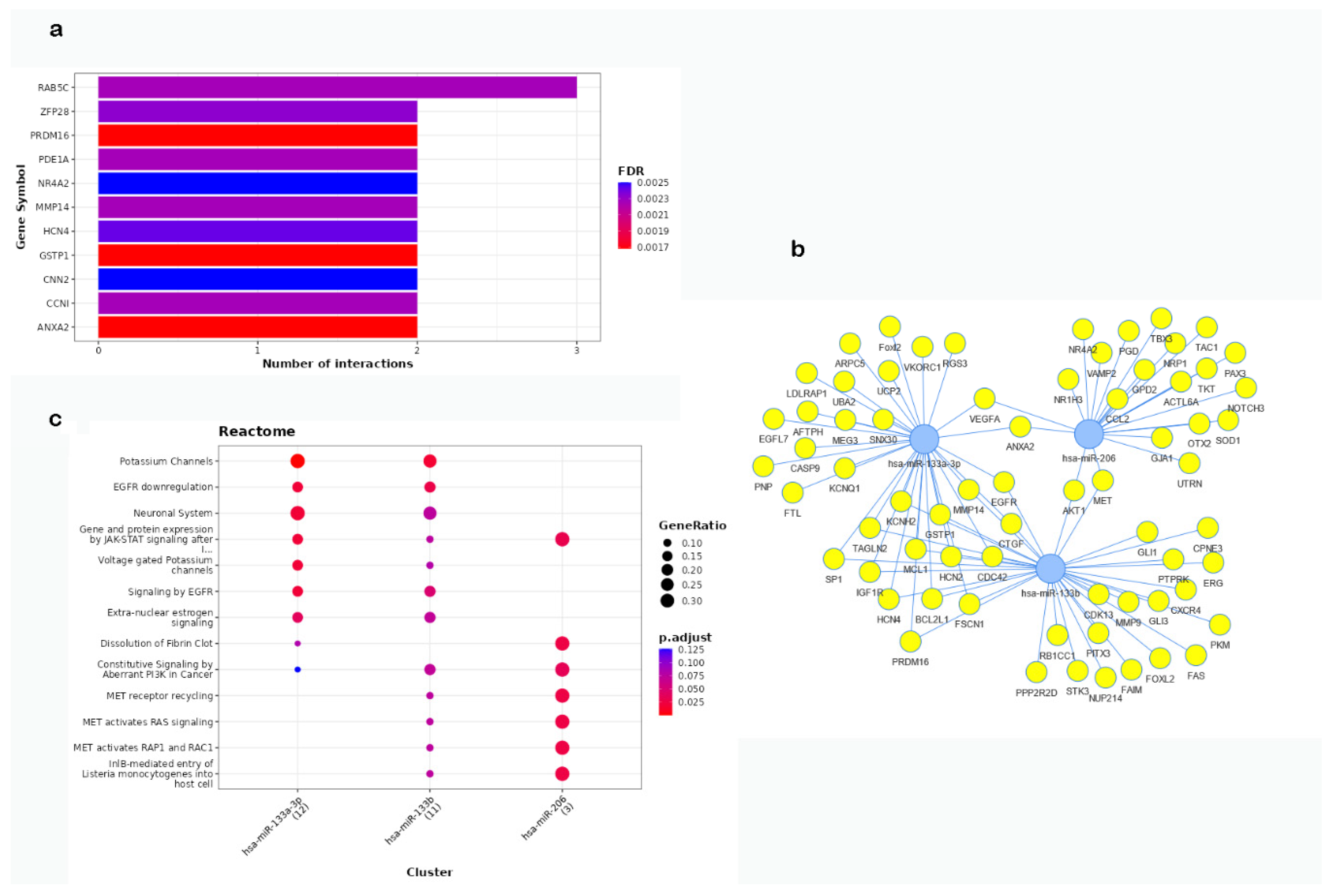

Table 1.
Demographic and clinical characteristics of participants stratified by CVD status.
| Variables | CVD- (N=119) |
CVD + (N=90) |
P-value |
|---|---|---|---|
| Age (mean, SD) | 82 (7.8) | 85 (6.3) | <0.001 |
| Sex (men, %) | 33 (28%) | 28 (31%) | 0.705 |
| BMI, Kg/m2 (mean, SD) | 26 (6.1) | 25 (6.3) | 0.813 |
| SBP, mmHg (mean, SD) | 128 (11) | 126 (13) | 0.353 |
| DBP, mmHg (mean, SD) | 75 (7.7) | 71 (8.2) | 0.010 |
| Total Cholesterol, mg/dl (mean, SD) | 164 (41) | 149 (39) | 0.009 |
| HDL Cholesterol, mg/dl (mean, SD) | 49 (15) | 49 (12) | 0.958 |
| LDL Cholesterol, mg/dl (mean, SD) | 92 (32) | 78 (32) | 0.004 |
| Triglycerides, mg/dl (mean, SD) | 120 (69) | 118 (64) | 0.882 |
| Statin users (Yes, %) | 22 (18%) | 26 (29%) | 0.109 |
| Fasting plasma glucose, mg/dL (mean, SD) | 103 (48) | 98 (28) | 0.333 |
| Glycated Haemoglobin A1c, % (mean, SD) | 6.2 (1.6) | 7 (8.6) | 0.385 |
| Total protein, g/dL (mean, SD) | 6.5 (0.59) | 6.5 (0.64) | 0.556 |
| Albumin, g/dL (mean, SD) | 5.3 (7.6) | 5.3 (6.7) | 0.787 |
| C-reactive Protein, mg/L (mean, SD) | 14 (23) | 17 (35) | 0.498 |
| Urea, mg/dL (mean, SD) | 46 (17) | 54 (29) | 0.019 |
| Creatinine, mg/dL (mean, SD) | 1 (0.3) | 1.2 (0.58) | 0.029 |
| Uric acid, mg/dL (mean, SD) | 4.4 (1.2) | 6 (7.5) | 0.030 |
| Sodium, mM/L (mean, SD) | 141 (2.7) | 140 (2.8) | 0.016 |
| Potassium, mM/L (mean, SD) | 4.4 (0.5) | 4.5 (0.64) | 0.365 |
| Chloride, mM/L (mean, SD) | 105 (3.9) | 101 (15) | 0.014 |
| Calcium, mg/dL (mean, SD) | 9.1 (0.62) | 9 (0.65) | 0.612 |
| Phosphorus, mg/dL (mean, SD) | 3.5 (0.92) | 3.5 (0.64) | 0.962 |
| Magnesium, mg/dL (mean, SD) | 1.9 (0.33) | 1.9 (0.34) | 0.247 |
| Iron, μg/dL (mean, SD) | 56 (26) | 49 (24) | 0.087 |
| Ferritin, ng/mL (mean, SD) | 189 (249) | 160 (228) | 0.432 |
| Total bilirubin, mg/dL (mean, SD) | 0.72 (0.39) | 0.64 (0.31) | 0.112 |
| Hypertension (Yes, %) | 80 (67%) | 73 (81%) | 0.037 |
| Ischemic cardiomyopathy (Yes, %) | - | 47 (52%) | - |
| Atrial fibrillation (Yes, %) | - | 26 (29%) | - |
| Heart failure (Yes, %) | - | 16 (18%) | - |
| Stroke (Yes, %) | - | 28 (31%) | - |
| Deceased (Yes, %) | 28 (27%) | 40 (49%) | 0.004 |
Abbreviations: SD, standard deviation; BMI, Body Mass Index.
Table 2.
Logistic regression analysis of miRNAs levels and CVD risk.
| Predictors | Odds Ratios | CI | p | Odds Ratios | CI | p | Odds Ratios | CI | p |
|---|---|---|---|---|---|---|---|---|---|
| (Intercept) | 0.04 | 0.00 – 2.97 | 0.146 | 0.07 | 0.00 – 5.18 | 0.226 | 0.02 | 0.00 – 2.48 | 0.147 |
| Age | 1.05 | 1.00 – 1.10 | 0.044 | 1.04 | 0.99 – 1.09 | 0.100 | 1.06 | 1.01 – 1.11 | 0.028 |
| Sex | 0.97 | 0.45 – 2.00 | 0.932 | 0.92 | 0.42 – 1.94 | 0.835 | 0.90 | 0.41 – 1.87 | 0.776 |
| Statin | 1.51 | 0.66-3.76 | 0.353 | 1.37 | 0.59 - 3.44 | 0.475 | 1.60 | 0.68 – 4.13 | 0.301 |
| miR-133a | 0.71 | 0.42 – 1.14 | 0.170 | ||||||
| miR-133b | 0.66 | 0.44 – 0.97 | 0.038 | ||||||
| miR-206 | 0.57 | 0.36 – 0.85 | 0.009 |
Table 3.
Association analysis of miRNAs with specific CVDs.
| Ischemic cardiomyopathy |
Atrial fibrillation |
Heart failure |
Stroke | |||||
|---|---|---|---|---|---|---|---|---|
| miRNA | OR (95% CI)# | p* | OR (95% CI)# | p* | OR (95% CI)# | p* | OR (95% CI)# | p* |
| miR-133a | 1.02 (0.63-1.70) | 0.943 | 1.16 (0.64-2.22) | 0.647 | 0.47 (0.24-0.95) | 0.031 | 1.02 (0.58-1.89) | 0.935 |
| miR-133b | 0.84 (0.57-1.23) | 0.378 | 0.85 (0.52-1.38) | 0.517 | 0.82 (0.46-1.44) | 0.495 | 0.63 (0.39-0.99) | 0.048 |
| miR-206 | 1.28 (0.89-1.89) | 0.202 | 1.06 (0.68- 1.73) | 0.795 | 0.64 (0.38-1.08) | 0.090 | 0.82 (0.54-1.24) | 0.332 |
*p-value are adjusted for age, sex, and statin use; p<0.05 was considered significant. # OR, odds ratio; CI, confidence interval.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



