
Submitted:
20 September 2024
Posted:
20 September 2024
You are already at the latest version
Abstract
Urbanization growth has intensified the challenge of managing and treating increasing amounts of municipal solid waste (MSW). Landfills are commonly utilized for MSW disposal because of their low construction and operation costs. However, this practice produces huge volumes of landfill leachate, a highly polluting liquid rich in ammoniacal nitrogen (NH3-N), organic compounds and various heavy metals, making it difficult to treat in conventional municipal wastewater treatment plants (WWTPs). In recent years, research has shown that microbial biofilms, developed on carriers of different materials and called “Moving Bed Biofilm Reactors” (MBBRs), may offer promising solutions for bioremediation. This study explored the biofilm development and the nitrification process of moving bed biofilms (MBBs) obtained from high am-monia-selected microbial communities. Using Crystal Violet Staining and Confocal Laser-Scanning Microscopy, we followed the biofilm formation stages correlating nitrogen removal to metagenomic analyses. Our results indicate that MBBs unveiled a 10-fold more enhanced nitrification rate than the dispersed microbial community pre-sent in the native sludge of the Porto Sant’Elpidio (Italy) WWTP. Four bacterial families, Chitinophagaceae, Comamonadaceae, Sphingomonadaceae, and Nitrosomona-daceae accumulate in structured biofilms and significantly contribute to the high ammonium removal rate of 80% in 24 hours as estimated in leachate-containing wastewaters.
Keywords:
Biological nitrogen removal
; Moving Bed Biofilm (MBB)
; Landfill leachate
; Microbial community
; Metagenomics
1. Introduction
Urbanization and population growth raised the issue of the production and, consequently, the need of appropriate treatment, of increasing amounts of municipal solid waste (MSW). Because of low building and operation costs, landfills are widely used in Italy as well as in developed countries, for MSW disposal. Inevitably, this generates huge volumes of landfill leachate, a highly pollutant liquid containing elevated concentrations of ammonium nitrogen (NH4+-N) and organic compounds in addition to a large spectrum of heavy metals [1,2,3,4,5]. Currently, most of leachates from Italian landfills are classified as intermediate (5-10 years old) and mature (≥ 15 years old) leachates [3]. They are characterized by low content of biodegradable organics due to two major factors: (a) their long staying in landfills; (b) the introduction, occurred in Italy about a decade ago, of the separate waste collection. Thus, these leachates show low biochemical oxygen demand/chemical oxygen demand ratio (BOD/COD ≤ 0.3) and elevated concentrations of ammonia (2000–4000 mg/L). Leachates with such a chemical composition are very hard to be treated in conventional municipal wastewater treatment plants (WWTPs) where nitrogen removal is mainly promoted by the microbial community of the activated sludge [2,6]. In fact, the nitrification-denitrification process involves first the conversion, by autotrophic ammonia-oxidizing bacteria (AOB), of ammonium ion (NH4+) to nitrite (NO2−) which is further oxidized to nitrate (NO3−) by nitrite-oxidizing bacteria (NOB). Finally, NO3− is reduced to gaseous nitrogen compounds by anoxic/anaerobic bacteria which use organic carbon as energy source [7].
In the last two decades, many studies explained why and how planktonic cells start colonizing living and/or inert surface and move to a biofilm lifestyle. A biofilm displays a three-dimensional structure with a highly ordered and complex extracellular matrix [8,9]. The development of mature biofilm confers to the cell population better fitness and protection against a broad range of environmental stress conditions including antimicrobial agents and sanitation processes of food and water. Thus, biofilm formation can generally increase the biological risk to human health [10,11,12]. On the other hand, bacterial biofilms are currently regarded as very promising, innovative and powerful tools in bioremediation techniques. In fact, microbial communities organized into biofilms are much more active than planktonic cells in degrading harmful pollutants such as inorganic and organic compounds, plastic and metals. According to this, microbial proliferation and biomass development are induced on carriers of different materials which are subsequently placed into the reactor. This technology, called Moving Bed Biofilm Reactor (MBBR), was first invented in Norway [13] and now is employed worldwide for the treatment of industrial and municipal wastewater. Biofilm reactors can operate in aerobic, anoxic, or anaerobic conditions and it has been extensively proven that microbes attached to carriers and organized in multi-layered biofilms show an enhanced nitrification-denitrification activity as compared with dispersed biomass. In fact, stratified biofilms allow, through an oxygen gradient, the simultaneous growth of nitrifiers and denitrifiers for complete nitrogen removal whereas organic matter is degraded by aerobic bacteria located mostly in the outer layers of biomass [14,15,16]. Thus, MBBRs are very efficient in ammonia removal in pollutants containing low carbon/nitrogen (C/N) ratio as landfill [17,18,19,20]. Unlike suspended activated sludge reactors, the moving bed biofilm process permits also to fully exploit the volumetric capacity of the tank through the concentration of biomass thereby maximizing the plant performance. In addition, MBBRs offer the advantages of reducing the clogging of bio-carriers and the number of head loss events because they do not require sludge recycling [21,22,23].
In a recent study, the bacterial population of the activated sludge derived from the WWTP of Porto Sant’Elpidio (FM, Italy) was subjected to repetitive ammonium stresses (NH4+-N ≅ 350 mg/L) in a minimal medium supplemented with leachate as only carbon source. These experiments, named named Repetitive Re-Inoculum Assay (RRIA) [24], allowed us to obtain an optimized microorganisms mixture able to efficiently abate the high concentration of ammonia contained in mature landfill leachates. Herein, the formation of moving bed biofilms from these NH4+ selected microbial communities was investigated by Crystal Violet Staining (CVS) and Confocal Laser-Scanning Microscopy (CLSM). Then, the nitrification process promoted by MBBs was estimated in leachate-based medium and correlated to metagenomic analyses performed by 16S rRNA gene next-generation sequencing. Taken together, our results reveal that, four bacterial families, Chitinophagaceae, Comamonadaceae, Sphingomonadaceae and Nitrosomonadaceae were predominant on structured biofilms derived from high NH4+ stress-selected bacterial consortia and largely contribute to the excellent ammonium removal rate (≅ 80 % in 24 hours) as estimated in wastewater-leachate mixtures.
2. Materials and Methods
2.1. Characteristics of Biofilm Carriers
The K1 moving bed biofilm carrier (Kaldnes), used in Bio-RRIA experiments (see below), is characterized by polyethylene high density (HDPE) hollow cylinders with an internal cross and 18 external fins; the diameter is 9.1 mm and the is length 7.2 mm. The density is 140 kg/m3 and the efficient surface area is 500 m2/m3.
Limited to LIVE/DEAD staining and Confocal Laser-Scanning Microscopy analysis, the carriers were HDPE sheets of 15-micron thickness and size of 1 cm2.
2.2. Preparation of Bio-Carriers
Bacterial adhesion to K1 carriers and HDPE sheets was obtained using both selected bacteria populations (G1-2, G5-2 and G5-4) and native sludge [24]. Aliquots of glycerol stock G1-2, G5-2 and G5-4 were plated on solid agar (1.7%) Leachate Minimal Medium (LMM) (see below) at the final concentration of NH4+ of 350 mg/L. After two days of incubation at 30 °C, the bacterial lawns formed were collected with a sterile cotton swab and suspended in 30 mL of LMM. Then, 30 pieces of K1 carriers were immersed in the bacterial suspension. The initial optical density (OD600) was ≅ 0.3 corresponding to ≅ 5x10 7cells/mL as determined by plating bacteria on Luria-Bertani (LB) solid medium after serial dilutions. The cultures were incubated in a rotary shaker at 28°C for 10-12 days, monitoring by Nessler’ test, the decrease of ammonia (NH3). For sludge bio-carriers, 10 mL of the activated sludge (150 mg, dry weight) were mixed with 20 mL of LMM, 30 pieces of K1 carriers and incubated at 28°C following the above-described protocol. The optical density of this culture could not be measured for the high turbidity due to the addition of the sludge and only the colony-forming units (CFU) were determined (≅ 2x106 cells/mL).
The LMM was prepared by combining 60 mL of leachate with 30 mL of MM medium and 90 mL of distilled water. The MM broth is composed of three solutions to be mixed together. Solution 1: 4.95 g (NH4)2SO4, 0.62 g KH2PO4, 0.27 g MgSO4, 0.04 g CaCl2, 0.5 mL FeSO4 (30 mM in 50 mM EDTA at pH 7.0), 0.0002 g CuSO4, 1.2 L distilled water. Solution 2: 8.2 g KH2PO4, 0.7 g NaH2PO4, 3 L distilled water, bring to pH 8.0. Solution 3: 0.6 g Na2CO3, 12 mL distilled water. The three solutions were sterilized by filtering and leachate by autoclave. The ammonia concentration of bacterial cultures was adjusted to the indicated values adding 1 M of (NH4)2SO4. Average composition of pre-treated leachates used in our experiments is reported in Supplementary Table S1.
For determination of colony-forming units (CFU) from bio-carriers, the HPDE supports were subjected to pulsed low-intensity sonication (three cycles of 45 sec at 3.5 Watts) in 3 mL of ice-cold 0.9% NaCl solution to completely remove the bacterial biofilm. Then cells, after adequate dilutions, were plated on LB solid medium and incubate over night at 28°C.
2.3. Crystal Violet Staining
Crystal Violet Staining was carried out essentially as previously described [25,26]. Briefly, K1 bio-carriers were rinsed with 0,9% NaCl solution to eliminate medium residual and incubated with 0.1% (w/v) crystal violet solution in distilled H2O. After 15 minutes at room temperature, the bio-carriers were rinsed four times with distilled H2O to remove unbound dye. The crystal violet was washed out from the biofilms with 4 ml of 95% ethanol and the color intensity was quantified spectrophotometrically at a wavelength of 600 nm [27].
2.4. LIVE/DEAD Staining and Confocal Laser-Scanning Microscopy (CLSM)
The HDPE sheets were fluorescence stained using the LIVE/DEAD®BacLightTM Bacterial Viability Kit according to manufacturer protocol (Invitrogen). Briefly, the two dyes, SYTO9 green fluorescent and propidium iodide (PI) red fluorescent were applied simultaneously as a 1:1000 dilution in distilled water and incubated for 15 minutes at room temperature in a dark room. After washing the stained carriers with distilled H2O to remove the dyes in excess, the fluorescent signals from both live (green) and dead (red) bacteria were visualized simultaneously by a confocal laser scanning microscopy CLSM (Nikon C2Si using the objective PLA APO λ 100x Oil).
2.5. Analytical Methods
Concentration determinations of total nitrogen, ammonia, nitrite, nitrate, biochemical oxygen demand (BOD) and chemical oxygen demand (COD) were carried out as previously reported [24].
2.6. Analysis of Microbial Communities by 16S rRNA Gene Next-Generation Sequencing
Bacterial cells from native sludge (aliquots of 10–20 mL) were harvested by centrifugation (8000 rpm for 20 min) whereas bio-carriers (4-6 tubes) were pulled out of culture medium, washed in 0.9% NaCl solution and subjected to a mild sonication for cells detachment as above described. Then, chromosomal DNA was extracted from all samples by the E.Z.N.A. Kit (Omega Bio-tek Inc., Norcross, GA, USA) according to the instructions given by the manufacturer. DNA concentration was estimated by NanoDrop (ThermoFisher Scientific, Waltham, MA, USA) and 16S rRNA gene next-generation sequencing by MiSeq Illumina Platform was performed as previously reported [24]. Operational taxonomic units (OTUs) were defined by clustering at a minimum pair-wise identity threshold of 97%. The NCBI 16S RefSeq database was employed for taxonomic classification.
3. Results and Discussion
3.1. Monitoring and Quantification of Biofilm Formation
In a recent study, financed by the Eco Elpidiense s.r.l., a private company that manages the municipal wastewater treatment plant of Porto Sant’Elpidio (FM, Italy), we developed a bacteria selection procedure named Repetitive Re-Inoculum Assay (RRIA). This assay, somewhat resembling the dilution to extinction approach [28], consists of consecutive re-inocula/dilutions (7-10 times) of the same bacterial community in a liquid leachate-based medium (LMM) at elevated NH4+-N content (≅ 350 mg/L). RRIA caused a drastic reduction (from ≅ 250 to ≅ 10) of bacterial species present in the activated sludge, causing a remarkable enrichment of those microorganisms characterized by elevated tolerance to recurring ammonia stresses coupled with high activity in nitrogen removal. Aliquots of RRIA cultures have been taken at different times during the two months assay for bacterial species identification by metagenomic analysis and cells storage at -80°C [24].
In the present work, we first established whether our selected microbial communities were able to develop organized biofilms. To this end, K1 HDPE bio-carriers were prepared from both native sludges and RRIA-derived multi-species suspensions to verify the microbial adhesion to these plastic supports by the crystal violet staining (CVS). This colorimetric method is easy, quick to perform and cost-effective although it has the disadvantages to be poorly quantitative and not distinguishing viable and not viable cells [29]. The result, shown in Supplementary Figure S1, was very promising and reveals that bacteria of all the processed samples stably colonize a large surface of K1 polyethylene supports as visible from the intense blue colour. In fact, G5-4, G1-2 and sludge bio-carriers, after ethanol wash and measure of related optical densities, displayed values of A600 nm about 6 times higher than that of controls (carriers not immersed in cell cultures) suggesting that this plastic material was a suitable carrier to be use in our planned MBBs experiments. According to Irankhah et al. [30] investigating MBB reactors from mixed cultures, we found comparable optical densities (A600 nm ≅ 2.0) in CVS assays in addition to the common observation of an evident cell adhesion at the air–liquid interface of the flasks wall in all experiments.
Furthermore, biomass accumulation was also estimated as a function of time during biofilm formation, for G5-2 and native sludge HDPE carriers, using LIVE/DEAD staining and Confocal Laser-Scanning Microscopy (CLSM) (Figure 1). For image acquisitions, bio-carriers were taken out from bacterial suspensions in LMM broth on the 5th, the 10th and the 20th days following the ammonia degradation. On the 20th day, the remaining bio-carriers were re-inoculated in a fresh medium (NH4+-N concentration ≅ 350 mg/L) and incubation was prolonged for additional 10 days at 28°C. The selective staining coupled with CLSM clearly visualized the predominance, across all samples, of green fluorescent labeled cells (living ones) demonstrating that bacteria were abundant and for a very large majority vital; otherwise, red fluorescent cells (dead ones) were almost completely absent. According to the random acquisition of images, the cells number seemed to be slightly higher for G5-2 than sludge MBBs in the first 20 days of biofilm development whereas such difference disappeared for longer times (25th and 30th days) when more structured biofilms were formed. Colony-Forming Units (CFU), determined by growing bacteria on LB agar plates after serial dilutions, supported the CLSM analysis revealing that the cells number was similar for both biofilms being ≅ 4x103/HPDE sheet on the 5th day to progressively increase up to ≅ 5x104/HPDE sheet on the 30th day. Sludge and G5-2 MBBs showed also a comparable partial nitrification activity. Thus, the NH4+-N concentration diminished from ≅ 350 to ≅ 60 mg/L in 6 days corresponding to a removal rate of 83% (not shown) which was consistent to that estimated with K1 bio-carriers in the initial phase of the Bio-Repetitive Re-Inoculum Assay (see next paragraph).
3.2. Ammonia Degradation by Biofilms Developed from Ammonia-Selected Bacterial Populations and Activated Sludges
Given that biofilm forming ability and biomass measurement was ascertained for our mixed cultures by different methods (Supplementary Figure S1 and Figure 1), G5-2 and G5-4 MBBs were tested for their capability to degrade ammonia in a minimal broth (LMM) supplemented with leachate as only carbon source (Figure 2). These bio-carriers were inoculated in LMM medium and ammonia concentration was estimated at the starting point (time 0) and at regular time intervals for the next 21 days. As bacterial biofilms began to oxidize ammonia and its concentration dropped to ≅ 80 mg/L (falls), bio-carriers were removed, washed and transferred to a fresh leachate broth so that the initial value of NH4+-N (≅ 360 mg/L) was restored (peaks). This step was repeated 10 times. This experiment is essentially the Repetitive Re-Inoculum Assay (RRIA) as previously described by Petrilli et al. [24] with the exception that MBBs were used instead of planktonic cells as in the original assay. Thus, it was named Bio-RRIA. As seen in Figure 2A, the two different MBBs showed basically a matching nitrogen removal patterns throughout the entire duration of the assay, characterized by an initial adaptation period in which a lower efficiency in ammonia degradation was observed. In fact, the NH4+ concentration was reduced from ≅ 360 to ≅ 80 mg/L (removal of 78%) in 5 days. From the 6th to the 15th days, the removal rate progressively increased and peaks, that identify the transfer of bio-carriers in a fresh leachate medium with consequent extra NH3 stress, became closer and closer. Thus, after five re-inocula (on the 15th day), MBBs degraded the same amount of ammonia (78%) in only 24 hours exhibiting a 10-fold higher removal activity than respective original microbial cultures. In fact, when G5-2 and G5-4 bacterial populations were grown in planktonic phase, under the same experimental conditions, about 9-11 days were necessary to obtain the ≅ 75% of ammonia oxidation [24].
During the last two decades, the performance of many biofilm-based reactors has been extensively investigated and a bibliometric analysis reveals that ammonia removal varies approximately between 60 and 95 % depending on biofilm system adopted, the type of wastewater, ammonium strength and bio-carried used. Most of these bioreactors operated with an initial ammonia concentration less than 100 mg/L [18,33,34,35,36,37,38]. In this context, our results are promising given that G5-2 and G5-4 MBBs show an excellent removal rate (≅ 80% in 24 hours) with high-strength ammonium (≅ 360 mg/L). According to our standard protocol for bio-carries preparation, the Bio-RRIA assays were highly reproducible with high ammonia-selected MBBs and occasionally small differences, limited to the length of the initial adaptation phase, were found. Another Bio-RRIA, in which the three biofilms G5-2 and G5-4 and G1-2 were compared, showed that, after the usual adaptation period, the one-day removal rate of 67% was achieved (Supplementary Figure S2). Nitrification was slightly lower due to the higher concentration of ammonia (≅ 425 mg/L) used in this Bio-RRIA.
In addition to NH4+, the levels of the most common nitrogenous compounds were estimated during the ammonia oxidation by the bacterial biofilms (Figure 2BC). A progressive accumulation of NO2- was observed for both G5-2 and G5-4 MBBs whereas appreciable amounts of NO3- (≥ 50 mg/L) were produced only in the final part of the Bio-RRIA (after the 15th day). Consistently, the total nitrogen (NT) was only slightly reduced as function of time and its level closely accounted for the sum of the ammonia remained, nitrites, nitrates and not biodegradable organic compounds from leachate. Under the experimental conditions used (leachate-based medium and presence of oxygen), these findings indicate that MBBs mainly promoted nitrification due to the predominant carrier adhesion, selective growth and action of autotrophic and heterotrophic nitrifying bacteria (see metagenomic analysis). Importantly, biofilm enables bacteria with different nutritional requirements to coexist by occupying distinct layers and niches within stratified biofilms where inorganic/organic carbon, oxygen and various substrates can be exchanged by bacteria. In this context, multicomponent biofilms, as utilized in our experiments, are more efficient than single-species biofilms in bioremediation.
The municipal wastewater treatment plant (WWTP) of the Porto Sant’Elpidio town is located in the central Italy near the Adriatic Sea coast where the climate is very mild and, particularly in the last years due to the global warming, the temperature in winter rarely falls below 5-10°C. A detailed description of this WWTP has been previously provided [24]. Thus, the effect of the temperature on the nitrification process by G1-2, G5-2, and G5-4 biofilms was investigated in the range 10-30°C (Figure 3). To this purpose, bio-carriers were taken from the final phase (≅ after 20 days) of a preparative Bio-RRIA where biofilms were already formed and the ammonia removal rate was optimal (≅ 78% in 24 hours). Then, MMBs were placed in fresh leachate-based medium to start a new Bio-RRIA at four different temperatures. The nitrogen removal rate remained constant at 30°C and 23°C whereas slowed down ≅ 3-times (78% in 72 hours) at 18°C for all the samples. By contrast, biofilms nitrification activity was considerably reduced at 10°C (75% in 14 days). Importantly, the wastewater temperature in the two nitrification/denitrification tanks (1,450 m3 each) of the Porto Sant’Elpidio WWTP drops down 15-20°C only in January and February as monitored by the Eco Elpidiense (FM, Italy), the private company that manages this municipal wastewater bioreactor. Thus, the finding that G1-2, G5-2 and G5-4 MBBs retain their maximum performance at 23°C with a limited loss of activity at 18°C, is very encouraging for their immediate application in the pilot reactor of 2,000 liters, already available at the Eco Elpidiense Company. Depending on pilot reactor results, we are confident that our approach derived from MBB technology, implemented and transferred on large-scale could be used to improve, in a near future, the Porto Sant’Elpidio municipal bioreactor performance by accelerating the biological conversion of ammonia to nitrite. This reaction is surely the most critical step of the entire nitrification-denitrification process by bacteria [39] and importantly, needs a lot of energy to make the aeration system, which injects air into the tanks of WWTP, operating.
Colorimetric assays and CFU (Figure 1 and supplementary Figure S1) showed that, in addition to high ammonia-selected cultures, also the bacterial community present in the native sludge from the Porto Sant’Elpidio MWWTP was able to adhere and form biofilm on HDPE supports. To obtain a representative picture, native sludges were sampled during the last two years taking into account the chemical analyses, performed by the Eco Control Laboratory (FM, Italy), to estimate the municipal bioreactor performance. In fact, the treatment capacity of a municipal plant can vary due to many factors (i.e. season, rainfall, change of equivalent people, etc.) but, in the specific case of the Porto Sant’Elpidio bioreactor, also depends on the quantity and quality of landfill leachate introduced (GP and KP, personal communication). As compared with the high reproducibility of G5-2 and G5-4 MBBs, Bio-RRIA carried out with different sludge-derived biofilms exhibited quite variable ammonia degradation curves (Figure 4 and Supplementary Figure S3). In fact, LS1 MBB plot was characterized by an initial ammonia removal rate of ≅ 60% in 10 days, a value more than 2-fold lower than that of G5-2 and G5-4 MBBs. After this acclimatization period, a progressive increase of the NH4+ oxidation activity and consequent shortening in time was observed as for G5-2 and G5-4 biofilms (Figure 2A). Diversely, SL4 MBB displayed a very poor nitrification capacity (NH4+ removal of ≅ 50% in 15 days) that remained low for the entire length of Bio-RRIA. Accordingly, this sludge sample was withdrawn when the Porto Sant’Elpidio WWTP was malfunctioning and dense and persistent foams were well visible on the surface of wastewater contained in the two nitrification/denitrification tanks. As seen in Figure 4BC, better NH4+ degradation patterns were obtained with SL2 and SL3 sludge-derived biofilms that approached the elevated ammonium removal rate of G5-2 and G5-4 MBBs. In fact, SL2 and SL3 biofilms were lacking a pronounced early adaptation phase and were characterized by an average removal of ≅ 76% in 5-6 days that did not change for all the duration of the Bio-RRIA experiment (30-40 days). All together, these results suggest that, unlike the high ammonia-selected G1-2, G5-2 and G5-4 MBBs, native sludge biofilms display a large variability and are much less reliable to produce biofilms with high efficiency in ammonia oxidation. As seen in Supplementary Figure S3, some sludge MBBs could be partially functional or even unsuccessful.
3.3. Composition of Biofilms Formed from Ammonia-Selected Microbial Communities and Activated Sludges
The microbial composition of G5-2 and G5-4 biofilms, used in the Bio-RRIA experiments, was investigated as a function of time by 16S rRNA NGS analysis. According to the near-coincident NH4+ degradation curves (Figure 2A), the metagenomic analysis of these MBBs led to comparable results in terms of bacterial species identified except for slight differences in their relative abundance. As seen in Figure 5AB, at family level, Chitinophagaceae, Comamonadaceae, Sphingomonadaceae and Nitrosomonadaceae substantially increased over the time becoming the most prevalent in both G5-2 and G5-4 biofilms. In particular, Chitinophagaceae, that did not exceed 1.5% in liquid bacterial cultures and newly formed MBBs (the 1st day), reached 13% and 9% for G5-2 and 25% and 15% for G5-4 on the 10th and 15th day, respectively. Similarly, the Nitrosomonadaceae fraction, which was less than 1% in the first stages of biofilm development (on the 1st day), became 16-18% in both MBBs after 10 days. Comamonadaceae and Sphingomonadaceae changed to a lower extent in G5-2 and G5-4 samples showing a 2-5-fold increment in structured biofilms (on the 10th and 15th days) as compared with planktonic growth cells and MMBs on the 1st day. By contrast, Alcaligenaceae and Xanthomonadaceae families, that were strongly predominant (≅ 10-30% of the all OTUs) in the early points of Bio-RRIA, dramatically decreased in MBBs for longer times (abundance ≤ 5% on the 10th and the 15th days). Notably, except for Chitinophagaceae, the bacterial families identified in our study were also found by Garcia et al. [40] investigating in bench-scale MBB bioreactors how size and geometry of plastic supports affects biofilm structure, function and relative microbial species abundance. Most of these families include many heterotrophic nitrifying bacteria and, particularly Alcaligenaceae and Comamonadaceae have been shown to perform simultaneous nitrification/denitrification in MBB reactors [41]. The biofilm composition at genus level of G5-2 and G5-4 samples is reported in Supplementary Figure S4.
Consistent with the taxa distribution at family and genus levels, we identified three principal bacterial species that exhibited a huge increase in frequency at the more advanced stages of biofilm formation and were reasonably responsible for the progressive increment of the nitrification process (Figure 5CD). Primarily, Nitrosomonas eutropha, one of the most investigated AOB, initially at limit of detection, reached 18% of all OTUs in both G5-2 and G5-4 MBBs on the 15th day. Nitrosomonas sp. was found to tolerate elevated NH4+ concentrations [42] and numerous studies indicated that this bacterium plays a key role in NH4+ degradation in different type of biofilm reactors for wastewater treatment [43,44,45,46,47]. Remarkably, N. eutropha is an autotrophic species which oxidizes ammonia in the presence of oxygen. In this respect, MBBs utilized in our Bio-RRIAs were operating in aerobiosis but in heterotrophic conditions due to the huge quantity of organic matter contained in leachate. Thus, the successful proliferation of N. eutropha can be explained by the fact that organized biofilms constitute a physicochemical barrier which, by allowing a selective penetration of nutrients (i.e. oxygen, inorganic carbon and nitrogen compounds), creates an optimized micro-environment for growth and activity of this autotrophic nitrifier. Furthermore, a considerable biomass accumulation was observed on the 10th also for two genera of Chitinophagaceae. In fact, Ferruginibacter lapsinanis came up to 6% and 11% on G5-2 and G5-4, respectively, and Flavitalea flava to 8% on G5-4. Chitinophagaceae includes many heterotrophic AOB that oxidize ammonia using as energy source the organic matter contained in leachate and it was one of the most represented family identified by Wang et al. [48] studying for several months the nitrogen removal in a pilot-scale reactor. All together our results suggest that, as above discussed, G5-2 and G5-4 biofilms, unlike planktonic cells, can provide multilevel metabolic environments that permit a concomitant autotrophic/heterotrophic nitrification. Finally, Comamonas faecalis, although identified at all times, reached 12% on 10-15 days biofilms possibly leading to the conversion of nitrate, first to nitrite, and then, although at low extent, to gaseous nitrogen compounds. In fact, the denitrification activity of bacteria belonging to this genus has been known for a long time [49] and recently, it has been demonstrated in removing several pollutants that produce NO2- and NO3- as by-products of their degradation [50,51,52,53]. Unexpectedly, the Nitrospira and Nitrobacter genera, including canonical nitrite-oxidizing bacteria and generally prevalent in activated sludges from MWWTPs [54,55,56] were not identified across all MBBs analyzed in this study. The lack of these NOB is possibly due to the unfavorable conditions (high concentrations of ammonium and organic compounds) used in biofilm formation and to the competion with heterotrophic species. Finally, other clades as Castellaniella hirudinis, Paracoccus koreensis, Paracoccus pantotrophus and Stenotrophomonas daejeonensis, were considerably represented at the starting points, while became almost undetectable for longer times of Bio-RRIA. In particular, the drastic OTUs fall of C. hirudinis and S. daejeonensis accounted for the disappearance, at family level, of Alcaligenaceae and Xanthomonadaceae, respectively, (Figure 5).
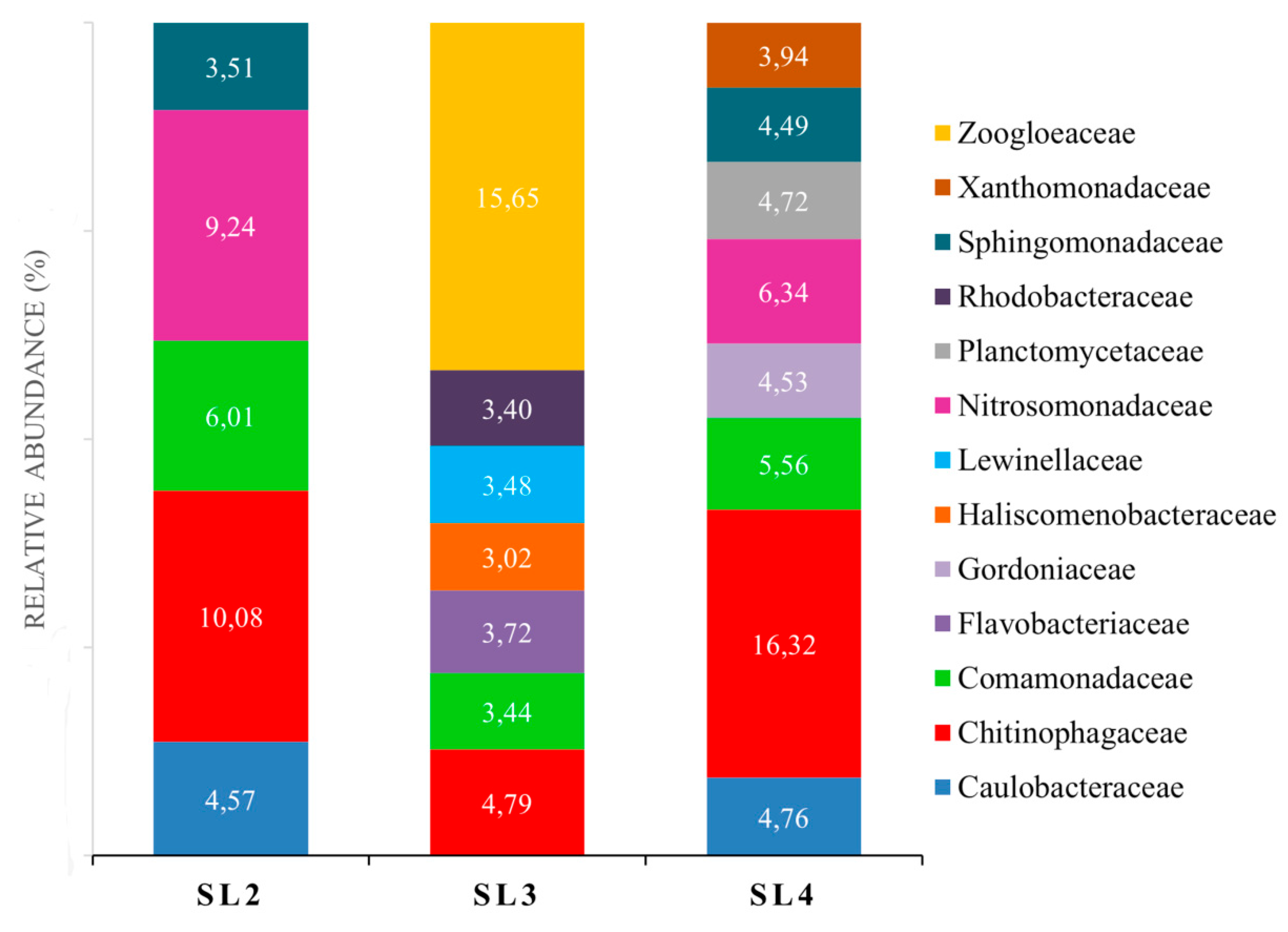
It is well established that, in most of the cases, the poor performance of municipal bioreactors is caused by significant alterations of the microbial population of activated sludges in terms of bacterial species and their relative abundances. Thus, we correlated the ammonia degradation efficiency of SL2, SL3 and SL4 biofilms derived from the Porto Sant’Elpidio MWWTP sludge with their taxa distribution as determined by the metagenomic analysis. As evident in Figure 6, at family level, SL3 MBB was quite dissimilar from the other two sludge biofilms. Specifically, SL3 MBB, which was very effective in NH4+ removal (Figure 4), exhibited the uncommon prevalence (≅ 16% of all the OTUs) of Zoogloeacea and the almost complete lack (< 1%) of Nitrosomonadaceae. Zoogloeacea was not detected either in sludge SL2 and SL4 MBBs or in G5-2 and G5-4 MBBs formed from NH4+-selected microbial communities. Nevertheless, Zoogloeacea accounted for 2.6% of total bacterial communities of activated sludges as shown by metagenomic analyses carried out in 14 MWWTPs in Asia and North America [57]. Notably, the two genera Zoogloea and Thauera represented the totality of members of this family in SL3 biofilm, being 12% and 4%, respectively (not shown). Zoogloea sp. is a heterotrophic/aerobic denitrifier which performs nitrogen removal when cultured in presence of nitrates and nitrites. In addition, Zoogloea. sp. was found to exhibit also a considerable nitrification capacity (NH4+ removal rate of 44%) without nitrate and nitrite production when was grown in a high ammonia-based medium [58]. Thus, the predominance of Zoogloea can account for the elevated ammonia oxidation potential of SL3 MBB in absence of Nitrosomonas sp. Other two families, the Chitinophagaceae and Comamonadaceae, which accumulated in G5-2 and G5-4 biofilms, were identified, at considerable levels, also in sludge MBBs. In particular, Chitinophagaceae, constituted by the only two genera Ferruginibacter and Flavitalea (not shown), became the most abundant family in SL2 (≅ 10%) and SL4 (≅ 16%) MBBs whereas Comamonadaceae was equally represented (≅ 4-6%) across all sludge biofilms (Figure 6). The possible role of these taxa in bioremediation has been already discussed relatively to G5-2 and G5-4 MBBs. As described in the 3.2 section, the SL4 biofilm had a reduced efficiency in ammonia degradation (Figure 4D) according to the fact that sludge sampling was done when the Porto Sant’Elpidio WWTP was not well-performing and an abundant foam was produced during the wastewater treatment. Notably, SL4 MBB contained considerable levels of two genera, Gordonia (4.5%) and Rubinisphaera (4.7%) not identified in the other sludge biofilms. While very little is known on Rubinisphaera (Planctomycetaceae), several members of Gordonia (Gordoniaceae) display a powerful catabolic activity in degrading hazardous pollutants as organic substances, hydrocarbons, sulfur compounds and rubber. Selection of this genus might have been caused by the peculiar and transient conditions related to the malfunctioning of the Porto Sant’Elpidio WWTP at the sampling time. Currently, Gordonia is emerging as a promising candidate, particularly in microbial consortia, to be used in targeted bioremediation techniques [59]. According to SL4 sample, Gordonia strains were detected in stable foams formed in wastewater treatment plants [60,61]. Foams denote a suffering of the bioreactor and usually create, as observed for the Porto Sant’Elpidio WWTP, serious operating problems. In addition, SL4 MBB showed a lower content than SL2 MBB (6% versus 9%) of Nitrosomonadaceae that could be another reason, somewhat explaining the malfunctioning of this sludge biofilm in ammonia oxidation. These findings provide further evidence that the bacterial composition of sludge biofilms strongly affects the nitrification process. Thus, monitoring the sludge microbial community can help to predict with a certain advance and eventually act to limit the possible malfunctioning of the bioreactor.
4. Conclusions
The present study represents an advancement of our recent work [24] in which, through multiple ammonia stresses (Repetitive Re-Inoculum Assay), we strongly enriched the bacterial community of activated sludges to produce an optimized mixture of microorganisms retaining high efficiency in nitrogen removal from landfill leachate. Herein, we proved, using two different techniques (CVS and CLSM), that these high ammonia-selected microbial populations developed stable and organized biofilms on plastic carriers. Importantly, these bio-carriers (G1-2, G5-2 and G5-4 MBBs), after an initial adaptation phase, achieved a very good ammonia removal rate (≅ 80% in 24 hours) in leachate-based medium with high-strength ammonium (≅ 360 mg/L). Metagenomic analysis revealed that four families, Chitinophagaceae (13-25%), Nitrosomonadaceae (16-18%), Comamonadaceae (16-18%) and Sphingomonadaceae (5-7%) accumulated in biofilms at advanced stages of development suggesting their crucial role in the nitrification process. Basically, MBBs exhibited a 10-fold higher activity in NH4+ degradation than respective planktonic growth bacteria due to the biofilm advantage of enabling a synchronous action of autotrophic and heterotrophic nitrifying bacteria.
Therefore, the performance of the Porto Sant’Elpidio MWWTP was monitored for more than two years and MBBs have been produced from the native sludge of this bioreactor. These biofilms evidenced huge variations in activity and a reduced NH4+ degradation as compared with G1-2, G5-2 and G5-4 MBBs. Intriguingly, the most active sludge biofilm obtained (SL3), was characterized by an ammonia removal rate of 78% in 3 days and by a quite rare predominance (16%) of the Zoogloeaceae family. These results, particularly those achieved with the ammonia selection-derived MBBs, are very encouraging and we are planning to quickly transfer this MBB-based technology initially, to the pilot reactor (2000 liters) available at the Eco Elpidiense Company and subsequently, to the Porto Sant’Elpidio plant.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Average composition of pre-treated leachates used in our experiments; Figure S1: Crystal Violet Staining; Figure S2: Bio-Repetitive Re-Inoculum Assay by G1-2 G5-2 and G5-4 MBBs; Figure S3: Bio-Repetitive Re-Inoculum Assay by sludge MBBs; Figure S4: Change of biofilm population from ammonia-selected microbial communities.
Author Contributions
Conceptualization, R. P., A. F., K. P., G. P., V. N. and M. F.; Funding acquisition, M. F.; Investigation, R. P. and M. F.; Methodology, R. P.; Project administration, G. P. and M. F.; Supervision, A. F., G. P. and M. F.; Writing – review & editing, A. F., K. P., G. P., V. N. and M. F.
Funding
This research was funded by the Eco Elpidiense Company, Porto Sant’Elpidio (FM), Italy, grant number [BVC000035- Convenzione Eco Elpidiense].
Acknowledgments
We thank Alessia Cappelli (University of Camerino) for her precious help in using the confocal laser scanning microscope and Dezemona Petrelli (University of Camerino) for providing us the crystal violet dye.
Conflicts of Interest
The authors declare no potential conflicts of interest.
References
- Costa, A.M.; de, R.G.; Alfaia, S.M.; Campos, J.C. Landfill leachate treatment in Brazil–an overview. J. Environ. Manag. 2019, 232, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Yang, G.; Tao, T.; Peng, Y. Recent advances in nitrogen removal from landfill leachate using biological treatments—A review. J. Environ. Manag. 2019, 235, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Wijekoon, P.; Koliyabandara, P.A.; Cooray, A.T.; Lam, S.S.; Athapattu, B.C.L.; Vithanage, M. Progress and prospects in mitigation of landfill leachate pollution: Risk, pollution potential, treatment and challenges. J. Hazard Mater. 2022, 421, 126627. [Google Scholar] [CrossRef] [PubMed]
- Lebron, Y.A.R.; Moreira, V.R.; Brasil, Y.L.; Silva, A.F.R.; Santos, L.V.S.; Lange, L.C.; Amaral, M.C.S. A survey on experiences in leachate treatment: Common practices, differences worldwide and future perspectives. J. Environ. Manag. 2021, 288, 112475. [Google Scholar] [CrossRef]
- Ma, S.; Zhou, C.; Pan, J.; Yang, G.; Sun, C.; Liu, Y.; Chen, X.; Zhao, Z. Leachate from municipal solid waste landfills in a global perspective: Characteristics, influential factors and environmental risks. J. Clean. Prod. 2022, 333, 130234. [Google Scholar] [CrossRef]
- Mai, W.; Chen, J.; Liu, H.; Liang, J.; Tang, J.; Wei, Y. Advances in Studies on Microbiota Involved in Nitrogen Removal Processes and Their Applications in Wastewater Treatment. Front. Microbiol. 2021, 12, 746293. [Google Scholar] [CrossRef]
- Ren, Y.; Hao Ngo, H.; Guo, W.; Wang, D.; Peng, L.; Ni, B.J.; Wei, W.; Liu, Y. New perspectives on microbial communities and biological nitrogen removal processes in wastewater treatment systems. Bioresour. Technol. 2020, 297, 122491. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: an emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563. [Google Scholar] [CrossRef] [PubMed]
- Sauer, K.; Stoodley, P.; Goeres, D.M.; Hall-Stoodley, L.; Burmølle, M.; Stewart, P.S.; Bjarnsholt, T. The biofilm life cycle: expanding the conceptual model of biofilm formation. Nat. Rev. Microbiol. 2022, 20, 608–620. [Google Scholar] [CrossRef]
- Shineh, G.; Mobaraki, M.; Perves Bappy, M.J.; Mills, D.K. Biofilm Formation, and RelatedImpacts on Healthcare, Food Processing and Packaging, Industrial Manufacturing, Marine Industries, and Sanitation–A Review. Appl. Microbiol. 2023, 3, 629–665. [Google Scholar] [CrossRef]
- Sharma, S.; Mohler, J.; Mahajan, S.D.; Schwartz, S.A.; Bruggemann, L.; Aalinkeel, R. Microbial Biofilm: A Review on Formation, Infection, Antibiotic Resistance, Control Measures, and Innovative Treatment. Microorganisms. 2023, 11, 1614. [Google Scholar] [CrossRef] [PubMed]
- Grinberg, M.; Orevi, T.; Kashtan, N. Bacterial surface colonization, preferential attachment and fitness under periodic stress. PLoS Comput. Biol. 2019, 15, e1006815. [Google Scholar] [CrossRef] [PubMed]
- McQuarrie, J.P.; Boltz, J.P. Moving bed biofilm reactor technology: process applications, design, and performance. Water Environ. Res. 2011, 83, 560–575. [Google Scholar] [CrossRef]
- Leyva-Díaz, J.C.; Martín-Pascual, J.; Poyatos, J.M. Moving bed biofilm reactor to treat wastewater. Int. J. Environ. Sci. Technol. 2017, 14, 881–910. [Google Scholar] [CrossRef]
- Di Capua, F.; Iannacone, F.; Sabba, F.; Esposito, G. Simultaneous nitrification-denitrification in biofilm systems for wastewater treatment: Key factors, potential routes, and engineered applications. Bioresour. Technol. 2022, 361, 127702. [Google Scholar] [CrossRef]
- Pan, D.; Shao, S.; Zhong, J.; Wang, M.; Wu, X. Performance and mechanism of simultaneous nitrification–denitrification and denitrifying phosphorus removal in long-term moving bed biofilm reactor (MBBR). Bioresour. Technol. 2022, 348, 126726. [Google Scholar] [CrossRef] [PubMed]
- Kulikowska, D.; Kaczówka, E.; Pokój, T.; Gusiatin, Z. Application of moving bed biofilm reactor (MBBR) for high-ammonium landfill leachate nitrification. New Biotechnol. 2009, 25, S351–S352. [Google Scholar] [CrossRef]
- Zulkifli, M.; Abu Hasan, H.; Sheikh Abdullah, SR.; Muhamad, M.H. A review of ammonia removal using a biofilm-based reactor and its challenges. J. Environ. Manage. 2022, 315, 115162. [Google Scholar] [CrossRef]
- Di Biase, A.; Kowalski, M.S.; Devlin, T.R.; Oleszkiewicz, J.A. Moving bed biofilm reactor technology in municipal wastewater treatment: A review. J. Environ. Manage. 2019, 247, 849–866. [Google Scholar] [CrossRef]
- Xiong, J.; Zheng, Z.; Yang, X.; He, J.; Luo, X.; Gao, B. Mature landfill leachate treatment by the MBBR inoculated with biocarriers from a municipal wastewater treatment plant. Process. Saf. Environ. Prot. 2018, 119, 304–310. [Google Scholar] [CrossRef]
- Madan, S.; Madan, R.; Hussain, A. Advancement in biological wastewater treatment using hybrid moving bed biofilm reactor (MBBR): a review. Appl. Water Sci. 2022, 12, 141. [Google Scholar] [CrossRef]
- Saini, S.; Tewari, S.; Dwivedi, J.; Sharma, V. Biofilm-mediated wastewater treatment: a comprehensive review. Mater. Adv. 2023, 4, 1415–1443. [Google Scholar] [CrossRef]
- Pratap, V.; Kumar, R.; Kumar, S.; Yadav, B.R. Optimization of moving bed biofilm reactors for the treatment of municipal wastewater. Environ. Res. 2024, 241, 117560. [Google Scholar] [CrossRef] [PubMed]
- Petrilli, R.; Fabbretti, A.; Cerretani, A.; Pucci, K.; Pagliaretta, G.; Picciolini, M.; Napolioni, V.; Falconi, M. ; Selection, Identification and Functional Performance of Ammonia-Degrading Microbial Communities from an Activated Sludge for Landfill Leachate Treatment. Microorganisms 2023, 11, 311. [Google Scholar] [CrossRef]
- Djordjevic, D.; Wiedmann, M.; McLandsborough, L. A. Microtiter plate assay for assessment of listeria monocytogenes biofilm formation. Appl.Environ.Microbiol. 2002, 68, 2950–2958. [Google Scholar] [CrossRef] [PubMed]
- Sabaeifard, P.; Abdi-Ali, A.; Soudi, M. R.; Dinarvand, R. Optimization of tetrazolium salt assay for pseudomonas aeruginosa biofilm using microtiter plate method. J. Microbiol. Methods 2014, 105, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Pantanella, F.; Valenti, P.; Natalizi, T.; Passeri, D.; Berlutti, F. Analytical techniques to study microbial biofilm on abiotic surfaces: Pros and cons of the main techniques currently in use. Ann. Ig. 2013, 25, 31–42. [Google Scholar] [CrossRef]
- Franklin, R.B.; Garland, J.L.; Bolster, C.H.; Mills, A.L. Impact of dilution on microbial community structure and functional potential: comparison of numerical simulations and batch culture experiments. Appl. Environ. Microbiol. 2001, 67, 702–712. [Google Scholar] [CrossRef]
- Doll, K.; Jongsthaphongpun, K.L.; Stumpp, N.S.; Winkel, A.; Stiesch, M. Quantifying implant-associated biofilms: Comparison of microscopic, microbiologic and biochemical methods. J. Microbiol. Methods. 2016, 130, 61–68. [Google Scholar] [CrossRef]
- Irankhah, S.; Abdi Ali, A.; Reza Soudi, M.; Gharavi, S.; Ayati, B. Highly efficient phenol degradation in a batch moving bed biofilm reactor: benefiting from biofilm-enhancing bacteria. World J. Microbiol. Biotechnol. 2018, 34, 164. [Google Scholar] [CrossRef]
- Berney, M.; Hammes, F.; Bosshard, F.; Weilenmann, H.-U. U.; Egli, T. Assessment and interpretation of bacterial viability by using the LIVE/DEAD BacLight kit in combination with flow cytometry. Appl. Environ. Microbiol. 2007, 73, 3283–3290. [Google Scholar] [CrossRef] [PubMed]
- Stiefel, P.; Schmidt-Emrich, S.; Maniura-Weber, K.; Ren, Q. Critical aspects of using bacterial cell viability assays with the fluorophores SYTO9 and propidium iodide. BMC Microbiol. 2015, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Chaali, M.; Naghdi, M.; Brar, S.K.; Avalos-Ramirez, A. A review on the advances in nitrifying biofilm reactors and their removal rates in wastewater treatment. J. Chem. Technol. Biotechnol. 2018, 93, 3113–3124. [Google Scholar] [CrossRef]
- Ren, J.; Cheng, W.; Jiao, M.; Wan, T.; Wang, M.; Li, D. Characteristics of oxygen mass transfer and its impact on pollutant removal performance and microbial community structure in an aerobic fluidized bed biofilm reactor for treatment of municipal wastewater. Bioresour. Technol. 2021, 323, 124552. [Google Scholar] [CrossRef]
- Wang, Y.; Du, Z.; Liu, Y.; Wang, H.; Xu, F.; Liu, B.; Zheng, Z. The nitrogen removal and sludge reduction performance of a multi-stage anoxic/oxic (A/O) biofilm reactor. Water Environ. Res. 2020, 92, 94–105. [Google Scholar] [CrossRef]
- Bassin, J.P.; Dias, I.N.; Cao, S.M.S.; Senra, E.; Laranjeira, Y.; Dezotti, M. Effect of increasing organic loading rates on the performance of moving-bed biofilm reactors filled with different support media: assessing the activity of suspended and attached biomass fractions. Process Saf. Environ. Protect. 2016, 100, 131–141. [Google Scholar] [CrossRef]
- Saidulu, D.; Majumder, A.; Gupta, A.K. A systematic review of moving bed biofilm reactor, membrane bioreactor, and moving bed membrane bioreactor for wastewater treatment: Comparison of research trends, removal mechanisms, and performance. J. Environ. Chem. Eng. 2021, 9, 106112. [Google Scholar] [CrossRef]
- Zhao, J.; Ni, G.; Piculell, M.; Li, J.; Hu, Z.; Wang, Z.; Guo, J.; Yuan, Z.; Zheng, M.; Hu, S. Characterizing and comparing microbial community and biofilm structure in three nitrifying moving bed biofilm reactors. J. Environ. Manage. 2022, 320, 115883. [Google Scholar] [CrossRef]
- Sun, H.W.; Peng, Y.Z.; Shi, X.N. Advanced treatment of landfill leachate using anaerobic-aerobic process: Organic removal by simultaneous denitritation and methanogenesis and nitrogen removal via nitrite. Bioresour. Technol. 2015, 177, 337–345. [Google Scholar] [CrossRef]
- Garcia, K.A.; McLee, P.; Schuler, A.J. Effects of media length on biofilms and nitrification in moving bed biofilm reactors. Biofilm. 2022, 4, 100091. [Google Scholar] [CrossRef]
- Janka, E.; Pathak, S.; Rasti, A.; Gyawali, S.; Wang, S. Simultaneous Heterotrophic Nitrification and Aerobic Denitrification of Water after Sludge Dewatering in Two Sequential Moving Bed Biofilm Reactors (MBBR). Int. J. Environ. Res. Public Health. 2022, 19, 1841. [Google Scholar] [CrossRef] [PubMed]
- Fujitani, H.; Kumagai, A.; Ushiki, N.; Momiuchi, K.; Tsuneda, S. Selective isolation of ammonia-oxidizing bacteria from autotrophic nitrifying granules by applying cellsorting and sub-culturing of microcolonies. Front. Microbiol. 2015, 6, 1159. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wang, Y.; Li, R.; Zhang, R.; Zhang, X.; Hua, S.; Peng, D. The differential proliferation of AOB and NOB during natural nitrifier cultivation and acclimation with raw sewage as seed sludge. RSC Adv. 2020, 10, 28277–28286. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Duan, H.; Zhang, Y.; Huang, X.; Yuan, Z.; Liu, Y.; Zheng, M. Adaptation of nitrifying community in activated sludge to free ammonia inhibition and inactivation. Sci. Total Environ. 2020, 728, 138713. [Google Scholar] [CrossRef]
- Phanwilai, S.; Kangwannarakul, N.; Noophan, P.; Kasahara, T.; Terada, A.; Munakata- Marr, J.; Figueroa, L.A. Nitrogen removal efficiencies and microbial communities in full-scale IFAS and MBBR municipal wastewater treatment plants at high COD:N ratio. Front. Environb. Sci. Eng. 2020, 14, 115. [Google Scholar] [CrossRef]
- Kruglova, A.; Kesulahti, J.; Minh Le, K.; Gonzalez-Martinez, A.; Mikola, A.; Vahala, R. Low-temperature adapted nitrifying microbial communities of Finnish wastewater treatment systems. Water 2020, 12, 2450. [Google Scholar] [CrossRef]
- Zhou, L.-J.; Han, P.; Yu, Y.; Wang, B.; Men, Y.; Wagner, M.; Wu, Q.L. Cometabolic biotransformation and microbial-mediated abiotic transformation of sulfonamides by three ammonia oxidizers. Water Res. 2019, 159, 444–453. [Google Scholar] [CrossRef]
- Wang, Z.; Yao, Y.; Steiner, N.; Cheng, H.; Wu, Y.; Woo, S.; Criddle, C.S. Impacts of nitrogen-containing coagulants on the nitritation/denitrification of anaerobic digester centrate. Environ. Sci.: Water Res. Technol. 2020, 6, 3451–3459. [Google Scholar] [CrossRef]
- Patureau, D.; Davison, J.; Bernet, N.; Moletta, R. Denitrification under various aeration conditions in Comamonas sp., strain SGLY2. FEMS Microbiol. Ecol. 1994, 14, 71–78. [Google Scholar] [CrossRef]
- Wang, Z.; Woo, S.; Yao, Y.; Cheng, H.; Wu, Y.; Criddle, C.S. Nitrogen removal as nitrous oxide for energy recovery: Increased process stability and high nitrous yields at short hydraulic residence times. Water Res. 2020, 173, 115575. [Google Scholar] [CrossRef]
- Hassan, H.; Jin, B.; Donner, E.; Vasileiadis, S.; Saint, C.; Dai, S. Microbial community and bioelectrochemical activities in MFC for degrading phenol and producing electricity: microbial consortia could make differences. Chem. Eng. J. 2018, 332, 647–657. [Google Scholar] [CrossRef]
- Shi, J.; Xu, C.; Han, Y.; Han, H. Enhanced anaerobic biodegradation efficiency and mechanism of quinoline, pyridine, and indole in coal gasification wastewater. Chem. Eng. J. 2019, 361, 1019–1029. [Google Scholar] [CrossRef]
- Zheng, M.; Zhu, H.; Han, Y.; Xu, C.; Zhang, Z.; Han, H. Comparative investigation on carbon-based moving bed biofilm reactor (MBBR) for synchronous removal of phenols and ammonia in treating coal pyrolysis wastewater at pilot-scale. Bioresour. Technol. 2019, 288, 121590. [Google Scholar] [CrossRef]
- Spieck, E.; Hartwig, C.; McCormack, I.; Maixner, F.; Wagner, M.; Lipski, A.; Daims, H. Selective enrichment and molecular characterization of a previously uncultured Nitrospira-like bacterium from activated sludge. Environ. Microbiol. 2006, 8, 405–415. [Google Scholar] [CrossRef]
- Mehrani, M.J.; Sobotka, D.; Kowal, P.; Ciesielski, S.; Makinia, J. The occurrence and role of Nitrospira in nitrogen removal systems. Bioresour. Technol. 2020, 303, 122936. [Google Scholar] [CrossRef]
- Cao, Y.; van Loosdrecht, M.C.; Daigger, G.T. Mainstream partial nitritation-anammox in municipal wastewater treatment: status, bottlenecks, and further studies. Appl. Microbiol. Biotechnol. 2017, 101, 1365–1383. [Google Scholar] [CrossRef]
- Zhang, T.; Shao, M.F.; Ye, L. 454 Pyrosequencing reveals bacterial diversity of activated sludge from 14 sewage treatment plants. ISME J. 2012, 6, 1137–1147. [Google Scholar] [CrossRef]
- Huang, T.L.; Zhou, S.L.; Zhang, H.H.; Bai, S.Y.; He, X.X.; Yang, X. Nitrogen Removal Characteristics of a Newly Isolated Indigenous Aerobic Denitrifier from Oligotrophic Drinking Water Reservoir, Zoogloea sp. N299. Int. J. Mol. Sci. 2015, 16, 10038–10060. [Google Scholar] [CrossRef]
- Sowani, H.; Kulkarni, M.; Zinjarde, S. Harnessing the catabolic versatility of Gordonia species for detoxifying pollutants. Biotechnol. Adv. 2019, 37, 382–402. [Google Scholar] [CrossRef]
- Tamura, T.; Saito, S.; Hamada, M.; Kang, Y.; Hoshino, Y.; Gonoi, T.; Mikami, Y.; Yaguchi, T. Gordonia crocea sp. nov. and Gordonia spumicola sp. nov. isolated from sludge of a wastewater treatment plant. Int. J. Syst. Evol. Microbiol. 2020, 70, 3718–3723. [Google Scholar] [CrossRef]
- Riesco, R.; Rose, J.J.A.; Batinovic, S.; Petrovski, S.; Sánchez-Juanes, F.; Seviour, R.J.; Goodfellow, M.; Trujillo, M.E. Gordonia pseudamarae sp. nov., a home for novel actinobacteria isolated from stable foams on activated sludge wastewater treatment plants. Int. J. Syst. Evol. Microbiol 2022, 72. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Analysis of biofilm formation by CLSM. Staining of sludge (A) and G5-2 (B) MBBs was carried out with SYTO9 and PI fluorescent dyes followed by CLSM analysis as described in Materials and Methods. Both dyes intercalate with nucleic acids but while SYTO9 penetrates both living and dead cells, PI can only pass through damaged membranes and displaces SYTO9 allowing differentiation between live (green) and dead (red) cells [31,32]. Stacks of images were taken at random areas on the HPDE sheets where each stack contained 10 images.
Figure 1.
Analysis of biofilm formation by CLSM. Staining of sludge (A) and G5-2 (B) MBBs was carried out with SYTO9 and PI fluorescent dyes followed by CLSM analysis as described in Materials and Methods. Both dyes intercalate with nucleic acids but while SYTO9 penetrates both living and dead cells, PI can only pass through damaged membranes and displaces SYTO9 allowing differentiation between live (green) and dead (red) cells [31,32]. Stacks of images were taken at random areas on the HPDE sheets where each stack contained 10 images.
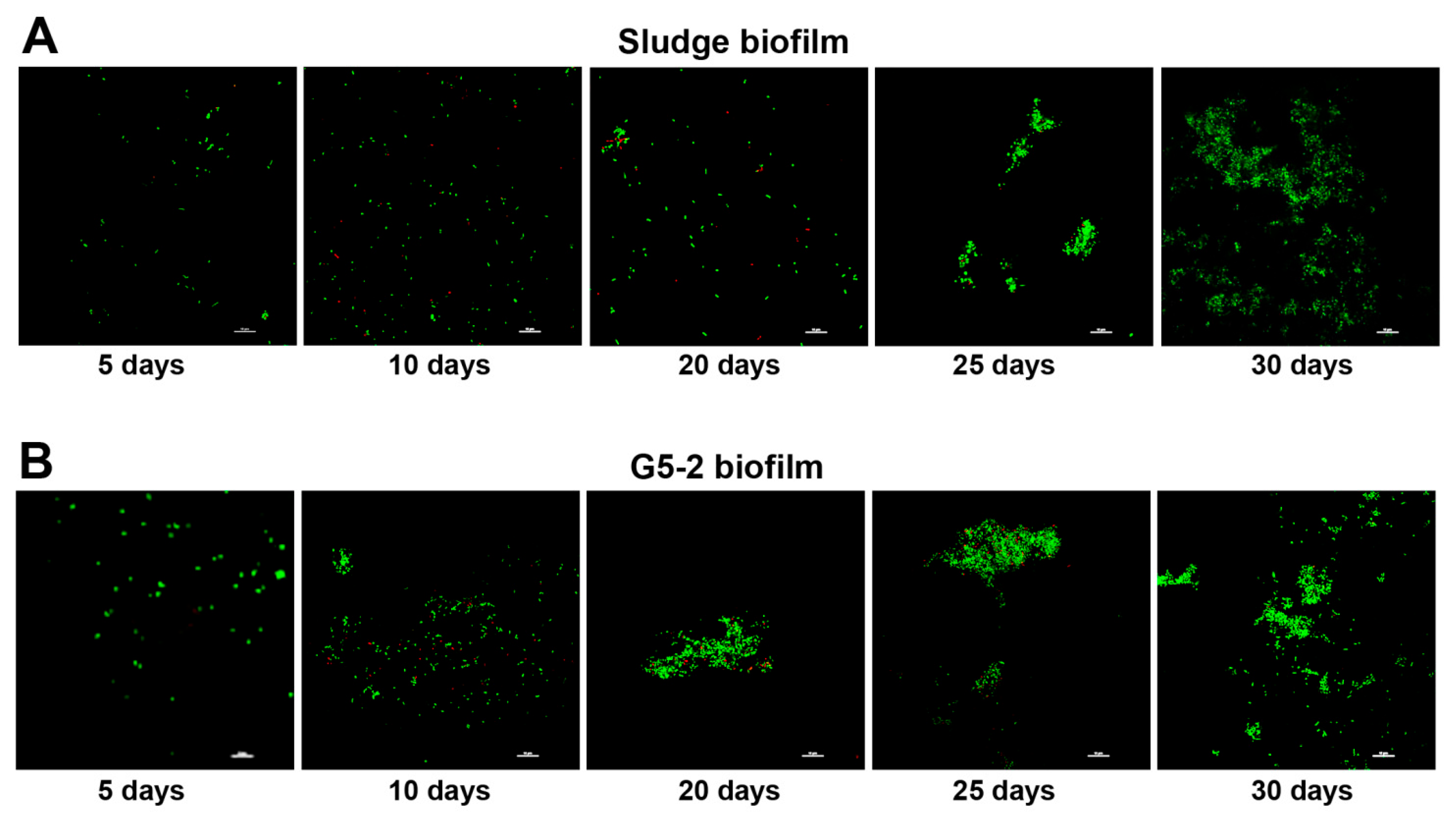
Figure 2.
Bio-Repetitive Re-Inoculum Assay by G5-2 and G5-4 MBBs. Bio-carriers of G5-2 and G5-4 (30 pieces each) were incubated in 30 mL of LMM at 28°C. The NH4+-N, was adjusted to ≅ 350 mg/L and monitored, using Nessler’s test, as a function of time for the whole duration of Bio-RRIA (A) (see body text). Samples from Bio-RRIA were withdrawn at indicated NH4+-N falls, for total nitrogen, nitrite and nitrate determinations of G5-2 (B) and G5-4 (C) biofilms. Total nitrogen is the sum of ammonia remained, nitrites, nitrates and organic nitrogen compounds from leachate. Leachate composition is reported in Supplementary Table S1.
Figure 2.
Bio-Repetitive Re-Inoculum Assay by G5-2 and G5-4 MBBs. Bio-carriers of G5-2 and G5-4 (30 pieces each) were incubated in 30 mL of LMM at 28°C. The NH4+-N, was adjusted to ≅ 350 mg/L and monitored, using Nessler’s test, as a function of time for the whole duration of Bio-RRIA (A) (see body text). Samples from Bio-RRIA were withdrawn at indicated NH4+-N falls, for total nitrogen, nitrite and nitrate determinations of G5-2 (B) and G5-4 (C) biofilms. Total nitrogen is the sum of ammonia remained, nitrites, nitrates and organic nitrogen compounds from leachate. Leachate composition is reported in Supplementary Table S1.
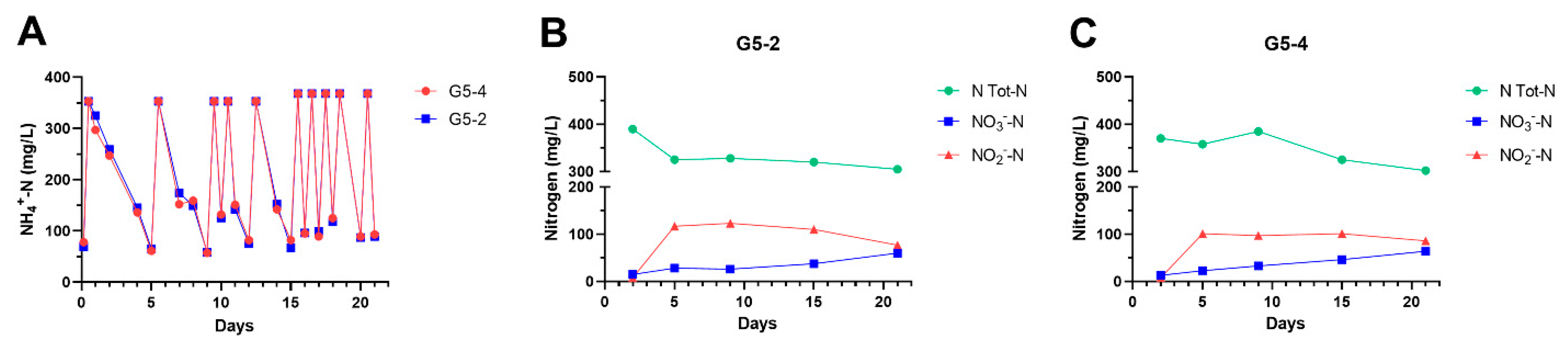
Figure 3.
Effect of the temperature on Bio-RRIA by G1-2, G5-2 and G5-4 MBBs. The G1-2 (red circle), G5-2 (blue square) and G5-4 (green triangle) MBBs were incubated at the four indicated temperatures and subjected to Bio-RRIA essentially as described in the body text and in the legend of Figure 2A.
Figure 3.
Effect of the temperature on Bio-RRIA by G1-2, G5-2 and G5-4 MBBs. The G1-2 (red circle), G5-2 (blue square) and G5-4 (green triangle) MBBs were incubated at the four indicated temperatures and subjected to Bio-RRIA essentially as described in the body text and in the legend of Figure 2A.
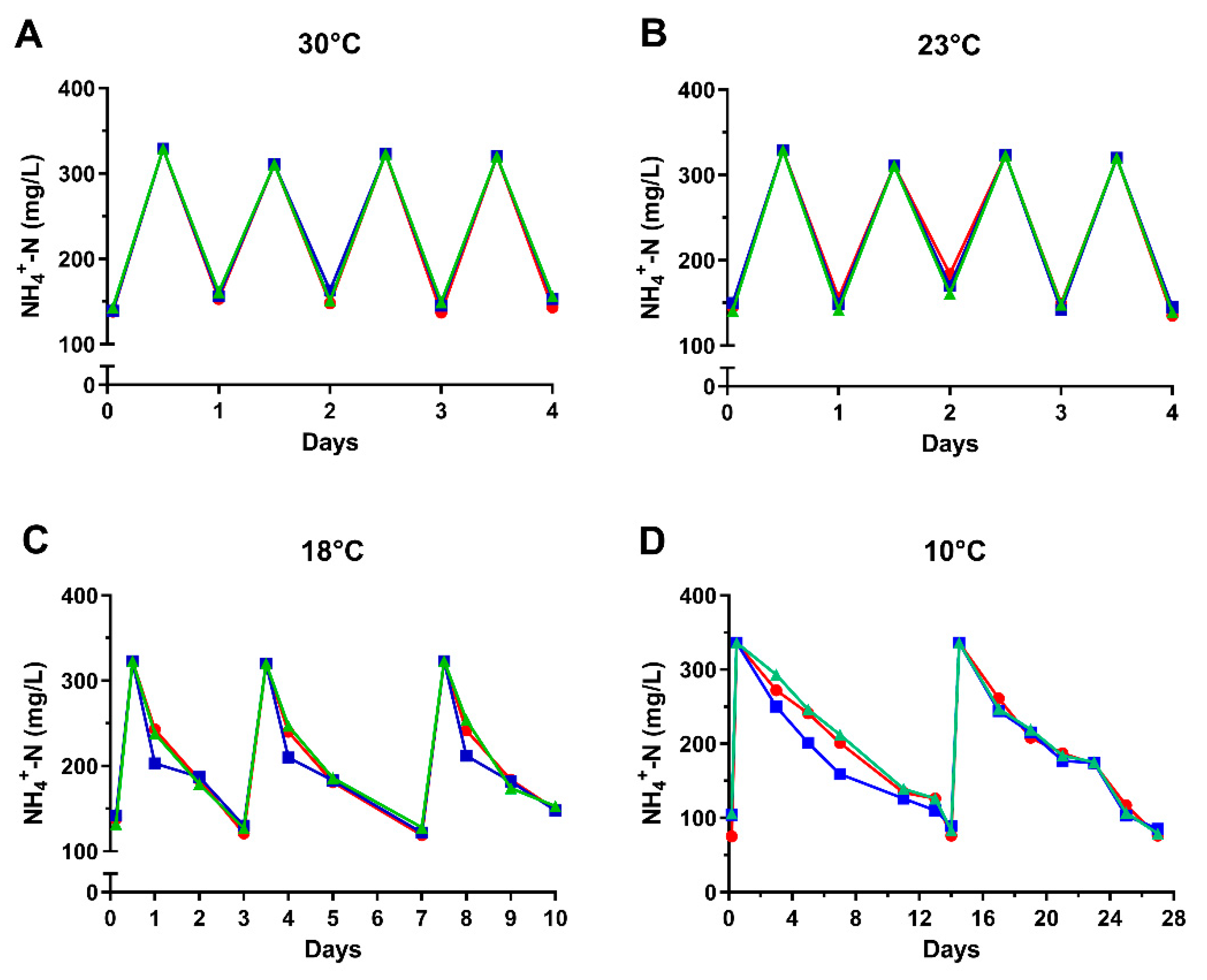
Figure 4.
Bio-Repetitive Re-Inoculum Assay by sludge MBBs. Sludge samples were taken at different periods from the Porto Sant’Elpidio MWWTP and used for K1 bio-carriers preparation. Biofilms were investigated for their efficiency in NH4+-N removing in Bio-RRIA experiments performed as described in the body text and in the Legend of Figure 2A. Four representative sludges, indicated with SL1-4, are shown.
Figure 4.
Bio-Repetitive Re-Inoculum Assay by sludge MBBs. Sludge samples were taken at different periods from the Porto Sant’Elpidio MWWTP and used for K1 bio-carriers preparation. Biofilms were investigated for their efficiency in NH4+-N removing in Bio-RRIA experiments performed as described in the body text and in the Legend of Figure 2A. Four representative sludges, indicated with SL1-4, are shown.
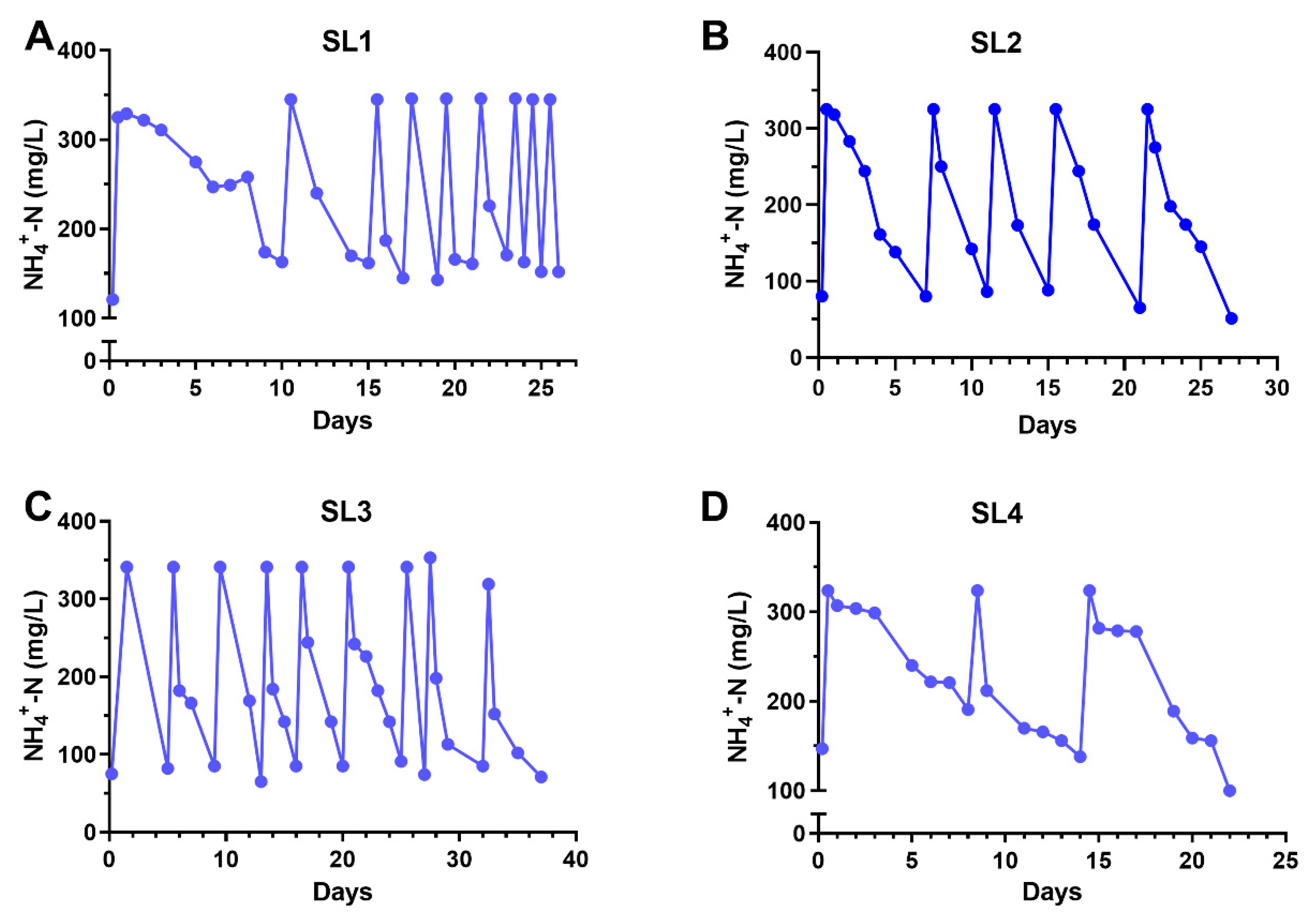
Figure 5.
Change of biofilm population from selected microbial communities. Relative abundance, expressed as percentage of total operational taxonomic units (OTUs), of the most prevalent families (A, B) and species (C, D) in the biofilms derived from G5-2 and G5-4 cultures are shown. Samples for species identification by 16S rRNA NGS were the bacterial cultures used for biofilm formation (grey bars) and MBBs taken on the 1st day (green bars), on the 10th day (red bars) and on the 15th day (dark blue bars) of Bio-RRIA reported in Figure 2A. The bar graph shows only families and species that contributed more than 4% to the total bacterial community in at least one point.
Figure 5.
Change of biofilm population from selected microbial communities. Relative abundance, expressed as percentage of total operational taxonomic units (OTUs), of the most prevalent families (A, B) and species (C, D) in the biofilms derived from G5-2 and G5-4 cultures are shown. Samples for species identification by 16S rRNA NGS were the bacterial cultures used for biofilm formation (grey bars) and MBBs taken on the 1st day (green bars), on the 10th day (red bars) and on the 15th day (dark blue bars) of Bio-RRIA reported in Figure 2A. The bar graph shows only families and species that contributed more than 4% to the total bacterial community in at least one point.
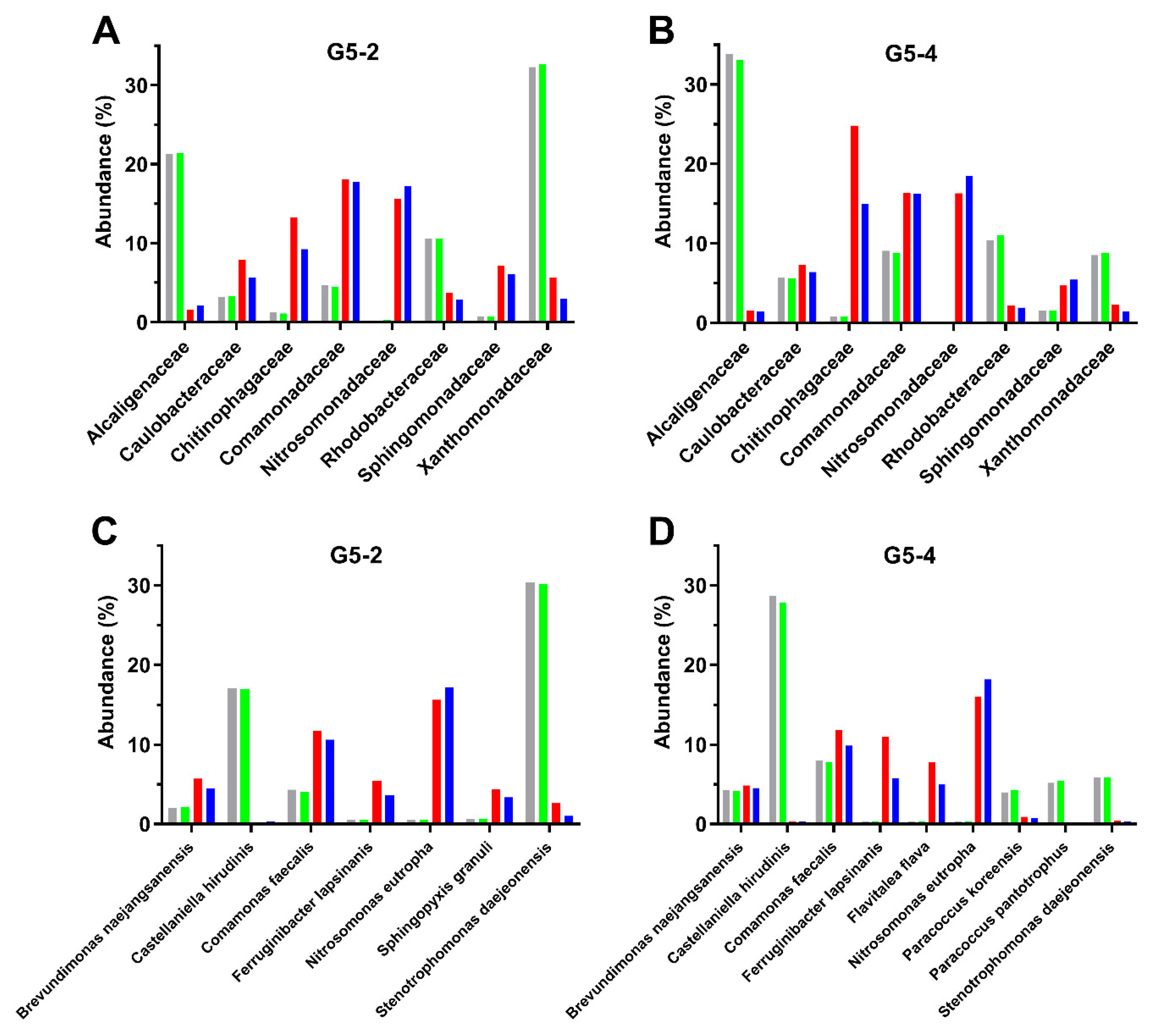
Figure 6.
Stacked bar-chart of MBBs from native sludges. Relative abundance, expressed as percentage of total operational taxonomic units (OTUs), of the most prevalent families in biofilm-derived sludges SL2, SL3 and LSL4. Bacteria identification was carried out by 16S rRNA NGS and values represent the average of at least three points taken during Bio-RRIA experiments as shown in Figure 4. Only families, that contributed more than 3% to the total bacterial community, are reported.
Figure 6.
Stacked bar-chart of MBBs from native sludges. Relative abundance, expressed as percentage of total operational taxonomic units (OTUs), of the most prevalent families in biofilm-derived sludges SL2, SL3 and LSL4. Bacteria identification was carried out by 16S rRNA NGS and values represent the average of at least three points taken during Bio-RRIA experiments as shown in Figure 4. Only families, that contributed more than 3% to the total bacterial community, are reported.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



