
Submitted:
20 September 2024
Posted:
23 September 2024
You are already at the latest version
Abstract
The emergence of antimicrobial resistance (AMR) in pathogens transmitted through food poses a significant threat to global public health, complicating infection treatment and increasing mortality rates. This review explores the role of resistome mapping as a crucial tool for understanding the transmission dynamics of antimicrobial resistance genes (ARGs) in foodborne pathogens, such as Salmonella, Escherichia coli, Listeria monocytogenes, and Campylobacter spp., as well as various techniques for resistome mapping, such as metagenomic sequencing, PCR-based methods, and whole-genome sequencing (WGS), highlighting the significance of horizontal gene transfer (HGT) as a key mechanism for ARG dissemination in foodborne pathogens. Additionally, we investigated the influence of agricultural practices and environmental factors on AMR development, highlighting the critical need for improved surveillance, antibiotic stewardship, and global collaboration to mitigate the spread of resistant ARGs through the food chain. The perceptions gained from resistome mapping play an essential role in developing effective approaches to address AMR and to ensure food safety.
Keywords:
antibiotic resistance genes (ARGs)
; horizontal gene transfer (HGT)
; metagenome sequencing
; whole genome sequencing
; antibiotic stewardship
; food safety
1. Introduction
Foodborne pathogens pose a significant threat to global public health owing to their ability to cause widespread outbreaks, induce serious illnesses, and potentially lead to fatalities. This challenge is exacerbated by the emergence and persistence of antimicrobial resistance (AMR) within these pathogens. AMR significantly compromises the efficacy of conventional treatment approaches, resulting in increased morbidity and mortality [1]. According to the World Health Organization (WHO), AMR infections are responsible for approximately 700,000 deaths annually. If not addressed, this figure could potentially increase to 10 million deaths annually by 2050 [2]. Moreover, the continued evolution of AMR strains in response to the selective pressure created by antibiotic usage further complicates this challenge. This burden is excessively high in low- and middle-income countries because of weak health systems, insufficient biosafety protocols, and the widespread misuse of antibiotics in agriculture and clinical contexts, which contribute to the rapid spread of AMR strains [3]. While pathogens such as Salmonella and Escherichia coli are well-known contributors to AMR [4], other foodborne pathogens, such as Listeria monocytogenes and Campylobacter spp., are highly identified as significant contributors [5] to the global “resistome”—the collection of all resistance genes present in both pathogenic and non-pathogenic bacteria [6]. Resistance genes can be transmitted at various points in the food supply chain. The complex nature of modern food production, processing, and distribution networks has created pathways for AMR bacteria to move from agricultural settings to consumers. Factors such as the overuse of antibiotics in animal husbandry, insufficient sanitation measures, and poor food safety practices further accelerate the spread of AMR pathogens [7].
Despite increased awareness, substantial knowledge gaps remain in the complete understanding of AMR transmission dynamics within foodborne pathogens. Previous studies have emphasized the importance of mobile genetic elements, such as plasmids and transposons, in facilitating the horizontal transfer of resistance genes. However, most of this research has focused on clinical contexts, leaving crucial gaps in the knowledge of AMR transmission and persistence in environmental and agricultural contexts [8,9]. Furthermore, there is an urgent need to enhance our knowledge of how specific agricultural practices, including the use of antibiotics in livestock and the application of manure as fertilizer, contribute to the enhancement and spread of AMR genes in foodborne pathogens. Additionally, while research has made progressive steps in identifying antibiotic resistance genes (ARG), a more holistic approach is necessary to identify the activation and expression of hidden resistance genes that may become clinically significant under specific environmental conditions [10]. Addressing these research gaps will enhance our understanding of AMR transmission dynamics and inform targeted strategies to mitigate the public health risks associated with foodborne AMR pathogens.
The purpose of this review is to delve into the complex resistome landscape of foodborne pathogens and examine the mechanisms that contribute to the development and spread of antimicrobial resistance. By analyzing current research and mapping resistance genes, this review aims to provide insights into the implications of resistance in foodborne pathogens, with the goal of guiding more effective surveillance, intervention strategies, and antibiotic stewardship practices. This work is crucial for addressing the growing public health challenges posed by antibiotic-resistant pathogens in the global food supply. This review explores the critical role of resistome mapping in foodborne pathogens and the mechanisms that contribute to the development and spread of AMR. This study underscores the considerable influence of AMR on public well-being by evaluating recent studies and scrutinizing the prevalence of AMR genes. The primary goal is to enhance the understanding of AMR transmission within the food chain and provide insights for improving surveillance systems, mitigation strategies, and antibiotic stewardship.
2. Global Impact of Foodborne Pathogens
Foodborne pathogens, including Salmonella, E. coli, Listeria, etc., cause a broad range of illnesses such as from mild gastrointestinal discomfort to severe, life-threatening conditions [11]. WHO estimates that globally, around 600 million illnesses each year from foodborne diseases, with approximately 420,000 fatalities. This case includes 30% of children, constituting 9% of the global population [12]. In addition, some foodborne infections can lead to chronic health problems such as kidney failure or arthritis, causing a long-term burden on individuals and healthcare systems [13]. Economically, the consequences of foodborne illnesses are highly significant. The expenditure on medical care for infected people, coupled with productivity loss due to contamination, causes substantial financial strain on countries [14]. Usually, agricultural or food industry outbreaks can lead to expensive recalls of products, which reduces consumer confidence and potential legal actions. Ultimately, this impacts the overall reputation of the industry. Additionally, foodborne disease outbreaks can impose international trade restrictions, affecting global markets and local economies [15].
The social impact of foodborne pathogens extends beyond health and economics, affecting public trust and social stability [16]. Susceptible people, especially old age and children, and those with weak immune systems, are extremely affected, which intensifies existing inequalities [14]. Beyond this, the most complex issue is addressing environmental concerns such as the spread of antibiotic resistance. The misuse and overuse of antibiotics in agriculture and food production significantly contributes to this problem, allowing resistant bacteria to disseminate in the environment. These resistant pathogens have the potential to contaminate soil, water, and other food sources, and subsequently transfer through the food chain to humans, which further intensifies the public health crisis [7]. The capacity of these pathogens to transfer their ARGs to other non-pathogenic bacteria through plasmids and other transposable elements and drive the development of drug-resistant pathogens is an additional concern [17].
3. Mechanisms of Antibiotic Resistance in Foodborne Pathogens
AMR in foodborne pathogens is acquired via numerous mechanisms (Figure 1) that permit bacteria to persist in antibiotic treatments and remain a public health concern. The most conspicuous mechanism is horizontal gene transfer (HGT), where bacteria exchange genetic material, including ARGs, through processes such as conjugation, transformation, and transduction [18]. In conjugation, bacteria transport resistance genes through cell–cell connections, frequently via plasmids that carry multiple resistance genes. This mechanism spreads resistance quickly across multiple bacterial species, specifically in environments such as farms or food-processing plants [19]. Transformation occurs when bacteria take up naked DNA from their environment, probably from dead bacteria that have released genetic material containing resistance genes [20]. Transduction involves bacteriophages (viruses that infect bacteria) that can carry resistance genes from one bacterium to another [21].
In addition to HGT, genetic mutations play a key role in AMR development. Bacteria can spontaneously obtain mutations by changing the antibiotic target sites, making the drug less effective or ineffective. For example, mutations in bacterial ribosomal RNA or DNA gyrase can inhibit the binding efficiency of antibiotics, such as macrolides and fluoroquinolones [22]. The development of efflux pumps is another mutation-driven mechanism that actively pushes antibiotics out from the bacterial cell, decreasing the concentration of the drug inside the cell and preventing it from moving to a lethal level. Another mutation-driven mechanism involves altering cell membrane permeability and preventing antibiotics from entering the cell [23]. In addition, bacteria can synthesize enzymes that can degrade or alter antibiotics. For example, beta-lactamases are enzymes that break down beta-lactam antibiotics, such as penicillins and cephalosporins, whereas other enzymes, such as aminoglycoside-modifying enzymes, chemically modify antibiotics to make them ineffective [24].Finally, bacteria can develop and use alternative metabolic pathways to bypass the antibiotic effect. For example, in the presence of sulfonamides (which inhibit folic acid synthesis), bacteria can develop new pathways to synthesize folic acid, thereby abolishing the effects of the drug [25]. These diverse mechanisms, combined with the selective pressure employed by the extensive use of antibiotics in agriculture and food production, create a difficult contest for controlling foodborne AMR pathogens. As resistance continues to develop and persist, addressing these AMR mechanisms via antibiotic stewardship, surveillance, and research is crucial for safeguarding public health and food safety.
4. Resistome Mapping: Concepts and Techniques
4.1. Definition and Importance of Resistome Mapping
Resistome mapping is the complete process of identifying, cataloging, and assessing an entire set of ARGs within a particular microbial community, environment, or organism. The concept of “resistome” comprehends the sum of the resistance genes, including those conferring antibiotic resistance, whether they are actively expressed or not. This involves not only the currently identified genes that are known to provide resistance to antibiotics but also those with the capability to do so, including cryptic or silent genes that may become activated under specific conditions[6]. In the context of foodborne pathogens, resistome mapping involves comprehensive analysis of the complete genetic content to identify ARGs that allow pathogens to resist antibiotics. This analytical approach is essential to identify the survival and flourishing of these pathogens in the presence of antibiotics.
Figure 2.
The workflow of resistome mapping (Created in BioRender.com).
Figure 2.
The workflow of resistome mapping (Created in BioRender.com).
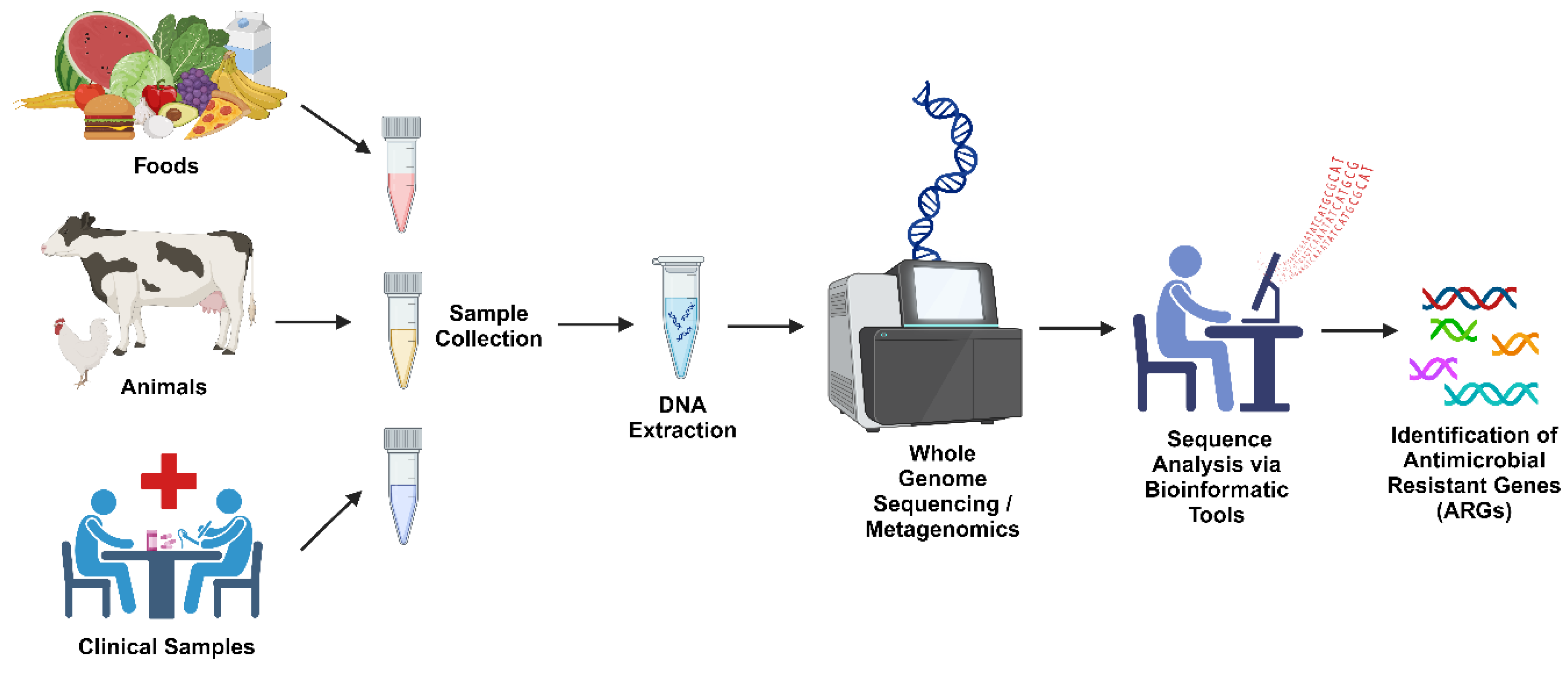
The significance of resistome mapping of foodborne pathogens cannot be overstated. Foodborne diseases are a major global public health concern, causing millions of illnesses and fatalities each year. Several pathogens, including Salmonella, E. coli, and Campylobacter, have emerged as resistant to numerous antibiotics, significantly complicating the treatment of infections [26]. Resistome mapping of these pathogens can advance the understanding of the detailed resistance mechanisms, including the genes that confer resistance and their acquisition, transmission, and expression patterns. Additionally, resistome mapping offers crucial information for developing more effective surveillance strategies to monitor the spread of ARGs across various environments, such as agricultural farms, food processing plants, and consumer markets [27]. This assists public health representatives in recognizing potential hotspots of resistance and executing targeted interventions to mitigate the spread of AMR pathogens. Resistome mapping can also contribute to the development of new diagnostic methods that can instantly identify AMR pathogens in food products, thereby preventing contaminated food from reaching consumers [28].
Resistome mapping plays a crucial role in antibiotic stewardship. By understanding the resistome of foodborne pathogens, policymakers and healthcare professionals can make more informed choices regarding antibiotic usage in both agriculture and medicine [29]. This is essential for reducing the excessive use of antibiotics in livestock, which is a significant driver of resistance development. Furthermore, resistome analysis assesses the impact of numerous agricultural practices on the emergence and spread of resistance [6]. Scientists can use the data produced from resistome studies to model the evolutionary dynamics of resistance, assisting them in predicting future trends in resistance development. It provides a detailed understanding of the genetic basis of resistance, which supports research aimed at addressing the global challenges of AMR [30]. With the increasing threat of AMR, resistome mapping has become crucial for ensuring food safety and public health.
4.2. Techniques for Resistome Mapping
Various techniques have been used to identify, quantify, and evaluate the dynamics of ARGs in microbes, providing unique insights into the resistance of foodborne pathogens. Each method offers unique benefits, ranging from high-resolution genome sequencing to specific gene detection, enabling a complete understanding of the dissemination and persistence of resistance genes.
Metagenomic sequencing is the most comprehensive and extensively used technique for mapping resistomes. It involves direct sequencing of the complete genetic material of a microbial community from environmental or clinical samples, without culturing individual organisms. High-throughput sequencing technologies, such as Illumina (which provides short-read sequencing with high accuracy) and Oxford Nanopore Technology (which offers long-read sequencing with real-time analysis), are widely utilized in this approach. This is particularly valuable because it captures both culturable and unculturable microorganisms, offering a complete view of the resistome in complex environments, including the human gut, animal microbiomes, agricultural soils, and food production settings, where diverse bacterial species coexist [31]. However, this approach has significant drawbacks. It may not provide specific information on which bacterial species harbor resistance genes without additional analyses, such as metagenome-assembled genomes (MAGs), which can reconstruct genomes from metagenomic data [32].
Metagenomics involves two approaches: shotgun metagenomics and amplicon sequencing. Shotgun metagenomics involves sequencing all DNA present in a sample, providing the entire resistome and other microbial functions. This method can also help identify mobile genetic elements (MGEs), such as plasmids, transposons, and integrons, which play a critical role in the HGT of ARGs. In contrast, amplicon sequencing targets specific gene regions, commonly the 16S rRNA gene, to identify microbial species and their relative abundance. Although this method is cost-effective and faster than shotgun metagenomics, it cannot identify ARGs unless specific primers for resistance genes are used [33].
Polymerase chain reaction (PCR)-based techniques, such as quantitative PCR (qPCR) and multiplex PCR, are commonly used in resistome research because of their high sensitivity and specificity. These methods are used to detect and quantify specific known ARGs in environmental, clinical, or food sources. qPCR can provide quantitative data, which are beneficial for monitoring changes in ARG prevalence over time or in different environments [34]. In contrast, multiplex PCR enables the detection of multiple ARGs in a single reaction, thereby enhancing the efficiency of surveillance efforts when multiple resistance genes are of concern. These methods are more cost-effective and require fewer computational resources than metagenomic approaches. However, the major limitation is their reliance on known ARG sequences, which means that they cannot identify novel or uncharacterized resistance genes. Furthermore, these techniques do not provide information on the genomic context of ARGs, such as whether they are located in mobile genetic elements [35].
Although metagenomics and PCR-based methods primarily detect the presence of resistance genes, functional metagenomics further analyzes the actual function of these genes. This method involves extracting DNA, cloning it into a vector, and introducing it into a model organism, typically E. coli. The transformed E. coli is subsequently exposed to antibiotics to determine if it expresses resistance, thus establishing genetic information for phenotypic resistance. This is valuable for discovering novel ARGs and understanding their functions in microbes. This method does not require prior knowledge of the resistance genes, making it an effective tool for identifying new resistance mechanisms. Moreover, it can reveal the genes that contribute to resistance through unexplored pathways. This approach has several limitations, including more labor-intensive and time-consuming techniques, the need for specialized laboratory setup and proficiency, and the possibility that not all genes will be expressed in the selected model organism. Nevertheless, functional metagenomics offers valuable insights into the functional aspects of the resistome [36,37].
Whole genome sequencing (WGS) is another efficient method for resistome mapping that involves sequencing the complete genome of a bacterial strain to detect ARGs and other associated elements. WGS provides detailed information on the genomic context of ARGs, permitting researchers to track the genetic pathways that lead to resistance and understand the role of mobile genetic elements in disseminating ARGs [38]. In comparative genomics, WGS data from various bacterial isolates can be analyzed to detect variations in resistance across different strains, species, or geographical regions. This method is useful in outbreak investigations because it helps identify the specific ARGs responsible for resistance. Such information is critical for public health responses and treatment approaches [39].
Bioinformatic tools and databases play a critical role in determining the success of resistome mapping by facilitating the identification and annotation of ARGs. Table 1 shows the list of all available AMR databases or resources and their information. These tools comprise carefully curated information on known resistance genes that helps in analyzing and translating high-throughput genomic sequencing data obtained through metagenomics and WGS to obtain meaningful insights into the distribution and function of ARGs.
5. Resistome Mapping Studies in Food Borne Pathogens
5.1. Salmonella
Resistome mapping of Salmonella isolates is critical for understanding AMR mechanisms. Salmonella, particularly S. enterica, is commonly found in poultry, eggs, and raw vegetables, and frequently carries ARGs that disseminate through the food chain [58]. Common resistance genes in Salmonella include those encoding resistance to β-lactams, tetracyclines, aminoglycosides, and sulfonamides. Remarkably, extended-spectrum beta-lactamase (ESBL)-producing Salmonella isolates with genes such as blaCTX-M and blaTEM pose significant threats because of their ability to degrade several β-lactam antibiotics [59]. Mobile genetic elements such as plasmids and transposons enable the dissemination of ARGs within Salmonella populations. In multidrug-resistant (MDR) strains, conjugative transfer of plasmids, such as IncFII and IncHI2, has been observed in isolates from both food and clinical settings. This highlights the role of HGT in the spread [59]. Geographic variations in ARG occurrence in Salmonella have been reported, with some regions exhibiting high levels of fluoroquinolone resistance due to qnr genes, whereas others show greater resistance to third-generation cephalosporins [60]. These differences indicate regional patterns of antibiotic use, especially in agriculture where antibiotics are used as both growth enhancers and prophylactics. Despite extensive data, inconsistencies exist in the AMR levels in Salmonella among different food sources. Although poultry is a well-recognized reservoir, seafood in some regions also shows high resistance levels [7]. These differences emphasize the complex nature of AMR in Salmonella, which is prompted by environmental factors, farming practices, and local antibiotic use. Incorporating resistome data into public health strategies is important to control Salmonella outbreaks by regulating antibiotic use in agriculture and enhancing food safety standards.
5.2. Escherichia coli
E. coli is another important foodborne pathogen with a complex resistome. Strains such as enterohemorrhagic E. coli (EHEC) are known to induce severe gastrointestinal disorders. Resistome mapping has revealed numerous AMR genes in E. coli isolated from food sources, particularly meat and dairy products. These include resistance genes for third-generation cephalosporins, aminoglycosides, and fluoroquinolones [61]. The incidence of ESBL-producing E. coli, particularly in poultry and cattle, is a rising concern, with genes such as blaCTX-M, blaSHV, and blaOXA frequently found on plasmids that facilitate rapid AMR dissemination [59]. Recent studies have also revealed colistin resistance in E. coli, with mcr genes emerging as a major concern because of its role as a last-line antibiotic for treating MDR infections [62]. HGT also plays a central role in disseminating resistance among E. coli populations and other Enterobacteriaceae species [63]. Regional variations in E. coli resistance profiles indicate local antibiotic use and stewardship policies [64]. Variations in the prevalence of resistance genes, such as variable levels of mcr genes, indicate that precise resistome mapping is essential for an accurate outbreak assessment [65].
5.3. Listeria monocytogenes
Listeria monocytogenes is a foodborne pathogen with a high mortality rate in vulnerable populations. While it generally exhibits lower levels of antibiotic resistance than other pathogens, resistome mapping has identified key resistance mechanisms, particularly against antibiotics used in agriculture, such as tetracyclines and macrolides [11,66]. The tetM and erm genes, which confer resistance to tetracyclines and macrolides, respectively, have been detected in isolates from various food products, including dairy and ready-to-eat meats [7,11]. A significant observation was the identification of MDR strains of L. monocytogenes that carry resistance genes on mobile genetic elements such as plasmids and transposons in both clinical and foodborne outbreaks [11]. The prevalence of resistance genes differs based on food origin, with dairy products exhibiting higher levels owing to widespread antibiotic usage in dairy farming [67]. Inconsistencies in the impact of antibiotic use on the L. monocytogenes resistome indicate that factors such as study design and geographic location may affect findings [68]. Continuous monitoring and surveillance are important to prevent the emergence of MDR strains and ensure effective treatment choices.
5.4. Campylobacter spp.
Resistome mapping of Campylobacter species, particularly Campylobacter jejuni and Campylobacter coli, has uncovered significant patterns of AMR relevant to foodborne illnesses. Campylobacter is a leading cause of bacterial gastroenteritis, and its resistome includes resistance to ciprofloxacin and tetracycline, primarily due to gyrA mutations and the tet(O) gene [69]. The high prevalence of fluoroquinolone-resistant Campylobacter strains is concerning, given ciprofloxacin’s critical role in treating severe campylobacteriosis [70]. This resistance has been linked to the extensive use of fluoroquinolones in veterinary medicine. Additionally, resistome mapping has identified MDR strains of Campylobacter carrying genes conferring resistance to macrolides, such as erm gene [71]. Regional variations in resistance profiles reflect local antibiotic use. Higher levels of tetracycline-resistant Campylobacter have been reported in regions with extensive tetracycline use in animal husbandry, whereas those with stringent antibiotic stewardship have lower resistance levels [72]. Conflicting results in these studies may arise from differences in sampling methods and testing protocols, highlighting the need for standardized approaches and comprehensive surveillance to accurately assess AMR in Campylobacter spp.
5.5. Vibrio cholerae
Vibrio cholerae, the causative agent of cholera, possesses a diverse resistome that changes with region and environmental conditions. Resistome mapping has identified resistance mechanisms against tetracyclines, sulfonamides, and aminoglycosides, with ARGs such as tetA, sul1, and aadA present in both clinical and environmental samples [73]. The spread of tetracycline resistance via tetA gene on plasmids or integrons is particularly alarming given tetracycline's past role in cholera treatment [74]. Recent studies have also shown resistance to quinolones and macrolides in V. cholerae, with resistance associated with peculiar genetic components and environmental conditions such as aquatic habitats [75]. Regional variations in the spread of resistance reflect variations in antibiotic use and environmental factors. Attempts to control AMR in V. cholerae must incorporate enhanced surveillance, environmental monitoring, and alternative treatment [76].
5.6. Clostridium perfringens
Clostridium perfringens, another significant pathogen in foodborne illnesses, is notably less studied but is an important resistome. Mapping studies have identified several resistance genes in C. perfringens, particularly those associated with foodborne outbreaks. Key resistance genes include those conferring resistance to tetracyclines, macrolides, and lincosamides [77]. The ermB gene confers resistance to macrolides and lincosamides and is commonly found on plasmids or integrons. The mobility of this gene facilitates its spread among bacterial strains, thereby contributing to the broader dissemination of AMR [78]. Additionally, the use of tetracyclines in agriculture and veterinary medicine contributes to tetracycline resistance in C. perfringens, which is often mediated by the tetM gene. This selective pressure can promote the persistence and dissemination of AMR strains [79]. However, differences in resistance levels reported in different studies on C. perfringens may result from differences in sampling methods and geographic areas. Some studies have indicated high resistance in isolates from certain food sources, while others have reported a lower prevalence [80]. These inconsistencies emphasize the need for consistent surveillance and monitoring of AMR in C. perfringens infections.
5.7. Shigella
AMR in Shigella species, including S. dysenteriae, S. flexneri, S. boydii, and S. sonnei, has been widely studied through resistome mapping. As a primary cause of bacterial dysentery, Shigella is resistant to several antibiotics including ampicillin, ciprofloxacin, and trimethoprim-sulfamethoxazole. Resistance mechanisms are mainly associated with plasmids or chromosomal elements [81]. The enzyme β-lactamase encoding blaTEM gene confers resistance to ampicillin-like antibiotics commonly found throughout different geographical regions, indicating extensive β-lactam use [82]. Similarly, qnr genes encoding quinolone resistance have been identified, raising concerns regarding the efficacy of fluoroquinolones in treating shigellosis [83]. sul1 and sul2 genes confer resistance to trimethoprim-sulfamethoxazole located in integrons, which can secure and disseminate these genetic elements [84]. Shigella showed a higher prevalence in South Asia and Africa, potentially linked to local antibiotic practices and healthcare setup [85]. Standardizing surveillance and improving data comparability are crucial for the effective management of AMR in Shigella.
5.8. Clostridium botulinum
Clostridium botulinum, the causative agent of botulism, possesses a diverse set of resistance mechanisms for survival in different environments. In addition to generating powerful neurotoxins, C. botulinum is resistant to antibiotics, such as clindamycin and vancomycin[86]. The ermB gene confers resistance to macrolides and lincosamides, and is commonly located on plasmids or integrons, facilitating its spread among bacterial populations [87]. Vancomycin resistance in C. botulinum, associated with the vanA and vanB genes, may be obtained from HGT from vancomycin-resistant enterococci [88]. The presence of resistance genes in both clinical and environmental C. botulinum strains highlights the adaptability of the pathogen. Regional differences in antibiotic resistance profiles are frequently associated with local antibiotic use and environmental factors. Higher resistance levels have been reported in areas with extensive antibiotic use, indicating the influence of selective pressure [89]. Addressing these inconsistencies requires uniform testing methods and comprehensive surveillance programs for effective management of AMR in C. botulinum.
5.9. Yersinia enterocolitica
Yersinia enterocolitica, a pathogen causing yersiniosis, shows resistance to antibiotics such as tetracycline, chloramphenicol, and trimethoprim-sulfamethoxazole [90]. Key ARGs include tetA, which confers tetracycline resistance, and catA, which confers resistance against chloramphenicol[91]. Plasmids often carry the tetA gene, enabling its transmission and contributing to the dissemination of resistance [91]. Trimethoprim-sulfamethoxazole resistance is linked to the sul1 and sul2 genes, which are often present in integrons. These genes contribute to the persistence of resistant strains in both clinical and environmental settings [92]. Higher resistance levels were noted in regions with intensive antibiotic use, indicating the influence of selective pressure [7]. Conflicting findings regarding resistance levels in Y. enterocolitica may result from variations in study methodologies and sampling locations [93]. Standardized surveillance practices and improved data comparability are essential to accurately assess and manage AMR in Y. enterocolitica.
6. Impact of Agricultural Practices on the Resistome
Resistomes are not only known for their ability to impart resistance to pathogens in the clinical setting but also to non-pathogenic organisms that predominate in the surrounding environment [94]. Antibiotic resistance in agricultural soils has been studied extensively. However, soil fertilization with manure and irrigation with wastewater are typically regarded as the primary sources of ARG contamination. The addition of significant volumes of organic matter to the soil found in manure and wastewater leads to the growth of AMR bacteria [95]. The potential contribution of AMR and ARG to soil through organic manure has drawn considerable interest. According to previous research, soil ARGs can be transferred to non-pathogenic and pathogenic bacteria that attach to or parasitize agricultural products. These products are frequently improperly cleaned before consumption, which poses a serious risk to public health [96]. Manure is the primary vehicle for the introduction of AMR and ARGs from animals into the environment. Consequently, manure management affects the transfer of AMR from agriculture to human clinical settings through soil, water, and food [97].
Wastewater from aquaculture and animal agriculture has been identified as another source of AMR dissemination. As several antibiotics used in aquaculture, including tetracycline, macrolides, and aminoglycosides, are vital for human treatment, prolonged use of these antibiotics in aquaculture has resulted in AMR in humans [98]. The impact of mineral fertilizers and pesticides, which are extensively used in conventional agriculture, on the development and dissemination of antibiotic resistance has received less attention. It has been demonstrated that herbicides, fungicides, and insecticides affect E. coli resistance to antibiotics [95]. Research findings indicate that herbicides can alter bacterial susceptibility to antibiotics, create resistance, and facilitate conjugative transfer of ARGs [99]. A significant portion of ARGs reaches river systems via agricultural nonpoint sources as a result of rainfall and runoff. This poses a major danger to water security, which is threatened by ARGs; they spread quickly and widely in river systems owing to the action of MGEs [100].
8. Challenges in Resistome Mapping
Although resistome mapping has immense potential for advancing our knowledge of AMR and influencing public health policies, it also encounters numerous significant challenges. These challenges include the technical and analytical difficulties associated with complex genomic data. Addressing these difficulties is crucial for fully exploiting resistome mapping as a tool in the global fight against AMR.
A major hurdle is that the analysis of large-scale sequencing data requires sophisticated bioinformatics tools that can efficiently process complex metagenomic datasets. A major obstacle is the difficulty in differentiating ARGs between pathogenic bacteria and commensal or environmental bacteria because these bacteria can coexist within a single sample [101,102]. Additionally, the presence of ARGs on mobile genetic elements, such as plasmids, adds complexity because they are more connected to specific genes in particular bacterial species [103]. To overcome these challenges, researchers are developing more advanced bioinformatics pipelines that can accurately identify, annotate, and quantify ARGs in complex datasets. However, these tools require continuous improvement and validation to ensure that they provide reliable insights into resistomes [104].
Another key challenge in resistome mapping is the limitation of the currently available AMR reference databases. Comprehensive and updated databases are essential for accurate identification and annotation of ARGs. Unfortunately, many existing databases are incomplete or lack adequate annotations, predominantly for novel or less studied resistance genes [105]. This leads to misidentification of ARGs, restricting the full understanding of AMR. Ongoing efforts aim to expand these databases by incorporating novel ARGs from various environments, but the development is slow and database content frequently fails to keep pace with new gene discoveries [105].
9. Future Directions to Overcome the Challenges
9.1. Integrative Approaches
Resistome mapping can be significantly enhanced by integrating with other scientific approaches, such as functional assays and epidemiological studies, which can achieve a more comprehensive insight into the development, spread, and public health effects of AMR.
Integrating epidemiological data permits tracing of the transmission routes of AMR bacteria across various environments and populations [106]. For example, analyzing resistome mapping of bacterial populations in hospitals, farms, and urban areas can aid in recognizing the movement patterns of ARGs between these systems. Resistome analysis combined with epidemiological data on disease outbreaks can help determine the connections between ARGs and clinical or public health threats [107]. This method is valuable for understanding AMR transmission dynamics, such as the impact of zoonotic transfer (where resistant bacteria are passed from animals to humans) or their spread through international trade and travel. Such integrative studies are essential for developing more focused interventions ranging from infection control measures in hospitals to policies on antibiotic use in agriculture. Although sequencing-based resistome mapping is important for detecting the occurrence of ARGs, it does not provide information on whether these genes actively confer resistance in their organisms. This gap can be addressed using functional assays. Through experimental validation of specific resistance genes, researchers can understand their effects on bacterial fitness, pathogenicity, and potential clinical relevance. For example, some ARGs may only provide minimal resistance under laboratory conditions but could evolve as significant clinical issues if bacteria develop additional mutations. Functional assays can also reveal which ARGs are the most prominent in spreading through HGT. [37,108]. Similarly, integrating functional assays with resistome mapping further enhances our ability to predict and manage future AMR threats and helps prioritize the ARGs that require immediate attention.
9.2. Policy and Global Collaboration
The battle against AMR extends beyond scientific research; it also requires organized efforts in policy-making and global collaboration. The data generated through resistome mapping can inform regulatory frameworks and encourage international collaboration, both of which are crucial for mitigating the spread of AMR on a global scale. Controlling the spread of AMR requires effective policies for the regulation of antibiotic use, particularly in agriculture and food production. Resistome mapping provides information for the implementation of such policies. For example, data from resistome studies can pinpoint hotspot areas for AMR development in agricultural settings, prompting the implementation of stringent protocols for antibiotic use in farm animals [109]. Moreover, resistome mapping can support efforts to limit the environmental dissemination of ARGs by identifying critical points where interventions are required, such as waste management practices in food production or hospitals [110]. These insights are necessary for developing regulatory frameworks that balance the use of antibiotics in certain industries and prioritize the mitigation of AMR.
AMR is a global health challenge that requires collective action from multiple nations and organizations. International collaboration and sharing of resistome data across borders are essential for enhancing global AMR surveillance [111]. Countries with advanced sequencing technologies can offer technology and expertise or share AMR or ARG trend data to support countries with limited resources. Sharing resistome data across countries can help identify emerging resistance threats before they spread globally, enabling more proactive measures to prevent outbreaks. Collaborative networks such as the WHO’s Global Antimicrobial Resistance Surveillance System (GLASS) are critical for coordinating a unified global response to AMR. By fostering international collaboration, resistome mapping can be integrated into global AMR surveillance programs to deliver real-time information to guide global public health interventions. By addressing technical challenges, promoting integrative scientific approaches, and encouraging global collaboration, we can advance the field of resistome mapping and better protect public health against the growing threat of AMR.
Author Contributions
Conceptualization, L.K.E; writing—original draft preparation, M.O.Y., C.E.J., A.J., and L.K.E.; writing—review and editing, L.K.E; supervision, L.K.E. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The authors are grateful to the American Society of Microbiology (ASM)Future Leader Mentorship Fellowship (FLMF) Program.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Almansour, A.M.; Alhadlaq, M.A.; Alzahrani, K.O.; Mukhtar, L.E.; Alharbi, A.L.; Alajel, S.M. The Silent Threat: Antimicrobial-Resistant Pathogens in Food-Producing Animals and Their Impact on Public Health. Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- https://www.who.int/news/item/29-04-2019-new-report-calls-for-urgent-action-to-avert-antimicrobial-resistance-crisis#:~:text=If%20no%20action%20is%20taken, food%20systems%20are%20increasingly%20precarious New Report Calls for Urgent Action to Avert Antimicrobial Resistance Crisis.
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare (Switzerland) 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Balbin, M.M.; Hull, D.; Guest, C.; Nichols, L.; Dunn, R.; Thakur, S. Antimicrobial Resistance and Virulence Factors Profile of Salmonella Spp. and Escherichia Coli Isolated from Different Environments Exposed to Anthropogenic Activity. J Glob Antimicrob Resist 2020, 22, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, K.; Islam, M.R.; Siddiky, N.A.; Samad, M.A.; Chowdhury, S.; Hossain, K.M.M.; Rume, F.I.; Hossain, M.K.; Mahbub-E-Elahi, A.T.M.; Ali, M.Z.; et al. Antimicrobial Resistance Profile of Common Foodborne Pathogens Recovered from Livestock and Poultry in Bangladesh. Antibiotics 2022, 11. [Google Scholar] [CrossRef]
- Kim, D.W.; Cha, C.J. Antibiotic Resistome from the One-Health Perspective: Understanding and Controlling Antimicrobial Resistance Transmission. Exp Mol Med 2021, 53, 301–309. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; Andremont, A.; Bengtsson-Palme, J.; Brandt, K.K.; de Roda Husman, A.M.; Fagerstedt, P.; Fick, J.; Flach, C.F.; Gaze, W.H.; Kuroda, M.; et al. Critical Knowledge Gaps and Research Needs Related to the Environmental Dimensions of Antibiotic Resistance. Environ Int 2018, 117, 132–138. [Google Scholar] [CrossRef]
- Vezeau, N.; Kahn, L. Current Understanding and Knowledge Gaps Regarding Wildlife as Reservoirs of Antimicrobial Resistance. Am J Vet Res 2024, 85. [Google Scholar] [CrossRef]
- Sauerborn, E.; Corredor, N.C.; Reska, T.; Perlas, A.; Vargas da Fonseca Atum, S.; Goldman, N.; Wantia, N.; Prazeres da Costa, C.; Foster-Nyarko, E.; Urban, L. Detection of Hidden Antibiotic Resistance through Real-Time Genomics. Nat Commun 2024, 15. [Google Scholar] [CrossRef]
- Dufailu, O.A.; Yaqub, M.O.; Owusu-Kwarteng, J.; Addy, F. Prevalence and Characteristics of Listeria Species from Selected African Countries. Trop Dis Travel Med Vaccines 2021, 7. [Google Scholar] [CrossRef]
- https://www.who.int/news-room/fact-sheets/detail/food-safety Food Safety.
- Batz, M.B.; Henke, E.; Kowalcyk, B. Long-Term Consequences of Foodborne Infections. Infect Dis Clin North Am 2013, 27, 599–616. [Google Scholar] [CrossRef]
- Elbehiry, A.; Abalkhail, A.; Marzouk, E.; Elmanssury, A.E.; Almuzaini, A.M.; Alfheeaid, H.; Alshahrani, M.T.; Huraysh, N.; Ibrahem, M.; Alzaben, F.; et al. An Overview of the Public Health Challenges in Diagnosing and Controlling Human Foodborne Pathogens. Vaccines (Basel) 2023, 11. [Google Scholar] [CrossRef]
- Hussain, M.A.; Dawson, C.O. Economic Impact of Food Safety Outbreaks on Food Businesses. Foods 2013, 2, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhang, A.; van Klinken, R.D.; Schrobback, P.; Muller, J.M. Consumer Trust in Food and the Food System: A Critical Review. Foods 2021, 10. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Yu, S.; Li, D.; Gillings, M.R.; Ren, H.; Mao, D.; Guo, J.; Luo, Y. Inter-Plasmid Transfer of Antibiotic Resistance Genes Accelerates Antibiotic Resistance in Bacterial Pathogens. ISME Journal 2024, 18. [Google Scholar] [CrossRef] [PubMed]
- Emamalipour, M.; Seidi, K.; Zununi Vahed, S.; Jahanban-Esfahlan, A.; Jaymand, M.; Majdi, H.; Amoozgar, Z.; Chitkushev, L.T.; Javaheri, T.; Jahanban-Esfahlan, R.; et al. Horizontal Gene Transfer: From Evolutionary Flexibility to Disease Progression. Front Cell Dev Biol 2020, 8. [Google Scholar] [CrossRef]
- Virolle, C.; Goldlust, K.; Djermoun, S.; Bigot, S.; Lesterlin, C. Plasmid Transfer by Conjugation in Gram-Negative Bacteria: From the Cellular to the Community Level. Genes (Basel) 2020, 11, 1–33. [Google Scholar] [CrossRef]
- Hasegawa, H.; Suzuki, E.; Maeda, S. Horizontal Plasmid Transfer by Transformation in Escherichia Coli: Environmental Factors and Possible Mechanisms. Front Microbiol 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.N.; Penadés, J.R.; Chen, J. Genetic Transduction by Phages and Chromosomal Islands: The New and Noncanonical. PLoS Pathog 2019, 15. [Google Scholar] [CrossRef]
- Muteeb, G.; Rehman, M.T.; Shahwan, M.; Aatif, M. Origin of Antibiotics and Antibiotic Resistance, and Their Impacts on Drug Development: A Narrative Review. Pharmaceuticals 2023, 16. [Google Scholar] [CrossRef]
- Gaurav, A.; Bakht, P.; Saini, M.; Pandey, S.; Pathania, R. Role of Bacterial Efflux Pumps in Antibiotic Resistance, Virulence, and Strategies to Discover Novel Efflux Pump Inhibitors. Microbiology (United Kingdom) 2023, 169. [Google Scholar] [CrossRef] [PubMed]
- Jian, Z.; Zeng, L.; Xu, T.; Sun, S.; Yan, S.; Yang, L.; Huang, Y.; Jia, J.; Dou, T. Antibiotic Resistance Genes in Bacteria: Occurrence, Spread, and Control. J Basic Microbiol 2021, 61, 1049–1070. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Cheng, W. The Mechanism of Bacterial Resistance and Potential Bacteriostatic Strategies. Antibiotics 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Mulchandani, R.; Zhao, C.; Tiseo, K.; Pires, J.; Van Boeckel, T.P. Predictive Mapping of Antimicrobial Resistance for Escherichia Coli, Salmonella, and Campylobacter in Food-Producing Animals, Europe, 2000-2021. Emerg Infect Dis 2024, 30, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Guitor, A.K.; Raphenya, A.R.; Klunk, J.; Kuch, M.; Alcock, B.; Surette, M.G.; McArthur, A.G.; Poinar, H.N.; Wright, G.D. Capturing the Resistome: A Targeted Capture Method to Reveal Antibiotic Resistance Determinants in Metagenomes. Antimicrob Agents Chemother 2020, 64. [Google Scholar] [CrossRef] [PubMed]
- A PUBLIC HEALTH PERSPECTIVE ON ANTIMICROBIAL RESISTANCE DIAGNOSTICS; 2016;
- Rahman, M.M.; Alam Tumpa, M.A.; Zehravi, M.; Sarker, M.T.; Yamin, M.; Islam, M.R.; Harun-Or-rashid, M.; Ahmed, M.; Ramproshad, S.; Mondal, B.; et al. An Overview of Antimicrobial Stewardship Optimization: The Use of Antibiotics in Humans and Animals to Prevent Resistance. Antibiotics 2022, 11. [Google Scholar] [CrossRef]
- Baquero, F.; Martínez, J.L.; Lanza, V.F.; Rodríguez-Beltrán, J.; Galán, J.C.; San Millán, A.; Cantón, R.; Coque, T.M. Evolutionary Pathways and Trajectories in Antibiotic Resistance. Clin Microbiol Rev 2021, 34. [Google Scholar] [CrossRef]
- Xia, Y.; Li, X.; Wu, Z.; Nie, C.; Cheng, Z.; Sun, Y.; Liu, L.; Zhang, T. Strategies and Tools in Illumina and Nanopore-Integrated Metagenomic Analysis of Microbiome Data. iMeta 2023, 2. [Google Scholar] [CrossRef]
- Setubal, J.C. Metagenome-Assembled Genomes: Concepts, Analogies, and Challenges. Biophys Rev 2021, 13, 905–909. [Google Scholar] [CrossRef]
- Usyk, M.; Peters, B.A.; Karthikeyan, S.; McDonald, D.; Sollecito, C.C.; Vazquez-Baeza, Y.; Shaffer, J.P.; Gellman, M.D.; Talavera, G.A.; Daviglus, M.L.; et al. Comprehensive Evaluation of Shotgun Metagenomics, Amplicon Sequencing, and Harmonization of These Platforms for Epidemiological Studies. Cell Reports Methods 2023, 3. [Google Scholar] [CrossRef]
- Anjum, M.F.; Zankari, E.; Hasman, H. Molecular Methods for Detection of Antimicrobial Resistance. Microbiol Spectr 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.; Fasolino, T.; Davis, N.J.; Ivankovic, D.; Brownlee, N. Multiplex Detection of Antimicrobial Resistance Genes for Rapid Antibiotic Guidance of Urinary Tract Infections. Microbiol Res (Pavia) 2023, 14, 591–602. [Google Scholar] [CrossRef]
- de Abreu, V.A.C.; Perdigão, J.; Almeida, S. Metagenomic Approaches to Analyze Antimicrobial Resistance: An Overview. Front Genet 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Apjok, G.; Számel, M.; Christodoulou, C.; Seregi, V.; Vásárhelyi, B.M.; Stirling, T.; Eszenyi, B.; Sári, T.; Vidovics, F.; Nagrand, E.; et al. Characterization of Antibiotic Resistomes by Reprogrammed Bacteriophage-Enabled Functional Metagenomics in Clinical Strains. Nat Microbiol 2023, 8, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Köser, C.U.; Ellington, M.J.; Peacock, S.J. Whole-Genome Sequencing to Control Antimicrobial Resistance. Trends in Genetics 2014, 30, 401–407. [Google Scholar] [CrossRef]
- Burnard, D.; Gore, L.; Henderson, A.; Ranasinghe, A.; Bergh, H.; Cottrell, K.; Sarovich, D.S.; Price, E.P.; Paterson, D.L.; Harris, P.N.A. Comparative Genomics and Antimicrobial Resistance Profiling of Elizabethkingia Isolates Reveal Nosocomial Transmission and in Vitro Susceptibility to Fluoroquinolones, Tetracyclines, and Trimethoprim-Sulfamethoxazole. J Clin Microbiol 2020, 58. [Google Scholar] [CrossRef]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The Comprehensive Antibiotic Resistance Database. Antimicrob Agents Chemother 2013, 57, 3348–3357. [Google Scholar] [CrossRef]
- Florensa, A.F.; Kaas, R.S.; Clausen, P.T.L.C.; Aytan-Aktug, D.; Aarestrup, F.M. ResFinder – an Open Online Resource for Identification of Antimicrobial Resistance Genes in next-Generation Sequencing Data and Prediction of Phenotypes from Genotypes. Microb Genom 2022, 8. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-Annot, a New Bioinformatic Tool to Discover Antibiotic Resistance Genes in Bacterial Genomes. Antimicrob Agents Chemother 2014, 58, 212–220. [Google Scholar] [CrossRef]
- Bonin, N.; Doster, E.; Worley, H.; Pinnell, L.J.; Bravo, J.E.; Ferm, P.; Marini, S.; Prosperi, M.; Noyes, N.; Morley, P.S.; et al. MEGARes and AMR++, v3.0: An Updated Comprehensive Database of Antimicrobial Resistance Determinants and an Improved Software Pipeline for Classification Using High-Throughput Sequencing. Nucleic Acids Res 2023, 51, D744–D752. [Google Scholar] [CrossRef]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D.G.J. BacMet: Antibacterial Biocide and Metal Resistance Genes Database. Nucleic Acids Res 2014, 42. [Google Scholar] [CrossRef] [PubMed]
- Arango-Argoty, G.; Garner, E.; Pruden, A.; Heath, L.S.; Vikesland, P.; Zhang, L. DeepARG: A Deep Learning Approach for Predicting Antibiotic Resistance Genes from Metagenomic Data. Microbiome 2018, 6. [Google Scholar] [CrossRef]
- Liu, B.; Pop, M. ARDB - Antibiotic Resistance Genes Database. Nucleic Acids Res 2009, 37. [Google Scholar] [CrossRef]
- Gibson, M.K.; Forsberg, K.J.; Dantas, G. Improved Annotation of Antibiotic Resistance Determinants Reveals Microbial Resistomes Cluster by Ecology. ISME Journal 2015, 9, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Míguez, A.; Beghini, F.; Cumbo, F.; McIver, L.J.; Thompson, K.N.; Zolfo, M.; Manghi, P.; Dubois, L.; Huang, K.D.; Thomas, A.M.; et al. Extending and Improving Metagenomic Taxonomic Profiling with Uncharacterized Species Using MetaPhlAn 4. Nat Biotechnol 2023, 41, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Franzosa, E.A.; McIver, L.J.; Rahnavard, G.; Thompson, L.R.; Schirmer, M.; Weingart, G.; Lipson, K.S.; Knight, R.; Caporaso, J.G.; Segata, N.; et al. Species-Level Functional Profiling of Metagenomes and Metatranscriptomes. Nat Methods 2018, 15, 962–968. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog Facilitate Examination of the Genomic Links among Antimicrobial Resistance, Stress Response, and Virulence. Sci Rep 2021, 11. [Google Scholar] [CrossRef]
- Gillespie, J.J.; Wattam, A.R.; Cammer, S.A.; Gabbard, J.L.; Shukla, M.P.; Dalay, O.; Driscoll, T.; Hix, D.; Mane, S.P.; Mao, C.; et al. Patric: The Comprehensive Bacterial Bioinformatics Resource with a Focus on Human Pathogenic Species. Infect Immun 2011, 79, 4286–4298. [Google Scholar] [CrossRef]
- Inouye, M.; Dashnow, H.; Raven, L.A.; Schultz, M.B.; Pope, B.J.; Tomita, T.; Zobel, J.; Holt, K.E. SRST2: Rapid Genomic Surveillance for Public Health and Hospital Microbiology Labs. Genome Med 2014, 6. [Google Scholar] [CrossRef]
- Bharat, A.; Petkau, A.; Avery, B.P.; Chen, J.; Folster, J.; Carson, C.A.; Kearney, A.; Nadon, C.; Mabon, P.; Thiessen, J.; et al. Correlation between Phenotypic and In Silico Detection of Antimicrobial Resistance in Salmonella Enterica in Canada Using Staramr. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Seemann T Abricate, Https://Github.Com/Tseemann/Abricate.
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In Silico Detection and Typing of Plasmids Using Plasmidfinder and Plasmid Multilocus Sequence Typing. Antimicrob Agents Chemother 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Che, Y.; Dang, C.; Zhang, M.; Zhang, X.; Sun, Y.; Li, X.; Zhang, T.; Xia, Y. Nanopore-Based Long-Read Metagenomics Uncover the Resistome Intrusion by Antibiotic Resistant Bacteria from Treated Wastewater in Receiving Water Body. Water Res 2022, 226. [Google Scholar] [CrossRef] [PubMed]
- Kleinheinz, K.A.; Joensen, K.G.; Larsen, M.V. Applying the ResFinder and VirulenceFinder Web-Services for Easy Identification of Acquired Antibiotic Resistance and E. Coli Virulence Genes in Bacteriophage and Prophage Nucleotide Sequences. Bacteriophage 2014, 4, e27943. [Google Scholar] [CrossRef]
- Samtiya, M.; Matthews, K.R.; Dhewa, T.; Puniya, A.K. Antimicrobial Resistance in the Food Chain: Trends, Mechanisms, Pathways, and Possible Regulation Strategies. Foods 2022, 11. [Google Scholar] [CrossRef]
- Husna, A.; Rahman, M.M.; Badruzzaman, A.T.M.; Sikder, M.H.; Islam, M.R.; Rahman, M.T.; Alam, J.; Ashour, H.M. Extended-Spectrum β-Lactamases (ESBL): Challenges and Opportunities. Biomedicines 2023, 11. [Google Scholar] [CrossRef]
- Sah, A.K.; Feglo, P.K. Plasmid-Mediated Quinolone Resistance Determinants in Clinical Bacterial Pathogens Isolated from the Western Region of Ghana: A Cross-Sectional Study. Pan Afr Med J 2022, 43, 207. [Google Scholar] [CrossRef]
- Davies, A.R.; Chisnall, T.; Akter, S.; Afrad, M.M.H.; Sadekuzzaman, M.; Badhy, S.C.; Hasan, M.Z.; Rahman, M.T.; Smith, R.P.; Card, R.M.; et al. Genomic Characterisation of Escherichia Coli Isolated from Poultry at Retail through Sink Surveillance in Dhaka, Bangladesh Reveals High Levels of Multi-Drug Resistance. Front Microbiol 2024, 15. [Google Scholar] [CrossRef]
- Mondal, A.H.; Khare, K.; Saxena, P.; Debnath, P.; Mukhopadhyay, K.; Yadav, D. A Review on Colistin Resistance: An Antibiotic of Last Resort. Microorganisms 2024, 12. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Chen, H.; Li, N.; Wang, T.; Liang, W. The Spread of Antibiotic Resistance Genes In Vivo Model. Canadian Journal of Infectious Diseases and Medical Microbiology 2022, 2022. [Google Scholar] [CrossRef]
- Assar, S.; Hassanshahi, G.; Darehkordi, A.; Falahati-Pour, S.K.; Zarandi, E.R. Resistance Pattern of Escherichia Coli to Levofloxacin in Iran: A Narrative Review; 2020; Vol. 12;
- Mmatli, M.; Mbelle, N.M.; Osei Sekyere, J. Global Epidemiology, Genetic Environment, Risk Factors and Therapeutic Prospects of Mcr Genes: A Current and Emerging Update. Front Cell Infect Microbiol 2022, 12. [Google Scholar] [CrossRef]
- Osek, J.; Lachtara, B.; Wieczorek, K. Listeria Monocytogenes – How This Pathogen Survives in Food-Production Environments? Front Microbiol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Tóth, A.G.; Csabai, I.; Krikó, E.; Tőzsér, D.; Maróti, G.; Patai, Á. V.; Makrai, L.; Szita, G.; Solymosi, N. Antimicrobial Resistance Genes in Raw Milk for Human Consumption. Sci Rep 2020, 10. [Google Scholar] [CrossRef]
- Kayode, A.J.; Okoh, A.I. Assessment of Multidrug-Resistant Listeria Monocytogenes in Milk and Milk Product and One Health Perspective. PLoS One 2022, 17. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, R.D.; Agunos, A.; Gow, S.P.; Varga, C. Assessing Antimicrobial Resistance in Campylobacter Jejuni and Campylobacter Coli and Its Association with Antimicrobial Use in Canadian Turkey Flocks. Epidemiol Infect 2023, 151. [Google Scholar] [CrossRef] [PubMed]
- NARMS Integrated Report: 2014 The National Antimicrobial Resistance Monitoring System: Enteric Bacteria;
- Liu, D.; Liu, W.; Lv, Z.; Xia, J.; Li, X.; Hao, Y.; Zhou, Y.; Yao, H.; Liu, Z.; Wang, Y.; et al. Emerging Erm(B)-Mediated Macrolide Resistance Associated with Novel Multidrug Resistance Genomic Islands in Campylobacter. 2019. [CrossRef]
- Premarathne, J.M.K.J.K.; Anuar, A.S.; Thung, T.Y.; Satharasinghe, D.A.; Jambari, N.N.; Abdul-Mutalib, N.A.; Yew Huat, J.T.; Basri, D.F.; Rukayadi, Y.; Nakaguchi, Y.; et al. Prevalence and Antibiotic Resistance against Tetracycline in Campylobacter Jejuni and C. Coli in Cattle and Beef Meat from Selangor, Malaysia. Front Microbiol 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- De, R. Mobile Genetic Elements of Vibrio Cholerae and the Evolution of Its Antimicrobial Resistance. Frontiers in Tropical Diseases 2021, 2. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance. Microbiology and Molecular Biology Reviews 2001, 65, 232–260. [Google Scholar] [CrossRef]
- Schmidt, K.; Scholz, H.C.; Appelt, S.; Michel, J.; Jacob, D.; Dupke, S. Virulence and Resistance Patterns of Vibrio Cholerae Non-O1/Non-O139 Acquired in Germany and Other European Countries. Front Microbiol 2023, 14. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Álvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; et al. Role Played by the Environment in the Emergence and Spread of Antimicrobial Resistance (AMR) through the Food Chain. EFSA Journal 2021, 19. [Google Scholar] [CrossRef]
- Kiu, R.; Hall, L.J. An Update on the Human and Animal Enteric Pathogen Clostridium Perfringens. Emerg Microbes Infect 2018, 7. [Google Scholar] [CrossRef]
- Lina, G.; Quaglia, A.; Reverdy, M.-E.; Leclercq, R.; Vandenesch, F.O.; Etienne, J. Distribution of Genes Encoding Resistance to Macrolides, Lincosamides, and Streptogramins among Staphylococci; 1999; Vol. 43;
- Beres, C.; Colobatiu, L.; Tabaran, A.; Mihaiu, R.; Mihaiu, M. Prevalence and Characterisation of Clostridium Perfringens Isolates in Food-Producing Animals in Romania. Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J. xin; Zheng, H. ran; Wang, Y. yuan; Bai, L. lu; Du, X. li; Wu, Y.; Lu, J. xing Molecular Characteristics and Phylogenetic Analysis of Clostridium Perfringens from Different Regions in China, from 2013 to 2021. Front Microbiol 2023, 14. [Google Scholar] [CrossRef]
- Asad, A.; Jahan, I.; Munni, M.A.; Begum, R.; Mukta, M.A.; Saif, K.; Faruque, S.N.; Hayat, S.; Islam, Z. Multidrug-Resistant Conjugative Plasmid Carrying MphA Confers Increased Antimicrobial Resistance in Shigella. Sci Rep 2024, 14. [Google Scholar] [CrossRef]
- Hussain, H.I.; Aqib, A.I.; Seleem, M.N.; Shabbir, M.A.; Hao, H.; Iqbal, Z.; Kulyar, M.F. e. A.; Zaheer, T.; Li, K. Genetic Basis of Molecular Mechanisms in β-Lactam Resistant Gram-Negative Bacteria. Microb Pathog 2021, 158. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J. Transferable Mechanisms of Quinolone Resistance from 1998 Onward; 2019;
- Ma, Q.; Zhu, C.; Yao, M.; Yuan, G.; Sun, Y. Correlation between the Sulfamethoxazole-Trimethoprim Resistance of Shigella Flexneri and the Sul Genes. Medicine (United States) 2021, 100, E24970. [Google Scholar] [CrossRef] [PubMed]
- Sadeghabadi, A.F.; Ajami, A.; Fadaei, R.; Zandieh, M.; Heidari, E.; Sadeghi, M.; Ataei, B.; Hoseini, S.G. Widespread Antibiotic Resistance of Diarrheagenic Escherichia Coli and Shigella Species; 2014;
- Rawson, A.M.; Dempster, A.W.; Humphreys, C.M.; Minton, N.P. Pathogenicity and Virulence of Clostridium Botulinum. Virulence 2023, 14. [Google Scholar] [CrossRef]
- Roberts, M.C. Environmental Macrolide-Lincosamide-Streptogramin and Tetracycline Resistant Bacteria. Front Microbiol 2011, 2. [Google Scholar] [CrossRef]
- Swenson, J.M.; Thornsberry, C.; Mccroskey, L.M.; Hatheway, C.L.; Dowell, V.R. Susceptibility of Clostridium Botulinum to Thirteen Antimicrobial Agents; 1980; Vol. 18;
- Douillard, F.P.; Derman, Y.; Woudstra, C.; Selby, K.; Mäklin, T.; Dorner, M.B.; Saxén, H.; Dorner, B.G.; Korkeala, H.; Lindström, M. Genomic and Phenotypic Characterization of Clostridium Botulinum Isolates from an Infant Botulism Case Suggests Adaptation Signatures to the Gut. mBio 2022, 13. [Google Scholar] [CrossRef]
- Karlsson, P.A.; Tano, E.; Jernberg, C.; Hickman, R.A.; Guy, L.; Järhult, J.D.; Wang, H. Molecular Characterization of Multidrug-Resistant Yersinia Enterocolitica From Foodborne Outbreaks in Sweden. Front Microbiol 2021, 12. [Google Scholar] [CrossRef]
- Gkouletsos, T.; Patas, K.; Lambrinidis, G.; Neubauer, H.; Sprague, L.D.; Ioannidis, A.; Chatzipanagiotou, S. Antimicrobial Resistance of Yersinia Enterocolitica and Presence of Plasmid PYV Virulence Genes in Human and Animal Isolates. New Microbes New Infect 2019, 32. [Google Scholar] [CrossRef]
- Fredriksson-Ahomaa, M.; Grönthal, T.; Heljanko, V.; Johansson, V.; Rantala, M.; Heikinheimo, A.; Laukkanen-Ninios, R. Enteropathogenic Yersinia with Public Health Relevance Found in Dogs and Cats in Finland. Pathogens 2024, 13. [Google Scholar] [CrossRef]
- Ray, L.C.; Payne, D.C.; Rounds, J.; Trevejo, R.T.; Wilson, E.; Burzlaff, K.; Garman, K.N.; Lathrop, S.; Rissman, T.; Wymore, K.; et al. Syndromic Gastrointestinal Panel Diagnostic Tests Have Changed Our Understanding of the Epidemiology of Yersiniosis - Foodborne Diseases Active Surveillance Network, 2010-2021. Open Forum Infect Dis 2024, 11. [Google Scholar] [CrossRef]
- Perry, J.A.; Westman, E.L.; Wright, G.D. The Antibiotic Resistome: What’s New? Curr Opin Microbiol 2014, 21, 45–50. [Google Scholar] [CrossRef]
- Khmelevtsova, L.; Azhogina, T.; Karchava, S.; Klimova, M.; Polienko, E.; Litsevich, A.; Chernyshenko, E.; Khammami, M.; Sazykin, I.; Sazykina, M. Effect of Mineral Fertilizers and Pesticides Application on Bacterial Community and Antibiotic-Resistance Genes Distribution in Agricultural Soils. Agronomy 2024, 14. [Google Scholar] [CrossRef]
- Zhang, W.G.; Wen, T.; Liu, L.Z.; Li, J.Y.; Gao, Y.; Zhu, D.; He, J.Z.; Zhu, Y.G. Agricultural Land-Use Change and Rotation System Exert Considerable Influences on the Soil Antibiotic Resistome in Lake Tai Basin. Science of the Total Environment 2021, 771. [Google Scholar] [CrossRef]
- Durso, L.M.; Cook, K.L. Impacts of Antibiotic Use in Agriculture: What Are the Benefits and Risks? Curr Opin Microbiol 2014, 19, 37–44. [Google Scholar] [CrossRef]
- Barathan, M.; Ng, S.L.; Lokanathan, Y.; Ng, M.H.; Law, J.X. Unseen Weapons: Bacterial Extracellular Vesicles and the Spread of Antibiotic Resistance in Aquatic Environments. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Zhu, S.; Yang, B.; Yu, F.; Zhang, J.; Wang, Z.; Liu, Y. Investigation of the Impact of Widely Used Pesticides on Conjugative Transfer of Multidrug Resistance Plasmids. J Hazard Mater 2024, 478. [Google Scholar] [CrossRef]
- Chen, T.; Wang, Z.; Ruan, X. Antibiotic Resistome Dynamics in Agricultural River Systems: Elucidating Transmission Mechanisms and Associated Risk to Water Security. Science of the Total Environment 2024, 951. [Google Scholar] [CrossRef]
- Oulas, A.; Pavloudi, C.; Polymenakou, P.; Pavlopoulos, G.A.; Papanikolaou, N.; Kotoulas, G.; Arvanitidis, C.; Iliopoulos, I. Metagenomics: Tools and Insights for Analyzing next-Generation Sequencing Data Derived from Biodiversity Studies. Bioinform Biol Insights 2015, 9, 75–88. [Google Scholar] [CrossRef]
- Roumpeka, D.D.; Wallace, R.J.; Escalettes, F.; Fotheringham, I.; Watson, M. A Review of Bioinformatics Tools for Bio-Prospecting from Metagenomic Sequence Data. Front Genet 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Inda-Díaz, J.S.; Lund, D.; Parras-Moltó, M.; Johnning, A.; Bengtsson-Palme, J.; Kristiansson, E. Latent Antibiotic Resistance Genes Are Abundant, Diverse, and Mobile in Human, Animal, and Environmental Microbiomes. Microbiome 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Lanza, V.F.; Baquero, F.; Martínez, J.L.; Ramos-Ruíz, R.; González-Zorn, B.; Andremont, A.; Sánchez-Valenzuela, A.; Ehrlich, S.D.; Kennedy, S.; Ruppé, E.; et al. In-Depth Resistome Analysis by Targeted Metagenomics. Microbiome 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Van Der Helm, E.; Imamovic, L.; Ellabaan, M.M.H.; Van Schaik, W.; Koza, A.; Sommer, M.O.A. Rapid Resistome Mapping Using Nanopore Sequencing. Nucleic Acids Res 2017, 45. [Google Scholar] [CrossRef] [PubMed]
- Martiny, H.M.; Munk, P.; Brinch, C.; Aarestrup, F.M.; Petersen, T.N. A Curated Data Resource of 214K Metagenomes for Characterization of the Global Antimicrobial Resistome. PLoS Biol 2022, 20. [Google Scholar] [CrossRef]
- Lee, K.; Kim, D.W.; Cha, C.J. Overview of Bioinformatic Methods for Analysis of Antibiotic Resistome from Genome and Metagenome Data. Journal of Microbiology 2021, 59, 270–280. [Google Scholar] [CrossRef]
- Liu, F.; Luo, Y.; Xu, T.; Lin, H.; Qiu, Y.; Li, B. Current Examining Methods and Mathematical Models of Horizontal Transfer of Antibiotic Resistance Genes in the Environment. Front Microbiol 2024, 15. [Google Scholar] [CrossRef]
- Yarygin, K.S.; Kovarsky, B.A.; Bibikova, T.S.; Melnikov, D.S.; Tyakht, A. V.; Alexeev, D.G. ResistoMap-Online Visualization of Human Gut Microbiota Antibiotic Resistome. In Proceedings of the Bioinformatics; Oxford University Press, July 15 2017; Vol. 33; pp. 2205–2206. [Google Scholar]
- Munkholm, L.; Rubin, O. The Global Governance of Antimicrobial Resistance: A Cross-Country Study of Alignment between the Global Action Plan and National Action Plans. Global Health 2020, 16. [Google Scholar] [CrossRef]
- Edelstein, M.; Lee, L.M.; Herten-Crabb, A.; Heymann, D.L.; Harper, D.R. Strengthening Global Public Health Surveillance through Data and Benefit Sharing. Emerg Infect Dis 2018, 24, 1324–1330. [Google Scholar] [CrossRef]
Figure 1.
Overview of mechanism of antimicrobial resistance in bacteria (Created in BioRender.com).
Figure 1.
Overview of mechanism of antimicrobial resistance in bacteria (Created in BioRender.com).
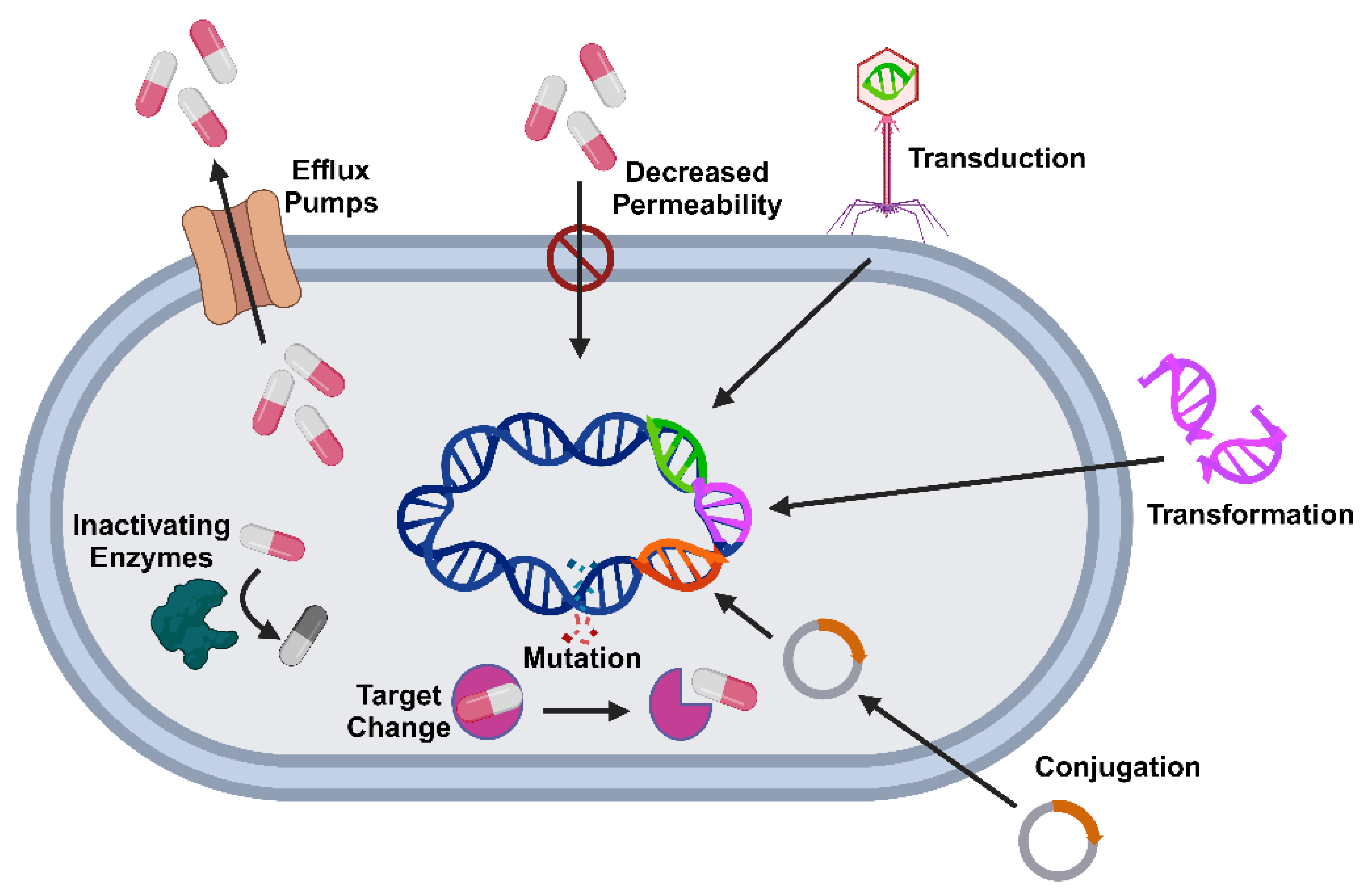

Table 1.
Bioinformatic tools and databases available for Resistome Mapping.
| Database/Tool | Role in Resistome Analysis | Website | Reference |
|---|---|---|---|
| CARD (Comprehensive Antibiotic Resistance Database) | A curated database containing information on resistance genes and their mechanisms, using for the detection and annotation of AMR genes. | CARD | [40] |
| ResFinder | Identifies acquired ARGs from bacterial genome data. | ResFinder | [41] |
| ARG-ANNOT | Antibiotic Resistance Gene-ANNOTation tool providing ARG identification based on curated sequences. | ARG-ANNOT | [42] |
| MEGARes | Database of ARGs used in resistome analysis of metagenomic datasets. | MEGARes | [43] |
| BacMet | A database of antibacterial biocide- and metal-resistance genes. | BacMet | [44] |
| DeepARG | A tool that uses deep learning to predict ARGs from metagenomic data. | DeepARG | [45] |
| Antibiotic Resistance Genes Database (ARDB) | A comprehensive resource of ARGs and their associated phenotypes. | ARDB | [46] |
| Resfams | A collection of protein families associated with antibiotic resistance, for the functional annotation of ARGs. | Resfams | [47] |
| Antibiotic Resistance Ontology (ARO) | A standardized vocabulary of resistance genes, phenotypes, and mechanisms, using for AMR classification | ARO | [40] |
| MetaPhlAn | A tool for profiling microbial communities, including the identification of ARGs. | MetaPhlAn | [48] |
| Humann2 | Tool for functional profiling of microbial communities, with ARG identification. | Humann2 | [49] |
| AMRFinderPlus | Identifies AMR genes, virulence factors, and other genes from whole genome sequences. | AMRFinderPlus | [50] |
| RGI (Resistance Gene Identifier) | A tool integrated with CARD to detect and annotate ARGs from genomic and metagenomic sequences. | RGI | [40] |
| PATRIC | A bacterial bioinformatics platform comparative genomics and AMR gene detection. | PATRIC | [51] |
| SRST2 (Short Read Sequence Typing for Bacterial Pathogens) | A tool for strain typing and detecting ARGs from short-read sequence data. | SRST2 | [52] |
| StarAMR | A tool for identifying ARGs from assembled genome sequences. | StarAMR | [53] |
| ABRicate | A command-line tool that screens bacterial genomes for ARGs using multiple databases (e.g., CARD, ResFinder). | ABRicate | [54] |
| PlasmidFinder | Identifies plasmid replicons in whole-genome sequencing data. Tools helps to identify the detection of plasmids related to ARGs. | PlasmidFinder | [55] |
| ARGpore | Detects ARGs from long-read (nanopore) sequencing data. | ARGpore | [56] |
| VirulenceFinder | Identifies virulence genes in bacterial genomes that may be associated with antimicrobial resistance. | VirulenceFinder | [57] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



