
Submitted:
21 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
Ceratitis capitata (Wiedemann) (medfly) strongly affects Argentinean fruit production and export. Augmentative biological control using the exotic parasitoid Diachasmimorpha longicaudata (Ashmead) is currently applied. The ability to find and parasitize medfly larvae on a wide diversity of fruit host species is a key issue to be analyzed. This research assessed the effect of the physical features of fruit on the preference of foraging D. longicaudata females and the influence of varying release density on parasitoid performance as a pest mortality factor in three fruit species. Trials were performed inside field cages under semi-arid environmental conditions in Argentina's central–western fruit-growing region. Sweet orange, peach, and fig were tested. Fruits were inoculated with third-instar larvae of the Vienna-8 temperature-sensitive lethal medfly strain. Naïve, 5-d-old mated D. longicaudata females were released in cages at 20, 40, 80 and 160 parasitoid densities. The highest levels of medfly mortality and parasitoid emergence were recorded in fig and peach, although D. longicaudata also induced mortality in orange, a fruit with few physical features favorable to parasitism. The medfly mortality in all fruit host species significantly increased with increased parasitoid females released into field cages. Diachsmimorpha longicaudata has high potential as a medfly biocontrol agent.
Keywords:
medfly
; host frutis
; semi-arid environmental
; larval parasitoid performance
; biological control
; augmentative parasitoid release
1. Introduction
Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) is an invasive dipterous species native to sub-Saharan Africa, although it is widespread throughout the African continent [1]. Commonly known as the Mediterranean fruit fly or just medfly, it is currently regarded as a cosmopolitan species due to its worldwide dispersal, mainly resulting from the increase in the global fruit trade [2]. The medfly is a highly polyphagous pest species that infests healthy, commercially valuable fruit [3]. The pattern of host relationships is mainly related to the fruits available locally in the region where the medfly occurs [4]. The economic damage caused by the medfly is related to its biological cycle [5]. After mating, females lay eggs inside healthy fruit, causing additional damage to the epidermis and allowing fruit-rotting organisms such as insects and/or pathogens to access them. Eggs give rise to larvae that pass through three instars while feeding on the fruit flesh, which leads to softening and rotting of the fruit. The mature larvae leave the fruit to pupate by burying themselves in the soil or within or beneath the fallen fruit. The adult emerges from the puparium and searches for a host plant to access water, food, and shelter, and the female searches for an optimal environment for mating and oviposition. From the first record of medfly infesting commercial peaches in Argentina in 1905 [6], this pest has been characterized by its rapid range expansion throughout all Argentinian fruit-growing regions, covering latitudes from 22° to 56°S. However, fruit-producing areas of the Patagonian Region (southern Argentina) and the Central and Southern Oases of Mendoza from the Cuyo region (southwestern Argentina) are nowadays fruit fly-free areas [7]. The medfly has been reported to infest 58 commercial and wild, exotic, and indigenous fruit species grown in fruit-producing regions throughout Argentina [8,9]. Given that Argentina is a relevant world producer of fresh fruits and vegetables due to its diverse ecosystems [10], the medfly widespread is a critical constraint for the country’s fruit-growing industry. In this regard, C. capitata is one of the main fruit pests that strongly affects the production, marketing, and export of fruits, which has a negative socioeconomic impact on Argentinean fruit systems [11]. This pest causes economic losses either by direct damage to fruits, such as the presence of larvae inside fruit or oviposition activity of females, or by indirect losses, which involve quarantine treatments or other measures that increase marketing costs in response to export restrictions imposed by importing countries [8]. Therefore, given the adverse effects of the medfly on the Argentinian economy, since 1994, the federal government put into operation the National Fruit Fly Control and Eradication Program (PROCEM, Spanish acronym) [12], which currently involves area-wide integrated medfly management approaches. The strategy applied through PROCEM has been based on the combined use of the sterile insect technique (SIT), cultural and air/ground chemical controls, trapping systems, a quarantine protection system, and the implementation of a phytosanitary emergency schedule for pest outbreaks in free- or low prevalence areas [13]. However, from 2014 to the present, augmentative biological control has been used by releasing parasitoids in fruit crops by the PROCEM in semi-arid fruit production valleys of the province of San Juan (central-western Argentina, Cuyo region) as a complementary tool in an environmentally friendly way [14].
The implementation of augmentative biological control against fruit flies in Argentina was achieved through the establishment and optimization of a mass rearing of the South Asian-native, larval parasitoid Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae) [15]. Such exotic braconid parasitoid is currently reared using cobalt-60 irradiated medfly larvae of the temperature-sensitive lethal (= tsl) genetic sexing Vienna-8 strain at the San Juan mass-rearing biofactory [16]. Recent D. longicaudata post-augmentative release data showed a significant decrease in the wild medfly population inhabiting a commercial multi-fruit farm [14]. These results, added to the previous ones reported by [15] on a medfly-infested fig crop where over 70% of pest mortality was due to parasitoid activity, encourage D. longicaudata use in San Juan’s fruit-growing oases. Furthermore, the need to promote sustainable and eco-friendly agricultural strategies in arid areas in a progressively warming and drying world, as pointed out by [17], highlights the need to focus on this issue. Likewise, the lack of resident parasitoid species that attack the pest in the region [18], reinforces the use of biologically plastic-imported species, i.e., those species capable of adapting to different environmental conditions, such as the braconid D. longicaudata [19]. Earlier studies [14,15] showed how D. longicaudata mass-reared at the San Juan biofactory can perform effectively as a biocontrol agent under semi-arid environmental conditions in ecologically isolated fruit-growing valleys of San Juan. In those areas, the climatic characteristics of arid lands, such as high daytime and low overnight temperatures, drought, strong winds, low relative humidity, sparse vegetation, and low alternative host densities, are considerably attenuated. Another essential factor that must be addressed involves host species diversity in the fruit-growing region and their influence on parasitoid effectiveness in finding and parasitizing host larvae. Large fruits, such as citrus, can limit parasitoid foraging behavior on fruit-infesting Tephritidae [20,21,22]. Therefore, the host fruit’s physical features have important practical implications for implementing medfly biological control in any fruit-producing region.
Several authors highlighted the ability of D. longicaudata to forage successfully on host larvae over a wide range of non-crop and crop fruit species [19,23,24,25,26,27]. Given this, it is hypothesized that the D. longicaudata female is particularly efficient at finding and killing medfly larvae infesting diverse commercial host fruit species ranging from orange, fig, and peach in irrigated fruit-growing areas of San Juan. In addition, it is also hypothesized that the parasitoid female becomes significantly more efficient at increasing its release density. This assumption is based on several open-field augmentative release trials using D. longicaudata for suppressing pest tephritid fruit fly populations worldwide [23,27,28,29,30,31]. Therefore, this research aims to evaluate the effect of the variation in fruit species preference by foraging D. longicaudata females, regarding the fruit’s physical features and the potential effect of varying release density on parasitoid performance as a pest mortality factor in three fruit species. Thus, this study discusses the findings of using D. longicaudata mass reared at the San Juan biofactory as a biological control agent against the medfly in one of Argentina’s most important fruit-growing regions recognized for its semi-arid climate marked by hot and cool extreme temperatures.
2. Materials and Methods
2.1. Insect Rearing Procedures
Diachasmimorpha longicaudata adult females were reared on 90 Gy-irradiated medfly larvae of the Vienna-8 tsl strain using 60 × 60 × 30 cm rectangular iron-framed voile screen-covered cages at the Parasitoid Rearing Laboratory of the San Juan mass-rearing biofactory. This laboratory belongs to the Plant, Animal, and Food Health Bureau of the government of the San Juan province, located in the central-western fruit-growing region of Argentina. Parasitoid cohorts were reared at 25 ± 1 ◦C, 60 ± 5% RH, 12:10 (L:D) photoperiod, and were fed honey and water ad libitum every other day. Medfly tsl larvae were reared at the Fruit Fly Rearing Laboratory of the San Juan biofactory based on Caceres’ [32] protocol. Medfly larvae radiation was performed in an IMO-1 mobile Gamacell irradiator with a Co-60 source of γ irradiation, located at the San Juan biofactory, and belongs to the National Atomic Energy Commission from Argentina. Naked irradiated host larvae (i.e., no artificial larval diet) were exposed daily to mated parasitoid females in 15 × 1.3-cm (diameter x height) voile screen-covered sandwich-type oviposition dishes. These devices were put on top of producing cages when parasitoid females were 5 d-old. Host exposure time was 4 h under a lighting condition of 2400 lux provided by twelve 36 W-fluorescent light tubes homogeneously dispersed throughout the rearing room.
2.2. Test site and weather conditions
The trials were performed inside field cages under uncontrolled environmental conditions at a 1,000-m2 experimental field of the Plant, Animal and Food Health Bureau (31°31′00′′S, 68°36′00′W, 710 m), located in the Rivadavia District, Tulum Valley, San Juan. A windbreak curtain of poplar trees (Populus alba L., Salicaceae) surrounds the experimental plot supplying natural shade within the site (Figure 1). According to Köeppen’s classification, the San Juan lowlands, in which the main fruit-producing valleys are found, possess a BWwka climate, i.e. a desert climate with a high concentration of rainfall in the summer, with a mean annual temperature below 18º C [33]. Mean relative humidity (= RH), mean air temperature (= AT), and precipitation were recorded during the study using a digital weather station (LUFT®, model WS80, China). Both HR and AT and accumulated rainfall recorded during trial dates are shown in Table 1.
2.3. Tested Host Fruit Species and Fruit Handling
Three fruit species were tested: Citrus sinensis (L.) Osbeck (“sweet orange”, Salustiana cultivar), Prunus persica (L.) Batsch (“peach”, Hesse cultivar), and Ficus carica L. (“fig”, Brown Turkey cultivar). The fruits were purchased at the market and, once brought to the laboratory, were washed and disinfected with a sodium benzoate and nipagin methylparaben solution. Before tests, each fruit was weighed, and rind thickness, flesh depth, and fruit diameter were measured, and the surface area (= 4πr 2) over which the female parasitoid will forage was also calculated. Fruits of similar size and ripeness were selected for the trials. Each washed fruit was artificially inoculated with non-irradiated third-instar larvae of the tsl medfly strain. Host larval density inoculated on fruit remained constant in all tests. Larval density per fruit species depended on the fruit surface area of each fruit species; therefore, a significantly equal larval density was used relative to fruit surface area. Thus, each orange or peach was inoculated with 130 medfly larvae to match larval proportions per fruit species, while the fig was inoculated with 100 host larvae (see Table 2). Inoculation was carried out by cutting the top of the fruit using a sterilized scalpel; half of the pulp was removed, and the fruit was filled with host larvae. They occupied 50% of the fruit volume. In the case of the peach, the stone was removed. Once the fruit was inoculated, the small upper cover (lid) and the large lower portion (fruit body) were attached with 2.5-cm wide Parafilm “M” ® sheets (Pechiney Plastic Packaging, Chicago, USA). Each infested fruit accounted for one oviposition unit.
2.4. Experimental Setup and Procedure
The study mainly assessed the host-killing capacity of the Diachasmimorpha longicaudata females mass-reared at the San Juan Biofactory by attacking medfly larvae on three primary host fruit species grown in the region. These tests were performed under a natural free-foraging scenario and with variation in parasitoid female release densities. Trials were carried out inside three 3.5 × 3.0-m (diameter × height) nylon field-cages throughout March 2019 and March 2020, corresponding to late summer and early autumn. Five 0.5 × 2.0-m (diameter × height) experimental tube voile mesh-covered cages, reinforced with 16-gauge galvanized wire distributed across the width and height of the experimental cage, were placed inside each major field cage. Each tube cage had a 1.5-m long zipper along its middle part to allow access to its interior. Inside the tube cages, a potted orange, peach, or fig seedling was placed according to the fruit species of the treatment. The tube cages were placed at 20 cm distance from each other inside the major field cage (Figure 1). Each field cage was protected from solar radiation by a 130-g/m2 black shade net and from rain by a 100-μ translucent high-density polyethylene cover (AGROREDES®, Buenos Aires, Argentina). Both coverings were 40 cm on top of the cage, allowing natural light and preventing rainwater. For each tube cage, three fruits inoculated with host larvae were placed. The fruit species used varied according to the treatment. The fruits were placed equidistant in a circle only in the upper section, at 1.5 m from the cage roof. The fruits were hanged from the top using a 0.5-mm nylon fishing-line attached to a metal clip inserted into the Parafilm strip wrapped around the fruit. Immediately below the fruit was placed a 10 × 2-cm (diameter × height) galvanized wire basket with a double voile covering, with a poplar shaving at the bottom as a pupation substrate. The device was held with four nylon fishing-lines tied to the main rope from which the fruit hung. Each major field cage accounted for one treatment as follows: treatment #1 involved only peach (= T1), treatment #2 sweet orange (= T2), treatment #3 fig (= T3). Each treatment was divided into four sub-treatments with different parasitoid release densities and one control. Sub-treatment locations were randomized to avoid the edge effects. All sub-treatments and the control were simultaneously performed. In sub-treatment “A” 20 parasitoid females were released (= TA), sub-treatment “B” involved 40 parasitoid females (= TB), sub-treatment “C” 80 parasitoids released (= TC), and sub-treatment “D” 160 parasitoids released (= TD) (Figure 2). Three controls were tested as follows: controls #1, #2, and #3, involved only peaches, oranges, and figs infested with medfly larvae, respectively, but no parasitoids were released (= C1, C2, and C3, respectively) (Figure 1). The host-parasitoid ratio per sub-treatment involving sweet orange and peach was 6.5, 3.3, 1.6, and 0.8 host larvae per 1 parasitoid female released, whereas in the fig sub-treatment was 5, 2.5, 1.3, and 0.6 host per 1 parasitoid female. The trials were conducted with Naïve (non-exposed to host or fruit or both) 5-d-old mated D. longicaudata females of 65th generations. Parasitoids were released into experimental tube cages on leaves of the potted tree, and they were allowed to forage infested fruit for 48-h, starting at 9 AM. Parasitoid females were only allowed to forage on fruit from the higher section of the experimental tube cage, close to the seedlings. This was due to previous studies showing a similar ability of D. longicaudata females to forage on infested fruit on the ground and the tree [34,35]. Once foraging time was over, the fruit, the pupation substrate, and parasitoids were removed from each tube cage. At the laboratory, fruits were dissected to remove living host larvae, and puparia were recovered from each basket. All hosts were placed in 10 × 8-cm (diameter × height) disposable plastic cups with sterilized poplar shaving at the bottom until insect emergence. Those devices were kept at 26 ± 1°C, 75 ± 5% RH, and darkness. Once insects emerged, the number and sex of the parasitoids and the number of adult flies were recorded from each sub-treatment and control test. Based on those data, biological parameters were determined as follows: (1) the host-killing parasitoid ability, (2) the parasitoid reproductive success, and (3) the parasitoid offspring sex ratio. The first parameter is equivalent to the parasitoid performance as a pest killer, and it was calculated using Abbott’s percent-corrected host mortality index [36]. This index allows determining the host-killing parasitoid female capacity by the interaction between host emergence from the treatment and that from the control. The parasitoid reproductive success was estimated as the number of emerged parasitoid offspring divided by the number of exposed larvae inside the fruit × 100. The sexual ratio of parasitoid offspring was calculated as the percentage of emerged females divided into all recovered adult parasitoids. Each sub-treatment and the related control were replicated 10 times. Each replicate involved new inoculated fruit and pupation substrate on each basket and a new parasitoid female cohort.
2.5. Data Analysis
The host killing, parasitoid reproductive success, and the parasitoid offspring sex ratio data were analyzed as nested models, with three-factor levels (year, fruit, and number of parasitoids realized), implementing Generalized Linear Mixed Models (GLMM) performed with the function “lme” form “nlme” [37,38] packages of R [39]. The final model was implemented using only significant factors at α= 0.05. For paired data, the comparison of host killing, reproductive success, and sex ratio throughout the experiment and within treatments, without considering factors, was made with a Wilcoxon signed rank test with continuity correction, with the “wilcox.test” function from the “stasts” based packages of R [39]. Comparisons of the fruit features were made using the Kruskal-Wallis rank sum test, “kruskal.test” function from the “stasts” based packages of R [39]. The R-4.4.1 software was used for statistical analysis [39]. The box plot figures show the median (horizontal line inside the box), interquartile range Q1–Q3 (bottom and top ends of the box), range (minimum: Q0, maximum: Q4; both ends of the whisker on the vertical line outside the box), and outliers (colored circles).
3. Results
3.1. Tested Host Fruit Species
Physical features between tested medfly-host fruit species were significantly different (weight, χ2= 79.12, df= 2, p < 0.01; diameter, χ2= 74.50, df= 2, p < 0.01; rind thickness, χ2= 79.12, df= 2, p < 0.01; pulp depth, χ2= 79.12, df= 2, p < 0.01; surface area, χ2= 62.79, df= 2, p < 0.01). The orange had the highest physical properties compared to peach and fig (Table 2). Peach differed significantly from fig in all physical traits, while the host density/cm2 of fruit surface used in the trials was significantly similar for all fruit species (Table 2).
3.2. Host-Killing Parasitoid Capacity
The medfly-killing parasitoid capacity parameter did not vary significantly between the two study years (Estimate = 1.20, SE = 0.94, Z = 1.27, p = 0.20). Therefore, the data from each testing year were pooled for analysis. Results for each tested host fruit species and by parasitoid release density are shown in Figure 2. The parasitoid females killed between 54 and 88% of host larvae in the fig, between 52 and 83% in the peach, and between 34 and 62% in the sweet orange. When all treatments were pooled, the host-killing parasitoid capacity by tested fruit species was significantly higher in the fig (71% marginal mean), followed by the peach (67%), and the lowest in the sweet orange (46%) (File S1). Overall, the host-killing parasitoid capacity was significantly higher as parasitoid release density increased, whatever tested fruit species. The marginal means of the host mortality percentage pooling all fruit species were ~47, 53, 69, and 78 for treatments #1, #2, #3 and #4, respectively (File S1).
3.3. Parasitoid Reproductive Success
In the case of parasitoid reproductive success, there is a small but significant difference between the two years studied (Estimate = 0.81 SE = 0.34, Z = -2.39, p = 0.02). Thus, the nested model was performed for this parameter at three levels (studying year, fruit species, and parasitoid release density). Results of the parasitoid reproductive success for each host fruit species and by parasitoid release density are shown in Figure 3. The percentage of offspring produced by D. longicaudata females from fig, peach, and sweet orange varied between 19 and 30%, 20 and 28%, and between 11 and 21%, respectively. Both fig and peach were the two host fruit species with the most substantial influence on increasing parasitoid offspring when pooling treatments (25 and 24% marginal means for fig and peach, respectively). In contrast, orange had the slightest impact on the parasitoid reproductive success significantly (16 % marginal mean) (File S3). When treatments were pooled, the parasitoid reproductive success significantly increased as parasitoid release density increased in all host fruit species. Marginal means of parasitoid offspring percentage were 17, 19, 24, and 27 for treatments #1, #2, #3, and #4, respectively (File S4).
3.4. Parasitoid Offspring Sex Ratio
The parasitoid offspring sex ratio did not vary significantly between the two study years (Estimate = 0.28, SE = 0.80, Z = 0.35, p = 0.72). Therefore, the data from each testing year were pooled for analysis. Results of emerged parasitoid females for each host fruit species and by parasitoid release density are shown in Figure 4. Diachasmimorpha longicaudata exhibited about 1:1 female:male sex ratio, as the percentage of females emerged from medfly puparia recovered from all three host fruit species ranged from 49 to 51%. There were no significant differences between marginal means of parasitoid offspring sex ratio when both tested factors, host fruit species (File S5) and parasitoid release densities (File S6), were compared.
4. Discussion
Augmentative biological control using parasitoids has become a relevant complementary tool with other eco-friendly techniques for medfly control [27,31]. Therefore, parasitoid species selected as potentially effective medfly biocontrol agents entail a prior and exhaustive assessment of their host foraging and killing abilities in different host fruits and environments. Understanding those biological aspects is strategically essential for developing and implementing a parasitoid mass-release biological control schedule. In this framework, the current study reports the ability of the introduced parasitoid D. longicaudata to kill the invasive C. capitata infesting main host fruit species through field-cage simulated augmentative releases. Consequently, the results highlighted three major findings. Firstly, D. longicaudata females successfully parasitized host larvae on all three tested medfly-multiplying fruit species. Secondly, although the highest levels of both medfly mortality and parasitoid reproductive success were recorded in those fruit species with physical features highly favorable to parasitism, such as fig and peach, D. longicaudata was also able to induce significant mortality in orange, which has a thicker rind and a deeper flesh that tends to enhance host larva protection. Thirdly, the medfly mortality recorded on all tested fruit host species significantly increased as the number of parasitoid females released into field cages increased.
The first finding highlighted the ability of D. longicaudata to successfully find and parasitize medfly larvae on the three most important host fruits occurring in the central-western fruit-producing region of Argentina. Such information verifies previous host fruit surveys, which found medfly larvae parasitized by D. longicaudata on figs and sweet oranges (unknown cultivars) [18] and on peaches (Elegant Lady variety) [15], following releases of this parasitoid throughout different fruit-growing valleys of San Juan. The fig, peach, and sweet orange are characterized mainly by their physical differences and are widespread in all the irrigated fruit-growing valleys of the San Juan province. Figs and peaches are not only grown commercially in central-western Argentina, but they are also common fruit trees, like sweet orange cultivars, in backyard and home orchards throughout urban and rural areas for domestic consumption, or for making jams or jellies for local trade [14]. The performance of D. longicaudata on medfly larvae infesting the main pest multiplying hosts is highly relevant given the temporal overlapping availability of orange, fig, and peach during the summer and early autumn (December - March) [40]. This scenario enables medfly population growth in the fruit-growing region of Cuyo, mostly in urban areas. Thus, it is essential to apply integrated management strategies against the medfly that minimize the environmental impact involving safety techniques for human health and, at the same time, ensure maximum sustainability. In this context, the most advisable approach is the biological control.
With regard to the second issue, host fruit species had a strong influence on the ability of parasitoids to kill host larvae. Medfly mortality was appreciably higher on fig and peach than on sweet orange, regardless of testing different parasitoid release densities in field cages. Particular physical characteristics of the fruit, such as rind thickness and flesh depth, appear to have been the main influences on the differences in medfly mortality rates found in the trials. A small fruit size, thin peel, and shallow flesh are often linked with a higher level of natural parasitism in a host fruit – frugivorous tephritid larva – parasitoid trophic relationship [21,22,26,41,42].
Based on the above, not surprisingly, the highest levels of medfly mortality due to the D. longicaudata ovipositional activity were found in figs. The fruit of this medfly-host species showed very suitable features to make possible high-level parasitoid effectiveness in killing the pest. In this regard, the thin skin, about 50- and 3-fold thinner than those of orange and peach, respectively, and a flesh 1.6- and 1.3-fold shallower than those of orange and peach, respectively, were apparently the most advantageous physical features for parasitism. Such an assertion is also supported by the host larval density/cm2 of fruit surface area ratio, which was significantly similar among the three host fruit species, suggesting no influence on the parasitoid foraging activity on fruits. A similar host density per cm2 of fruit surface area balanced out the differences in fruit size, mainly characterized by fruit weight, diameter, and external fruit surface area. The physical features of the fig propitious to parasitism may also be associated with the markedly high reproductive success of D. longicaudata females. However, the parasitoid offspring rate found in this host fruit species was not different from that recorded in peach but was highly different from orange. The D. longicaudata female also exhibited a high ability to seek, locate, and kill medfly larvae on peach, although the parasitoid performance was lower than that recorded in fig. Peach is a host fruit species that also has suitable physical properties to facilitate parasitoid ovipositional activity on frugivorous dipteran larvae [43]. The orange was the fruit species with both the lowest medfly mortality and parasitoid offspring rates compared to the other two-tested host fruit species. Lower values of both parameters may be mainly due to the thicker rind and deeper flesh, which further limit parasitism [21]. However, medfly mortality in sweet oranges by D. longicaudata is highly relevant. Citrus sinensis, like other citrus species, provides a bridge for medfly during the cold season, late autumn and winter, when peaches and figs are unavailable in central-western Argentina. Although a previous study showed that D. longicaudata females may be more attracted to peach volatiles than those of orange [44], chemical stimuli are unlikely to have played a role in the medfly-killing parasitoid capacity in the current study. The above is due to D. longicaudata females strongly preferring infested fruit based on chemical cues from the tephritid larvae feeding on rotting fruit substrate, regardless of host fruit species [45,46]. Therefore, any effect of volatiles emanating from a particular fruit species on the attraction of D. longicaudata females was likely homogenized due to the presence of medfly larvae in all fruit exposed to parasitoids.
The third finding showed a similar pattern of increasing both host mortality and parasitoid offspring in the three fruit species when the ratio of released D. longicaudata females to medfly larvae was gradually increased. Thus, the higher host-killing capacity and reproductive success achieved by D. longicaudata occurred at the highest ratios of parasitoid females released into the tube cages, i.e., 80 and 160. In this regard, 160 parasitoids released per experimental tube cage compared to 20 and 40 released parasitoids substantially increased medfly mortality 1.4- to 1.7-fold in peach, 1.5- to 1.6-fold in fig and 1.6- to 1.9-fold in orange, taking into account the two-year study period. When comparing the two maximum parasitoid release densities (80 vs. 160), there was less difference between them than with the two lower parasitoid densities. However, there was a substantial increase of 7-8% in fig, 8-9% in peaches, and 9-14% in oranges towards 160 parasitoids released per tube cage. The last data is relevant because the increase to 160 parasitoid females released in the trials involving orange yielded the maximum effectiveness of D. longicaudata killing medfly larvae infesting this citrus species. Similarly, the number of offspring produced by D. longicaudata increased at higher parasitoid female densities, reaching its maximum value at 160 parasitoids/cage tube density. However, the parasitoid reproductive success in fig and peach was substantially similar and higher than in orange, most likely due to the physical properties that favor parasitism in both fruit species. Such data from field-cage natural conditions, i.e., restricting the parasitoid dispersal and host search area, suggest that medfly control may improve substantially with an increase in parasitoid release density per hectare in an infested fruit crop. Previous augmentative releases of D. longicaudata on fruit farms in irrigated valleys of San Juan used parasitoid release densities averaging between 1600 and 2300 parasitoids/hectare with a sex ratio of 0.8-1.1 females/male [14,15]. Therefore, based on results of this study, the host-killing D. longicaudata capacity at higher open-field parasitoid release densities needs to be tested in future studies.
In brief, the study supports D. longicaudata as a biocontrol agent able to produce high mortality to medfly in all three primary host fruit species, such as fig, peach, and orange, which are key hosts responsible for growing and supporting medfly populations in all the irrigated fruit-growing valleys of central-western Argentina. Particularly, the capacity of D. longicaudata to kill medfly larvae infesting a Citrus species with more challenging physical characteristics for parasitism, like orange, is a potentially valuable trait when using this exotic parasitoid in open-field largescale releases. The results of the current study are critical for designing area-wide medfly management approach involving both small orchards and extensive commercial crops in irrigated semi-arid fruit-producing areas of Argentina.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Host-killing parasitoid capacity by fruit species; Table S2: Host-killing parasitoid capacity by treatment; Table S3: Parasitoid reproductive success by fruit species; Table S4: Parasitoid reproductive success by treatment; Table S5: Parasitoid offspring sex ratio by fruit species; Table S6: Parasitoid offspring sex ratio by treatment; Database_Statistical analysis S7: Overviews of statistical models applied for tested biological parameters_HK_PRS_POSR.
Author Contributions
Conceptualization, L.d.C.S., S.R.N.-C., F.M., F.R.M.G. and S.M.O.; Data curation, L.d.C.S., S.R.N.-C. and S.M.O.; funding acquisition, L.d.C.S., S.R.N.-C., F.M. and S.M.O.; Formal Analysis, S.R.N.-C.; Investigation, L.d.C.S., S.R.N.-C., F.M., F.R.M.G. and S.M.O.; Methodology, L.d.C.S., S.R.N.-C., F.M., F.R.M.G. and S.M.O.; project administration, L.d.C.S., S.R.N.-C., F.M. and S.M.O.; Resources, L.d.C.S., S.R.N.-C., F.M. and S.M.O.; Software, S.R.N.-C.; Supervision, L.d.C.S., S.R.N.-C., F.M., F.R.M.G. and S.M.O.; Validation, L.d.C.S., S.R.N.-C., F.M., F.R.M.G. and S.M.O.; Visualization, L.d.C.S., S.R.N.-C., F.M., F.R.M.G. and S.M.O.; writing—original draft preparation; L.d.C.S., S.R.N.-C., F.M., F.R.M.G. and S.M.O.; writing—review and editing, L.d.C.S., S.R.N.-C., F.M., F.R.M.G. and S.M.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Agencia Nacional de Promoción de la Investigación, el Desarrollo Tecnológico y la Innovación (Argentina). The number of the grant is PICT2020-01050.
Data Availability Statement
The data presented in this study are available in Supplementary Material here (File S7).
Acknowledgments
We thank to the authorities of the Plant, Animal, and Food Health Bureau of the government of the San Juan province for providing us with biological material, study site, and technical staff to perform trials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- De Meyer, M.; Robertson, M.P.; Peterson, A.T.; Mansell, M.W. Ecological niches and potential geographical distributions of Mediterranean fruit fly (Ceratitis capitata) and Natal fruit fly (Ceratitis rosa). J. Biogeo. 2008, 35, 270–281. https://doi.org/ 10.1111/j.1365-2699.2007.01769.x. [CrossRef]
- Malacrida, A.R.; Gomulski, L.M.; Bonizzoni, M.; Bertin, S.; Gasperi, G.; Guglielmino, C.R. Globalization and fruit fly invasion and expansion: the medfly paradigm. Genetica 2007, 131, 1-9. https://doi.org/ 10.1007/s10709-006-9117-2. [CrossRef]
- Liquido, N.J.; Shinoda, L.A.; Cunningham, R.T. 1991. Host plants of the Mediterranean fruit fly (Diptera: Tephritidae): an annotated world review. Misc. Publ. Entomol. Soc. America 1991, 77, 1-52.
- Wernicke, M.; Egartner, A.; Blümel, S.; Moraiti, C. A.; Papadopoulos, N. T. Overwintering potential of the Mediterranean fruit fly (Diptera: Tephritidae) in Austria. J. Econ. Entomol. 2024, toae180, https://doi.org/10.1093/jee/toae180. [CrossRef]
- He, Y.; Xu, Y.; Chen, X. Biology, ecology and management of tephritid fruit flies in China: a review. Insects 2023, 14, 196. ; https://doi.org/10.3390/insects14020196. [CrossRef]
- Vergani, A. R. La mosca del mediterráneo. Publ. Inst. Sanidad Vegetal de Argentina Serie B, 1952, 22, 1-12.
- Villarreal, P.; Mongabure, A.; Borges, C. A.; Gómez-Segade, C. Evaluación del impacto económico del Programa Nacional de Control y Erradicación de Mosca de los Frutos, PROCEM Patagonia. FunBaPa Ediciones: Viedma, Río Negro, Argentina, 2023, p. 88.
- Guillén, D.; Sánchez, R. Expansion of the National Fruit Fly Control Programme in Argentina. In Area-Wide Control of Insects Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 653–660.
- Bouvet, J.P.; Segade, G. Arándanos y Cítricos: Anastrepha fraterculus (Wiedemann) y Ceratitis capitata (Wiedemann). In Plagas Cuarentenarias de frutales de la República Argentina: Avances en los Resultados; Rossini, M., Agostini, J.P., Dummel, D.M., Eds.; Ediciones del Instituto Nacional de Tecnología Agropecuaria, Centro Regional Patagonia Norte, Estación Experimental Agropecuaria Alto Valle: Río Negro, Argentina, 2015; pp. 31–43.
- Ernst, B. Fruticultura Argentina: características e importancia. 2020. Available online: https://www.revistainternos.com.ar/v2/wpcontent/uploads/2021/04/FRUTICULTURA-ARGENTINA-Estudio-Comparativo-2009vs.2019-2-COMPLETO-FDA.pdf (accessed on 2 September 2024).
- SENASA; Servicio Nacional de Sanidad y Calidad Agroalimentaria. Moscas de los Frutos. 2017. Available online: http://www.senasa.gob.ar/cadena-vegetal/frutales/produccion-primaria/programas-fitosanitarios/mosca-de-los-frutos-0 (accessed on 2 September 2024).
- Quiroga, D.; Ramirez, W.; Ruiz, C. National Fruit Fly Control and Eradication Program (PROCEM) Argentina. In Proceedings of the 8th International Symposium on Fruit Flies of Economic Importance, Valencia, Spain, 26 September–1 October 2010; Sabater-Muñóz, B., Navarro-Llopis, V., Urbaneja, A., Eds.; Editorial Universitat Politécnica de Valéncia: Valencia, Spain, 2010; p. 100. Available online: http://hdl.handle.net/10251/11200 (accessed on 2 September 2024).
- SENASA; Servicio Nacional de Sanidad y Calidad Agroalimentaria. Control de 1 Millón de Hectáreas para Prevenir la Mosca de los Frutos. 2018. Available online: http://www.senasa.gob.ar/senasa-comunica/infografias/control-de-1-millon-de-hectareaspara-prevenir-la-mosca-de-los-frutos (accessed on 2 September 2024).
- Suárez, L.; Biancheri, M.J.B.; Murúa, F.; Ordano, M.; Wang, X.; Cancino, J.; Garcia, F.R.M.; Sánchez, G.; Beltrachini, S.; Kulichevsky, L.E.; Ovruski, S.M. Medfly population suppression through augmentative release of an introduced parasitoid in an irrigated multi-fruit orchard of central–western Argentina. Insects 2023a, 14, 387. https://doi.org/10.3390/insects14040387. [CrossRef]
- Sánchez, G.; Murúa, F.; Suárez, L.; Van Nieuwenhove, G.; Taret, G.; Pantano, V.; Ovruski, S.M. Augmentative releases of Diachasmimorpha longicaudata (Hymenoptera: Braconidae) for Ceratitis capitata (Diptera: Tephritidae) control in a fruit-growing region of Argentina. Biol. Control 2016, 103, 101–107. https://doi.org/10.1016/j.biocontrol.2016.08.002. [CrossRef]
- Suárez, L.; Buonocore Biancheri, M.J.; Murúa, F.; Bilbao, M.; García, M.; Cancino, J.; Martín, O.; Molina, D.; Laría, O.; Ovruski, S.M. Effects of host age and radiation dose in Diachasmimorpha longicaudata (Hymenoptera: Braconidae) mass reared on Medfly larvae of the tsl Vienna 8 genetic sexing strain. Biol. Control 2019, 130, 51–59. https://doi.org/10.1016/j.biocontrol.2018.12.013. [CrossRef]
- Saabna, N.; Keasar, T. Parasitoids for biological control in dryland agroecosystems. Curr. Opin. Insect Sci., 2024, 64, 101226. https://doi.org/10.1016/j.cois.2024.101226. [CrossRef]
- Suárez, L.; Murúa, F.; Lara, N.; Escobar, J.; Taret, G.; Rubio, J.L.; Van Nieuwenhove, G.; Bezdjian, L.; Schliserman, P.; Ovruski, S.M. Biological control of Ceratitis capitata (Diptera: Tephritidae) in Argentina: releases of Diachasmimorpha longicaudata (Hymenoptera: Braconidae) in fruit-producing semi-arid areas of San Juan. Nat. Sci. 2014, 6, 664–675. https://doi.org/10.4236/ns.2014.69066. [CrossRef]
- Cancino, J.; Ruiz, L.; López E.; Aguilar, E.; Galvez, C.; Montoya, P.; Liedo, P. Suppression of Ceratitis capitata (Wied.) (Diptera: Tephritidae) populations in coffee in the Mexico-Guatemala border region through the augmentative releases of Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae). Biocontrol Sci. Tech. 2019, 29, 822–826. https://doi.org/10.1080/09583157.2019.1608507. [CrossRef]
- Hoffmeister, T. Factors determining the structure and diversity of parasitoid complexes in tephritids fruit flies. Oecologia 1992, 89, 288-297. https://doi.org/10.1007/BF00317230. [CrossRef]
- López, M.; Aluja, M.; Sivinski, J. Hymenopterous larval-pupal and pupal parasitoids of Anastrepha flies (Diptera: Tephritidae) in Mexico. Biol. Control 1999, 15, 119–129. https://doi.org/10.1006/BCON.1999.0711. [CrossRef]
- Wang, X-G.; Johnson, M.W.; Daane, K.M.; Yokoyama, V.Y. Larger Olive Fruit Size Reduces the Efficiency of Psyttalia concolor as a parasitoid of the Olive fruit fly. Biol. Control 2009, 49, 45-51. https://doi.org/10.1016/j.biocontrol.2009.01.004. [CrossRef]
- Purcell, M.F. Contribution of biological control to integrated pest management of tephritid fruit flies in the tropics and subtropics. Integ. Pest. Manag. Rev. 1998, 3, 63–83. https://doi.org/10.102 3/a:1009647429498. [CrossRef]
- Messing, R.H.; Jang, E.B. (1992) Response of the fruit fly parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae) to host-fruit stimuli. Environ. Entomol. 1992, 21, 1189-1195. https://doi. org/10.1603/0046-225X-34.3.576.
- Montoya, P.; Liedo, P.; Benrey, B.; Barrera, J.F.; Cancino, J.; Sivinski, J.; Aluja, M. Biological control of Anastrepha spp. (Diptera: Tephritidae) in mango orchards through augmentative releases of Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae). Biol. Control 2000, 18, 212–224. https://doi.org/10.1006/bcon.2000.0819. [CrossRef]
- Montoya, P.; López, P.; Cruz Bustos, J. Effect of Diachasmimorpha longicaudata releases on the native parasitoid guild attacking Anastrepha spp. larvae in disturbed zones of Chiapas, Mexico. BioControl 2017, 62, 581-593. https://doi.org/10.1007/s10526-017-9826-8. [CrossRef]
- Garcia, F.R.M.; Ovruski, S.M.; Suárez, L.; Cancino, J.; Liburd, O.E. Biological control of tephritid fruit flies in the Americas and Hawaii: A review of the use of parasitoids and predators. Insects 2020, 11, 662. https://doi.org/10.3390/insects11100662. [CrossRef]
- Montoya, P.; Cancino, J.; Zenil, M.; Santiago, G.; Gutierrez, J.M. The augmentative biological control component in the Mexican national campaign against Anastrepha spp. fruit flies. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 661–670.
- de Pedro, L.; Tormos, J.; Harbi, A.; Ferrara, F.; Sabater-Muñoz, B.; Asís, J.D.; Beitia, F. Combined use of the larvo-pupal parasitoids Diachasmimorpha longicaudata and Aganaspis daci for biological control of the medfly. Ann. Appl. Biol. 2019, 174, 40–50. https://doi.org/10.1111/aab.12468. [CrossRef]
- Sivinski, J. Augmentative biological control: research and methods to help make it work. CAB Rev. 2013, 8, 1–11. https://doi.org/10.1079/PAVSNNR20138026. [CrossRef]
- Dias, N.P.; Montoya, P.; Nava, D. E. A 30-year systematic review reveals success in tephritid fruit fly biological control research. Entomol Exp Appl. 2022, 170, 370–384. https://doi.org/10.1111/eea.13157. [CrossRef]
- Cáceres, C. Mass rearing of temperate sensitive genetic sexing strains in the Mediterranean fruit fly (Ceratitis capitata). Genetica 2002, 116, 107–116. https://doi.org/10.1023/A:1020967810703. [CrossRef]
- Campos, V. E.; Gatica, G.; Andino, N.; Fernández Maldonado, V. N.; Cardús, A. Land surface temperature in an arid city: assessing spatio-temporal changes. 2023. Available online: https://ri.conicet.gov.ar/bitstream/handle/11336/232349/CONICET_Digital_Nro.a5dc7937-042d-43de-9076-72ffe5c2679b_B.pdf?seq (accessed on 2 September 2024).
- García-Medel, D.; Sivinski, J.; Dıáz-Fleischer, F.; Ramirez-Romero, R.; Aluja, M. Foraging behavior by six fruit fly parasitoids (Hymenoptera: Braconidae) released as single- or multiple-species cohorts in field cages: influence of fruit location and host density. Biol. Control 2007, 43, 12–22. https://doi.org/10.1016/j.biocontrol.2007.06.008. [CrossRef]
- Harbi, A.; Beitia, F.; Ferrara, F.; Chermiti, B.; Sabater-Muñoz, B. Functional response of Diachasmimorpha longicaudata (Ashmead) over Ceratitis capitata (Wiedemann): influence of temperature, fruit location and host density. Crop. Prot. 2018, 109, 115–122. https://doi.org/10.1016/j.cropro.2018.03.010. [CrossRef]
- Rosenheim, J.A.; Hoy, M. A. Confidence intervals for the Abbott’s formula correction of bioassay data for control response. J. Econ. Entomol. 1989, 82, 331-335. https://doi.org/10.1093/jee/82.2.331. [CrossRef]
- Pinheiro, J.C.; Bates, D. M. nlme: Linear and Nonlinear Mixed Effects Models; R package version 3.1-166, 2024. Available online: https://CRAN.R-project.org/package=nlme (accessed on 10 September 2024).
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer, New York, USA, 2000. https://doi.org/10.1007/b98882. [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing, Vienna, Austria, 2024. Available online: https://www.R-project.org/ (accessed on 10 September 2024).
- Suárez, L.; Biancheri, M.J.B.; Díaz-Nieto, L.M.; Schliserman, P.; Murúa, F.; Rull, J.; Lasa, R.; Pantano, V.; Ovruski, S.M. Dynamic seasonal response of Ceratitis capitata (Diptera: Tephritidae) to fruit juice-based lures in fig orchards. Int. J. Pest Manag. 2023b, 64, 346-358. https://doi.org/10.1080/09670874.2021.1918360. [CrossRef]
- Sivinski, J.; Aluja, M. The evolution of ovipositor length in the Parasitic Hymenoptera and the search for predictability in biological control. Fla. Entomol. 2003, 86, 143–150. https://doi.org/10.1653/0015-4040(2003)086[0143:TEOOLI]2.0.CO;2. [CrossRef]
- Sivinski, J.; Aluja, M. The roles of parasitoid foraging for hosts, food and mates in the augmentative control of Tephritidae. Insects 2012, 3, 668–691. https://doi.org/10.3390/insects3030668. [CrossRef]
- Buonocore-Biancheri, M.J.; Wang, X.; Núñez-Campero, S.R.; Suárez, L.; Schliserman, P.; Ponssa, M.D.; Kirschbaum, D.S.; Garcia, F.R.M.; Ovruski, S.M. The population dynamics and parasitism rates of Ceratitis capitata, Anastrepha fraterculus, and Drosophila suzukii in non-crop hosts: Implications for the management of pest Fruit flies. Insects 2024, 15, 61. https://doi.org/10.3390/ insects15010061. [CrossRef]
- Segura, D.F.; Nussenbaum, A.L.; Viscarret, M.M.; Devescovi, F.; Bachmann, G.E.; Corley, J.C.; Ovruski, S.M.; Cladera, J.L. Innate host habitat preference in the parasitoid Diachasmimorpha longicaudata: functional significance and modifications through learning. PLoS One 2016, 11, e0152222. https://doi.org/10.1371/journal. pone.0152222. [CrossRef]
- Eben, A.; Benrey, B.; Sivinski, J.; Aluja, M. Host species and host plant effects on preference and performance of Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Environ. Entomol. 2000, 29, 87–94. https://doi.org/10.1603/0046-225X-29.1.87. [CrossRef]
- Stuhl, C.; Sivinski, J.; Teal, P.; Aluja, M. Responses of multiple species of tephritid (Diptera) fruit fly parasitoids (Hymenoptera: Braconidae: Opiinae) to sympatric and exotic fruit volatiles. Fla. Entomol. 2012, 95, 1031–1039. https://doi.org/10.1653/024.095.0432. [CrossRef]
Figure 1.
Location of the three major field cages and their five internal tube cages, at the experimental trial yard from the Plant, Animal and Food Health Bureau, government of the San Juan province, Rivadavia district, San Juan province, Argentina. Specifications: large colorless circle = main experimental field cage; small blue, red, and green circles: inner tube cages; small black circles: poplar tree windbreak. Treatments: T1 = Infested peaches; T2 = Infested oranges fruit; T3 = Infested figs. Sub-treatments: TA = 20 released female parasitoids (RFP), TB = 40 RFP; TC = 80 RFP, TD = 160 RFP. Control tests: C1, control test from T1; C2, control from T2; C3, control from T3.
Figure 1.
Location of the three major field cages and their five internal tube cages, at the experimental trial yard from the Plant, Animal and Food Health Bureau, government of the San Juan province, Rivadavia district, San Juan province, Argentina. Specifications: large colorless circle = main experimental field cage; small blue, red, and green circles: inner tube cages; small black circles: poplar tree windbreak. Treatments: T1 = Infested peaches; T2 = Infested oranges fruit; T3 = Infested figs. Sub-treatments: TA = 20 released female parasitoids (RFP), TB = 40 RFP; TC = 80 RFP, TD = 160 RFP. Control tests: C1, control test from T1; C2, control from T2; C3, control from T3.
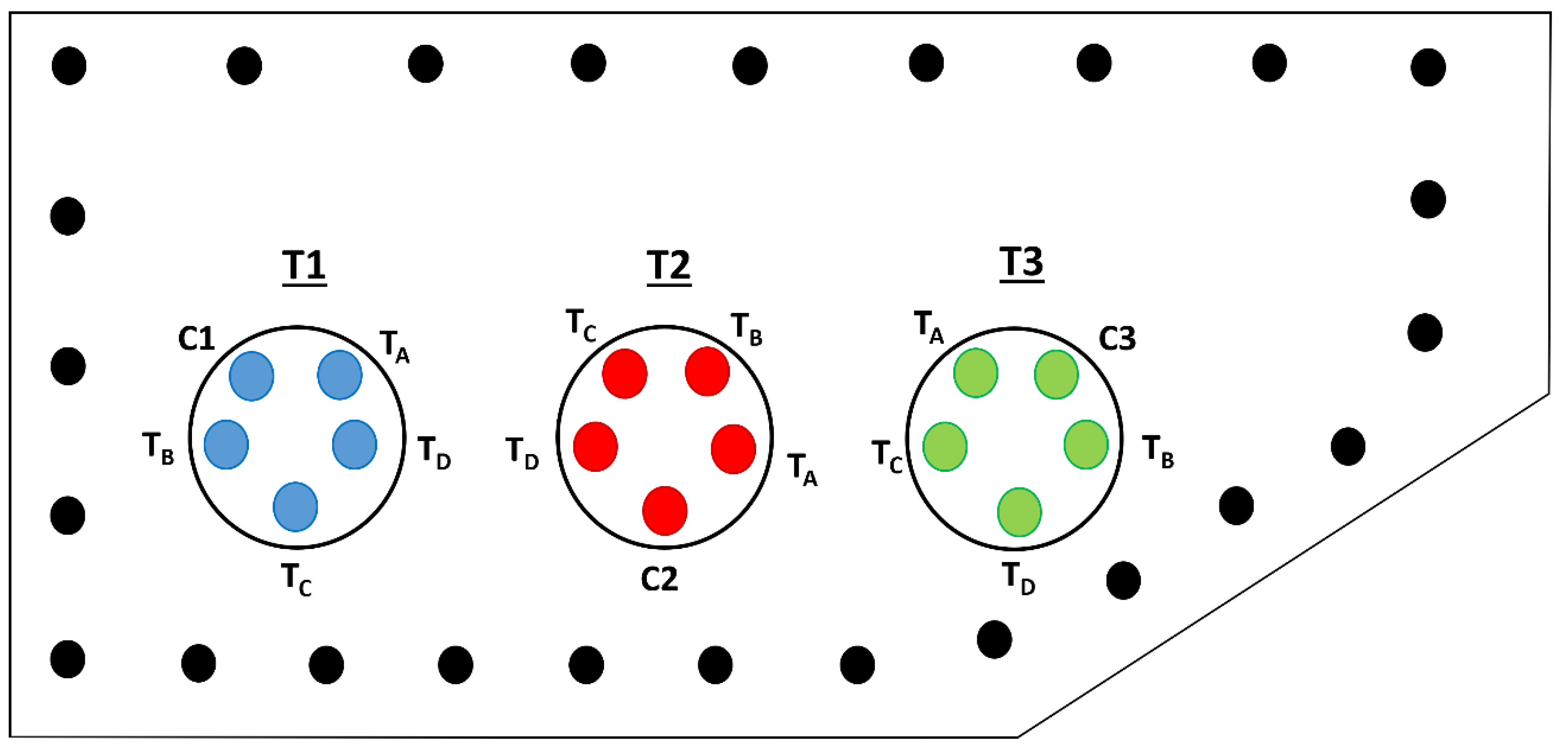
Figure 2.
Percentage of the medfly-killing ability of Diachasmimorpha longicaudata on three fruit host species (fig, sweet orange, and peach) at four parasitoid release densities (20, 40, 80, and 160 females) under field cage conditions in Tulum fruit-growing valley, San Juan, central-western Argentina.
Figure 2.
Percentage of the medfly-killing ability of Diachasmimorpha longicaudata on three fruit host species (fig, sweet orange, and peach) at four parasitoid release densities (20, 40, 80, and 160 females) under field cage conditions in Tulum fruit-growing valley, San Juan, central-western Argentina.
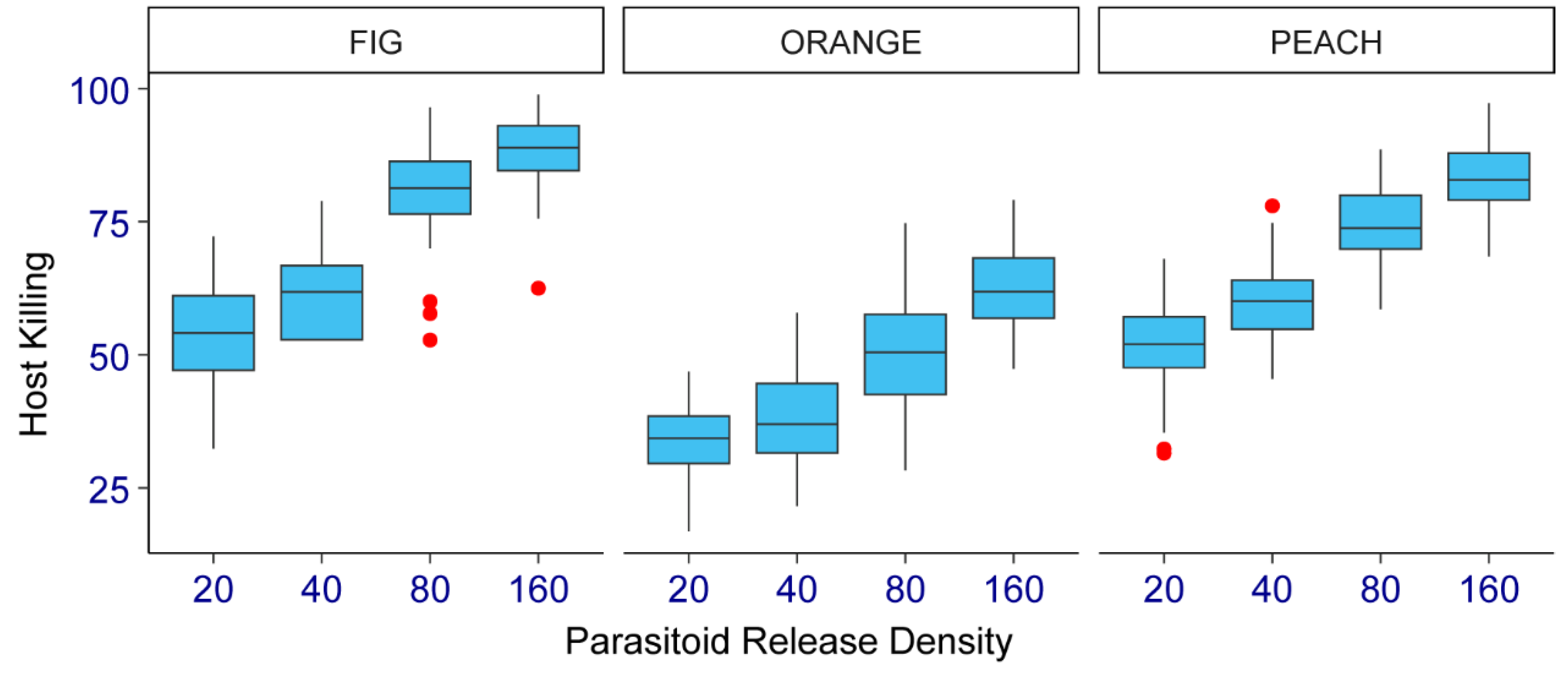
Figure 3.
Percentage of offspring produced by Diachasmimorpha longicaudata (parasitoid reproductive success) when parasitizing medfly larvae infesting three fruit host species (fig, sweet orange, and peach) at four parasitoid release densities (20, 40, 80, and 160 females) in studying years 2019 and 2020 under field cage conditions in Tulum fruit-growing valley, San Juan, central-western Argentina.
Figure 3.
Percentage of offspring produced by Diachasmimorpha longicaudata (parasitoid reproductive success) when parasitizing medfly larvae infesting three fruit host species (fig, sweet orange, and peach) at four parasitoid release densities (20, 40, 80, and 160 females) in studying years 2019 and 2020 under field cage conditions in Tulum fruit-growing valley, San Juan, central-western Argentina.
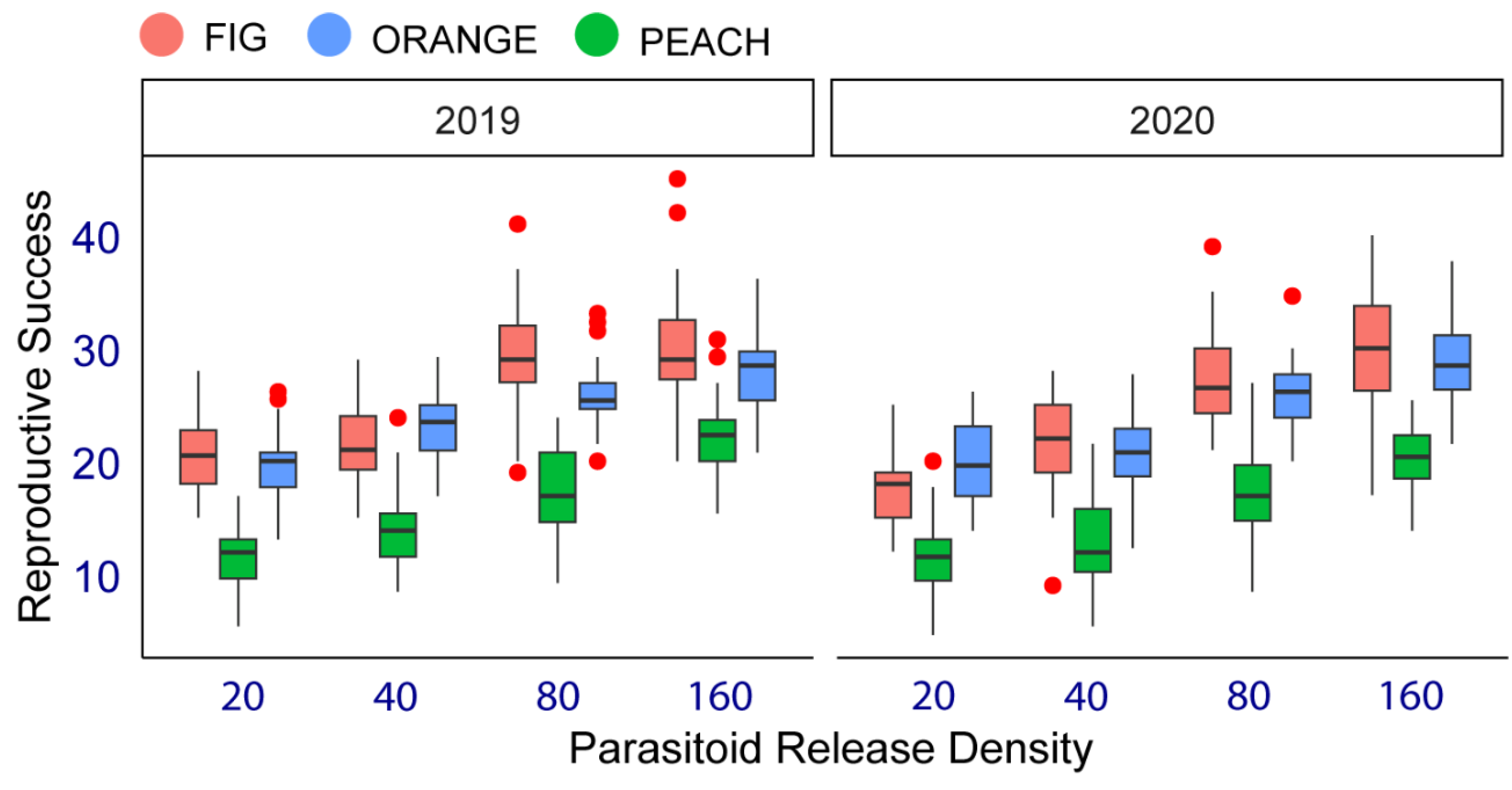
Figure 4.
Percentage of emerged Diachasmimorpha longicaudata females from medfly puparia recovered from three fruit host species (fig, sweet orange, and peach) at four parasitoid release densities (20, 40, 80, and 160 females) under field cage conditions in Tulum fruit-growing valley, San Juan, central-western Argentina.
Figure 4.
Percentage of emerged Diachasmimorpha longicaudata females from medfly puparia recovered from three fruit host species (fig, sweet orange, and peach) at four parasitoid release densities (20, 40, 80, and 160 females) under field cage conditions in Tulum fruit-growing valley, San Juan, central-western Argentina.
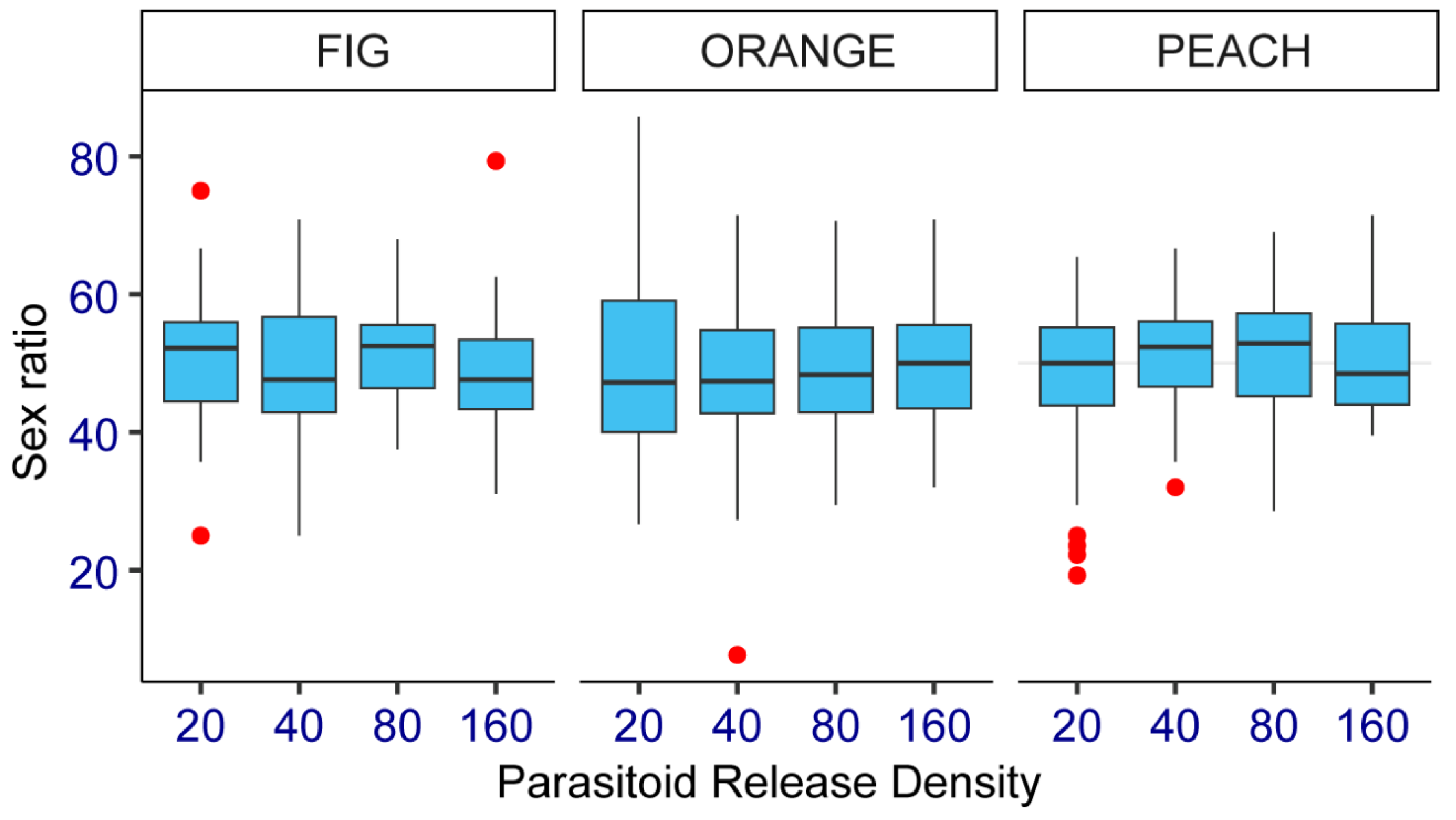

Table 1.
Minimum (= Min), maximum (=Max) and average temperature and relative humidity, and accumulated rainfall recorded during trial dates in March 2019 and 2020 (late summer/early autumn).
Table 1.
Minimum (= Min), maximum (=Max) and average temperature and relative humidity, and accumulated rainfall recorded during trial dates in March 2019 and 2020 (late summer/early autumn).
| Study year | Temperature (°C) |
Relative Humidity (%) |
Precipitation (mm) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Min. | Max. | Average monthly | Min. | Max. | Average monthly | Accumulated rainfall | |||
| 2019 | 9.9 | 36.3 | 21.1 | 10 | 86 | 56.2 | 0.2 | ||
| 2020 | 10.9 | 36.5 | 22.8 | 23 | 89 | 57.1 | 0.3 | ||
Table 2.
Physical features of medfly-host fruit species, Citrus sinensis (sweet orange), Prunus persica (peach), and Ficus carica (fig), and density of Ceratitis capitata larvae per cm2 of fruit surface area.
Table 2.
Physical features of medfly-host fruit species, Citrus sinensis (sweet orange), Prunus persica (peach), and Ficus carica (fig), and density of Ceratitis capitata larvae per cm2 of fruit surface area.
| Host fruits | Fruit physical features (n = 30) (mean ± SE) |
Medfly larvae | ||||||
|---|---|---|---|---|---|---|---|---|
| Weight (g) |
Diameter (cm) |
Rind thickness (cm) | Pulp depth (cm) | Surface area (cm2) |
Density * (cm2) (mean ± SE) |
Larvae per fruit | ||
| Sweet orange | ||||||||
| 200.2 ± 4.8a | 7.1 ± 0.2a | 4.6 ± 0.2a | 3.1 ± 0.1a | 44.0 ± 8.0a | 3.08 ± 0.06a | 130 | ||
| Peach | 156.9 ± 1.5b | 6.6 ± 0.1b | 0.3 ± 0.1b | 2.5 ± 0.1b | 41.9 ± 0.4b | 3.10 ± 0.03a | 130 | |
| Fig | 57.2 ± 0.8c | 5.1 ± 0.2c | 0.1 ± 0.1c | 2.0 ± 0.1c | 32.3 ± 1.1c | 3.11 ± 0.01a | 100 | |
*Inoculated host larvae ratio per cm2 of fruit surface area (density). Means followed by different letters within the same column are significantly different (Kruskal-Wallis rank sum test, p = 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



