
Submitted:
20 September 2024
Posted:
23 September 2024
You are already at the latest version
Abstract
Cirsium japonicum, a traditional herb, exhibits significant anti-inflammatory activity; however, the main components and potential mechanisms of C. japonicum remain unclear. The aim of this study is to investigate the anti-inflammatory mechanism of Cirsium japonicum through network pharmacology and cellular experiments. The effective components and potential targets for anti-inflammatory activity of C. japonicum were identified using traditional Chinese medicine systematic pharmacology database, TCMSP analysis platform, and GeneCards database. The drug-component-target-disease network diagram was constructed using Cytoscape3.8.0 software, while the protein interaction network diagram was created using STRING database and Cytoscape3.8.0 software. Gene ontology (GO) enrichment and KEGG pathway enrichment analysis were carried out using DAVID database. Molecular docking between key targets and active components was constructed with AutoDock software to determine the best binding target. Results revealed that 14 active components of C. japonicum targeted 171 anti-inflammatory proteins. GO function enrichment analysis yielded 173 items, while KEGG pathway enrichment analysis identified 48 signaling pathways related to inflammation regulation. Molecular docking showed that strong affinity between sitosterol, stigmasterol, and other components with key targets such as peroxisome proliferator-activated receptor α recombinant protein (PPARA) and cyclooxygenase-2 (PTGS2). Vanillin, one active ingredient of C. japonicum, inhibited the release of lipopolysaccharide (LPS)-induced inflammatory factors in RAW264.7 cells. These findings suggest that C. japonicum may exert its anti-inflammatory effects by modulating the phosphatidylinositol 3-kinase (PI3K)/ protein kinase B (Akt) signal pathway (PI3K-Akt) and apoptin signal pathway, highlighting the multi-component, multi-target, and multi-channel molecular mechanism underlying its anti-inflammatory properties. Finally, the anti-inflammatory effect of vanillin, an effective component in C. japonicum, was verified by cell experiments. This study provides a new understanding of the pharmacological mechanisms of C. japonicum in treatment of inflammatory conditions.
Keywords:
Cirsii japonicum
; network pharmacology
; anti-inflammatory activity
; molecular docking
; cellular experiment
1. Introduction
The traditional herb Cirsium japonicum (Radix Cirsii Japonici) has been widely used for the treatment of hemorrhagic fever, hepatitis, coagulation disorders, and urinary tract disorders [1,2,3,4,5]. Several studies have demonstrated that the aqueous extract of C. japonicum exhibits effective therapeutic potential against infectious jaundice and chronic hepatitis [6]. Additionally, it displays inhibitory activity against Mycobacterium tuberculosis, Neisseria meningitidis, Mycobacterium diphtheriae, Staphylococcus aureus, Mycobacterium typhi, Mycobacterium paratyphi, and Mycobacterium anthracis [7]. Furthermore, it has been found to ameliorate metabolic disorders associated with steatohepatic conditions induced by high-fat diets [8]. Aqueous, alkaline, and acidic alcoholic extracts of C. japonicum, as well as aqueous extracts of leaves, demonstrate antihypertensive effects. Clinical reports have also documented the use of root tablets for hypertension treatment. Additionally, C. japonicum has been reported to enhance immunity, promote lipid metabolism, exhibit diuretic and hepatoprotective properties, enhance ethanol metabolizing enzyme activity, and reduce lipid peroxidation [9]. However, the main components and potential mechanisms of C. japonicum remain unclear.
With the rapid development of systems biology, bioinformatics, and pharmacology, network-based drug discovery has emerged as a promising method for developing effective drugs. In 2007, Hopkins et al. introduced the concept of "network pharmacology", which utilizes systematic biology to analyze drug intervention and potential therapeutic targets for diseases [10,11,12,13,14,15]. Network pharmacology emphasizes a shift from the traditional "one target, one drug" strategy to a novel "network target, multi-component" strategy [16]. In the field of traditional Chinese medicine research, it is widely employed due to its holistic and systematic nature that aligns with the principles of traditional Chinese medicine prescriptions [17,18,19]. Molecular docking is also extensively utilized in material basis research on traditional Chinese medicine as computer-aided drug design method that relies on interactions and affinities between targets and active compounds [20,21,22,23,24,25].
Network pharmacology employs network methods to analyze the intricate interplay among drugs, diseases, and targets, while also investigating the synergistic effects of multiple components on diseases. This approach closely aligns with the theoretical framework of holistic concepts and diagnosis and treatment in Chinese medicine, serving as a contemporary scientific methodology for elucidating the material basis of traditional Chinese medicines’ efficacy and mechanisms of action. It facilitates visualization analysis of compound-target-signaling pathways through multidisciplinary approaches such as systems biology, bioinformatics, and multi-omics linkage. To some extent, this analysis reveals the therapeutic effectiveness of each active ingredient in natural plants [26,27,28,29]. In recent years, network pharmacology has been widely used to predict the mechanism of action between natural active ingredients and diseases.
The present study employed network pharmacology and molecular docking methods to investigate the underlying network mechanism of the anti-inflammatory effects of active ingredients in C. japonicum, elucidating their synergistic actions through multiple targets and pathways. In addition, the anti-inflammatory effect of vanillin, a potent component of C. japonicum, was also assessed in mouse macrophages.
2. Materials and Methods
2.1. Software and Network Databases
TCMSP (https://tcmsp-e.com), UniProt (https://www.uniprot.org)database, Cytoscape3.8.0, software, GeneCards (https://www.genecards.org) database, STRING (https://string-db.org) database, DAVID (https://david.ncifcrf.gov) database, OmicShare (https://www.omicshare.com) online tool, and AutoDockTools-1.5.6 software were used in this study.
2.2. Prediction and Identification of Active Ingredients and Potential Targets of C. Japonicum
The active ingredient screening of C. japonicum was performed using TCMSP [30] (https://tcmsp-e.com), a comprehensive pharmacology database and analytical platform for traditional Chinese medicine. The selection criteria included an oral bioavailability (OB) threshold of ≥20% and drug-like properties (DL) threshold of ≥0.1, based on the parameter information and standards provided by TCMSP. Subsequently, the target names were converted into corresponding Gene symbols using the UniProt database (https://www.uniprot.org). Cytoscape 3.8.0 software was employed to construct a network diagram illustrating the interactions between active ingredients and targets in C. japonicum.
2.3. Construction of Networks and Pathway Analysis
The GeneCards database (https://www.genecards.org) was used to perform a search for relevant antimicrobial targets, while the keyword "Anti-inflammation" was employed to explore disease genes of significance. The drug-component-target-disease network diagram was constructed by intersecting the retrieved anti-inflammatory gene target with the active ingredient gene target of C. japonicum.
By importing the target genes corresponding to the active ingredients of C. japonicum and the target genes of inflammation into the Draw Venn Diagram website, a Venn diagram was generated to identify common targets for disease treatments.
The anti-inflammatory targets of C. japonicum were imported into the STRING database (https://string-db.org) to construct protein interaction networks. Subsequently, the obtained data were visualized and analyzed using Cytoscape 3.8.0 software.
The potential anti-inflammatory targets of C. japonicum were retrieved and inputted into the DAVID database (https://david.ncifcrf.gov) for Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) signaling pathway analysis. The identifiers were selected as "OFFICIAL-GENE-SYMBOL" and the species was specified as "Homo sapiens". The selection of “Home sapiens" aimed to predict the functional distribution of the targets. GO analysis included Biological Process (BP), Cellular Component (CC), and Molecular Function (MF), with the top ten items chosen for each category to facilitate visualization. Additionally, KEGG signaling pathway analysis was visualized using OmicShare online tool (https://www.omicshare.com).
2.4. Molecular Docking
The structural formulas of the active ingredients of C. japonicum were downloaded from TCMSP. The top five active ingredients with the highest degree values and the top five core targets were selected. Subsequently, their corresponding structures were acquired by querying the target names on the Uniprot website. Additionally, PDB structural formulas of these targets were obtained from the RCSB PDB website. The active ingredients underwent energy minimized using Chem3D 19.0 and were exported as ligand files in pdb format. On Discovery Studio 2019, receptor proteins underwent dehydrogenation and deligation processed before being hydrogenated again and exported as receptor files in MOL2 format.
The AutoDockTools-1.5.6 software was utilized to conduct molecular docking analysis between the active ingredient and the core target, aiming to determine the binding energy and validate the therapeutic potential of C. japonicum’s key active ingredient in inflammation treatment [31]. A lower binding energy indicates a more stable binding conformation and a higher probability of ligand-receptor interaction.
2.5. Verification of Anti-inflammatory Activity of Active Ingredients of C. japonicum in RAW264.7 Cells
After overnight culture in a 6-well plate (1 × 105 cells/well, 2 mL medium/well), the cells were pre-treated with vanillin, one of active ingredients of C. japonicum, for 3, 6, 9, and 12 h, respectively. This was followed by lipopolysaccharide (LPS) treatment for an additional 24 h. At the end of scheduled experiments, the culture supernatant from each well was collected and the inflammatory cytokines, including tumor necrosis factor α (TNF-α), interleukin 6 (IL-6), interleukin 1β (IL-1β), and interleukin 10 (IL-10), respectively. They were measured by using commercial assay kits (Rpworld (Beijing) Co., Ltd, Beijing, China) according to the manufacturer’s instructions. The absorbance at a wavelength of 520 nm was then measured using a microplate reader.
3. Results
3.1. The Identification of Active Ingredients in C. japonicum and the Prediction of Their Molecular Targets
The total of 18 active ingredients were obtained from the TCMSP database with an OB of ≥ 20% (Table 1). Among them, 14 active ingredients were found to have corresponding targets, resulting in the identification of 171 targets. However, rhodopsin, β-amyl acetate, cyclopropane, and pectin did not show any corresponding targets. To convert these identified targets into gene names specific to Homo Sapiens, a search was conducted using "Homo Sapiens" as the keyword in the UniProt database. After removing duplicate genes from the results obtained, a total of 85 unique genes remained. The interaction between active ingredients and their respective targets in C. japonicum sapiens were visualized using Cytoscape 3.8.0 software (Figure 1). To identify target genes associated with inflammation, searches were performed using "Inflammation" as a keyword in both OMIM and GeneCards databases. This filtering process resulted in the identification of a total of 2,924 disease-related targets.
3.2. Screening of Potential Anti-inflammatory Targets of C. japonicum
A search in the GeneCards database yielded 1,809 identified inflammatory targets. Through an intersection analysis between antibacterial and active ingredient targets of C. japonicum, visualized using a Venn diagram generated by the Draw Venn Diagram (ugent.be) online tool, we identified a total of 32 potential anti-inflammatory targets (Figure S1). Subsequently, it was imported into Cytoscape3.8.0 software to draw the Drug - Ingredient – Target - Disease network diagram (Figure 2).
3.3. Construction of Protein-Protein Interaction (PPI) Network and Results of Core Gene Screening
The 32 identified anti-inflammatory genes were inputted into the STRING online database, resulting in the acquisition of a PPI network map consisting of 29 anti-inflammatory genes after excluding unrelated genes (Figure S2). While Network analyzer function analysis was employed to identify potential key targets associated with C. japonicum’s anti-inflammatory properties, such as PPARG, PTGS2, BDNF, GCG, and PPARA. This identification was made by evaluating the Closeness, Betweenness, and Degree values.
3.4. GO and KEGG Enrichment Function Analysis
A total of 32 common targets of active ingredients of C. japonicum and inflammation were subjected to GO enrichment analysis using the David database. This analysis revealed 20 cellular components, including the extracellular region, extracellular space, and extracellular exosome. Furthermore, this analysis identified 23 molecular processes that were significantly enriched, primarily involving heme binding, peroxidase activity, protein binding, and prostaglandin binding. Exosome-related processes were also observed. Moreover, a total of 130 biological processes were discovered through this analysis encompassing the regulation of blood pressure, inflammatory response, prostaglandin biosynthesis, nitric oxide synthesis, and nitric oxide-mediated signal transduction (Figure 3). KEGG enrichment analysis further revealed the involvement of various pathways including the relaxin signaling pathway, AGE-RAGE signaling pathway in diabetic complications, pathways associated with neurodegeneration - multiple diseases, HIF-1 signaling pathway, etc. (Figure 4).
3.5. Molecular Docking Validation
The four active ingredients (including oleic acid, methyl linoleate, leguminol, and sterol) were subjected to molecular docking with six key targets (such as PPARG, PTGS2, BDNF, GCG, PPARA, and EDN1), and the binding energies after docking are presented in Table S1. Lower AutoDock docking scores indicate a higher ability of the molecules to bind to the targets compared to the target itself. A lower AutoDock score signifies stronger binding ability between the molecule and the target as well as reduced energy requirement for binding. It is evident that certain active components of C. japonicum (such as sterol and stigmasterol) exhibit robust affinity towards several key targets, including PPARA, PPARG, PTGS2, and EDN1. Molecular docking results demonstrate that sitosterol binds to the active sites of PPARA and PTGS2 proteins while forming hydrogen bond interactions with ASN-299, THR-60 and LYS-546, respectively (Figure 5 -a, b). Vanillin binds to the active sites of BDNF and PPARA proteins, while forming hydrogen bond interactions with ARG-88, TY-52, ARG-87, ARG-409, and ASP-419, respectively (Figure 5-c, d). Stigmasterol binds specifically to the active site of PPARA protein while forming a hydrogen bond interaction with GLY-296 (Figure 5 -e).
3.6. Effect of Vanillin on Inflammatory Factors in RAW264.7 Cells Challenged with LPS
The levels of inflammatory factors, such as TNF-α, IL-6, IL-1β, and IL-10, were assessed using competitive enzyme-linked immunosorbent assay (ELISA). In comparison to the blank control group, the levels of TNF-α, IL-6, and IL-1β in the model group were significantly increased (P < 0.005), while IL-10 showed a significant decrease (P < 0.01) (Figure 6). These findings indicate a substantial alteration in the levels of inflammatory cytokines within the cells due to LPS-induced inflammation. However, upon addition of vanillin, there was a notable reduction in TNF-α, IL-1β and IL-6 levels within the cells (P < 0.005) (Figure 6 -a,b,c). Notably, 0.2 mg/mL of vanillin demonstrated the most effective inhibition on IL-1β (P < 0.0001), while 0.05 mg/mL of vanillin displayed optimal inhibitory effects on both IL-6 and TNF-α; furthermore, 0.2 mg/mL of vanillin significantly increased the concentration of IL-10 (Figure 6 -d).
4. Discussion
C. japonicum, a widely accessible and cost-effective Chinese herbal medicine, possesses various functions such as bacteriostasis, anti-inflammation, blood coagulation and hemostasis, blood pressure reduction, and anti-tumor effects [32]. However, the precise mechanism underlying its anti-inflammatory action remains unclear. This study employs network pharmacology and molecular docking to investigate the anti-inflammatory effects of C. japonicum, followed by in vitro experimental verification at the cellular level.
Flavonoids, widely distributed in Cirsium, is also the most abundant component and the main active ingredient of Cirsium. It possesses various biological activities such as anti-oxidation, anti-tumor, anti-inflammation, and liver, cardiovascular, and cerebrovascular protection [33,34,35,36]. In this study, 14 effective components of C. japonicum were obtained from the TCMSP website, primarily including methyl linoleate, oleic acid, stigmasterol, and sitosterol. Previous studies have demonstrated that stigmasterol exhibits potent anti-cancer properties, while also showing effects against osteoarthritis and inflammation. Moreover, it displays potent activity against parasites, fungi, and bacteria, while also exhibiting immunomodulatory and neuroprotective effects through its antioxidant properties [37,38,39,40,41]. One study conducted by Feng et al. [42] has revealed that stigmasterol significantly inhibits colon shortening and reduces colitis severity by suppressing pro-inflammatory IL-1β, IL-6, and cyclooxygenase-2 (COX-2) monocyte chemotactic protein release. Additionally, stigmasterol improves intestinal function and regulates fat metabolism to alleviate hepatic steatosis in rats by fortifying the intestinal barrier and enhancing bile acid metabolism [43]. β-sitosterol, a phytosterol with anti-inflammatory properties, exerts regulatory effects on blood glucose metabolism [44]. Xiao et al. [45] demonstrated that β-sitosterol reduces serum TNF-α levels in rats, thereby decelerating the progression of gastric mucosa damage through decreased release and aggregation of inflammatory factors within the gastric mucosa. Moreover, the combination of β-sitosterol and aspirin can enhance the anti-inflammatory efficacy of aspirin [46].
Based on network pharmacology, we conducted a systemic analysis on the active components, targets, related pathways, and biological processes of C. japonicum. Through analyzing the relevant database of network pharmacology, a total of 14 active components and their corresponding 171 gene targets were identified for C. japonicum. The drug-component-target-disease network diagram was constructed to reveal that 14 effective active components in C. japonicum can synergistically act on 32 anti-inflammatory targets (Figure 2). The 14 anti-inflammatory components primarily consist of methyl linoleate, oleic acid, β-starch acetate, and sterols (such as stigmasterol sitosterol). Among them, oleic acid can inhibit the LPS-induced inflammatory reaction by down-regulating the expression of the nuclear factor kappa-B (NF-κB) signaling pathway [47]. Methyl linoleate effectively inhibits the expression of IL-1βin THP-1 cells.
In the PPI network visualization analysis, ADRB2, PTGS2, NOS3, BDNF, and PPARG have been identified as potential core targets for the anti-inflammatory effects of C. japonicum (Figure S1). Among them, ADRB2 is a crucial β2-adrenergic receptor involved in maintaining hepatocellular carcinoma cell proliferation and survival. Additionally, it has been found to attenuate osteoarthritis-like defects in temporomandibular joints when conditionally detected in mice [48,49]. PTGS2, also known as COX-2 enzyme, plays a significant role in the inflammatory response by catalyzing arachidonic acid conversion to prostate H2, which triggers the inflammatory cascade [50]. The expression of PTGS2 is regulated by various stress-related factors and serves as an important regulator; up-regulated PTGS2 significantly contributes to inflammation regulation through glucagon production [51]. Nitric oxide (NO) plays a crucial role in regulating various aspects of vascular function, including smooth muscle cell proliferation and migration, vascular tone, endothelial permeability, and endothelial-leukocyte interactions. It serves as a key anti-atherogenic factor in the endothelium [52]. Endothelial-type nitric oxide synthase 3 (NOS3), encoded by a gene located on chromosome 7q35-36, is responsible for maintaining vascular homeostasis and regulating endothelial function. NOS3 genetic polymorphisms have been demonstrated to exert an impact on NO levels, lipid profiles, and are associated with hypertension [53] as well as diabetic foot ulcers [54]. BDNF represents a crucial class of neurotrophic factors that play an essential role in regulating neuronal proliferation, differentiation, maturation, and pro-neuronal regeneration; it constitutes a fundamental factor in ongoing depression research [55]. BDNF and its receptor, tyrosine kinase receptor B (TrκB), have been implicated in the pathogenesis of various neurological disorders [56]. Furthermore, activation of the BDNF/TrκB signaling pathway has shown potential for ameliorating memory deficits in rats with Alzheimer's disease [57]. PPARG belongs to the nuclear transcription factor superfamily as a subtype of peroxisome proliferator-activated receptor and has been demonstrated to mitigate inflammatory responses by inhibiting the NF-κB signaling pathway, making it a promising therapeutic molecular target for diverse malignant tumors [58].
In the GO and KEGG pathway analyses of 32 targets related to anti-inflammatory effects of C. japonicum, it was revealed that the biological processes primarily involved in C. japonicum include response to LPS, negative regulation of cellular regulatory processes, response to hypoxia, negative regulation of macrophage-derived foam cell differentiation, and response to activity (Figure 3). The anti-inflammatory activity of C. japonicum mainly involves signaling pathway processes, such as relaxin signaling pathway, arginine biosynthesis pathway, prostate cancer pathway, AGE-RAGE pathway in diabetic complications, apocynin signaling pathway, and PI3K-Akt signaling pathway (Figure 4). Among these pathways, the PI3K/Akt signaling pathway is a crucial intracellular mechanism that responds to extracellular signals and regulates various cellular and molecular functions, including metabolism, survival, growth, and angiogenesis. Its involvement in gastritis has also gained significant attention in recent years due to its role in cell growth, proliferation, apoptosis, as well as blood glucose regulation [59]. Several studies have demonstrated that cytokines such as TNF-α and IL-6 can attenuate inflammatory responses by modulating the PI3K-Akt signaling pathway [60]. Molecular docking results further confirmed that the ability of active ingredients from C. japonicum, including sitosterol, vanillin, stigmasterol, etc., to bind key targets (such as BNDF, PPARA, PPARA, etc.) and form hydrogen bonding interactions (Figure 5). Notably, vanillin exhibited a higher binding affinity than oleic acid with binding energies ranging from -5.54 to -3.35 kal (Table S1). Moreover, vanillin effectively suppressed the expression of pro-inflammatory factors (such as IL-6 and IL-1β) in mouse macrophages while promoting the expression of anti-inflammatory factor IL-10 (Figure 6). These findings suggest that C. japonicum possesses significant preventive and therapeutic potential against LPS-induced inflammation in mouse macrophages.
5. Conclusions
The network pharmacology analysis showed that 14 active ingredients of C. japonicum targeted 171 anti-inflammatory proteins, including ADRB2, PTGS2, NOS3, BDNF, and PPARG. Moreover, it was found that C. japonicum has the potential to modulate the PI3K-Akt signaling and apoptogens signaling pathways in inflammation regulation. Notably, one of the active ingredients of C. japonicum exhibited remarkable anti-inflammatory activity in macrophages. This study provides a theoretical and scientific basis for further understanding the anti-inflammatory mechanism of C. japonicum as well as its potential development and application.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Venn diagram of intersecting targets between anti-inflammatory genes and active ingredients of C. japonicum.; Table S1: Binding energies of key anti-inflammatory protein targets docked to active ingredients of C. japonicum; Figure S2: Protein-protein interaction (PPI) network analysis.
Author Contributions
Conceptualization, J.X.W. and X.M.W.; Methodology, H.T., Z.L.W., Y.Z., and B.H.; Software, W.A.; Writing—original draft preparation, J.X.W.; Writing—review and editing, J.X.W. and X.Z.S.; Visualization, J.X.W.; Supervision, X.M.W. and J.Q.W.; Project administration, J.X.W.; Funding acquisition, X.M.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work has received funding support from the program of the Third Xinjiang Scientific Expedition Program (Grant No. 2022xjkk0404).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are contained within the manuscript and Supplementary Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wu, L.C.; Li, J.F.; Zhang, T.T.; Tao, F.F.; Liu, W.H. Discusses the Anti-inflammatory Effect of Stigmasterol Based on Network Pharmacology and Cell Experiments. Chin. Patent Med. 2022, 44, 7. [Google Scholar] [CrossRef]
- Park, J.Y.; Kim, H.Y.; Shibamoto, T.; Jang, T.S.; Lee, S.C.; Shim, J.S.; Hahm, D.H.; Lee, H.J.; Lee, S.; Kang, K.S. Beneficial Effects of a Medicinal Herb, Cirsium Japonicum var. Maackii, Extract and its Major Component, Cirsimaritin on Breast Cancer Metastasis in MDA-MB-231 Breast Cancer Cells. Bioorg. Med. Chem. Lett. 2017, 17, 3968–3973. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.L.; Shao, H.; Chen, Y.; Ding, N.; Yang, A.N.; Tian, J.; Jiang, Y.X.; Li, G.Z.; Jiang, Y.D. In Renal Hypertension, Cirsium Japonicum Strengthens Cardiac Function via the Intermedin/Nitric Oxide Pathway. Biomed. Pharmacother. 2018, 101, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Wagle, A.; Seong, S.H.; Shrestha, S.; J, H.A.; Choi, J.S. Korean Thistle (Cirsium Japonicum Var. Maackii (Maxim.) Matsum.): a Potential Dietary Supplement Against Diabetes and Alzheimer's Disease. Molecules 2019, 24, 649. [Google Scholar] [CrossRef]
- Yoon, S.; Kim, M.; Shin, S.; Woo, J.; Son, D.; Ryu, D.; Yoo, J.; Park, D.; Jung, E. Effect of Cirsium Japonicum Flower Extract on Skin Aging Induced by Glycation. Molecules 2022, 27, 2093. [Google Scholar] [CrossRef]
- Cho, C.; Kang, L.J.; Jang, D.; Jeon, J.; Lee, H.; Choi, S.; Han, S.J.; Oh, E.; Nam, J.; Kim, C.S.; Park, E.; Jeong, S.Y.; Park, C.H.; Shin, Y.S.; Eyun, S.I.; Yang, S. Cirsium Japonicum Var. Maackii and Apigenin Block Hif-2α-induced Osteoarthritic Cartilage Destruction. J. Cell. Mol. Med. 2019, 8, 5369–5379. [Google Scholar] [CrossRef]
- Meng, L.; Liu, R.; Sun, A.; Wu, S.J.; Liu, N.N. Separation and Purification of Rutin and Acaciin from the Chinese Medicinal Herb Herba Cirsii by Combination of Macroporous Absorption Resin and High-speed Counter-current Chromatography. J. Chromatogr. Sci. 2009, 47, 329–332. [Google Scholar] [CrossRef]
- Ye, L.; Yang, F.Q.; Liang, J.; Zhao, S.H.; Gao, Y.L. In Vitro Antibacterial Activity of Different Polar Parts of Cirsium Japonicum Extract from Ningxia Against Four Candida Species. Chin. J. Exper. Trad. Med. Form. 2011, 17, 222–223. [Google Scholar] [CrossRef]
- Кравченкo, О.В. Artichoke Extracts: Physiological Effects, Use in Obstetric Practice. Reprod. Endocrinol 2018, 40, 80–86. [Google Scholar] [CrossRef]
- Che, D.N.; Shin, J.Y.; Kang, H.J.; Cho, B.O.; Park, J.H.; Wang, F.; Hao, S.P.; Sim, J.S.; Sim, D.J.; Jang, S.I. Ameliorative Effects of Cirsium Japonicum Extract and Main Component Cirsimaritin in Mice Model of Hgh-fat Diet-induced Metabolic Dysfunction-associated Fatty Liver Disease. Food Sci. Nutr. 2021, 11, 6060–6068. [Google Scholar] [CrossRef]
- Li, T.; Zhang, W.; Hu, E.; Sun, Z.; Li, P.; Yu, Z.; Zhu, X.; Zheng, F.; Xing, Z.; Xia, Z.; He, F.; Luo, J.; Tang, T.; Wang, Y. Integrated Metabolomics and Network Pharmacology to Reveal the Mechanisms of Hydroxysafflor Yellow A Against Acute Traumatic Brain Injury Comput. Struct. Biotechnol. J 2021, 19, 1002–1013. [Google Scholar] [CrossRef]
- Shang, L.R.; Wang, Y.C.; Li, J.X.; Zhou, F.Y.; Xiao, K.M.; Liu, Y.H.; Zhang, M.Q.; Wang, S.H.; Yang, S.L. Mechanism of Sijunzi Decoction in the Treatment of Colorectal Cancer Based on Network Pharmacology and Experimental Validation. J. Ethnopharmacol. 2022, 302, 115876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, D.F.; Zhou, W.A.; Wang, L.; Wang, B.Y.; Zhang, T.Y.; Li, S. Network Pharmacology: Towards the Artificial Intelligence-based Precision Traditional Chinese Medicine. Brief Bioinform. 2023, 25. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, H.; Li, N.; Chen, J.M.; Xu, H.; Wang, Y.J.; Liang, Q.Q. Network Pharmacology, A Promising Approach to Reveal the Pharmacology Mechanism of Chinese Medicine Formula. J. Ethnopharmacol. 2023, 116306. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Z.Q.; Liao, J.; Chen, Q.; Lu, X.Y.; Fan, X.H. Network Pharmacology Approaches for Research of Traditional Chinese Medicines. Chin. J Nat. Medicines. 2023, 21, 323–332. [Google Scholar] [CrossRef]
- Zhong, Y.; Luo, J.; Tang, T.; Li, P.; Liu, T.; Cui, H.; Wang, Y.; Huang, Z. Exploring Pharmacological Mechanisms of Xuefu Zhuyu Decoction in the Treatment of Traumatic Brain Injury via a Network Pharmacology Approach. Evid.-Based Compl. Alt. 2019, 2018, 7518374. [Google Scholar] [CrossRef]
- Ma, C.; Xu, T.; Sun, X.; Zhang, S.; Liu, S.; Fan, S.; Lei, C.; Tang, F.; Zhai, C.; Li, C.; Luo, J.; Wang, Q.; Wei, W.; Wang, X.; Cheng, F. Network Pharmacology and Bioinformatics Approach Reveals the Therapeutic Mechanism of Action of Baicalein in Hepatocellular Carcinoma. Evid.-Based Compl. Alt. 2019, 2019, 1–15. [Google Scholar] [CrossRef]
- Ma, Q.Y.; Liu, Y.C.; Zhang, Q.; Yi, W.D.; Sun, Y.; Gao, X.D.; Zhao, X.; Wang, H.W.; Lei, K.; Luo, W.J. Integrating Network Pharmacology, Molecular Docking and Experimental Verification to Reveal the Mechanism of Artesunate in Inhibiting Choroidal Melanoma. Front. Pharmacol. 2024, 151448381–1448381. [Google Scholar] [CrossRef]
- Y, J.H.; Li, L.; Hu, Z.X. Exploring the Molecular Mechanism of Action of Yinchen Wuling Powder for the Treatment of Hyperlipidemia, Using Network Pharmacology, Molecular Docking, and Molecular Dynamics Simulation. BioMed. Res. Int. 2021, 9965906. [Google Scholar] [CrossRef]
- Yuan, Z.Z.; Pan, Y.Y.; Leng, T.; Chu, Y.; Zhang, H.J.; Ma, J.R; Ma, X.J. Progress and Prospects of Research Ideas and Methods in the Network Pharmacology of Traditional Chinese Medicine. J. Pharm. Pharm. Sci. 2022, 25218–25226. [Google Scholar] [CrossRef]
- Aminu, K.S.; Uzairu, A.; Abechi, S.E.; Shallangwa, G.A.; Umar, A.B. Activity Prediction, Structure-based Drug Design, Molecular Docking, and Pharmacokinetic Studies of 1,4-dihydropyridines Derivatives as α-amylase inhibitors. J. Talbah Univ. Med. Sci. 2024, 19, 270–286. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.K.; Tao, B.; Xue, X.; Feng, C.X.; Ren, Y.T.; Ma, H.Y.; Zhang, J.L.; Si, Y.F.; Zhang, S.S.; Liu, S.; Li, H.; Zhou, J.H.; Li, G.; Wang, Z.F.; X, J.P.; Zhu, Z.L. Molecular Mechanism of Epicedium Treatment for Depression Based on Network Pharmacology and Molecular Docking Technology. BMC Complement Med. 2021, 21, 222. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.L.; Cui, Y.P. Study on the Mechanism of Action of RoucongrongTang in Treating Functional Constipation based on Network Pharmacology and Molecular Docking. J. Contemporary Med. 2024, 6, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Larisa, I.; Mati, K. The Impact of Software Used and the Type of Target Protein on Molecular Docking Accuracy. Molecules 2022, 27, 9041. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Narang, B.K.; Gupta, M.K.; Abbot, V.; Singh, V.; Rawal, R.K. Molecular Docking Studies on Isocytosine Analogues as Xanthine Oxidase Inhibitors. Drug. Res. 2018, 68, 395–402. [Google Scholar] [CrossRef]
- Zhu, W.T.; Fan, X.M.; Wei, H.; Wang, S.M. Mechanism Research of Apatinib-treated Breast Cancer Based on Network Pharmacology. Chin. Pharm. J. 2016, 51, 1860–1865. [Google Scholar] [CrossRef]
- Liu, Y.; Ju, Y.; Qin, X. Studies on the Compatibility Mechanism and Material Basis of Danggui Buxue Decoction Against Anemia Mice Using Metabonomics and Network Pharmacology. J. Pharm. Pharmacol. 2021, 73, 767–777. [Google Scholar] [CrossRef]
- Gong, P.Y.; Guo, Y.J.; Li, X.M.; Wang, N.; Gu, J. Study on the Potential Pharmacodynamic Substances of Jinhua Qinggan Granule for Prevention and Treatment of Novel Coronavirus Based on Network Pharmacology and Molecular Docking Technology. Chin. Herb Med. 2020, 51, 1685–1693. [Google Scholar] [CrossRef]
- Ding, H.X.; Zhao, Y.; Feng, J. Research progress of plant extracts in the field of feed additives based on network pharmacology. J. Anim. Nutr. 2021, 33, 3065–3071. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, Y.; Jiang, J.; Cui, X.; Cheng, S.; Liu, L.; Huang, Z.; Liao, R.; Zhao, P.; Yu, J.; Wang, J.; Jia, Y.; Jin, W.; Zhou, F. Stigmasterol Attenuates Hepatic Steatosis in Rats by Strengthening the Intestinal Barrier and Improving Bile Acid Metabolism. Npj. Sci. Food. 2022, 27, 38. [Google Scholar] [CrossRef]
- Liu, D.; Chen, Y.F.; Liu, T.J.; Tang, T.; Yu, S.J.; Liu, G.M.; Tang, Y.B. Z.Q. Network Pharmacology and Experimental Study of Jiawei Guizhi Fuling Pill in the Treatment of Benign Prostatic Hyperplasia. Res. Dev. Natur. Products. 2022, 34, 1234–1249. [Google Scholar] [CrossRef]
- Gao, M.M.; Chen, Y.L.; Hao, Y.K.; Guo, J. Metabonomics Study of Cirsium Japonicum Extract in Improving Hypercholesterolemia Model Mice. Chin. Ph. 2023, 34, 1590–1595. [Google Scholar] [CrossRef]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as Anti-inflammatory Agents. Proc. Nutr. Soc. 2010, 69, 27–28. [Google Scholar] [CrossRef] [PubMed]
- Mehrabani, M.; Sargazi, M.L.; Amirkhosravi, A.; Farhadi, S.; Vasei, S.; Raeiszadeh, M.; Mehrabani, M. The Influence of Harvest Time on Total Phenolic and Flavonoid Contents, Antioxidant, Antibacterial and Cytotoxicity of Rheum Khorasanicum Root Extract. Ann. Pharm. Fr. 2023, 81, 475–483. [Google Scholar] [CrossRef]
- Shi, M.Y.; Chen, Z.X.; Gong, H.; Peng, Z.L.; Sun, Q.; Luo, K.P.; Wu, B.Y.; Wen, C.B.; Lin, W. Luteolin, a Flavone Ingredient: Anticancer Mechanisms, Combined Medication Strategy, Pharmacokinetics, Clinical Trials, and Pharmaceutical Researches. Phytother. Res. 2024, 38, 880–911. [Google Scholar] [CrossRef]
- Naik, K.K.; Thangavel, S.; Alam, A.; Kumar, S. Flavone Analogues as Antimicrobial Agents. Recent Patents Infla. 2017, 11, 53–63. [Google Scholar] [CrossRef]
- Chen, Y.P.; Xie, T.; Zhang, H.; Zhou, Y.M. Physiological Function of β -sitosterol and Its Application in Animal Production. J. Anim. Physiol. An. N. 2022, 34, 11. [Google Scholar] [CrossRef]
- Liu, Y.X.; Wang, Z.W.; Yao, P.S.; Li, X.y.; Han, R.M.; Zhang, D.Q.; Zhao, Z.J.; Wang, Y.P.; Zhang, J.P. Antioxidation Activity Enhancement by Intramolecular Hydrogen Bond and Non-Browning Mechanism of Active Ingredients in Rosemary: Carnosic Acid and Carnosol. J. Phys. Chem. C. 2024, 128, 7627–7638. [Google Scholar] [CrossRef]
- Hou, J.Z.; Xiong, W.; Shao, X.Y.; Long, L.; Chang, Y.; Chen, G.H.; Wang, L.; Wang, Z.C.; Huang, Y.Z. Liposomal Resveratrol Alleviates Platelet Storage Lesion via Antioxidation and the Physical Buffering Effect. ACS Appl. Mater. 2023, 15, 45658–45667. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.C.; Gu, D.Y.; Liu, C.; Wang, Y.; Tang, S.S.; Yin, Y.X.; Tian, J. Fermentation of Robinia Pseudoacacia Flower for Improving the Antioxidation: Optimized Conditions, Active Composition, Mechanism, and Biotransformation Process. PREP Biochem Biotech. 2023, 53, 11–13. [Google Scholar] [CrossRef]
- Li, Z.J.; Xue, Y.B.; Li, M.X.; Guo, Q.B.; Sang, Y.X.; Wang, C.L.; Luo, C. The Antioxidation of Different Fractions of Dill (Anethum graveolens) and Their Influences on Cytokines in Macrophages RAW264.7. J. Oleo Sci. 2018, 67, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.B.; Liu, X.L.; Cheng, R.Q.; Jia, H.X.; Wang, X.Y.; Kong, L.N.; Li, Y.J.; Cui, T.T.; Qu, H.D.; Feng, C. Effect of β-sitosterol on The Side Effects and Pharmacological Effects of Aspirin-induced Gastric Mucosal Injury. Chin. J. Ex. Trad. Med. Formulae. 2016, 22, 148–152. [Google Scholar] [CrossRef]
- Long, B.y. Effect of Oleic Acid on Lipopolysaccharide-induced Inflammatory Response of Mouse Macrophages and Its Mechanism. D. Univ. South Chin. 2019. [Google Scholar] [CrossRef]
- Sun, J.L.; Yan, J.F.; Li, J.; Wang, W.R.; Yu, S.B.; Zhang, H.Y.; Huang, F.; Niu, L.N.; Jiao, K. Conditional Deletion of Adrb2 in Mesenchymal Stem Cells Attenuates Osteoarthritis-like Defects in Temporomandibular Joint. Bone 2020, 33, 115229. [Google Scholar] [CrossRef] [PubMed]
- Kunzmann, A.T.; Murray, L.J.; Cardwell, C.R.; McShane, C.M.; McMenamin, U.C.; Cantwell, M.M. PTGS2 (Cyclooxygenase-2) Expression and Survival among Colorectal Cancer Patients: A Systematic Review. Cancer Epidemiol Biomarkers Prev. 2013, 22, 1490–1497. [Google Scholar] [CrossRef]
- Mousavi, S.E.; Saberi, P.; Ghasemkhani, N.; Fakhraei, N.; Mokhtari, R.; Dehpour, A.R. Licofelone Attenuates LPS-induced Depressive-like Behavior in Mice: A Possible Role for Nitric Oxide. Pharm. Sci. 2018, 21, 184–194. [Google Scholar] [CrossRef]
- Hellmann, J.; Tang, Y.; Zhang, M.J.; Hai, T.; Bhatnagar, A.; Srivastava, S.; Spite, M. Atf3 Negatively Regulates Ptgs2/Cox2 Expression During Acute Inflammation. Prostag Oth. Lipid. M. 2015, 116–117, 49–56. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and Inflammation. Ioscler Thromb Vasc. Biol. 2011, 5, 986–1000. [Google Scholar] [CrossRef]
- Neto, A.B.L.; Vasconcelos, N.B.R.; Santos, T.R.D.; Duarte, L.E.C.; Assunção, M.L.; Sales-Marques, C.; Silva Ferreira, H. Prevalence of IGFBP3, NOS3 and TCF7L2 Polymorphisms and Their Association with Hypertension: A Population-based Study with Brazilian Women of African Descent. BMC Res. Notes 2021, 17, 186. [Google Scholar] [CrossRef]
- Sadati, S.M.; Radfar, M.; Kakavand Hamidi, A.; Abdollahi, M.; Qorbani, M.; Nasli Esfahani, E.; Amoli, M.M. Association Between the Polymorphism of Glu298Asp in Exon 7 of the eNOS Gene with Foot Ulcer and Oxidative Stress in Adult Patients with Type 2 Diabetes. Can. J. Diabetes 2018, 42, 18–22. [Google Scholar] [CrossRef]
- Björkholm, C.; Monteggia, L.M. BDNF - A Key Transducer of Antidepressant Effects. Neuropharmacology 2016, 102, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Ming, Y.; Zhou, X.N.; Liu, G.; Abudupataer, M.; Zhu, S.C.; Xiang, B.T.; Yin, X.J.; Lai, H.; Sun, Y.X.; Wang, C.S.; Li, J.; Zhu, K. PM2.5 Exposure Exacerbates Mice Thoracic Aortic Aneurysm and Dissection by Inducing Smooth Muscle Cell Apoptosis via the MAPK Pathway. Chemosphere. 2023, 313, 137500. [Google Scholar] [CrossRef] [PubMed]
- Jin, W. Regulation of BDNF-TrkB Signaling and Potential Therapeutic Strategies for Parkinson's Disease. Clin. Med. Insights-Pe. 2020, 17, 257. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Zhu, X.Y.; Si, J.R. Angelica Polysaccharide Ameliorates Memory Impairment in Alzheimer's Disease Rat Through Activating BDNF/TrkB/CREB Pathway. Exp. Biol. Med (Maywood) 2020, 245, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nava-Villalba, M.; Nuñez-Anita, R.E.; Bontempo, A.; Aceves, C. Activation of Peroxisome Proliferator-activated Receptor Gamma is Crucial for Antitumoral Effects of 6-iodolactone. Mol. Cancer Res. 2015, 7, 168. [Google Scholar] [CrossRef]
- Wang, B.; Song, Z.J.; Qiu, L.; Ling, J.P. Experimental Study on Yiqi Huayu Zhiwei Recipe Relieving Chronic Atrophic Gastritis by Regulating PI3K/AKT Signaling Pathway. J. Zhejiang Uni. Trad. Chin. Med. 2023, 47, 1123–1128. [Google Scholar] [CrossRef]
- Sun, J.P.; Shi, L.; Wang, F.; Qin, J.; Ke, B. Modified Linggui Zhugan Decoction Ameliorates Glycolipid Metabolism and Inflammation via PI3K-Akt/mTOR-S6K1/AMPK-PGC-1 α Signaling Pathways in Obese Type 2 Diabetic Rats. Chin. J. Integr. Med. 2022, 28, 52–59. [Google Scholar] [CrossRef]
- Dobrovolskaia, M.A.; Vogel, S.N. Toll Receptors, CD14, and Macrophage Activation and Deactivation by LPS. Microbes Infect. 2002, 4, 903–914. [Google Scholar] [CrossRef]
- Maldonado, R.F.; Sá-Correia, L.; Valvano, M.A. Lipopolysaccharide Modification in Gram-negative Bacteria During Chronic Infection. FEMS Microbiol. Rev. 2016, 40, 480–493. [Google Scholar] [CrossRef]
- Olivry, T.; Foster, A.P.; Mueller, R.S.; McEwan, N.A.; Chesney, C.; Williams, H.C. Interventions for Atopic Dermatitis in Dogs: A Systematic Review of Randomized Controlled Trials. Vet. Dermatol. 2010, 21, 4–22. [Google Scholar] [CrossRef]
Figure 1.
Active ingredient - target network diagram.
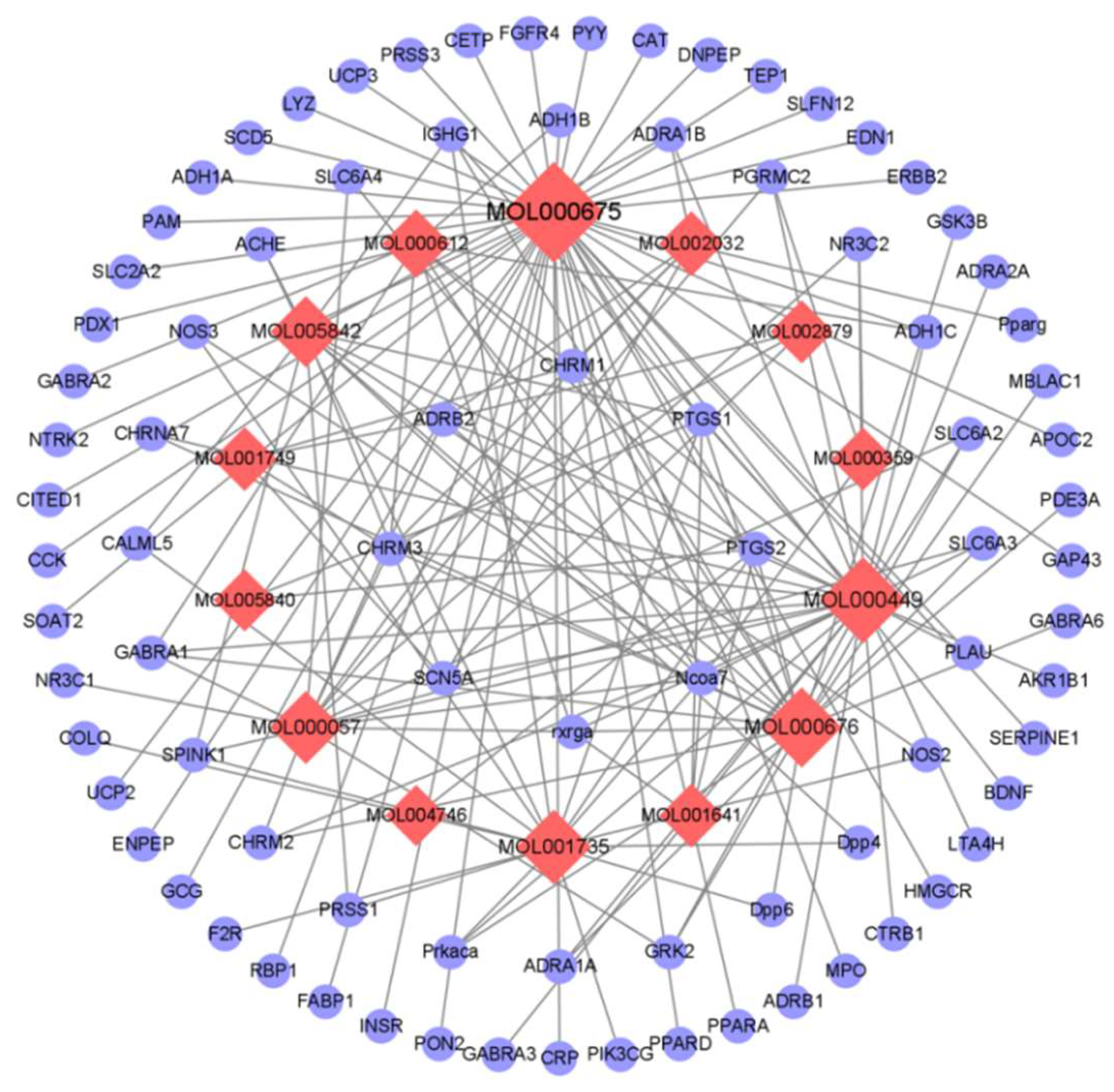
Figure 2.
Drug - Ingredient - Target - Disease Network Diagram.
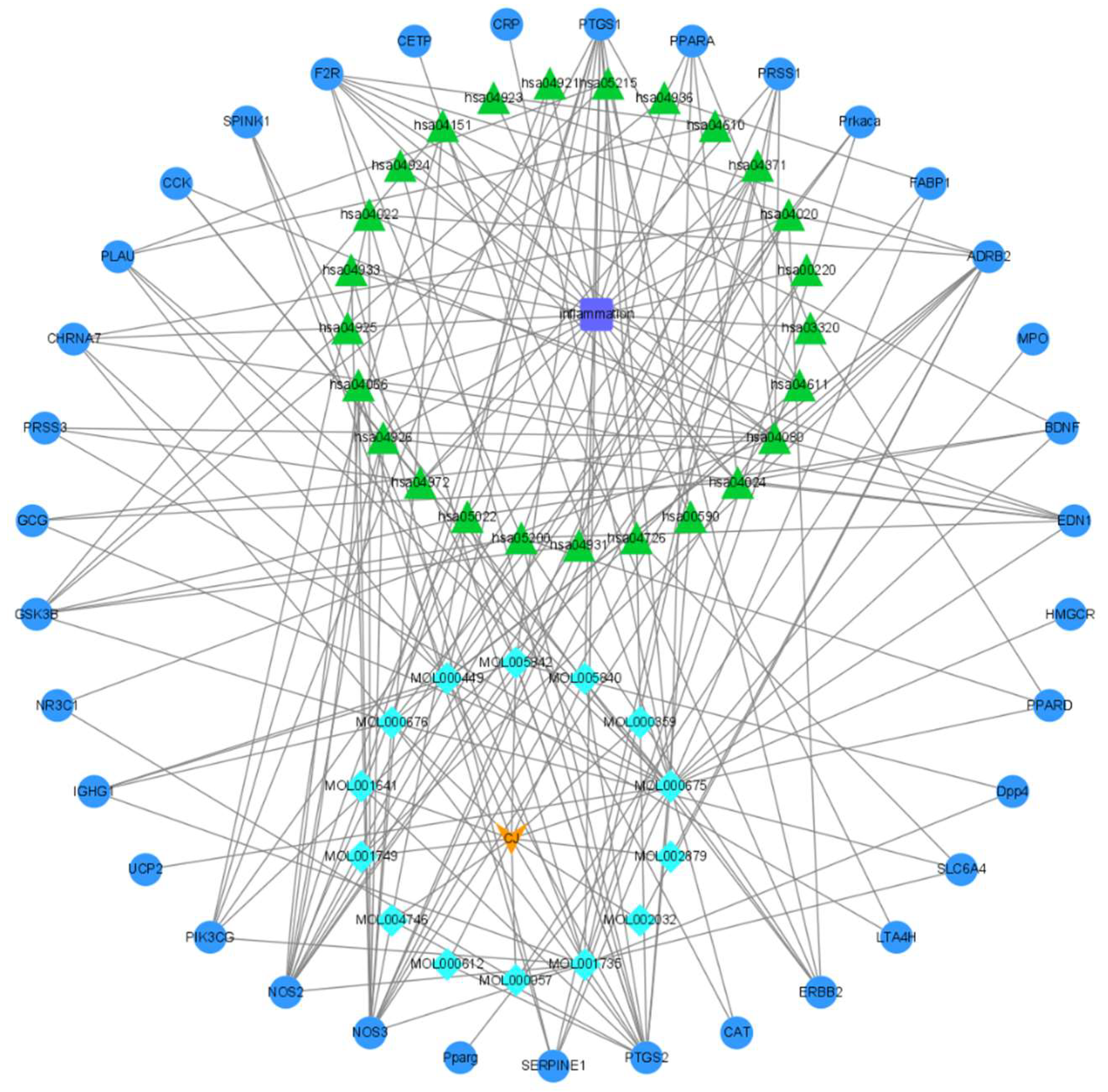
Figure 3.
GO enrichment analysis.
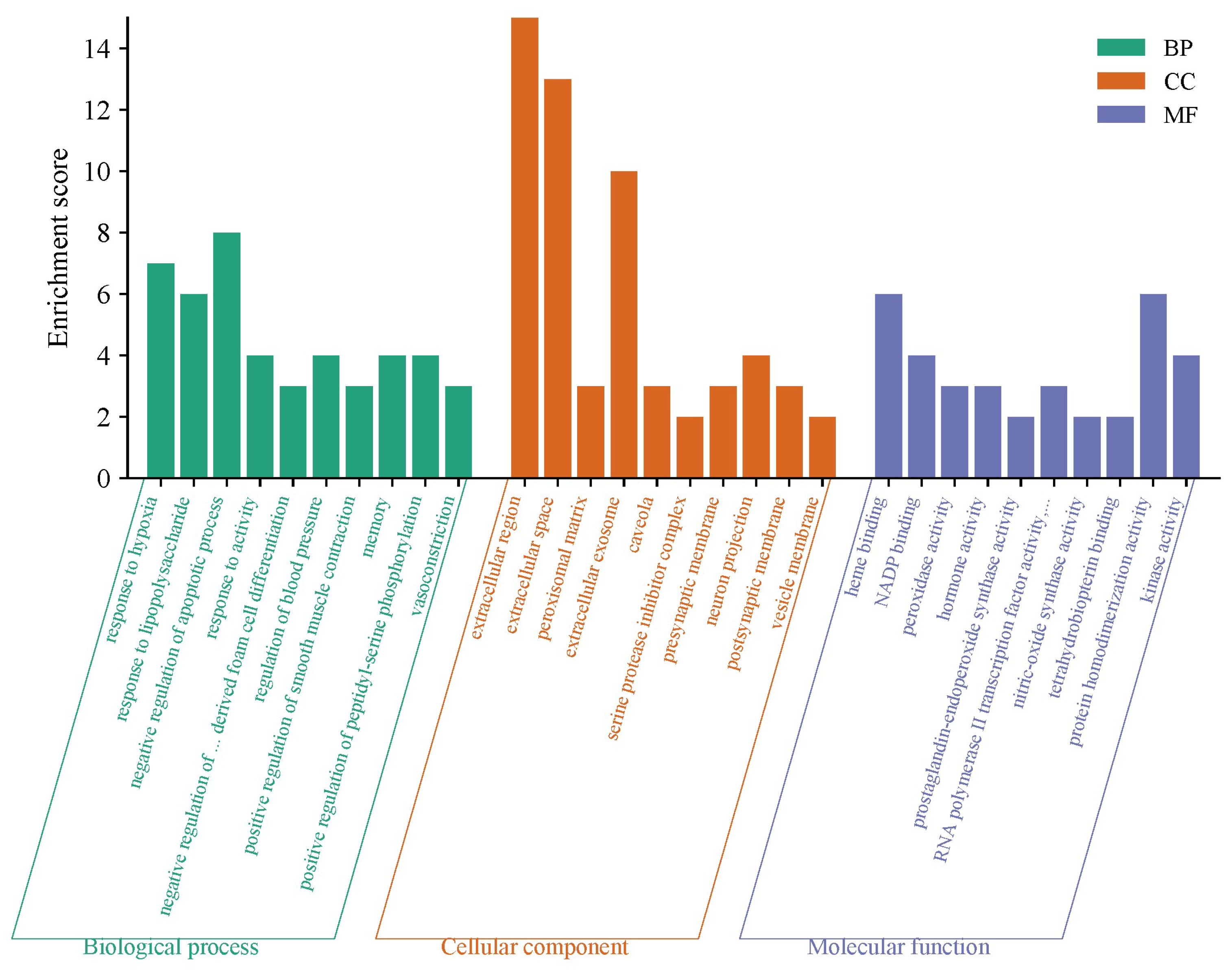
Figure 4.
KEGG enrichment analysis and key pathway network.
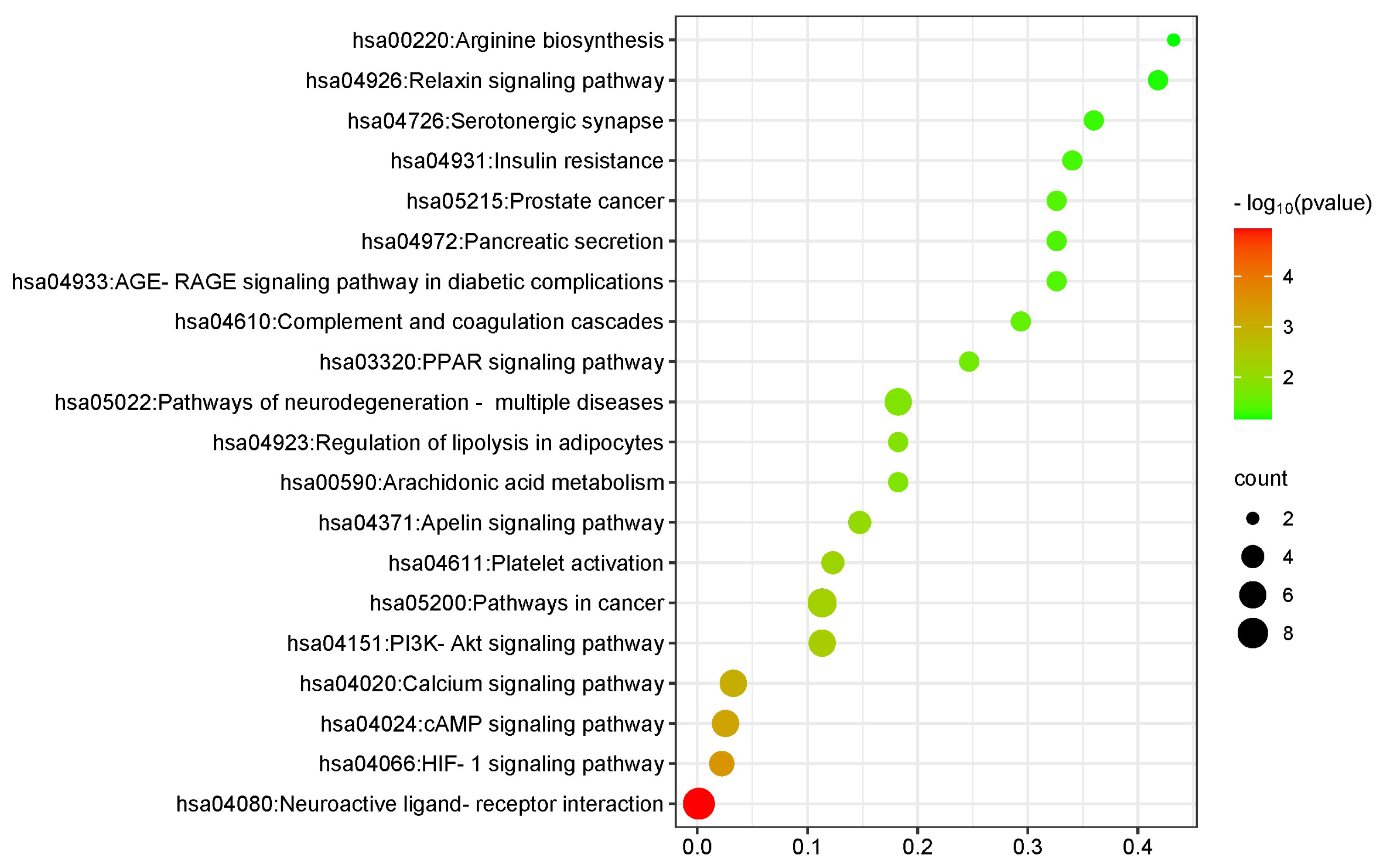
Figure 5.
Molecular docking patterns of key targets and specific active components of C. japonicum. (a) Sitosterol and PPARA; (b) Sitosterol and PTGS2; (c) Vanillin and BDNF; (d) Vanillin and PPARA; (e) Stigmasterol and PPARA.
Figure 5.
Molecular docking patterns of key targets and specific active components of C. japonicum. (a) Sitosterol and PPARA; (b) Sitosterol and PTGS2; (c) Vanillin and BDNF; (d) Vanillin and PPARA; (e) Stigmasterol and PPARA.
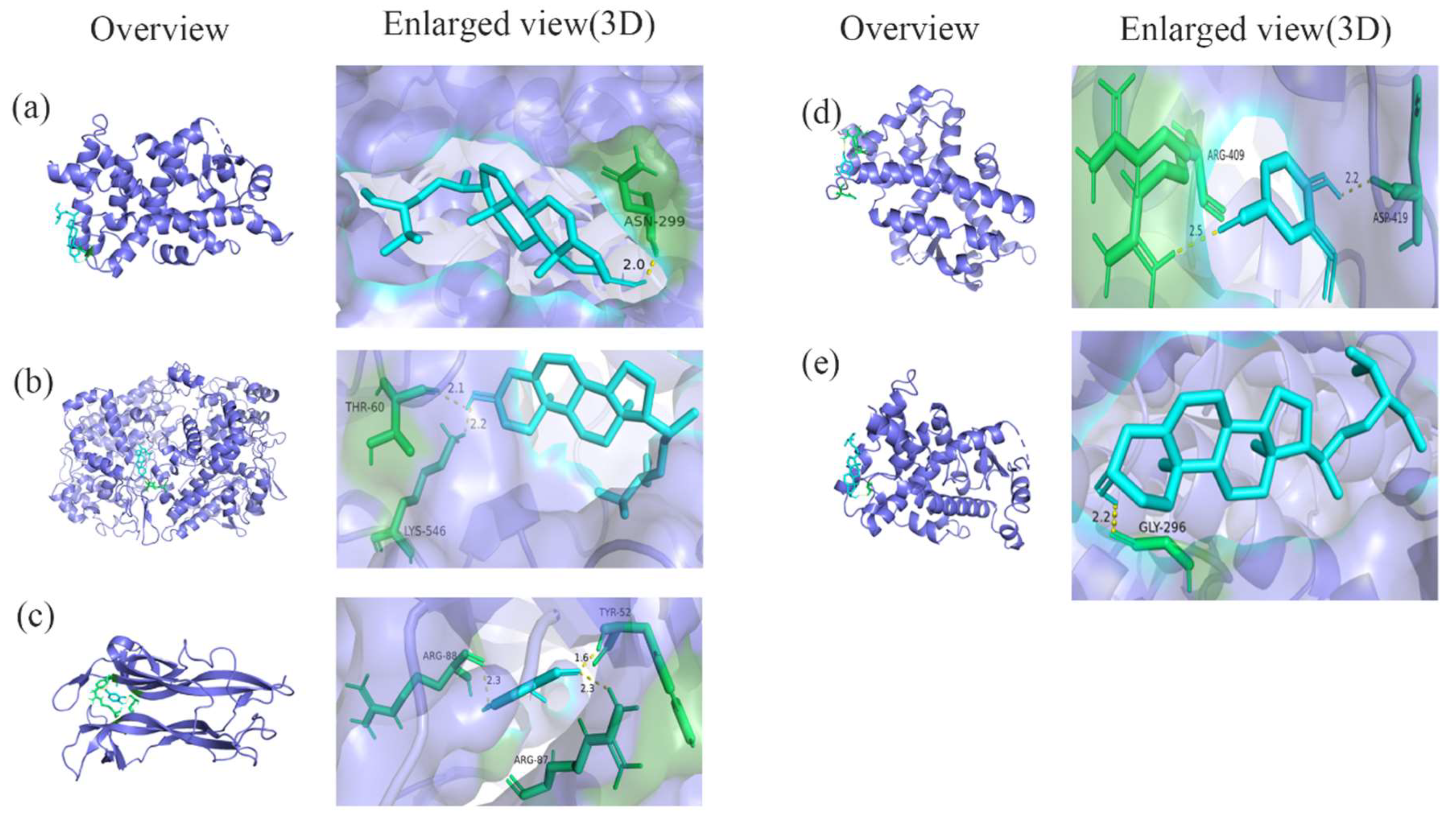
Figure 6.
Effects of vanillin on inflammatory factors in RAW264.7 cells stimulated by LPS. Mouse macrophages were divided into five groups: CON group, LPS group, LPS+0.4 mg/mL vanillin group, LPS+0.2 mg/mL vanillin group, and LPS+0.1 mg/mL vanillin group. After 24 h of DMEM culture, vanillin was added into cells stimulated by LPS (1 μg/mL) after 3 h, and the supernatants and proteins were taken after 24 h. (a-d) The expressions of cytokines. (a) IL-6; (b) IL-1β; (c) TNF-α; (d) IL-10. The levels of IL-6, IL-1β, TNF-α, and IL-10 were detected by ELISA kit. The data are expressed by the average standard deviation (n=3).
Figure 6.
Effects of vanillin on inflammatory factors in RAW264.7 cells stimulated by LPS. Mouse macrophages were divided into five groups: CON group, LPS group, LPS+0.4 mg/mL vanillin group, LPS+0.2 mg/mL vanillin group, and LPS+0.1 mg/mL vanillin group. After 24 h of DMEM culture, vanillin was added into cells stimulated by LPS (1 μg/mL) after 3 h, and the supernatants and proteins were taken after 24 h. (a-d) The expressions of cytokines. (a) IL-6; (b) IL-1β; (c) TNF-α; (d) IL-10. The levels of IL-6, IL-1β, TNF-α, and IL-10 were detected by ELISA kit. The data are expressed by the average standard deviation (n=3).
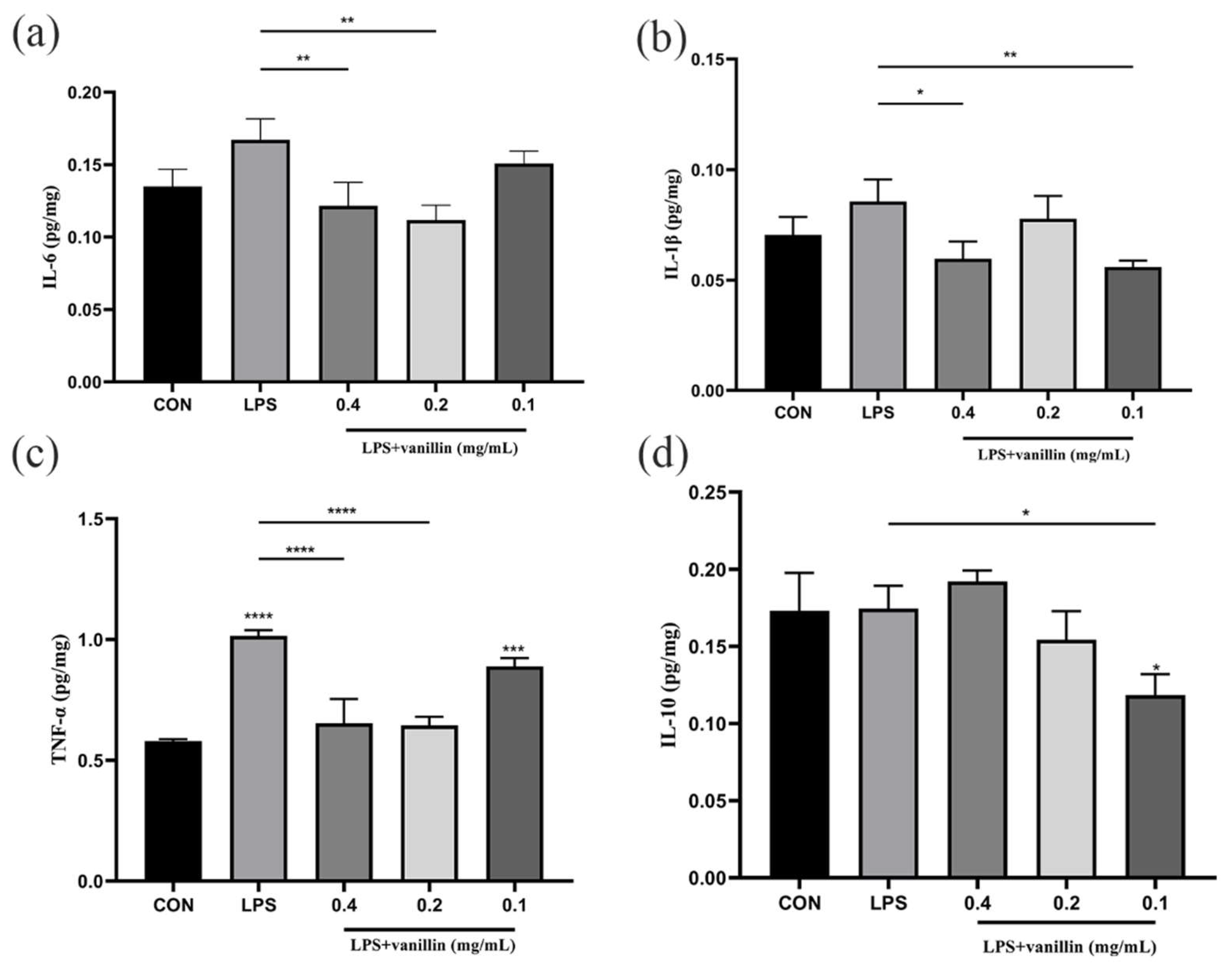

Table 1.
Screening of active components of C. japonicum. .
| Molecular ID | Molecule name | OB (%) | DL |
|---|---|---|---|
| MOL001641 MOL001735 MOL001749 MOL002032 MOL002879 MOL003180 MOL003344 MOL000359 MOL000449 MOL004746 MOL000057 MOL005736 MOL005840 MOL005842 MOL005846 MOL000612 MOL000675 |
Methyl linoleate Dinatin ZINC03860434 DNOP Diop Widdrene β-amyrin acetate Sitosterol Stigmasterol (E,7S,11R)-3,7,11,15-tetramethylhexadec-2-en-1-ol DIBP Cyperene PANA Pectolinarigenin Pectolinarin (-)-Alpha-cedrene Oleic acid |
41.93 30.97 43.59 40.59 43.59 53.81 42.06 36.91 43.83 49.63 51.87 50.35 41.17 47.62 43.08 55.56 33.13 |
0.17 0.27 0.35 0.4 0.39 0.12 0.74 0.75 0.76 0.13 0.13 0.11 0.13 0.3 0.65 0.1 0.14 |
| MOL000635 | Vanillin | 51.99 | - |
| MOL000676 | DBP | 64.54 | 0.13 |
-: no data.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



