
Submitted:
20 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
Amphibians, which are essential components of ecosystems, are susceptible to pharmaceutical contamination, a phenomenon of increasing concern owing to the widespread consumption and detection of pharmaceutical compounds in environmental matrices. This review investigates oxidative stress (OS) as the primary mechanism of drug toxicity in these organisms. The evidence gathered reveals that various pharmaceuticals, from antibiotics to anesthetics, induce OS by altering biomarkers of oxidative damage and antioxidant defense. These findings underscore the deleterious effects of pharmaceuticals on amphibian health and development and emphasize the necessity of incorporating OS biomarkers into ecotoxicological risk assessments. Although further studies on diverse amphibian species, drug mixtures, and field studies are required, OS biomarkers offer valuable tools for identifying sublethal risks. Furthermore, the development of more refined OS biomarkers will facilitate early detection of adverse effects, which are crucial for protecting amphibians and their ecosystems. Ultimately, this review calls for continued research and mitigation strategies to safeguard biodiversity from pharmaceutical contamination.
Keywords:
biomarkers
; bioindicators
; pharmaceuticals
; redox imbalance
; xenobiotics
1. Introduction
1.1. Amphibians
The taxonomic category Amphibia comprises 72 families and 562 genera, encompass over 8350 species. Approximately 3% of the total are caecilians, 9% are caudates, and 88% are frogs [1]. Most amphibians exhibit a biphasic life cycle characterized by the presence of both aquatic and terrestrial stages. The larval phase, which is typically aquatic, transitions to the adult phase, which can reside in either water or land environments. During their development into adults, larvae experience metamorphosis, which involves the transition from using gills to breathing air. Amphibians typically rely on moist skin as a secondary respiratory surface [1].
The Gymnophiona, Caudata, and Anura clades contain all the amphibian species [2]. Caecilians are legless burrowing amphibians belonging to the order Gymnophiona, which inhabit humid tropical areas in the Americas, Asia, and Africa. Most caecilians are less than 50 cm in length and are markedly similar to earthworms. They have small, highly ossified heads with annular grooves along the body, and degenerated eyes [3]. Caecilians are a diverse group of organisms, with approximately 200 different species identified to date. Despite their elusive nature, much of their biology remains unclear [4,5].
Salamanders belong to the order Caudata, which consists of over 700 species and is categorized into three groups: sirens (eel-like amphibians), basal salamanders (including hellbenders), and derived salamanders (mudpuppies, amphiumas, axolotls, newts, and various terrestrial species) [3,6]. Although it is widely believed that salamanders begin their lives as larvae in water and eventually transform into adult forms that live on land, this notion is not accurate for most salamander species [7]. Specifically, two-thirds of salamander species belong to the lungless salamander family (Plethodontidae), and these animals hatch directly from their eggs into terrestrial forms. Moreover, certain species, such as Necturus maculosus (commonly known as mudpuppy) and Ambystoma mexicanum (axolotl), exhibit neoteny, the retention of juvenile characteristics into adulthood. In these species, individuals can reach sexual maturity while still in their larval form and may never undergo metamorphosis [2].
The order Anura, which comprises toads and frogs, is the amphibian order with the greatest number of extant species, totaling 7708 [8]. These animals are distinguished by their small, tail-free bodies, damp and porous skin, large eyes, and long and powerful hind legs, which enable them to jump effectively. In most cases, their life cycle includes an aquatic larval stage with gills that transforms into a terrestrial adult with lungs. Anurans are found in a diverse range of ecosystems, encompassing tropical forests and deserts, and fulfill a vital function as both predators and prey for other species [3,4]. The significance of this order as an essential subject of study arises from its taxonomic and functional diversity, which enables researchers to understand the evolution and adaptation of vertebrates to various ecosystems. The order Anura comprises 55 families that are currently recognized and can be categorized into 11 superfamilies: Hyloidea, Dendrobatoidea, Microhyloidea, Ranoidea, Pelobatoidea, Pipoidea, Rhinophrynoidea, Scaphiopodoidea, Discoglossoidea, Pelodytoidea, and Leiopelmatoidea [8,9,10]. The ongoing classification of amphibians is subject to change as new phylogenetic research emerges, and additional species are uncovered.
1.2. Emerging Contaminants
The origin of Emerging Contaminants (EC) can be traced back to the Industrial Revolution, which introduced a plethora of new chemicals into the environment. Although the concept of these substances as potential environmental contaminants has recently gained traction, it was not a major concern during the earlier periods. Throughout the Industrial Revolution, until the late 20th century, conventional pollutants were the primary focus, with the recognition and comprehension of ECs undergoing substantial development in the latter half of the 20th century. The evolution of analytical techniques in the early 21st century has played a crucial role in the detection of low concentrations of new chemicals, leading to a significant shift in the perception of EC [11].
Emerging contaminants, which are substances that possess characteristics such as biotoxicity, environmental persistence, and bioaccumulation, are released into the environment and pose risks to both the ecological environment and human health. Despite these risks, their regulations and management have not been comprehensively addressed or effectively implemented [12].
Depending on their chemical characteristics, use, and origin, CE can be classified into several categories: I) pharmaceuticals and personal care products (PPCPs). This group of substances, intended to enhance health, hygiene, and aesthetics, is of increasing concern because of their environmental presence. It includes various compounds, such as prescription and over-the-counter medications, as well as personal care and grooming products, such as lotions, cosmetics, and fragrances [11]. II) Endocrine-disrupting chemicals (EDCs). Endocrine-disrupting compounds (EDCs) can impair the normal function of glands and hormones by blocking, mimicking, or disrupting their natural actions. Consequently, EDCs may adversely affect vital functions such as growth, development, and reproduction. [13]. III) Polyfluoroalkyl substances (PFAS). These compounds possess numerous carbon-fluorine bonds, imparting unique properties, such as resistance to heat, water, and oil. Their chemical and thermal stabilities have facilitated their use in diverse products, including firefighting foams, waterproof coatings, nonstick cookware, and various manufacturing processes [14]. IV) Microplastics (MPs). These tiny plastic fragments, less than 5 mm in length, include primary PMs, originally manufactured at this size, and secondary PMs, which result from the degradation of larger macroplastics (> 5 mm) under various environmental conditions. The widespread presence of microplastics in the air, soil, water, and organisms significantly threatens human health and ecosystem balance [15]. V) Nanomaterials. Materials sized between 1-100 nm exhibit unique physical, chemical, and biological properties owing to their nanometer scale. This category comprises carbon nanotubes, graphene, quantum dots, metal nanoparticles, and ceramic nanofibers. Nonetheless, their small size and high surface-to-volume ratio raise concerns about their potential adverse effects on human health and the environment [11]. VI) Industrial chemicals and by-products (ICBs). This category encompasses manufacturing chemicals and persistent environmental toxins such as dioxins. Additionally, heavy metals, such as lead and mercury, can contaminate soil and water, harming ecosystems and human health. Improper disposal of organic solvents and pesticides can disrupt natural cycles and the food chain [16,17].
1.3. Pharmaceuticals in the Environment
Pharmaceutically active compounds (PhACs), natural or synthetic chemical compounds with specific biological activities, are mainly used to prevent, diagnose, treat, or alleviate disease symptoms [18]. The traditional definition of "drug" suggests that most of these compounds act in a similar way in any biological system, depending on their mechanism of action. However, it is important to recognize that amplified or unexpected effects may arise in certain species owing to physiological variations, including differences in the manner in which the drug is metabolized and interacts with the organism [19].
PhACs enter the environment primarily through two pathways, both through their use in humans and animals and their subsequent metabolic excretion in urine and feces (including unmetabolized parent drugs, drug conjugates, and bioactive metabolites), and improper disposal of leftover or expired medications in the sewage system [20]. PhACs that undergo incomplete degradation in wastewater treatment plants (WTPs) are released into treated effluents, causing the detection of these compounds in surface water, seawater, groundwater, and even drinking water at concentrations ranging from ngL-1 to μgL-1 [21,22,23,24,25,26,27,28,29,30,31]. In addition, irrigation with treated effluents or the application of sludge from WTPs (biosolids) in crop fields is a common pathway for soil contamination with PhACs [32,33,34].
PhACs are present at low concentrations in the environment, typically ranging from ngL-1 to µgL-1, but their pervasiveness poses a potential threat to organisms in both aquatic and terrestrial ecosystems [35,36,37]. The persistent nature of several PhACs increases the possibility of bioaccumulation in different organisms, which can lead to serious physiological disorders [38,39,40]. Although some other PhACs do not present this persistence characteristic, their continuous release and constant presence in the environment confers them a "pseudopersistent" behavior, representing a risk to living organisms [41,42].
Wildlife can play a key role in assessing environmental contamination by PhACs, acting as sentinels, monitoring bioaccumulation, and serving as a bioindicator of potential adverse effects depending on the environmental fate and mechanism of action of each PhAC. In addition, we must recognize the importance of the interconnection between human, animal, and environmental health, highlighting the need to further investigate the impact of PhACs on wildlife, especially considering their role in complex food webs involving humans [18].
1.4. Oxidative Stress
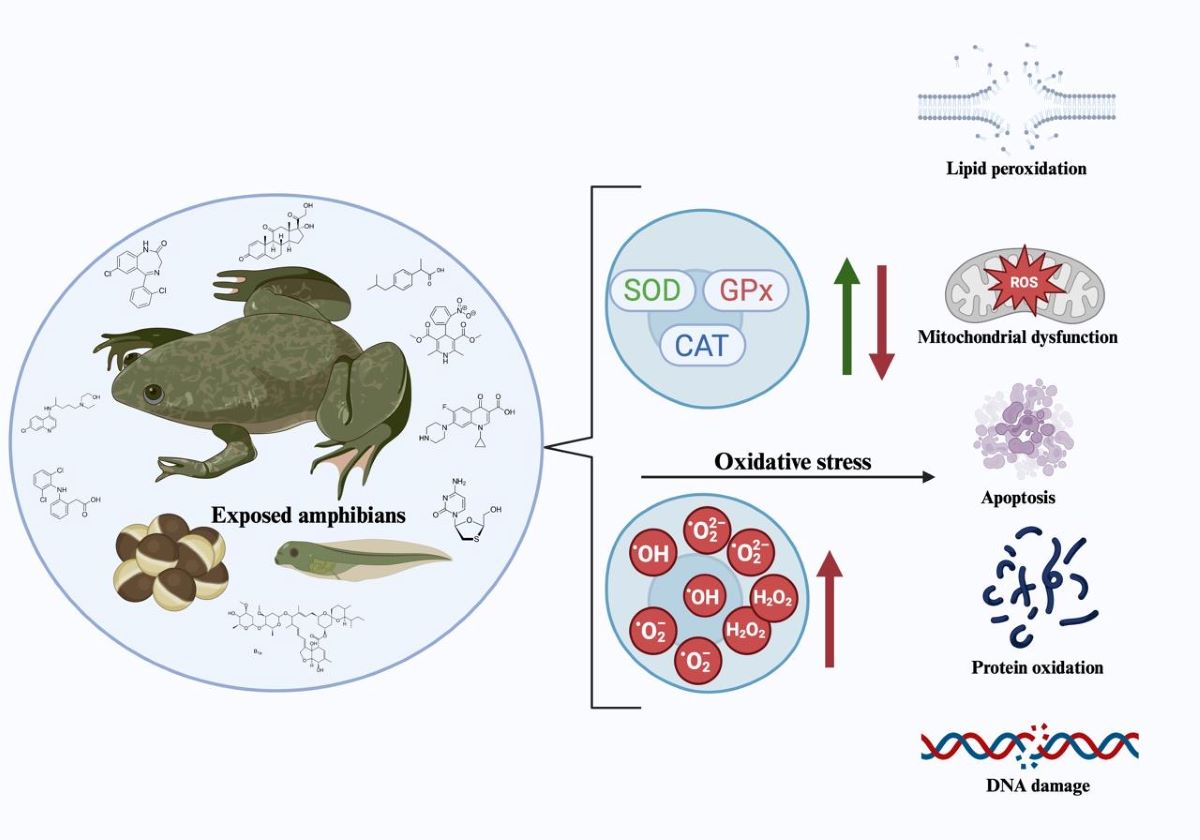
Oxidative stress (OS) is characterized by an imbalance between oxidative and antioxidant species, favoring the former. This imbalance disturbs signaling and redox control and can lead to damage at the molecular level [43]. A healthy redox system is based on the balance between oxidation and reduction, which implies a net-zero electron flow at the end of the biological pathways. Alterations in this redox steady state trigger molecular changes that, depending on their magnitude, can result in damage at the molecular, cellular, and/or tissue levels. Both endogenous and exogenous sources lead to constant OS exposure [44].
Various environmental pollutants, including pharmaceuticals, can increase ROS production in amphibians and other aquatic organisms, inducing oxidative stress. Xenobiotics can elevate intracellular levels of ROS through various mechanisms, such as increased basal metabolism with consequent intensification of mitochondrial activity, alteration of the redox cycle, increased generation of ROS as by-products of reactions mediated by cytochrome P450 enzymes (CYPs), or increased Fenton and Haber-Weiss reactions due to excess copper and iron ions. Oxidative stress biomarkers, which are first-line responses in animals and are highly sensitive to pollutants even at low concentrations, are useful for the early detection of environmental contamination [45].
1.4.1. Oxidative Stress Biomarkers
1.4.1.1. Oxidative Damage
Excessive ROS can damage various biomolecules including DNA, lipids, proteins, and carbohydrates (Figure 1). This damage can be detected using OS-specific biomarkers, which identify the molecular fingerprints left by ROS [46].
Lipids: When the concentration of ROS inside the cell increases lipid peroxidation (LPO) is triggered. This process is initiated by the attack of a free radical on the methylene group of the fatty acid to extract a hydrogen atom. Consequently, a carbon-centered lipid radical (L•) is formed, which generates a lipid peroxide radical (LOO•) upon reaction with molecular oxygen. Subsequently, lipid peroxide undergoes a cyclization reaction to form an endoperoxide, which eventually decomposes into toxic end products such as malondialdehyde (MDA), 4-oxy-2-nonenal (ONE) and 4-hydroxyl nonenal (4-HNE) [44]. These end products are detrimental to the cell as they can cause damage to proteins and DNA. In addition, lipid peroxidation compromises the functionality of the cell membrane, decreasing its fluidity and inactivating cell membrane-anchored receptors and enzymes [47].
Proteins: Protein oxidation involves the participation of various ROS and propagating radicals. These reactions lead to oxidative modifications in amino acid side chains, ROS-mediated peptide fragmentation, reactions between peptides and lipids or carbohydrate oxidation products, and formation of carbonyl derivatives of proteins [44]. Carbonylation of protein residues has been established as a key biomarker to assess protein damage caused by ROS, affecting specific amino acid residues, such as lysine, threonine, proline, and arginine [48,49].
DNA and RNA: Free radicals and ROS can induce specific modifications and hydroxylation in the purine and pyrimidine bases of DNA, damage the deoxyribose-phosphate backbone, and disrupt protein-DNA cross-linkage formation [47]. Mitochondrial DNA is more vulnerable to this type of damage than nuclear DNA due to its closer proximity to the site of ROS generation. In addition, there is scientific evidence that RNA is more susceptible to oxidative damage than DNA, due to factors such as its proximity to mitochondria (the main source of ROS), its single-stranded structure, the lack of an active repair mechanism for oxidative damage, and lower protection by proteins compared to DNA [44]. One of the most investigated biomarkers of damage is 7,8-dihydro-8-deoxyguanosine (8-oxoG) [50,51].
1.4.1.2 Antioxidant Defenses
Constant exposure to oxidizing factors, both internal and external, has led aerobic organisms to develop complex antioxidant mechanisms, including enzymes and other molecules, to maintain the redox balance. The study of these antioxidant enzymes has made it possible to assess an organism's ability to cope with OS and, therefore, can be used as a biomarker [52].
The enzymes superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) constitute the first line of cellular antioxidant defense. These enzymes are functionally interrelated because hydrogen peroxide (H2O2), the product of the reaction catalyzed by SOD, is the substrate for both CAT and GPx [53].
SOD is a crucial metalloenzyme involved in cellular defence against ROS. It acts as the first line of detoxification and is the most potent antioxidant within cells [54].Its main function is to catalyze the conversion of two highly reactive superoxide anions (•O2-) into hydrogen peroxide (H2O2) and molecular oxygen (O2), thus reducing its damaging potential. SOD activity depends on the presence of a specific metal cofactor, resulting in different enzyme forms depending on the type of metal ion required [55].
CAT, which is widely distributed in living tissues that utilize oxygen, uses iron or manganese as a cofactor. Its main function is to catalyze the decomposition of hydrogen peroxide (H2O2) into water and O2, thus completing the detoxification process initiated by SOD [56].
GPx is an important intracellular enzyme that breaks down H2O2 into water and lipid peroxides into their corresponding alcohols, mainly in the mitochondria and occasionally in the cytosol. In most cases, its activity depends on selenium (Se) as a cofactor. Therefore, GPx is often referred to as selenocysteine peroxidase. This enzyme plays a crucial role in inhibiting the lipid peroxidation process, thus protecting OS cells [54,57].
Amphibians, owing to their sensitivity to chemical contaminants during their life cycles in water, are considered excellent bioindicators of environmental pollution. The decline in their populations in recent decades has been attributed to the combined effects of pollution, changes in human activity, and climate [58,59]. Despite their importance as indicators of environmental health, toxicological research on amphibians has been limited compared to that on other vertebrates [60,61]. The current study seeks to provide a comprehensive and critical analysis of the scientific literature to evaluate the existing evidence on the use of oxidative stress biomarkers as indicators of toxicity caused by various pharmaceutical compounds in different species of amphibians. This review aims to be exhaustive in its scope and provide a critical examination of the available evidence on this topic. The objective of analyzing these biomarkers is to identify response patterns, assess the sensitivity of various species, and determine the relevance of these biomarkers in conducting ecotoxicological risk assessments.
2. Methodology
An exhaustive bibliographic search was carried out in the electronic databases Web of Science, PubMed, and Scopus, covering the period 2000 to 2024. Combinations of the following search terms were used: oxidative stress, amphibians, drugs, pharmaceuticals, biomarkers, oxidative damage, SOD, CAT and GPx.
The inclusion criteria were as follows:
- Experimental investigations analyzing the impact of drug exposure on oxidative stress biomarkers in amphibians.
- Articles published in peer-reviewed scientific journals in English.
- Studies providing data on at least one oxidative stress biomarker and drug-specific class.
- Research that clearly identifies the amphibian species used in this study.
Excluded:
- Studies using plant extracts or natural compounds instead of synthetic drugs.
- Studies that did not provide quantitative data on biomarkers of oxidative stress.
- Studies that focus exclusively on drug bioaccumulation without evaluating biomarkers of oxidative stress.
- Studies that do not provide sufficient information on experimental conditions.
Data extraction was performed systematically, documenting the following information for each study: authors, year of publication, amphibian species used, type of drug, concentration used, exposure time, main results, and conclusions.
3. Results
This review identified several studies that have investigated the effects of various pharmaceuticals on biomarkers of oxidative stress in amphibians. The results revealed complex and varied responses depending on both the type of drug and amphibian species studied (Table 1). It is important to note that all studies to date have focused on frog and toad species, such as Rhinella arenarum, Xenopus laevis, and Lithobates catesbeianus. However, amphibian groups of salamanders and caecilians remain largely unexplored in this context, representing a substantial gap in our understanding of the impact of pharmaceuticals on amphibian biodiversity.
3.1. Antibiotics
Antibiotics are pharmaceutical agents extensively utilized in both human and veterinary medicine to combat infectious diseases. Furthermore, they are administered at subtherapeutic doses as feed additives in animal husbandry to promote growth [78]. The global consumption of antibiotics is estimated to range between 100,000 and 200,000 metric tons. Nevertheless, a substantial proportion, ranging from 70% to 90%, is not fully metabolized within an organism and is subsequently excreted in its original form or as active metabolites, thereby entering the environment [79]. Previous studies have provided information on the toxicity of antibiotics to algae [80,81], microcrustaceans [82,83,84], mollusk bivalves [85,86,87], and fish [88,89,90,91]. Nevertheless, research on the impact of antibiotics on amphibian populations remains limited. The lethal and sublethal effects of oxytetracycline (OTC) were evaluated in Rhinella arenarum embryos exposed to 10–115 mgL-1 for a period of 96 h. These findings indicated that OTC exposure significantly altered the activity of antioxidant enzymes, including decreased CAT, SOD, and GST activity. In addition, an increase in GSH levels was observed. Although no oxidative damage was evident, the authors suggested that continued exposure to OTC might have long-term negative effects [71]. In another study, da Luz et al. (2021) [69] utilized Physalaemus cuvieri tadpoles to assess the acute toxicity of azithromycin (AZT) and hydroxychloroquine (HCQ). Tadpoles were exposed to a concentration of 12.5 μgL-1 of the drugs for 72 h. This concentration was selected to simulate the potential increase in the environmental drug concentrations attributable to the COVID-19 pandemic. The study found no evidence of increased oxidative damage in tadpoles exposed to the drug, as assessed by biomarkers, such as nitrite, TBARS, ROS and H2O2. However, an increase in SOD and CAT activities was observed, suggesting an adaptive response to counteract potential oxidative stress. In addition, the toxicity of two veterinary antibiotics, enrofloxacin (ENR) and ciprofloxacin (CPX), was evaluated in R. arenarum larvae exposed to environmentally relevant concentrations of both drugs (1-1000 μgL-1) for 96 hours under standard laboratory conditions. Concentrations higher than 10 μgL-1 of both antibiotics induced detrimental effects on larvae, mainly on development, growth and antioxidant enzyme activity. Specifically, CPX at 1000 μgL-1 induced a significant increase in GST activity and ENR at 1000 μgL-1 inhibited both GST and CAT. These findings suggest that the tested antibiotics can trigger EO and affect antioxidant defense mechanisms in R. arenarum larvae [65]. The effects of two commonly used antibiotics, sulfamethoxazole (SMX) and oxytetracycline (OTC), were evaluated in tadpoles of Lithobates catesbeianus, which were exposed to 20, 90 and 460 ngL-1 of both antibiotics for 16 days. OTC, especially at the highest concentrations, led to a decrease in SOD, GPx and glucose 6-phosphate dehydrogenase (G6PDH) activities. In addition, increased levels of carbonylated proteins were observed in the liver of tadpoles exposed to the highest concentration of OTC. In contrast, SMX did not significantly affect the evaluated biomarkers [72]. Finally, one study evaluated the toxicity of a commercial formulation of monensin (CFM), a polyether antibiotic isolated from Streptomyces cinnamonensis, on embryos and larvae of R. arenarum exposed to 4, 12 and 20 μgL-1 following acute exposure for 96 h. In embryos, a significant decrease in GST activity was observed at 12 and 20 μgL-1. In larvae, GST activity was significantly decreased at 20 μgL-1. In addition, low GSH levels were observed in larvae exposed to 12 and 20 μgL-1, and an increase in lipid peroxidation was observed at all concentrations tested [76].
3.2. Nonsteroidal Anti-Inflammatory Drugs
Nonsteroidal anti-inflammatory drugs (NSAIDs) are a group of drugs with analgesic, anti-inflammatory, and antipyretic properties. Their widespread use worldwide, with billions of prescriptions dispensed each year, makes them one of the most widely consumed types of drugs [92]. In addition, NSAIDs have been shown to be resistant to degradation, persistent, pharmacologically active and toxic to non-target organisms such as algae [93,94], molluscs [95,96] and fish [97,98,99]. As for the studies in amphibians, embryos and larvae of Trachycephalus typhonius and Physalaemus albonotatus were exposed to concentrations between 125 and 2000 μgL-1 for 22 days. In T. typhonius, GST activity was inhibited at low concentrations (125μgL-1) but induced at high concentrations (500, 1000 and 2000 μgL-1). In P. albonotatus, GST activity was inhibited at both low and high concentrations, exhibiting a hormesis-type response pattern, with maximal activity at intermediate concentrations. GST activity shows complex responses and varies by species, highlighting the importance of considering species-specific sensitivity when assessing the ecological risk of these compounds [66]. In another study, adult male Pelophylax ridibundus frogs were exposed to 250 ngL-1 ibuprofen for 14 days. These findings indicate that ibuprofen exposure caused an increase in oxyradicals and glutathione levels (both reduced and oxidized); however, SOD activity was not affected. Additionally, the concentration of lipofuscin, a marker of oxidative damage, decreased with drug exposure
3.3. Antivirals
Antivirals (AV), drugs designed to combat viral infections by limiting or preventing viral replication, have increased in consumption. Consequently, an increase in the concentrations of these compounds and their active metabolites in the environment has been observed [100]. The presence of AV in ecosystems poses a potential risk, because they can interfere with the normal functions of biological systems [101]. Rhinella arenarum tadpoles were exposed to four antiretrovirals (lamivudine (3TC), stavudine (d4T), zidovudine (AZT), and nevirapine (NVP)) at concentrations ranging from 0.5 to 4 mgL-1 for a period of 48 h to determine the acute toxicity of these compounds. Among the four compounds, 3TC showed the lowest bioaccumulation. A statistically significant increase in GST activity was observed only at the highest concentration tested. The bioaccumulation of d4T was slightly higher than that of 3TC was. In addition, the increase in GST activity was statistically significant at 1, 2 and 4 mgL-1. AZT showed low bioaccumulation in tadpoles. GST activity similarly increased at concentrations of 1, 2 and 4 mgL-1. Finally, NVP exhibited the highest bioaccumulation of all tested compounds, suggesting a high permeability to tadpoles. The increase in GST activity was statistically significant at all the four concentrations tested. The observed increase in GST activity indicates overproduction of ROS, which can cause oxidative damage in R. arenarum tadpoles [68]. A recent study evaluated the toxicity of two VA, favipiravir (32.9-250 mgL-1) and oseltamivir (8.2-62.5 mgL-1), in Xenopus laevis embryos and tadpoles for a 96-h period. Favipiravir inhibits the activity of GR and CAT enzymes in embryos; however, it increases GST activity and decreases MDA levels. En embriones expuestos a oseltamivir no se observaron cambios significativos en la actividad de CAT ni en los niveles de MDA. In tadpoles, both GST and GR activities and MDA levels decreased. The results suggest that VA may affect antioxidant defense systems in X. laevis, and that the response varies depending on the developmental stage of the organism [74]
3.4. Antihypertensive
The recurrent detection of antihypertensive drugs in the environment reflects the accelerated growth of the pharmaceutical industry and the high consumption of these drugs globally [102]. Additionally, the presence of these drugs in the environment can have harmful effects on organisms, affecting their health and survival [103,104,105]. A study was conducted on adult male frogs of the species Pelophylax ridibundus to evaluate the toxicity of nifedipine at a concentration of 10 μM for a period of 14 days. Drug exposure causes oxidative stress, as evidenced by a considerable increase in the rate of ROS generation and SOD, GSH, and GSSG levels [63].
3.5. Glucocorticoids
Glucocorticoids (GC) are widely used in the treatment of various diseases, such as rheumatoid arthritis, asthma, and Crohn's disease, owing to their potent anti-inflammatory properties [106]. Both natural and synthetic GCs exert their main action through the glucocorticoid receptor (GR), which modulates the expression of specific genes. In addition, they can act through non-genomic mechanisms, binding directly to the GR without affecting gene expression [107]. The increase in the human population, together with its aging, has led to a greater dependence on GCs, as is the case with other drugs. This growing demand raises concerns regarding the potential environmental impacts of the consumption and presence of these drugs in the environment [108]. Among the studies that have evaluated the toxicity of glucocorticoids in amphibians, Cuzziol Boccioni et al. (2020) [67] evaluated the chronic toxicity of dexamethasone at concentrations of 1-1000 μgL-1 in Rhinella arenarum larvae for an exposure period of 22 days. The results showed a significant increase in GST activity in the larvae exposed to dexamethasone, indicating a response to oxidative stress. In addition, the authors related the histological alterations observed in different tissues, such as intestinal dysplasia and epithelial cells, to oxidative damage induced by the drug. Rutkoski et al. (2024) [77] evaluated the toxicity of two glucocorticoids, prednisone (PD) and prednisolone (PL), on Aquarana catesbeianus tadpoles exposed to concentrations of 0.1, 1 and 10 μgL-1 for 16 days. PD exposure caused an increase in the MDA levels. In addition, both drugs caused an increase in CAT and GPx activities, while PD exposure also elevated SOD, GST, and G6PDH activities. These findings suggest that GCs induce oxidative stress in tadpoles of A. catesbeianus.
3.6. Pharmaceutical Mixture
Research has demonstrated that individual pharmaceutical compounds can influence diverse molecular and cellular pathways in various non-target organisms. Furthermore, the nature and intensity of the effects were directly correlated with the dosage and specific compound. However, it is crucial to consider that under real conditions, organisms are often exposed to low doses of multiple drugs simultaneously. This combined exposure can modulate the overall toxicity through different pathways; however, they often interact with each other, adding to the complexity of assessing the environmental impact of pharmaceuticals [109]. A study was conducted to evaluate the effect of a pharmaceutical mixture comprising diclofenac, naproxen, atenolol, and gemfibrozil on Limnodynastes peronii tadpoles. The subjects were exposed to various concentrations of the mixture (0.1, 1, 10, 100, and 1000 μgL-1) for 30-day period. The results showed that peroxidase activity increased significantly in tadpoles exposed to 1000 μgL-1 concentration, suggesting an increase in oxidative stress. However, no significant effects on SOD or RBC activity were observed at any of the tested concentrations [62].
3.7. Anesthetic
Tricaine methanesulfonate (MS-222) is the anesthetic of choice for amphibians and fish, with applications ranging from simple procedures requiring sedation, such as morphometric measurements, to its use as part of euthanasia protocols, either as a first step or as the sole agent [110,111]. A study conducted by Gavrilović et al. (2024) [73] investigated the potential impact of MS-22 as an anesthetic/euthanizing agent on experimental outcomes in studies examining biomarkers of oxidative stress. Hyla arborea tadpoles were reared at two different temperatures (20°C and 25°C) to induce variations in their antioxidant capacities. Subsequently, the tadpoles were exposed to 0.1, 1 and 5 gL-1 of MS-222 for 15 min. The results of this study indicated that MS-222 can significantly alter GSH levels and GSH/thiol-related parameters. Furthermore, specimens from different temperature groups exhibited varying responses to MS-222, suggesting a potential correlation with initial levels of antioxidant capacity. The biomarkers of oxidative damage and CAT activity were not significantly affected by MS-222 exposure.
3.8. Benzodiazepines
Benzodiazepines (BDZ), which are psychotropic drugs widely prescribed worldwide, are used to treat mental disorders, including anxiety, panic disorder, and insomnia. Its therapeutic action is due to its interaction with the γ-aminobutyric acid (GABA) receptor, which results in an increase in ionic conduction and the consequent manifestation of anxiolytic, hypnotic, and sedative effects [112,113]. The presence of BDZ in the environment poses a potential risk due to its ability to be absorbed by organisms. Several studies have demonstrated the ability of these compounds to bioaccumulate and cause adverse effects in various aquatic organisms [114,115,116,117]. In amphibians, limited research has been conducted on the effects of BDZ on these organisms. Fogliano et al. (2022) [70] exposed X. laevis embryos to 1, 5 and 10 μgL-1 of delorazepam (DLZ) from the 4-8 cell stage to stage 45-46, simulating early and prolonged exposure to the drug in a natural environment. DLZ exposure caused a significant increase in ROS production and lipid hydroperoxide levels, indicating oxidative damage at the cellular level. In response to stress, an increase in GPx and GR activity was detected, although this response was not sufficient to completely counteract the oxidative damage induced by the drug, especially at the highest concentration.
3.9. Antiparasitic
The increasing use of macrocyclic lactones for the control of parasitic infections has been accompanied by significant historical developments. The first macrocyclic lactones used to combat parasites (roundworms and arthropods) were avermectin and its chemical derivative ivermectin (IVM) (C.Campbell, 2012). IVM, owing to its broad-spectrum anthelmintic activities, is used in the treatment of various diseases caused by parasitic nematodes [119]. Previous studies have shown adverse effects of IVM on non-target organisms in both terrestrial and aquatic ecosystems [120,121,122,123,124]. Embryos and larvae of Rhinella arenarum were exposed to different concentrations of IVM, both the active ingredient and a commercial formulation, varying between 1.5, 5 and 10 μgL-1 for 96 h. Exposure to IVM, both in its active ingredient form and in a commercial formulation, induced significant changes in oxidative stress biomarkers. CAT activity was increased in embryos exposed to the commercial formulation and in larvae exposed to both forms of IVM. GST activity increased in embryos and larvae exposed to the commercial formulation but was inhibited in embryos exposed to the active ingredient. In addition, GSH levels were decreased in embryos exposed to the active ingredient. Los niveles de TBARS aumentaron en embriones y larvas expuestos a la formulación comercial. This study highlighted that the commercial formulation of IVM is more toxic than the active ingredient, underscoring the importance of considering the effects of excipients in environmental risk assessments [75].
4. Conclusions and Future Research
This review provides a comprehensive analysis of the role of oxidative stress in the mechanism underlying drug-induced toxicity in amphibians. Research has demonstrated that a diverse array of pharmaceutical compounds, including antibiotics, nonsteroidal anti-inflammatory drugs, antivirals, antihypertensives, glucocorticoids, anesthetics, and benzodiazepines can elicit oxidative stress in these organisms. Alterations observed in the biomarkers of oxidative damage include increased ROS production, lipid peroxidation, and protein oxidation, along with changes in the activity of antioxidant enzymes, highlighting the detrimental effects of these compounds on amphibian health and development. The results of this review emphasize the significance of considering oxidative stress in the ecotoxicological risk assessments of pharmaceuticals. The use of oxidative stress biomarkers can provide valuable information regarding the sublethal effects of drug exposure and can facilitate the identification of potential risks to amphibian populations.
However, research in this field is still limited and there is room for improvement. It is crucial to expand the species spectrum as no studies have been performed on salamanders (order Caudata) or caecilians (order Gymnophiona), which limits our understanding of the effects of drugs on amphibian diversity. This review identified only one study that evaluated the effects of a drug mixture. Most of the reviewed studies have focused on the short-term effects of individual drugs under laboratory conditions, which limits their extrapolation to real situations where amphibians are exposed to multiple contaminants and stressors. Further research is required to investigate the effects of pharmaceutical combinations and conduct field studies to adequately assess the impact of pharmaceuticals on the natural environment. Additionally, the development of more sensitive and specific biomarkers is essential for early detection of adverse drug effects in amphibians.
Finally, it is imperative to continue researching the effects of drugs on amphibians and other aquatic organisms as well as to develop effective strategies to mitigate their negative effects. Only through a comprehensive understanding of the risks associated with pharmaceutical pollution can we safeguard the health of ecosystems and ensure the preservation of the species inhabiting them.
Author Contributions
J.D.C.V., H.I.F., I.P.A. and N.D.C, writing—original draft preparation, review and editing. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Berkovitz, B.; Shellis, P. Amphibians. The Teeth of Non-Mammalian Vertebrates 2023, 203–257. [CrossRef]
- O’Rourke, D.P.; Rosenbaum, M.D. Biology and Diseases of Amphibians. Laboratory Animal Medicine: Third Edition 2015, 931–965. [CrossRef]
- Vitt, L.J.; Caldwell, J.P. Herpetology: An Introductory Biology of Amphibians and Reptiles: Fourth Edition; 2013.
- Wells, K.D. The Ecology and Behavior of Amphibians; 2013.
- Gower, D.J.; Wilkinson, M. Conservation Biology of Caecilian Amphibians. Conservation Biology 2005, 19. [Google Scholar] [CrossRef]
- Rozenblit, F.; Gollisch, T. What the Salamander Eye Has Been Telling the Vision Scientist’s Brain. Semin Cell Dev Biol 2020, 106, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Elinson, R.P.; del Pino, E.M. Developmental Diversity of Amphibians. Wiley Interdiscip Rev Dev Biol 2012, 1. [Google Scholar] [CrossRef] [PubMed]
- Frost, D.R. Amphibian Species of the World. Available at Http://Research.Amnh.Org/Herpetology/Amphibia/Index.Html. American Museum of Natural History, New York, USA. Accessed on 09 Sep 2023. 2023.
- Rivera-Correa, M.; Baldo, D.; Candioti, F.V.; Orrico, V.G.D.; Blackburn, D.C.; Castroviejo-Fisher, S.; Chan, K.O.N.N.; Gambale, P.; Gower, D.J.; Quah, E.S.H.; et al. Amphibians in Zootaxa: 20 Years Documenting the Global Diversity of Frogs, Salamanders, and Caecilians. Zootaxa 2021, 4979. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, D.C.; Wake, D.B. Class Amphibia Gray, 1825. In: Zhang, Z.-Q. (Ed.) Animal Biodiversity: An Outline of Higher-Level Classi fication and Survey of Taxonomic Richness. Zootaxa 2011, 3148. [Google Scholar] [CrossRef]
- Li, X.; Shen, X.; Jiang, W.; Xi, Y.; Li, S. Comprehensive Review of Emerging Contaminants: Detection Technologies, Environmental Impact, and Management Strategies. Ecotoxicol Environ Saf 2024, 278, 116420. [Google Scholar] [CrossRef]
- Li, Z.; Yuan, D. Global Performance and Trends of Research on Emerging Contaminants in Sewage Sludge: A Bibliometric Analysis from 1990 to 2023. Ecotoxicol Environ Saf 2024, 281, 116597. [Google Scholar] [CrossRef]
- Ahn, C.; Jeung, E.B. Endocrine-Disrupting Chemicals and Disease Endpoints. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Evich, M.G.; Davis, M.J.B.; McCord, J.P.; Acrey, B.; Awkerman, J.A.; Knappe, D.R.U.; Lindstrom, A.B.; Speth, T.F.; Tebes-Stevens, C.; Strynar, M.J.; et al. Per- and Polyfluoroalkyl Substances in the Environment. Science (1979) 2022, 375. [Google Scholar] [CrossRef]
- Kye, H.; Kim, J.; Ju, S.; Lee, J.; Lim, C.; Yoon, Y. Microplastics in Water Systems: A Review of Their Impacts on the Environment and Their Potential Hazards. Heliyon 2023, 9, e14359. [Google Scholar] [CrossRef]
- Tong, Y.; Gao, J.; Yue, T.; Zhang, X.; Liu, J.; Bai, J. Distribution, Chemical Fractionation, and Potential Environmental Risks of Hg, Cr, Cd, Pb, and As in Wastes from Ultra-Low Emission Coal-Fired Industrial Boilers in China. J Hazard Mater 2023, 446, 130606. [Google Scholar] [CrossRef]
- Sorvari, J.; Wahlström, M. Industrial By-Products. Handbook of Recycling: State-of-the-art for Practitioners, Analysts, and Scientists 2024, 259–285. [CrossRef]
- Bean, T.G.; Chadwick, E.A.; Herrero-Villar, M.; Mateo, R.; Naidoo, V.; Rattner, B.A. Do Pharmaceuticals in the Environment Pose a Risk to Wildlife? Environ Toxicol Chem 2024, 43. [Google Scholar] [CrossRef]
- Toutain, P.L.; Ferran, A.; Bousquet-Mélou, A. Species Differences in Pharmacokinetics and Pharmacodynamics. Handb Exp Pharmacol 2010, 199. [Google Scholar]
- Fernández, C.; Beltrán, E.M.; Tarazona, J. V. Pharmaceuticals Effects in the Environment. Encyclopedia of Toxicology: Third Edition 2014, 844–848. [CrossRef]
- Xu, M.; Huang, H.; Li, N.; Li, F.; Wang, D.; Luo, Q. Occurrence and Ecological Risk of Pharmaceuticals and Personal Care Products (PPCPs) and Pesticides in Typical Surface Watersheds, China. Ecotoxicol Environ Saf 2019, 175, 289–298. [Google Scholar] [CrossRef]
- Kim, S.D.; Cho, J.; Kim, I.S.; Vanderford, B.J.; Snyder, S.A. Occurrence and Removal of Pharmaceuticals and Endocrine Disruptors in South Korean Surface, Drinking, and Waste Waters. Water Res 2007. [Google Scholar] [CrossRef]
- Ebele, A.J.; Oluseyi, T.; Drage, D.S.; Harrad, S.; Abou-Elwafa Abdallah, M. Occurrence, Seasonal Variation and Human Exposure to Pharmaceuticals and Personal Care Products in Surface Water, Groundwater and Drinking Water in Lagos State, Nigeria. Emerg Contam 2020. [Google Scholar] [CrossRef]
- Shen, X.; Chang, H.; Sun, Y.; Wan, Y. Determination and Occurrence of Natural and Synthetic Glucocorticoids in Surface Waters. Environ Int 2020, 134, 105278. [Google Scholar] [CrossRef] [PubMed]
- Ashfaq, M.; Li, Y.; Rehman, M.S.U.; Zubair, M.; Mustafa, G.; Nazar, M.F.; Yu, C.P.; Sun, Q. Occurrence, Spatial Variation and Risk Assessment of Pharmaceuticals and Personal Care Products in Urban Wastewater, Canal Surface Water, and Their Sediments: A Case Study of Lahore, Pakistan. Science of the Total Environment 2019, 688, 653–663. [Google Scholar] [CrossRef]
- Mainero Rocca, L.; Gentili, A.; Caretti, F.; Curini, R.; Pérez-Fernández, V. Occurrence of Non-Steroidal Anti-Inflammatory Drugs in Surface Waters of Central Italy by Liquid Chromatography–Tandem Mass Spectrometry. Int J Environ Anal Chem 2015, 95, 685–697. [Google Scholar] [CrossRef]
- Praveena, S.M.; Shaifuddin, S.N.M.; Sukiman, S.; Nasir, F.A.M.; Hanafi, Z.; Kamarudin, N.; Ismail, T.H.T.; Aris, A.Z. Pharmaceuticals Residues in Selected Tropical Surface Water Bodies from Selangor (Malaysia): Occurrence and Potential Risk Assessments. Science of The Total Environment 2018, 642, 230–240. [Google Scholar] [CrossRef]
- López-Serna, R.; Jurado, A.; Vázquez-Suñé, E.; Carrera, J.; Petrović, M.; Barceló, D. Occurrence of 95 Pharmaceuticals and Transformation Products in Urban Groundwaters Underlying the Metropolis of Barcelona, Spain. Environmental Pollution 2013. [Google Scholar] [CrossRef] [PubMed]
- Björlenius, B.; Ripszám, M.; Haglund, P.; Lindberg, R.H.; Tysklind, M.; Fick, J. Pharmaceutical Residues Are Widespread in Baltic Sea Coastal and Offshore Waters – Screening for Pharmaceuticals and Modelling of Environmental Concentrations of Carbamazepine. Science of the Total Environment 2018. [Google Scholar] [CrossRef] [PubMed]
- Alygizakis, N.A.; Gago-Ferrero, P.; Borova, V.L.; Pavlidou, A.; Hatzianestis, I.; Thomaidis, N.S. Occurrence and Spatial Distribution of 158 Pharmaceuticals, Drugs of Abuse and Related Metabolites in Offshore Seawater. Science of the Total Environment 2016. [Google Scholar] [CrossRef]
- Lolić, A.; Paíga, P.; Santos, L.H.M.L.M.; Ramos, S.; Correia, M.; Delerue-Matos, C. Assessment of Non-Steroidal Anti-Inflammatory and Analgesic Pharmaceuticals in Seawaters of North of Portugal: Occurrence and Environmental Risk. Science of the Total Environment 2015, 508, 240–250. [Google Scholar] [CrossRef]
- Li, Y.; Sallach, J.B.; Zhang, W.; Boyd, S.A.; Li, H. Insight into the Distribution of Pharmaceuticals in Soil-Water-Plant Systems. Water Res 2019, 152. [Google Scholar] [CrossRef]
- Martínez-Piernas, A.B.; Plaza-Bolaños, P.; García-Gómez, E.; Fernández-Ibáñez, P.; Agüera, A. Determination of Organic Microcontaminants in Agricultural Soils Irrigated with Reclaimed Wastewater: Target and Suspect Approaches. Anal Chim Acta 2018. [Google Scholar] [CrossRef]
- Lees, K.; Fitzsimons, M.; Snape, J.; Tappin, A.; Comber, S. Pharmaceuticals in Soils of Lower Income Countries: Physico-Chemical Fate and Risks from Wastewater Irrigation. Environ Int 2016. [CrossRef] [PubMed]
- Fonseca, V.F.; Duarte, I.A.; Duarte, B.; Freitas, A.; Pouca, A.S.V.; Barbosa, J.; Gillanders, B.M.; Reis-Santos, P. Environmental Risk Assessment and Bioaccumulation of Pharmaceuticals in a Large Urbanized Estuary. Science of the Total Environment 2021, 783. [Google Scholar] [CrossRef] [PubMed]
- Gunnarsson, L.; Snape, J.R.; Verbruggen, B.; Owen, S.F.; Kristiansson, E.; Margiotta-Casaluci, L.; Österlund, T.; Hutchinson, K.; Leverett, D.; Marks, B.; et al. Pharmacology beyond the Patient – The Environmental Risks of Human Drugs. Environ Int 2019, 129, 320–332. [Google Scholar] [CrossRef]
- Jacob, R.S.; Araújo, C.V.M.; Santos, L.V. de S.; Moreira, V.R.; Lebron, Y.A.R.; Lange, L.C. The Environmental Risks of Pharmaceuticals beyond Traditional Toxic Effects: Chemical Differences That Can Repel or Entrap Aquatic Organisms. Environmental Pollution 2021, 268. [Google Scholar] [CrossRef]
- Gómez-Regalado, M. del C.; Martín, J.; Santos, J.L.; Aparicio, I.; Alonso, E.; Zafra-Gómez, A. Bioaccumulation/Bioconcentration of Pharmaceutical Active Compounds in Aquatic Organisms: Assessment and Factors Database. Science of The Total Environment 2023, 861, 160638. [Google Scholar] [CrossRef] [PubMed]
- Grabicova, K.; Grabic, R.; Fedorova, G.; Fick, J.; Cerveny, D.; Kolarova, J.; Turek, J.; Zlabek, V.; Randak, T. Bioaccumulation of Psychoactive Pharmaceuticals in Fish in an Effluent Dominated Stream. Water Res 2017. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Lu, G.; Yan, Z.; Liu, J.; Wang, P.; Wang, Y. Bioaccumulation and Trophic Transfer of Pharmaceuticals in Food Webs from a Large Freshwater Lake. Environmental Pollution 2017. [Google Scholar] [CrossRef] [PubMed]
- Sengar, A.; Vijayanandan, A. Human Health and Ecological Risk Assessment of 98 Pharmaceuticals and Personal Care Products (PPCPs) Detected in Indian Surface and Wastewaters. Science of the Total Environment 2022, 807. [Google Scholar] [CrossRef] [PubMed]
- Teran-Velasquez, G.; Helm, B.; Krebs, P. High Spatiotemporal Model-Based Tracking and Environmental Risk-Exposure of Wastewater-Derived Pharmaceuticals across River Networks in Saxony, Germany. Water (Switzerland) 2023, 15. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: Eustress and Distress in Redox Homeostasis. In Stress: Physiology, Biochemistry, and Pathology Handbook of Stress Series, Volume 3; 2019.
- Singh, A.K.; Rana, H.K.; Pandey, A.K. The Oxidative Stress: Causes, Free Radicals, Targets, Mechanisms, Affected Organs, Effects, Indicators. In Antioxidants Effects in Health: The Bright and the Dark Side; 2022.
- Freitas, J.S. Pollutants and Oxidative Stress in Tadpoles. In Toxicology of Amphibian Tadpoles; 2024.
- El-SiKaily, A.; Shabaka, S. Biomarkers in Aquatic Systems: Advancements, Applications and Future Directions. Egypt J Aquat Res 2024, 50, 169–182. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlahogianni, T.; Dassenakis, M.; Scoullos, M. Molecular Biomarkers of Oxidative Stress in Aquatic Organisms in Relation to Toxic Environmental Pollutants. Ecotoxicol Environ Saf 2006, 64. [Google Scholar] [CrossRef]
- Klran, T.R.; Otlu, O.; Karabulut, A.B. Oxidative Stress and Antioxidants in Health and Disease. Journal of Laboratory Medicine 2023, 47. [Google Scholar]
- Kisty, E.A.; Falco, J.A.; Weerapana, E. Redox Proteomics Combined with Proximity Labeling Enables Monitoring of Localized Cysteine Oxidation in Cells. Cell Chem Biol 2023, 30. [Google Scholar] [CrossRef]
- Chiorcea-Paquim, A.M. 8-Oxoguanine and 8-Oxodeoxyguanosine Biomarkers of Oxidative DNA Damage: A Review on HPLC–ECD Determination. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Castillo-González, C.; Barcenilla, B.B.; Young, P.G.; Hall, E.; Shippen, D.E. Quantification of 8-OxoG in Plant Telomeres. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Lee, T.H. Antioxidant Enzymes as Redox-Based Biomarkers: A Brief Review. BMB Rep 2015, 48. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, S.; Singh, S.; Sharma, P. Role of Antioxidants in Neutralizing Oxidative Stress. Nutraceutical Fruits and Foods for Neurodegenerative Disorders 2024, 353–378. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alexandria Journal of Medicine 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Stephenie, S.; Chang, Y.P.; Gnanasekaran, A.; Esa, N.M.; Gnanaraj, C. An Insight on Superoxide Dismutase (SOD) from Plants for Mammalian Health Enhancement. J Funct Foods 2020, 68, 103917. [Google Scholar] [CrossRef]
- Jing, M.; Han, G.; Wan, J.; Zhang, S.; Yang, J.; Zong, W.S.; Niu, Q.; Liu, R. Catalase and Superoxide Dismutase Response and the Underlying Molecular Mechanism for Naphthalene. Science of The Total Environment 2020, 736, 139567. [Google Scholar] [CrossRef]
- Flohé, L.; Toppo, S.; Orian, L. The Glutathione Peroxidase Family: Discoveries and Mechanism. Free Radic Biol Med 2022, 187, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, C.; Schueler, F.W.; Rollinson, N. A 91% Decline in a Common Anuran in an Otherwise Stable Amphibian Community Inferred from 17 Years of Rapid Road Surveys. Anim Conserv 2024, 27. [Google Scholar] [CrossRef]
- Campbell Grant, E.H.; Amburgey, S.M.; Gratwicke, B.; Acosta-Chaves, V.; Belasen, A.M.; Bickford, D.; Brühl, C.A.; Calatayud, N.E.; Clemann, N.; Clulow, S.; et al. Priority Research Needs to Inform Amphibian Conservation in the Anthropocene. Conserv Sci Pract 2023, 5. [Google Scholar] [CrossRef]
- Johnson, M.S.; Aubee, C.; Salice, C.J.; Leigh, K.B.; Liu, E.; Pott, U.; Pillard, D. A Review of Ecological Risk Assessment Methods for Amphibians: Comparative Assessment of Testing Methodologies and Available Data. Integr Environ Assess Manag 2017, 13. [Google Scholar] [CrossRef]
- Seneviratne, A.; Dissanayake, M.R.; Sumanasekara, V. Review on Use of Amphibian Taxa as a Bio-Indicator for Watershed Health and Stresses. Environmental Studies and Services Division, NBRO 2015.
- Melvin, S.D. Oxidative Stress, Energy Storage, and Swimming Performance of Limnodynastes Peronii Tadpoles Exposed to a Sub-Lethal Pharmaceutical Mixture throughout Development. Chemosphere 2016, 150, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Falfushynska, H.; Gnatyshyna, L.; Horyn, O.; Sokolova, I.; Stoliar, O. Endocrine and Cellular Stress Effects of Zinc Oxide Nanoparticles and Nifedipine in Marsh Frogs Pelophylax Ridibundus. Aquatic Toxicology 2017, 185. [Google Scholar] [CrossRef]
- Falfushynska, H.I.; Gnatyshyna, L.L.; Horyn, O.; Stoliar, O.B. Vulnerability of Marsh Frog Pelophylax Ridibundus to the Typical Wastewater Effluents Ibuprofen, Triclosan and Estrone, Detected by Multi-Biomarker Approach. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2017, 202, 26–38. [Google Scholar] [CrossRef]
- Peltzer, P.M.; Lajmanovich, R.C.; Attademo, A.M.; Junges, C.M.; Teglia, C.M.; Martinuzzi, C.; Curi, L.; Culzoni, M.J.; Goicoechea, H.C. Ecotoxicity of Veterinary Enrofloxacin and Ciprofloxacin Antibiotics on Anuran Amphibian Larvae. Environ Toxicol Pharmacol 2017, 51. [Google Scholar] [CrossRef] [PubMed]
- Peltzer, P.M.; Lajmanovich, R.C.; Martinuzzi, C.; Attademo, A.M.; Curi, L.M.; Sandoval, M.T. Biotoxicity of Diclofenac on Two Larval Amphibians: Assessment of Development, Growth, Cardiac Function and Rhythm, Behavior and Antioxidant System. Science of the Total Environment 2019, 683. [Google Scholar] [CrossRef] [PubMed]
- Cuzziol Boccioni, A.P.; Peltzer, P.M.; Martinuzzi, C.S.; Attademo, A.M.; León, E.J.; Lajmanovich, R.C. Morphological and Histological Abnormalities of the Neotropical Toad, Rhinella Arenarum (Anura: Bufonidae) Larvae Exposed to Dexamethasone. J Environ Sci Health B 2020, 56. [Google Scholar] [CrossRef]
- Fernández, L.P.; Brasca, R.; Attademo, A.M.; Peltzer, P.M.; Lajmanovich, R.C.; Culzoni, M.J. Bioaccumulation and Glutathione S-Transferase Activity on Rhinella Arenarum Tadpoles after Short-Term Exposure to Antiretrovirals. Chemosphere 2020, 246. [Google Scholar] [CrossRef]
- da Luz, T.M.; Araújo, A.P. da C.; Estrela, F.N.; Braz, H.L.B.; Jorge, R.J.B.; Charlie-Silva, I.; Malafaia, G. Can Use of Hydroxychloroquine and Azithromycin as a Treatment of COVID-19 Affect Aquatic Wildlife? A Study Conducted with Neotropical Tadpole. Science of The Total Environment 2021, 780, 146553. [Google Scholar] [CrossRef]
- Fogliano, C.; Motta, C.M.; Venditti, P.; Fasciolo, G.; Napolitano, G.; Avallone, B.; Carotenuto, R. Environmental Concentrations of a Delorazepam-Based Drug Impact on Embryonic Development of Non-Target Xenopus Laevis. Aquatic Toxicology 2022, 250. [Google Scholar] [CrossRef]
- Lourido, M.; Peluso, J.; Aronzon, C.M. Lethal and Sublethal Effects of the Emerging Contaminant Oxytetracycline on the Embryo-Larval Development of Rhinella Arenarum. Environ Toxicol Pharmacol 2022, 89, 103783. [Google Scholar] [CrossRef]
- Rutkoski, C.F.; Grott, S.C.; Israel, N.G.; Carneiro, F.E.; de Campos Guerreiro, F.; Santos, S.; Horn, P.A.; Trentini, A.A.; Barbosa da Silva, E.; Coelho de Albuquerque, C.A.; et al. Hepatic and Blood Alterations in Lithobates Catesbeianus Tadpoles Exposed to Sulfamethoxazole and Oxytetracycline. Chemosphere 2022, 307. [Google Scholar] [CrossRef] [PubMed]
- Gavrilović, B.R.; Despotović, S.G.; Petrović, T.G.; Radovanović, T.B.; Gavrić, J.P.; Mirč, M.; Anđelković, M.; Vukov, T.; Tomašević Kolarov, N.; Prokić, M.D. Does the Anesthetic Tricaine Methanesulfonate (MS-222) Distort Oxidative Status Parameters in Tadpoles? Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2024, 278, 109859. [Google Scholar] [CrossRef]
- Laçin, C.; Turhan, D.O.; Güngördü, A. Assessing the Impact of Antiviral Drugs Commonly Utilized during the COVID-19 Pandemic on the Embryonic Development of Xenopus Laevis. J Hazard Mater 2024, 472, 134462. [Google Scholar] [CrossRef]
- Peluso, J.; Martínez Chehda, A.; Olivelli, M.S.; Aronzon, C.M. Ecotoxicological Effects of the Emerging Contaminant Ivermectin on Rhinella Arenarum: A Comparative Study of Active Ingredient and Commercial Formulation. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2024, 283, 109965. [Google Scholar] [CrossRef]
- Peluso, J.; Chehda, A.M.; Aronzon, C.M. A Multi-Approach Analysis of the Toxicity of a Commercial Formulation of Monensin on Rhinella Arenarum Embryos and Larvae. Environ Toxicol Pharmacol 2024, 108, 104454. [Google Scholar] [CrossRef]
- Rutkoski, C.F.; Grott, S.C.; Israel, N.G.; Guerreiro, F. de C.; Carneiro, F.E.; Bitschinski, D.; Warsneski, A.; Horn, P.A.; Lima, D.; Bastolla, C.L.V.; et al. Prednisone and Prednisolone Effects on Development, Blood, Biochemical and Histopathological Markers of Aquarana Catesbeianus Tadpoles. Aquatic Toxicology 2024, 268. [Google Scholar] [CrossRef]
- Alishiri, M.; Abdollahi, S.A.; Neysari, A.N.; Ranjbar, S.F.; Abdoli, N.; Afsharjahanshahi, M. Removal of Ciprofloxacin and Cephalexin Antibiotics in Water Environment by Magnetic Graphene Oxide Nanocomposites; Optimization Using Response Surface Methodology. Results in Engineering 2023, 20, 101507. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, Y.; Chen, S.; Guan, X.; Zhong, Y.; Yang, Q. Occurrence, Risk Assessment, and in Vitro and in Vivo Toxicity of Antibiotics in Surface Water in China. Ecotoxicol Environ Saf 2023, 255, 114817. [Google Scholar] [CrossRef]
- Wat, C.C.Y.; Xin, X.; Lai, R.W.S.; Mao, X.; Leung, K.M.Y. Impact of Environmental Factors Changes Induced by Marine Heatwaves and Heavy Precipitation on Antibiotic Toxicity to Isochrysis Galbana: Implications for Climate Change Adaptation. Mar Pollut Bull 2024, 203, 116453. [Google Scholar] [CrossRef]
- Fu, L.; Huang, T.; Wang, S.; Wang, X.; Su, L.; Li, C.; Zhao, Y. Toxicity of 13 Different Antibiotics towards Freshwater Green Algae Pseudokirchneriella Subcapitata and Their Modes of Action. Chemosphere 2017, 168, 217–222. [Google Scholar] [CrossRef]
- Martins, A.; Guimarães, L.; Guilhermino, L. Chronic Toxicity of the Veterinary Antibiotic Florfenicol to Daphnia Magna Assessed at Two Temperatures. Environ Toxicol Pharmacol 2013, 36, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Yisa, A.G.; Chia, M.A.; Gadzama, I.M.K.; Oniye, S.J.; Sha’aba, R.I.; Gauje, B. Immobilization, Oxidative Stress and Antioxidant Response of Daphnia Magna to Amoxicillin and Ciprofloxacin. Environ Toxicol Pharmacol 2023, 98, 104078. [Google Scholar] [CrossRef]
- Bawa-Allah, K.A.; Ehimiyein, A.O. Ecotoxicological Effects of Human and Veterinary Antibiotics on Water Flea (Daphnia Magna). Environ Toxicol Pharmacol 2022, 94, 103932. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Shen, Z.; Ding, J.; Qu, M.; Li, T.; Tong, M.; Di, Y. Sulfamethoxazole Induced Systematic and Tissue-Specific Antioxidant Defense in Marine Mussels (Mytilus Galloprovincialis): Implication of Antibiotic’s Ecotoxicity. Chemosphere 2021, 279, 130634. [Google Scholar] [CrossRef] [PubMed]
- Lacaze, E.; Pédelucq, J.; Fortier, M.; Brousseau, P.; Auffret, M.; Budzinski, H.; Fournier, M. Genotoxic and Immunotoxic Potential Effects of Selected Psychotropic Drugs and Antibiotics on Blue Mussel (Mytilus Edulis) Hemocytes. Environmental Pollution 2015, 202, 177–186. [Google Scholar] [CrossRef]
- Giannessi, J.; De Marchi, L.; Meucci, V.; Intorre, L.; Monni, G.; Baratti, M.; Pretti, C. Subcellular Tissue-Specific Responses of Mytilus Galloprovincialis to Fluoroquinolone Antibiotics. Environ Toxicol Pharmacol 2023, 104, 104306. [Google Scholar] [CrossRef]
- Liang, X.; Wang, F.; Li, K.; Nie, X.; Fang, H. Effects of Norfloxacin Nicotinate on the Early Life Stage of Zebrafish (Danio Rerio): Developmental Toxicity, Oxidative Stress and Immunotoxicity. Fish Shellfish Immunol 2020, 96, 262–269. [Google Scholar] [CrossRef]
- Wang, X.; Hu, M.; Gu, H.; Zhang, L.; Shang, Y.; Wang, T.; Wang, T.; Zeng, J.; Ma, L.; Huang, W.; et al. Short-Term Exposure to Norfloxacin Induces Oxidative Stress, Neurotoxicity and Microbiota Alteration in Juvenile Large Yellow Croaker Pseudosciaena Crocea. Environmental Pollution 2020, 267, 115397. [Google Scholar] [CrossRef]
- Rodrigues, S.; Antunes, S.C.; Nunes, B.; Correia, A.T. Histological Alterations in Gills and Liver of Rainbow Trout (Oncorhynchus Mykiss) after Exposure to the Antibiotic Oxytetracycline. Environ Toxicol Pharmacol 2017, 53, 164–176. [Google Scholar] [CrossRef]
- Rodrigues, S.; Antunes, S.C.; Correia, A.T.; Golovko, O.; Žlábek, V.; Nunes, B. Assessment of Toxic Effects of the Antibiotic Erythromycin on the Marine Fish Gilthead Seabream (Sparus Aurata L.) by a Multi-Biomarker Approach. Chemosphere 2019, 216, 234–247. [Google Scholar] [CrossRef]
- Zhou, Y.; Lin, J. yuan; Bian, Y.; Ren, C. jie; Xiao-li, N.; Yang, C. yu; Xiao-xue, X.; Feng, X. song Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) in the Environment: Updates on Pretreatment and Determination Methods. Ecotoxicol Environ Saf 2023, 267, 115624. [Google Scholar] [CrossRef]
- Russo, C.; Nugnes, R.; Orlo, E.; di Matteo, A.; De Felice, B.; Montanino, C.; Lavorgna, M.; Isidori, M. Diclofenac Eco-Geno-Toxicity in Freshwater Algae, Rotifers and Crustaceans. Environmental Pollution 2023, 335, 122251. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jin, M.; Mao, W.; Chen, C.; Fu, L.; Li, Z.; Du, S.; Liu, H. Photosynthetic Toxicity of Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) on Green Algae Scenedesmus Obliquus. Science of The Total Environment 2020, 707, 136176. [Google Scholar] [CrossRef] [PubMed]
- Mezzelani, M.; Gorbi, S.; Da Ros, Z.; Fattorini, D.; d’Errico, G.; Milan, M.; Bargelloni, L.; Regoli, F. Ecotoxicological Potential of Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) in Marine Organisms: Bioavailability, Biomarkers and Natural Occurrence in Mytilus Galloprovincialis. Mar Environ Res 2016, 121, 31–39. [Google Scholar] [CrossRef]
- Piedade, F.; Bio, S.; Nunes, B. Effects of Common Pharmaceutical Drugs (Paracetamol and Acetylsalicylic Acid) Short Term Exposure on Biomarkers of the Mussel Mytilus Spp. Environ Toxicol Pharmacol 2020, 73, 103276. [Google Scholar] [CrossRef] [PubMed]
- Chabchoubi, I. Ben; Bouchhima, R.A.; Louhichi, N.; Baanannou, A.; Masmoudi, S.; Hentati, O. Short-Term Effects of Various Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) on Danio Rerio Embryos. MethodsX 2023, 10, 102215. [Google Scholar] [CrossRef]
- Mikula, P.; Hollerova, A.; Hodkovicova, N.; Doubkova, V.; Marsalek, P.; Franc, A.; Sedlackova, L.; Hesova, R.; Modra, H.; Svobodova, Z.; et al. Long-Term Dietary Exposure to the Non-Steroidal Anti-Inflammatory Drugs Diclofenac and Ibuprofen Can Affect the Physiology of Common Carp (Cyprinus Carpio) on Multiple Levels, Even at “Environmentally Relevant” Concentrations. Science of The Total Environment 2024, 917, 170296. [Google Scholar] [CrossRef]
- Stancová, V.; Ziková, A.; Svobodová, Z.; Kloas, W. Effects of the Non-Steroidal Anti-Inflammatory Drug(NSAID) Naproxen on Gene Expression of Antioxidant Enzymes in Zebrafish (Danio Rerio). Environ Toxicol Pharmacol 2015, 40, 343–348. [Google Scholar] [CrossRef]
- Wang, R.; Luo, J.; Li, C.; Chen, J.; Zhu, N. Antiviral Drugs in Wastewater Are on the Rise as Emerging Contaminants: A Comprehensive Review of Spatiotemporal Characteristics, Removal Technologies and Environmental Risks. J Hazard Mater 2023, 457, 131694. [Google Scholar] [CrossRef]
- Silva, S.R.; Barbosa, F.A.R.; Mol, M.P.G.; Magalhães, S.M.S. Toxicity for Aquatic Organisms of Antiretroviral Tenofovir Disoproxil. J Environ Prot (Irvine, Calif) 2019, 10. [Google Scholar] [CrossRef]
- Godoy, A.A.; Kummrow, F.; Pamplin, P.A.Z. Occurrence, Ecotoxicological Effects and Risk Assessment of Antihypertensive Pharmaceutical Residues in the Aquatic Environment - A Review. Chemosphere 2015, 138, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Andrade de Sousa, J.; Hermes Pusceddu, F.; Dos Santos Barbosa Ortega, A.; Ueda de Carvalho, M.U. de C.; Amaral Gomes dos Santos, R.; Moledo de Souza Abessa, D.; Dias Seabra Pereira, C.; Alves Maranho, L. Biological Effects Caused by the Pharmaceuticals Losartan and Diclofenac, and Their Mixture on Marine Organisms. Ecotoxicology and Environmental Contamination 2022, 17. [Google Scholar] [CrossRef]
- Godoy, A.A.; Kummrow, F.; Pamplin, P.A.Z. Ecotoxicological Evaluation of Propranolol Hydrochloride and Losartan Potassium to Lemna Minor L. (1753) Individually and in Binary Mixtures. Ecotoxicology 2015, 24. [Google Scholar] [CrossRef]
- Reque, R.; Carneiro, R.D.; Yamamoto, F.Y.; Ramsdorf, W.A.; Martins, L.R.; Guiloski, I.C.; de Freitas, A.M. Ecotoxicity of Losartan Potassium in Aquatic Organisms of Different Trophic Levels. Environ Toxicol Pharmacol 2021, 87. [Google Scholar] [CrossRef]
- Timmermans, S.; Souffriau, J.; Libert, C. A General Introduction to Glucocorticoid Biology. Front Immunol 2019, 10. [Google Scholar] [CrossRef]
- Hardy, R.S.; Raza, K.; Cooper, M.S. Therapeutic Glucocorticoids: Mechanisms of Actions in Rheumatic Diseases. Nat Rev Rheumatol 2020, 16. [Google Scholar] [CrossRef]
- Hamilton, C.M.; Winter, M.J.; Margiotta-Casaluci, L.; Owen, S.F.; Tyler, C.R. Are Synthetic Glucocorticoids in the Aquatic Environment a Risk to Fish? Environ Int 2022, 162, 107163. [Google Scholar] [CrossRef]
- Mezzelani, M.; Peruzza, L.; d’Errico, G.; Milan, M.; Gorbi, S.; Regoli, F. Mixtures of Environmental Pharmaceuticals in Marine Organisms: Mechanistic Evidence of Carbamazepine and Valsartan Effects on Mytilus Galloprovincialis. Science of The Total Environment 2023, 860, 160465. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Soldado, N.; Mora-Medina, R.; Lora-Benítez, A.J.; Gonçalves Reis, L. de P.; Molina-López, A.M.; Moyano-Salvago, M. del R. Comparative Study of Tricaine Methanesulfonate (MS-222) and Eugenol as Euthanasia Agents in Zebrafish (Danio Rerio) as an Experimental Model. Lab Anim 2023, 57. [Google Scholar] [CrossRef]
- Balko, J.A.; Posner, L.P.; Chinnadurai, S.K. Immersion in Tricaine Methanesulfonate (MS-222) Is Not Sufficient for Euthanasia of Smokey Jungle Frogs (Leptodactylus Pentadactylus). Journal of Zoo and Wildlife Medicine 2019, 50. [Google Scholar] [CrossRef]
- Engin, E. GABAA Receptor Subtypes and Benzodiazepine Use, Misuse, and Abuse. Front Psychiatry 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Soyka, M.; Wild, I.; Caulet, B.; Leontiou, C.; Lugoboni, F.; Hajak, G. Long-Term Use of Benzodiazepines in Chronic Insomnia: A European Perspective. Front Psychiatry 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Fogliano, C.; Carotenuto, R.; Panzuto, R.; Spennato, V.; De Bonis, S.; Simoniello, P.; Raggio, A.; Avallone, B.; Agnisola, C.; Motta, C.M. Behavioral Alterations and Gills Damage in Mytilus Galloprovincialis Exposed to an Environmental Concentration of Delorazepam. Environ Toxicol Pharmacol 2023, 97, 104030. [Google Scholar] [CrossRef]
- Lebreton, M.; Sire, S.; Carayon, J.L.; Malgouyres, J.M.; Vignet, C.; Géret, F.; Bonnafé, E. Low Concentrations of Oxazepam Induce Feeding and Molecular Changes in Radix Balthica Juveniles. Aquatic Toxicology 2021, 230, 105694. [Google Scholar] [CrossRef]
- Cerveny, D.; Brodin, T.; Cisar, P.; McCallum, E.S.; Fick, J. Bioconcentration and Behavioral Effects of Four Benzodiazepines and Their Environmentally Relevant Mixture in Wild Fish. Science of The Total Environment 2020, 702, 134780. [Google Scholar] [CrossRef]
- Pohl, J.; Ahrens, L.; Carlsson, G.; Golovko, O.; Norrgren, L.; Weiss, J.; Örn, S. Embryotoxicity of Ozonated Diclofenac, Carbamazepine, and Oxazepam in Zebrafish (Danio Rerio). Chemosphere 2019. [Google Scholar] [CrossRef] [PubMed]
- C. Campbell, W. History of Avermectin and Ivermectin, with Notes on the History of Other Macrocyclic Lactone Antiparasitic Agents. Curr Pharm Biotechnol 2012, 13. [CrossRef]
- Sulik, M.; Antoszczak, M.; Huczyński, A.; Steverding, D. Antiparasitic Activity of Ivermectin: Four Decades of Research into a “Wonder Drug. ” Eur J Med Chem 2023, 261, 115838. [Google Scholar] [CrossRef] [PubMed]
- Conforti, S.; Dietrich, J.; Kuhn, T.; Koppenhagen, N. van; Baur, J.; Rohner, P.T.; Blanckenhorn, W.U.; Schäfer, M.A. Comparative Effects of the Parasiticide Ivermectin on Survival and Reproduction of Adult Sepsid Flies. Ecotoxicol Environ Saf 2018, 163, 215–222. [Google Scholar] [CrossRef]
- Garric, J.; Vollat, B.; Duis, K.; Péry, A.; Junker, T.; Ramil, M.; Fink, G.; Ternes, T.A. Effects of the Parasiticide Ivermectin on the Cladoceran Daphnia Magna and the Green Alga Pseudokirchneriella Subcapitata. Chemosphere 2007, 69, 903–910. [Google Scholar] [CrossRef]
- González-Tokman, D.; Martínez M., I.; Villalobos-Ávalos, Y.; Munguía-Steyer, R.; Ortiz-Zayas, M. del R.; Cruz-Rosales, M.; Lumaret, J.P. Ivermectin Alters Reproductive Success, Body Condition and Sexual Trait Expression in Dung Beetles. Chemosphere 2017, 178, 129–135. [Google Scholar] [CrossRef]
- Powrie, Y.; Strydom, M.; Aucamp, M.; Schellack, N.; Steenkamp, V.; Smith, C. Zebrafish Behavioral Response to Ivermectin: Insights into Potential Neurological Risk. Med Drug Discov 2022, 16, 100141. [Google Scholar] [CrossRef]
- Lorente, C.J.; Mesa, L.; Montalto, L.; Gutiérrez, M.F.; Miró, M.V.; Lifschitz, A. Ivermectin Bioaccumulation and Transfer through Developmental Stages in Culex Pipiens (Diptera: Culicidae). Chemosphere 2023, 322, 138106. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mechanism and Main Consequences of Oxidative Stress at the Cellular Level.
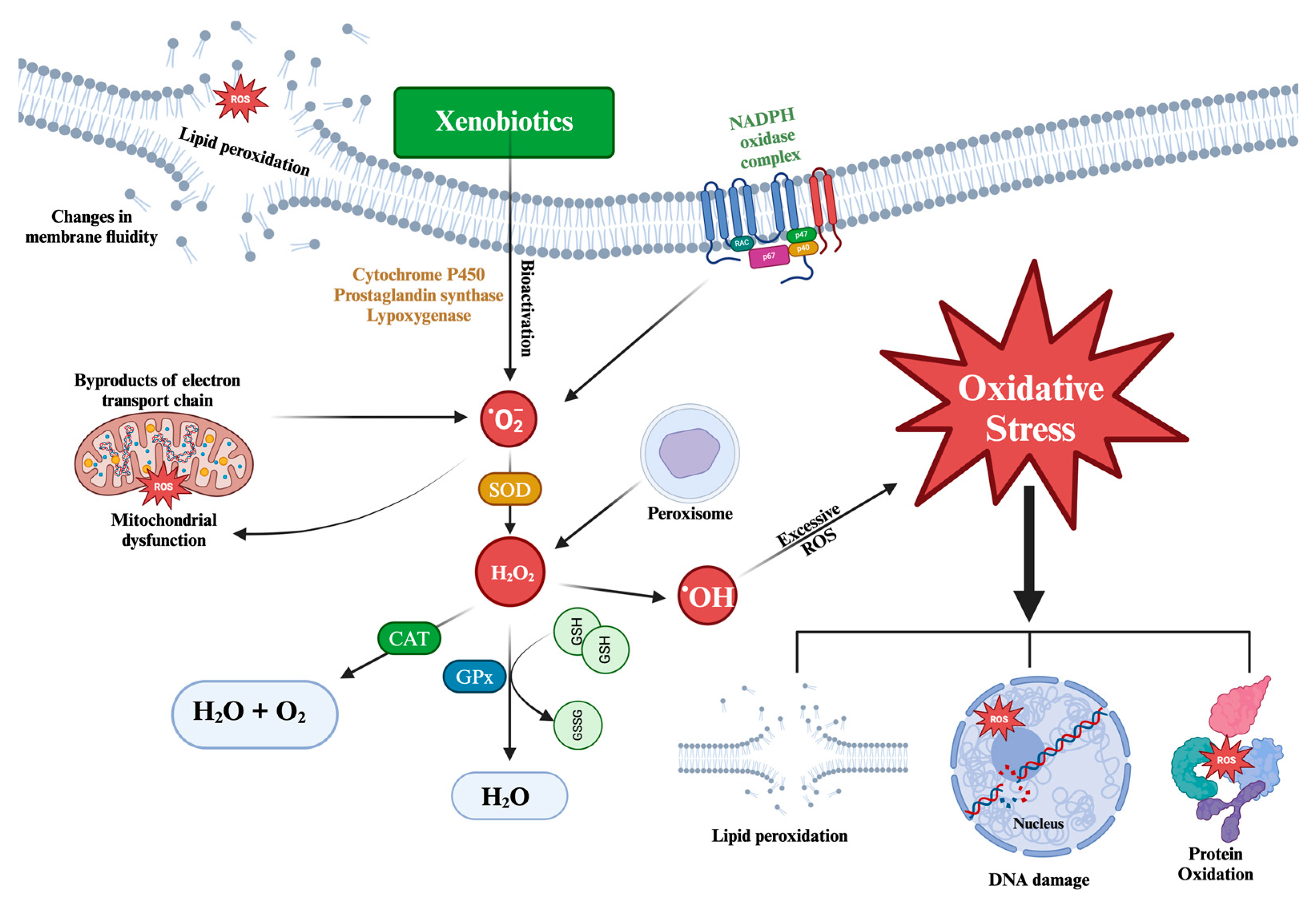

Table 1.
Effect of pharmaceutical products on biomarkers of oxidative stress in amphibians.
| Specie | Pharmaceutical | Concentration | Time of exposure | Main findings | References |
|---|---|---|---|---|---|
| Limnodynastes peronii | Diclofenac Naproxen Atenolol Gemfibrozil |
0.1, 1, 10 and 100 μgL-1 | 30 days | A significant increase in peroxidase activity was observed at the highest concentration of the drug mixture. | [62] |
| Pelophylax ridibundus | Nifedipine | 10 μM | 14 days | Increased ROS production, elevated SOD activity, and higher GSH and GSSG levels. | [63] |
| Pelophylax ridibundus | Ibuprofen Estrone |
250 100 ngL-1 |
14 days | Exposure can induce oxidative stress, although the magnitude of this effect varies depending on the compound. | [64] |
| Rhinella arenarum | Enrofloxacin Ciprofloxacin |
1, 10, 100 and 1000 μgL-1 | 96 h | An increase in LPO, decrease in CAT activity, and increase in GST activity was observed, particularly at the highest exposure concentrations. | [65] |
|
Trachycephalus typhonius Physalaemus albonotatus |
Diclofenac | 125 to 4000 μgL-1 125 to 2000 μgL-1 |
96 h 22 and 20 days |
An imbalance between ROS production and antioxidant systems was observed in both species, whereas GST activity exhibited interspecies variation. | [66] |
| Rhinella arenarum | Dexamethasone | 1-1000μgL-1 | 22 days | GST activity significantly increased in larvae exposed to the drug. | [67] |
| Rhinella arenarum | Lamivudine Stavudine Zidovudine Nevirapine |
0.5, 1, 2 and 4 μgmL-1 | 48 h | Biochemical imbalance between ROS production and induction of antioxidant systems. | [68] |
| Physalaemus cuvieri | Hydroxychloroquine Azithromycin |
12.5 μgL-1 | 72 h | Exposure to drugs did not elicit a significant oxidative stress response in tadpoles, potentially because of the activity of antioxidant enzymes. | [69] |
| Xenopus laevis | Delorazepam | 1, 5 and 10 μgL-1 | 96 h | Delorazepam alters redox equilibrium in embryos, potentially resulting in adverse effects on their development and viability. | [70] |
| Rhinella arenarum | Oxytetracycline | 10, 30 and 60 mgL-1 | 96 h | Exposure induced oxidative stress in both embryos and larvae, as evidenced by increased lipoperoxidation and altered antioxidant enzyme activities. | [71] |
| Lithobates catesbeianus | Sulfamethoxazole Oxytetracycline |
20, 90 and 460 ngL-1 | 16 days | Drug exposure induced OS in tadpoles as evidenced by the inhibition of antioxidant enzymes and increased oxidative damage to proteins. | [72] |
| Hyla arborea | Ethyl 3-aminobenzoate methanesulfonate (MS-222) | 0.1, 1 and 5 gL-1 | 15 min | MS-222 may potentially interfere with investigations of OS biomarkers, particularly those associated with GSH. | [73] |
| Xenopus laevis | Favipiravir Oseltamivir |
32.9 to 250 mgL-1 8.2 to 62.5 mgL-1 |
96 h | Biomarker responses indicate distinct detoxification and oxidative stress processes during organogenesis and the subsequent developmental stages. | [74] |
| Rhinella arenarum | Ivermectin | 1.25, 10 and 100 μgL-1 | 96 h | Induced OS, even at low concentrations, and the commercial formulation may exhibit higher toxicity than the active ingredient alone. | [75] |
| Rhinella arenarum | Monensin | 4, 12 and 120 μgL-1 | 96 h | A decrease in GST activity and GSH levels was observed, which was accompanied by an increase in TBARS levels. | [76] |
| Aquarana catesbeianus | Prednisone Prednisolone |
0.1, 1 and 10 μgL-1 | 16 days | Elevated SOD, CAT, GPx, and GST activities as well as increased MDA levels were observed in tadpoles exposed to prednisone. | [77] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



