
Submitted:
23 September 2024
Posted:
23 September 2024
You are already at the latest version
Abstract
This article presents the results of isolating and identifying EPF species and screening thermotolerant and highly virulent EPF strains that are promising for controlling Colorado potato beetle populations. Soil samples were collected from southern Kazakhstan for the isolation of fungi used in the experiments. Using the insect bait method, 41 isolates of EPF were obtained from the soils of southern Kazakhstan. PCR analysis based on ITS and TEF loci (elongation factor Ef1a) showed that all the isolates belong to the species B. bassiana. However, analysis of the ITS locus revealed two cryptic species: B. bassiana and B. pseudobassiana. No other fungal species were detected, likely due to the insufficient informativeness of the ITS and TEF loci in distinguishing species. Further studies with other loci are required. As a result of selecting natural isolates for virulence and thermotolerance, the following strains were identified: B13, B14, B15, B22, B23, B24, B25, B23-23, Bc4, Bc7, and Bc8. These strains showed high virulence (100% on the 11th day after treatment) and tolerance to suboptimal temperatures (+15°C and +30°C). Therefore, they are considered the most promising for use in the context of climate change. The data suggest that, despite the source of the isolates, EPF have great potential for controlling Colorado potato beetle populations, as the majority of isolates lead to fatal outcomes. Their geographical origin primarily influences their temperature preferences, which negatively affect their effectiveness in field conditions.
Keywords:
Colorado potato beetle
; climate change
; biological plant protection
; entomopathogenic fungi
; virulence
; thermotolerance
; B. bassiana
; B. pseudobassiana
1. Introduction
The Colorado potato beetle (Leptinotarsa decemlineata Say) was first described in 1824 and initially named the nightshade leaf beetle. It now occupies a special place among the phytophagous insects that damage cultivated plants [1]. It is one of the most striking examples of zoological species that have thrived in artificial ecosystems, illustrating the biological features of harmful insect phytophages under conditions of anthropogenic agroecosystem transformation [2].
This beetle became widely known primarily due to its “ecological explosion”, with an unprecedented territorial expansion in both rate and scale. This began in the mid-19th century when the previously harmless leaf beetle shifted from wild thorny nightshades to cultivated potatoes [3]. It causes significant damage to potatoes and eggplants (Solanum species), and, to a lesser extent, tomatoes (Lycopersicon species), which are primarily consumed by adults. The beetle is also potentially dangerous to pepper, physalis, melon pear, and tobacco crops [4].
The beetle’s preference for potatoes over other nightshade crops results in an increased concentration of pests in potato fields, higher female fertility when feeding on potato leaves, and high larval survival rates during development. As a result, the pest remains a major scourge of potato farming in many countries, including Kazakhstan [5].
In Kazakhstan, the Colorado potato beetle was first detected in 1973 in the Atyrau region, in 1977 in the Aktobe region, in 1978 in the Kostanay region, and in 1989 in the Almaty region. Frequent stormy winds contributed to the widespread dispersal of the pest in the summer. For example, in 1982, a stormy wind from the west spread the Colorado potato beetle across one-quarter of the North Kazakhstan region. In 1986, pest outbreaks were recorded in potato fields in Zhezkazgan and Karaganda, and in 1988-89 in the Almaty, Zhambyl, and Chimkent regions. By the early 1990s, the Colorado potato beetle had spread throughout all regions and had become the most dangerous pest of potato crops. Since then, many areas of the country have seen massive pest population outbreaks, leading to catastrophic potato crop losses and making the issue of protecting potatoes from this pest a matter of national importance [6].
This pest has high economic significance due to its high voracity and fertility [1,7]. Additionally, its extreme ecological plasticity, determined by genetic and physiological polymorphism, allows the Colorado potato beetle to easily adapt to environmental changes and technological impacts, particularly pesticides, while maintaining high potential viability and reproduction rates [8].
The need for phytosanitation is exacerbated by climate change, which significantly alters the pest’s behavior and distribution. As a result, scientists have begun to model the possible impacts of climate change on the pest and describe potential consequences: 1) Global warming may allow the Colorado potato beetle to spread to regions where it was previously not found. 2) Rising average temperatures may extend the growing season and increase the number of beetle generations per year, leading to larger population sizes. 3) The beetles may adapt their behavior in response to climate change, such as changing their feeding locations or seeking shade during particularly hot days to avoid heat stress. 4) Variations in rainfall amounts and distribution may affect plant health, making them more susceptible to beetle damage. For example, plants may be less resistant to pest attacks in drought conditions [9,10,11,12,13,14,15,16,17].
These impacts highlight the importance of developing integrated and sustainable pest control strategies in the context of climate change. Currently, the Colorado potato beetle is primarily controlled using chemical and agrotechnical methods [18]. The list of insecticides approved for use on potatoes for Colorado potato beetle control is quite extensive, but it is well known that chemical methods have a number of significant drawbacks, the most important of which are environmental pollution and the accumulation of initial compounds, metabolites, and degradation products in soil, water, the atmosphere, and living organisms.
Moreover, among harmful insects, the Colorado potato beetle shows the fastest resistance to chemical insecticides, which exacerbates the problem related to anti-resistance protection strategies [19,20,21,22,23]. The negative impact of pesticides on the biosphere and the need to reduce chemical usage in agriculture have led to the search for alternative pest control methods. The most attractive alternative to chemical pesticides is natural agents (entomophages and microorganisms), which are inherently safe for humans and the environment [24,25,26].
Compared to other biological agents, entomopathogenic fungi (EPF) have the greatest potential as they cause severe epizootics in insects, more so than bacteria and viruses. Additionally, the use of EPF offers several advantages, including the following: 1) fungi are easily cultivated and can be produced in large quantities at a relatively low cost; 2) they infect insects through direct penetration of the cuticle and can therefore act as contact insecticides; 3) there are no issues related to resistance among insect populations due to the complex pathogenesis process; 4) fungi have the ability to degrade chemical pesticide residues; 5) there is potential for enhancing fungal stability in the environment to ensure long-term effectiveness; 6) fungi can colonize plants and establish symbiotic relationships as endophytes, thereby increasing plant resistance to pests and diseases; 7) a lack of impact on mammals, thus reducing environmental toxicity; and 8) compatibility with insecticides [27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44].
To date, around 1,000 species of EPF are known worldwide, many of which have adapted to a lifestyle reliant on insects [44,45,46]. They belong to various taxonomic groups such as Oomycetes, Microsporidia, Chytridiomycota, Entomophthoromycota, Basidiomycota, and Ascomycota [46]. Among these, anamorphic ascomycetes such as Beauveria, Metarhizium, Lecanicillium, and Isaria, which infect a wide range of insects and are widely distributed, have significant potential in applied work [48,49,50].
EPF have been used to control economically important pests for nearly 200 years, and today, the number of mycoinsecticides based on them exceeds 100 [49,50,51,52]. However, the total share of biopesticides on the global pesticide market does not exceed 6%, which is significantly lower than the share of chemical insecticides [53]. To expand the range of mycoinsecticides and mycoacaricides, more work is being carried out to search for supervirulent strains with a selective spectrum of action [54,55,56,57,58,59]. Special attention is paid to the ability of strains to retain their pathogenic properties under unfavorable environmental conditions [60,61,62,63,64,65].
Due to the high sensitivity of EPF to environmental conditions, their use in the field may not yield the desired effect. Their effectiveness can be improved by using biopolymers (polysaccharides, polypeptides, and proteins), which can extend the viability of conidia in the field [66,67]. Cellulose, chitosan, alginate, and starch are often used for this purpose [68,69,70,71].
In this regard, the goal of this study is to select thermotolerant and highly virulent strains of EPF that are promising for controlling Colorado potato beetle populations, as well as finding methods to preserve their viability in the sharply continental and arid climate of Kazakhstan to expand the adaptability of agriculture to climate change.
2. Materials and Methods
2.1. Sample Collection and Fungal Isolation
EPF can be isolated from insect cadavers, plants, and soil [49,52,72,73,74,75]. Since soil is the natural habitat for most species of entomopathogenic fungi, soil samples were collected from various locations in southern Kazakhstan in 2023-2024 using a soil probe to a depth of 20 cm and then placed in sterile ziplock bags. The collected soil was mechanically ground and sieved to remove foreign materials. It was then stored in a refrigerator at 4°C for further analysis.
There are two methods for isolating entomopathogenic fungi: selective media and insect baits [76,77]. The most effective method involves using susceptible larvae of Galleria mellonella (Lepidoptera: Pyralidae) and Tenebrio molitor (Coleoptera: Tenebioidae) [78,79,80,81,82,83]. Larvae of the large mealworm beetle Tenebrio molitor were used to isolate entomopathogenic fungi. From each collected soil sample, 250 grams were placed in a plastic container (with 5 replicates). Five Tenebrio molitor larvae were also added along with food to prevent premature death without infection. The containers were stored at 25°C. Every 3-4 days, the larvae were monitored for signs of mycosis to confirm infection. Infected larvae were placed in a moist chamber (at 25°C for 7 days) to promote the appearance of mycosis. When signs of mycosis appeared, conidia were collected from the surface of the larvae and transferred to Sabouraud dextrose agar for further growth. Re-seeding was performed as needed until a pure culture was obtained.
2.2. EPF Species Identification
EPF species identification was carried out using classical light microscopy and molecular genetic methods. The isolates were cultured on Sabouraud dextrose agar for 14 days [84]. Highly informative molecular markers were used in a multilocus approach to identify species of Beauveria, such as translation elongation factor-1α (TEF-1α) and internal transcribed spacer (ITS) [85,86,87,88]. The results were compared with deposited cultures in GenBank using the BLAST method via the NCBI server.
2.3. Determining the Effect of Temperature on the Radial Growth of Anamorphic Ascomycetes
To determine the effect of temperature on the radial growth intensity of fungi in the surface culture, Petri dishes with Sabouraud dextrose agar were prepared 48 hours in advance [89]. The fungi were inoculated by pricking the center of the Petri dish with an even layer of medium. The dishes were placed in an incubator at fixed temperatures (+10°C, +15°C, +20°C, +25°C, +30°C, and +35°C). For 29 days, colony measurements were taken every 2 days in two perpendicular directions (crosswise) to determine colony diameter. The experiments were repeated four times.
2.4. Determining the Viability of Conidia
The germination rate of conidia of the studied fungal strains was determined in a water droplet. Two drops of conidial suspension of the tested strain were placed on a coverslip and placed in a moist chamber (Petri dish with a damp filter) in an incubator at +20°C and +25°C. The slides were observed every 12 hours under a microscope in five fields of view for 72 hours. For greater accuracy, conidia were counted in multiple drops of the suspension, with four replicates.
2.5. Preparation of the Working Suspension and Determination of Titer Using the Goryaev Chamber
To prepare the working suspension, dry fungal biomass was scraped from the Sabouraud dextrose agar medium using a scalpel. The biomass was placed in a glass test tube, covered with 2 ml of cold sterile water, and thoroughly mixed. From this mixture, 1 ml was taken using a pipette and added to a prepared sterile test tube containing 9 ml of cold sterile water. The resulting suspension was thoroughly mixed until it became cloudy. A drop of the semi-preparative form was applied to the Goryaev chamber, and excess liquid was removed with filter paper. The spore titer in 1 ml of suspension was determined by counting the spores in large squares using the following formula:
Т—spore titer in spores/ml;
∑ spore —the sum of spores counted in large squares;
k—the number of large squares with counted spores.
If necessary, the suspension was diluted with water to the required concentration or a new batch was prepared. The prepared solution was used within 1-2 hours, as further germination of fungal conidia reduces the effectiveness of the treatment [90].
2.6. Laboratory Evaluation of Biological Activity
To determine the biological activity of EPF against Colorado potato beetle larvae, test insects without mechanical or other visible damage were placed in cages at 5 individuals per cage. Each cage was a 1000 ml plastic container covered with milling gauze. The insects were infected at room temperature. The experiments were repeated four times.
First, a control group was treated with distilled water to create identical conditions for all insects, and then the test insects were treated with a fungal suspension. A conidial titer of 1×10⁷ was used in the preliminary screening tests. The containers were inspected daily for 20 days after treatment, and dead individuals were collected. The cadavers were then placed on slides in a moist chamber to determine the cause of death and the level of fungal colonization.
Biological effectiveness was calculated using Abbott’s formula [91]:
С—percentage of pest mortality;
А—average number of individuals before treatment;
В—average number of individuals after treatment.
2.7. Statistical Analysis
Statistical analysis was performed using the analysis of variance method using Microsoft Office Excel spreadsheets and the IBM statistical application package.
3. Results
3.1. Species Composition of Entomopathogenic Fungi
To isolate entomopathogenic micromycetes, more than 150 soil samples were collected from various points of agrocenoses in the region. In laboratory conditions, larvae of the yellow mealworm (Tenebrio molitor L.) were placed in these soil samples as bait for EPF (Figure 1). After 29 days, 647 out of 865 larvae died. Signs of mycosis appeared in more than 60 larvae from different soil samples out of the 647 dead larvae. In total, 41 isolates of EPF were obtained in pure culture.
An analysis of the species composition of the isolated cultures based on morphological characteristics showed that most of the isolates belonged to the genus Beauveria. During the isolation of EPF into pure culture, the cultural and morphological features of new isolates were also examined.
It was established that all newly isolated strains grow well on Sabouraud nutrient medium. Most of the isolates exhibited morphology characteristic of the genus Beauveria, specifically round colonies of white or beige color with a darker center. The mycelium is fluffy or felt-like and white in color (Figure 2).
The size of conidia from the isolated strains was assessed to identify cryptic species. However, no significant differences in spore size were found. Therefore, PCR analysis was conducted on the ITS and TEF loci to clarify the taxonomic status of natural isolates. PCR analysis of the nuclear DNA locus tef (elongation factor EF1α) showed that all isolated strains belong to the species B. bassiana (Table 1). However, analysis of the ITS locus revealed two cryptic species: B. bassiana and B. pseudobassiana. The overwhelming majority of the isolates belong to B. bassiana, 85% (35 isolates), with B. pseudobassiana being the second most common at 15% (6 isolates). No other fungal species were detected. This is likely because their distribution is limited by specific environmental conditions or hosts.
3.2. Influence of Air Temperature on the Growth AND Viability of EPF
An important aspect of selecting promising producer strains is determining their temperature preferences. The resistance of fungal cultures to suboptimal temperatures significantly affects the efficacy of mycoinsecticides. Depending on the timing and location of application, strains with varying degrees of thermotolerance are required.
To determine the effect of temperature on the radial growth of fungi in surface culture, temperatures in the range of +10°C, +15°C, +20°C, +25°C, +30°C, and +35°C were used.
At +10°C, the best growth was shown by isolates B10, B11, B12, B24, B25, and B26. In the overall sample of cultures, almost all isolates exhibited growth at this temperature. At +15°C, the same trend was observed as at the previous temperature, but the colony diameter was significantly larger than at +10°C (Figure 3). At +20°C, on the 29th day after sowing, the maximum colony growth was observed in isolates B18, B19, B20, and Bc6. The colony diameter at this temperature ranged from 44.1 mm to 69.6 mm. For this sample of isolates, the maximum colony growth was observed at +25°C. By the end of the cultivation period—on the 29th day—the best results were shown by isolates B9, B10, B14, B18, B19, B20, B25, B29, B30, B23-13, B23-14, B23-23, Bc4, Bc5, and Bc6. The diameter of these colonies exceeded 80 mm. At +30°C, the intensity of radial growth decreased compared to +25°C. As a result, at +30°C, the maximum colony growth was shown by isolates B13, B14, B15, B16, B18, B19, B20, B22, B23, B24, B25, B26, B53, B23-2, B23-3, B23-11, B23-13, B23-14, B23-23, Bc2, Bc3, Bc4, Bc5, Bc6, Bc7, and Bc8. Therefore, in nature, even at a constant temperature of 30°C for a month, the fungus is capable of not only maintaining viability but also growing quite vigorously. At +35°C, the lowest growth intensity was observed. Analysis of the obtained data showed that the isolates with increased tolerance to this temperature include B13, B14, B15, B22, B23, B24, B25, B23-23, Bc4, Bc7, and Bc8.
The observations showed that the most favorable temperatures for the growth of entomopathogenic fungal isolates on modified Sabouraud medium were +20°C and +25°C. There were no exceptions among the isolates. However, we were interested in isolates that maintained viability at elevated temperatures. As a result, the promising isolates based on thermotolerance are B13, B14, B15, B22, B23, B24, B25, B23-23, Bc4, Bc7, and Bc8.
Temperature is one of the environmental factors that limits the widespread use of EPF as mycoinsecticides. To mitigate the impact of adverse environmental factors, sodium alginate—a biopolymer that forms a capsule to preserve fungal viability—was used. Microcapsules were obtained by adding droplets of CaCl2 solution (0.25 mL) to liquid sodium alginate. The advantages of using sodium alginate include its slow dissolution in water, forming a gel that withstands temperature changes. Therefore, it is widely used in cosmetics, medicine, and the food industry. Thus, the obtained formulation is environmentally safe for humans.
3.3. Viability of Fungal Conidia at Different Temperature Regimes
The level and rate of conidia germination were determined at two air temperatures, +20°C and +25°C, over three days with 12-hour intervals. Analysis of the obtained data showed high variability in the tested cultures regarding this indicator. The final proportion of germinated spores (after 72 hours) varied between 3.7% and 74.1% on average across the entire sample of cultures (Figure 4).
For most of the studied cultures, the most optimal temperature was +25°C for both temperature regimes. However, for several isolates, the germination rate at +20°C and +25°C was higher than that of other cultures, reaching up to 99.1%. Based on the observations, a group of isolates with increased germination rates was identified: B21, B22, B23, B24, B25, B26, B27, B28, B29, B30, Bc3, Bc4, Bc5, Bc7, and Bc8. Some of these isolates not only exhibit increased conidia germination rates but also increased radial growth; thus, they are considered thermotolerant isolates.
3.4. Assessment of the Virulence of Natural Entomopathogenic Fungal Isolates against Colorado Potato Beetle Larvae
An important aspect of evaluating the potential of producer strains is assessing their biological activity. Observations showed significant variability among the studied fungal cultures in terms of virulence (Figure 5). The analysis of the results indicates that the high-virulence group includes the following isolates: B13, B14, B15, B16, B17, B18, B19, B20, B21, B22, B23, B24, B25, B26, B27, B28, B29, B53, B23-2, B23-11, B23-13, B23-14, B23-18, B23-23, B23-25, Bc2, Bc3, Bc4, Bc5, Bc6, Bc7, and Bc8. For these isolates, the final mortality rate (on the 11th day after inoculation) ranged from 90% to 100%. The best mortality rates were observed in isolates B13, B14, B15, B22, B23, B24, B25, B23-23, Bc4, Bc7, and Bc8. The mortality rate after infection with these cultures reached 50-60% on the 7th day and up to 100% on the 11th day, while for other cultures the mortality rate at this time varied from 30% to 40% and reached up to 80-90% on the 11th day.
The assessment of fungal colonization of insect cadavers by specific isolates showed a positive correlation (r = 0.88) (Figure 6). This indicates that, as mortality increases, the level of mycosis in Colorado potato beetle larvae also increases. For most isolates, the proportion of dead individuals with clear signs of mycosis exceeded 61%.
On average, there was a trend that higher virulence of an isolate was associated with an increased level of mycosis. With 100% virulence, the level of mycosis exceeded 80%. The results indicate high effectiveness of these isolates.
4. Discussion
Climate change, soil degradation, and the indiscriminate use of chemical pesticides and fertilizers pose risks to the sustainability of food and agricultural systems. Specifically, excessive use of chemical insecticides leads to the emergence of resistant pest populations. In this context, EPF have significant potential for controlling Colorado potato beetle populations, as their complex pathogenic process reduces the likelihood of developing resistant populations. Identifying promising EPF strains involves understanding the pathogen, along with assessing the virulence, thermotolerance, and productivity of these strains.
To assess the potential of promising producer strains in continental climates, new isolates from soils in southern Kazakhstan were selected and used for experiments. The results support the notion that soil is a natural habitat for EPF [92,93,94]. Research indicates that the species richness of EPF in Kazakhstani soils is rather modest. Most of the isolated fungi belong to the genus Beauveria, which aligns with the idea that these fungi are widely distributed in nature [95,96,97]. According to Kazartsev and Lednev, B. pseudobassiana is more common in forest ecosystems, while B. bassiana is more frequently found in agricultural areas [98]. Our results are consistent with this theory, as B. bassiana predominated in the isolated cultures from the soil. However, previous studies by us showed that B. bassiana was more prevalent than B. pseudobassiana in forest ecosystems in the Czech Republic [99]. Therefore, it is not possible to definitively correlate the distribution of a specific fungal species with its geographic origin.
It is known that different fungal strains may exhibit varying degrees of tolerance to limiting abiotic environmental factors, particularly high or low temperatures. Thus, finding strains of EPF with different temperature preferences is relevant from an applied perspective, as high temperatures in continental and arid climates are a major limiting factor for the use of entomopathogenic fungi-based microbial insecticides [100]. In our experiment assessing tolerance to suboptimal temperatures, no significant differences between fungal species were found. This is a topic of considerable debate in the literature. Some authors suggest that EPF can be adapted not only to climatic zones as a whole but also to specific habitats within those zones [61]. Others state that isolates from agroecosystems are more adapted to high temperatures (+37°C), while isolates from forest and arctic regions are adapted to low temperatures (+8°C) [62]. Additionally, Lednev and colleagues report that B. pseudobassiana cultures are inclined towards temperate latitudes and forest landscapes and thus tend to be psychrophilic [101]. In contrast, B. bassiana is primarily associated with arid zones and tends to be thermophilic. Moreover, Kryukov and colleagues note that the thermotolerance of fungal isolates significantly increases from north to south [102]. Our results align with these findings, as a group of isolates capable of maintaining viability at high temperatures was identified. However, a general trend of decreased growth rate for isolates at suboptimal temperatures was observed.
To mitigate the impact of negative environmental factors, we decided to use the biopolymer sodium alginate, which is often used for encapsulation and the production of granules and nano- or microspheres containing conidia of entomopathogenic fungi. The literature describes how this polymer can enhance the characteristics of EPF by prolonging conidia survival and activity in field conditions [66,67,68,69,70,71]. Therefore, further research is planned to evaluate the effectiveness of these microcapsules in field conditions.
A common issue in scientific studies is whether the virulence of EPF depends on the source of isolation or geographic origin. Some authors suggest that the host species from which the EPF were isolated is more important than their geographic origin. They show that EPF isolated from Colorado potato beetle populations are more virulent and promising for pest control compared to strains isolated from other sources [103,104]. Conversely, other authors argue that despite the source of isolation, other entomopathogenic fungal isolates can also be lethal and considered promising for Colorado potato beetle control [105,106]. Our results support the latter view. The isolates we obtained from soils in southern Kazakhstan exhibited high virulence against Colorado potato beetles, with mortality rates ranging from 90% to 100% on the 11th day post-treatment. However, the potential of EPF in Colorado potato beetle control is still not fully understood, and efforts to improve their effectiveness are ongoing. Some studies have even experimented with using EPF in combination with nematodes to enhance efficacy, though the results are mixed [107,108]. Currently, it is clear that most EPF are highly virulent against Colorado potato beetles regardless of the source of isolation.
5. Conclusions
In this study, 41 isolates of EPF were obtained from soils in southern Kazakhstan to study the effectiveness of EPF in controlling Colorado potato beetle populations. Preliminary identification of isolates was performed using keys. Analysis of the species composition of the isolated cultures based on morphological traits revealed that most isolates belonged to the genus Beauveria. PCR analysis was conducted for the ITS and TEF loci to clarify the taxonomic status of natural isolates. PCR analysis of the nuclear DNA locus tef (elongation factor Ef1a) showed that all isolates belong to B. bassiana. However, the ITS locus analysis revealed two cryptic species: B. bassiana and B. pseudobassiana. The majority of isolated cultures were B. bassiana – 85% (35 isolates), with B. pseudobassiana being the second most common at 15% (6 isolates). No other fungal species were detected. This may be due to the ITS and TEF loci being insufficiently informative. Therefore, further research with other loci (Bloc, RPB1, RPB2, nrSSU, nrLSU) is required.
The optimum temperature for radial growth of natural fungal isolates on Sabouraud medium is between +20°C and +25°C. The highest tolerance to suboptimal temperatures (+15°C and +30°C) was observed in 17 out of the 41 studied cultures. The assessment of conidia germination rates at two temperature regimes, +20°C and +25°C, showed that +25°C was optimal for most isolates. Isolates B21, B22, B23, B24, B25, B26, B27, B28, B29, B30, Bc3, Bc4, Bc5, Bc7, and Bc8 not only exhibited increased conidia germination rates but also increased radial growth, making them the most thermotolerant isolates.
Most natural isolates of EPF exhibited high virulence against Colorado potato beetles, with mortality rates ranging from 90% to 100% on the 11th day post-treatment. However, isolates B13, B14, B15, B22, B23, B24, B25, B23-23, Bc4, Bc7, and Bc8 not only showed high virulence but also high tolerance to suboptimal temperatures. Therefore, they are considered the most promising for use in the sharply changing continental climate of Kazakhstan and in other arid zones.
Author Contributions
R.A.: project leader, main investigator, funding; M.M., Z.A., N.T., B.M., A.K.: formal analysis, statistical processing of data, assistance in methodology and in writing the article; G.A.: isolation of entomopathogenic fungi and laboratory experiments; N.A.: inoculation of larvae Colorado potato beetle and observation. All authors agree with the final version of the article.
Funding
This research has been funded by the Science Committee of the Ministry of Education and Science of the Republic of Kazakhstan (Grant № AP14972820 Study of the biodiversity of entomopathogenic fungi common in the agrocenoses of southern Kazakhstan).
Data Availability Statement
All data is presented in this article.
Acknowledgments
We thank the farmers for the opportunity to collect soil samples.
Conflicts of Interest
All authors declare no conflicts of interest.
References
- Weber, D. Colorado beetle: Pest on the move. Pesticide Outlook 2003, 14, 256–269. [Google Scholar] [CrossRef]
- Weber, D. Biological Control of Potato Insect Pests. Insect Pests of Potato. Academic Press is an imprint of Elsevier, 2012, 399-437.
- Alyokhin, A.V.; Baker, M.; Mota-Sanchez, D.; Dively, G.; Grafius, E. Colorado Potato Beetle Resistance to Insecticides. American Journal of Potato Research 2008, 85, 395–413. [Google Scholar] [CrossRef]
- Li, C.; Liu, H.; Huang, F.; Cheng, D.; Wang, J.; Zhang, Y.; Sun, J.; Guo, W. Effect of Temperature on the Occurrence and Distribution of Colorado Potato Beetle (Coleoptera: Chrysomelidae) in China. Environmental Entomology 2014, 43, 511–519. [Google Scholar] [CrossRef]
- Khairushev, E.K. Potato Colorado potato beetle and ways to combat it. Almaty, 1992, 1-6.
- Alyokhin, A.; Rondon, S.I.; Gao, Y. Insect pests of potato: global perspectives on biology and management. Academic Press, 2022, 501.
- Guo, W.; Li, C.; Ahemati, T.; Jiang, W.; Li, G.; Wu, G.; Fu, K. Colorado Potato Beetle Leptinotarsa decemlineata (Say). Biological Invasions and Its Management in China. Invading Nature - Springer Series in Invasion Ecology, Springer, Dordrecht, 2017, 11, 195-217.
- Wang, C.; Xu, H.; Pan, X. Management of Colorado potato beetle in invasive frontier areas. Journal of Integrative Agriculture 2020, 19, 360–366. [Google Scholar] [CrossRef]
- Almarinez, B.J.M.; Amalin, D.M.; Aviso, K.B.; Cabezas, H.; Lao, A.R.; Tan, R.R. Network Modeling for Post-Entry Management of Invasive Pest Species in the Philippines: The Case of the Colorado Potato Beetle, Leptinotarsa decemlineata (Say, 1824) (Coleoptera: Chrysomelidae). Insects 2023, 14, 731. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Hawthorne, D.; Qin, Y.; Pan, X.; Li, Z.; Zhu, S. Impact of climate and host availability on future distribution of Colorado potato beetle. Scientific Reports 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Liao, J.; Liu, J.; Li, C. Effects of Repeated Short-Term Heat Exposure on Life History Traits of Colorado Potato Beetle. Insects 2022, 13, 455. [Google Scholar] [CrossRef]
- Pulatov, B.; Hall, K.; Linderson, M.L.; Jönsson, A.M. Effect of climate change on the potential spread of the Colorado potato beetle in Scandinavia: an ensemble approach. Clim Res 2014, 62, 15–24. [Google Scholar] [CrossRef]
- Pulatov, B.; Jönsson, A.M.; Wilcke, R.A.; Linderson, M.; Hall, K.; Bärring, L. Evaluation of the phenological synchrony between potato crop and Colorado potato beetle under future climate in Europe. Agriculture, Ecosystems & Environment 2016, 224, 39–49. [Google Scholar]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Kroschel, J.; Mujica, N.; Okonya, J.; Alyokhin, A. Insect Pests Affecting Potatoes in Tropical, Subtropical, and Temperate Regions. The Potato Crop. Springer, Cham, 2020, 251-306.
- Skendžic ́,S. ; Zovko, M.; Živkovic ́, I.P.; Lešic ́, V.; Lemic ́, D. The Impact of Climate Change on Agricultural Insect Pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Sablon, L.; Dickens, J.C.; Haubruge, É.; Verheggen, F.J. Chemical Ecology of the Colorado Potato Beetle, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae), and Potential for Alternative Control Methods. Insects 2013, 4, 31–54. [Google Scholar] [CrossRef]
- Bastarache, P.; Timani, K.; Ben Youssef, M.; Omakele, E.; Vickruck, J.L.; Morin, P.J. Cyantraniliprole and Thiamethoxam Exposure Changes Expression of Transcripts Associated with Small Non-Coding RNA Processing in the Colorado Potato Beetle. Insects 2024, 15, 147. [Google Scholar] [CrossRef]
- Bouafoura, R.; Bastarache, P.; Ouédraogo, B.C.; Dumas, P.; Moffat, C.E.; Vickruck, J.L.; Morin, P.J. Characterization of Insecticide Response-Associated Transcripts in the Colorado Potato Beetle: Relevance of Selected Cytochrome P450s and Clothianidin. Insects 2022, 13, 505. [Google Scholar] [CrossRef] [PubMed]
- Kadoić Balaško, M.; Mikac, K.M.; Bažok, R.; Lemic, D. Modern Techniques in Colorado Potato Beetle (Leptinotarsa decemlineata Say) Control and Resistance Management: History Review and Future Perspectives. Insects 2020, 11, 581. [Google Scholar] [CrossRef]
- Kaleem Ullah, R.M.; Gökçe, A.; Bakhsh, A.; Salim, M.; Wu, H.Y.; Naqqash, M.N. Insights into the Use of Eco-Friendly Synergists in Resistance Management of Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Insects 2022, 13, 846. [Google Scholar] [CrossRef]
- Timani, K.; Bastarache, P.; Morin, P.J. Leveraging RNA Interference to Impact Insecticide Resistance in the Colorado Potato Beetle, Leptinotarsa decemlineata. Insects 2023, 14, 418. [Google Scholar] [CrossRef]
- Liu, D.; Smagghe, G.; Liu, T.-X. Interactions between Entomopathogenic Fungi and Insects and Prospects with Glycans. J. Fungi 2023, 9, 575. [Google Scholar] [CrossRef]
- Venkatesh, G.; Sakthi Priya, P.; Anithaa, V.; Dinesh, G.K.; Velmurugan, S.; Abinaya, S.; Karthika, P.; Sivasakthivelan, P.; Soni, R.; Thennarasi, A. Role of entomopathogenic fungi in biocontrol of insect pests. Plant Protection: From Chemicals to Biologicals 2022, 505-548.
- Sujeetha, J.A.R.P.; Sahayaraj, K. Role of Entomopathogenic Fungus in Pest Management. Basic and Applied Aspects of Biopesticides. Springer, New Delhi, 2014, 36-41.
- Charnley, A.K.; Collins, S.A. Entomopathogenic fungi and their role in pest control. The Mycota IV: Environmental and Microbial Relationships, 2nd edition, Springer, 2007, 159-187.
- Lacey, L.A.; Kaya, H.K. Field Manual of Techniques in Invertebrate Pathology. Application and Evaluation of Pathogens for Control of Insects and Other Invertebrate Pests, 2nd Edition, Springer, Dordrecht, 2007, 868.
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insecticide. World J Microbiol Biotechnol 2016, 32, 177. [Google Scholar] [CrossRef]
- Liu, D.; Smagghe, G.; Liu, T.X. Interactions between Entomopathogenic Fungi and Insects and Prospects with Glycans. J. Fungi 2023, 9, 575. [Google Scholar] [CrossRef] [PubMed]
- Mannino, M.C.; Pedrini, N. Is the Insect Cuticle the only Entry Gate for Fungal Infection? Insights into Alternative Modes of Action of Entomopathogenic Fungi. Journal of Fungi 2019, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lee, M.R.; Kim, S.; Kim, J.C.; Park, S.E.; Li, D.; Shin, T.Y.; Nai, S.; Kim, J.S. Genomic Analysis of the Insect-Killing Fungus Beauveria bassiana JEF-007 as a Biopesticide. Scientific Reports 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Jiang, X.; Wang, G.; Luo, Z.; Fan, Y.; Wu, Z.; Pei, Y. Requirement of a Mitogen-Activated Protein Kinase for Appressorium Formation and Penetration of Insect Cuticle by the Entomopathogenic Fungus Beauveria bassiana. Applied and Environmental Microbiology 2010, 76, 2262–2270. [Google Scholar] [CrossRef] [PubMed]
- Ahirwar, N.K.; Singh, R. Assessing the Safety and E ciency of Entomopathogenic Fungi as Bioinsec cides: A Brief Review. Journal of Earth & Environmental Waste Management 2023, 1, 1–6. [Google Scholar]
- Fernandes, E.K.K.; Bittencourt, V.R.E.P.; Roberts, D.W. Perspectives on the potential of entomopathogenic fungi in biological control of ticks. Exp Parasitol 2012, 130, 300–305. [Google Scholar] [CrossRef]
- Khan, S.; Guo, L.; Maimaiti, Y.; Mijit, M.; Qiu, D. Entomopathogenic fungi as microbial biocontrol agent. Mol Plant Breed 2012, 3, 63–79. [Google Scholar] [CrossRef]
- Valero-Jiménez, C.A.; Wiegers, H.; Zwaan, B.J.; Koenraadt, C.J.M.; van Kan, J.A.L. Genes involved in virulence of the entomopathogenic fungus Beauveria bassiana. Journal of Invertebrate Pathology 2016, 133, 41–49. [Google Scholar] [CrossRef]
- Sui, L.; Lu, Y.; Zhou, L.; Li, N.; Li, Q.; Zhang, Z. Endophytic Beauveria bassiana promotes plant biomass growth and suppresses pathogen damage by directional recruitment. Frontiers in Microbiology 2023, 14, 1227269. [Google Scholar] [CrossRef]
- González-Mas, N.; Valverde-García, R.; Gutiérrez-Sánchez, F.; Quesada-Moraga, E. Effect of passage through the plant on virulence and endophytic behavioural adaptation in the entomopathogenic fungus Beauveria bassiana. Biological Control 2021, 160, 104687. [Google Scholar] [CrossRef]
- Abidin, A.F.; Ekowati, N.; Ratnaningtyas, N.I. Insecticide compatibility to the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae. Scripta Biologica 2017, 4, 273–279. [Google Scholar] [CrossRef]
- Ondráčková, E.; Seidenglanz, M.; Šafář, J. Effect of seventeen pesticides on mycelial growth of Akanthomyces, Beauveria, Cordyceps and Purpureocillium strains. Czech Mycology 2019, 71, 123–135. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Kitsiou, F.; Natsiopoulos, D.; Eliopoulos, P.A. Entomopathogenic Fungi: Interactions and Applications. Encyclopedia 2022, 2, 646–656. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Marinho-Prado, J.S.; Assalin, M.R.; Martins, L.G.; Braga, E.S.; Tasic, L.; Dita, M.; Lopes, R.B. Natural occurrence of Beauveria caledonica, pathogenicity to Cosmopolites sordidus and antifungal activity against Fusarium oxysporum f. Sp. Cubense. Pest Management Science 2022, 78, 4458–4470. [Google Scholar] [CrossRef]
- St. Leger, R.J.; Wang, C. Genetic engineering of fungal biocontrol agents to achieve greater efficacy against insect pests. Appl.Microbiol.Biotechnol 2010, 85, 901–907. [Google Scholar] [CrossRef]
- Araújo, J.; Hughes, D. Diversity of Entomopathogenic Fungi: Which Groups Conquered the Insect Body? Advances in Genetics 2015, 94, 1–39. [Google Scholar]
- Mora, M.A.E.; Castilho, A.M.C.; Fraga, M.E. Classification and infection mechanism of entomopathogenic fungi. Arquivos Do Instituto Biológico 2017, 84, 1–10. [Google Scholar] [CrossRef]
- Bihal, R.; Al-Khayri, J.M.; Banu, A.N.; Kudesia, N.; Ahmed, F.K.; Sarkar, R.; Arora, A.; Abd-Elsalam, K.A. Entomopathogenic Fungi: An Eco-Friendly Synthesis of Sustainable Nanoparticles and Their Nanopesticide Properties. Microorganisms 2023, 11, 1617. [Google Scholar] [CrossRef]
- Humber, R. Seeking stability for research and applied uses of entomopathogenic fungi as biological control agents. J Asia Pac Entomol 2010, 19, 1019–1025. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Science and Technology 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Meyling, N.V.; Hajek, A.E. Principles from community and metapopulation ecology: application to fungal entomopathogens. The Ecology of Fungal Entomopathogens. Springer, Dordrecht, 2009, 39-54.
- Faria, M.R.D.; Wraight, S.P. Mycoinsecticides and Mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biological Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Jaronski, S.; Mascarin, G. Mass Production of Fungal Entomopathogens. Microbial Control of Insect and Mite Pests 2016, 141–155. [Google Scholar]
- Smagghe, F.; Spooner-Hart, R.; Chen, Z.; Donovan-Mak, M. Biological control of arthropod pests in protected cropping by employing entomopathogens: Efficiency, production and safety. Biological Control 2023, 186, 105337. [Google Scholar] [CrossRef]
- Wang, C.; Fan, M.; Li, Z.; Butt, T.M. Molecular monitoring and evaluation of the application of the insect-pathogenic fungus Beauveria bassiana in southeast China. Journal of Applied Microbiology 2004, 96, 861–870. [Google Scholar] [CrossRef]
- Li, Z.; Alves, S.B.; Roberts, D.W.; Fan, M.; Delalibera, I.; Tang, J.; Rangel, D.E.N. Biological control of insects in Brazil and China: history, current programs and reasons for their successes using entomopathogenic fungi. Biocontrol Science and Technology 2009, 20, 117–136. [Google Scholar] [CrossRef]
- Kryukov, V.Y.; Yaroslavtseva, O.N.; Levchenko, M.V. Phenotypic variability of environmental isolates of the entomopathogenic fungus Beauveria bassiana. Microbiology 2010, 79, 265–269. [Google Scholar] [CrossRef]
- Valero-Jiménez, C.A.; Wiegers, H.; Zwaan, B.J.; Koenraadt, C.J.; Van Kan, J.A. Genes involved in virulence of the entomopathogenic fungus Beauveria bassiana. Journal of Invertebrate Pathology 2015, 133, 41–49. [Google Scholar] [CrossRef]
- Clifton, E.H.; Castrillo, L.A.; Jaronski, S.T.; Hajek, A.E. Cryptic diversity and virulence of Beauveria bassiana recovered from Lycorma delicatula (spotted lanternfly) in eastern Pennsylvania. Front. Insect Sci 2023, 3, 1127682. [Google Scholar] [CrossRef]
- Liu, J.; Ling, Z.; Wang, J.; Xiang, T.; Xu, L.; Gu, C.; Liu, R.; Xu, J.; Xu, C.; Zhou, W.; Liu, Y.; Jin, Z.; Wan, Y. In vitro transcriptomes analysis identifies some special genes involved in pathogenicity difference of the Beauveria bassiana against different insect hosts. Microbial Pathogenesis 2021, 154, 104824. [Google Scholar] [CrossRef]
- St Leger, R.J. Integument as a Barrier to Microbial Infections. Physiology of the Insect Epidermis. Commonwealth Scientific and Industrial Research Organization, Melbourne, 1991, 286-308.
- Fargues, J.; Goettel, M.S.; Smits, N.; Ouedraogo, A.; Rougier, M. Effect of Temperature on Vegetative Growth of Beauveria bassiana Isolates from Different Origins. Mycologia 1997, 89, pp. 383–392.
- Fernandes, É.K.; Rangel, D.E.; Moraes, Á.M.; Bittencourt, V.R.; Roberts, D.W. Cold activity of Beauveria and Metarhizium, and thermotolerance of Beauveria. Journal of Invertebrate Pathology 2008, 98, 69–78. [Google Scholar] [CrossRef]
- de la Cruz Quiroz, R.; Cruz Maldonado, J.; Rostro Alanis, M. Fungi-based biopesticides: shelf-life preservation technologies used in commercial products. J Pest Sci 2019, 92, 1003–1015. [Google Scholar] [CrossRef]
- Tong, M.; Feng, G. Molecular basis and regulatory mechanisms underlying fungal insecticides’ resistance to solar ultraviolet irradiation. Pest Management Science 2021, 78, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Bayramoğlu, Z. The UV protectant properties of tea extracts on entomopathogenic fungus spores and their lethal effect on Galleria mellonella (L., 1758) (Lepidoptera: Pyralidae). Turkish Journal of Entomology 2023, 47, 363–372. [Google Scholar] [CrossRef]
- Friuli, M.; Pellegrino, R.; Lamanna, L.; Nitti, P.; Madaghiele, M.; Demitri, C. Materials Engineering to Help Pest Control: A Narrative Overview of Biopolymer-Based Entomopathogenic Fungi Formulations. J. Fungi 2023, 9, 918. [Google Scholar] [CrossRef] [PubMed]
- Holka, M.; Kowalska, J. The Potential of Adjuvants Used with Microbiological Control of Insect Pests with Emphasis on Organic Farming. Agriculture 2023, 13, 1659. [Google Scholar] [CrossRef]
- Gerding-González, M.; France, A.; Sepulveda, M.E.; Campos, J. Use of chitin to improve a Beauveria bassiana alginate-pellet formulation. Biocontrol Science and Technology 2007, 17, 105–110. [Google Scholar] [CrossRef]
- Nong, X.; Wang, G.; Zhang, X.; Liu, X.; Zhang, Z. Alginate Granule formulation promotes recovery of metarhizium anisopliae population. Chinese Journal of Biological Control 2016, 32, 762. [Google Scholar]
- Batista, D.P.C.; De Oliveira, I.N.; Ribeiro, A.R.B.; Fonseca, E.J.S.; Santos-Magalhães, N.S.; de Sena-Filho, J.G.; Dornelas, C.B. Encapsulation and release of Beauveria bassiana from alginate–bentonite nanocomposite. RSC advances 2017, 7, 26468–26477. [Google Scholar] [CrossRef]
- Shah, S.; Ash, G.J.; L. Wilson, B.A. Resporulation of Metarhizium anisopliae granules on soil and mortality of Tenebrio molitor: Implications for wireworm management in sweetpotato. Annals of Applied Biology 2022, 182, 65–76. [Google Scholar] [CrossRef]
- Keyser, C.A.; De Fine Licht, H.H.; Steinwender, B.M.; Meyling, N.V. Diversity within the entomopathogenic fungal species Metarhizium flavoviride associated with agricultural crops in Denmark. BMC Microbiology 2015, 15, 1–11. [Google Scholar] [CrossRef]
- Imoulan, A.; Hussain, M.; Kirk, P.M.; El Meziane, A.; Yao, Y. Entomopathogenic fungus Beauveria: Host specificity, ecology and significance of morpho-molecular characterization in accurate taxonomic classification. Journal of Asia-Pacific Entomology 2017, 20, 1204–1212. [Google Scholar] [CrossRef]
- Lovett, B.; St. Leger, R.J. The insect pathogens. Microbiol. Spectr 2017, 5, 923–943. [Google Scholar] [CrossRef] [PubMed]
- Imoulan, A.; Wei, X.D.; Wang, W.J. Distribution and genetic diversity of Beauveria species at different soil depths in natural and agricultural ecosystems. Mycol Progress 2019, 18, 1241–1252. [Google Scholar] [CrossRef]
- Meyling, N.V.; Eilenberg, J. Occurrence and distribution of soil borne entomopathogenic fungi within a single organic agroecosystem. Agriculture, Ecosystems & Environment 2006, 113, 336–341. [Google Scholar]
- Meyling, N.V. Methods for isolation of entomopathogenic fungi from the soil environment. Laboratory manual, Denmark, 2007, 1-18.
- Zimmermann, G. The ‘Galleria bait method’ for detection of entomopathogenic fungi in soil. Journal of Applied Entomology 1986, 102, 213–215. [Google Scholar] [CrossRef]
- Keller, S.; Kessler, P.; Schweizer, C. Distribution of insect pathogenic soil fungi in Switzerland with special reference to Beauveria brongniartii and Metharhizium anisopliae. BioControl 2003, 48, 307–319. [Google Scholar] [CrossRef]
- Montesinos, M.R.; Viniegra Gonzalez, G.; Alatorre Rosas, R.; Loera, O. Relationship between virulence and enzymatic profiles in the cuticle of Tenebrio molitor by 2-deoxy-D-glucose-resistant mutants of Beauveria bassiana (Bals.) Vuill. World J. Microbiol Biotechnol 2011, 27, 2095–2102. [Google Scholar] [CrossRef]
- Juliya, R.F. Genetic diversity of Beauveria bassiana in semi natural and agricultural habitats and its biocontrol potential against cowpea aphid, Aphis craccivora Koch. Braz J Microbiol 2019, 50, 697–704. [Google Scholar] [CrossRef]
- Karaborklu, S.; Altin, N.; Keskin, Y. Native entomopathogenic fungi isolated from Duzce, Turkey and their virulence on the mealworm beetle [Tenebrio molitor L. (Coleoptera: Tenebrionidae)]. Journal Article 2019, 4395.
- Gebremariam, A.; Chekol, Y.; Assefa, F. Phenotypic, molecular, and virulence characterization of entomopathogenic fungi, Beauveria bassiana (Balsam) Vuillemin, and Metarhizium anisopliae (Metschn.) Sorokin from soil samples of Ethiopia for the development of mycoinsecticide. Heliyon 2021, 7, e07091. [Google Scholar] [CrossRef]
- Enkerli, J.; Widmer, F. Molecular ecology of fungal entomopathogens: molecular genetic tools and their applications in population and fate studies. BioControl 2010, 55, 17–37. [Google Scholar] [CrossRef]
- Rehner, S.A. Phylogenetics of the insect pathogenic genus Beauveria. Insect-fungal associations: Ecology and evolution. Oxford University Press, Oxford, 2005, 3-27.
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Minnis, A.M.; Sung, G.H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef] [PubMed]
- Humber, R.A. Identification of entomopathogenic fungi. Manual of Techniques in Invertebrate Pathology. Cornell University, 2nd Edition, 2011, 151-187.
- Kryukov, V.Y.; Yaroslavtseva, O.N.; Elisaphenko, E.A. Change in the temperature preferences of Beauveria bassiana sensu lato isolates in the latitude gradient of Siberia and Kazakhstan. Microbiology 2012, 81, 453–459. [Google Scholar] [CrossRef]
- Makarov, E.M.; Baimagambetov, E.J.; Lednev, G.R. Methodological guidelines for assessing the biological effectiveness of strains of the entomopathogenic fungus Beauveria bassiana S.L. (Bals.) Vuill in relation to locusts. Astana, 2015, 17.
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. econ. Entomol 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Tuininga, A.R.; Miller, J.L.; Morath, S.U.; Daniels, T.J.; Falco, R.C.; Marchese, M.; Sahabi, S.; Rosa, D.; Stafford, K.C. Isolation of Entomopathogenic Fungi From Soils and Ixodes scapularis (Acari: Ixodidae) Ticks: Prevalence and Methods. Journal of Medical Entomology 2009, 46, 557–565. [Google Scholar] [CrossRef]
- Korosi, G.A.; Wilson, B.A.; Powell, K.S.; Ash, G.J.; Reineke, A.; Savocchia, S. Occurrence and diversity of entomopathogenic fungi (Beauveria spp. And Metarhizium spp.) in Australian vineyard soils. Journal of Invertebrate Pathology 2019, 164, 69–77. [Google Scholar] [CrossRef]
- Berestetskiy, A.O.; Ivanova, A.N.; Petrova, M.O. Comparative Analysis of the Biological Activity and Chromatographic Profiles of the Extracts of Beauveria bassiana and B. pseudobassiana Cultures Grown on Different Nutrient Substrates. Microbiology 2018, 87, 200–214. [Google Scholar] [CrossRef]
- Imoulan, A.; Hussain, M.; Kirk, P.M.; El Meziane, A.; Yao, Y. Entomopathogenic fungus Beauveria: Host specificity, ecology and significance of morpho-molecular characterization in accurate taxonomic classification. Journal of Asia-Pacific Entomology 2017, 20, 1204–1212. [Google Scholar] [CrossRef]
- Imoulan, A.; Wu, H.; Lu, W.; Li, Y.; Li, B.; Yang, R.; Wang, W.; Wang, X.; Kirk, P.M.; Yao, Y. Beauveria medogensis sp. Nov., a new fungus of the entomopathogenic genus from China. Journal of Invertebrate Pathology 2016, 139, 74–81. [Google Scholar] [CrossRef]
- Meyling, N.V.; Pilz, C.; Keller, S.; Widmer, F.; Enkerli, J. Diversity of Beauveria spp. isolates from pollen beetles Meligethes aeneus in Switzerland. J Invertebr Pathol 2012, 109, 76–82. [Google Scholar] [CrossRef]
- Kazartsev, I.A.; Lednev, G.R. Distribution and Diversity of Beauveria in Boreal Forests of Northern European Russia. Microorganisms 2021, 9, 1409. [Google Scholar] [CrossRef] [PubMed]
- Abdukerim, R.; Lednev, G.; Trýzna, M.; Ryšánek, P.; Zouhar, M. Comparison Analysis of Species Composition of Entomopathogenic Fungi Isolated from Bark Beetles (Coleoptera: Scolytidae). OnLine Journal of Biological Sciences 2018, 18, 62–68. [Google Scholar] [CrossRef]
- Vidal, C.; Fargues, J. Climatic constraints for fungal bioinsecticides. Use of Entomopathogenic Fungi in Biological Pest Management, 2007, 39–55.
- Lednev, G.R.; Uspanov, A.M.; Tokarev, Y.S.; Kazartsev, I.A.; Levchenko, M.V.; Sabitova, M.N.; Smagulova, S.B.; Duisembekov, B.A. Biological features of strains of two species of fungi of the genus Beauveria of northern Eurasia. Proceedings of the All-Russian conference with international participation “Biodiversity and ecology of fungi and mushroom-like organisms of Northern Eurasia”. Yekaterinburg, 2015, 139-141.
- Kryukov, V.Y.; Yaroslavtseva, O.N.; Elisaphenko, E.A. Change in the temperature preferences of Beauveria bassiana sensu lato isolates in the latitude gradient of Siberia and Kazakhstan. Microbiology 2012, 81, 453–459. [Google Scholar] [CrossRef]
- Zemek,R. ; Konopická,J.; Jozová, E.; Skoková Habuštová, O. Virulence of Beauveria bassiana Strains Isolated from Cadavers of Colorado Potato Beetle, Leptinotarsa decemlineata. Insects 2021, 12, 1077. [Google Scholar] [CrossRef] [PubMed]
- Yıldırım, K.; Eski, A.; Biryol, S. Isolation, Characterization, and Formulation of Indigenous Beauveria bassiana Fungus Against Colorado Potato Beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Potato Res 2024, 67, 583–601. [Google Scholar] [CrossRef]
- Polat, İ.; Yanar, Y.; Yanar, D. Efficacy of local entomopathogenic fungi isolated from forestlands in Tokat Province (Türkiye) against the Colorado potato beetle, Leptinotarsa decemlineata (Say, 1824) (Coleoptera: Chrysomelidae). Turkish Journal of Entomology 2022, 46, 159–173. [Google Scholar] [CrossRef]
- Kryukov, V.Y. Adaptations of entomopathogenic ascomycetes (Ascomycota, Hypocreales) host insects and environmental factors in the continental climate of western Siberia and Kazakhstan. Dissertation for the degree of Doctor of Biological Sciences. Novosibirsk, 2014, P. 249. https://www.dissercat.com/content/adaptatsii-entomopatogennykh-askomitsetov-ascomycota-hypocreales-k-nasekomym-khozyaevam-i.
- Uzun, Y.A.; Göze Özdemir, F.G.; Demirözer, O.; Nayır, T. Efficacy of single and combined applications of entomopathogenic fungi and nematodes against the pupae of colorado potato beetle (Leptinotarsa decemlineata [Say]), (Coleoptera: Chrysomelidae). Egyptian Journal of Biological Pest Control 2022, 32, 1–6. [Google Scholar] [CrossRef]
- Pu ̊ža,V. ; Nermut’,J.; Konopická, J.; Skoková Habuštová, O. Efficacy of the Applied Natural Enemies on the Survival of Colorado Potato Beetle Adults. Insects 2021, 12, 1030. [Google Scholar] [CrossRef]
Figure 1.
Soil samples with mealworm larvae (Tenebrio molitor L).

Figure 2.
Morphology of colonies of entomopathogenic micromycetes isolated from soils of agrocenoses of the Turkestan region on Sabouraud nutrient medium.
Figure 2.
Morphology of colonies of entomopathogenic micromycetes isolated from soils of agrocenoses of the Turkestan region on Sabouraud nutrient medium.
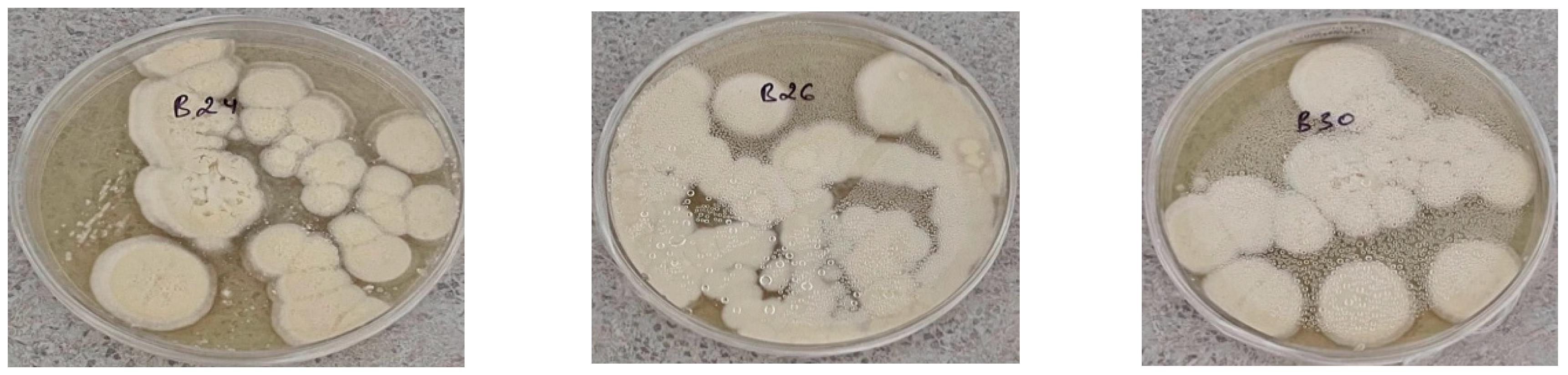
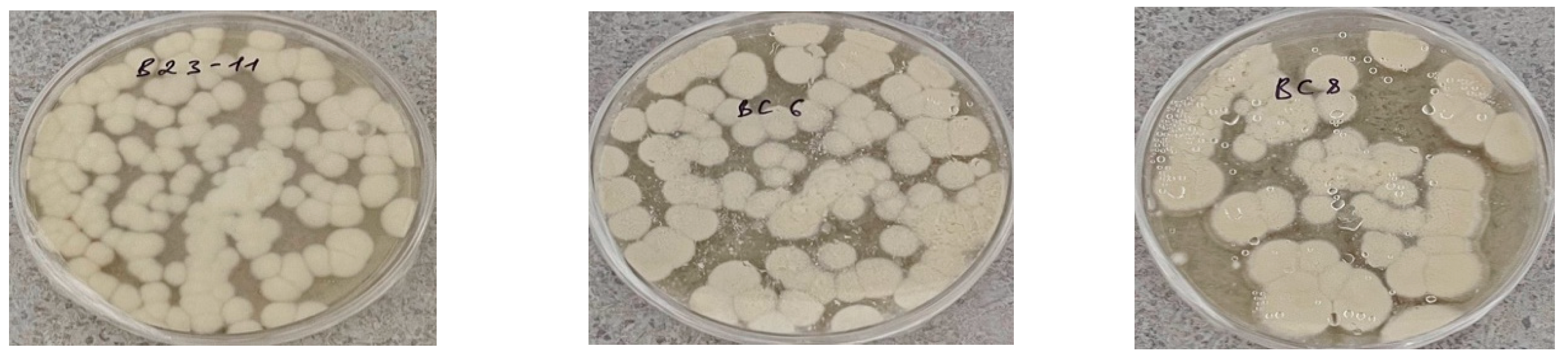
Figure 3.
Growth of colonies of natural isolates of EPF isolated from the soils of southern Kazakhstan on Sabouraud nutrient medium on the 29th day after sowing at different air temperatures.
Figure 3.
Growth of colonies of natural isolates of EPF isolated from the soils of southern Kazakhstan on Sabouraud nutrient medium on the 29th day after sowing at different air temperatures.
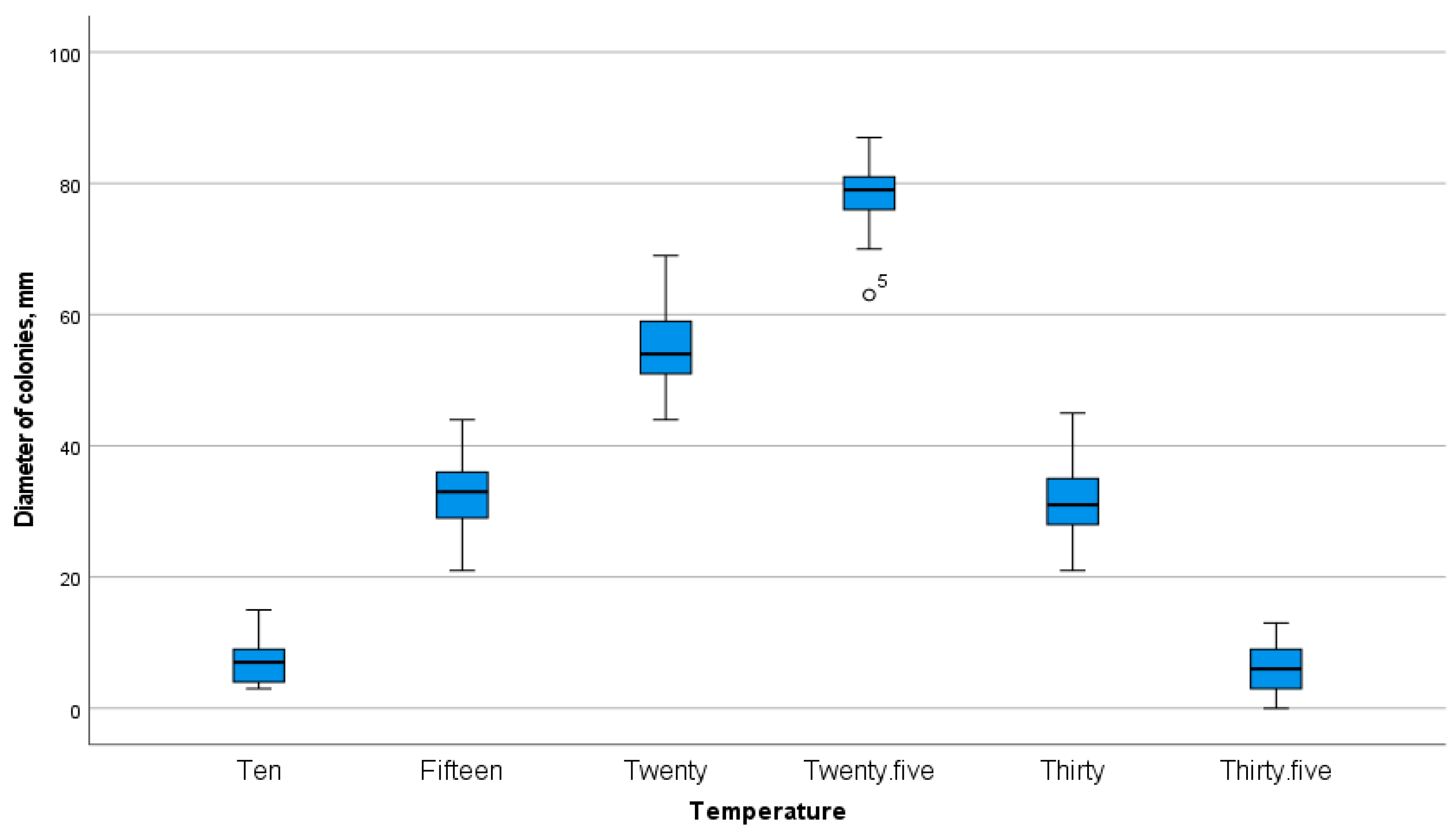
Figure 4.
Germination of conidia of natural isolates of EPF isolated from the soils of southern Kazakhstan in a drop of water at different temperatures.
Figure 4.
Germination of conidia of natural isolates of EPF isolated from the soils of southern Kazakhstan in a drop of water at different temperatures.
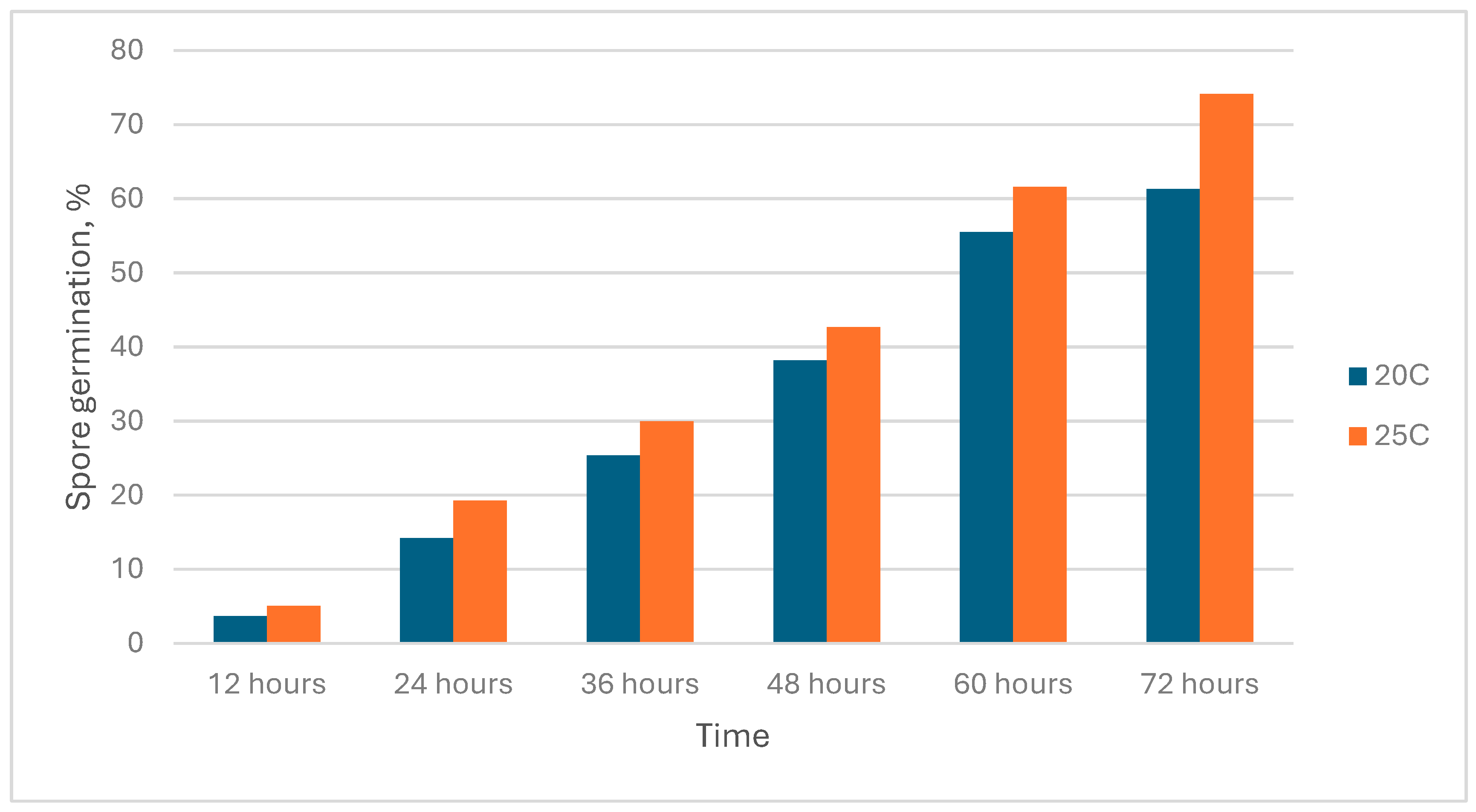
Figure 5.
Virulence of natural isolates of EPF isolated from soils of southern Kazakhstan in relation to second- and third-instar larvae of the Colorado potato beetle.
Figure 5.
Virulence of natural isolates of EPF isolated from soils of southern Kazakhstan in relation to second- and third-instar larvae of the Colorado potato beetle.
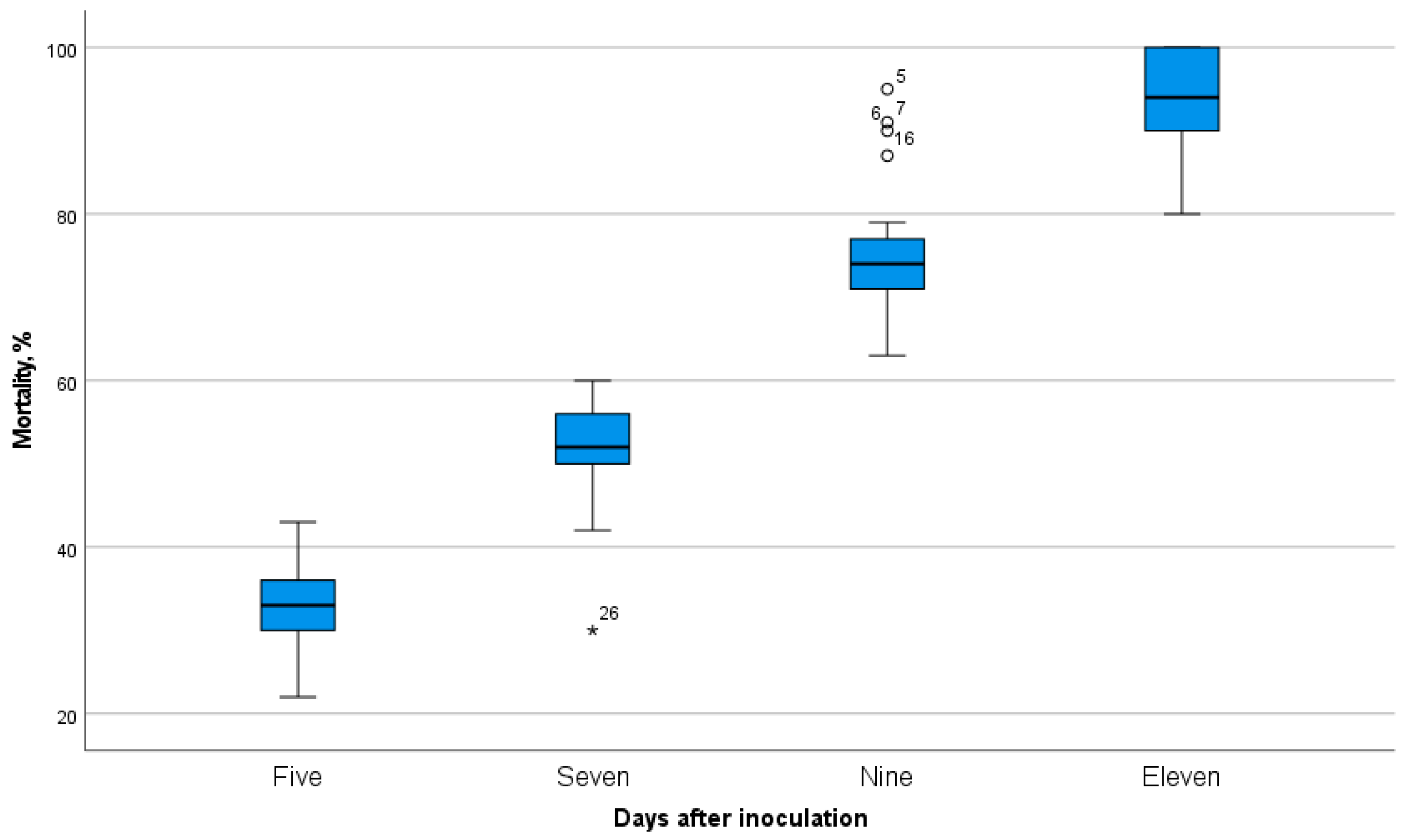
Figure 6.
Relationship between the level of mycosis of Colorado potato beetle larvae and the mortality rate.
Figure 6.
Relationship between the level of mycosis of Colorado potato beetle larvae and the mortality rate.
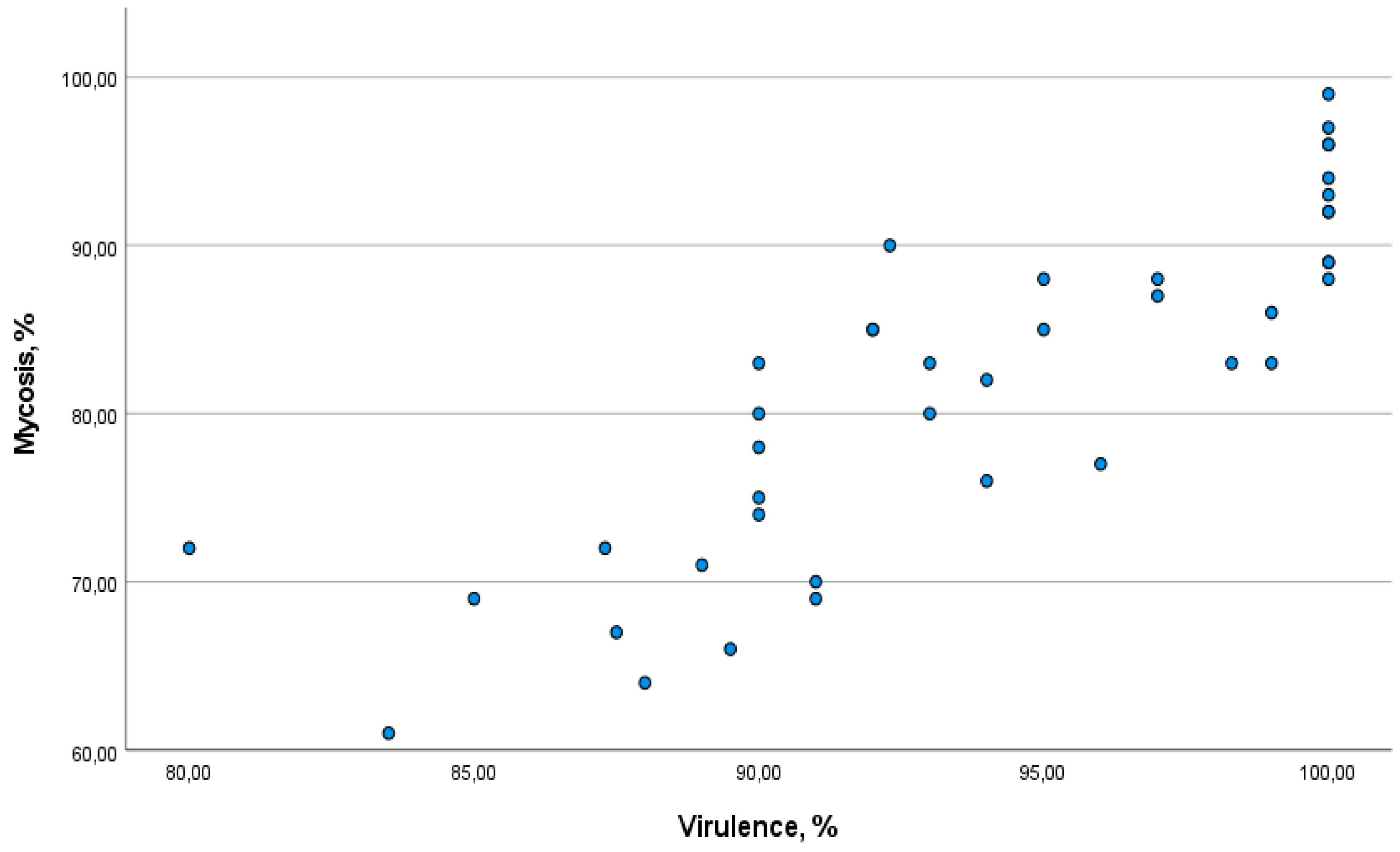

Table 1.
Species composition of natural isolates of EPF isolated from the soils of southern Kazakhstan.
Table 1.
Species composition of natural isolates of EPF isolated from the soils of southern Kazakhstan.
| № | Isolates | Type species in Genbank by ITS locus |
Number of genes read | Other similar species | Type species in Genbank for TEF locus | Number of genes read | Other similar species |
| 1 | B9 |
B. pseudobassiana ARSEF 3405 |
614 |
B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 | 1251 |
B. bassiana isolate ARSEF 815 Beauveria sp. KVL_03_136 |
| 2 | B10 |
B. pseudobassiana ARSEF 3405 |
940 |
B. majiangensis GZAC GZU1214 B. kipukae ARSEF 7032 |
B. bassiana ARSEF 2860 | 1250 |
B. bassiana isolate ARSEF 815 Beauveria sp. KVL_03_136 |
| 3 | B11 |
B. pseudobassiana ARSEF 3405 |
657 |
B.majiangensis GZAC GZU1214 B. bassiana ARSEF 1564 B.australis ARSEF 4598 |
B. bassiana ARSEF 2860 |
1244 |
B. bassiana isolate ARSEF 815 Beauveria sp. KVL_03_136 |
| 4 | B12 | B. bassiana ARSEF 1564 | 352 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B.majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 | 1016 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 5 | B13 | B. bassiana ARSEF 1564 | 588 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B.majiangensis GZAC GZU1214 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 | 1018 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 6 | B14 | B. bassiana ARSEF 1564 | 701 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. pseudobassiana ARSEF 3405 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 |
1314 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 7 | B15 | B. bassiana ARSEF 1564 | 465 |
B.australis ARSEF 4598 B.varroae ARSEF 8257 B.majiangensis GZAC GZU1214 B.amorpha ARSEF 2641 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 |
1029 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 8 | B16 | B. bassiana ARSEF 1564 | 491 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 |
1028 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 9 | B17 | B. bassiana ARSEF 1564 | 636 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. lii ARSEF 11741 B.amorpha ARSEF 2641 B. malawiensis IMI 228343 |
B. bassiana ARSEF 2860 |
1020 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 10 | B18 | B. bassiana ARSEF 1564 | 690 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 |
1014 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 11 | B19 | B. bassiana ARSEF 1564 | 343 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 |
1017 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 12 | B20 | B. bassiana ARSEF 1564 | 314 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 |
1025 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 13 | B21 | B. bassiana ARSEF 1564 | 646 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. lii ARSEF 11741 B. malawiensis IMI 228343 B.amorpha ARSEF 2641 |
B. bassiana ARSEF 2860 |
1015 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 14 | B22 | B. bassiana ARSEF 1564 | 319 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 |
1020 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 15 | B23 | B. bassiana ARSEF 1564 | 462 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 | 1016 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 16 | B24 | B. bassiana ARSEF 1564 | 646 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 Beauveria majiangensis GZAC GZU1214 B. lii ARSEF 11741 B. malawiensis IMI 228343 B.amorpha ARSEF 2641 |
B. bassiana ARSEF 2860 | 1028 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 17 | B25 | B. bassiana ARSEF 1564 | 437 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 |
1019 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 18 | B26 | B. bassiana ARSEF 1564 | 524 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 | 1023 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 19 | B28 | B. bassiana ARSEF 1564 | 384 |
B. australis ARSEF 4598 B. majiangensis GZAC GZU1214 B. varroae ARSEF 8257 |
B. bassiana ARSEF 2860 | 1017 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 20 | B29 | B. bassiana ARSEF 1564 | 575 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B.amorpha ARSEF 2641 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 | 1016 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 21 | B30 | B. bassiana ARSEF 1564 | 425 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 | 1028 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 22 | B53 | B. bassiana ARSEF 1564 | 352 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 |
1011 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 23 | B23-1 | B. bassiana ARSEF 1564 | 472 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 | 1019 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 24 | B23-2 | B. bassiana ARSEF 1564 | 296 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. pseudobassiana ARSEF 3405 |
B. bassiana ARSEF 2860 | 1020 |
B. bassiana isolate ARSEF 815 Beauveria sp. KVL_03_136 |
| 25 | B23-3 |
B. pseudobassiana ARSEF 3405 |
724 |
B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 | 1012 |
B. bassiana isolate ARSEF 815 Beauveria sp. KVL_03_136 |
| 26 | B23-9 |
B. pseudobassiana ARSEF 3405 |
574 |
B. bassiana ARSEF 1564 B. majiangensis GZAC GZU1214 B. australis ARSEF 4598 B. varroae ARSEF 8257 B.a malawiensis IMI 228343 B. kipukae ARSEF 7032 B. brongniartii ARSEF 617 |
B. bassiana ARSEF 2860 | 1021 |
B. bassiana isolate ARSEF 815 Beauveria sp. KVL_03_136 |
| 27 | B23-11 | B. bassiana ARSEF 1564 | 266 |
B. pseudobassiana ARSEF 3405 B. australis ARSEF 4598 B. majiangensis GZAC GZU1214 B. varroae ARSEF 8257 |
B. bassiana ARSEF 2860 | 1015 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 28 | B23-13 | B. bassiana ARSEF 1564 | 394 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 | 1013 |
B. bassiana isolate ARSEF 815 Beauveria sp. KVL_03_136 |
| 29 | B23-14 | B. bassiana ARSEF 1564 | 640 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 |
1022 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 30 | B23-17 | B. bassiana ARSEF 1564 | 755 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. malawiensis IMI 228343 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 |
1029 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 31 | B23-18 | B. bassiana ARSEF 1564 | 846 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B.amorpha ARSEF 2641 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 |
1022 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 32 | B23-23 | B. bassiana ARSEF 1564 | 846 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B.amorpha ARSEF 2641 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 |
1016 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 33 | B23-25 |
B. pseudobassiana ARSEF 3405 |
882 |
B. majiangensis GZAC GZU1214 B. kipukae ARSEF 7032 |
B. bassiana ARSEF 2860 | 1013 |
B. bassiana isolate ARSEF 815 Beauveria sp. KVL_03_136 |
| 34 | B23-11 | B. bassiana ARSEF 1564 | 825 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B.amorpha ARSEF 2641 B. malawiensis IMI 228343 B. lii ARSEF 11741 B. caledonica BCRC 32867 |
B. bassiana ARSEF 2860 | 1017 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 35 | Bc2 | B. bassiana ARSEF 1564 | 432 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B.amorpha ARSEF 2641 |
B. bassiana ARSEF 2860 | 1018 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 36 | Bc3 | B. bassiana ARSEF 1564 | 394 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. lii ARSEF 11741 |
B. bassiana ARSEF 2860 | 1019 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 37 | Bc4 | B. bassiana ARSEF 1564 | 652 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B. lii ARSEF 11741 B.amorpha ARSEF 2641 |
B. bassiana ARSEF 2860 |
1024 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 38 | Bc5 | B. bassiana ARSEF 1564 | 343 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 | 1014 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 39 | Bc6 | B. bassiana ARSEF 1564 | 329 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 | 1020 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 40 | Bc7 | B. bassiana ARSEF 1564 | 626 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 B.amorpha ARSEF 2641 B. malawiensis IMI 228343 |
B. bassiana ARSEF 2860 | 1022 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
| 41 | Bc8 | B. bassiana ARSEF 1564 | 386 |
B. australis ARSEF 4598 B. varroae ARSEF 8257 B. majiangensis GZAC GZU1214 |
B. bassiana ARSEF 2860 | 1026 |
B. bassiana isolate ARSEF 815 B. bassiana isolate Bb5 B. bassiana isolate BbDm-1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



