
Submitted:
20 September 2024
Posted:
23 September 2024
You are already at the latest version
Abstract
Variable retention harvesting (VRH) was designed for timber purposes and biodiversity conservation in natural forests. This system was globally tested, but few studies are related to soil microbial components. The objective was to evaluate different retention types (aggregated and dispersed retention) considering different years-after-harvesting (6, 9, 16 YAH) on soil microbial community attributes compared with unmanaged primary forests (PF) in Nothofagus pumilio forests of Tierra del Fuego (Argentina). We also evaluated the influence of climate, soil and understory vegetation. Results showed that aggregated retention increased microbial biomass carbon (MBC), microbial biomass nitrogen (MBN) and soil basal respiration (SBR) compared to dispersed retention, but with similar values than PF. However, harvested areas decreased MBC:MBN values compared with PF. We found an overall decreasing of microbial biomass and activity in 9 YAH stands, with a positive recovery at 16 YAH. Soil pH, mean annual temperature and understory vegetation cover showed a positive relationship with MBC, MBN, and SBR. The recovery after 16 YAH reaching to a different microbial community. Therefore, maintenance of retention components in managed stands for longer periods is needed. Our results highlight some advantages of VRH as a tool for conservation of forest-dwelling soil microorganisms (microbial biomass and activity).
Keywords:
microbial biomass carbon
; microbial biomass nitrogen
; soil basal respiration
; forest recovery
; sustainable forest management
1. Introduction
There is an increasing need for sustainable forest management practices to maintain and enhance economic, ecological, and social values of native forests for future generations [1,2]. Thus, forest ecosystems provide many services for society, such as wood production, supporting biodiversity maintenance and conservation (above- and below-ground), as well as carbon (C) sequestration to mitigate climate change [3,4]. Although management modifies many components of forest ecosystems, such as vegetation structure and composition, soil properties, litter, and microclimate, most of the research has been focused on above-ground components [5,6,7,8,9,10,11]. Recent studies emphasized the forest management impacts on below-ground soil biology changes and the interactions among microorganisms, plants, and soil components [12,13,14,15,16]. These studies are crucial to understand the long-term impact of harvesting on soil microbial properties, because microorganisms are directly related to productivity and sustainability of natural forest ecosystems [13,14,16,17,18].
In forests and other terrestrial ecosystems, active soil living microorganisms represent only 0.1-2.0% of the total soil volume [19], but they are involved in near 90% of soil ecosystem functions [18,20]. Certainly, soil microorganisms provide multiple functions, and regulating and supporting ecosystem services, such as biogeochemical cycles, mineralization of organic matter, plant growth, CO2 fluxes to the atmosphere, and play key roles in the development and maintenance of soil structure and fertility [21,22,23,24]. Additionally, characterization of soil microbial variables is commonly used to determine soil biological health, including the response of soils to environmental stress such as climate change, anthropogenic disturbances, and as an indicator of ecosystem resilience [25,26,27]. Therefore, soil microbial variables could act as a useful indicator of forest sustainability, ecosystem health and conservation status under different management practices. The magnitude of forest management depends on direct and indirect effects of soil properties such as bulk density, soil nutrients, C cycling, soil fertility, understory plant species, substrate (e.g., litterfall and rhizodeposition) for the soil microorganisms, and microclimate conditions that affect microorganism activity and abundance [12,13,14,28,29].
Variable retention harvesting (VRH) is considered an effective tool to support stand-level biodiversity and used as an indicator for sustainable forest monitoring: e.g., STEMS in British Columbia (Canada), DEMO in Washington-Oregon (USA), and EMEND in Alberta (Canada) [14,16,29,30,31,32,33,34,35]. Several regeneration methods have been proposed for Southern Patagonian Nothofagus pumilio (lenga) forests [36,37,38] based on the natural regeneration [39,40]. These forests have been mainly managed from selection cuts to clear-cuts, including shelterwood cuts and VRH [37,38]. In Tierra del Fuego (Argentina), VRH proposes to leave 30% of the forest area as aggregated retention and 15-20% of the original basal area as dispersed retention [8]. The main goals of VRH are maintain the economic values of the harvesting, facilitate the in-situ conservation and connectivity for biodiversity, and provide faster post-harvest forest recovery [41,42]. Although not is well documented, there are reasons to believe that VRH will provide similar benefits to below-ground biodiversity and function components. The aggregate retention may facilitate soil microbes by maintaining below-ground plant-soil-microorganism interactions [24,43,44,45]. Furthermore, retained trees can provide refugia for above- and below-ground biota being important to maintain carbon and nutrient cycling dynamics [46,47,48]. Additionally, retained trees may maintain physical and chemical soil properties, and supply C through symbiotic interactions, litterfall and rhizodeposition than the surrounding harvest areas [15,16,49]. Also, aggregates in VRH buffer environmental extremes microclimatic conditions by providing shade and maintaining higher soil pH values [13,18,41,46]. Nevertheless, the impact of VRH and years-after-harvesting (YAH) on soil microorganisms was less documented, where soil microbial biomass, fungi and bacteria richness, microbial community structure and microbial activity may be reduced, increased or unaffected in openings harvested areas relative to retention patches [13,16,30,50,51,52,53,54,55].
Below-ground plant-soil interactions have been increasingly recognized as key factor for ecosystem functioning [56,57], and this is considered a knowledge gap for retention forestry [44,47]. Indeed, the soil microbial community in ecosystem forests may influence post disturbance rates of nutrient cycling and, ultimately, plants growth and regeneration success [5,9,11,17,45,48,58]. Therefore, it is important to understand how harvesting influences over soil microbial community in stands with forest managed by VRH across the YAH. To accomplish this, we used an experiment in pure N. pumilio forests in Tierra del Fuego (Argentina). This species is a deciduous tree, cold-tolerant that occurs at lower (sea level) and higher elevations (tree-line), with a remarkable commercial importance, make it an ideal species to evaluate the impacts of forest managements. To our knowledge, few studies have evaluated the response of soil microbial variables, such as biomass and soil respiration, and soil microbial coefficients for VRH across the YAH. Based on further studies describing impacts of VRH on N. pumilio forests, we formulated the following hypothesis related to forest management: soil microbial variables greatly impacted in dispersed retention and conserved at higher levels of retention (e.g., aggregates), and both recover after harvesting but following different trends (e.g., dispersed retention do not reach the same values of aggregates across the years during the recovery). Therefore, the objective was to analyze the influence of different retention types in VRH (aggregated and dispersed retention) across the first 16 YAH on soil microbial community attributes, considering microbial biomass (carbon and nitrogen), soil basal respiration, and microbial indexes, and compare them with control unmanaged primary forests (PF). We also evaluated how climate, soil, and vegetation factors drive the different soil microbial variables by determining the relationships between microbial biomass and soil basal respiration with the main environmental factors. Such knowledge would allow a better understanding of the anthropic, biotic, and abiotic factors that modulate the biomass and function of the soil microorganisms, and ultimately, the soil fertility and conservation of forest ecosystems.
2. Materials and Methods
2.1. Study Area and Experimental Design
The study was carried out in three pure natural N. pumilio forests in one east-west gradient that cover different environmental conditions in Tierra del Fuego (Argentina): (i) Lenga Patagonia S.A. (-54.482 S, -66.802 W); (ii) Campo Chico ranch (-54.581 S, -66.911 W); and (iii) Los Cerros ranch (-54.381 S, -67.862 W). These forests have a rolling topography ranging in elevation from 157 to 265 m.a.s.l. We followed a two-factor random design, with VRH and YAH as main factors. We study two retention treatments: (i) aggregated retention (AR) consisting in one patch of 30 m radius per hectare, where original forest structure was retained, and (ii) dispersed retention (DR) that left remaining trees after harvesting distributed between the aggregates (10-15 m2 ha−1 basal area). YAH factor had three levels: 6 YAH (Lenga Patagonia S.A.), 9 YAH (Campo Chico ranch), and 16 YAH (Los Cerros ranch). At each forest landscape, eight stands were selected (>2 ha each): four harvested stands where the two retention types were applied (AR and DR), and four control stands, defined as primary unmanaged forests (PF) (Figure 1). Control stands were selected for comparisons, considering that they have a good representation of the original soil microbial biomass and activity before harvesting. The final design included 36 stands (3 YAH × 3 forest treatments × 4 replicates). AR and DR were located at least 20 m apart from the aggregate edge, and less than 500 m from PF. Soil samples (n = 4) were randomly collected within the aggregates, DR and PF.
2.2. Characterization of Environmental and Vegetation Variables
The climate parameters (1960-2010) for each area were derived from the WorldClim 2 data set [59], including mean annual precipitation (MAP) and mean annual temperature (MAT) at a resolution of 0.008333° (near 1 km). We also include elevation (m.a.s.l.) derived from Farr et al. [60] due to their direct influence in the regional climate. Tierra del Fuego climate is characterized by short and cool summers, and long snowy winters. MAP varied from 419 to 438 mm yr-1, and MAT fluctuated from 4.0 to 4.4°C at Campo Chico ranch, and from 4.7 to 5.1°C at Los Cerros ranch (Table 1). Only three months are free of mean daily air temperatures below 0°C, being the growing season of approximately five months (November to March) [37].
Forest structure was characterized by two sub-plots, using the point sampling method (BAF = 1-5) [61] with a Criterion RD-1000 (Laser Technology, NY, USA). At each sampling-point we measured: (i) dominant height of the stand, using a TruPulse 200 laser clinometer and distance rangefinder (Laser Technology, NY, USA) by averaging the height of the two taller trees per transect, (ii) tree density, (iii) basal area (BA), and (iv) volume annual growth at stand level. Associated to these forest structure, we also measured tree regeneration: (v) recruitment (seedlings) in two plots of 1 m², (vi) initial regeneration including seedlings and saplings <1.3 m height in two plots of 1 m², and (vii) advanced regeneration including saplings with more than 1.3 m height and <5.0 cm diameter at breast height in two plots of 5 m². For methodology and modelling see Martínez Pastur et al. [62] and Chaves et al. [45]. To evaluate the understory, we used the point-intercept method [63] with 50 intercept points (every 1 m) along the same transects at each sampling plot. We recorded vascular plants (dicots, monocots, ferns) including tree regeneration less than 1.30 m height, identifying dominant and suppressed layers. We also recorded the presence data of the species occurring on transects but that were not intercepted by the points. These data were used to determine richness and forest floor covers, following Moore [64] and Correa [65]. Cover data was sorted by vascular plant species, bare soil, cover, bryophyte (mosses and liverworts), woody debris (>1 cm diameter), and litter. Species richness was calculated as the total number of vascular plant species identified at each plot. Understory vegetation biomass was determined at the peak of the growing season by clipping up to the soil surface all above-ground biomass in two quadrats of 0.2 × 0.5 m. Then, samples were dried (60°C) for 48-h to obtain dry matter plant biomass (total, green, and dead components) expressed in kg dry matter (DM) ha−1.
Nothofagus forest soils in Tierra del Fuego have been classified as podzols according to IUSS Working Group WRB [66]. The soil in the study area is characterized by an organic uppermost layer up to 2 cm thick (O horizon), followed by a mineral layer of less than 40 cm depth where most roots develop (mostly A horizon), with a variable proportion of stony material. The first soils layers in N. pumilio forests (O horizons at <5 cm and A horizons <30 cm) account for most of the soil fertility [11,48,67]. At each plot, four composite soil samples (0-10 cm) were randomly collected using soil auger cores (5.6 cm in diameter) after removing the litter layer. Soil samples were placed in plastic bags and stored in refrigerated containers until transport to the laboratory. The four samples were used for laboratory analysis, including: (i) soil pH determined in 1:2.5 water soil extract; (ii) soil organic carbon (SOC) concentration were derived from the dry combustion (induction furnace) method, using a conversion factor of 0.58 [68]; (iii) Kjeldahl digestion was used to determine soil total nitrogen (N) [69]; and (iv) Olsen methodology was employed to determine available soil phosphorus (P) [70].
2.3. Soil Microbial Biomass and Activity
Soil microbial variables were sampled and determined during February 2022 (coincident with the peak plant growth). We used a subsample of 600 g of soil, which were sieved (2 mm mesh) and then refrigerated at 4ºC during a week until microbial variables analyses. Microbial biomass carbon (MBC) was estimated using the chloroform-fumigation extraction method [71] calculated as: MBC = (OCf - OCnf)/Kec, where OCf = organic C extracted from fumigated samples, OCnf = C extracted from non-fumigated samples, and kEC = fumigation efficiency constant (0.45 MBC expressed as mg C kg-1 dry soil). Microbial biomass nitrogen (MBN) was determined by a modification of the fumigation-incubation method [72]. Liquid chloroform (1 mL) was added directly to 30 g moist soil samples, stirred, and left for 20-h in sealed beakers located in desiccators. Chloroform was then removed using a vacuum pump. Fumigated and non-fumigated samples were incubated at field capacity for 10 days at 25°C. After incubation, samples were extracted with 2M KCl and analyzed for NH4-N by the Berthelot reaction. MBN was calculated as the difference in N between fumigated and non-fumigated samples, divided by a correction factor (kN). Non-extractable amount of microbial N was compensated for by a correction factor of kN = 0.54. MBN was expressed as mg N kg-1 dry soil. Soil basal respiration (SBR) was estimated by quantifying the carbon dioxide (CO2) released in the process of basal respiration (heterotrophic) during 7 days of incubation at 25°C [73]. This was done by placing 75 g fresh soil (moistened to 60% of field capacity) into 1.5 L capacity glass containers with hermetic lids, together with a smaller flask containing 20 mL 0.2 M NaOH to capture the released CO2. The released CO2 was determined by titration with 0.1 M HCl, after precipitation of the barium carbonate formed by adding barium chloride (BaCl2) aqueous solution to the NaOH solution, utilizing phenolphthalein as an indicator. The mean soil respiration (accumulated SBR/7) during the incubation time was expressed as mg C kg-1 soil day-1. Soil microbial metabolic quotient (qCO2) is defined as soil basal respiration per unit of biomass and was calculated as the C-CO2 evolution (SBR) per unit MBC and per unit time. Finally, we calculated the ratio of MBC to total C (qMC = MBC/SOC) and MBC:MBN ratio that indicated shifts in the fungi:bacteria [74,75,76,77,78].
2.4. Statistical Analyses
Data were tested for normal distribution (Shapiro-Wilk test) and homogeneity of variances (Levene’s test). The microbial variables (MBC, MBN, MBC:MBN, SBR, qCO2, qMC) were analyzed with a two-way analysis of variance (ANOVA) using forest treatments (AR, DR, PF) and YAH (6, 9, 16) as main factors. Significant differences were compared with Tukey test with a significance level of p <0.05. A principal component analysis (PCA) was used to analyze the relationships among microbial (MBC, MBN, SBR, MBC:MBN ratio, qCO2), climatic and topographic (MAP, MAT, elevation), soil physical-chemical characteristics (soil pH, SOC, soil N, soil P), and vegetation variables (species richness, understory vegetation biomass, initial and advanced regeneration, dominant overstory height, forest growth, vegetation cover, bare soil, tree density, regeneration recruitment). Simple linear regressions were carried out among microbial (MBC, MBN, SBR, MBC:MBN), soil (SOC, soil pH, soil N), climatic and topographic (MAT, elevation, MAP) and vegetation variables (species richness, vegetation cover, forest growth) to determine correlations between forest treatments and YAH. The dependent variables were soil, topographic, climatic, and vegetation, while independent variables were MBC, MBN, SBR and MBC:MBN. All the statistical analyses were performed using INFOSTAT software [79].
3. Results
3.1. Environmental and Vegetation Variables of the Studied Forest Areas
Vegetation cover was higher at 9 YAH stands compared to 6 and 16 YAH stands (Table 1). In addition, vegetation cover in PF were 78.5 to 107.5%, which contrasted with AR and DR treatments that showed values of 141.5% (6 YAH), 209.0% (9 YAH), and 189.0% (16 YAH), being higher in AR than DR. Bare soil fluctuated from 2.5 to 37.5% in DR and PF (Table 1). The studied forests presented a species richness between 13 and 27, which varied according to YAH and forest treatments. The dominant and most representative plant species were Osmorhiza depauperata, Poa pratensis associated with Viola magellanica, Schizeilema ranunculus, Galium aparine and Taraxacum officinale that varied among YAH and forest treatments. Understory vegetation biomass was higher in DR compared to PF. AR had similar understory vegetation biomass values to PF, except at 9 YAH stands, that showed an increase in the biomass due to tree regeneration. The soil pH values were acidic, decreasing from 5.2 in 16 YAH stands to 4.2 in 6 YAH stands. In contrast, SOC and soil N contents were higher at 9 YAH stands. Additionally, soil P varied among areas and forest treatments (Table 1).
3.2. Response of Soil Microbial Variables to Retention Types and Years-after-Harvesting
Soil microbial variables varied (p <0.05) depending on YAH and forest treatments (Table 2). AR significantly increased MBC and MBN (15-35% and 37%, respectively) compared to DR and PF. Besides, DR significantly decreased SBR (25%) and MBC:MBN ratio (30%) compared to PF. We detected no effects of forest treatments on qCO2 and qMC. Regarding YAH, MBN was significantly higher at 16 YAH stands compared to 6 YAH (57%) and 9 YAH (43%). On the other hand, SBR and MBC:MBN were significantly different between 9 and 16 YAH. We detected no effects of YAH on MBC and qCO2. In contrast, qMC decreased (~50%) at 9 YAH compared to 6 YAH and 16 YAH, where significant interactions were detected between factors for MBC and SBR (Table 2).
The interaction between forest treatments and YAH for MBC showed that 6 YAH and 16 YAH stands did not present statistical differences, while at 9 YAH the DR decreased in MBC compared with AR and PF (Figure 2A). However, the interaction between forest treatments and YAH for SBR, determined that AR had higher values at 6 and 9 YAH compared with DR and PF. In addition, the magnitude of change in SBR was higher at 9 YAH than 6 YAH (DR < PF < AR), but without differences at 16 YAH (Figure 2B).
3.3. Relationships of Soil Microbial Variables with Environmental Factors
Principal component analyses (PCA) determined that the first two axes explained 47-56% of the total variance of the samples (30-37%, and 18-19% for PC1 and PC2, respectively). There was a clear split between DR and AR or PF (Figure 3A). This pattern was explained because DR presented higher species richness, vegetation cover, understory vegetation biomass, and advance regeneration, while AR or PF exhibited higher tree density, stand forest growth, initial regeneration, SBR and MBC. In addition, PCA showed a clear separation between 9 and 16 YAH stands (Figure 3B), being strongly correlated with SOC, soil N and elevation for 9 YAH stands, and MBN, SBR, and MAT for 16 YAH stands.
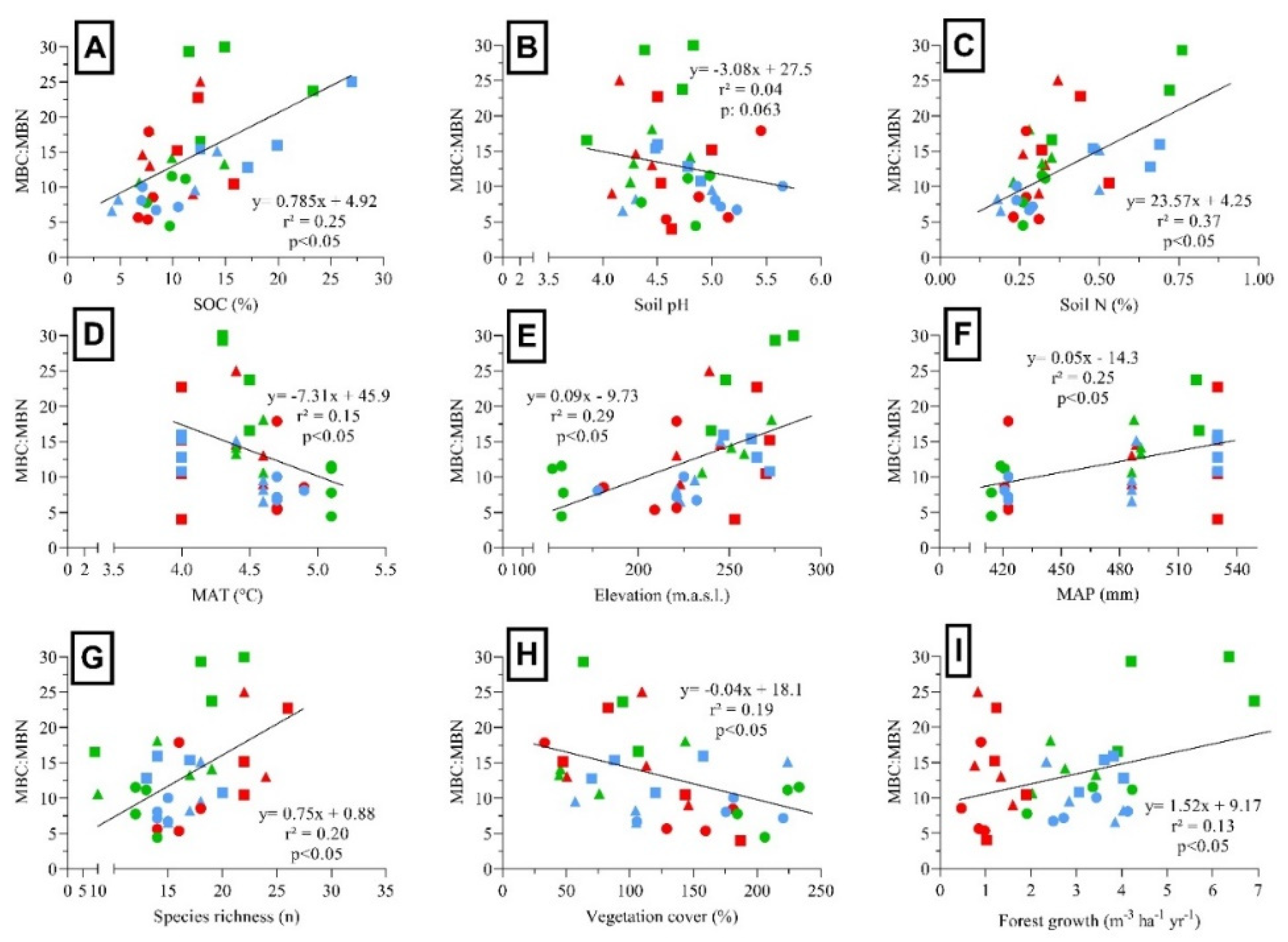
There was a separation between forest treatments (see Appendix Figure A1-A) where AR presented higher MAT, soil pH, MBC, MBN and SBR values, and DR were related to areas with higher elevation. In the analyses, there was also a separation between 16 YAH related to MBC, MBN, SBR, and qCO2, and 6 with 9 YAH related to MBC:MBN, vegetation cover, species richness, and understory vegetation biomass (see Appendix Figure A1-B). To better visualize the relationship between soil microbial and environmental variables (soil physicochemical, climate, understory vegetation), we analyzed correlations by using the most significant variables obtained from PCA. MBC was positively correlated with the SOC, soil pH and N (Figure 4A, B and C). In contrast, MBN was negatively correlated with the SOC and N (Figure 4D and F), while MBN showed positively correlated with soil pH (Figure 4E). On the contrary, SBR showed a weak and not significant response to SOC and N (Figure 4G and I), whereas SBR was positively correlated with soil pH (Figure 4H).
MBC, MBN, and SBR were positively correlated with the MAT (Figure 5A, D and G) and negatively correlated with elevation (Figure 5B, E and H). MBN showed a negative correlation with MAP (Figure 5F), while MBC and SBR showed a weak and not significant response to MAP (Figure 5C and I). MBC, MBN, and SBR declined significantly with the increased species richness (Figure 6A, D and G). In addition, MBC, MBN, and SBR showed positively related to vegetation cover (Figure 6B, E and H). Finally, MBC and SBR showed a positively correlated with forest growth (Figure 6C and I). MBC:MBN ratio was positively correlated with SOC and soil N. In addition, MBC:MBN ratio was negatively correlated with the MAT, while showed positively correlated with elevation and MAP. However, MBC:MBN ratio showed a weak response to soil pH. In contrast, MBC:MBN ratio was positively correlated with species richness, but negatively correlated with vegetation cover. Finally, MBC:MBN ratio was positively correlated with forest growth (see Appendix Figure A2).
4. Discussion
We determined that variable retention harvesting in N. pumilio forests caused shifts on soil microbial biomass and activity. Overall, our results showed that aggregated retention increased MBC, MBN and SBR compared to DR, but conserved similar values that PF (except for MBN). The interaction between forest treatments and YAH showed that the soil biomass (MBC and MBN) and microbial activity (SBR) were most pronounced at 6 and 9 YAH stands. We found that the responses of MBC, MBN, and SBR were strongly dependent on soil, climatic and vegetation variables. Soil pH, MAT and vegetation cover showed a positive relationship with MBC, MBN, and SBR compared with elevation and species richness that showed a negative relationship. Soil microbial biomass and activity is closely related to soil fertility, being one of the most sensitive indicators to disturbances and ecosystems management [25,26]. In our study, retention of living trees in N. pumilio forests as aggregates was successful in conserving and increasing the soil microbial biomass (MBC, MBN, and SBR) compared to primary forests. Although microbial biomass was unaffected by dispersed retention, there was a shift in MBC and MBN caused by aggregated retention (with a trend to decrease). This was consistent with other studies in other forest ecosystems [12,16,80]. However, there are antecedents that showed a negative effect of timber harvesting on soil microbial biomass [28,30,53,81], and other studies that did not find changes in MBC and MBN [18,29]. This inconsistency might be caused by differences in soil type, litter quantity and quality and climate conditions among forest ecosystems, such as MAP, MAT, and elevation compared with our forest growing at these latitudes [29,30,81,82]. Another explanation for the differences found in the present study can be related to the understory vegetation, as a very competitive organisms for inorganic nutrients, and the high uptake rates and extensive root systems [83,84,85,86].
Our results showed that N. pumilio forests increased the understory species richness and vegetation cover in the dispersed retention treatments compared to aggregate retention. Thus, understory vegetation may restrict soil microbial activity via competition for soil available nutrients. Our study found a negative relationship between species richness and microbial biomass (MBC and MBN) and activity (SBR). The competitive dynamic between plants and soil microbes for nutrients has been demonstrated in the forest and other ecosystems [82,86,87,88]. Furthermore, in our study, the dispersed retention increased understory vegetation cover and biomass, while soil microorganism biomass and activity decreased. However, aggregate retention in N. pumilio forests did not modify the understory structure (richness, biomass, vegetation cover), but changes in soil biomass and microbial activity occurred for this forest treatment. In our study, soil microbial growth and activity during late summer (soil sampling in February) might be limited by soil available nutrients due to high understory plant nutrients uptake. Lencinas et al. [9] and Toro Manríquez et al. [11] found that variable retention modified original understory plant assemblages with greater effects in DR than in AR, and where plant distribution was driven by microclimatic conditions. On the other hand, the impacts of harvesting could be influenced by below-ground resources, since roots contribute with similar amounts of litter than above-ground litter as a source of organic matter in forest soils. Average rates of fine-root production in forests range from 3.1 to 6.0 ton ha-1 y-1 [89], which is similar to leaf litter production (4.62 ton ha-1 yr-1) [84]. Similarly, a global meta-analysis demonstrated that root litter accounted for 48% of annual plant-litter inputs in forests, compared to 41% from leaf litter [90]. Soil microbial may be enhanced within patches of living trees after forest harvesting, e.g., at STEMS experiment in British Columbia and EMEND in Alberta (Canada), soil microbial [16,29,30], fungal communities [55], and decomposition of fine roots [34] maintained their levels in retention patches, but not in the dispersed retention. Prescott and Grayston [91] showed that the organic compounds exuded from living roots and associated mycorrhizal fungi were essential for soil biodiversity. It is now evident that a substantial fraction of soil biota depends directly on photosynthate produced from the tree canopy [92]. Thus, our results highlight some advantages of aggregated retention as a tool for conservation of forest-dwelling soil microorganisms. In Douglas-fir forests, Simard et al. [93] found smaller reductions in forest-floor C one year after harvesting in areas where living trees were retained either as single trees (25 large stems ha-1) or in 30 m diameter patches, compared with harvested areas. Martínez Pastur et al. [44,62] showed the importance of preserving N. pumilio forest connectivity to maintain both above- and below-ground forest ecosystem functioning. The response of soil microbial may be the result of trees-soil-microorganism interactions, where N. pumilio forests with high tree density, regeneration and growth rate, may increase SOC (aggregated retention), and therefore improve microbial biomass and activity (MBC, MBN, SBR). This is consistent with previous study, where microbial biomass (bacterial and fungal) increased with SOC, soil N and soil pH [24,57,82]. The effects of these three factors are strongly linked to each other, since SOC and soil N are the major soil resource components, and soil pH influences over the availability of these nutrients. In particular, several studies have identified soil pH as a key environmental variable driving soil microbial communities [94]. In our N. pumilio forests, soil microbial biomass increased with soil pH (4.2 to 5.2). Differences in soil pH can arise from many factors, including vegetation type, soil characteristics, and climate variables [27,57,94,95]. Regardless of the mechanism, our results demonstrated that soil pH was a good predictor of soil microbial variables (MBC, MBN, SBR) in the studied forests. In addition, another possible explanation for the response of microbial community to silvicultural treatments in N. pumilio forests could be related to changes the microorganism communities. Our results demonstrated that the retention treatments tended to decrease the MBC:MBN values compared with the primary unmanaged forests. Soil microbial C:N stoichiometry can reflect microbial community structures. Changes in MBC:MBN ratio indicated shifts in the fungi:bacteria ratio [75,77,78]. In our study, we determined low MBC:MBN values when the forests were harvested (AR and DR), which suggest an increase of bacteria or a decrease in soil fungi [76]. Because fungi (mycorrhizae) are dependent on tree photosynthates for carbon, it is expected to be particularly susceptible to harvesting [16,32,43,53,96]. Girdling studies have shown that removal of below-ground photosynthates reduces the fungal, and to a lesser extent of bacterial communities [14,55,88,97]. Similarly, Luoma et al. [50], found a decrease in ectomycorrhizal in sites with lower trees retention in both aggregated and dispersed harvesting regimes. Also, there are antecedents of a positive relationship between tree cover and the soil myco-biota in N. pumilio forests [43]. Therefore, although aggregate patches cover a smaller area (0.28 ha), it appears that dispersed trees (80 tree ha-1) maintain the conditions necessary for persistence of soil fungi in early stand development stage.
Forest management significantly affects the above- and below-ground structure of the primary N. pumilio forest over time [9,11,36,37]. The canopy opening allows a rapid growth of forest regeneration, due to the increase of light intensity at understory level and soil water availability [6,40,98]. However, these microclimatic changes produce an impact over the natural cycles and soil microorganisms [11,44,45,48,80]. Understanding changes or recovery of soil microbial community of harvested forests is essential to evaluate the potential impact of variable retention forest management. We found an overall decreasing trend of microbial biomass (MBN) and activity in 9 YAH stands, but we found some evidence of a positive recovery in 16 YAH stands, suggesting that forest management after 16 years maintained greater soil microbial biomass and activity possible positive effects on microbially-mediated processes that feed back into forest productivity. Webster et al. [99] found that herbaceous litter, labile needle and bark residues, serve as an important short-term substrate for microbial respiration, while coarser residues (e.g., large branches and stumps) provide substrate availability for soil microorganism in the medium term (5-15 years) bridging a gap until the regenerating forest can produce substantial quantities of litter [100]. Possibility, in austral forests the initial establishment phase largely governed by microclimatic conditions, followed by an accelerated growth phase associated with crown development and understory reflecting increased nutrient demand and greater competence with soil microorganisms.
In Southern Patagonian forests, the increase in bacterial dominance (lower MBC:MBN) in 16-YAH stands was likely related to accelerated microbial turnover and SOC cycling rate, which may increase both soil N level and microbial-derived C accumulation [46,101,102]. Moreover, a lower MBC:MBN ratio is indicative of bacterial dominance (copiotrophs) over fungi (oligotrophs) [78]. The possible increase in soil bacteria due to treatments has been reported in other studies conducted in different forest types and time intervals. For example, bacterial abundance was observed 1-2 years post-harvest in a Norway Spruce stands [28], 4 YAH in oak/hickory stands [103], and 8 YAH in a Douglas-fir stands [104]. Particularly, gram-negative bacteria abundance increase following the harvest treatment may be caused by greater resource availability from rhizodeposition processes. Gram-negative are common single-celled soil bacteria that utilize recent plant C sources such as rhizodeposition [14,105]. Thus, the increase in bacterial dominance (lower MBC:MBN ratios) in N. pumilio forests may likely be related to accelerated microbial turnover and SOC cycling rate, which may increase both in situ organic N level and microbial-derived C [102]. However, previous studies also reported the non-significant effect on MBC:MBN [106] or unchanged fungi:bacteria ratios [107]. Philpott et al. [34] found similar species compositions in the 13 and 70 years-old Douglas-fir stands, indicating that VRH has only marginal short-term effects on β-diversity in the fungi community. Indeed, he not found enough evidence to support the hypothesis that ectomycorrhizal fungi were more abundant under either AR or DR or even in uncut stands. Besides, Varenius et al. [108] found that ectomycorrhizal fungi and fungal communities were significantly altered by forests harvesting but recovering after a >20 YAH period. Therefore, these results may imply a dominance of bacterial community and soil N regulation and driving soil C accumulation in our N. pumilio forests. Moretto and Martínez Pastur [109] indicated that this phenomenon could be explained by microorganism immobilization due to the scarce N soil availability which was evident in our study. In addition, we determined an increase in the microbial quotient (qMC) 16-YAH compared to 6 (15 %) and 9 (56 %) YAH. The qMC ratio was affected by YAH in N. pumilio forests. Thus, we found that the contribution of MBC to SOC increased up to 56% during seven year (from 9 to 16 YAH) suggesting that MBC acts as an important indicator in ecosystem recovery for austral forest ecosystems [25]. Therefore, maintenance of >10 years forest harvested stands (AR and DR) is essential for soil microbial biomass and activity recovery in N. pumilio forests under management. Spake et al. [110] reported that 90 years was the average time for recovery of soil microorganisms’ richness of old-growth forest. Similarly, Holden and Treseder [111] found that microbial responses were negative for the first 15 years following disturbance in boreal forests. Thus, the recovery of soil microbial biomass following harvesting may be controlled by the recovery of forest primary productivity and SOC accumulation.
In our study, significant reductions soil microbial biomass had been observed in dispersed retention at the site 9 YAH. However, seems that microbial biomass and activity recovered 16 YAH. Thus, forest soils of 16 YAH stands supported a greater microbial biomass and activity, but a different microbial communities (as indicated by MBC:MBN) than 9 YAH or 6 YAH forest soils. There are antecedents that the composition of the forest soil microbial community can rebound to pre-harvest levels within 5-10 YAH [28]. It is possible that the efforts to reduce soil disturbance and the facilitation of natural regeneration and biodiversity of N. pumilio forests preserved the resilience of the forest floor microbial communities to disturbance [2,8,9,24]. If soil microbial biomass and activity do not recover after severe harvested N. pumilio forests, a limiting nutrients availability could limit the ecosystems productivity. Our study showed that forest growth was related to MCB and SBR. Therefore, future forest productivity will depend on the ability of soil microbial community to recover activity previous harvesting. However, future studies on the composition of microbial community structure, soil enzyme activities, and bacterial and fungal biodiversity of forest soils are needed to validate the long-term effect of harvesting, as has been studied in other forest ecosystems [14,16,29,30,31,32,33,34,35].
5. Conclusions
Our results showed that aggregated retention increased MBC, MBN and SBR compared to dispersed retention, but decreased MBC:MBN values compared with the primary unmanaged forests. We found an overall decreasing trend of microbial biomass and activity in dispersed retention at the site 9 years after harvested, but we found some evidence of a positive recovery in the sites 16 YAH. Thus, forest soils of 16 YAH stands supported a greater microbial biomass and activity, but a different microbial community than 9 or 6 YAH forest soils. Therefore, more than 10 years are needed without forest intervention to recuperate the microbial soil ecology (oil microbial biomass and activity) in N. pumilio forests under management (aggregated and dispersed retention forest). Also, nutrient retention in microbial biomass would help in ecosystem recovery following disturbance. Therefore, our results highlight some advantages of aggregated retention forest management as a tool for conservation of forest-dwelling soil microorganisms. Such knowledge would allow a better understanding of the anthropic, biotic and abiotic factors that modulate the growth and function of the soil microorganisms and, ultimately, the soil fertility and conservation of forest ecosystems.
Author Contributions
ST: Investigation, Methodology, Formal analysis, Writing-review draft, Writing-review and editing. GMP: Investigation, Project administration, Methodology and Writing-review and editing. JRS: Data curation, Project administration, Investigation, Methodology and Resources. PLP: Conceptualization, Investigation, Writing-review and editing. ST, GMP, JRS, PLP: Drafted the manuscript, and all authors were involved in critical revision and approval of the final version.
Funding
This work was supported by the National Institute of Agricultural Technology (INTA) of Argentina.
Data Availability Statement
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Appendixes
Figure A1.
Principal components analyses (PCA) including microbial variables (MBC = microbial biomass carbon, MBN = microbial biomass nitrogen, SBR = soil basal respiration, MBC:MBN ratio, qCO2 = metabolic quotient), climate and topographic variables (MAP = mean annual precipitation, MAT = mean annual temperature, elevation), soil characteristics (soil pH, SOC = soil organic carbon, N = soil nitrogen, P = soil phosphorus) and understory vegetation variables (species richness, understory vegetation biomass, advanced regeneration, dominant height, recruitment, initial regeneration, forest growth, bare soil, tree density) in N. pumilio forests of Tierra del Fuego. (A) Forest treatments including primary unmanaged forests (PF, green), aggregated retention (AR, blue) and dispersed retention (DR, red), and (B) Years-after-harvesting including 6 YAH (white), 9 YAH (grey), and 16 YAH (black).
Figure A1.
Principal components analyses (PCA) including microbial variables (MBC = microbial biomass carbon, MBN = microbial biomass nitrogen, SBR = soil basal respiration, MBC:MBN ratio, qCO2 = metabolic quotient), climate and topographic variables (MAP = mean annual precipitation, MAT = mean annual temperature, elevation), soil characteristics (soil pH, SOC = soil organic carbon, N = soil nitrogen, P = soil phosphorus) and understory vegetation variables (species richness, understory vegetation biomass, advanced regeneration, dominant height, recruitment, initial regeneration, forest growth, bare soil, tree density) in N. pumilio forests of Tierra del Fuego. (A) Forest treatments including primary unmanaged forests (PF, green), aggregated retention (AR, blue) and dispersed retention (DR, red), and (B) Years-after-harvesting including 6 YAH (white), 9 YAH (grey), and 16 YAH (black).
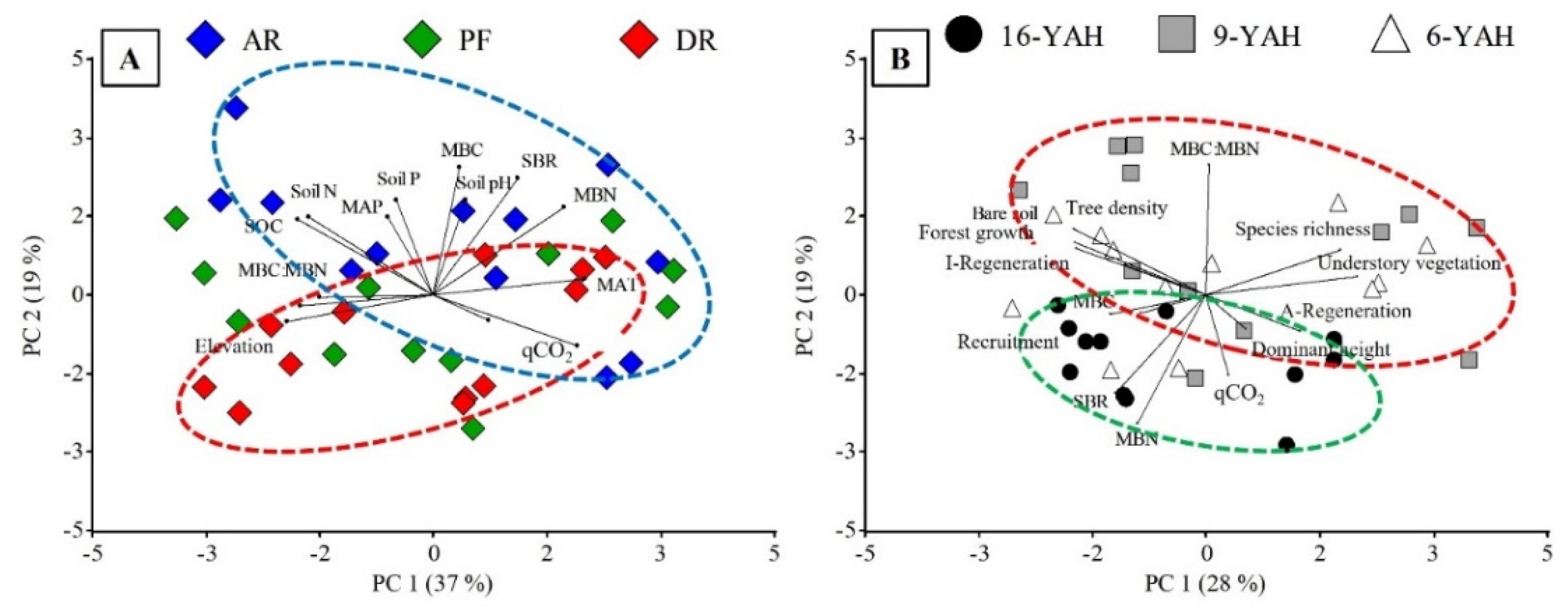
Figure A2.
Relationships among MBC:MBN ratio and soil variables, climatic variables, and vegetation variables in N. pumilio forests of Tierra del Fuego. (A) MBC:MBN and soil organic carbon (SOC), (B) MBC:MBN and soil pH, (C) MBC:MBN and soil nitrogen (N), (D) MBC:MBN and mean annual temperature (MAT), (E) MBC:MBN and elevation, (F) MBC:MBN and mean annual precipitation (MAP), (G) MBC:MBN and species richness, (H) MBC:MBN and understory vegetation cover, and (I) MBC:MBN and forests growth. Dots were classified as primary unmanaged forests (green), aggregated retention (blue) and dispersed retention (red) considering different years-after-harvesting (6 = triangles, 9 = squares, 16 = circles). Lines showed the linear regression, where r² = adjusted parameter, and p = probability.
Figure A2.
Relationships among MBC:MBN ratio and soil variables, climatic variables, and vegetation variables in N. pumilio forests of Tierra del Fuego. (A) MBC:MBN and soil organic carbon (SOC), (B) MBC:MBN and soil pH, (C) MBC:MBN and soil nitrogen (N), (D) MBC:MBN and mean annual temperature (MAT), (E) MBC:MBN and elevation, (F) MBC:MBN and mean annual precipitation (MAP), (G) MBC:MBN and species richness, (H) MBC:MBN and understory vegetation cover, and (I) MBC:MBN and forests growth. Dots were classified as primary unmanaged forests (green), aggregated retention (blue) and dispersed retention (red) considering different years-after-harvesting (6 = triangles, 9 = squares, 16 = circles). Lines showed the linear regression, where r² = adjusted parameter, and p = probability.
References
- Peri, P.L.; Martínez Pastur, G.; Schlichter, T. Uso sustentable del bosque: Aportes desde la silvicultura Argentina; Ministerio de Ambiente y Desarrollo Sostenible de la Nación Argentina: Buenos Aires, Argentina, 2021. http://hdl.handle.net/20.500.12123/10343.
- Soler, R.M.; Lorenzo, C.; González, J.; Carboni, L.; Delgado, J.; Díaz, M.; Huertas Herrera, A. The politics behind scientific knowledge: Sustainable forest management in Latin America. For. Pol. Econ. 2021, 131, e102543. [CrossRef]
- Muys, B. Forest ecosystem services. In Life on land: Encyclopedia of the UN Sustainable Development Goals; Leal Filho, W., Azul, A.M., Brandli, L., Lange Salvia, A., Wall, T., Eds.; Springer: Cham, Switzerland, 2021. [CrossRef]
- Peri, P.L.; Martínez Pastur, G.; Nahuelhual, L. Ecosystem services in Patagonia: A multi-criteria approach for an integrated assessment; Springer Nature: Charm, Switzerland, 2021. [CrossRef]
- Lencinas, M.V.; Martínez Pastur, G.; Gallo, E.; Cellini, J.M. Alternative silvicultural practices with variable retention to improve understory plant diversity conservation in southern Patagonian forests. For. Ecol. Manage. 2011, 262, 1236-1250. [CrossRef]
- Martínez Pastur, G.; Cellini, J.M.; Lencinas, M.V.; Barrera, M.; Peri, P.L. Environmental variables influencing regeneration of Nothofagus pumilio in a system with combined aggregated and dispersed retention. For. Ecol. Manage. 2011, 261, 178-186. [CrossRef]
- Gallo, E.A.; Lencinas, M.V.; Martínez Pastur, G. Site quality influences over understory plant diversity in old-growth and harvested Nothofagus pumilio forests. For. Syst. 2013, 22, 25-38. [CrossRef]
- Soler, R.M.; Schindler, S.; Lencinas, M.V.; Peri, P.L.; Martínez Pastur, G. Why biodiversity increases after variable retention harvesting: A meta-analysis for southern Patagonian forests. For. Ecol. Manage. 2016, 369, 161-169. [CrossRef]
- Lencinas, M.V.; Sola, F.J.; Martínez Pastur, G. Variable retention effects on vascular plants and beetles along a regional gradient in Nothofagus pumilio forests. For. Ecol. Manage. 2017, 406, 251-265. [CrossRef]
- Huertas Herrera, A.; Cellini, J.M.; Barrera, M.D.; Lencinas, M.V.; Martínez Pastur, G. Environment and anthropogenic impacts as main drivers of plant assemblages in forest mountain landscapes of Southern Patagonia. For. Ecol. Manage. 2018, 430, 380-393. [CrossRef]
- Toro Manríquez, M.D.; Cellini, J.M.; Lencinas, M.V.; Peri, P.L.; Peña Rojas, K.; Martínez Pastur, G. Suitable conditions for natural regeneration in variable retention harvesting of southern Patagonian Nothofagus pumilio forests. Ecol. Processes 2019, 8, 1-12. [CrossRef]
- Foote, J.A.; Boutton, T.W.; Scott, D.A. Soil C and N storage and microbial biomass in US southern pine forests: Influence of forest management. For. Ecol. Manage. 2015, 355, 48-57. [CrossRef]
- Lewandowski, T.E.; Forrester, J.A.; Mladenoff, D.J.; D’Amato, A.W.; Palik, B.J. Response of the soil microbial community and soil nutrient bioavailability to biomass harvesting and reserve tree retention in northern Minnesota aspen-dominated forests. Appl. Soil Ecol. 2016, 99, 110-117. [CrossRef]
- Lewandowski, T.E.; Forrester, J.A.; Mladenoff, D.J.; D’Amato, A.W.; Fassnacht, D.S.; Padley, E.; Martin, K.J. Do biological legacies moderate the effects of forest harvesting on soil microbial community composition and soil respiration. For. Ecol. Manage. 2019, 432, 298-308. [CrossRef]
- Singh, S.B.; Saha, S.; Dutta, S.K.; Singh, A.R.; Boopathi, T. Impact of secondary forest fallow period on soil microbial biomass carbon and enzyme activity dynamics under shifting cultivation in North Eastern Hill region, India. Catena 2017, 156, 10-17. [CrossRef]
- Churchland, C.; Bengtson, P.; Prescott, C.E.; Grayston, S.J. Dispersed variable-retention harvesting mitigates N losses on harvested sites in conjunction with changes in soil microbial community structure. Front. For. Glob. Change 2021, 3, e609216. [CrossRef]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: are there any links? Ecology 2003, 84(8), 2042-2050. [CrossRef]
- Li, F.; Zi, H.; Sonne, C.; Li, X. Microbiome sustains forest ecosystem functions across hierarchical scales. Eco-Environ. Health 2023, 2(1), 24-31. [CrossRef]
- Blagodatskaya, E.; Kuzyakov, Y. Active microorganisms in soil: Critical review of estimation criteria and approaches. Soil Biol. Biochem. 2013, 67, 192-211. [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54(4), 655-670. [CrossRef]
- Bardgett, R.D.; Freeman, C.; Ostle, N.J. Microbial contributions to climate change through carbon cycle feedbacks. ISME J. 2008, 2(8), 805-814. [CrossRef]
- Manzoni, S.; Porporato, A. Soil carbon and nitrogen mineralization: Theory and models across scales. Soil Biol. Biochem. 2009, 41(7), 1355-1379. [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Comm. 2016, 7, e10541. [CrossRef]
- Gargaglione, V.; Gonzalez Polo, M.; Birgi, J.; Toledo, S.; Peri, P.L. Silvopastoral use of Nothofagus antarctica forests in Patagonia: Impact on soil microorganisms. Agrofor. Syst. 2022, 96(7), 957-968. [CrossRef]
- Marcos, M.S.; Olivera, N.L. Microbiological and biochemical indicators for assessing soil quality in drylands from Patagonia. In Biology and biotechnology of Patagonian microorganisms; Olivera, N., Libkind, D., Donati, E., Eds.; Springer: Cham, Switzerland, 2016. [CrossRef]
- Schloter, M.; Nannipieri, P.; Sørensen, S.J.; van Elsas, J.D. Microbial indicators for soil quality. Biol. Fert. Soils 2018, 54(1), 1-10. [CrossRef]
- Fierer, N.; Wood, S.A.; de Mesquita, C.P.B. How microbes can, and cannot, be used to assess soil health. Soil Biol. Biochem. 2021, 153, e108111. [CrossRef]
- Siira-Pietikäinen, A.; Pietikäinen, J.; Fritze, H.; Haimi, J. Short-term responses of soil decomposer communities to forest management: Clear felling versus alternative forest harvesting methods. Can. J. For. Res. 2001, 31(1), 88-99. [CrossRef]
- Hannam, K.D.; Quideau, S.A.; Kishchuk, B.E. Forest floor microbial communities in relation to stand composition and timber harvesting in northern Alberta. Soil Biol. Biochem. 2006, 38(9), 2565-2575. [CrossRef]
- Lindo, Z.; Visser, S. Microbial biomass, nitrogen and phosphorus mineralization, and mesofauna in boreal conifer and deciduous forest floors following partial and clear-cut harvesting. Can. J. For. Res. 2003, 33(9), 1610-1620. [CrossRef]
- Jones, M.D.; Twieg, B.D.; Durall, D.M.; Berch, S.M. Location relative to a retention patch affects the ECM fungal community more than patch size in the first season after timber harvesting on Vancouver Island, British Columbia. For. Ecol. Manage. 2008, 255(3-4), 1342-1352. [CrossRef]
- Hartmann, M.; Lee, S.; Hallam, S.J.; Mohn, W.W. Bacterial, archaeal and eukaryal community structures throughout soil horizons of harvested and naturally disturbed forest stands. Environ. Microbiol. 2009, 11(12), 3045-3062. [CrossRef]
- Kranabetter, J.M.; De Montigny, L.; Ross, G. Effectiveness of green-tree retention in the conservation of ectomycorrhizal fungi. Fungal Ecol. 2013, 6(5), 430-438. [CrossRef]
- Philpott, T.J.; Barker, J.S.; Prescott, C.E.; Grayston, S.J. Limited effects of variable-retention harvesting on fungal communities decomposing fine roots in coastal temperate rainforests. Appl. Environ. Microbiol. 2018, 84(3), e02061-17. [CrossRef]
- Sultaire, S.M.; Benucci, G.; Longley, R.; Kroll, A.; Verschuyl, J.; Bonito, G.; Roloff, G. Using high-throughput sequencing to investigate summer truffle consumption by chipmunks in relation to retention forestry. For. Ecol. Manage. 2023, 549, e121460. [CrossRef]
- Martínez Pastur, G.; Cellini, J.M.; Peri, P.L.; Vukasovic, R.; Fernández, M.C. Timber production of Nothofagus pumilio forests by a shelterwood system in Tierra del Fuego (Argentina). For. Ecol. Manage. 2000, 134(1-3), 153-162. [CrossRef]
- Martínez Pastur, G.; Lencinas, M.V.; Cellini, J.M.; Peri, P.L.; Soler, R. Timber management with variable retention in Nothofagus pumilio forests of Southern Patagonia. For. Ecol. Manage. 2009, 258, 436-443. [CrossRef]
- Gea Izquierdo, G.; Martínez Pastur, G.; Cellini, J.M.; Lencinas, M.V. Forty years of silvicultural management in southern Nothofagus pumilio (Poepp. et Endl.) Krasser primary forests. For. Ecol. Manage. 2004, 201(2-3), 335-347. [CrossRef]
- Martínez Pastur, G.; Peri, P.L.; Fernández, M.C.; Staffieri, G.; Rodríguez, D. Desarrollo de la regeneración a lo largo del ciclo del manejo forestal de un bosque de Nothofagus pumilio: Incidencia de la cobertura y el aprovechamiento o cosecha. Bosque 1999, 20(2), 39-46. [CrossRef]
- Rosenfeld, J.M.; Cerrillo, R.N.; Alvarez, J.G. Regeneration of Nothofagus pumilio (Poepp. et Endl.) Krasser forests after five years of seed tree cutting. J. Environ. Manage. 2006, 78(1), 44-51. [CrossRef]
- Franklin, J.F.; Berg, D.R.; Thornburgh, D.A.; Tappeiner, J.C. Alternative silvicultural approaches to timber harvesting: Variable retention systems. In Creating a forestry for the 21st century: The science of ecosystem management; Kohm, K.A., Franklin, J.F., Eds.; Island Press: Washington, USA, 1997; pp. 111-139.
- Mitchell, S.J.; Beese, W.J. The retention system: Reconciling variable retention with the principles of silvicultural systems. For. Chron. 2002, 78(3), 397-403. [CrossRef]
- Elíades, L.A.; Cabello, M.N.; Pancotto, V.; Moretto, A.; Ferreri, N.A.; Saparrat, M.C.; Barrera, M.D. Soil mycobiota under managed and unmanaged forests of Nothofagus pumilio in Tierra del Fuego, Argentina. NZ J. For. Sci. 2019, 49, e7. [CrossRef]
- Martínez Pastur, G.; Vanha-Majamaa, I.; Franklin, J.F. Ecological perspectives on variable retention forestry. Ecol. Processes 2020, 9, 1-6. [CrossRef]
- Chaves, J.E.; Aravena Acuña, M.C.; Rodríguez-Souilla, J.; Cellini, J.M.; Rappa, N.J.; Lencinas, M.V.; Martínez Pastur, G. Carbon pool dynamics after variable retention harvesting in Nothofagus pumilio forests of Tierra del Fuego. Ecol. Processes 2023, 12(1), e5. [CrossRef]
- Jerabkova, L.; Prescott, C.E.; Titus, B.D.; Hope, G.D.; Walters, M.B. A meta-analysis of the effects of clearcut and variable-retention harvesting on soil nitrogen fluxes in boreal and temperate forests. Can. J. For. Res. 2011, 41(9), 1852-1870. [CrossRef]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Martínez Pastur, G.; Messier, C.; Neyland, M.; Palik, B.; Sverdrup-Thygeson, A.; Volney, A.; Wayne, A.; Franklin, J.F. Retention forestry to maintain multifunctional forests: A world perspective. BioScience 2012, 62(7), 633-645. [CrossRef]
- Martínez Pastur, G.; Aravena Acuña, M.C.; Chaves, J.E.; Cellini, J.M.; Silveira, E.M.; Rodríguez-Souilla, J.; Peri, P.L. Nitrogenous and phosphorus soil contents in Tierra del Fuego forests: Relationships with soil organic carbon, climate, vegetation and landscape metrics. Land 2023, 12(5), e983. [CrossRef]
- Paterson, E. Importance of rhizodeposition in the coupling of plant and microbial productivity. Eur. J. Soil Sci. 2003, 54(4), 741-750. [CrossRef]
- Luoma, D.L.; Stockdale, C.A.; Molina, R.; Eberhart, J.L. The spatial influence of Pseudotsuga menziesii retention trees on ectomycorrhiza diversity. Can. J. For. Res. 2006, 36(10), 2561-2573. [CrossRef]
- Bach, L.H.; Grytnes, J.A.; Halvorsen, R.; Ohlson, M. Tree influence on soil microbial community structure. Soil Biol. Biochem. 2010, 42(11), 1934-1943. [CrossRef]
- Mitchell, R.J.; Campbell, C.D.; Chapman, S.J.; Cameron, C.M. The ecological engineering impact of a single tree species on the soil microbial community. J. Ecol. 2010, 98(1), 50-61. [CrossRef]
- Hartmann, M.; Howes, C.G.; Van Insberghe, D.; Yu, H.; Bachar, D.; Christen, R.; Mohn, W.W. Significant and persistent impact of timber harvesting on soil microbial communities in Northern coniferous forests. ISME J. 2012, 6(12), 2199-2218. [CrossRef]
- Teste, F.P.; Lieffers, V.J.; Strelkov, S.E. Ectomycorrhizal community responses to intensive forest management: Thinning alters impacts of fertilization. Plant Soil 2012, 360, 333-347. [CrossRef]
- Churchland, C.; Grayston, S.J.; Bengtson, P. Spatial variability of soil fungal and bacterial abundance: Consequences for carbon turnover along a transition from a forested to clear-cut site. Soil Biol. Biochem. 2013, 63, 5-13. [CrossRef]
- Bardgett, R.D.; Bowman, W.D.; Kaufmann, R.; Schmidt, S.K. A temporal approach to linking aboveground and belowground ecology. Trends Ecol. Evol. 2005, 20(11), 634-641. [CrossRef]
- Fierer, N.; Strickland, M.S.; Liptzin, D.; Bradford, M.A.; Cleveland, C.C. Global patterns in belowground communities. Ecol. Let. 2009, 12(11), 1238-1249. [CrossRef]
- Bartelt-Ryser, J.; Joshi, J.; Schmid, B.; Brandl, H.; Balser, T. Soil feedbacks of plant diversity on soil microbial communities and subsequent plant growth. Perspec. Plant Ecol. Evol. Syst. 2005, 7(1), 27-49. [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Clim. 2017, 37(12), 4302-4315. [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodríguez, E.; Roth, L.; Seal, D.; Shaffer, S.; Shimada, J.; Umland, J.; Werner, M.; Oskin, M.; Burbank, D.; Alsdorf, D. The shuttle radar topography mission. Rev. Geophys. 2007, 45, RG2004. [CrossRef]
- Bitterlich, W. The Relascope Idea: Relative Measurements in Forestry; CAB: London, England, 1984.
- Martínez Pastur, G.; Rosas, Y.M.; Toro Manríquez, M.; Huertas Herrera, A.; Miller, J.A.; Cellini, J.M.; Lencinas, M.V. Knowledge arising from long-term research of variable retention harvesting in Tierra del Fuego: Where do we go from here? Ecol. Processes 2019, 8, 1-16. [CrossRef]
- Levy, E.G.; Madden, E.A. The point method of pasture analyses. NZ J. Agric. 1933, 46, 267-379.
- Moore, D. Flora of Tierra del Fuego; Anthony Nelson: London, England, 1983.
- Correa, M.N. Flora Patagónica; INTA, Buenos Aires, Argentina, 1969-1998.
- IUSS Working Group WRB. World reference base for soil resources 2014, update 2015: International soil classification system for naming soils and creating legends for soil maps; World Soil Resources Reports, FAO, Rome, Italy, 2015; n° 106.
- Peña-Rodríguez, S.; Moretto, A.S.; Pontevedra-Pombal, X.; Oro, N.; García Rodeja Gayoso, E.; Rodríguez-Salgado, I.; Rodríguez-Racedo, J.; Escobar, J.; Nóvoa-Muñoz, J.C. Trends in nutrient reservoirs stored in uppermost soil horizons of subantarctic forests differing in their structure. Agrofor. Syst. 2013, 87, 1273-1128. [CrossRef]
- Peri, P.L.; Rosas, Y.M.; Ladd, B.; Toledo, S.; Lasagno, R.G.; Martínez Pastur, G. Modelling soil carbon content in South Patagonia and evaluating changes according to climate, vegetation, desertification and grazing. Sustainability 2018, 10(2), e438. [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen-total. In Methods of soil analysis: Part 2, chemical and microbiological properties; Page, A.L., Ed.; Agronomy Monographs, American Society of Agronomy: NY, USA, 1983. [CrossRef]
- Olsen, S.R. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. US Department of Agriculture: NY, USA, 1954; n° 939.
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703-707. [CrossRef]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEN value. Soil Biol. Biochem. 1996, 28(1), 33-37. [CrossRef]
- Robertson, G.P.; Wedin, D.; Groffman, P.M.; Blair, J.M.; Holland, E.A.; Nadelhoffer, K.J.; Harris, D. Soil Carbon and nitrogen availability nitrogen mineralization, nitrification and soil respiration potentials. In Standard soil methods for long term ecological research; Robertson, G.P., Coleman, D.C. Bledsoes, C.S., Sollins, P., Eds.; Oxford University Press: NY, USA, 1999.
- Anderson, T.H.; Domsch, K.H. Application of eco-physiological quotients (qCO2 and qMC) on microbial biomasses from soils of different cropping histories. Soil Biol. Biochem. 1990, 22(2), 251-255. [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochem. 2007, 85, 235-252. [CrossRef]
- Sun, Y.; Liao, J.; Zou, X.; Xu, X.; Yang, J.; Chen, H.Y.; Ruan, H. Coherent responses of terrestrial C:N stoichiometry to drought across plants, soil, and microorganisms in forests and grasslands. Agric. For. Meteorol. 2020, 292, e108104. [CrossRef]
- Wan, P.; He, R.; Wang, P.; Cao, A. Implementation of different forest management methods in a natural forest: Changes in soil microbial biomass and enzyme activities. For. Ecol. Manage. 2022, 520, e120409. [CrossRef]
- Wang, M.; Xu, Z.; Huang, Z. Soil carbon accrual under harvest residue retention modulated by the copiotroph-oligotroph spectrum in bacterial community. J. Soils Sed. 2022, 22, 2459-2474. [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; González, L.; Tablada, M.; Robledo, C.W. Infostat; Grupo InfoStat, Universidad Nacional de Córdoba, Córdoba, Argentina, 2013.
- Chatterjee, A.; Vance, G.F.; Pendall, E.; Stahl, P.D. Timber harvesting alters soil carbon mineralization and microbial community structure in coniferous forests. Soil Biol. Biochem. 2008, 40(7), 1901-1907. [CrossRef]
- Das, S.; Deb, S.; Sahoo, S.S.; Sahoo, U.K. Soil microbial biomass carbon stock and its relation with climatic and other environmental factors in forest ecosystems: A review. Acta Ecol. Sinica 2023, 43(6), 933-945. [CrossRef]
- Toledo, S.; Bondaruk, V.F.; Yahdjian, L.; Oñatibia, G.R.; Loydi, A.; Alberti, J.; Peri, P.L. Environmental factors regulate soil microbial attributes and their response to drought in rangeland ecosystems. Sci. Tot. Environ. 2023, 892, e164406. [CrossRef]
- Kuzyakov, Y.; Xu, X. Competition between roots and microorganisms for nitrogen: mechanisms and ecological relevance. New Phytologist 2013, 198(3), 656-669. [CrossRef]
- Jevon, F.V.; Polussa, A.; Lang, A.K.; William Munger, J.; Wood, S.A.; Wieder, W.R.; Bradford, M.A. Patterns and controls of aboveground litter inputs to temperate forests. Biogeochem. 2022, 161(3), 335-352. [CrossRef]
- Morikawa, Y.; Hayashi, S.; Negishi, Y.; Masuda, C.; Watanabe, M.; Watanabe, K.; Seiwa, K. Relationship between the vertical distribution of fine roots and residual soil nitrogen along a gradient of hardwood mixture in a conifer plantation. New Phytol. 2022, 235(3), 993-1004. [CrossRef]
- Zhang, S.; Yang, X.; Li, D.; Li, S.; Chen, Z.; Wu, J. A meta-analysis of understory plant removal impacts on soil properties in forest ecosystems. Geoderma 2022, 426, e116116. [CrossRef]
- Xiao, R.; Man, X.; Duan, B.; Cai, T.; Ge, Z.; Li, X.; Vesala, T. Changes in soil bacterial communities and nitrogen mineralization with understory vegetation in boreal larch forests. Soil Biol. Biochem. 2022, 166, e108572. [CrossRef]
- Fernández, M.; Vincent, G.; Dorr, E.; Bakker, S.; Lerch, T.Z.; Leloup, J.; Bazot, S. Does forest stand density affect soil microbial communities? Appl. Soil Ecol. 2024, 195, e105244. [CrossRef]
- Finér, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics. For. Ecol. Manage. 2011, 262(11), 2008-2023. [CrossRef]
- Freschet, G.T.; Cornwell, W.K.; Wardle, D.A.; Elumeeva, T.G.; Liu, W.; Jackson, B.G.; Cornelissen, J.H. Linking litter decomposition of above- and below-ground organs to plant–soil feedbacks worldwide. J. Ecol. 2013, 101(4), 943-952. [CrossRef]
- Prescott, C.E.; Grayston, S.J. TAMM review: Continuous root forestry, living roots sustain the belowground ecosystem and soil carbon in managed forests. For. Ecol. Manage. 2023, 532, e120848. [CrossRef]
- Högberg, M.N.; Briones, M.J.; Keel, S.G.; Metcalfe, D.B.; Campbell, C.; Midwood, A.J.; Högberg, P. Quantification of effects of season and nitrogen supply on tree below-ground carbon transfer to ectomycorrhizal fungi and other soil organisms in a boreal pine forest. New Phytol. 2010, 187(2), 485-493. [CrossRef]
- Simard, S.W.; Roach, W.J.; Defrenne, C.E.; Pickles, B.J.; Snyder, E.N.; Robinson, A.; Lavkulich, L.M. Harvest intensity effects on carbon stocks and biodiversity are dependent on regional climate in Douglas-fir forests of British Columbia. Front. For. Glob. Change 2020, 3, e88. [CrossRef]
- Liu, J.; Wang, Q.; Ku, Y.; Zhang, W.; Zhu, H.; Zhao, Z. Precipitation and soil pH drive the soil microbial spatial patterns in the Robinia pseudoacacia forests at the regional scale. Catena 2022, 212, e106120. [CrossRef]
- Chen, Y.L.; Ding, J.Z.; Peng, Y.F.; Li, F.; Yang, G.B.; Liu, L.; Yang, Y.H. Patterns and drivers of soil microbial communities in Tibetan alpine and global terrestrial ecosystems. J. Biogeogr. 2016, 43(10), 2027-2039. [CrossRef]
- Busse, M.D.; Beattie, S.E.; Powers, R.F.; Sanchez, F.G.; Tiarks, A.E. Microbial community responses in forest mineral soil to compaction, organic matter removal, and vegetation control. Can. J. For. Res. 2006, 36(3), 577-588. [CrossRef]
- Mummey, D.L.; Clarke, J.T.; Cole, C.A.; O’Connor, B.G.; Gannon, J.E.; Ramsey, P.W. Spatial analysis reveals differences in soil microbial community interactions between adjacent coniferous forest and clearcut ecosystems. Soil Biol. Biochem. 2010, 42(7), 1138-1147. [CrossRef]
- Rodríguez-Souilla, J.; Cellini, J.M.; Lencinas, M.V.; Roig, F.A.; Chaves, J.E.; Aravena Acuña, M.C.; Martínez Pastur, G. Variable retention harvesting and climate variations influence over natural regeneration dynamics in Nothofagus pumilio forests of Southern Patagonia. For. Ecol. Manage. 2023, 544, e121221. [CrossRef]
- Webster, K.L.; Wilson, S.A.; Hazlett, P.W.; Fleming, R.L.; Morris, D.M. Soil CO2 efflux and net ecosystem exchange following biomass harvesting: Impacts of harvest intensity, residue retention and vegetation control. For. Ecol. Manage. 2016, 360, 181-194. [CrossRef]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137(3-4), 253-268. [CrossRef]
- Bingham, A.H.; Cotrufo, M.F. Organic nitrogen storage in mineral soil: implications for policy and management. Sci. Total Environ. 2016, 551, 116-126. [CrossRef]
- Soares, M.; Rousk, J. Microbial growth and carbon use efficiency in soil: links to fungal-bacterial dominance, SOC-quality and stoichiometry. Soil Biol. Biochem. 2019, 131, 195-205. [CrossRef]
- Ponder, F.; Tadros, M. Phospholipid fatty acids in forest soil four years after organic matter removal and soil compaction. Appl. Soil Ecol. 2002, 19(2), 173-182. [CrossRef]
- Moore-Kucera, J.; Dick, R.P. PLFA profiling of microbial community structure and seasonal shifts in soils of a Douglas-fir chronosequence. Microbial Ecol. 2008, 55, 500-511. [CrossRef]
- Kramer, C.; Gleixner, G. Soil organic matter in soil depth profiles: Distinct carbon preferences of microbial groups during carbon transformation. Soil Biol. Biochem. 2008, 40(2), 425-433. [CrossRef]
- Burton, J.; Chen, C.; Xu, Z.; Ghadiri, H. Soil microbial biomass, activity and community composition in adjacent native and plantation forests of subtropical Australia. J. Soils Sed. 2010, 10, 1267-1277. [CrossRef]
- Busse, M.D.; Sanchez, F.G.; Ratcliff, A.W.; Butnor, J.R.; Carter, E.A.; Powers, R.F. Soil carbon sequestration and changes in fungal and bacterial biomass following incorporation of forest residues. Soil Biol. Biochem. 2009, 41(2), 220-227. [CrossRef]
- Varenius, K.; Lindahl, B.D.; Dahlberg, A. Retention of seed trees fails to lifeboat ectomycorrhizal fungal diversity in harvested Scots pine forests. FEMS Microbiol. Ecol. 2017, 93(9), e105. [CrossRef]
- Moretto, A.S.; Martínez Pastur, G. Litterfall and leaf decomposition in Nothofagus pumilio forests along an altitudinal gradient in Tierra del Fuego, Argentina. J. For. Sci. 2014, 60(12), 500-510. https://10.17221/74/2014-JFS.
- Spake, R.; Ezard, T.H.; Martin, P.A.; Newton, A.C.; Doncaster, C.P. A meta-analysis of functional group responses to forest recovery outside of the tropics. Conserv. Biol. 2015, 29(6), 1695-1703. [CrossRef]
- Holden, S.R.; Treseder, K.K. A meta-analysis of soil microbial biomass responses to forest disturbances. Front. Microbiol. 2013, 4, e52720. [CrossRef]
Figure 1.
(A) Study area in the gradient of N. pumilio forests distribution (green) in Tierra del Fuego (Argentina). (B) Satellite image showing variable retention harvesting design in Los Cerros ranch (AR = aggregated retention, DR = dispersed retention, PF = unmanaged primary forests). (C) Diagram of the experimental design and soil sampling points (stars) at each forest stand (AR = blue circles, DR = red area, PF = green area).
Figure 1.
(A) Study area in the gradient of N. pumilio forests distribution (green) in Tierra del Fuego (Argentina). (B) Satellite image showing variable retention harvesting design in Los Cerros ranch (AR = aggregated retention, DR = dispersed retention, PF = unmanaged primary forests). (C) Diagram of the experimental design and soil sampling points (stars) at each forest stand (AR = blue circles, DR = red area, PF = green area).
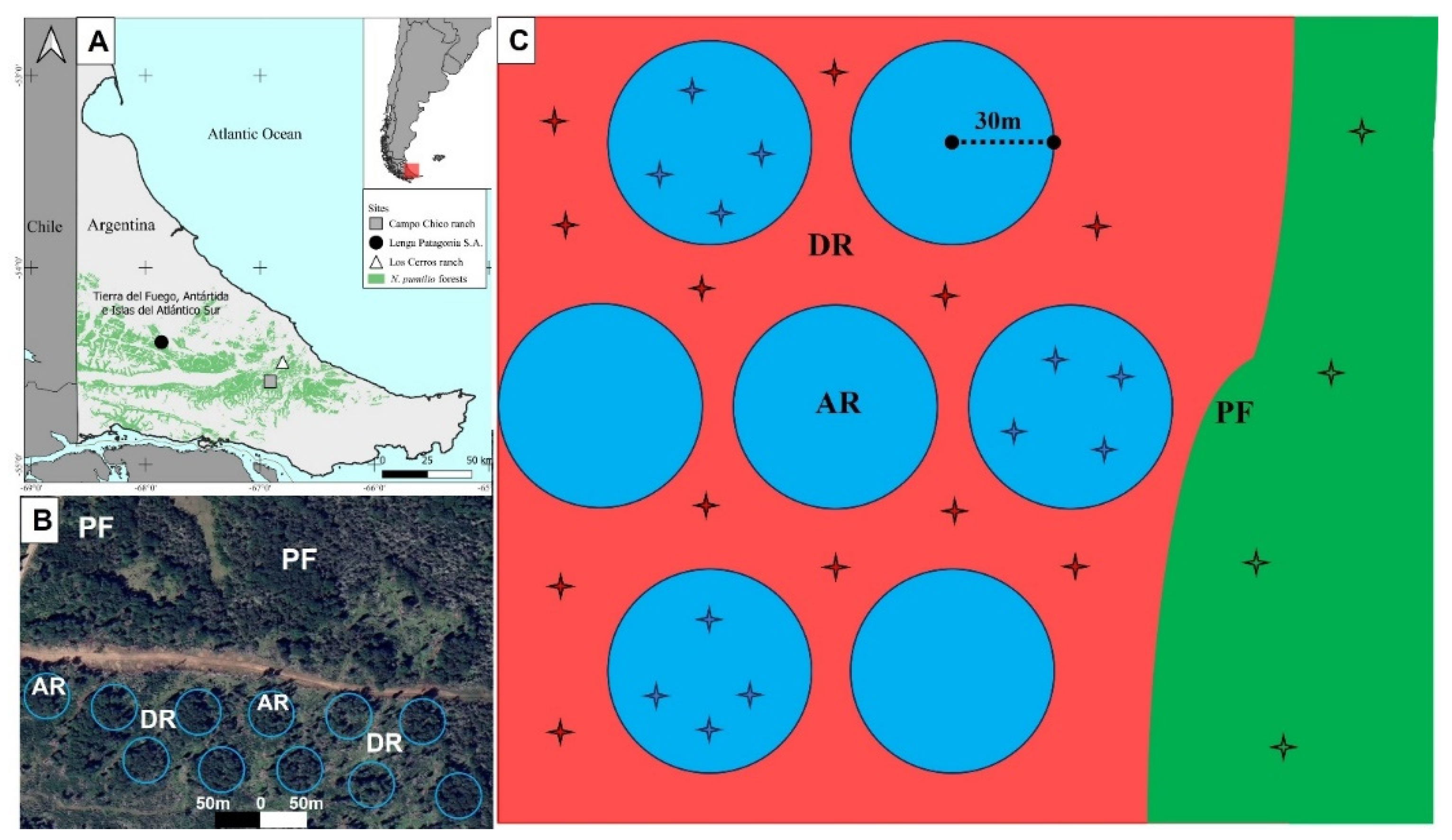
Figure 2.
Response of (A) microbial biomass carbon (MBC), and (B) soil basal respiration (SBR) for the primary unmanaged forests (PF) and managed stands (AR = aggregated retention, DR = dispersed retention) across the different years-after-harvesting (YAH = 6, 9, 16) in N. pumilio forests of Tierra del Fuego. Different letters indicate significant differences (p <0.05).
Figure 2.
Response of (A) microbial biomass carbon (MBC), and (B) soil basal respiration (SBR) for the primary unmanaged forests (PF) and managed stands (AR = aggregated retention, DR = dispersed retention) across the different years-after-harvesting (YAH = 6, 9, 16) in N. pumilio forests of Tierra del Fuego. Different letters indicate significant differences (p <0.05).
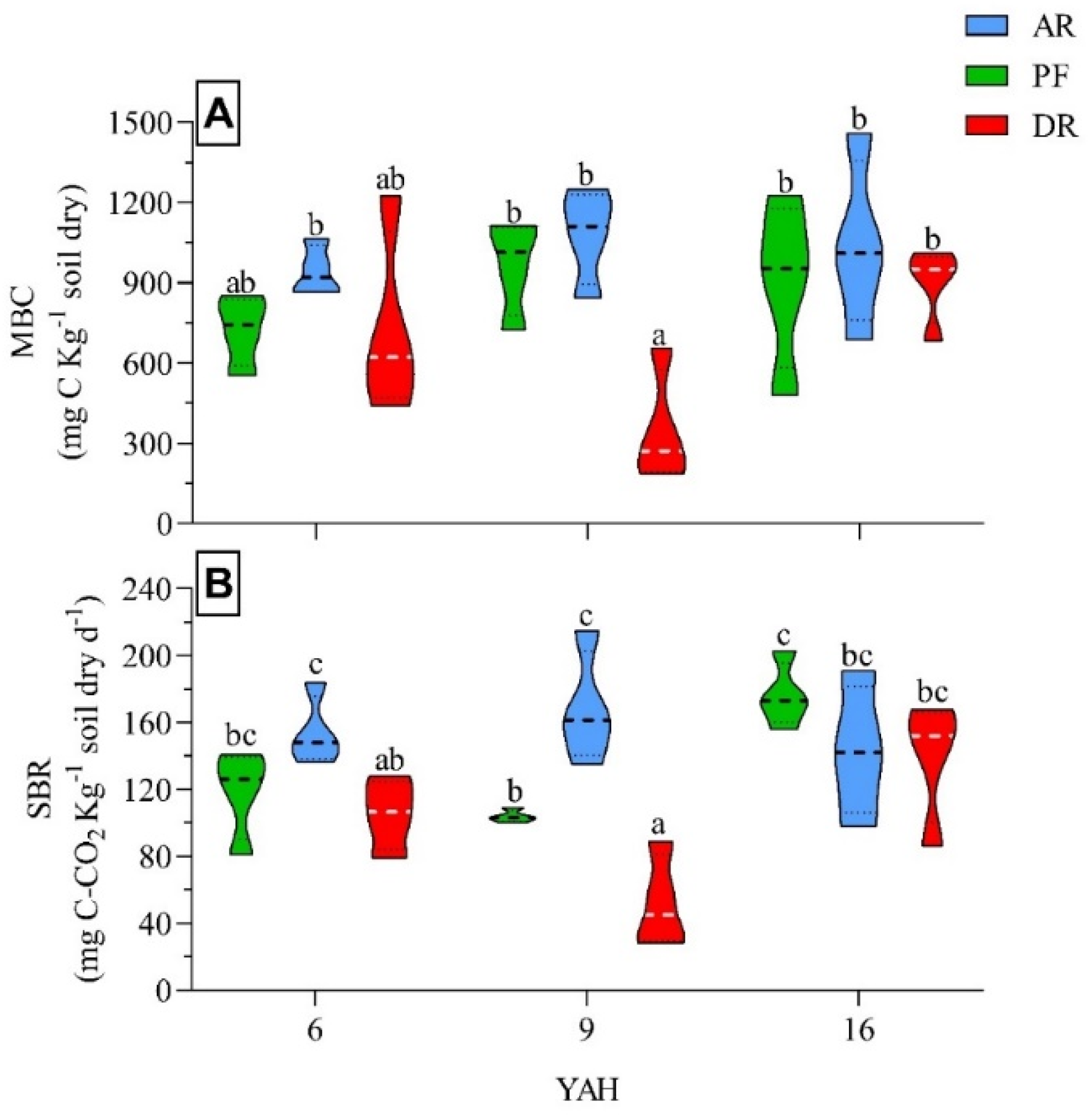
Figure 3.
Principal components analyses (PCA) including microbial variables (MBC = microbial biomass carbon, MBN = microbial biomass nitrogen, SBR = soil basal respiration, MBC:MBN ratio, qCO2 = metabolic quotient), climate and topographic variables (MAP = mean annual precipitation, MAT = mean annual temperature, elevation), soil characteristics (soil pH, SOC = soil organic carbon, N = soil nitrogen, P = soil phosphorus) and understory vegetation variables (species richness, understory vegetation biomass, advance regeneration, dominant height, recruitment, initial regeneration, forest growth, bare soil, tree density) in N. pumilio forests of Tierra del Fuego. (A) Forest treatments including primary unmanaged forests (PF, green), aggregated retention (AR, blue), and dispersed retention (DR, red). (B) Years-after-harvesting including 6 YAH (white), 9 YAH (grey), and 16 YAH stands (black). The ellipses indicate clustering of treatments: green for PF and 16 YAH, red for DR and 9 YAH, and blue for AR and 6 YAH.
Figure 3.
Principal components analyses (PCA) including microbial variables (MBC = microbial biomass carbon, MBN = microbial biomass nitrogen, SBR = soil basal respiration, MBC:MBN ratio, qCO2 = metabolic quotient), climate and topographic variables (MAP = mean annual precipitation, MAT = mean annual temperature, elevation), soil characteristics (soil pH, SOC = soil organic carbon, N = soil nitrogen, P = soil phosphorus) and understory vegetation variables (species richness, understory vegetation biomass, advance regeneration, dominant height, recruitment, initial regeneration, forest growth, bare soil, tree density) in N. pumilio forests of Tierra del Fuego. (A) Forest treatments including primary unmanaged forests (PF, green), aggregated retention (AR, blue), and dispersed retention (DR, red). (B) Years-after-harvesting including 6 YAH (white), 9 YAH (grey), and 16 YAH stands (black). The ellipses indicate clustering of treatments: green for PF and 16 YAH, red for DR and 9 YAH, and blue for AR and 6 YAH.
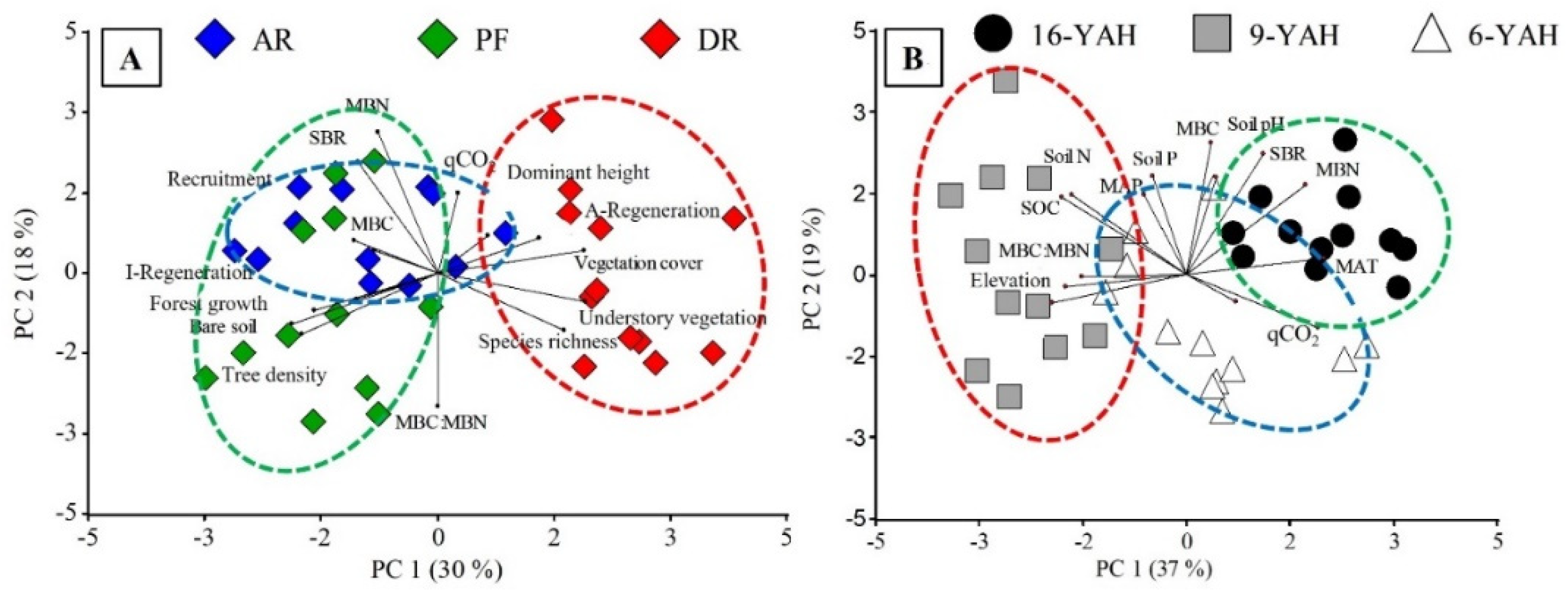
Figure 4.
Relationships among soil microbial biomass and soil activity variables in N. pumilio forests of Tierra del Fuego. (A) Microbial biomass carbon (MBC) and soil organic carbon (SOC), (B) MBC and soil pH, (C) MBC and soil nitrogen (N), (D) microbial biomass nitrogen (MBN) and SOC, (E) MBN and soil pH, (F) MBN and soil N, (G) soil basal respiration (SBR) and SOC, (H) SBR and soil pH, and (I) SBR and soil N. Dots were classified as primary unmanaged forests (green), aggregated retention (blue) and dispersed retention (red) considering different years-after-harvesting (6 = triangles, 9 = squares, 16 = circles). Lines showed a linear regression, where r² = adjusted parameter, and p = probability.
Figure 4.
Relationships among soil microbial biomass and soil activity variables in N. pumilio forests of Tierra del Fuego. (A) Microbial biomass carbon (MBC) and soil organic carbon (SOC), (B) MBC and soil pH, (C) MBC and soil nitrogen (N), (D) microbial biomass nitrogen (MBN) and SOC, (E) MBN and soil pH, (F) MBN and soil N, (G) soil basal respiration (SBR) and SOC, (H) SBR and soil pH, and (I) SBR and soil N. Dots were classified as primary unmanaged forests (green), aggregated retention (blue) and dispersed retention (red) considering different years-after-harvesting (6 = triangles, 9 = squares, 16 = circles). Lines showed a linear regression, where r² = adjusted parameter, and p = probability.
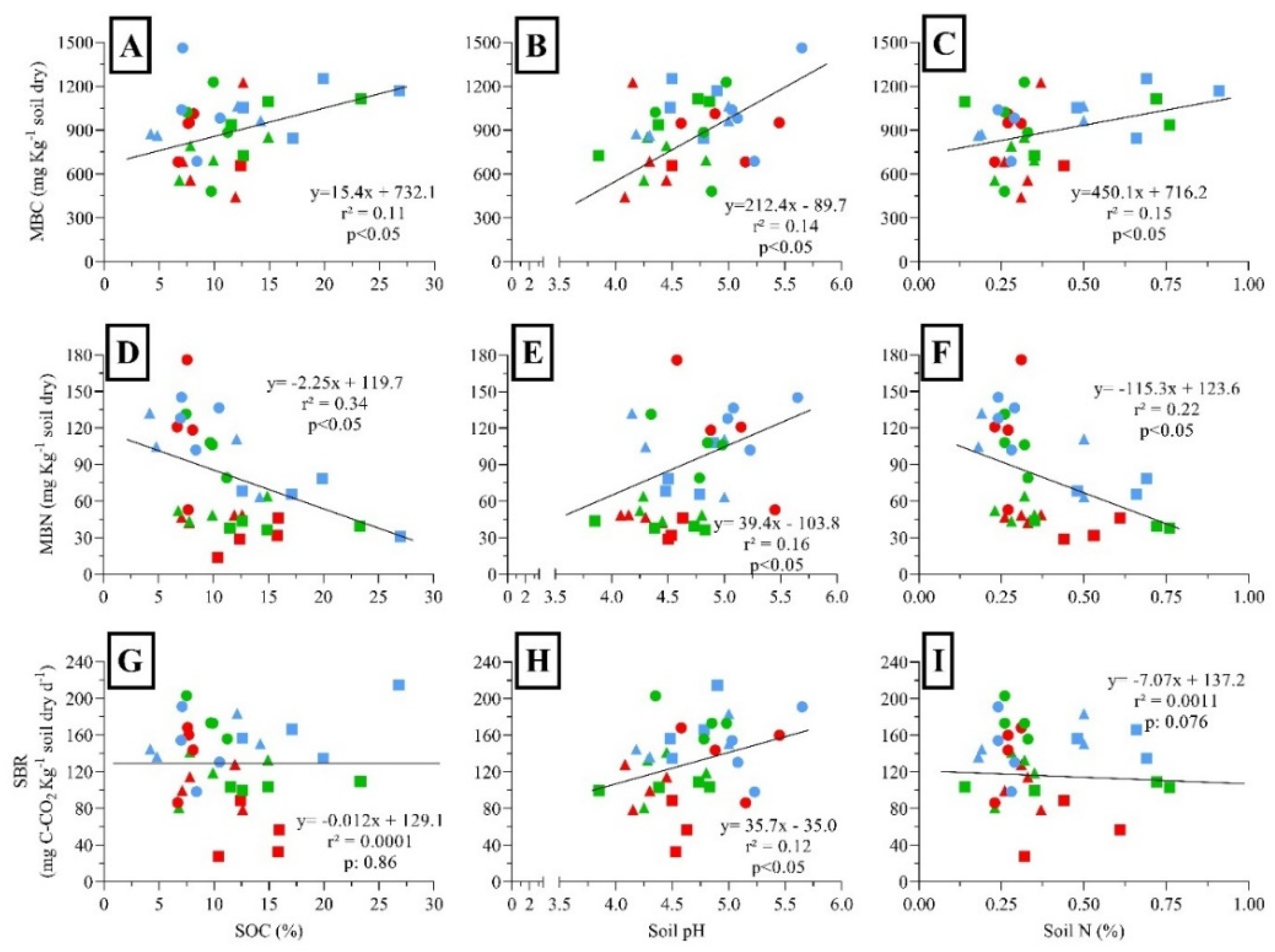
Figure 5.
Relationships among soil microbial biomass and activity and climatic variables in N. pumilio forests of Tierra del Fuego. (A) Microbial biomass carbon (MBC) and mean annual temperature (MAT), (B) MBC and elevation, (C) MBC and mean annual precipitation (MAP), (D) microbial biomass nitrogen (MBN) and MAT, (E) MBN and elevation, (F) MBN and MAP, (G) soil basal respiration (SBR) and MAT, (H) SBR and elevation, and (I) SBR and MAP. Dots were classified as primary unmanaged forests (green), aggregated retention (blue) and dispersed retention (red) considering different years-after-harvesting (6 = triangles, 9 = squares, 16 = circles). Lines showed the linear regression, where r² = adjusted parameter, and p = probability.
Figure 5.
Relationships among soil microbial biomass and activity and climatic variables in N. pumilio forests of Tierra del Fuego. (A) Microbial biomass carbon (MBC) and mean annual temperature (MAT), (B) MBC and elevation, (C) MBC and mean annual precipitation (MAP), (D) microbial biomass nitrogen (MBN) and MAT, (E) MBN and elevation, (F) MBN and MAP, (G) soil basal respiration (SBR) and MAT, (H) SBR and elevation, and (I) SBR and MAP. Dots were classified as primary unmanaged forests (green), aggregated retention (blue) and dispersed retention (red) considering different years-after-harvesting (6 = triangles, 9 = squares, 16 = circles). Lines showed the linear regression, where r² = adjusted parameter, and p = probability.
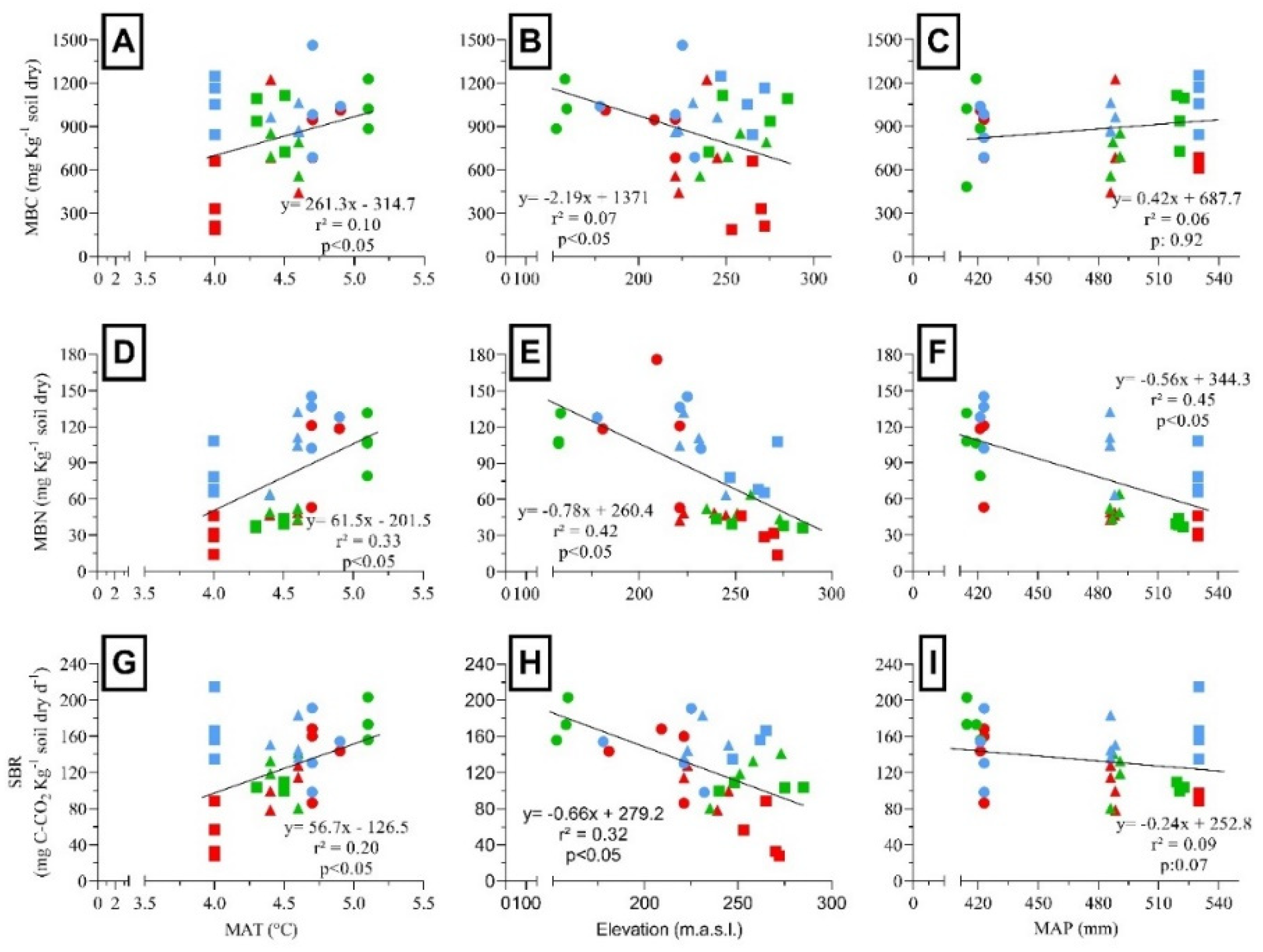
Figure 6.
Relationships among soil microbial biomass and activity and vegetation variables in N. pumilio forests of Tierra del Fuego. (A) Microbial biomass carbon (MBC) and species richness, (B) MBC and vegetation cover, (C) MBC and forest growth, (D) microbial biomass nitrogen (MBN) and species richness, (E) MBN and understory vegetation cover, (F) MBN and forest growth, (G) soil basal respiration (SBR) and species richness, (H) SBR and understory vegetation cover, and (I) SBR and forest growth. Dots were classified as primary unmanaged forests (green), aggregated retention (blue) and dispersed retention (red) considering different years-after-harvesting (6 = triangles, 9 = squares, 16 = circles). Lines showed the linear regression, where r² = adjusted parameter, and p = probability.
Figure 6.
Relationships among soil microbial biomass and activity and vegetation variables in N. pumilio forests of Tierra del Fuego. (A) Microbial biomass carbon (MBC) and species richness, (B) MBC and vegetation cover, (C) MBC and forest growth, (D) microbial biomass nitrogen (MBN) and species richness, (E) MBN and understory vegetation cover, (F) MBN and forest growth, (G) soil basal respiration (SBR) and species richness, (H) SBR and understory vegetation cover, and (I) SBR and forest growth. Dots were classified as primary unmanaged forests (green), aggregated retention (blue) and dispersed retention (red) considering different years-after-harvesting (6 = triangles, 9 = squares, 16 = circles). Lines showed the linear regression, where r² = adjusted parameter, and p = probability.

Table 1.
Characterization of the study areas across a regional gradient of the natural distribution of N. pumilio forests in Tierra del Fuego (Argentina). Geographic location, description of climatic variables, soil physicochemical, and understory vegetation in the studied forest treatments (AR = aggregated retention, DR = dispersed retention, PF = primary unmanaged forests) and years-after-harvesting (YAH = 6, 9, 16). MAP = mean annual precipitation (mm yr-1), MAT = mean annual temperature (°C), ELE = elevation (m.a.s.l.), DH = dominant height (m), TD = tree density (n ha-1), BA = basal area (m2 ha-1), FG = forest growth (m3 ha-1 yr-1), I-R = initial regeneration (n 103 ha-1), A-R = advanced regeneration (n 103 ha-1), REC = recruitment (n 103 ha-1), SP = species richness, BIO = biomass (kg ha-1), DS = dominant species, COV = cover (%), BS = bare soil (%), pH = soil acidity, SOC = soil organic carbon (%), SN = soil nitrogen (%), SP = soil phosphorus(µg g-1), FEMA = Festuca magellanica, GAAP = Galium aparine, HIRE = Hierochloë redolens, MAGR = Macrachaenium gracile, OSDE = Osmorhiza depauperata, PHAL = Phleum alpinum, POPR = Poa pratensis, SCRA = Schizeilema ranunculus, TAOF = Taraxacum officinale, VIMA = Viola magellanica. Values showed mean and standard deviation (±SD).
Table 1.
Characterization of the study areas across a regional gradient of the natural distribution of N. pumilio forests in Tierra del Fuego (Argentina). Geographic location, description of climatic variables, soil physicochemical, and understory vegetation in the studied forest treatments (AR = aggregated retention, DR = dispersed retention, PF = primary unmanaged forests) and years-after-harvesting (YAH = 6, 9, 16). MAP = mean annual precipitation (mm yr-1), MAT = mean annual temperature (°C), ELE = elevation (m.a.s.l.), DH = dominant height (m), TD = tree density (n ha-1), BA = basal area (m2 ha-1), FG = forest growth (m3 ha-1 yr-1), I-R = initial regeneration (n 103 ha-1), A-R = advanced regeneration (n 103 ha-1), REC = recruitment (n 103 ha-1), SP = species richness, BIO = biomass (kg ha-1), DS = dominant species, COV = cover (%), BS = bare soil (%), pH = soil acidity, SOC = soil organic carbon (%), SN = soil nitrogen (%), SP = soil phosphorus(µg g-1), FEMA = Festuca magellanica, GAAP = Galium aparine, HIRE = Hierochloë redolens, MAGR = Macrachaenium gracile, OSDE = Osmorhiza depauperata, PHAL = Phleum alpinum, POPR = Poa pratensis, SCRA = Schizeilema ranunculus, TAOF = Taraxacum officinale, VIMA = Viola magellanica. Values showed mean and standard deviation (±SD).
| Factor | 6-YAH | 9-YAH | 16-YAH | |||||||
| AR | DR | PF | AR | DR | PF | AR | DR | PF | ||
| Latitude | -54.483 | -54.480 | -54.482 | -54.580 | -54.581 | -54.569 | -54.382 | -54.381 | -54.365 | |
| Longitude | -66.828 | -66.823 | -66.802 | -66.907 | -66.907 | -66.927 | -67.864 | -67.862 | -67.869 | |
| Climate | MAP | 486.6 ± 1.7 | 487.2 ± 1.2 | 488.7 ± 2.1 | 530.0 ± 1.1 | 530.0 ± 1.1 | 520.6 ± 1.3 | 422.4 ± 1.0 | 425.4 ± 1.0 | 427.1 ± 3.6 |
| MAT | 4.7 ± 0.1 | 4.6 ± 0.1 | 4.6 ± 0.1 | 4.0 ± 0.1 | 4.0 ± 0.1 | 4.4 ± 0.1 | 4.7 ± 0.1 | 4.7 ± 0.1 | 5.0 ± 0.1 | |
| ELE | 230 ± 11 | 232 ± 12 | 254 ± 16 | 261 ± 10 | 265 ± 8 | 262 ± 21 | 214 ± 24 | 208 ± 19 | 157 ± 3 | |
| Forest Structure | DH | 23.6 ± 3.9 | 21.9 ± 1.9 | 19.1 ± 1.6 | 23.3 ± 2.2 | 24.4 ± 2.1 | 22.4 ± 2.9 | 21.7 ± 2.0 | 21.7 ± 1.3 | 22.7 ± 2.7 |
| TD | 229 ± 111 | 120 ± 87 | 376 ± 89 | 234 ± 172 | 61 ± 37 | 671 ± 172 | 354 ± 170 | 45 ± 20 | 350 ± 114 | |
| BA | 51.0 ± 10.4 | 18.3 ± 6.8 | 55.5 ± 5.2 | 49.5 ± 7.5 | 14.9 ± 3.0 | 63.8 ± 11.1 | 51.0 ± 11.5 | 12.8 ± 3.2 | 50.3 ± 9.0 | |
| FG | 3.3 ± 0.8 | 1.1 ± 0.4 | 2.7 ± 0.6 | 3.6 ± 0.4 | 1.3 ± 0.4 | 5.3 ± 1.5 | 3.2 ± 0.7 | 0.8 ± 0.2 | 3.3 ± 1.0 | |
| I-R | 603 ± 220 | 101 ± 71 | 1078 ± 357 | 446 ± 95 | 94 ± 40 | 195 ± 31 | 181 ± 61 | 31 ± 21 | 170 ± 39 | |
| A-R | 0 ± 0 | 0 ± 0 | 0 ± 0 | 2 ± 1 | 3.3 ± 2.9 | 0 ± 0 | 0 ± 0 | 4.0 ± 2.7 | 0 ± 0 | |
| REC | 198 ± 27 | 21 ± 13 | 56 ± 30 | 0 ± 0 | 3 ± 1 | 0 ± 0 | 31 ±18 | 0 ± 0 | 138 ± 25 | |
| Understory vegetation | SR | 17 ± 1 | 27 ± 4 | 15 ± 4 | 16 ±3 | 23 ± 2 | 17 ± 6 | 15 ± 1 | 16 ± 2 | 13 ± 1 |
| BIO | 406 ± 131 | 3613 ± 178 | 779 ± 236 | 1771 ± 487 | 2493 ± 1216 | 859 ± 184 | 611 ± 267 | 1968 ± 727 | 524 ± 136 | |
| DS | OSDE, VIMA, POPR | GAAP, OSDE, TAOF | OSDE, VIMA, TAOF | OSDE, SCRA, PHAL | OSDE, SCRA, VIMA | MAGR, SCRA, VIMA | GAAP, OSDE, TAOF | OSDE SCRA, HIRE | GAAP, OSDE, FEMA | |
| COV | 103.2 ± 39.2 | 141.5 ± 34.0 | 78.5 ± 38.1 | 174.2 ± 49.3 | 209.0 ± 24.6 | 89.7 ± 56.0 | 63.0 ± 14.8 | 189.0 ± 33.6 | 107.5 ± 27.9 | |
| BS | 25.5 ± 13.7 | 16.0 ± 7.3 | 37.5 ± 17.9 | 7.5 ± 2.4 | 5.5 ± 2.5 | 33.0 ± 22.1 | 35.5 ± 6.2 | 2.5 ± 1.6 | 21.5 ± 10.6 | |
| Soil | pH | 4.62 ± 0.44 | 4.24 ± 0.17 | 4.44 ± 0.25 | 4.66 ± 0.21 | 4.66 ± 0.23 | 4.44 ± 0.44 | 5.24 ± 0.28 | 5.01 ± 0.37 | 4.74 ± 0.27 |
| SOC | 8.81 ± 5.10 | 9.85 ± 2.79 | 9.85 ± 3.62 | 19.11 ± 5.94 | 13.63 ± 2.69 | 15.59 ± 5.37 | 8.24 ± 1.62 | 7.52 ± 0.57 | 9.56 ± 1.55 | |
| SN | 0.34 ± 0.18 | 0.32 ± 0.04 | 0.29 ± 0.06 | 0.69 ± 0.17 | 0.48 ± 0.13 | 0.49 ± 0.30 | 0.26 ± 0.03 | 0.27 ± 0.03 | 0.29 ± 0.04 | |
| SP | 55.9 ± 36.7 | 52.1 ± 17.2 | 48.3 ± 34.5 | 66.7 ± 34.9 | 39.0 ± 21.8 | 62.3 ± 28.2 | 62.9 ± 17.9 | 58.4 ± 27.0 | 47.5 ± 9.6 | |
Table 2.
Analyses of variance for soil biological variables for forest treatments (FT) (PF = primary unmanaged forests, AR = aggregated retention, DR = dispersed retention) and years-after-harvesting (YAH) (6, 9, 16) as main factors in N. pumilio forests of Tierra del Fuego. MBC = microbial biomass carbon (mg kg-1 dry soil), MBN = microbial biomass nitrogen (mg kg-1 dry soil), SBR = soil basal respiration (mg C-CO2 kg-1 dry soil d-1), MBC:MBN = ratio between MBC and MBN, qCO2 = metabolic quotient (µgC-CO2 mg-1 MBC d-1), qMC = microbial quotient (%). Mean values and standard deviation are shown. Different letters indicate significant differences (p <0.05).
Table 2.
Analyses of variance for soil biological variables for forest treatments (FT) (PF = primary unmanaged forests, AR = aggregated retention, DR = dispersed retention) and years-after-harvesting (YAH) (6, 9, 16) as main factors in N. pumilio forests of Tierra del Fuego. MBC = microbial biomass carbon (mg kg-1 dry soil), MBN = microbial biomass nitrogen (mg kg-1 dry soil), SBR = soil basal respiration (mg C-CO2 kg-1 dry soil d-1), MBC:MBN = ratio between MBC and MBN, qCO2 = metabolic quotient (µgC-CO2 mg-1 MBC d-1), qMC = microbial quotient (%). Mean values and standard deviation are shown. Different letters indicate significant differences (p <0.05).
| Treatments | Levels | MBC | MBN | SBR | MBC:MBN | qCO2 | qMC |
| FT | PF | 865±228 ab | 66±33 a | 133±37 b | 13±9 b | 0.17±0.07 a | 0.81±0.29 a |
| AR | 1021±206 b | 104±42 b | 155±31 b | 10±4 a | 0.15±0.02 a | 1.09±0.60 a | |
| DR | 658±331 a | 65±50 a | 99±46 a | 10±8 a | 0.17±0.07 a | 0.75±0.44 a | |
| F(p) | 7.50(<0.05) | 8.62(<0.05) | 12.82(<0.05) | 5.58(<0.05) | 0.15(0.86) | 3.00(0.06) | |
| YAH | 6 | 835±226 a | 75±47 a | 129±30 ab | 11±7 ab | 0.17±0.05 a | 1.08±0.50 b |
| 9 | 712±378 a | 55±32 a | 109±55 a | 13±9 b | 0.16±0.06 a | 0.43±0.26 a | |
| 16 | 970±256 a | 122±39 b | 141±34 b | 8±4 a | 0.15±0.06 a | 1.26±0.40 b | |
| F(p) | 1.70(0.20) | 11.20(<0.05) | 8.28(<0.05) | 6.24(<0.05) | 0.67(0.52) | 9.38(<0.05) | |
| FT x YAH | 2.93(<0.05) | 0.47(0.75) | 5.36(<0.05) | 1.79(0.15) | 1.62(0.19) | 1.25(0.31) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



