Submitted:
23 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
Music has been proposed for a long time as a treatment for mental disorders. However, some special types of music or stimulating sounds might induce negative emotions. Herein, 8-week-old male mice were exposed to two specific genres of music, hip-hop (HH) and heavy metal (HM), and construction noise (CN), and a series of behavioral experiments were performed to evaluate the impact of these music or sounds on the emotion of mice. The experiment lasted 1 week in which 8-week-old male mice were used as animal models. We found that long term exposure HM, HH, and CN induced depressive symptoms with a decrease in dopamine receptor 1 and BDNF in the mixture of hippocampus and frontal cortex of mice. Our results have highly addressed that certain special types of sound or music might induce negative emotions and loss of dopaminergic neurons in mice, and it is necessary to intervene in adolescences who are exposed to these music or sounds for a long time.
Keywords:
adolescence
; depression
; anxiety
; heavy-metal music
; hip-hop
; construction noise
Introduction
Music has often been used as an effective treatment for psychological diseases. The neuroscientific approach proved music exerted a strong influence on the complex neurobiology procedures of the brain, reducing symptoms of mental disorders including depression, anxiety, autism, schizophrenia, and dementia, etc (Lin et al., 2011). It has been reported that music was able to prevent depression and anxiety that are induced by chronic unpredictable mild stress by restoring the levels of BDNF (Fu et al., 2023). Notably, music therapy is found to alleviate both physical pain and psychological pain for terminally ill patients (Gao et al., 2019), while music therapy combined with standardized treatments gives better outcomes in improving depression, (Erkkila et al., 2011) anxiety, (Zhu et al., 2021) schizophrenia, (Mossler, Chen, Heldal, & Gold, 2011) obsessive-compulsive disorder (OCD) (Shiranibidabadi & Mehryar, 2015) in patients.
Nevertheless, most of the previous studies specifically focus on producing soothing and comforting effects by music. However, different music tracks give different levels of relaxation, with loud beats of drums detrimental to the level of relaxation (Elliott, 2011). There are doubts about whether a different musical composition will have the exact opposite effects on a human’s emotional state.
Hip-hop music and heavy metal music are two controversial types of music that appeal to teenagers more than other age groups (Recours, Aussaguel, & Trujillo, 2009). Studies show a general correlation between antisocial behaviors and listening to rap, which includes hip-hop, and heavy metal music (F. Baker & Bor, 2008). Rap music including hip-hop is found to be associated with deviant behaviors (violence, theft, street gangs, mild drug use, and hard drug use) in adolescents (Miranda, 2004). Relationship is found between heavy metal music and depression, delinquency, risk-taking and drug-taking behaviors, suicidal thinking, and deliberate self-harm, despite that the underlying mechanisms is unknown (Arnett, 1991; Hines & McFerran, 2014) due to influences of other factors such as family situations (Martin, 1993). Indeed, heavy metal’s usual hopeless theme might be the main cause of it usually becoming the victim to be blamed for teenager’s psychological health (Arnett, 1991). Still, other studies claim that the miserable themes prevalent in heavy metal music provide an emotional outlet for adolescents, improving their well-being (C. Baker & Brown, 2016), and fans of heavy metal exhibit no anxiety-like or depressive behaviors. Therefore, it is reasonable to doubt the validity of the correlation between heavy metal music and mental disorders (Recours et al., 2009).
Moreover, significant associations were found between noise annoyance and depression, anxiety disorder, and general mental health (Gong et al., 2022). 4 weeks of daily exposure to nontraumatic white noise is found to have anxiogenic effects through elevating the lateral amygdala’s neuronal activity (Peng et al., 2023). Furthermore, the risk of depression was increased by exposure to aircraft noises (Hegewald et al., 2020). However, the chronic effect of construction noise on anxiety or depression was never deeply investigated in the previous studies.
Teenage brains are characterized by a variety of modulations, including a decrease in gray matter accompanied by an increase in white matter (Giedd, 1999), a decrease in neuronal density and synaptic pruning (Peter R., 1979), a shift from diffuse to focal of cortical activity (Durston, 2006), an increase in ventral striatum activity (Pfeifer et al., 2011), amygdala hyperactivation with relatively weaker amygdala-hippocampus connectivity (Guyer, 2008), etc, and most of which show a controversial correlation with adolescent’s impulsive behaviors (Paus, 2008). Some studies attribute teenage impulsive actions to an imbalance of development between earlier maturation of subcortical regions, specifically, the striatum, and less mature prefrontal cortex (one indication of development is the increase in density of dopamine receptors D1 and D2, which is said to account for adolescents’ sensitivity to rewards) (Casey, 2010). Other studies explain that the remodeling of the prefrontal cortex and the dopaminergic system during adolescence (Wahlstrom, White, & Luciana, 2010) leads to anticipations of more abstract and distant types of rewards, which is also more likely to be frustrated, causing adolescents to be more vulnerable to depression (Davey, Yucel, & Allen, 2008).
Social stressors (including social instability and social defeat) applied during adolescence have been shown to promote anxious and depressive behaviors in rodent models (McCormick & Green, 2013). Both acute and chronic stresses act via diverse neuronal circuits to induce anxiety and depression, while the early life environment greatly influences stressor reactivity throughout the life span (Anisman, Merali, & Stead, 2008). 3-weeks of chronic mild stress (CMS) protocol is shown to induce depression through the observation of a decrease in the sucrose preference and an increase in the immobility time in the forced swim test (Kompagne et al., 2008; Rygula et al., 2005), and shown to induce anxiety through the observation of a reduction in open arm exploration in elevated plus maze test (Gameiro et al., 2006; Vyas, Pillai, & Chattarji, 2004). Nonetheless, few studies have been made concerning acoustic stress during adolescence.
Thus, heavy metal music, hip-hop music, or construction noise were applied in our study to evaluate the effects of those voice on emotional state (mainly depression and anxiety) on teenage mice. Therefore, 8-week mice as models for young teenagers were used to measure how acoustic stress stimuli may alter the composition and development of the brain, which is then reflected in the behavior. Several behavioral tests including the sucrose preference test (SPT), the nest building (NB), the open field test (OFT), the elevated plus maze (EPM), the tail suspension test (TST), and the forced swim test (FST) were performed to determine the anxiety and depression level of rodent models. The underlying mechanisms of those voice regulating depressive or anxious behaviors were slightly investigated by measuring the dopamine receptor, serotonin, brain-derived neurotrophic factors, and Trk B’s level in the brain of mice.
Material and Methods
Experimental Design
The study aims to explore the impact of long-term exposure to acoustic stimuli over one week on mice in adolescence (8 weeks old). To minimize variation in the parameters of interest, mice were litter-matched, age-matched, and sex-matched in all animal trials. The chosen acoustic stimuli are intended to focus on intense sounds that may arouse anxiety-like or depressive emotions after long and constant exposure. Two groups of music, hip-hop, and heavy metal, and construction noise were applied in the study. There were four groups of mice, with ten mice per group. Each of them was placed in identical chambers with similar treatments except that three groups were put under three distinct chronic sound stress from 9 am to 12 pm and from 2 pm to 5 pm every day, giving them a total of six hours of acoustic stimuli each day. The voice box is stuck to the top of the box about 0.25m from the mice. Every day, at around 1 pm, each mouse is weighed and the Sucrose preference test (SPT), the Nest building (NB), the Open field test (OFT), the Elevated plus maze (EPM), the Tail suspension test (TST), and the Forced swim test (FST) are conducted to evaluate anxiety or depression-like behaviors in mice. The frontal lobe and hippocampus were collected and the levels of protein expression (dopamine receptor 1-type, D1R; BDNF; Trk B) and neurotransmitter (serotonin, 5-HT) were estimated by ELISA and western blot assays.
Animal Experiment
All animal studies were carried out in strict conformity with the Nanjing University of Chinese Medicine’s institutional ethical standards on animal care. Every specific pathogen-free (SPF) animal was given sterile SPF pellet rodent feed and sterile water. They were also kept in standard environments, with a room temperature of 22 °C and a 12-hour light/dark cycle.
Wild-type C57BL/6 male mice (8 weeks old) were used. Experimental groups were divided into groups of WT mice without any intentional acoustic stimuli (WT, n=10), mice under hip-hop stimuli (HH, n=10), mice under heavy metal stimuli (HM, n=10), and mice under construction noise stimuli (CN, n=10).
Sound Exposure
The music lists for hip-hop and heavy metal are below and with an average of 3-4 min per song and a total of about 30 min for all songs in either hip-hop group or heavy metal group. The mice were exposed with different sounds from 9 am to 12 pm and from 2 pm to 5 pm every day for one week.
Body Weight
The initial weights of mice were recorded and the mice were weighted every day at 2 pm for one week. The average of the weights of mice were calculated daily and the trends of the weights over the week were graphed.
Sucrose Preference Test (SPT)
The sucrose preference test is conducted to evaluate the sound exposure-induced anhedonia in mice models (Liu et al., 2018). The mice were given two bottles, with one 100 mL of 5% sucrose solution and one with an equivalent amount of water, at the start of the trial. The water and sucrose solution were provided on a 24-hour basis every day. The residual volume of each bottle was measured every day at around 2 pm for estimation of the total volume of water and sucrose solution consumed in each group. After the one-week trial, the sucrose preference (%) for each group mice was calculated as the amount of sucrose solution intake over the total liquid intake each day. Data consisting of six data corresponding to six days were obtained for the sucrose preferences for mice under four different sound exposure.
Nest Building (NB)
The nest-building behavioral test was performed according to the previously published approach (Deacon, 2006). Briefly, each mouse was individually placed in an open chamber of 15 cm × 40 cm × 20 cm and given available water and two pieces of cotton. The mice were given a night time to build a nest with the cotton, the results were photographed and scored basing on a rudimentary scale of 4 (Deacon, 2006).
Open Field Test (OFT)
The open field test was performed according to the previously published approach (Kraeuter, Guest, & Sarnyai, 2019). The mice were placed in an open field chamber (length, width, and height: 25 cm × 25 cm × 40 cm) for 10 min. Briefly, the mice were initially placed in the center and their movements were recorded by a video tracking system. The open field is divided into a central field and an outer field. The time mice spent in both fields and the total distance traveled for each mouse were recorded. After each trial, the apparatus is cleaned with 70% ethanol to remove smells that may interfere with the following mice’s performances.
Elevated Plus Maze (EPM)
The elevated plus-maze and the testing procedure were performed according to the previously published approach (File, 1986). The elevated plus maze (EPM) consisted of two open arms (50 cm × 10 cm) and two closed arms (50 cm × 10 cm) elevated 50 cm above the ground. Briefly, mice were initially placed in the center or the conjunction of the four arms facing a closed arm and allowed to explore for 5 min freely. A video tracking system tracked their movements and recorded the time mice spent in open and closed arms and the number of entrances made to each arm. After each trial, the apparatus is cleaned with 70% ethanol to remove smells that may interfere with the following mice’s performances.
Tail Suspension Test (TST)
The tail suspension test was performed according to the previously published approach (Adem Can 2012). Mice were suspended by the tails about 50 cm above the ground for 6 min. The mouse tail was secured to the hook at the top by adhesive tape placed approximately 1 cm below the tail tip. The immobility time was recorded with a video tracking system.
Forced Swim Test (FST)
The forced swim test was performed according to the previously published approach (Roger D. Porsolt, 1978). Mice were placed in Plexiglas cylinders (50 cm high × 20 cm internal diameter) filled with water (23-25 °C) to a depth of 15 cm water. The immobility time was recorded with a video tracking system over the 6 min session.
Western Blot
Western blot was performed to detect the protein levels of BDNF, TrkB and p-TrkB in the frontal lobe and the hippocampal region of the mice. The frontal lobe and the hippocampal region were homogenized in a RIPA solution containing a complete protease inhibitor cocktail using an ultrasound machine. The homogenates were then centrifuged at 12,000 rpm at 4 °C for 30 min, and supernatants were collected and the protein concentration were determined by a BCA detection kit (Beyotime, China). Eventually, the protein samples were combined 1:1 with 2× loading buffer and boiled with 95 °C for 15 min for the following assay.
ELISA Assay
ELISA assay was performed to detect the level of dopamine receptor and serotonin in the frontal lobe and the hippocampal region of the mice by following the manufacturer’s protocol.
Statistical Analysis
GraphPad Prism 8.0.1 was used for statistical analysis and graph plotting. Student’s t-test was used for analyzation of significant differences between two sets of data. The differences in multiple sets of data were analyzed through one-way ANOVA. All data are presented as mean ± SEM. Significance levels were indicated as *p<0.05, **p<0.01, ***p<0.001.
Results
Long-Term Exposure of HH, HM, or CN Induced Depression-Like Behaviors in Mice
Sucrose preference, TST and FST tests were performed to evaluate the depression-like behaviors in mice. For Sucrose preference, the test was conducted per day for all groups over six days. The results showed that a significant reduction in sucrose preferences between WT and HM mice (Figure 1, p<0.01) or CN mice (Figure 1, p<0.05), while no difference was observed in sucrose preference between WT and HH mice.
Furthermore, TST and FST tests were conducted after sound exposure for one week. Notably, results of TST also indicated that WT mice showed a significantly greater mobility time than HH mice (Figure 2, p<0.01), HM mice (Figure 2, p<0.001), and CN mice (Figure 2, p<0.001) while in FST, no difference was observed between the mobility time of WT mice and the mobility time of the other three groups (HH mice, HM mice, or CN mice) (Figure 3).
All results thus demonstrated that long-term exposure of HH mice, HM mice, or CN mice induced depression-like behaviors in mice.
Nest building (NB) behavioral test is also performed due to its correlation with the detrimental state of mice. (Jirkof, 2014) No difference was observed between the quality of nest built by WT mice and the nest built by the other three groups (HH mice, HM mice, or CN mice) (Figure 4).
Long-Term Exposure of HH, HM, or CN Alleviated the Fear and Anxiety-Like Behaviors of Mice
Additionally, EPM and OFT were also performed after sound exposure for one week to evaluate the fear and anxiety-like behaviors towards the new environment.
In the EPM, WT mice showed significantly less time spent in open arms than HH mice (Figure 5a, p<0.05), HM mice (Figure 5a, p<0.01), and CN mice (Figure 5a, p<0.05). Correspondingly, the total distance traveled in the open arm is also significantly less for WT mice than for HH mice (Figure 5b p<0.05), HM mice (Figure 5b, p<0.01), CN mice (Figure 5b, p<0.05). Moreover, HM mice showed a non-significant increase in time spent in the center (Figure 6a) and distance traveled (Figure 6b) compared to the WT mice.
All results suggested a mitigation of anxiety and fear induced by a long-term exposure of strong acoustic stimuli including HH, HM and CN.
Reduction of DR1 and BDNF Might be Responsible for Long-Term Exposure of Strong Acoustic Stimuli-Induced Emotional Changes
Dopamine receptor 1 (DR1) mediates behavior related to anxiety and depression. Given that, the protein level of DR1 was detected to explore the underlying mechanism of long-term exposure of strong acoustic stimuli-induced emotional changes. Results of ELISA indicated long-term exposure of HH (Figure 7, p<0.001) or CN (Figure 7, p<0.001) reduced the protein level of DR1 in brain of mice, while an insignificant decrease in DR1 concentration between WT mice and HM mice were observed. (Figure 7).
Additionally, BDNF, as a key neurotrophic factor, was also considered to be closely related to emotional changes in humans. Then, the effect of long-term exposure of strong acoustic stimuli on BDNF/Trk B pathway was also detected by western blot assay. The results showed that long-term exposure of HH (Figure 8b, p<0.05) and CN (Figure 8c, p<0.01) significantly reduced the protein level of BDNF in brain of mice, and there was no difference In BDNF level between HM mice and WT mice (Figure 8a). Unexpectedly, no difference in Trk B level between WT mice and all other three experiment groups (HM mice, HH mice, CN mice) (Figure 9a, b, c).
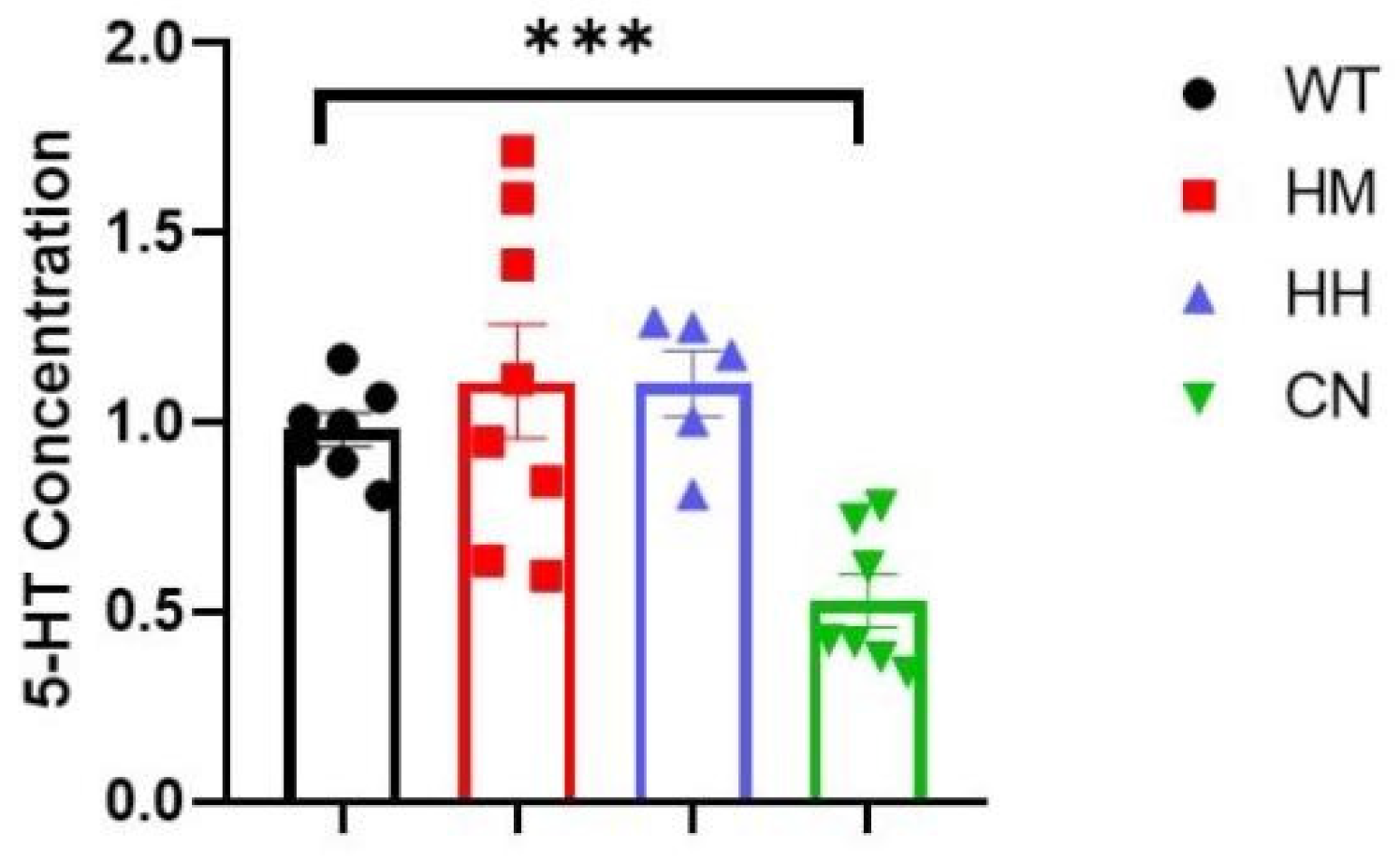
Finally, the effect of long-term exposure of strong acoustic stimuli on 5-HT level was also detected due to its involvement in emotional regulation. However, no significant difference was found in 5-HT concentration between WT mice and HM mice or HH mice, while a significant decrease in 5-HT concentration for CN mice was observed (Figure 10, p<0.001).
Discussion
The main results of this study were that the application of long-term acoustic stimulus, including HH, HM and CN, might induce depression-like behaviors as proven by a significant decrease in sucrose preferences in HM and CN mice, a significant decrease in mobility time in the tail suspension test for HM, HH, and CN mice. Notably, long-term exposure of HH, HM, and CN also alleviated the fear and anxiety-like behaviors of mice as indicated by an increase in time spent and distance traveled in the open arm in the elevated plus maze. Furthermore, our results suggested a significant reduction in DR1 and BDNF levels in the hippocampus and frontal lobe might be responsible for long-term exposure of strong acoustic stimuli-induced emotional changes.
The expression level of D1R is related to stress stimuli. Chronic stress exposure decreases dopamine levels in the mPFC (medial prefrontal cortex). (Hare et al., 2019) Repeated social defeat stress reduces the mRNA level of D1 receptors in mPFC, which also plays a significant role in suppressing stress susceptibility. (Shinohara et al., 2018)
BDNF is also influenced by stress stimuli. Early life stress may exert differential alterations to the expression of BDNF and CREB transcripts in the hippocampus, contributing to individual differences in hippocampal vulnerability to stress which influences mood. (Nair et al., 2007) Chronic social defeat stress induces lasting downregulation of BDNF which is reversed by the antidepressant imipramine. (Tsankova et al., 2006) Chronic stress reduces the expression of BDNF in the dentate gyrus to induce neurogenesis in the hippocampus, preventing depression, (Warner-Schmidt & Duman, 2006) but does not affect the expression of trkB, (Mark A. Smith, 1995) which may explain the reduction in BDNF accompanied by a constant concentration of trkB for HH and CN mice.
D1-like receptors, which include D1R and D5R, are associated with the regulation of depressive symptoms. It has been proven that D1 but not D2 receptor activation increases protein synthesis by eEF2 (Eukaryotic elongation factor 2) dephosphorylation through inhibiting of eEF2K (eEF2 kinase). (David, Barrera, Gould, Gal-Ben-Ari, & Rosenblum, 2020) Notably, the antidepressant effects of ketamine are also attributed to the inhibition of spontaneous glutamate release-driven NMDA receptor activity, following by a decrease in eEF2K activity, thus increasing protein synthesis. (Adaikkan, Taha, Barrera, David, & Rosenblum, 2018) This implies that DR1’s involvement in the regulation of depressive symptoms might be similar to that of ketamine. Moreover, a strong connection is established between ketamine and DR1 in the sense that acute ketamine administration is associated with significantly increased dopamine levels in the cortex. (Kokkinou, Ashok, & Howes, 2018) Therefore, DR1 has been proposed as a possible target for treatments of depression, which yields promising results. Similarly, D1-receptor stimulation by D1 receptor agonists is shown to relieve pain-related depression. (Lazenka, Freitas, Henck, & Negus, 2017) Injections of MPTP (1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine), which reduces dopaminergic neurons, impairs the DR1-mediated neurogenesis in v-DG (ventral dentate gyrus) in the hippocampus, induced depression-like symptoms. (Zhang, Hong, Di, & Chen, 2016) Moreover, the addiction to abuse quetiapine (an effective atypical antipsychotic in the management of mental disorders including depression and anxiety) is blocked by pretreatment of D1 receptor antagonist, suggesting the modulatory role of D1 receptor in the antidepressant pathway of quetiapine. (Althobaiti, 2021)
BDNF level is also related to the mediation of the depression pathway from previous research. Studies prove that the up-regulation of BDNF level in the hippocampus produced antidepressant effects. (Kozisek, Middlemas, & Bylund, 2008; Yukihiko Shirayama, 2002) Serum BDNF is lower in patients with major depressed patients (Karege, 2002) and antidepressant treatments restore BDNF level (Shimizu et al., 2003). However, the mechanisms of the antidepressive role of BDNF are still unclear (Dias, Banerjee, Duman, & Vaidya, 2003)
Nevertheless, the role that BDNF plays in the depression pathway is ambiguous; it may be required for activity-dependent plasticity linked to depression. (Castren, Voikar, & Rantamaki, 2007) The simultaneous decreases in DR1 and BDNF for HH and CN mice suggested a possible relationship between BDNF and DR1. SKF 83959, a D5R and D1-D2 receptor heteromer agonist, induces a 70% increase in the expression of BDNF in the PFC (prefrontal cortex) and elevates Akt signaling, acting like an antidepressant. (Perreault, Jones-Tabah, O’Dowd, & George, 2013) Nevertheless, controversial evidence explains how the activation of dopamine D1-D2 receptor heteromer by dopamine or agonist SKF 83959 leads to intracellular calcium mobilization, resulting in CaMKIIα activation which leads to enhanced BDNF production in the nucleus accumbens, (Ahmed Hasbi, 2009) while BDNF exerts antidepressant-like effects in the hippocampus and pro-depressant effects in the nucleus accumbens. (Kozisek et al., 2008) The interrelation between BDNF and DR1 can exert opposite influences on mood disorders depending on specific regions of the brain, but the connection between the two is evident.
The reduction of DR1 in the mixture of the hippocampus and frontal lobe can be responsible for the anxiolytic effects of acoustic stimuli. A dopaminergic pathway, such as the projection of the ventral tegmentum area to the interpeduncular nucleus, is found to mediate anxiety-like behaviors. Specifically, the excitation of the release of dopamine in VTA→IPN increased, while the inhibition of this pathway reduced, the anxiety behaviors. (DeGroot et al., 2020) Nicotine, a dopamine agonist that stimulates dopamine neurons of the ventral tegmentum area, induces anxiety. (Nguyen et al., 2021)
The decrease in anxiety suggested by the behavioral tests may also be related to the corresponding decrease in BDNF level. The BDNF-TrkB pathway is essential for the consolidation of fear conditioning in the amygdala, as BDNF deletions greatly impair the consolidation of fear conditioning. (Heldt et al., 2014) A single-nucleotide polymorphism (SNP) in the BDNF gene (BDNF Val66Met) that produces Met/Met rats demonstrates a decrease in BDNF secretion (Dincheva, Pattwell, Tessarollo, Bath, & Lee, 2014) and a deficit in forming fear memory. (Jaehne et al., 2022) Additionally, under a stressful setting, Met/Met mice exhibited increased anxiety-related behaviors. (Zhe-Yu Chen, 2006) These findings suggest that anxiety is influenced by the status of the BDNF Val/Met allele.
Furthermore, a relationship is found between BDNF and serotonin (5-HT). The decrease in basal BDNF levels in the hippocampus is not reversed with the administration of fluoxetine, a selective serotonin reuptake inhibitor (SSRI) that normally produce anxiolytic and antidepressant effects, (Bath et al., 2012) suggesting that the function of SSRIs may depend on BDNF. The reduction in BDNF due to BDNF Met polymorphism results in increased anxiety behaviors and decreased 5-HT fiber density. (Duman, 2017) This may explain the anxiolytic effect of construction noise, exemplified by a decrease in time spent and total distance traveled in the open arm during the EPM, and a significant decrease in BDNF level and 5-HT level. However, the anxiolytic effects of music including heavy metal and hip-hop show more correlation with the decrease in BDNF concentration and are not related to 5-HT level.
This study has several limitations. The first limitation was the time frame. One week of acoustic stimuli might not be enough to induce any significant mental disorder or verify the long-term effects of HH, HM, or CN on adolescents. Another limitation was the exclusiveness of the male sex. Female mice may yield completely different results. For adult ovariectomized female rats, dopamine agonists produced depressive-like symptoms while dopamine antagonists exerted antidepressive-like effects, (Fedotova & Ordyan, 2011) implying the complexity of dopamine receptors’ role in inducing or reducing depression depending on different gender. Additionally, female patients were more depressed and expressed less BDNF. (Karege, 2002) The disorder of D1-D2 receptor heteromer in females may significantly promote females’ susceptibility to depression and anxiety disorders. (Hasbi et al., 2020) Lastly, this study slightly explored the possibility of inducing anxiety or depression through HH, HM, or CN, and other mental disorders and molecular changes untested might have taken place. Additionally, dopamine acts at two different receptor families: D1-like receptors (D1 and D5), and D2-like receptors (D2, D3, and D4). This study focused solely on D1-like receptors, while D2-like receptors are also involved in depression, especially for D2 and D3. Several DR2 and DR3 agonists, including 7-OH-DPAT, BP 897, and pramipexole, produced anxiolytic- and antidepressant-like effects in the animal model. (Hori & Kunugi, 2012; Zofia Rogó, 2004) D3R deficiency also results in chronic depression and anxiety. (Moraga-Amaro, Gonzalez, Pacheco, & Stehberg, 2014) Moreover, the downregulation of D3R in the nucleus accumbens shifted microglia to the pro-inflammatory stage and contributes to the development of depressive-like behaviors through Akt signaling pathway, (Wang et al., 2022) marking a shared pathway that is regulated by both DR1 and DR3. Therefore, further experiments should be performed to measure different dopamine receptors, D1-like or D2-like, in different brain regions, hippocampus or nucleus accumbens, to determine the role of acoustic stimuli in inducing depression or anxiety.
Conclusion
In conclusion, acoustic stimuli (heavy metal music, hip-hop music, and construction noise) applied chronically over the course of a week induces depression while alleviates anxiety in 8 weeks old mice models. The sound stimuli also reduces the amount of dopamine receptors 1 and BDNF in the mixture of the frontal lobe and hippocampus. Nevertheless, additional research needs to confirm this finding due to its limited time period and exclusiveness on males. Further research awaits to be conducted to ascertain the relationship between the decrease in protein on a molecular level and the observed behaviors related to depression and anxiety.
Author contributions:
J.L. and J.R. designed the study, performed the animal and cell experiments, and analyzed the interpreted data. J.R. wrote the manuscript and J.L. revised it. Both authors approved the manuscript.
Data availability statement:
Upon a reasonable request, the corresponding author will provide the data sets used and/or analyzed in the current study.
Acknowledgement:
This work was conducted without external funding support.
Conflict of Interest statement:
The authors declare that they have no conflict of interest. The care and use of laboratory animals were conducted in accordance with all institutional and national guidelines.
References
- Adaikkan, C. , Taha, E., Barrera, I., David, O.; Rosenblum, K. Calcium/Calmodulin-Dependent Protein Kinase II and Eukaryotic Elongation Factor 2 Kinase Pathways Mediate the Antidepressant Action of Ketamine. Biol. Psychiatry 2018, 84, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Adem Can, D.T.D. , Chantelle, E.; Terrillion, Sean, C.; Piantadosi, Shambhu Bhat, Todd, D. Gould. The Tail Suspension Test. J. Vis. Exp. 2012, 59. [Google Scholar]
- Ahmed Hasbi, T.F. , Mohammad Alijaniaram, Tuan Nguyen, Melissa, L.; Perreault, Brian, F. O’Dowd, Susan, R. George. Calcium signaling cascade links dopamine D1–D2 receptor heteromer to striatal BDNF production and neuronal growth. Proc. Natl. Acad. Sci. USA.
- Althobaiti, Y.S. Quetiapine-Induced Place Preference in Mice: Possible Dopaminergic Pathway. Pharmaceuticals 2021, 14, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Anisman, H. , Merali, Z.; Stead, J.D.H. Experiential and genetic contributions to depressive- and anxiety-like disorders: Clinical and experimental studies. Neurosci. Biobehav. Rev. 2008, 32, 1185–1206. [Google Scholar] [CrossRef] [PubMed]
- Arnett, J. Adolescents and Heavy Metal Music. Youth Soc. 1991, 23. [Google Scholar] [CrossRef]
- Baker, C.; Brown, B. Suicide. Self-Harm and Survival Strategies in Contemporary Heavy Metal Music: A Cultural and Literary Analysis. J. Med. Humanit. 2016, 37, 1–17. [Google Scholar] [CrossRef]
- Baker, F.; Bor, W. Can music preference indicate mental health status in young people? Australas. Psychiatry 2008, 16, 284–288. [Google Scholar] [CrossRef]
- Bath, K.G. , Jing, D.Q., Dincheva, I., Neeb, C.C., Pattwell, S.S., Chao, M.V.; Ninan, I. BDNF Val66Met Impairs Fluoxetine-Induced Enhancement of Adult Hippocampus Plasticity. Neuropsychopharmacology 2012, 37, 1297–1304. [Google Scholar] [CrossRef]
- Casey, B.J.; Jones, R.M. Neurobiology of the Adolescent Brain and Behavior: Implications for Substance Use Disorders. J. Am. Acad. Child Adolesc. Psychiatry 2010, 49. [Google Scholar] [CrossRef]
- Castren, E. , Voikar, V.; Rantamaki, T. Role of neurotrophic factors in depression. Curr. Opin. Pharmacol. 2007, 7, 18–21. [Google Scholar] [CrossRef]
- Davey, C.G. , Yucel, M.; Allen, N.B. The emergence of depression in adolescence: Development of the prefrontal cortex and the representation of reward. Neurosci. Biobehav. Rev. 2008, 32, 1–19. [Google Scholar] [CrossRef] [PubMed]
- David, O. , Barrera, I., Gould, N., Gal-Ben-Ari, S.; Rosenblum, K. D1 Dopamine Receptor Activation Induces Neuronal eEF2 Pathway-Dependent Protein Synthesis. Front. Mol. Neurosci. 2020, 13, 67. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M. Assessing nest building in mice. Nat. Protoc. 2006, 1, 1117–1119. [Google Scholar] [CrossRef]
- DeGroot, S.R. , Zhao-Shea, R., Chung, L., Klenowski, P.M., Sun, F., Molas, S.; Tapper, A.R. Midbrain Dopamine Controls Anxiety-like Behavior by Engaging Unique Interpeduncular Nucleus Microcircuitry. Biol. Psychiatry 2020, 88, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Dias, B.G. , Banerjee, S.B., Duman, R.S.; Vaidya, V.A. Differential regulation of brain derived neurotrophic factor transcripts by antidepressant treatments in the adult rat brain. Neuropharmacology 2003, 45, 553–563. [Google Scholar] [CrossRef]
- Dincheva, I. , Pattwell, S.S., Tessarollo, L., Bath, K.G.; Lee, F.S. BDNF modulates contextual fear learning during adolescence. Dev. Neurosci. 2014, 36, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S. BDNF, 5-HT, and Anxiety: Identification of a Critical Periadolescent Developmental Period. Am. J. Psychiatry 2017, 174, 1137–1139. [Google Scholar] [CrossRef]
- Durston, S. , Davidson, M.C., Tottenham, N., Galvan, A., Spicer, J., Fossella, J.A.; Casey, B.J. A shift from diffuse to focal cortical activity with development. Dev. Sci. 2006, 9. [Google Scholar]
- Elliott, D. , Polman, R.; McGregor, R. Relaxing Music for Anxiety Control. J. Music Ther. 2011, 48. [Google Scholar]
- Erkkila, J. , Punkanen, M., Fachner, J., Ala-Ruona, E., Pontio, I., Tervaniemi, M.; Gold, C. Individual music therapy for depression: Randomised controlled trial. Br. J. Psychiatry 2011, 199, 132–139. [Google Scholar] [CrossRef]
- Fedotova, J.; Ordyan, N. Involvement of D1 receptors in depression-like behavior of ovariectomized rats. Acta Physiol. Hung. 2011, 98, 165–176. [Google Scholar] [CrossRef] [PubMed]
- File, S.P. a. S. E. Anxiolytic and Anxiogenic Drug Effects on Exploratory Activity in an Elevated Plus-Maze: A Novel Test of Anxiety in the Rat. Pharmacol. Biochem. 1986, 24. [Google Scholar]
- Fu, Q. , Qiu, R., Chen, L., Chen, Y., Qi, W.; Cheng, Y. Music prevents stress-induced depression and anxiety-like behavior in mice. Transl. Psychiatry 2023, 13, 317. [Google Scholar] [CrossRef] [PubMed]
- Gameiro, G.H. , Gameiro, P.H., Andrade Ada, S., Pereira, L.F., Arthuri, M.T., Marcondes, F.K.; Veiga, M.C. Nociception- and anxiety-like behavior in rats submitted to different periods of restraint stress. Physiol. Behav. 2006, 87, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y. , Wei, Y., Yang, W., Jiang, L., Li, X., Ding, J.; Ding, G. The Effectiveness of Music Therapy for Terminally Ill Patients: A Meta-Analysis and Systematic Review. J. Pain. Symptom Manag. 2019, 57, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Giedd, J.N. , Blumenthal, J., Jeffries, N.O., Castellanos, F.X., Liu, H., Zijdenbos, A., Rapoport, J.L. Brain development during childhood and adolescence: A longitudinal MRI study. Nat. Neurosci. 1999, 2. [Google Scholar]
- Gong, X. , Fenech, B., Blackmore, C., Chen, Y., Rodgers, G., Gulliver, J.; Hansell, A.L. Association between Noise Annoyance and Mental Health Outcomes: A Systematic Review and Meta-Analysis. Int J Environ Res Public Health 2022, 19. [CrossRef]
- Guyer, A.E. , Monk, C.S., McClure-Tone, E.B., Nelson, E.E., Roberson-Nay, R., Adler, A.D., … Ernst, M. A Developmental Examination of Amygdala Response to Facial Expressions. J. Cogn. Neurosci. 2008, 20. [Google Scholar]
- Hare, B.D. , Shinohara, R., Liu, R.J., Pothula, S., DiLeone, R.J.; Duman, R.S. Optogenetic stimulation of medial prefrontal cortex Drd1 neurons produces rapid and long-lasting antidepressant effects. Nat. Commun. 2019, 10, 223. [Google Scholar] [CrossRef]
- Hasbi, A. , Nguyen, T., Rahal, H., Manduca, J.D., Miksys, S., Tyndale, R.F.; George, S.R. Sex difference in dopamine D1-D2 receptor complex expression and signaling affects depression- and anxiety-like behaviors. Biol. Sex. Differ. 2020, 11, 8. [Google Scholar] [CrossRef]
- Hegewald, J. , Schubert, M., Freiberg, A., Romero Starke, K., Augustin, F., Riedel-Heller, S.G.; Seidler, A. Traffic Noise and Mental Health: A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public. Health 2020, 17. [Google Scholar] [CrossRef]
- Heldt, S.A. , Zimmermann, K., Parker, K., Gaval, M., Weinshenker, D.; Ressler, K.J. BDNF deletion or TrkB impairment in amygdala inhibits both appetitive and aversive learning. J. Neurosci. 2014, 34, 2444–2450. [Google Scholar] [CrossRef]
- Hines, M.; McFerran, K.S. Metal made me who I am: Seven adult men reflect on their engagement with metal music during adolescence. Int. J. Community Music 2014, 7, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Kunugi, H. The efficacy of pramipexole, a dopamine receptor agonist, as an adjunctive treatment in treatment-resistant depression: An open-label trial. ScientificWorldJournal 2012, 2012, 372474. [Google Scholar] [CrossRef] [PubMed]
- Jaehne, E.J. , Kent, J.N., Antolasic, E.J., Wright, B.J., Spiers, J.G., Creutzberg, K.C.; van den Buuse, M. Behavioral phenotyping of a rat model of the BDNF Val66Met polymorphism reveals selective impairment of fear memory. Transl. Psychiatry 2022, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Jirkof, P. Burrowing and nest building behavior as indicators of well-being in mice. J. Neurosci. Methods 2014, 234, 139–146. [Google Scholar] [CrossRef]
- Karege, F. , Perret, G., Bondolfi, G., Schwald, M., Bertschy, G., Aubry, J.-M. Decreased serum brain-derived neurotrophic factor levels in major depressed patients. Psychiatry Res. 2002, 109. [Google Scholar]
- Kokkinou, M. , Ashok, A.H.; Howes, O.D. The effects of ketamine on dopaminergic function: Meta-analysis and review of the implications for neuropsychiatric disorders. Mol. Psychiatry 2018, 23, 59–69. [Google Scholar] [CrossRef]
- Kompagne, H. , Bardos, G., Szenasi, G., Gacsalyi, I., Harsing, L.G.; Levay, G. Chronic mild stress generates clear depressive but ambiguous anxiety-like behaviour in rats. Behav. Brain Res. 2008, 193, 311–314. [Google Scholar] [CrossRef]
- Kozisek, M.E. , Middlemas, D.; Bylund, D.B. Brain-derived neurotrophic factor and its receptor tropomyosin-related kinase B in the mechanism of action of antidepressant therapies. Pharmacol. Ther. 2008, 117, 30–51. [Google Scholar] [CrossRef]
- Kraeuter, A.K. , Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-Like Behavior. Methods Mol. Biol. 2019, 1916, 99–103. [Google Scholar] [CrossRef]
- Lazenka, M.F. , Freitas, K.C., Henck, S.; Negus, S.S. Relief of Pain-Depressed Behavior in Rats by Activation of D1-Like Dopamine Receptors. J. Pharmacol. Exp. Ther. 2017, 362, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.T. , Yang, P., Lai, C.Y., Su, Y.Y., Yeh, Y.C., Huang, M.F.; Chen, C.C. Mental health implications of music: Insight from neuroscientific and clinical studies. Harv. Rev. Psychiatry 2011, 19, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y. , Yin, C.Y., Zhu, L.J., Zhu, X.H., Xu, C., Luo, C.X.; Zhou, Q.G. Sucrose preference test for measurement of stress-induced anhedonia in mice. Nat. Protoc. 2018, 13, 1686–1698. [Google Scholar] [CrossRef] [PubMed]
- Mark, A.; Smith, S.M. , Richard Kvetnansky, and Robert, M. Post. Stress and glucocorticoids affect the expression of brain-derived neurotrophic factor and neurotrophin-3 mRNAs in the hippocampus. J. Neurosci. 1995, 15. [Google Scholar]
- Martin, G. , Clarke, M.; Pearce, C. Adolescent Suicide: Music Preference as an Indicator of Vulnerability. J. Am. Acad. Child Adolesc. Psychiatry 1993, 32. [Google Scholar]
- McCormick, C.M.; Green, M.R. From the stressed adolescent to the anxious and depressed adult: Investigations in rodent models. Neuroscience 2013, 249, 242–257. [Google Scholar] [CrossRef]
- Miranda, D.; Claes, M. Rap Music Genres and Deviant Behaviors in French-Canadian Adolescents. J. Youth Adolesc. 2004, 33. [Google Scholar] [CrossRef]
- Moraga-Amaro, R. , Gonzalez, H., Pacheco, R.; Stehberg, J. Dopamine receptor D3 deficiency results in chronic depression and anxiety. Behav. Brain Res. 2014, 274, 186–193. [Google Scholar] [CrossRef]
- Mossler, K. , Chen, X., Heldal, T.O.; Gold, C. Music therapy for people with schizophrenia and schizophrenia-like disorders. Cochrane Database Syst Rev 2011, 12, CD004025. [Google Scholar] [CrossRef]
- Nair, A. , Vadodaria, K.C., Banerjee, S.B., Benekareddy, M., Dias, B.G., Duman, R.S.; Vaidya, V.A. Stressor-specific regulation of distinct brain-derived neurotrophic factor transcripts and cyclic AMP response element-binding protein expression in the postnatal and adult rat hippocampus. Neuropsychopharmacology 2007, 32, 1504–1519. [Google Scholar] [CrossRef]
- Nguyen, C. , Mondoloni, S., Le Borgne, T., Centeno, I., Come, M., Jehl, J.; Faure, P. Nicotine inhibits the VTA-to-amygdala dopamine pathway to promote anxiety. Neuron 2021, 109, 2604–2615. [Google Scholar] [CrossRef] [PubMed]
- Paus, T. , Keshavan, M. . Nature Reviews Neuroscience 2008, 9. [Google Scholar]
- Peng, X. , Mao, Y., Tai, Y., Luo, B., Dai, Q., Wang, X.; Wang, H. Characterization of Anxiety-Like Behaviors and Neural Circuitry following Chronic Moderate Noise Exposure in Mice. Environ. Health Perspect. 2023, 131, 107004. [Google Scholar] [CrossRef] [PubMed]
- Perreault, M.L. , Jones-Tabah, J., O’Dowd, B.F.; George, S.R. A physiological role for the dopamine D5 receptor as a regulator of BDNF and Akt signalling in rodent prefrontal cortex. Int. J. Neuropsychopharmacol. 2013, 16, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Peter, R., H. Synaptic density in human frontal cortex — Developmental changes and effects of aging. Brain Research 1979, 163. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, J.H. , Masten, C.L., Moore, W.E., 3rd, Oswald, T.M., Mazziotta, J.C., Iacoboni, M.; Dapretto, M. Entering adolescence: Resistance to peer influence, risky behavior, and neural changes in emotion reactivity. Neuron 2011, 69, 1029–1036. [Google Scholar] [CrossRef]
- Recours, R. , Aussaguel, F.; Trujillo, N. Metal music and mental health in France. Cult. Med. Psychiatry 2009, 33, 473–488. [Google Scholar] [CrossRef]
- Roger, D.; Porsolt, G.A.; Nadine, B.; Maurice, J. Behavioural despair in rats: A new model sensitive to antidepressant treatments. Eur. J. Pharmacol. 1978, 47. [Google Scholar]
- Rygula, R. , Abumaria, N., Flugge, G., Fuchs, E., Ruther, E.; Havemann-Reinecke, U. Anhedonia and motivational deficits in rats: Impact of chronic social stress. Behav. Brain Res. 2005, 162, 127–134. [Google Scholar] [CrossRef]
- Shimizu, E. , Hashimoto, K., Okamura, N., Koike, K., Komatsu, N., Kumakiri, C.; Iyo, M. Alterations of serum levels of brain-derived neurotrophic factor (BDNF) in depressed patients with or without antidepressants. Biol. Psychiatry 2003, 54, 70–75. [Google Scholar] [CrossRef]
- Shinohara, R. , Taniguchi, M., Ehrlich, A.T., Yokogawa, K., Deguchi, Y., Cherasse, Y.; Furuyashiki, T. Dopamine D1 receptor subtype mediates acute stress-induced dendritic growth in excitatory neurons of the medial prefrontal cortex and contributes to suppression of stress susceptibility in mice. Mol. Psychiatry 2018, 23, 1717–1730. [Google Scholar] [CrossRef] [PubMed]
- Shiranibidabadi, S.; Mehryar, A. Music therapy as an adjunct to standard treatment for obsessive compulsive disorder and co-morbid anxiety and depression: A randomized clinical trial. J. Affect. Disord. 2015, 184, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Tsankova, N.M. , Berton, O., Renthal, W., Kumar, A., Neve, R.L.; Nestler, E.J. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat. Neurosci. 2006, 9, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Vyas, A. , Pillai, A.G.; Chattarji, S. Recovery after chronic stress fails to reverse amygdaloid neuronal hypertrophy and enhanced anxiety-like behavior. Neuroscience 2004, 128, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Wahlstrom, D. , White, T.; Luciana, M. Neurobehavioral evidence for changes in dopamine system activity during adolescence. Neurosci. Biobehav. Rev. 2010, 34, 631–648. [Google Scholar] [CrossRef]
- Wang, J. , Lai, S., Wang, R., Zhou, T., Dong, N., Zhu, L.; Chen, Y. Dopamine D3 receptor in the nucleus accumbens alleviates neuroinflammation in a mouse model of depressive-like behavior. Brain Behav. Immun. 2022, 101, 165–179. [Google Scholar] [CrossRef]
- Warner-Schmidt, J.L.; Duman, R.S. Hippocampal neurogenesis: Opposing effects of stress and antidepressant treatment. Hippocampus 2006, 16, 239–249. [Google Scholar] [CrossRef]
- Yukihiko Shirayama, A.C.-H. C. , Shin Nakagawa, David S Russell, Ronald S Duman. Brain-derived neurotrophic factor produces antidepressant effects in behavioral models of depression. J. Neurosci. 2002, 22. [Google Scholar]
- Zhang, T. , Hong, J., Di, T.; Chen, L. MPTP Impairs Dopamine D1 Receptor-Mediated Survival of Newborn Neurons in Ventral Hippocampus to Cause Depressive-Like Behaviors in Adult Mice. Front. Mol. Neurosci. 2016, 9, 101. [Google Scholar] [CrossRef]
- Zhe-Yu Chen, D.J. , Kevin, G.; Bath, Alessandro Ieraci, Tanvir Khan, ChiaJen Siao, Daniel, G.; Herrera, Miklos Toth, Chingwen Yang, Bruce, S. McEwen, Barbara, L. Hempstead, and Francis, S. Lee. Genetic Variant BDNF (Val66Met) Polymorphism Alters AnxietyRelated Behavior. Science 2006, 314. [Google Scholar]
- Zhu, Y. , Wang, R., Tang, X., Li, Q., Xu, G.; Zhang, A. The effect of music, massage, yoga and exercise on antenatal depression: A meta-analysis. J. Affect. Disord. 2021, 292, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Zofia Rogó, G. y. S. , Aleksandra K³odziñs. Anxiolytic- and Antidepressant-like Effects of 7-OH-DPAT, Preferential Dopamine D3 Receptor Agonist, in Rats. Pol. J. Pharmocology 2004, 56. [Google Scholar]
Figure 1.
Sucrose preference is calculated per day for all four groups over six days. There is a reduction in sucrose preferences between WT and HM mice or CN mice, while no difference was observed in sucrose preference between WT and HH mice. All values were presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
Figure 1.
Sucrose preference is calculated per day for all four groups over six days. There is a reduction in sucrose preferences between WT and HM mice or CN mice, while no difference was observed in sucrose preference between WT and HH mice. All values were presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
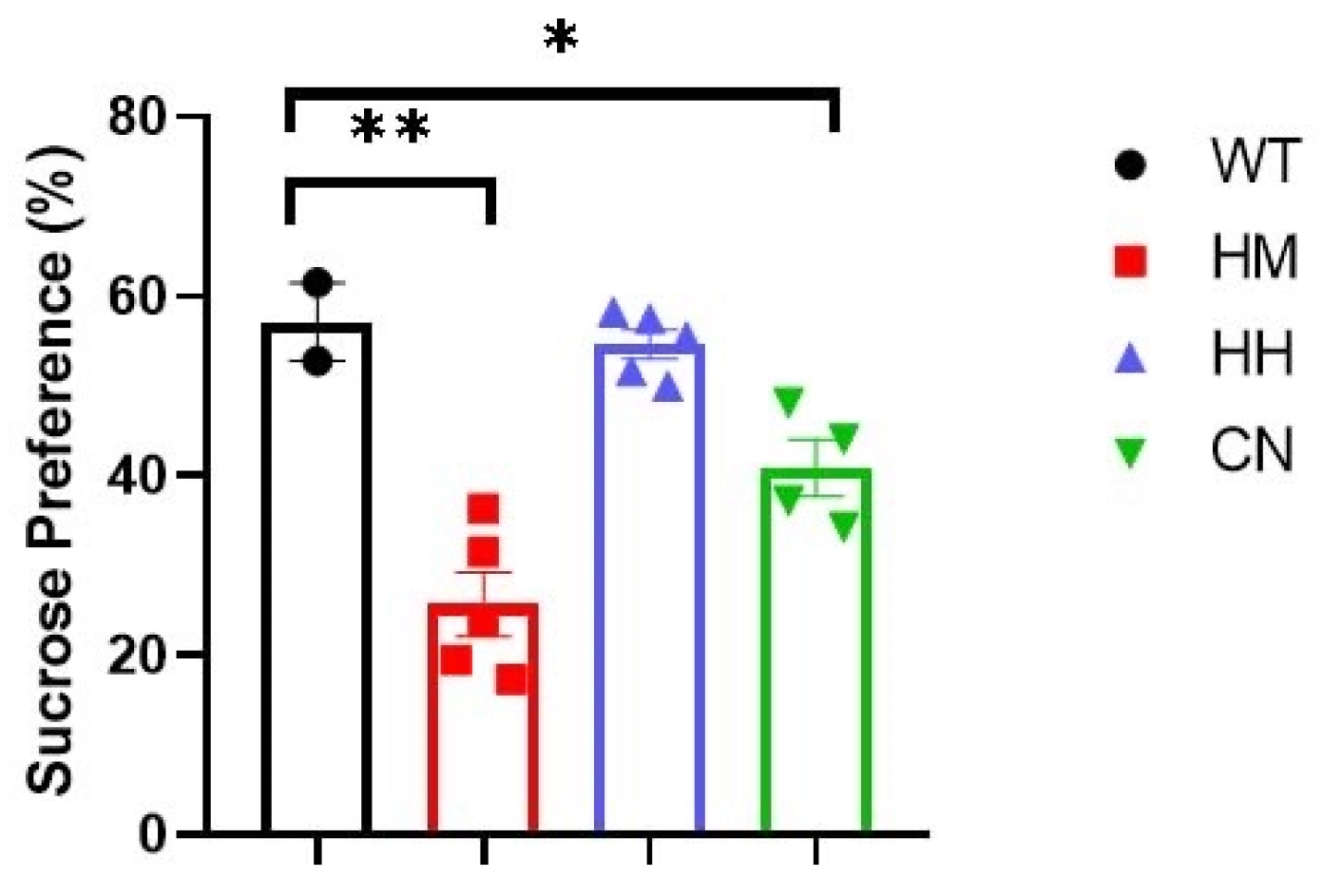
Figure 2.
Tail suspension test (TST) is performed for all four groups after sound exposure for one week. WT mice showed longer mobility time than HH mice, HM mice, and CN mice. All values were presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
Figure 2.
Tail suspension test (TST) is performed for all four groups after sound exposure for one week. WT mice showed longer mobility time than HH mice, HM mice, and CN mice. All values were presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
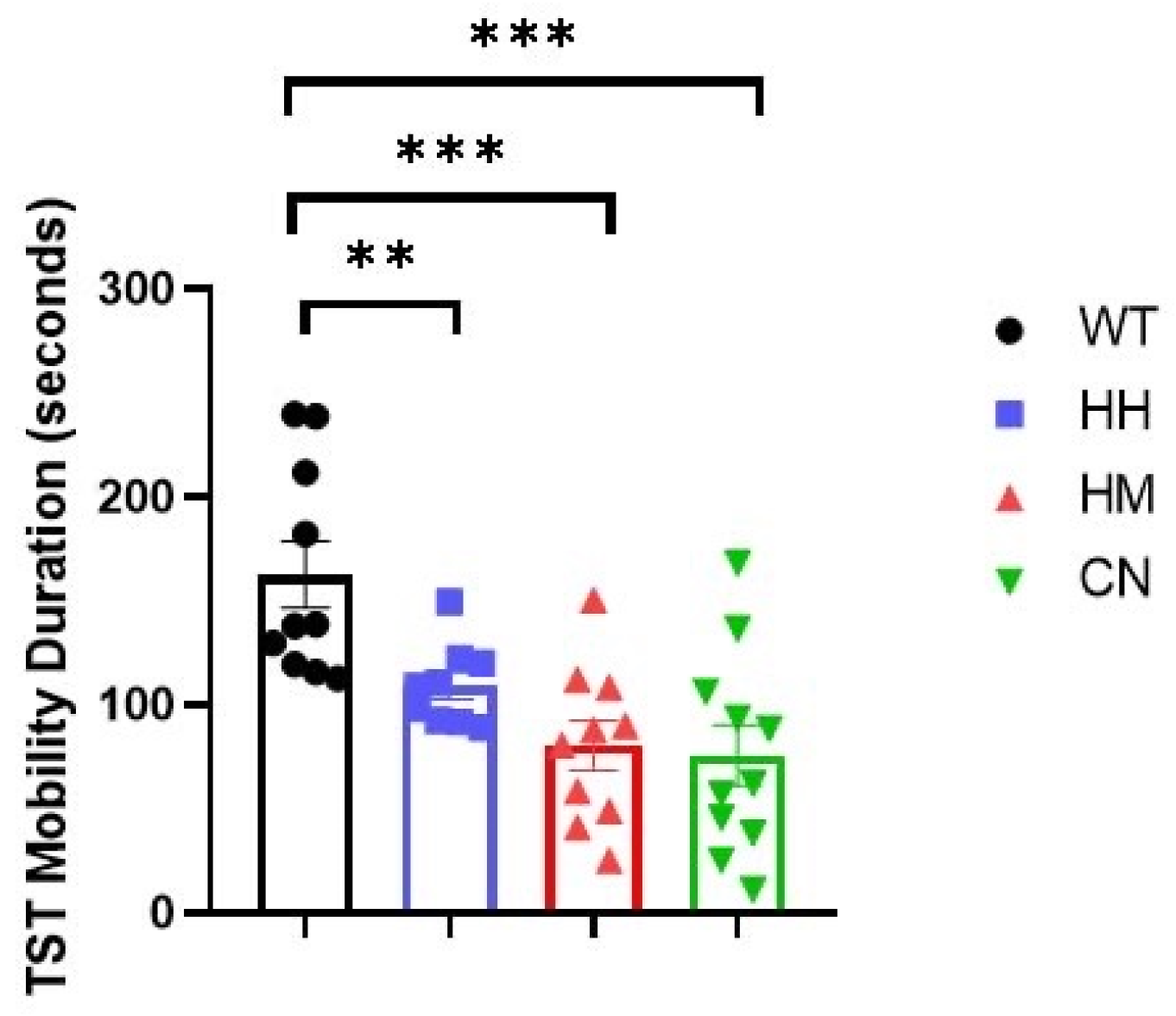
Figure 3.
Forced swim test (FST) is performed for all four groups after sound exposure for one week. No difference was observed between the mobility time of WT mice and the mobility time of the other three groups (HH mice, HM mice, or CN mice). All values were presented as mean ± SEM.
Figure 3.
Forced swim test (FST) is performed for all four groups after sound exposure for one week. No difference was observed between the mobility time of WT mice and the mobility time of the other three groups (HH mice, HM mice, or CN mice). All values were presented as mean ± SEM.
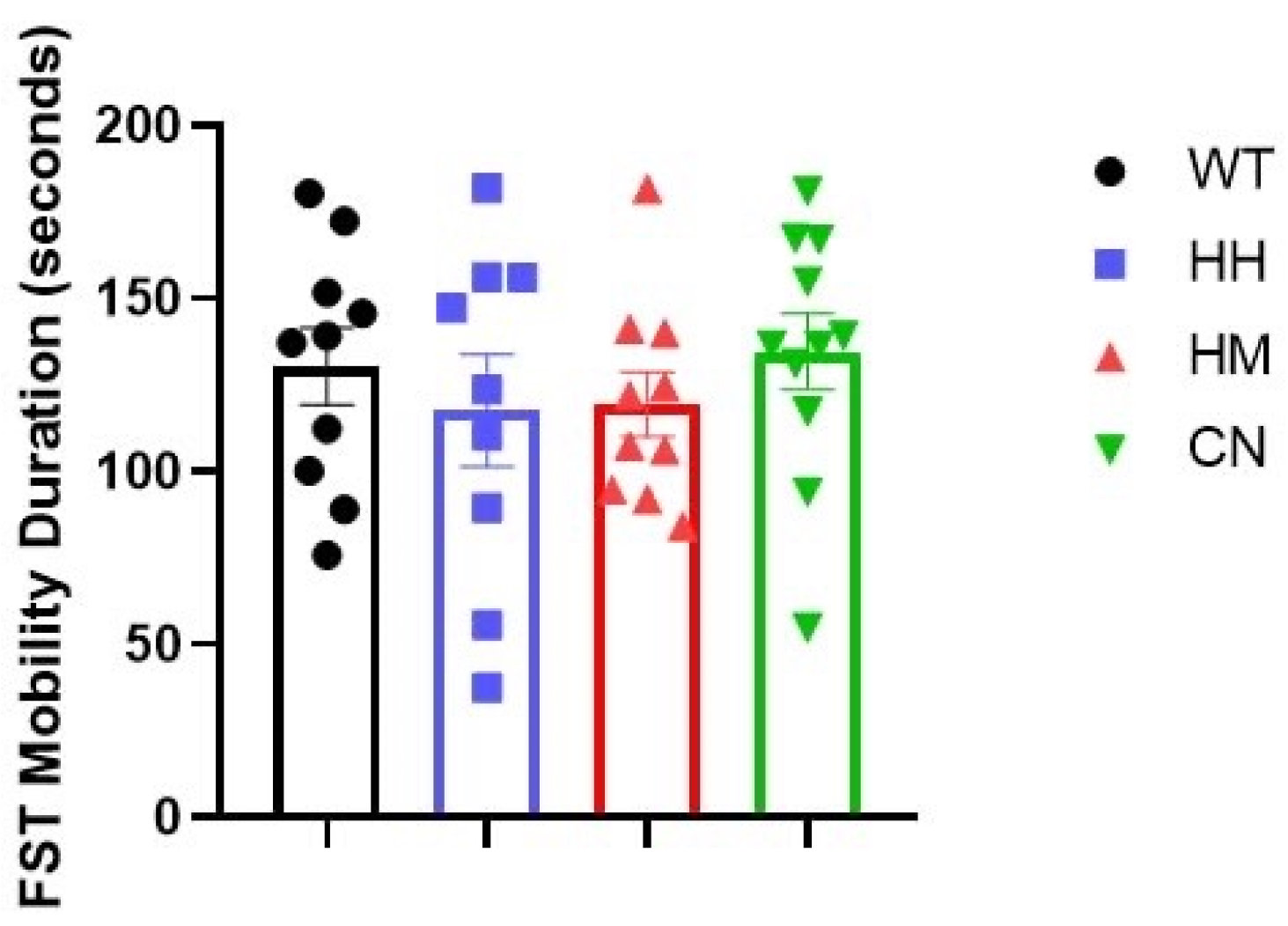
Figure 4.
Nest Building (NB) is performed for all four groups after sound exposure for one week. No difference in performances evaluated by the standard rating scale was observed between WT mice and the other three groups (HH mice, HM mice, or CN mice). All values were presented as mean ± SEM.
Figure 4.
Nest Building (NB) is performed for all four groups after sound exposure for one week. No difference in performances evaluated by the standard rating scale was observed between WT mice and the other three groups (HH mice, HM mice, or CN mice). All values were presented as mean ± SEM.
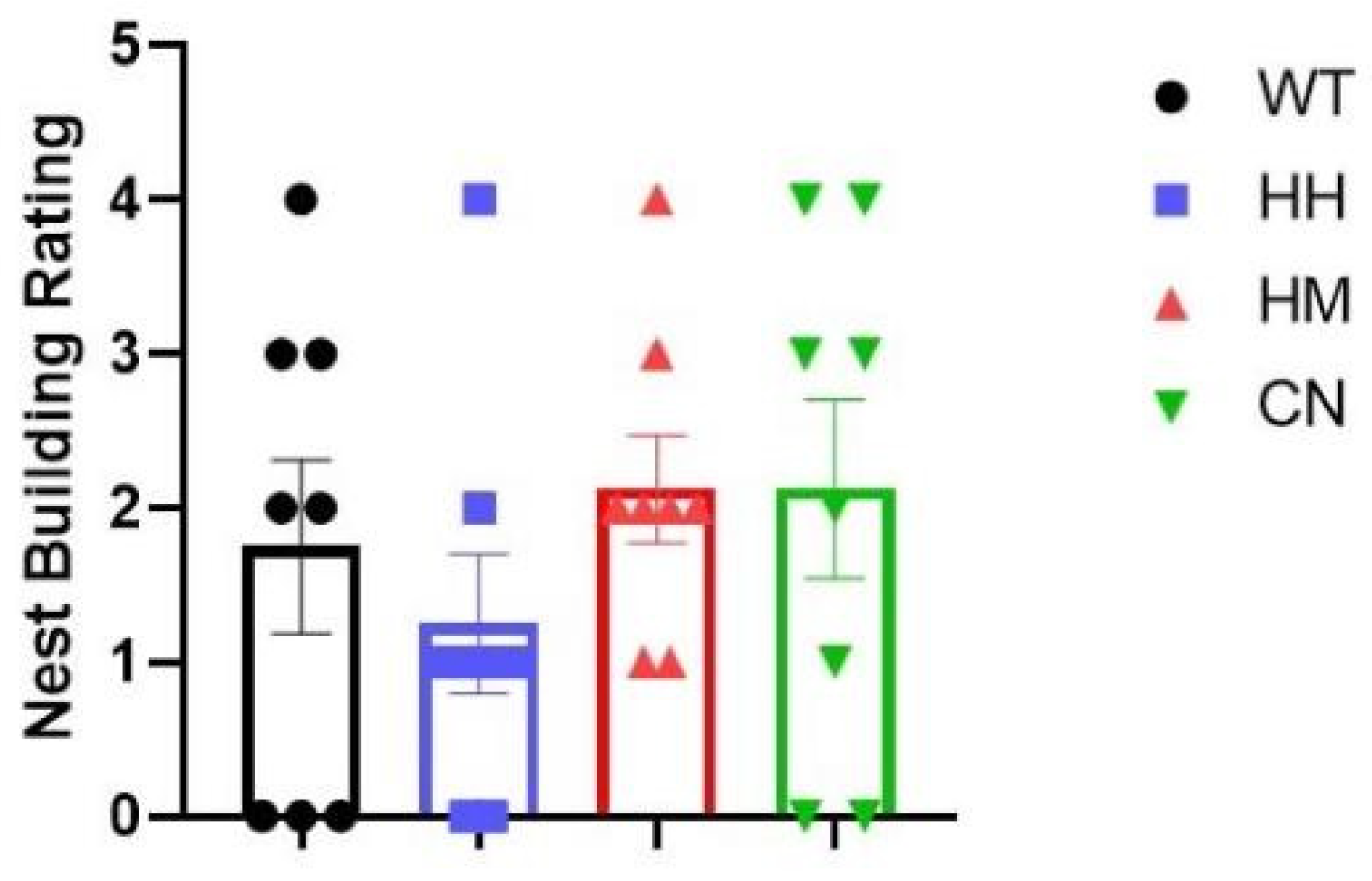
Figure 5.
Elevated plus maze (EPM) is performed for all four groups after sound exposure for one week. (a) WT mice showed shorter time spent in the open arms compared with the other three groups (HH mice, HM mice, and CN mice). (b) WT mice showed shorter distance traveled in the open arms compared with the other three groups (HH mice, HM mice, and CN mice) All values were presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
Figure 5.
Elevated plus maze (EPM) is performed for all four groups after sound exposure for one week. (a) WT mice showed shorter time spent in the open arms compared with the other three groups (HH mice, HM mice, and CN mice). (b) WT mice showed shorter distance traveled in the open arms compared with the other three groups (HH mice, HM mice, and CN mice) All values were presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
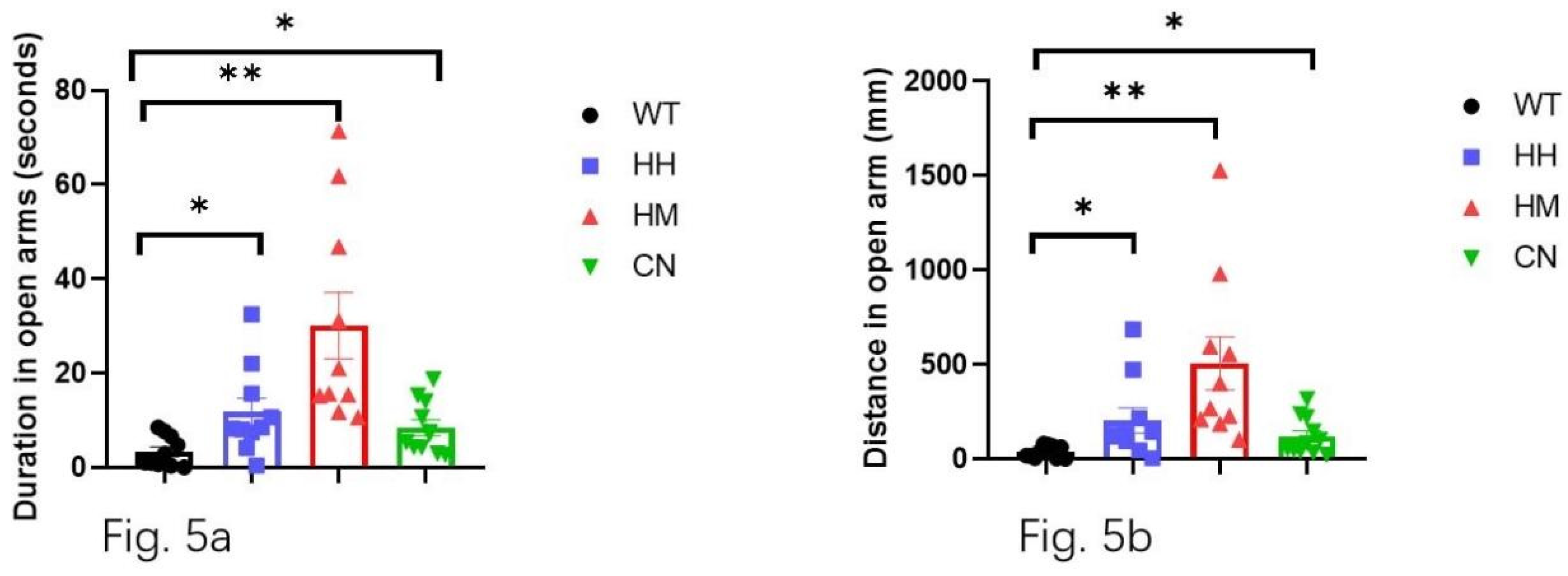
Figure 6.
Open field test (OFT) is performed for all four groups after sound exposure for one week. (a) WT mice showed no significant difference in time spent in the center of the field compared with the other three groups (HH mice, HM mice, or CN mice). (b) WT mice showed no significant difference in distance traveled in the center of the field compared with the other three groups (HH mice, HM mice, or CN mice) All values were presented as mean ± SEM.
Figure 6.
Open field test (OFT) is performed for all four groups after sound exposure for one week. (a) WT mice showed no significant difference in time spent in the center of the field compared with the other three groups (HH mice, HM mice, or CN mice). (b) WT mice showed no significant difference in distance traveled in the center of the field compared with the other three groups (HH mice, HM mice, or CN mice) All values were presented as mean ± SEM.
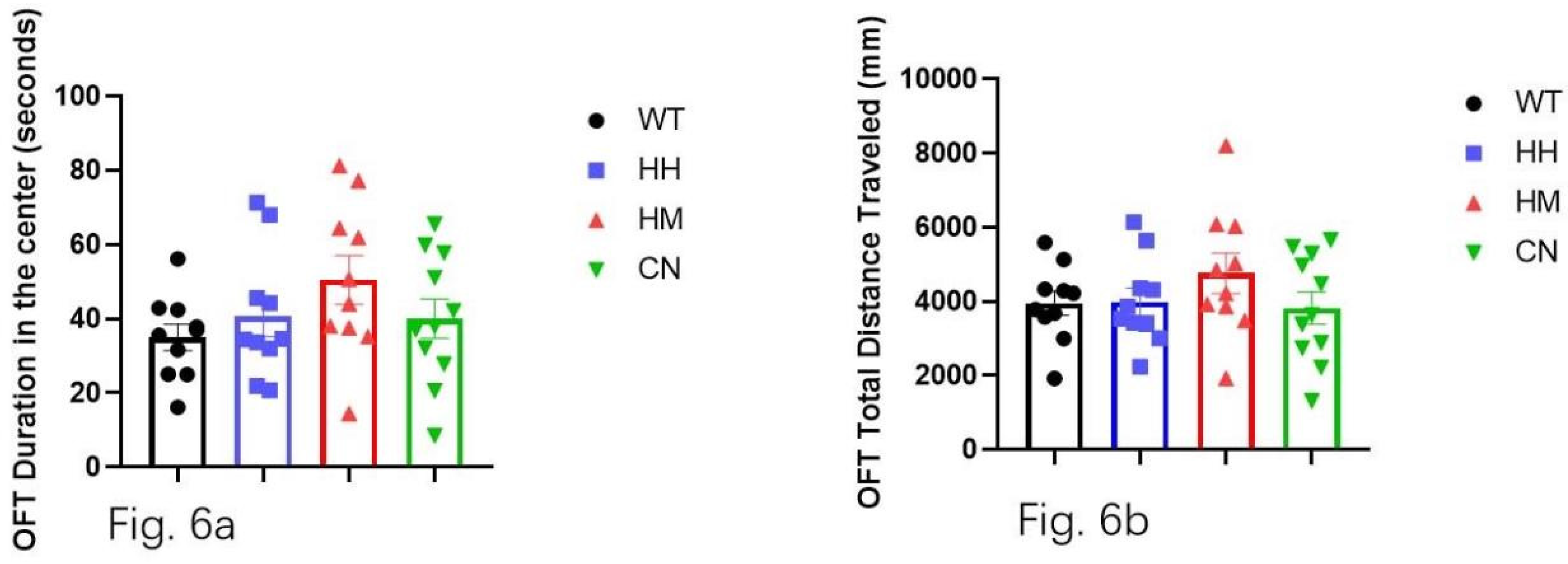
Figure 7.
DR1 concentration in the mixture of the frontal lobe and the hippocampus is measured after sound exposure for one week. HH mice and CN mice indicated a reduction in DR1 while HM mice showed no significant difference in DR1 concentration compared with WT mice. All values were presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
Figure 7.
DR1 concentration in the mixture of the frontal lobe and the hippocampus is measured after sound exposure for one week. HH mice and CN mice indicated a reduction in DR1 while HM mice showed no significant difference in DR1 concentration compared with WT mice. All values were presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001.
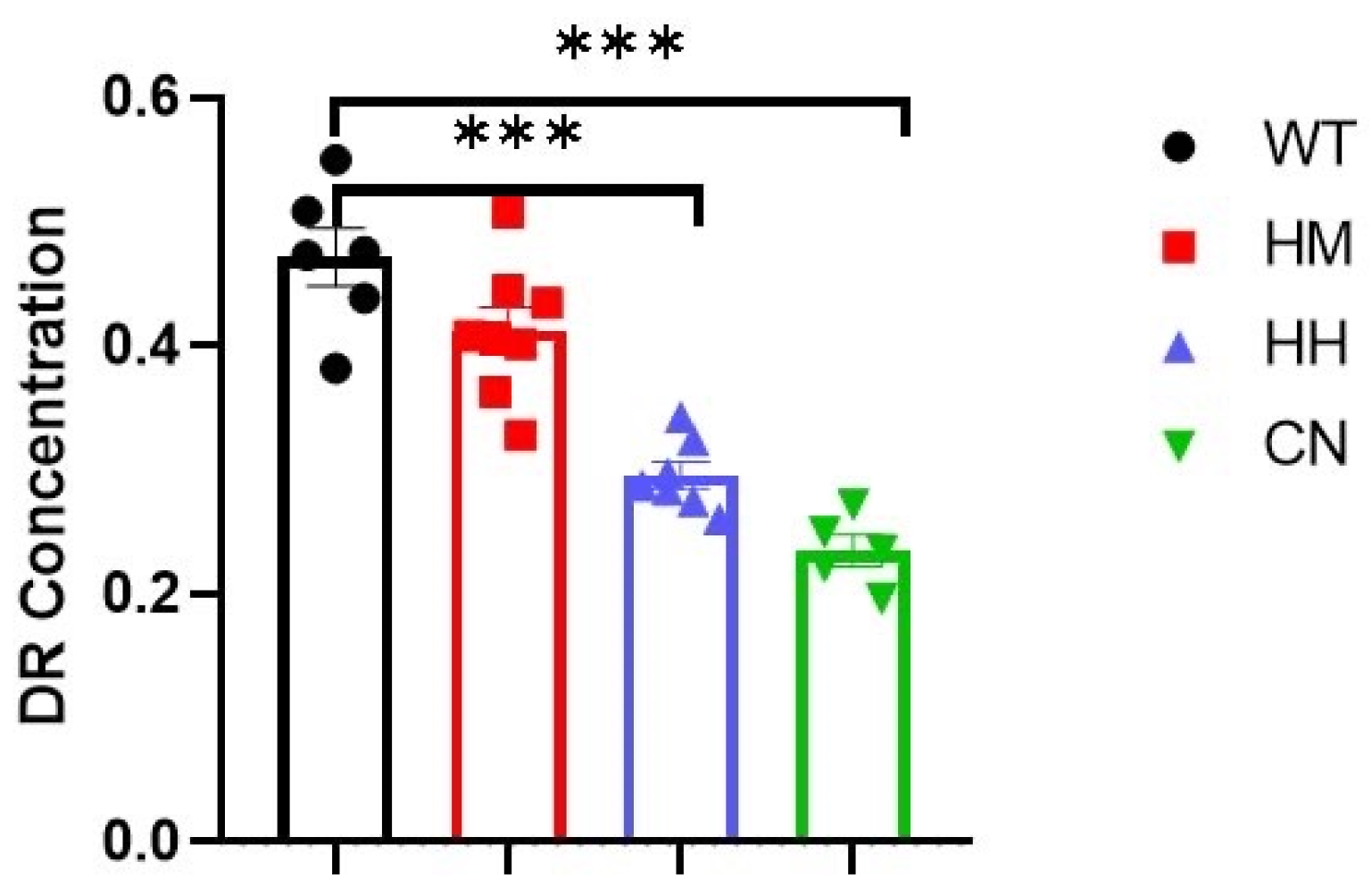
Figure 8.
BDNF concentration in the mixture of the frontal lobe and the hippocampus is measured after sound exposure for one week. The results indicate a reduction in BDNF in (b) HH mice and (c) CN mice compared with WT mice. (a) However, no significant difference is found in BDNF concentration between HM mice and WT mice.
Figure 8.
BDNF concentration in the mixture of the frontal lobe and the hippocampus is measured after sound exposure for one week. The results indicate a reduction in BDNF in (b) HH mice and (c) CN mice compared with WT mice. (a) However, no significant difference is found in BDNF concentration between HM mice and WT mice.
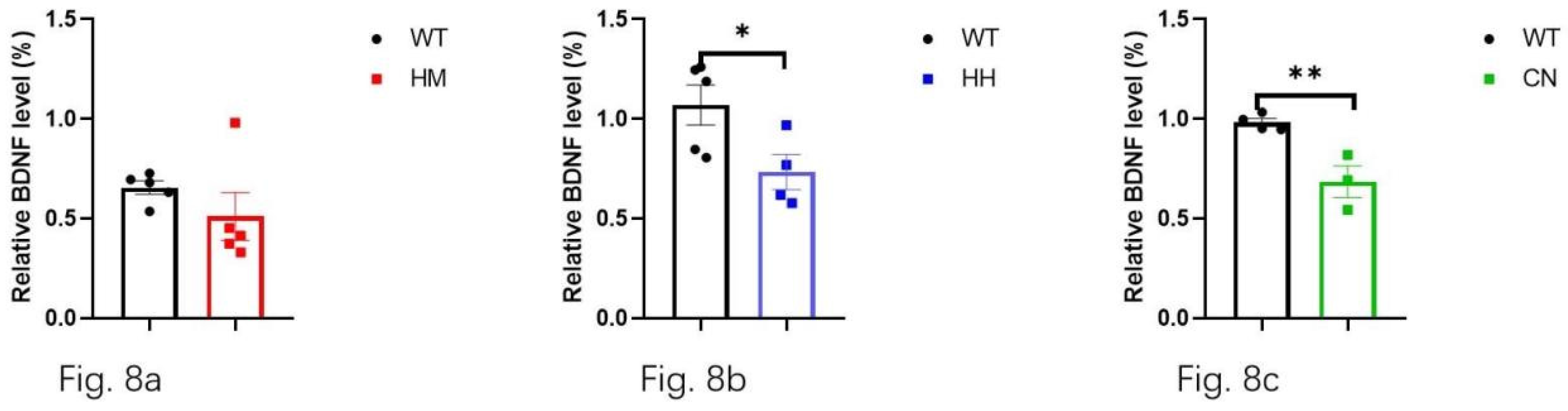
Figure 9.
Trk B concentration in the mixture of the frontal lobe and the hippocampus is measured after sound exposure for one week. No difference in Trk B concentration is found between WT mice and (a) HM mice, (b) HH mice, or (c) CN mice.
Figure 9.
Trk B concentration in the mixture of the frontal lobe and the hippocampus is measured after sound exposure for one week. No difference in Trk B concentration is found between WT mice and (a) HM mice, (b) HH mice, or (c) CN mice.
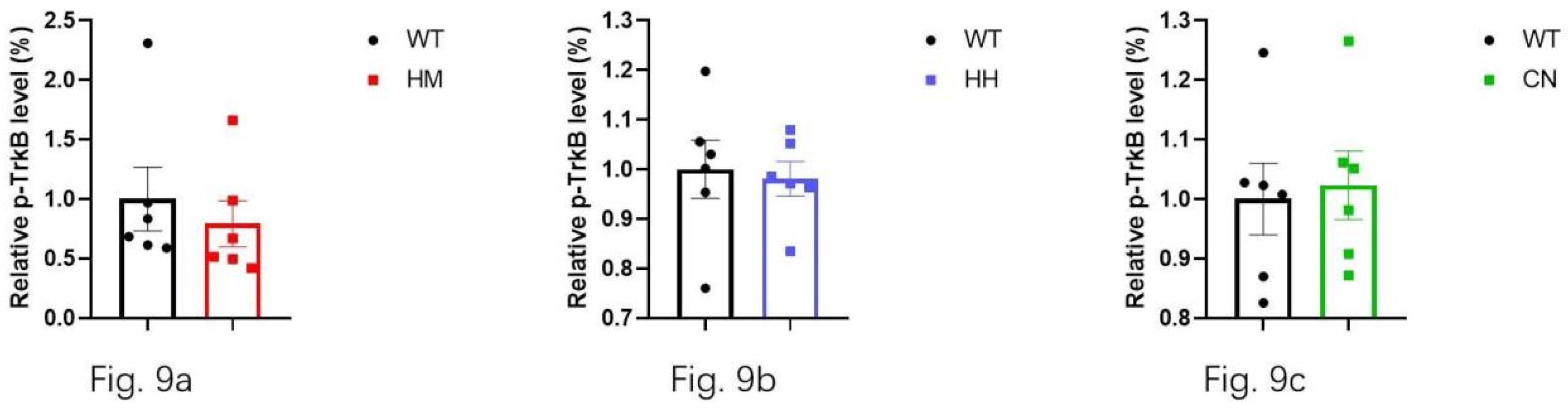
Figure 10.
5-HT concentration in the mixture of the frontal lobe and the hippocampus is measured after sound exposure for one week. No difference in 5-HT concentration is found between WT mice and HM mice, HH mice. A significant decrease in 5-HT concentration is found in CN mice compared with WT mice.
Figure 10.
5-HT concentration in the mixture of the frontal lobe and the hippocampus is measured after sound exposure for one week. No difference in 5-HT concentration is found between WT mice and HM mice, HH mice. A significant decrease in 5-HT concentration is found in CN mice compared with WT mice.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



