
Submitted:
23 September 2024
Posted:
24 September 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Adolescents most often experience mild acute COVID-19 but may still face fatigue and persistent symptoms such as post-COVID-19 condition (PCC) and post-infective fatigue syndrome (PIFS). We explored the fecal microbiota in a prospective controlled cohort of 136 non-hospitalized SARS-CoV-2-positive and 32 SARS-CoV-2-negative controls aged 12–25 years. With a targeted qPCR approach, the quantities of 100 fecal bacterial taxa were measured at baseline (early convalescent stage) and after six months. Results: Faecalibacterium prausnitzii M21.2 and Gemmiger formicilis (both p<0.001) were enriched in the SARS-CoV-2-positive participants compared to controls at baseline. Lower baseline abundance of Faecalibacterium prausnitzii M21/2 (p=0.013) and higher abundance of Clostridium spiroforme (p=0.006), Sutterella wadsworthensis (p<0.001) and Streptococcus thermophilus (p=0.039) were associated with six-month fatigue. Sutterella wadsworthensis and Streptococcus thermophilus enrichment was additionally associated with PCC in the SARS-CoV-2-positive group (p<0.001 and 0.042 respectively). Conclusions: In mild SARS-CoV-2 infection in adolescents and young adults, beneficial Faecalibacterium prausnitzii M21/2 and Gemmiger formicilis were enriched compared to SARS-CoV-2 negative controls. Additionally, both known (Faecalibacterium prausnitzii, Streptococcus thermophilus) and new (Clostridium spiroforme, Sutterella wadsworthensis) bacteria were associated with persistent symptoms in the COVID-19 infected group, warranting further exploration of the role of these bacteria in COVID-19 disease and PCC pathophysiology.
Keywords:
SARS-CoV-2
; COVID-19
; fatigue
; post-COVID-19 condition
; post-infective fatigue syndrome
; microbiota
; Faecalibacterium prausnitzii
; Sutterella wadsworthensis
1. Introduction
Individuals affected by COVID-19 (SARS-CoV-2 infection) can exhibit a broad spectrum of post-COVID problems. Common persistent symptoms include fatigue, dyspnea, and cognitive dysfunction, and have an impact on everyday functioning [1]. Up to three-quarters of patients affected by COVID-19 describe at least one persisting symptom six months after infection, with fatigue being the most frequently reported [2,3,4]. The World Health organization (WHO) has defined long-lasting symptoms following confirmed or suspected COVID-19 infection, with no alternative diagnosis to explain them and with a negative impact on everyday functioning, as post-COVID-19 condition (PCC) [5]. PCC exhibits significant clinical overlap with post-infective fatigue syndrome (PIFS) [6], which is defined as persistent fatigue affecting daily activities, in addition to other symptoms, following an acute infectious event [7]. Several studies indicate that the gut microbiota might contribute to COVID-19 pathogenesis and disease outcomes [8]. SARS-CoV-2 enters cells primarily through binding to angiotensin-converting enzyme 2 (ACE2) receptors which are expressed both in the respiratory and gastrointestinal tract [9]. Patients with gut microbiome dysbiosis have an increased expression of ACE2 on their mucosal cells [10], making them more vulnerable to an infection by fecal SARS-CoV-2 [11]. SARS-CoV-2 infection has been associated with altered intestinal microbiota, in adult patients characterized by enrichment of opportunistic pathogens and depletion of beneficial commensals [11,12,13]. Several gut commensals with known immunomodulatory potential such as Faecalibacterium prausnitzii (F. prausnitzii), Eubacterium rectale and Bifidobacteria species have been shown to be under-represented in adult patients infected with SARS-CoV-2 [14]. Healthy children and adolescents are usually mildly affected by SARS-CoV-2 in the acute phase of the infection and do not require hospitalization [15,16,17,18]. Children may still face post-acute illness, as individuals with mild disease seem to have the same risk of post-COVID-19 condition as hospitalized patients [19]. There is increasing evidence of an association between certain gut bacterial profiles and severity of acute COVID-19 in adult patients [14,20,21]. In addition, gut dysbiosis seem to persist after disease resolution [22,23]. Knowledge on microbiota alterations in children and young adults during and after mild COVID-19 is limited. The aims of the present study were to investigate associations between selected gut microbiota taxa and SARS-CoV-2 status in the early convalescent stage among non-hospitalized adolescents and young adults. Further, in the SARS-CoV-2 positive cases, we aimed to assess the strengths of associations between bacterial taxa at baseline and the development of fatigue, PCC and PIFS at six months follow-up.
2. Materials and Methods
The data in the present study is part of the Long-Term Effects of COVID-19 in Adolescents (LoTECA) project, a longitudinal observational cohort study of SARS-CoV-2 positive and negative non-hospitalized adolescents and young adults (12-25 years of age), with a total follow-up of 12 months (Clinical Trials ID: NCT04686734). Details of the LoTECA study design are previously published [24]. Clinical examination and laboratory testing, fecal sampling and completion of questionnaires, were performed at baseline (early convalescent stage) and at six months follow-up. Only participants who submitted a fecal sample at baseline were asked to deliver a follow-up sample at six months.
2.1. Study population
From December 24th, 2020, until May 18th, 2021, individuals 12-25 years old were consecutively recruited through two accredited microbiological laboratories. Individuals with laboratory-confirmed SARS-CoV-2 infection (detected by upper respiratory tract swabs followed by reverse-transcription polymerase chain reaction (RT-PCR)) were eligible for enrolment after completing quarantine (10 days). Individuals having approximately the same distribution of sex and age as the SARS-CoV-2-infected cases, but with a negative SARS-CoV-2 test from the same microbiological laboratories during the same period, were recruited as controls. Some of the controls had undergone testing due to infectious symptoms, while others were asymptomatic but had been exposed to COVID-19. The baseline assessment and inclusion took place within 28 days of a positive or negative COVID-19 test. A follow-up examination was scheduled six months after inclusion. The SARS-CoV-2 negative status was confirmed by the absence of anti-nucleocapsid antibodies at inclusion and at six months follow-up. During the first five weeks of the recruitment period, different genetic variants of the SARS-CoV-2 virus belonging to the B.1 lineage were present in this geographical area. From late-February 2021, the B.1.1.7 (alpha) variant became dominant for the remaining part of the recruitment period. Participants with gastrointestinal diseases (e.g., inflammatory bowel disease) or medications that might impact the gut microbiota (e.g., proton pump inhibitors, antibiotics) were excluded.
2.2. Investigational Program
Participants were summoned to a one-day investigational program at Akershus University Hospital, Norway. Participants underwent a clinical examination, spirometry, blood sampling, fecal sampling and completed several questionnaires, including enquiries about demographic background details and symptoms experienced during the illness episode, described in detail elsewhere [25]. The Chalder Fatigue Questionnaire (CFQ), a well-validated 11-item inventory for mental and physical fatigue [26] was used to assess fatigue at baseline and at follow-up. Each item is rated on a four-point Likert scale; in the present study, the bimodal scoring (0-0-1-1) was applied, and a sum score ≥ 4 was classified as fatigue caseness in line with current litterature recommendation [27]. The application of the WHO definition PCC [5] and the case definition for PIFS [7] at six months follow-up was performed as previously described [24]. All participants were categorized as either fatigue, PCC and PIFS cases or non-cases. This assessment was conducted independently by two researchers who were blinded to the participants’ initial SARS-CoV-2 status.
2.3. Precision fecal microbiota profiling
Fecal samples from the participants were collected on Bio-Me filter cards (Bio-Me, Oslo, Norway). The cards were frozen at - 80oC until analysis. Three 6 mm discs were punched out from each sample filter card into a MagMAX Microbiome Bead Plate (ThermoFisher Scientific). Microbiome MagMAX Ultra kit (ThermoFisher Scientific) was used for Microbial DNA extraction. Bacterial cell walls were first disrupted using the VWR Star Beater (VWR, Radnor, PA, USA) for 2 min at 30Hz and then using the KingFisher™ Flex (ThermoFisher Scientific, Waltham, MA, USA), following the manufacturer’s instructions with modifications as described by [28]. Purified DNA was eluted in 200 µL MagMAX Elution Buffer and quantified using Quant-iT PicoGreen dsDNA reagent (ThermoFisher Scientific) and a F200 Infinite plate reader (Tecan, Männedorf, Switzerland). Bio-Me’s Precision Microbiome Profiling platform (PMP™), a target method for microbiome analysis, was used to measure the absolute quantities, genome/ng DNA, of 108 targets, including 100 different bacterial species and strains (hereby referred to as bacterial taxa) (For details see Supplementary Table S1). These taxa, known from the literature to be important for gut health and disease, were measured using quantitative PCR with reference to known concentrations of each target bacteria [29,30]. The microbiome profile was assessed using abundance of single taxa and diversity measures. Richness was assessed with PMP-alpha diversity (meaning the hits among the targeted bacteria, count of detected bacteria in each sample).
2.4. Statistical analyses
The microbiota in fecal samples were analyzed at baseline and at six months follow-up from SARS-CoV-2 positive and SARS-CoV-2 negative participants. The data were preprocessed, and probes/variables where bacteria were detected in less than 15% of the samples, had a minimum standard deviation less than 450 genomes/ng DNA and a maximum standard deviation less than 5000 genomes/ng DNA, or had an interquartile range (IQR) and median of 0(0.0), were excluded, leaving 95 bacterial taxa for further analyses. Categorical variables are presented as counts and percentages. Continuous variables are presented with median and range. Crude differences between groups were assessed using Mann-Whitney U and χ2, or Fisher’s exact tests. Pairs of variables assessed at baseline and six months were analyzed with Wilcoxon signed ranks test. P-values <0.05 were considered statistically significant. The study was regarded as exploratory; therefore, we did not adjust for multiple testing. All statistical analyses were performed using SPSS statistical software and Stata version D16.
2.5. Ethical considerations
The project was conducted according to the Helsinki declaration. The project was approved by the Regional Committee for Ethics in Medical Research (ref. #203645). Written informed consent was obtained from legal guardians for participants < 16 years, or from the participants themselves when older than 16 years.
3. Results
At baseline, fecal samples were collected from 145 (36%) out of 404 SARS-CoV-2 positive and from 32 (30%) out of 105 SARS-CoV-2 negative participants. At six months, among those who provided baseline samples, 107 (74%) of the 145 SARS-CoV-2 positive and 19 (59%) of the 32 SARS-CoV-2 negative participants submitted a follow-up fecal sample. After excluding three participants with other diseases and/or medications, and six participants whose samples did not pass the laboratory quality control steps, we analyzed the fecal microbiota of 136 SARS-CoV-2 positive and 32 SARS-CoV-2 negative at baseline and of 102 SARS-CoV-2 positive and 17 SARS-CoV-2 negative at six months follow-up (Table 1).
Principal component analysis did not reveal any bacteria clusters separating the SARS-CoV-2 positive and negative participants (Supplementary Figure S1), nor was there a difference in bacterial richness between the two groups of participants at baseline (Supplementary Figure S2). However, the abundances of 14 bacterial taxa were statistically different between samples from SARS-CoV-2 positive compared to SARS-CoV-2 negative participants (Table 2), with an enrichment of six taxa, including F. prausnitzii M21.2 (Figure 1A), Gemmiger formicilis (G. formicilis) (Figure 1B), Holdemanella biformis (H. biformis), Flavonifractor plauti (F. plauti) and Eggerthella lenta (E. lenta) in the SARS-CoV-2 positive participants. Eight species had significantly reduced bacterial abundances (Table 2), including Phocaeicola massiliensis (P. massiliensis), Odoribacter splanchnicus (O. splanchnicus), Alistipes shahii and Bifidobacterium longum (B. longum).
3.1. Fecal bacterial abundances and SARS-CoV-2 status at six months
In fecal samples collected at six months follow-up, seven bacterial taxa revealed diverging abundances between SARS-COV-2 positive participants and controls (Table 2). These taxa were different from those found separating the two groups at baseline.

Table 2.
Bacterial taxa with an increased (↑) or a decreased (↓) abundance (ng/genome DNA) in SARS-CoV-2 positive participants compared to SARS-CoV-2 negative controls at baseline and at six months follow-up.
Table 2.
Bacterial taxa with an increased (↑) or a decreased (↓) abundance (ng/genome DNA) in SARS-CoV-2 positive participants compared to SARS-CoV-2 negative controls at baseline and at six months follow-up.
| Bacterial taxa with deviating abundance in SARS-CoV-2 positive compared to SARS-CoV-2 negative | ↑ or ↓ in SARS-CoV-2 positive at baseline |
↑ or ↓ in SARS-CoV-2 positive at six months |
p-value |
|---|---|---|---|
| Faecalibacterium prausnitzii M21.2 | ↑ | ns | <0.001 |
| Gemmiger formicilis | ↑ | ns | <0.001 |
| Gordonibacter pamelaeae | ↑ | ns | 0.003 |
| Holdemanella biformis | ↑ | ns | 0.005 |
| Flavonifractor plautii | ↑ | ns | 0.031 |
| Phocaeicola massiliensis | ↓ | ns | 0.012 |
| Holdemanella filiformis | ↓ | ns | 0.005 |
| Eggerthella lenta | ↑ | ns | 0.021 |
| Odoribacter splanchnicus | ↓ | ns | 0.028 |
| Alistipes shahii | ↓ | ns | 0.032 |
| Alistipes finegoldii | ↓ | ns | 0.035 |
| Bacteroidesuniformis | ↓ | ns | 0.035 |
| Clostridium citroniae | ↓ | ns | 0.045 |
| Bifidobacterium longum | ↓ | ns | 0.046 |
| Bifidobacterium animalis | ns | ↓ | 0.007 |
| Faecalibacterium prausnitzii CNCM4575 | ns | ↓ | 0.014 |
| Streptococcus anginosus | ns | ↓ | 0.017 |
| Bacteroides stercoris | ns | ↓ | 0.019 |
| Clostridium nexile | ns | ↓ | 0.038 |
| Parabacterium merdae | ns | ↓ | 0.042 |
| Eubacterium eligens | ns | ↑ | 0.048 |
3.2. Associations between bacterial taxa at baseline and fatigue, PCC and PIFS among SARS-CoV-2 positive participants at six months.
Higher baseline abundance of Sutterella wadsworthensis (S. wadsworthensis) was associated with fatigue caseness at inclusion (approximately half of the SARS -CoV-2 positive participants) (Figure 2A). The 46 (35%) SARS-CoV-2 positive participants classified as fatigue cases at six months follow-up had lower abundances of F. prausnitzii M21/2 (Figure 3A) and Ruminococcus bicirculans (R. bicirculans) and higher abundances of five bacterial taxa including Clostridium spiroforme (C. spiroforme) (Figure 3B), S. wadsworthensis (Figure 2B) and Streptococcus thermophilus (S. thermophilus) at baseline compared to non-cases (Table 3).
The 43% of SARS-CoV-2 positive participants with PCC (56/130) at six months had higher abundances of S. wadsworthensis (Figure 2C) and S. thermophilus at baseline compared to those without PCC (Table 3). The baseline abundance of S. wadsworthensis was also higher in the 15 (11.5%) SARS-CoV-2 positive participants with PIFS at six months, although this association failed to reach statistical significance (p=0.09), (Figure 2D). There were no bacterial taxa at baseline that were associated with PIFS status (15 (11.5%) SARS-CoV-2 positive participants) at six months.
4. Discussion
The main finding of the present study was an enrichment of the beneficial short chain fatty acid (SCFA) producers F. prausnitzii M21.2 and G. formicilis in the SARS-CoV-2 positive adolescents and young adults compared to SARS-CoV-2 negative controls at baseline. Six months post-infection this enrichment was no longer evident. Furthermore, in the SARS-CoV-2 positive group, lower baseline levels of F. prausnitzii M21/2 and R. bicirculans and higher abundance of C. spiroforme, S. wadsworthensis and S. thermophilus were associated with fatigue at six months. Additionally, enrichment of S. wadsworthensis and S. thermophilus at baseline was associated with PCC.
4.1. SARS-CoV-2 status
The increased abundance of F. prausnitzii and G. formicilis in the SARS-CoV-2 positive participants at baseline conflicts with results from adult COVID-19 cohorts where these SCFA producing and beneficial species are reported being depleted [14,31,32]. However, the enrichment of F. prausnitzii is consistent with gut microbiota studies of children with COVID-19 [33]. Our findings support the hypothesis of F. prausnitzii as a hallmark species enriched in pediatric COVID-19 disease, and that it may play a protective role against severe disease manifestations and outcomes [33,34]. Another player in maintaining a healthy gut microbiota, the anti-inflammatory bacterium H. biformis, was also found to be more abundant in our SARS-CoV-2 positive participants than in controls [35,36]. However, although F. prausnitzii, G. formicilis and H. biformis were enriched in our SARS-Cov-2 positive participants, several other commensal bacteria were depleted in comparison with controls. P. massiliensis, (former Bacteroides massiliensis) [37] a beneficial, mucine degrading member of the gut microbiota with abilities to utilize glycans [38], was decreased in our COVID-19 patients, alongside a depletion of B. longum. Bifidobacteria interact with human immune cells and modulate specific pathways and may decrease proinflammatory cytokines and restore intestinal barrier integrity [39]. Depletion of Bifidobacteria are reported in adult patients with COVID-19 [14] as well as in pediatric patients [34], and together with F. prausnitzii [40,41,42] there is evidence for an inverse association between disease severity and abundance [43]. F. plautii, enriched in our SARS-CoV-2 positive participants, has in previous studies been found to be increased in pediatric inflammatory bowel disease (IBD) and in irritable bowel syndrome (IBS) patients [44,45]. Similarly, we found an increased abundance of E. lenta, a bacterium associated with autoimmunity and previously found enriched in the gut microbiota of children with multisystem inflammatory syndrome (MIS-C) [46]. Taken together, we find evidence of both anti- and pro-inflammatory differences in the microbiota of SARS-COV-2 positive versus SARS-COV-2 negative controls at baseline following mild COVID-19 disease in adolescents and young adults.
4.2. SARS-CoV-2 infection and severity
One hypothesis regarding the less severe course of COVID-19 disease in children compared with adults is that their gastrointestinal microbiota differs from that of adults [47], and that pre-infection differences in the gut microbiota, such as enrichment of SCFA producers and associated immune responses, may determine COVID-19 disease presentation, severity and outcome [17,48]. The SARS-CoV-2 positive adolescents and young adults in our study were not hospitalized and did not have severe manifestations of COVID-19. Maybe the enrichment of the SCFA producers F. prausnitzii and G. formicilis contributed to the relatively mild disease course. Lower levels of F. prausnitzii in populations at risk for severe COVID-19 disease, as the elder, obese and diabetic populations [43], are further supporting this theory. In line with these results, we found an association of fatigue caseness at six months with lower baseline abundances of F. prausnitzii M21/2 and R. bicirculans, [49] a beneficial SCFA producer and one of the most abundant members of the healthy gut microbiota. These findings support the hypothesis that the gut microbiome after COVID-19 disease may have an impaired capacity to synthesize SCFAs, and that low SCFAs levels may contribute to persistence of symptoms such as fatigue and PCC [50]. Similarly, we found an association between lower abundance of Bifidobacteria species with SARS- CoV-2 status as well as with fatigue caseness, a finding that has been reported both in adults [43] and in children [34] with COVID-19.
An interesting finding in the present study is the enrichment of the Gram-negative, microaerophilic bacterium S. wadsworthensis at baseline in the SARS-COV-2 positive participants with fatigue at six months, as well as in those fulfilling the criteria for PCC. This bacterial species is a potential pathogen associated with pediatric IBD [51] as well as pediatric obesity [52]. S. wadsworthensis has not been implicated in PCC or PIFS before, but the ability of Sutterella species to adhere to intestinal epithelial cells could indicate an immunomodulatory role [53]. Similarly, S. thermophilus was enriched at baseline in Sars-CoV-2 patients with fatigue and PCC at six months. In line with this, Li et al. found S. thermophilus to be enriched in COVID-19 patients compared to healthy and that this species correlated with severity of the disease [54].
Another novel association discovered in the present study was the higher abundance of C. spiroforme in participants with fatigue following COVID-19 infection compared to those without fatigue post-infection. C. spiroforme has recently been linked to the development and progression of colorectal cancer in [55]. Another study from 2024 found that the presence of this species in the feces may aid in the diagnosis of IBD [56] indicating that C. spiroforme may act as a pathogen.
The bacterial taxa with a deviating abundance in those with PCC and PIFS were not the same in the SARS-CoV-2 positive as in the SARS-CoV-2 negative participants although the SARS-CoV-2 negative participants had a similarly high symptom load as the participants who tested positive for SARS-CoV-2. It is not possible to determine whether the differences in bacterial taxa levels observed between our SARS-CoV-2 positive participants and controls preceded or followed the infection with COVID-19. While we cannot attribute a causal role of bacterial species in the pathogenesis of SARS-CoV-2 infection in our study, our data support a potential role of the gut microbiome in modulating the disease course, as hypothesized earlier by Rochi et al. [57].
4.3. Strengths and limitations
One of the strengths of the study is the well-defined group of non-hospitalized symptomatic young individuals with COVID-19 and a comparable SARS-CoV-2 negative control group. All participants were subjected to rigorous and systematic examinations, they were from the same catchment area, with a similar socio-economic status and were not treated with medications during their COVID-19 illness. SARS-CoV-2 antibody testing was conducted both at inclusion and at six months follow-up, and participants displaying antibodies indicative of prior COVID-19 infection were excluded as SARS-CoV-2 negative controls.
Our study has several limitations. The sample size of the study, particularly regarding SARS-CoV-2 negative participants, was small, with only one in three of the LoTECA enrolled participants being willing to deliver fecal samples, rendering them eligible for this sub-study. Due to the small sample size and no correction for multiple testing, our findings with deviating species abundance in fatigue and PCC could be coincidental and must be interpreted with caution. In addition, there is a potential risk of self-selection bias, as those who joined the study and provided a fecal sample may not be representative for the targeted population of children/adolescents. As species and strains to be detected were selected a priori, we may have missed novel bacteria with previously unknown beneficial / harmful effects. The sample material used in the study is microbial DNA, hence the results represent active, dormant and dead microbes. Also, we have no influence or knowledge of the participants’ diet and lifestyle, which is known to influence the microbiota and could be an important confounder in our analyses.
5. Conclusions
Adolescents and young adults with mild acute COVID-19 infection had increased fecal abundance of the beneficial SCFA producers F. prausnitzii and G. formicilis species compared to SARS-CoV-2 negative participants in the early convalescent stage. Additionally, we found that both known (F. Prausnitzii, S. thermophilus) and new (C. spiroforme and S. wadsworthensis) bacteria were associated with persistent symptoms in the COVID-19 infected group, warranting further exploration of the role of these bacteria in COVID-19 and PCC pathophysiology.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: List of bacterial taxa used in PMP™ panels, Figure S1: Principal component analysis of SARS-CoV-2 positive and negative participants at baseline, Figure S2: Boxplot of bacterial richness between SARS-CoV-2 positive and negative participants at baseline.
Author Contributions
Conceptualization, Christine Olbjørn, Aina Elisabeth Fossum Moen, Joel Selvakumar and Vegard Bruun Bratholm Wyller; Methodology, Christine Olbjørn, Milada Hagen, Aina Elisabeth Fossum Moen and Vegard Bruun Bratholm Wyller; Software, Milada Hagen and Joel Selvakumar; Validation, Milada Hagen; Formal analysis, Christine Olbjørn, Milada Hagen, Lise Beier Havdal and Joel Selvakumar; Investigation, Lise Beier Havdal, Silke Lauren Sommen, Lise Lund Berven, Tonje Stiansen-Sonerud, Joel Selvakumar and Vegard Bruun Bratholm Wyller; Resources, Christine Olbjørn, Milada Hagen, Aina Elisabeth Fossum Moen, Lise Beier Havdal, Silke Lauren Sommen, Lise Lund Berven, Tonje Stiansen-Sonerud, Joel Selvakumar and Vegard Bruun Bratholm Wyller; Data curation, Christine Olbjørn, Milada Hagen, Aina Elisabeth Fossum Moen, Lise Beier Havdal, Tonje Stiansen-Sonerud, Joel Selvakumar and Vegard Bruun Bratholm Wyller; Writing – original draft, Christine Olbjørn; Writing – review & editing, Christine Olbjørn, Milada Hagen, Aina Elisabeth Fossum Moen, Lise Beier Havdal, Silke Lauren Sommen, Lise Lund Berven, Espen Thiis-Evensen, Tonje Stiansen-Sonerud, Joel Selvakumar and Vegard Bruun Bratholm Wyller; Visualization, Christine Olbjørn, Milada Hagen and Aina Elisabeth Fossum Moen; Supervision, Christine Olbjørn and Vegard Bruun Bratholm Wyller; Project administration, Christine Olbjørn and Vegard Bruun Bratholm Wyller; Funding acquisition, Christine Olbjørn, Bio-Me. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by an internal strategic grant from Akershus University Hospital, grant number 262983 and from Regionalt forskningsfond, RFF Oslo, project number 324123. The APC was funded by Akershus University Hospital.
Data Availability Statement
The raw datasets generated and analyzed during the current study are not publicly available in order to protect participant confidentiality. The data files are stored by Akershus University Hospital, Lørenskog, Norway, on a server dedicated to research. The security follows the rules given by The Norwegian Data Protection Authority, P.O. Box 8177 Dep. NO-0034 Oslo, Norway. The data are available on request to the authors.
Acknowledgments
We thank Ann Cathrin Pettersen, Helene Larssen, LoTECA participants, Bio-Me and other laboratory personnel that was involved in the analysis.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Behnood, S.A.; Shafran, R.; Bennett, S.D.; Zhang, A.X.; O'Mahoney, L.L.; Stephenson, T.J.; Ladhani, S.N.; De Stavola, B.L.; Viner, R.M.; Swann, O.V. Persistent symptoms following SARS-CoV-2 infection amongst children and young people: A meta-analysis of controlled and uncontrolled studies. J. Infect. 2022, 84, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, R.; Chiappini, E.; Licari, A.; Galli, L.; Marseglia, G.L. Prevalence and clinical presentation of long COVID in children: A systematic review. Eur. J. Pediatr. 2022, 181, 3995–4009. [Google Scholar] [CrossRef] [PubMed]
- Ceban, F.; Ling, S.; Lui, L.M.W.; Lee, Y.; Gill, H.; Teopiz, K.M.; Rodrigues, N.B.; Subramaniapillai, M.; Di Vincenzo, J.D.; Cao, B.; et al. Fatigue and cognitive impairment in Post-COVID-19 Syndrome: A systematic review and meta-analysis. Brain Behav. Immun. 2022, 101, 93–135. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.K.; Malone, L.A.; Kokorelis, C.; Petracek, L.S.; Eastin, E.F.; Lobner, K.L.; Neuendorff, L.; Rowe, P.C. Long-Term COVID 19 Sequelae in Adolescents: The Overlap with Orthostatic Intolerance, M.E./.C.F.S. Curr. Pediatr. Rep. 2022, 10, 31–44. [Google Scholar] [CrossRef]
- Soriano, J.B.; Murthy, S.; Marshall, J.C.; Relan, P.; Diaz, J.V. A clinical case definition of post-COVID-19 condition by a Delphi consensus. Lancet Infect. Dis. 2022, 22, e102–e107. [Google Scholar] [CrossRef]
- Wong, T.L.; Weitzer, D.J. Long COVID and Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS)-A Systemic Review and Comparison of Clinical Presentation and Symptomatology. Medicina 2021, 57, 418. [Google Scholar] [CrossRef]
- Hickie, I.; Davenport, T.; Wakefield, D.; Vollmer-Conna, U.; Cameron, B.; Vernon, S.D.; Reeves, W.C.; Lloyd, A. Post-infective and chronic fatigue syndromes precipitated by viral and non-viral pathogens: Prospective cohort study. BMJ (Clin. Res. Ed) 2006, 333, 575. [Google Scholar] [CrossRef]
- Zhang, F.; Lau, R.I.; Liu, Q.; Su, Q.; Chan, F.K.L.; Ng, S.C. Gut microbiota in COVID-19: Key microbial changes, potential mechanisms and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 2022, 20, 323–337. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Petersen, C.; Round, J.L. Defining dysbiosis and its influence on host immunity and disease. Cell. Microbiol. 2014, 16, 1024–1033. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.Y.; Yeoh, Y.K.; Li, A.Y.L.; Zhan, H.; Wan, Y.; Chung, A.C.K.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients With COVID-19 During Time of Hospitalization. Gastroenterology 2020, 159, 944–955.e8. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Wu, J.; Zeng, Y.; Li, J.; Yan, J.; Meng, W.; Han, H.; Feng, F.; He, J.; Zhao, S.; et al. SARS-CoV-2 triggered oxidative stress and abnormal energy metabolism in gut microbiota. MedComm 2022, 3, e112. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Liu, Q.; Zhang, F.; Lui, G.C.; Tso, E.Y.; Yeoh, Y.K.; Chen, Z.; Boon, S.S.; Chan, F.K.; Chan, P.K.; et al. Depicting SARS-CoV-2 faecal viral activity in association with gut microbiota composition in patients with COVID-19. Gut 2021, 70, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S.; et al. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut 2021, 70, 698–706. [Google Scholar] [CrossRef]
- Brodin, P. SARS-CoV-2 infections in children: Understanding diverse outcomes. Immunity 2022, 55, 201–209. [Google Scholar] [CrossRef]
- Ludvigsson, J.F. Systematic review of COVID-19 in children shows milder cases and a better prognosis than adults. Acta Paediatr. 2020, 109, 1088–1095. [Google Scholar] [CrossRef]
- Brodin, P. Immune determinants of COVID-19 disease presentation and severity. Nat. Med. 2021, 27, 28–33. [Google Scholar] [CrossRef]
- Castagnoli, R.; Votto, M.; Licari, A.; Brambilla, I.; Bruno, R.; Perlini, S.; Rovida, F.; Baldanti, F.; Marseglia, G.L. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection in Children and Adolescents: A Systematic Review. JAMA Pediatr. 2020, 174, 882–889. [Google Scholar] [CrossRef]
- Ailioaie, L.M.; Ailioaie, C.; Litscher, G. Infection, Dysbiosis and Inflammation Interplay in the COVID Era in Children. Int. J. Mol. Sci. 2023, 24, 10874. [Google Scholar] [CrossRef]
- Villapol, S. Gastrointestinal symptoms associated with COVID-19: Impact on the gut microbiome. Transl. Res. J. Lab. Clin. Med. 2020, 226, 57–69. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, W.; Guo, M.; Xiao, C.; Fu, Z.; Yu, S.; Jiang, L.; Wang, S.; Ling, Y.; Liu, F.; et al. Integrated analysis of gut microbiome and host immune responses in COVID-19. Front. Med. 2022, 16, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gu, S.; Chen, Y.; Lu, H.; Shi, D.; Guo, J.; Wu, W.-R.; Yang, Y.; Li, Y.; Xu, K.-J.; et al. Six-month follow-up of gut microbiota richness in patients with COVID-19. Gut 2022, 71, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Mak, J.W.Y.; Su, Q.; Yeoh, Y.K.; Lui, G.C.; Ng, S.S.S.; Zhang, F.; Li, A.Y.L.; Lu, W.; Hui, D.S.-C.; et al. Gut microbiota dynamics in a prospective cohort of patients with post-acute COVID-19 syndrome. Gut 2022, 71, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, J.; Havdal, L.B.; Drevvatne, M.; Brodwall, E.M.; Lund Berven, L.; Stiansen-Sonerud, T.; Einvik, G.; Leegaard, T.M.; Tjade, T.; Michelsen, A.E.; et al. Prevalence and Characteristics Associated With Post-COVID-19 Condition Among Nonhospitalized Adolescents and Young Adults. JAMA Netw. Open 2023, 6, e235763. [Google Scholar] [CrossRef] [PubMed]
- Lund Berven, L.; Selvakumar, J.; Havdal, L.; Stiansen-Sonerud, T.; Einvik, G.; Leegaard, T.M.; Tjade, T.; Michelsen, A.E.; Mollnes, T.E.; Wyller, V.B.B. Inflammatory Markers, Pulmonary Function, and Clinical Symptoms in Acute COVID-19 Among Non-Hospitalized Adolescents and Young Adults. Front. Immunol. 2022, 13, 837288. [Google Scholar] [CrossRef]
- Morriss, R.K.; Wearden, A.J.; Mullis, R. Exploring the validity of the Chalder Fatigue scale in chronic fatigue syndrome. J. Psychosom. Res. 1998, 45, 411–417. [Google Scholar] [CrossRef]
- White, P.D.; Goldsmith, K.A.; Johnson, A.L.; Potts, L.; Walwyn, R.; DeCesare, J.C.; Baber, H.L.; Burgess, M.; Clark, L.V.; Cox, D.L.; et al. Comparison of adaptive pacing therapy, cognitive behaviour therapy, graded exercise therapy, and specialist medical care for chronic fatigue syndrome (PACE): A randomised trial. Lancet 2011, 377, 823–836. [Google Scholar] [CrossRef]
- Husakova, M.; Kralik, P.; Babak, V.; Slana, I. Efficiency of DNA Isolation Methods Based on Silica Columns and Magnetic Separation Tested for the Detection of Mycobacterium avium Subsp. Paratuberculosis in Milk and Faeces. Materials 2020, 13, 5112. [Google Scholar] [CrossRef]
- Farup, P.G.; Maseng, M.G. Are Faecal Microbiota Analyses on Species-Level Suitable Clinical Biomarkers? A Pilot Study in Subjects with Morbid Obesity. Microorganisms 2021, 9, 664. [Google Scholar] [CrossRef]
- Derosa, L.; Iebba, V.; Silva, C.A.C.; Piccinno, G.; Wu, G.; Lordello, L.; Routy, B.; Zhao, N.; Thelemaque, C.; Birebent, R.; et al. Custom scoring based on ecological topology of gut microbiota associated with cancer immunotherapy outcome. Cell 2024, 187, 3373–3389.e16. [Google Scholar] [CrossRef]
- Gu, S.; Chen, Y.; Wu, Z.; Chen, Y.; Gao, H.; Lv, L.; Guo, F.; Zhang, X.; Luo, R.; Huang, C.; et al. Alterations of the Gut Microbiota in Patients With Coronavirus Disease 2019 or H1N1 Influenza. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2020, 71, 2669–2678. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, L.; Wang, Y.; Dai, T.; Qin, Z.; Zhou, F.; Zhang, L. Alterations in microbiota of patients with COVID-19: Potential mechanisms and therapeutic interventions. Signal Transduct. Target. Ther. 2022, 7, 143. [Google Scholar] [CrossRef] [PubMed]
- Romani, L.; Del Chierico, F.; Macari, G.; Pane, S.; Ristori, M.V.; Guarrasi, V.; Gardini, S.; Pascucci, G.R.; Cotugno, N.; Perno, C.F.; et al. The Relationship Between Pediatric Gut Microbiota and SARS-CoV-2 Infection. Front. Cell. Infect. Microbiol. 2022, 12, 908492. [Google Scholar] [CrossRef] [PubMed]
- Piazzesi, A.; Pane, S.; Del Chierico, F.; Romani, L.; Campana, A.; Palma, P.; Putignani, L. The pediatric gut bacteriome and virome in response to SARS-CoV-2 infection. Front. Cell. Infect. Microbiol. 2024, 14, 1335450. [Google Scholar] [CrossRef]
- Zhang, C.; Liang, D.; Li, X.; Liu, J.; Fan, M.; Jing, M.; Wang, Y.; Zhang, Y.; Fang, Y.; Li, D. Characteristics of Gut Microbial Profiles of Offshore Workers and Its Associations With Diet. Front. Nutr. 2022, 9, 904927. [Google Scholar] [CrossRef]
- Romaní-Pérez, M.; López-Almela, I.; Bullich-Vilarrubias, C.; Rueda-Ruzafa, L.; Gómez Del Pulgar, E.M.; Benítez-Páez, A.; Liebisch, G.; Lamas, J.A.; Sanz, Y. Holdemanella biformis improves glucose tolerance and regulates GLP-1 signaling in obese mice. FASEB J. 2021, 35, e21734. [Google Scholar] [CrossRef]
- García-López, M.; Meier-Kolthoff, J.P.; Tindall, B.J.; Gronow, S.; Woyke, T.; Kyrpides, N.C.; Hahnke, R.L.; Göker, M. Analysis of 1,000 Type-Strain Genomes Improves Taxonomic Classification of Bacteroidetes. Front. Microbiol. 2019, 10, 2083. [Google Scholar] [CrossRef]
- Pudlo, N.A.; Urs, K.; Crawford, R.; Pirani, A.; Atherly, T.; Jimenez, R.; Terrapon, N.; Henrissat, B.; Peterson, D.; Ziemer, C.; et al. Phenotypic and Genomic Diversification in Complex Carbohydrate-Degrading Human Gut Bacteria. mSystems 2022, 7, e0094721. [Google Scholar] [CrossRef]
- Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Sánchez, B.; Margolles, A. Bifidobacteria and Their Molecular Communication with the Immune System. Front. Microbiol. 2017, 8, 2345. [Google Scholar] [CrossRef]
- Lin, R.; Xiao, M.; Cao, S.; Sun, Y.; Zhao, L.; Mao, X.; Chen, P.; Tong, X.; Ou, Z.; Zhu, H.; et al. Distinct gut microbiota and health outcomes in asymptomatic infection, viral nucleic acid test re-positive, and convalescent COVID-19 cases. mLife 2022, 1, 183–197. [Google Scholar] [CrossRef]
- Farsi, Y.; Tahvildari, A.; Arbabi, M.; Vazife, F.; Sechi, L.A.; Shahidi Bonjar, A.H.; Jamshidi, P.; Nasiri, M.J.; Mirsaeidi, M. Diagnostic, Prognostic, and Therapeutic Roles of Gut Microbiota in COVID-19: A Comprehensive Systematic Review. Front. Cell. Infect. Microbiol. 2022, 12, 804644. [Google Scholar] [CrossRef] [PubMed]
- Schult, D.; Reitmeier, S.; Koyumdzhieva, P.; Lahmer, T.; Middelhoff, M.; Erber, J.; Schneider, J.; Kager, J.; Frolova, M.; Horstmann, J.; et al. Gut bacterial dysbiosis and instability is associated with the onset of complications and mortality in COVID-19. Gut Microbes 2022, 14, 2031840. [Google Scholar] [CrossRef] [PubMed]
- Hazan, S.; Stollman, N.; Bozkurt, H.S.; Dave, S.; Papoutsis, A.J.; Daniels, J.; Barrows, B.D.; Quigley, E.M.; Borody, T.J. Lost microbes of COVID-19: Bifidobacterium, Faecalibacterium depletion and decreased microbiome diversity associated with SARS-CoV-2 infection severity. BMJ Open Gastroenterol. 2022, 9, e000871. [Google Scholar] [CrossRef]
- Armstrong, H.; Alipour, M.; Valcheva, R.; Bording-Jorgensen, M.; Jovel, J.; Zaidi, D.; Shah, P.; Lou, Y.; Ebeling, C.; Mason, A.L.; et al. Host immunoglobulin G selectively identifies pathobionts in pediatric inflammatory bowel diseases. Microbiome 2019, 7, 1. [Google Scholar] [CrossRef]
- Hollister, E.B.; Oezguen, N.; Chumpitazi, B.P.; Luna, R.A.; Weidler, E.M.; Rubio-Gonzales, M.; Dahdouli, M.; Cope, J.L.; Mistretta, T.-A.; Raza, S.; et al. Leveraging Human Microbiome Features to Diagnose and Stratify Children with Irritable Bowel Syndrome. J. Mol. Diagn. JMD 2019, 21, 449–461. [Google Scholar] [CrossRef]
- Suskun, C.; Kilic, O.; Yilmaz Ciftdogan, D.; Guven, S.; Karbuz, A.; Ozkaya Parlakay, A.; Kara, Y.; Kacmaz, E.; Sahin, A.; Boga, A.; et al. Intestinal microbiota composition of children with infection with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and multisystem inflammatory syndrome (MIS-C). Eur. J. Pediatr. 2022, 181, 3175–3191. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. Why is COVID-19 less severe in children? A review of the proposed mechanisms underlying the age-related difference in severity of SARS-CoV-2 infections. Arch. Dis. Child. 2020, 106, 429–439. [Google Scholar] [CrossRef]
- Li, J.; Richards, E.M.; Handberg, E.M.; Pepine, C.J.; Raizada, M.K. Butyrate Regulates COVID-19-Relevant Genes in Gut Epithelial Organoids From Normotensive Rats. Hypertension 2021, 77, e13–e16. [Google Scholar] [CrossRef]
- Wegmann, U.; Louis, P.; Goesmann, A.; Henrissat, B.; Duncan, S.H.; Flint, H.J. Complete genome of a new Firmicutes species belonging to the dominant human colonic microbiota (‘Ruminococcus bicirculans’) reveals two chromosomes and a selective capacity to utilize plant glucans. Environ. Microbiol. 2014, 16, 2879–2890. [Google Scholar] [CrossRef]
- Zhang, F.; Wan, Y.; Zuo, T.; Yeoh, Y.K.; Liu, Q.; Zhang, L.; Zhan, H.; Lu, W.; Xu, W.; Lui, G.C.; et al. Prolonged Impairment of Short-Chain Fatty Acid and L-Isoleucine Biosynthesis in Gut Microbiome in Patients With COVID-19. Gastroenterology 2022, 162, 548–561.e4. [Google Scholar] [CrossRef]
- Kaakoush, N.O. Sutterella Species, IgA-degrading Bacteria in Ulcerative Colitis. Trends Microbiol. 2020, 28, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Squillario, M.; Bonaretti, C.; La Valle, A.; Di Marco, E.; Piccolo, G.; Minuto, N.; Patti, G.; Napoli, F.; Bassi, M.; Maghnie, M.; et al. Gut-microbiota in children and adolescents with obesity: Inferred functional analysis and machine-learning algorithms to classify microorganisms. Sci. Rep. 2023, 13, 11294. [Google Scholar] [CrossRef] [PubMed]
- Hiippala, K.; Kainulainen, V.; Kalliomaki, M.; Arkkila, P.; Satokari, R. Mucosal Prevalence and Interactions with the Epithelium Indicate Commensalism of Sutterella spp. Front. Microbiol. 2016, 7, 1706. [Google Scholar] [CrossRef]
- Li, S.; Yang, S.; Zhou, Y.; Disoma, C.; Dong, Z.; Du, A.; Zhang, Y.; Chen, Y.; Huang, W.; Chen, J.; et al. Microbiome Profiling Using Shotgun Metagenomic Sequencing Identified Unique Microorganisms in COVID-19 Patients With Altered Gut Microbiota. Front. Microbiol. 2021, 12, 712081. [Google Scholar] [CrossRef]
- Kim, N.; Gim, J.A.; Lee, B.J.; Choi, B.I.; Yoon, H.S.; Kim, S.H.; Joo, M.K.; Park, J.-J.; Kim, C. Crosstalk between mucosal microbiota, host gene expression, and sociomedical factors in the progression of colorectal cancer. Sci. Rep. 2022, 12, 13447. [Google Scholar] [CrossRef]
- Wu, X.; Li, Y.; Shang, Y.; Wang, W.; Wu, L.; Han, L.; Wang, Q.; Wang, Z.; Xu, H.; Liu, W. Application of two-dimensional polymerase chain reaction to detect four types of microorganisms in feces for assisted diagnosis of I.B.D. Clin. Chim. Acta Int. J. Clin. Chem. 2024, 555, 117802. [Google Scholar] [CrossRef]
- Rocchi, G.; Giovanetti, M.; Benedetti, F.; Borsetti, A.; Ceccarelli, G.; Zella, D.; Altomare, A.; Ciccozzi, M.; Guarino, M.P.L. Gut Microbiota and COVID-19: Potential Implications for Disease Severity. Pathogens 2022, 11, 1050. [Google Scholar] [CrossRef]
Figure 1.
Boxplots showing abundance (genomes/ng DNA) of A) Faecalibacterium prausnitzii M21.2 and B) Gemmiger formicilis in SARS-CoV-2 positive participants compared to SARS-CoV-2 negative at baseline.
Figure 1.
Boxplots showing abundance (genomes/ng DNA) of A) Faecalibacterium prausnitzii M21.2 and B) Gemmiger formicilis in SARS-CoV-2 positive participants compared to SARS-CoV-2 negative at baseline.
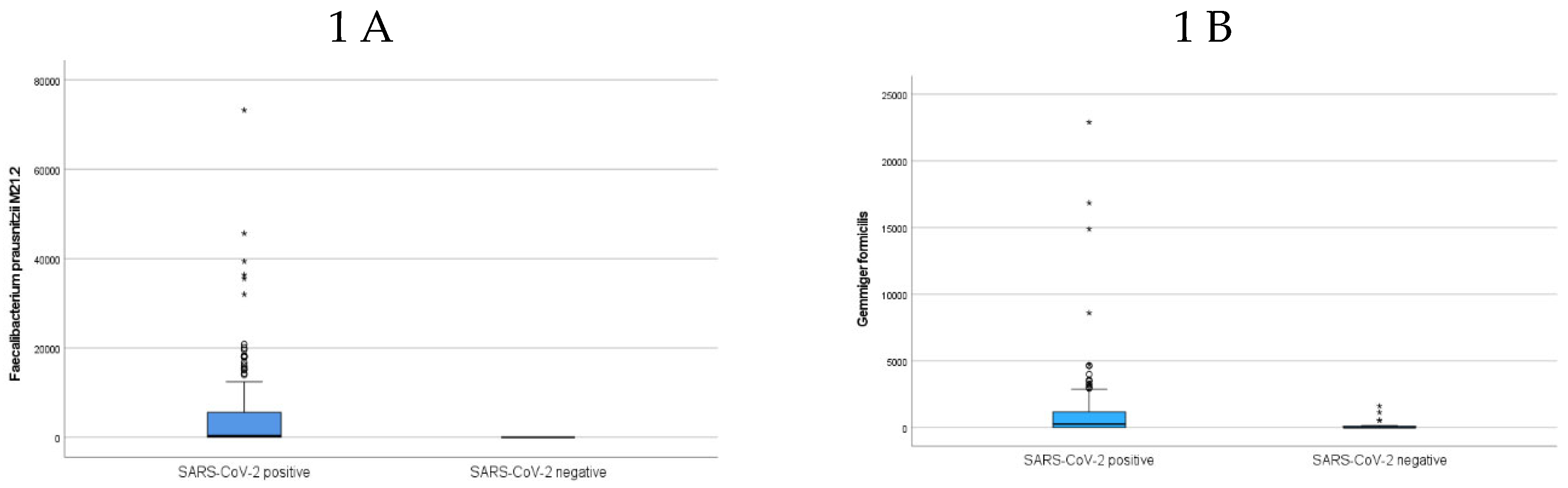
Figure 2.
Boxplots showing baseline abundance (genomes/ng DNA) of Sutterella wadsworthensis in SARS-CoV-2 positive participants with A) fatigue case versus non-case at baseline, B) fatigue case versus non-case at six months, C) PCC case versus non-case at six months, D) PIFS case versus non-case at six months. Abbreviations: PCC (post-COVID-19 condition), PIFS (post-infective fatigue syndrome).
Figure 2.
Boxplots showing baseline abundance (genomes/ng DNA) of Sutterella wadsworthensis in SARS-CoV-2 positive participants with A) fatigue case versus non-case at baseline, B) fatigue case versus non-case at six months, C) PCC case versus non-case at six months, D) PIFS case versus non-case at six months. Abbreviations: PCC (post-COVID-19 condition), PIFS (post-infective fatigue syndrome).
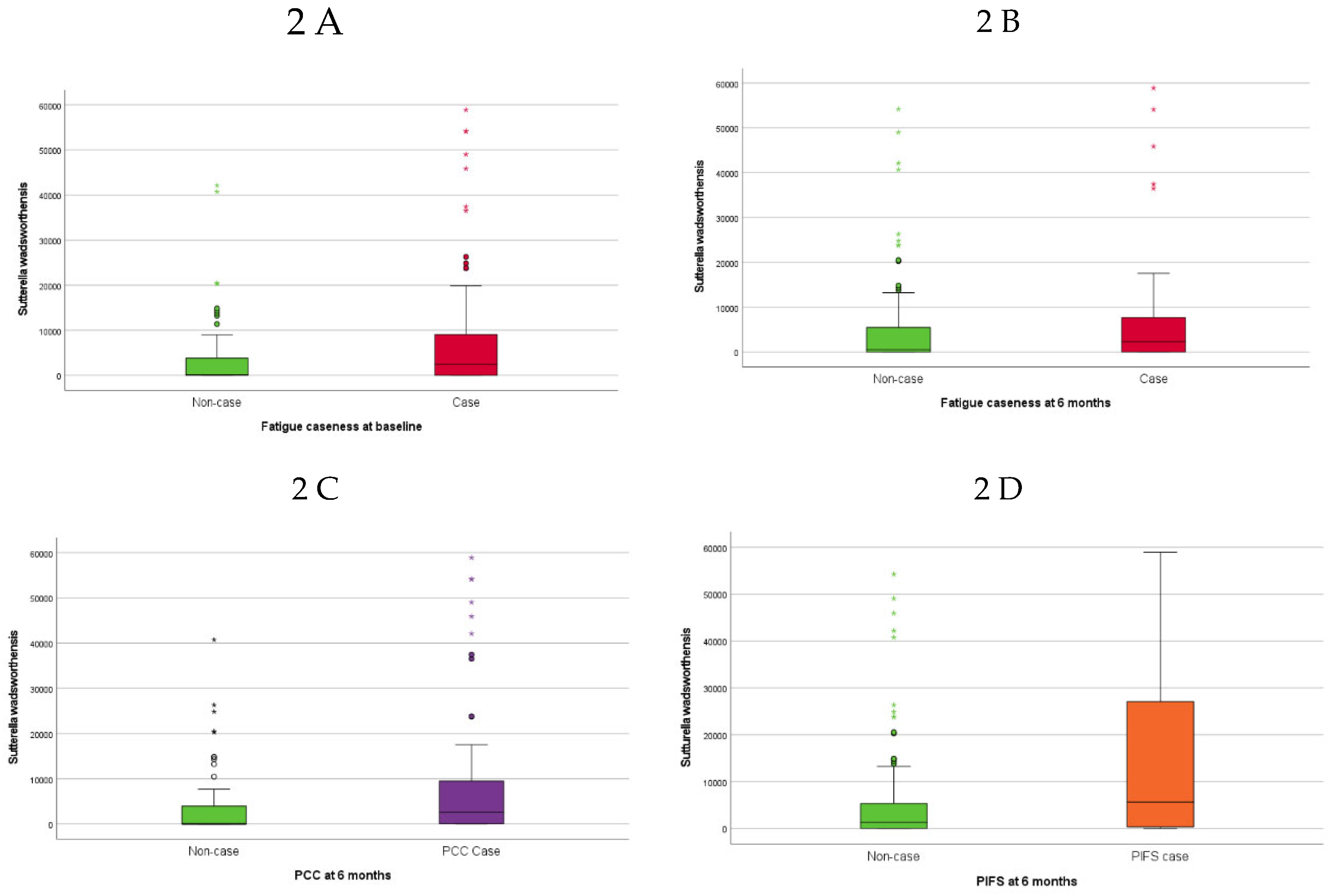
Figure 3.
Boxplots showing baseline abundance (genomes/ng DNA) of A) Faecalibacterium prausnitzii M21.2 and B) Clostridium spiroforme in SARS-CoV-2 positive participants with fatigue case compared to non-case at six months. Abbreviations: PCC (post-COVID-19 condition), PIFS (post-infective fatigue syndrome).
Figure 3.
Boxplots showing baseline abundance (genomes/ng DNA) of A) Faecalibacterium prausnitzii M21.2 and B) Clostridium spiroforme in SARS-CoV-2 positive participants with fatigue case compared to non-case at six months. Abbreviations: PCC (post-COVID-19 condition), PIFS (post-infective fatigue syndrome).
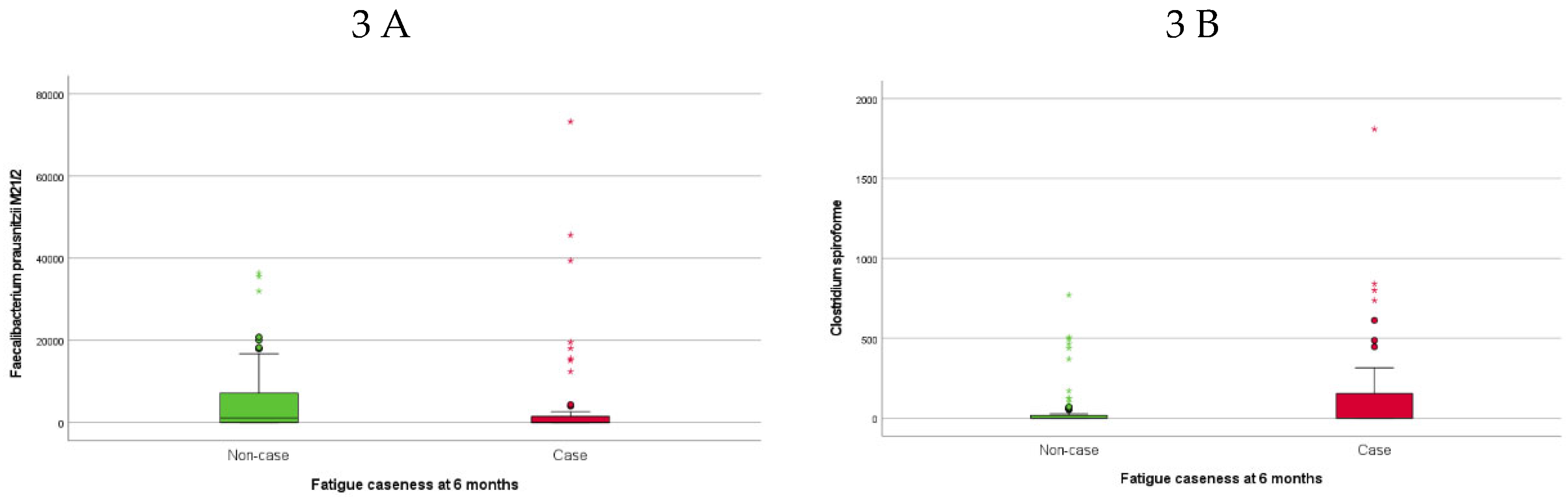
Table 1.
Characteristics of SARS-CoV-2 positive and SARS-CoV-2 negative participants including fatigue caseness (positive CFQ bimodal score) at baseline and six months follow-up, and post-COVID-19 condition (PCC) and post-infective fatigue syndrome (PIFS) at six months follow-up. Abbreviations: IQR (interquartile range), BMI (body mass index), CFQ (Chalder fatigue questionnaire).
Table 1.
Characteristics of SARS-CoV-2 positive and SARS-CoV-2 negative participants including fatigue caseness (positive CFQ bimodal score) at baseline and six months follow-up, and post-COVID-19 condition (PCC) and post-infective fatigue syndrome (PIFS) at six months follow-up. Abbreviations: IQR (interquartile range), BMI (body mass index), CFQ (Chalder fatigue questionnaire).
| Baseline | Six months follow-up | |||
|---|---|---|---|---|
| Characteristics |
SARS-CoV-2 positive (n=136) | SARS-CoV-2 negative (n=32) | SARS-CoV-2 Positive (n=102) |
SARS-CoV-2 negative (n=17) |
| Female –n (%) | 78 (57) | 19 (59) | 46 (45) | 12 (71)) |
| Age median [IQR] | 17.0 [14-21] | 16.5 [15-20] | 17.0 [14-22.5] | 18.0 [16-21] |
| BMI – median [IQR] | 22.4 [19.9-25.3] | 21.8 [20.2-24.4] | 22.3 [19.8-25.3] | 21.6 [20-23.8] |
| CFQ fatigue caseness -n (%) | 71 (51) | 14 (45) | 36 (35) | 4 (24) |
| PCC caseness-n (%) | 42 (42) | 7 (41) | ||
| PIFS caseness-n (%) | 12 (12) | 1 (6) | ||
Fecal bacterial abundances and SARS-CoV-2 status at baseline
Table 3.
Bacterial taxa with a significantly increased (↑) or decreased (↓) abundance at baseline in SARS-CoV-2 positive with fatigue, PCC and PIFS case at six months follow-up compared to SARS-CoV-2 positive without fatigue, PCC and PIFS case.
Table 3.
Bacterial taxa with a significantly increased (↑) or decreased (↓) abundance at baseline in SARS-CoV-2 positive with fatigue, PCC and PIFS case at six months follow-up compared to SARS-CoV-2 positive without fatigue, PCC and PIFS case.
|
Bacterial taxa with deviating abundance at baseline in case compared to no case |
SARS-CoV-2 positive participants (n= 130) | |||
|---|---|---|---|---|
| Baseline ↑ or ↓ (p value) in case n (%) |
Six-months follow-up ↑ or ↓ (p value) in case n (%) case vs no case n (%) |
|||
| Fatigue 68 (52%) |
Fatigue 46 (53%) |
PCC 56 (43%) |
PIFS 15 (12%) |
|
| Bacteroides thetaiomicron | ↑ (0.026) | ns | ns | ns |
| Sutterella wadsworthensis | ↑ (0.047) | ↑ (<0.047) | ↑ (<0.001) | ns |
| Alistipes putredenis | ↑ (0.049) | ns | ns | ns |
| Bifidobact angulatum | ↓ (0.014) | ns | ns | ns |
| Phocoeicola massiliensis | ↓(0.014) | ns | ns | ns |
| Bacteroides stercoris | ↓ (0.025) | ns | ns | ns |
| Clostridium spiroforme | ns | ↑ (0.006) | ns | ns |
| Faecalibacterium prausnitzii CNCM75 | ns | ↑ (0.024) | ns | ns |
| Streptococcus thermophilus | ns | ↑ (0.039) | ↑ (0.042) | ns |
| Roseburia intestinalis | ns | ↑ (0.046) | ns | ns |
| Faecalibacterium prausnitzii M21/2 | ns | ↓ (0.013) | ns | ns |
| Ruminococcus bicirculans | ns | ↓(0.045) | ns | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



