
Submitted:
23 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
The genetic transformation of Coffea arabica L. is an alternative strategy for obtaining plants with agronomic traits of interest that is less time-consuming than conventional breeding methods. Given the importance of coffee cultivation in Colombia, this study evaluated the main factors interfering with the genetic transformation of C. arabica using Agrobacterium tumefaciens. An effi-cient and reproducible method was accordingly established that involved the propagation of “early” embryogenic tissue in a liquid proliferation medium, supplemented with 3 mg l−1 BAP for eight months, which was subsequently sonicated for 300 sec in a suspension of LBA4405 OD600 of 0.5 harboring pC1301 and incubated in this same suspension for 1 hr. This plasmid contained the uidA gene under control of the 35S promoter. The Agrobacterium suspen-sion was subsequently removed from the embryogenic tissue using a micropipette, after which the tissue was deposited on filter paper to remove the remaining Agrobacterium suspension. The embryogenic tissue was co-cultured for four days in a solid differentiation medium supple-mented with 100 µM acetosyringone on filter paper. Subsequently, the tissue was post-cultured for four days in liquid differentiation medium under orbital shaking at 100 rpm with 300 mg l−1 Claforan (Hoechst), followed by selection with 50 mg l-1 hygromycin at 26 °C in the dark, with subcultures at 20-day intervals until somatic embryos were formed for subsequent culturing in germination medium. Molecular analysis confirmed the presence of the uidA gene in coffee seedlings transformed with strains LBA4405 and EHA105 and vectors pC1301 and pC2301. This method successfully enables the stable integration of genes of interest in the coffee plant ge-nome
Keywords:
genetic transformation
; Agrobacterium tumefaciens
; somatic embryogenesis
; Coffea arabica
; coffee
1. Introduction
Coffee crops cover approximately 12 million hectares (ha) in 82 countries in the tropics and subtropics, mainly in Brazil and Colombia (Latin America), Vietnam and Indonesia (Asia), and Ethiopia and Ivory Coast (Africa). Approximately 10 million tons of green coffee beans are produced globally, making it one of the most traded commodities worldwide [1]. The International Coffee Organization (ICO) estimated that in 2022 between 12.5 and 25 million agricultural households worldwide depended on coffee production for a living [2]. Coffea belongs to the Rubiaceae family. To date, there are 130 known coffee plant species [3], of which Coffea arabica L. accounts for 60% of global production, followed by Coffea canephora Pierre ex A.Froehner with 40% [4], and Coffea liberica Bull ex Hiern with less than 1%. Coffea arabica (2n=4x=44) is the only tetraploid and autogamous species of the genus. The low diversity of cultivated varieties of this species can be attributed to its allotetraploid origin, reproductive biology (self-compatible) and recent evolution [5]. In Colombia, C. arabica occupies the largest planted area in the country (842,399 ha), with a harvest value of COP$11,143,095 in 2023 [6]. As a result, it is the leading agricultural export product, with 548,546 coffee growers deriving their livelihood from this crop. In other words, in Colombia approximately two million people currently depend on coffee for subsistence [7].
When conventional breeding methods are used, it can take between 25–30 years to produce a new variety of C. arabica with attributes of interest, such as resistance to Hypothenemus hampei, Ferrari (Coleoptera: Curculionidae: Scolytinae), commonly known as the coffee berry borer (CBB). This beetle depends on coffee berries for its survival, infests all coffee species in different proportions and causes the greatest economic losses to coffee crops, not only in Colombia but worldwide [8]. Genetic transformation, however, presents an alternative for developing pest-resistant C. arabica plants in less time than that required by conventional breeding by allowing gene transfer between different plant species and the subsequent expansion of their genetic pool. Yet an in vitro plant regeneration system is necessary to produce transgenic coffee plants. Because the multiplication potential of somatic embryogenesis (SE) is significantly higher than that of other regeneration methods [9,10,11], SE has been the method of choice not only for developing transgenic coffee plants [12,13,14,15,16], but also for the multiplication of F1 hybrids [9,17] and the propagation and conservation of elite genotypes with high genetic and epigenetic stability [9]. SE is traditionally defined as a process by which plant somatic cells can be dedifferentiated into totipotent embryogenic stem cells and redifferentiated into a somatic embryo capable of regenerating plants under appropriate culture conditions [18]. Another study conducted by Campos et al. [19] suggests that cells that are capable of differentiating into somatic embryos do not undergo dedifferentiation. These meristematic cells therefore retain totipotency and, with the appropriate stimulus, undergo multiplication and differentiation, forming new viable embryos.
Although somatic embryos have been obtained from coffee plant cells since 1970 [20], with numerous studies documenting the regeneration of somatic embryos over the last 50 years [9,17], the embryogenic capacity of coffee plant cells depends mainly on the genotype, which leads to the almost empirical development of specific protocols for each variety or clone [11,19,22]. Furthermore, other factors also influence the embryogenic response such as culture medium, growth regulators and gelling agents [25]. Embryos can be obtained directly and indirectly by SE. In direct SE (DSE), from 1 to 10 somatic embryos are regenerated directly on the explant without forming non-embryogenic callus [22]. The process is called low frequency SE due to the low number of embryos produced [21]. In contrast, indirect SE (ISE) is characterized by the formation of several clusters of friable embryogenic tissue (ET) that originate between 50–100 or more somatic embryos, which are produced from a disorganized tissue called non-embryogenic callus [11]. ISE can be considered as a type of high frequency SE because it produces many embryos [21].
Agrobacterium tumefaciens has been the preferred method for the genetic transformation of different plant species, including perennial species such as coffee [26] because it allows a low, stable copy number of the gene of interest to be efficiently inserted in the plant genome. Although T-DNA does not present preferential integration in specific chromosomal sequences or regions of gene expression [27], a common characteristic is a slight local A + T motif enrichment at the pre-integration site and microhomology between the T-DNA border sequences and the pre-integration site [28]. Because of the low efficiency of the genetic transformation of C. arabica and C. canephora [12,26,29], very few genes of interest, such as Cry1Ac from Bacillus thuringiensis that confers resistance to the coffee leaf miner Perileucoptera spp. [12], have been incorporated into the coffee plant genome. In the field, transgenic plants of C. canephora expressing the Cry1Ac protein presented fewer lesions than the susceptible controls and exhibited stable resistance [13]. In addition, transgenic plants of C. arabica expressing a gene encoding the α-amylase inhibitor (α-AI1) of Phaseolus vulgaris L., an active inhibitor of the digestive α-amylases of the CBB, have been produced [14]. Similarly, the Cry10Aa gene from B. thuringiensis has been transferred to the coffee plant genome, conferring resistance to the CBB [15]. The berries that express this Cry10Aa protein cause mortality in first-instar larvae, with less than 9% seed damage as compared with 100% damage in untransformed berries [16]. However, no information is available on the offspring of the transformed plants or commercial transgenic coffee varieties.
In C. arabica, the importance of evaluating the factors that influence the genetic transformation of C. arabica is clearly evidenced not only by the limitations faced in ET formation and the use of complex and inefficient genetic transformation methods, but also by the restrictions identified for successful regeneration and transformation (high genotypic dependence, need for exogenous hormones, Agrobacterium strain, method of infection and culture conditions pre- and post-inoculation with Agrobacterium). This study therefore aimed to develop an efficient and reproducible method for the genetic transformation of C. arabica by evaluating the factors that affect the integration of the uidA gene, which encodes GUS, into its genome.
2. Materials and Methods
2.1. In Vitro Culture Conditions
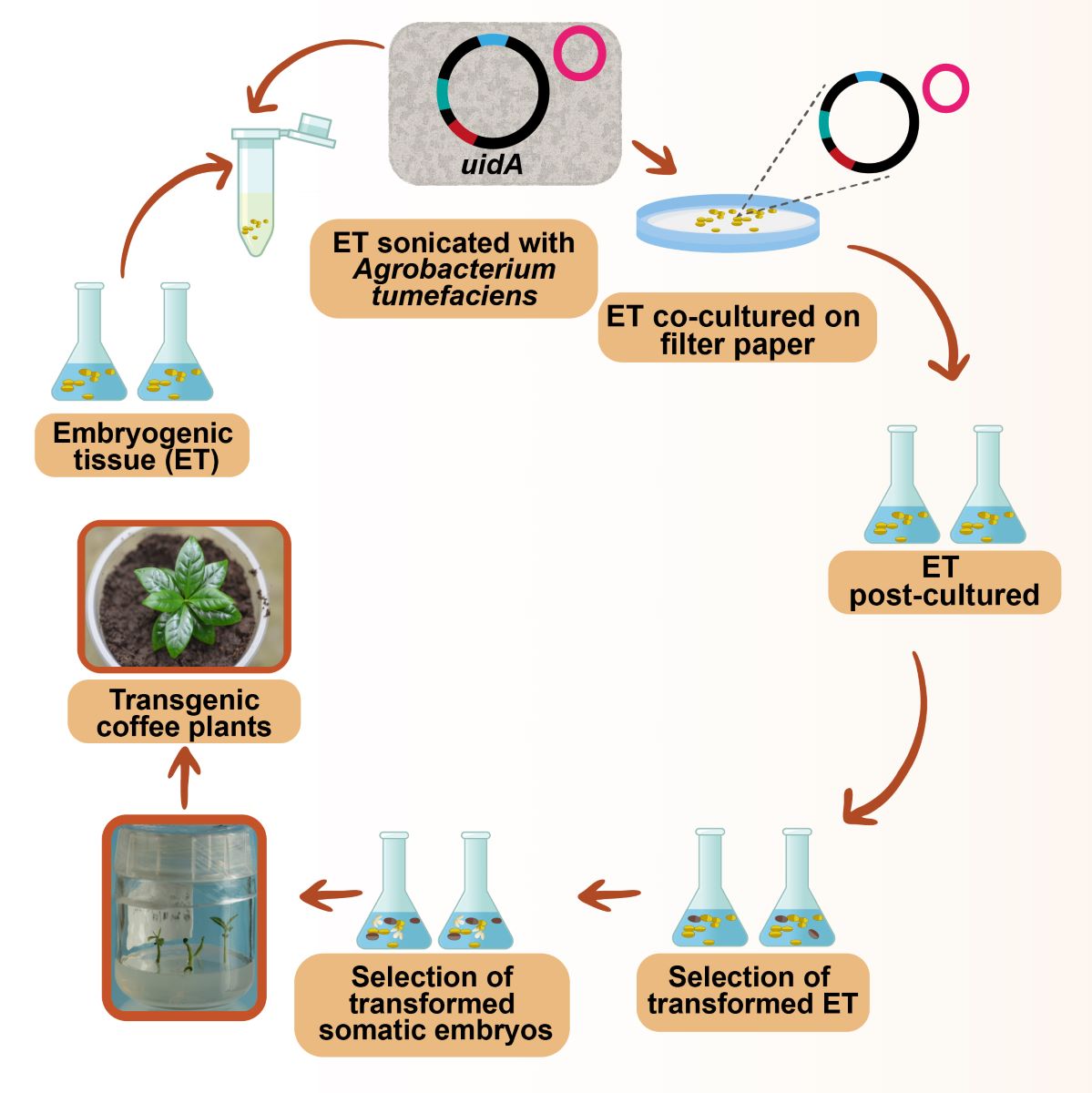
To induce indirect SE, leaves were collected at the Naranjal Experiment Station (Chinchiná, department of Caldas, Colombia) from coffee trees of genotypes BK.620 and BI.625 produced by the crossing of C. arabica cv. Caturra and the Timor hybrid. Climatic conditions at this site are as follows: average annual temperature, 20.8 °C; average relative humidity, 78%; annual rainfall, 2,686 mm; solar brightness, 1,817 hr yr-1. The ET was regenerated as described by Molina et al. [22]. Young leaves were first disinfected with commercial sodium hypochlorite supplemented with 1% Tween 80 for 10 min, followed by three washes with sterile distilled water, after which 1 cm2 explants were placed in culture medium supplemented with Murashige and Skoog (MS) [53] salts, 1 mg l−1 thiamine, 1 mg l−1 nicotinic acid, 1 mg l−1 pyridoxine, 100 mg l−1 myoinositol, 37 mg l−1 cysteine, 1 mg l−1 2,4-dichlorophenoxyacetic acid (2,4-D), 8 mg l−1 kinetin, 30 g l−1 sucrose and 8 g l−1 Bacto Difco Agar. After 1 month of culturing, explants was subcultured to a similar secondary culture medium supplemented with half-strength MS salts (MS/2) containing 3 mg l−1 6-benzylaminopurine (BAP) instead of kinetin and 2,4-D. Cultures were left in the dark at 25±2 °C until ET was formed. Within 4–6 months, friable ET developed on explants on the secondary medium. At that time, 1 g ET was transferred to 250-ml Erlenmeyer flasks containing 100 ml MS/2 liquid proliferation medium, 10 mg l−1 thiamine, 0.2 mg l−1 myoinositol, 0.04 mg l−1 cysteine, 3 mg l−1 6-BAP and 30 g l−1 sucrose. The cell suspensions were propagated by orbital shaking at 100 rpm in the dark at 25±2 °C, subculturing at 20-day intervals. “Early” ET was maintained for 6–8 months in liquid proliferation medium, and “differentiated” ET was maintained for 12–14 months in the same liquid medium. During pre-culturing, co-culturing and post-culturing, the ET was maintained in a differentiation medium with MS/2 salts, 10 mg l−1 thiamine, 0.1 mg l−1 myoinositol, 0.04 mg l−1 cysteine, 0.5 mg l−1 kinetin and 30 g l−1 sucrose. The solid culture medium was solidified with 2.8 g l−1 Phytagel, and cultures were left in the dark at 25±2 °C.
2.2. Agrobacterium Tumefaciens Strains and Vectors
Agrobacterium tumefaciens strains EHA105 and LBA4404 were used, and the Center for the Application of Molecular Biology to International Agriculture (CAMBIA), located in Canberra, Australia, provided the vectors pC1301 and pC2301. The T-DNA region comprises the reporter gene uidA, which encodes the protein GUS in these vectors. This reporter gene contains an intron for specific expression in eukaryotes and this gene is under control of the cauliflower mosaic virus promoter CaMV35S. In addition, in these vectors the genes for selection for resistance in plants, the hygromycin phosphotransferase selection gene (hptII) for pC1301, and gene selection for neomycin phosphotransferase (nptII) for pC2301, are under the control of the same promoter. A mixture was prepared with 30 µl electrocompetent cells of A. tumefaciens strains EHA105 and LBA4404 and 1 µl DNA (100 ng/µl) from vectors pC1301 or pC2301. The mixture was carefully shaken and transferred to a previously chilled electroporation cuvette (0.2-cm gap), which was then placed in the cuvette holder. Electroporation was performed in a Bio-Rad electroporator under the following conditions: 25 µF, 2.5 volts, and 200 W. Immediately after the electrical pulse, 800 µl super optimal broth with catabolite repression (SOC) medium was added, and the mixture was transferred to a microcentrifuge tube and incubated at 28 °C under orbital shaking at 225 rpm for 1 hr. After this time, dilutions of 10−0, 10−1, 10−2 and 10−3 were made in selective Luria-Bertani (LB) medium supplemented with kanamycin (100 mg l−1). Subsequently, individual colonies of each vector were selected in LB medium supplemented with kanamycin (100 mg l−1) at 28 °C for 12 hr. After confirming the introduction of the vectors into the A. tumefaciens strains by restriction enzyme digestion, competent A. tumefaciens cells were prepared containing vectors pC1301 and pC2301 in 10% glycerol.
2.3. Agrobacterium Transformation
Agrobacterium strains transformed with the abovementioned vectors were cultured in 5 ml LB medium supplemented with 50 µg ml−1 kanamycin and 5 g l−1 sucrose for 22 hr at 28 °C under orbital shaking conditions at 200 rpm. Strains were subsequently centrifuged at 3000 rpm for 10 min, and the precipitate was resuspended by adjusting the A600 to 0.5 in LB medium supplemented with 50 µg ml−1 kanamycin, 5 g l−1 glucose and 100 µM acetosyringone, and then incubated for 22 hr at 28 °C under orbital shaking conditions at 100 rpm. The precipitate was finally resuspended by adjusting the A600 to 0.5 in the differentiation medium. The ET was pre-cultured for 0–8 days in a solid differentiation medium supplemented with 100 µM acetosyringone. Subsequently, 1.0 ml of the A. tumefaciens suspension containing the transformation vectors was added to microcentrifuge tubes at a packed cell volume of 0.5 ml ET. Tubes were after placed individually in the center of a sonicator bath (Ultrasonic Cleaner, Cole -Parmer) and sonicated four times (0, 60, 120 and 300 sec) at 60 Hz. After 1 hr incubation in the bacterial suspension, the Agrobacterium suspension was then removed from the embryogenic tissue using a fine-tip micropipette, after which the tissue was deposited on sterile Whatman® filter paper (42.5 mm) to eliminate excess Agrobacterium. Embryogenic cells were subsequently co-cultured with 100 µM acetosyringone for four days in either 50-ml Erlenmeyer flasks with 25 ml liquid differentiation medium or Petri dishes with 25 ml solid differentiation medium on sterile Whatman® filter paper (42.5 mm). Co-culturing time on the solid medium was evaluated between 0–8 days. A 4-day post-culturing period followed co-culturing during which the ET was transferred to a liquid differentiation medium supplemented with 300 mg l-1 Claforan® (Hoechst). The selection process then began by adding 50 mg l-1 hygromycin for vector pC1301 and 20 mg l-1 geneticin for vector pC2301. Cultures were maintained at 26 °C under orbital shaking conditions at 100 rpm in the dark, sub-culturing at 20-day intervals.
The following tests were performed to evaluate the factors influencing the genetic transformation of coffee plant embryogenic cells:
- Sonication time: Early ET of genotype BI.625 without pre-culturing was sonicated in a suspension of A. tumefaciens LBA4405 containing vector pC1301 for 0, 60, 120 and 300 sec. After 1 hr incubation in the suspension, the ET was co-cultured for four days in both solid and liquid differentiation media.
- Co-culturing time: The ET of genotype BK.620 without pre-culturing was sonicated in a suspension of A. tumefaciens EHA105 containing vector pC1301 for 300 sec. After 1 hr incubation in the suspension, the ET was co-cultured in solid differentiation medium for 0–8 days.
- Pre-culturing time: The ET of genotype BK.620 was pre-cultured in the dark in solid differentiation medium supplemented with 100 µM acetosyringone for 0–8 days, followed by sonication of embryogenic cells for 300 sec in a suspension of A. tumefaciens EHA105 containing vector pC2301. After 1 hr incubation in the suspension, the ET was co-cultured in a solid differentiation medium for four days.
- Age of ET: Transformation efficiency of the uidA gene was evaluated using early and differentiated ET of genotype BI.625 without pre-culturing, followed by sonication for 300 sec of embryogenic cells in a suspension of A. tumefaciens LBA4404 containing vector pC1301. After 1 hr incubation in the suspension, the ET was co-cultured in a solid differentiation medium for four days.
- Agrobacterium strain: A. tumefaciens strains LBA4404 and EHA105 were evaluated with the vector pC1301.
- Transformation vector: Vectors pC1301 and pC2301 previously introduced into strain LBA4404 were compared.
- Coffee genotype: The early ETs of genotypes BI.625 and BK.620 were co-cultured with A. tumefaciens strain LBA4404 containing the vector pC1301. The protocol described for evaluating the age of ET was followed to evaluate Agrobacterium strains, transformation vectors and coffee plant genotypes.
2.4. Stable Transformation
2.4.1. Development of Transgenic Somatic Embryos
Proembryos were subcultured at 20-day intervals in a liquid proliferation medium with MS/2 salts, supplemented with selection agents until somatic embryos were formed. These embryos were then cultured in a germination medium with MS/2 salts, 1 mg l-1 thiamine, 1 mg l-1 nicotinic acid, 0.1 g l-1 myoinositol, 20 g l-1 sucrose, 7 g l-1 agar (Plant TC, PhytoTechnology), to which 300 mg l-1 Claforan® was added in addition to 50 mg l-1 hygromycin or 20 mg l-1 geneticin, depending on the vector used. The embryos remained in the dark until they produced the first pair of cotyledonary leaves. Plants were afterwards submitted to a 12-hr photoperiod (light intensity of 20 µmol.m-2sec-1) at 26 °C. When seedlings had produced four pairs of leaves, they were transferred to plastic pots containing a substrate composed of soil and kept in greenhouse conditions.
2.4.2. Histochemical GUS Assay
The method proposed by Jefferson [54] was used to perform histochemical testing for GUS. Transformed embryogenic cells as well as leaves and roots of transformed plants were incubated for 24 hr at 37 °C in an X-Gluc buffer solution (2 mM X-Gluc, 100 mM sodium phosphate buffer pH 8.0, 10 mM EDTA, 1 mM potassium ferricyanide, 1 mM potassium ferrocyanide and 20% v/v methanol). The tissue was cleaned using 95% ethanol (v/v). The transient expression of the uidA gene was determined as the total number of blue spots per gr dry weight of co-cultured ET using a binocular stereoscope. Blue coloration on both leaves and roots of transgenic plants was evaluated.
2.4.3. PCR and Southern Blot Analysis of Transgenic Plants
The DNA of a randomly selected transformed plant was extracted from leaf tissue frozen at -80 °C and vacuum dehydrated in a lyophilizer (Labconco, FreeZone 6 L) using the CTAB method [55]. This involved incubating 80 mg macerated tissue in 500 µl 2x CTAB buffer (CTAB 2% w/v, Tris 0.1 M, EDTA 0.02 M, NaCl 8.2% w/v, polyvinylpyrrolidone 2% w/v and ß-mercaptoethanol 1.5% v/v) and subsequent stirring at 65 °C for 30 min, after which a 500 µl of chloroform:isoamyl alcohol (24:1) was added and centrifuged at 14,000 rpm for 15 min at 4 °C for DNA purification. The DNA was precipitated by adjusting the suspension to a concentration of 0.18 M sodium acetate, 0.7 M NaCl and 500 µl of isopropyl alcohol. The DNA collected was washed with 70% ethanol and resuspended by incubation for 30 min at 37 °C in 100 µl ultrapure water with 2.5 µg RNase-A. The primers 5´-GATCAGCGTTGGTGGAAAGCGCG-3´ and 5´-CACCGAAGTTCATGCCAGTCCAGCG-3´ were used to amplify the uidA gene. The reaction mixture for PCR was composed of sterile millipore water, 1X PCR buffer, 3.0 mM MgCl2, 0.2 mM of each dinucleotide (dNTP), 0.8 µM primer, 2.5 U Taq polymerase (Gibco BRL) and 200 ng DNA for a final volume of 25 µl per tube. The mixture was incubated in a thermal cycler (PTC-200 MJ Research) under the following conditions: 1 cycle at 94 °C for 5 min; 40 cycles at 94 °C for 1 min, 60 °C for 1 min 30 sec and 72 °C for 3 min 30 sec; and an extension at 72 °C for 5 min. Eleven plants of each of the two transformation vectors, derived from independent transformation events, were selected among the plants that tested positive by PCR for molecular analysis via Southern blotting to determine the number of integration sites of the uidA gene in the coffee genome. The protocol reported by Noir et al. [56] was followed, and the DNA was digested with BglII and BsteII. The DNA fragments were then separated on 0.8% agarose gels, transferred to nylon membranes and hybridized with radioactive probes.
2.5. Experimental Design and Data Analysis
A series of seven experiments was conducted to evaluate the following factors affecting the genetic transformation of coffee: age of embryogenic cells, Agrobacterium strain, transformation vector, genotype, sonication times (0, 60, 120 and 300 sec) and co-culturing medium (solid and liquid), pre-culturing time and co-culturing time. For all experiments, the experimental unit consisted of either a 50-ml Erlenmeyer flask with 25 ml liquid differentiation medium or a Petri dish with 25 ml solid differentiation medium, totaling three experimental units per treatment. The response variable was the transient expression of the uidA gene measured as the number of blue spots per gr dry weight ET. Treatment effect was evaluated using a completely randomized experiment design in all experiments, except those of sonication time and co-culturing medium, in which an analysis of variance for completely randomized designs was performed, with a 4x2 factorial arrangement. Duncan's multiple comparison test was applied in the case of those experiments where the analysis of variance showed differences and the variation factors were qualitative. The regression or orthogonal polynomial test was used to identify differences between treatment groups for experiments with quantitative factors of variation.
3. Results
3.1. Age of ET
During the long-term maintenance of competent ETs for use as explants in genetic transformation, the cell cultures produced two types of embryogenic clusters: (1) early ET, composed of small, friable light-yellow clusters that were propagated in the proliferation medium between months 6 and 8 by subculturing at 20-day intervals (Figure 1a); and (2) differentiated ET, composed not only of yellow cell clusters larger than those of early ET but also of pro-embryos and small somatic embryos, which were cultured for 12–14 months in the proliferation medium and then subcultured at 20-day intervals (Figure 1b). The analysis of variance showed that the expression of the enzymatic marker GUS was significantly higher in early ET (P ≤0.0001) than in differentiated ET (Table 1).
3.2. Pre-Culturing Time
Pre-culturing time refers to culturing ET in a differentiation medium with the addition of acetosyringone to promote cell division before co-culturing with Agrobacterium [30]. The purpose of pre-culturing is to ensure that the tissues are sufficiently competent to resist bacterial infection and to increase the expression of virulence genes (vir) once the Agrobacterium suspension enters in contact with the ET, facilitating the cleavage and insertion of the T-DNA into the embryogenic cells [31].
The regression of the expression of the uidA gene according to pre-culturing time was linear, negative and significant (Figure 2). In other words, the ET of coffee plants propagated in liquid culture medium without pre-culturing presented the highest transient expression of GUS as compared with coffee embryogenic cells pre-cultured for 1–8 days in solid differentiation medium with acetosyringone, in which the expression of the uidA gene decreased (Figure 2).
3.3. Agrobacterium Strain, Transformation Vector and Coffee Genotype
The ability of Agrobacterium to infect different genotypes differs among strains, affecting transformation efficiency [30]. The strain LBA4404 was more effective than the strain EHA105 at infecting coffee embryogenic cells because the transient expression of the enzymatic marker GUS was significantly higher (Table 1). Additionally, vector pC1301 showed significantly higher transient expression of the uidA gene (Table 1, Figure 1c,f) than did vector pC2301 (Table 1, Figure 1d,g). The genotypes BK.620 and BI.625 did not differ in the number of blue spots per gr dry weight (Table 1), possibly because they have a common genetic origin [22].
3.4. Sonication Time and Co-Culturing Medium
The transient expression of GUS increased when the embryogenic cells in the Agrobacterium suspension were exposed to sonication between 60 and 300 sec in both liquid and solid culture media (Figure 3), followed by 1 hr incubation without shaking in the same bacterial suspension. A significantly higher number of blue spots (P ≤0.0001) was obtained with the longest sonication time tested (300 sec) compared with the number of blue spots per gr dry weight in embryogenic cells without exposure to sonication and the shortest sonication time evaluated (Figure 3).
GUS expression was significantly higher in ET co-cultured in the dark in solid differentiation medium supplemented with 100 µM acetosyringone on filter paper for 4 days (P ≤0.0001) than in the ET co-cultured in liquid culture medium, which presented a lower number of blue spots per gr dry weight (Table 1). The transient expression of GUS according to sonication time was higher with increasing sonication time between 60 and 300 sec in both culture media (Figure 3). The most significant transitory expression of GUS was obtained when embryogenic cells were subjected to 300 sec sonication followed by co-culturing in solid culture medium with MS/2 salts and 100 µM acetosyringone on filter paper, in comparison with the co-culturing in liquid culture medium, which showed a lower number of blue spots per gr dry weight at all sonication times tested (Figure 3).
3.5. Co-Culturing Time
The regression of the uidA gene expression in relation to co-culturing time was quadratic and significant (Figure 4) because GUS expression increased between 1–4 days, reaching its highest expression (P ≤0.0001) after 4 days co-culturing in the dark (R2=0.89). Prolonged periods (between 5–8 days) decreased the expression of the uidA gene due to Agrobacterium overgrowth.
3.6. Regeneration of Transgenic Plants and Molecular Analysis
It took one year from the development of pro-embryos until the formation of somatic embryos in liquid differentiation medium supplemented with 50 mg l-1 hygromycin for vector pC1301 and 20 mg l-1 geneticin for vector pC2301. The inhibition of untransformed cells was higher in ET samples with 50 mg l-1 hygromycin (Figure 1c) than in those with 20 mg l-1 geneticin (Figure 1d). Embryos were then cultured for 3 months in the dark in a germination medium without growth regulators and the selection agents hygromycin and geneticin (Figure 1e), depending on the vector used, until the first pair of dicotyledonous leaves were produced, after which plants were incubated under a 12-hr photoperiod for 4 months. The leaves of hygromycin-selected transgenic plants (Figure 1f) as well as those selected with geneticin (Figure 1g) showed blue coloration when reacting with X-Gluc. The leaves of untransformed plants, however, did not show blue coloration with X-Gluc (Figure 1h). Fifty transformed seedlings derived from independent transformation events with vector pC1301 y pC2301 with four pairs of dicotyledonous leaves and good root development were transferred to sterile soil under greenhouse conditions (Figure 1i,j). The presence of a 1600-bp amplicon corresponding to the uidA gene was confirmed in all plants by PCR. A total of 25 plants were transformed with vector pC2301 and the remaining 25 plants were transformed with vector pC1301 (Figure 5). However, no amplicons were detected in the untransformed control plants. Hybridization by Southern blotting confirmed the presence of a band corresponding to the uidA gene in coffee seedlings transformed with vectors pC1301 and pC2301. The presence of the GUS gene-coding region fragment was confirmed in 9 of the 11 plants transformed with vector pC2301 by digestion with enzymes Bgl II and Bste II (Figure 6 and Figure 7). Similarly, a coding region fragment of the gene hptII was confirmed in eight of the 11 plants transformed with vector pC1301 and digested with the enzyme XhoI (Figure 8 and Figure 9).
4. Discussion
To improve genetic transformation efficiency of Coffea arabica, this study evaluated the following factors that affect the development of transgenic plants: age of ET, pre-culturing time, Agrobacterium strain, transformation vector, coffee plant genotype, sonication time, co-culturing medium and co-culturing time.
An initial density of 10 gr fresh weight l-1 of ET proved optimal, renewing completely the culture medium at 20-day intervals. These results were similar to those obtained by Maciel et al., [32]. The age of ET was found to influence genetic transformation efficiency of coffee embryogenic cells. Early ET, which was cultured from 6 to 8 months in liquid proliferation medium, presented higher transformation efficiency of the uidA gene (P≤0.0001) as compared with differentiated ET, which was cultured from 12 to 14 months in the same culture medium (Table 1) This could be possibly attributed to the fact that early ET (Figure 1a) is composed of clusters of cells undergoing division with irregular, gapped and incomplete cell walls [33] and, as a result, susceptible to the formation of a higher number of deeper wounds that induce the release of phenolic compounds by sonication [31]. These, in turn, activate the expression of vir genes, which direct the cleavage of T-DNA and its subsequent integration into the coffee plant genome. However, in differentiated ET, which is composed of pro-embryos and small somatic embryos (Figure 1b), fewer and more superficial wounds are produced than in early ET. The infection by Agrobacterium is therefore lower, as is the expression of vir genes and T-DNA cleavage. Similarly, the yellow embryogenic callus of C. arabica variety Caturra propagated for seven months in semi-solid proliferation medium presented the highest transformation efficiency of the GFP5 reporter gene [26]. On the contrary, primary ET presented a very low transformation efficiency and, as the age of the embryogenic culture increased, the transformation efficiency gradually increased until it reached its maximum potential between 7–9 months [26]. These results are similar to those of the study carried out by Etienne et al. [9], in which two development stages were characterized during the long-term maintenance of embryogenic cultures to obtain F1 coffee hybrids: (1) cell aggregation in the multiplication phase and embryo generation in the differentiation phase, both stages of which have high embryogenic potential.
These studies demonstrate that the age of ET is one of the most decisive factors for improving genetic transformation efficiency in coffee plants. In addition, one of the main obstacles for the genetic transformation of perennial species such as coffee is the low availability of abundant ET in optimal physiological conditions that ensures the highest transformation efficiency. Although in the case of coffee it is possible to produce ET year-round [26], results of this study indicated that between 6–8 months are necessary for propagation, which is similar to that reported by Ribas et al. [26].
On the other hand, the composition of the nutritive medium used for ET proliferation prior to transformation can influence its physiological condition and transformation efficiency; the half-strength macro- and micronutrients of MS salts (MS/2) used in this study for the long-term propagation of ET maintained it in optimal conditions for its genetic transformation. Similarly, the MS/2 culture medium allowed the efficient proliferation of embryogenic callus of variety Caturra for its subsequent genetic transformation [26] as compared with full-strength (MS) and quarter-strength (MS/4) salts that affected the capacity of ET to regenerate somatic embryos [26].
In some species, such as Cicer arietinum L., a pre-culturing time in a culture medium is required for explants to reach their optimal physiological condition for Agrobacterium colonization, without which it is difficult to differentiate the embryogenic cells transformed into seedlings [30]. In this study, coffee plant ET without pre-culturing presented the highest expression of the uidA gene as compared with coffee embryogenic cells pre-cultured between 1–8 days in a medium supplemented with 100 µM acetosyringone (Figure 2). These findings indicate that the embryogenic cells of C. arabica undergoing cell division, maintained for 6–8 months in liquid proliferation medium supplemented with BAP as the only growth hormone, are suitable for Agrobacterium infection and production of transgenic plants. This corroborates that the multiplication of embryogenic cells in liquid medium increases their capacity to produce somatic embryos [9] and that cytokinins are essential for ET proliferation without requiring auxins after the induction of totipotent cells [34]. Similarly, embryogenic calli of variety Caturra propagated for 7–9 months in semi-solid medium without pre-culturing are suitable for gene transfer [26]. In Hevea brasiliensis Müll. Arg, the highest GUS transformation efficiency was also observed in explants that were not pre-cultured in a medium supplemented with 100 µM acetosyringone [35]. In contrast, in C. arietinum the transient expression of GUS was 76.67% higher in explants pre-cultured for 5 days in a medium without acetosyringone [36]. Whereas in another study conducted by Sadhu et al. [37] (2022), wounded embryo axes of C. arietinum pre-cultured for 3 days in shoot induction medium showed maximum transformation frequency. Moreover, the induction of competent cells of embryo axes of P. vulgaris required a 12-week pre-culturing time without acetosyringone, which was critical to its genetic transformation success [38].
The use of sonication in various species, such as H. brasiliensis, C. arietinum, Glycine max L., and Passiflora cincinnata Mast., among others [35,37,39,40,41], as well as in C. canephora [42], increased the genetic transformation efficiency of the uidA gene. Likewise, this study revealed that micro-wounds of coffee embryogenic cells exposed to 300 sec sonication were larger, more numerous and located deeper than those of cells exposed to 60 and 120 sec sonication. These results are similar to those observed in G. max, where more extensive and deeper micro-wounds were produced with increasing sonication time, with a transient expression so high that individual foci could not be distinguished in some tissues [39], favoring the transfer and integration of the uidA gene in the coffee genome. In a study recently carried out by Saravanan et al. [40], the highest transformation efficiency of apical meristems of G. max was achieved using 600-sec sonication compared with other times tested, while a combination of vacuum infiltration and sonication, each for 600 sec, produced the highest efficiency transformation. Additionally, in another study conducted by Wu et al. [43] in Vitis vinifera cv. Chardonnay, a sonication time of 240 sec followed by a vacuum infection of 300 sec favored A. tumefaciens colonization. In addition, transgenic plants of Pisum sativum L. were developed using 30 sec sonication and 3 min vacuum infiltration [44]. In contrast, agroinfiltration by injection followed by abrasion of the abaxial epidermis of explants of C. arabica variety Catuaí resulted in significantly higher transient expression of the uidA gene than when the epidermis was not rubbed or when sonication was used for 60 sec [45]. Despite the differences in the methods used to produce wounds in the tissues, these studies showed that wounds increased the infection by Agrobacterium and, consequently, the number of transformed cells.
Similarly, these micro-wounds generated during sonication in coffee embryogenic cells stimulate the expression of vir genes, such as acetosyringone and a variety of monosaccharides, including glucose, galactose and arabinose [31], as well as the culture medium used for the growth of Agrobacterium, which explains why the incubation of coffee ET with a bacterial suspension for 1 hr post-sonication without agitation increased the excision and transfer of the uidA gene to embryogenic cells, as previously shown in H. brasiliensis embryogenic cells incubated in bacterial suspension for 18 min post-sonication [35]. Sonicated coffee embryogenic cells also showed normal growth up through the formation of transgenic plants, confirming the observations of Trick and Finer [39], who established that sonication could affect the growth of ET immediately after exposure. However, after two weeks, the ET proliferated under conditions similar to those of the control without sonication. These results demonstrated that the sonication-assisted Agrobacterium transformation increases GUS expression in the embryogenic cells of C. arabica.
Different Agrobacterium strains, such as LBA4404, C58, EHA101 and EHA105, are suitable for the transformation of coffee ET [26]. In this study, the expression of the uidA gene was higher with A. tumefaciens strain LBA4404 as compared with strain EHA105 (Table 1). Similarly, the highest transformation efficiency of Cyclamem persicum Mill. was achieved by inoculating explants of variety Pure White with A. tumefaciens strain LBA4404 [46]. Moreover, C. canephora and C. arabica plants transformed with A. tumefaciens strain LBA4404, which contains the cry1Ac gene, presented resistance to Perileucoptera spp. [12]. In contrast, leaf explants of C. arabica variety Catuaí agroinfiltrated with A. tumefaciens strain GV3101 showed a higher expression of the uidA gene than did leaves agroinfiltrated with strains LBA4404 and ATHV [45]. In several studies carried out with Cucumis sativus L., Agrobacterium strain EHA105 presented a transformation efficiency up to three times higher than that of strain LBA4404 [30]. On the other hand, in the present study there were no significant differences in GUS expression between the two coffee genotypes BK.620 and BI.625, probably because they are advanced lines of the Colombia variety obtained by the crossing of variety Caturra and the Timor Hybrid that share a common genetic origin (Table 1).
Co-culturing of ET using a solid differentiation medium with MS/2 salts and 100 µM acetosyringone on filter paper for four days resulted in the highest expression of the uidA gene (Table 1). In addition, in C. sativus the co-culturing in solid medium on filter paper suppressed the explant necrosis, which led to a higher regeneration efficiency [47]. On the other hand, studies conducted by Duan et al. [48] in immature embryos of the inbred maize line Qi319 evidenced that a high transformation efficiency was achieved with Agrobacterium by co-culturing on dry filter paper in solid culture medium. Whereas Pavlichenko and Protopopova [49] established a simplified transformation method in Berlin poplar (Populus x berolinensis K. Koch), using internodal sections without axillary buds co-cultured directly on the solid medium surface in bacterial suspension drops, without rinsing, and then drying the suspension with sterile paper post-inoculation. The lower GUS expression efficiency in ET co-cultured in liquid medium can probably be attributed to the constant agitation, which restricts bacteria colonization to the areas between ET lobes. In contrast, co-culturing in solid culture medium favors the growth of Agrobacterium over the entire cell suspension surface. Co-culturing on filter paper also benefits the transfer of T-DNA to embryogenic cells, because Agrobacterium tends to increase the integration of T-DNA to embryogenic cells, attributed to the low availability of nutrients on filter paper.
A 4-day co-culturing time of C. arabica ET in solid medium with acetosyringone on filter paper produced the highest transient expression of GUS. Moreover, the genetic transformation of embryogenic calli was achieved in C. canephora with a 5-day co-culturing time [50]; similar to what occurred in C. arabica variety Caturra [26]. Different co-culturing times have been used in other species. For example, the highest transformation frequency was produced in calli of Oriza sativa L. co-cultured for 2 days in co-culturing medium with a concentration of 300 µM acetosyringone [51]. Similarly, in C. arietanum a 3-day co-culturing time was optimal for seed transformation, with 78.33% GUS expression and 72.2% regeneration [36]. These results agree with those found by Sadhu et al. [37] who found that a 3-day co-culturing period in a medium containing 100 µM acetosyringone was optimum for efficient chickpea transformation. Whereas, in the case of H. brasiliensis, a 3.5-day co-culturing time in the dark showed the highest number of blue spots per somatic embryo [35].
This study evaluated most of the factors affecting the genetic transformation of coffee embryogenic suspensions. A method to transfer genes conferring characteristics of interest to coffee plants, including resistance to pests such as the CBB, was developed with a 1.40% transformation efficiency. An alternative to developing varieties with resistance to H. hampei is genetic transformation by the transfer of genes that encode insecticidal proteins that inhibit digestive enzymes, which in turn digest proteins and carbohydrates in the midgut. One such inhibitor is Lupinus bogotensis aspartic protease inhibitor (LbAPI), which was found to be highly effective in inhibiting CBB aspartic proteases in vitro, with a mean inhibitory concentration (IC50) of 2.9 µg. In vivo, the concentration of recombinant LbAPI required to cause 50% mortality in H. hampei larvae in artificial diets was 0.91% [52]. In addition, the α-amylase inhibitor from P. vulgaris caused an 88% inhibition of the α-amylase activity of the CBB, and transgenic plants expressing this inhibitor presented delayed borer development [14]. Although the development of transgenic plants and their subsequent planting in the field can be challenging, this is indeed an alternative for controlling H. hampei in coffee crops.
Author Contributions
Conceptualization, D.M. and R.A.; methodology, D.M. and R.A.; validation, D.M. and R.A.; formal analysis, D.M.; investigation, D.M.; R.A.; writing —original draft preparation, D.M.; writing —review and editing, D.M.; R.A.; project administration, D.M.; R.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Coffee Research Center (Cenicafé) (Crossref Funder ID 100019597).
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Acknowledgments
We sincerely appreciate to Denis L. Osorio, Claudia Velásquez and Hernán Diaz for their assistance and to Dr. Germán Moreno and Hernando Cortina for their invaluable collaboration in different aspects of this study.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results
References
- FAOSTAT. Food and Agriculture Organization of the United Nations, Statistics Division. 2024, http://faostat.fao.org [Accessed July 26, 2024].
- International Coffee Organization. Coffee Development Report 2022. 2022, www.ico.org/documents/cy2022-23/annual-review-2021-2022-e.pdf. [Accessed June 24, 2024].
- Davis, A.P.; Rakotonasolo, F. Six new species of coffee (Coffea) from northern Madagascar. Kew Bull. 2021, 76, 497–511. [CrossRef]
- International Coffee Organization. Coffee Market Report. www.ico.org/documents/cy2022-23/cmr-1222-e.pdf. 2023, [Accessed April 24, 2024].
- Scalabrin, S.; Magris, G.; Liva, M.; Vitulo, N.; Vidotto, M.; Scaglione, D.; Del Terra, L.; Ruosi, M.R.; Navarini, L.; Pellegrino, G.; Mier y Teran, J.C.; Toniutti, L.; Suggi, F.; Cerutti, M.; Di Gaspero, G.; Morgante, M. A chromosome-scale assembly reveals chromosomal aberrations and exchanges generating genetic diversity in Coffea arabica germplasm. Nat Commun. 2024, 15, 463. [CrossRef]
- Federación Nacional de Cafeteros de Colombia. Estadísticas cafeteras. 2024, http://federaciondecafeteros.org/wp/estadisticascafeteras [Accessed April 24, 2024].
- Federación Nacional de Cafeteros de Colombia. Sistema de información cafetero de Colombia. 2024, http://sica.cafedecolombia.com [Accessed April 24, 2024].
- Molina, D.; Moncada, M.P.; Cortina, H.; Benavides, P. Searching for a coffee variety with antibiosis effect to Hypothenemus hampei Ferrari (Coleoptera: Curculionidae). Euphytica. 2022, 218, 97. [CrossRef]
- Etienne, H.; Breton, D.; Breitler, J-C.; Bertrand, B.; Déchamp, E.; Awada, R.; Marraccini, P.; Léran, S.; Alpizar, E.; Campa, C.; Cuurtel, P.; Georget, F.; Ducos, J-P. Coffee somatic embryogenesis: How did research, experience gained and innovations promote the commercial propagation of elite clones from the two cultivated species?. Front. Plant Sci. 2018, 9, 1630. [CrossRef]
- Méndez-Hernández, H.A.; Galaz-Ávalos, R.M.; Quintana-Escobar, A.O.; Pech-Hoil, R.; Collí-Rodríguez, A.M.; Salas-Peraza, I.Q.M. In vitro conversion of Coffea spp. somatic embryos in SETIS™ bioreactor system. Plants. 2023, 12, 3055. [CrossRef]
- Avila-Victor, C.M.; Ordaz-Chaparro, V.M.; Arjona-Suárez, E.d.J.; Iracheta-Donjuan, L.; Gómez-Merino, F.C.; Robledo-Paz, A.. In vitro mass propagation of coffee plants (Coffea arabica L. var. Colombia) through indirect somatic embryogenesis. Plants. 2023, 12, 1237. [CrossRef]
- Leroy, T.; Henry, A.M.; Royer, M.; Altosaar, I.; Frutos, R.; Duris, D.; Philippe, R. Genetically modified coffee plants expressing the Bacillus thuringiensis cry1Ac gene for resistance to leaf miner. Plant Cell Rep. 2000, 19, 382–389. [CrossRef]
- Perthuis, B.; Vassal, J.M.; Fenouillet, C.; Leroy, T. Cry1Ac insecticidal protein levels in genetically modified Coffea canephora Pierre coffee plants were negatively correlated with the growth speed in a field experiment. Euphytica. 2014, 202, 373–383. [CrossRef]
- Albuquerque, É.V.; Bezerra, C.A.; Romero, J.V.; Valencia, J.W.; Valencia-Jiménez, A.; Pimenta, L.M., Barbosa, A.E.; Silva, M.C.; Meneguim, A.M.; Sá, M.E.; Engler, G.; de Almeida-Engler, J.; Fernandez, D.; Grossi-de-Sá, M.F. Seed-specific stable expression of the α-AI1 inhibitor in coffee grains and the in vivo implications for the development of the coffee berry borer. Trop. Plant Biol. 2015, 8, 98–107. [CrossRef]
- Valencia-Lozano, E.; Cabrera-Ponce, J.L.; Gómez-Lim, M.A.; Ibarra, J.E. Development of an efficient protocol to obtain transgenic coffee, Coffea arabica L., expressing the Cry10Aa toxin of Bacillus thuringiensis. Int. J. Mol. Sci. 2019, 20, 5334. [CrossRef]
- Valencia-Lozano, E.; Cabrera-Ponce, J.L.; Noa-Carrazana, J.C.; Ibarra, J.E. Coffea arabica L. resistant to coffee berry borer (Hypothenemus hampei) mediated by expression of the Bacillus thuringiensis Cry10Aa protein. Front. Plant Sci. 2021, 12, 765292. [CrossRef]
- Aguilar, M.E.; Wang, X.Y.; Escalona, M.; Yan, L.; Huang, L.F. Somatic embryogenesis of Arabica coffee in temporary immersion culture: Advances, limitations, and perspectives for mass propagation of selected genotypes. Front. Plant Sci. 2022, 13, 994578. [CrossRef]
- Salaün, C.; Lepiniec, L.; Dubreucq, B. Genetic and molecular control of somatic embryogenesis. Plants (Basel). 2021, 10, 1467. [CrossRef]
- Campos, N.A.; Panis, B.; Carpentier, S.C. Somatic embryogenesis in coffee: the evolution of biotechnology and the integration of omics technologies offer great opportunities. Front. Plant Sci. 2017, 8, 1460. [CrossRef]
- Staritsky, G. Embroid formation in calllus tissues of Coffea. Acta Bot. Neerl. 1970, 19, 509–514.
- Sondahl, M.R.; Sharp, W.R. High frecuency induction of somatic embryos in cultured leaf explants of Coffea arabica L. Z. Pflanzenphysiologie. 1977, 81, 395–408. [CrossRef]
- Molina, D.; Aponte, M.E.; Cortina, H.; Moreno, L.G. The effect of genotype and explant age on somatic embryogenesis of coffee. Plant Cell Tiss. Org. Cult. 2002, 71, 117–123. [CrossRef]
- Arimarsetiowati, R.; Daryono, B.S.; Astuti, Y.T.M.; Prastowo, E.; Semiarti, E. Regeneration and development of Coffea arabica L. plants through indirect somatic embryogenesis. Coffee Sci. 2023, 18, e182078. [CrossRef]
- Ibrahim, M.S.D.; Hartati, R.S.; Rubiyo, R.; Reflinur, R.; Purwito, A.; Sudarsono, S. Exploring indirect somatic embryogenesis and somaclonal variation for propagation of three Coffea arabica L. cultivars. Chil. J. Agric. Res. 2024, 84, 15–27. [CrossRef]
- Avila-Victor, C.M.; Arjona-Suárez, E.d.J.; Iracheta-Donjuan, L.; Valdez-Carrasco, J.M.; Gómez-Merino, F.C.; Robledo-Paz, A. Callus type, growth regulators, and phytagel on indirect somatic embryogenesis of coffee (Coffea arabica L. var. Colombia). Plants. 2023, 12, 3570. [CrossRef]
- Ribas, A.F.; Dechamp, E.; Champion, A.; Bertrand, B.; Combes, M-C.; Verdeil, J-L.; Lapeyre, F.; Lashermes, P.; Etienne, H. Agrobacterium-mediated genetic transformation of Coffea arabica L. is greatly enhanced by using established embryogenic callus cultures. BMC Plant Biol. 2011, 11, 92. [CrossRef]
- Shilo, S.; Tripathi, P.; Melamed-Bessudo, C.; Tzfadia, O.; Muth, T.R.; Levy, A.A. T-DNA-genome junctions form early after infection and are influenced by the chromatin state of the host genome. PLoS Genet. 2017, 13, e1006875. [CrossRef]
- Gelvin, S.B. Plant DNA repair and Agrobacterium T-DNA integration. Int. J. Mol. Sci. 2021, 22, 8458. [CrossRef]
- Canche-Moo, R.L.R.; Ku-Gonzalez, A.; Burgeff, C.; Loyola-Vargas, V.M.; Rodriguez-Zapata, L.C.; Castaño, E. Genetic transformation of Coffea canephora by vacuum infiltration. Plant Cell Tiss. Org. Cult. 2006, 84, 373–377. [CrossRef]
- Tan, J.; Lin, L.; Luo, H.; Zhou, S.; Zhu, Y.; Wang, X.; Miao, L.; Wang, H.; Zhang, P. Recent progress in the regeneration and genetic transformation system of cucumber. Appl. Sci. 2022, 12, 7180. [CrossRef]
- Gordon, J.E.; Christie, P. J. The Agrobacterium Ti plasmids. Microbiol. Spectr. 2014, 2, 10.1128/microbiolspec.plas-0010-2013. [CrossRef]
- Maciel, A.L.; Rodrigues, F.A.; Pasqual, M.; Carvalho, C.H. Large-scale, high-efficiency production of coffee somatic embryos. Crop. Breed. Appl. Biotechnol. 2016, 16, 102–107. [CrossRef]
- Parra-Vega, V.; Corral-Martínez, P.; Rivas-Sendra, A.; Seguí-Simarro, J.M. Induction of embryogenesis in Brassica napus microspores produces a callosic subintinal layer and abnormal cell walls with altered levels of callose and cellulose. Front Plant Sci. 2015, 6, 1018. [CrossRef]
- Martínez, N.; Dávila, C.; Morales, J.; Castro, K.; Martínez, N.; López, H.; Villalobos, E. 6-benzylaminopurine induces somatic embryogenesis in the staminodia of new genotypes of Theobroma cacao L. from the Papaloapan Basin of Mexico and differs from that of T. bicolor Bonpl. Plant Cell Tiss. Organ Cult. 2024, 157, 64. [CrossRef]
- Udayabhanu, J.; Huang, T.; Xin, S.; Cheng, J.; Hua, Y.; Huang, H. Optimization of the transformation protocol for increased efficiency of genetic transformation in Hevea brasiliensis. Plants. 2022, 11, 1067. [CrossRef]
- Srivastava, J.; Datta, S.; Mishra, S.P. Development of an efficient Agrobacterium-mediated transformation system for chickpea (Cicer arietinum). Biologia. 2017, 72, 153–160. [CrossRef]
- Sadhu, S.K.; Jogam, P.; Gande, K.; Banoth, R.; Suprasanna, P.; Peddaboina, V. Optimization of different factors for an Agrobacterium-mediated genetic transformation system using embryo axis explants of chickpea (Cicer arietinum L.). J. Plant Biotechnol. 2022, 49, 61–73. [CrossRef]
- Song, G-Q.; Han, X.; Wiersma, A.T.; Zong, X.; Awale, H.E.; Kelly, J.D. Induction of competent cells for Agrobacterium tumefaciens-mediated stable transformation of common bean (Phaseolus vulgaris L.). PLoS ONE. 2020, 15, e0229909. [CrossRef]
- Trick H.N.; Finer J.J. Sonication-assisted Agrobacterium-mediated transformation of soybean [Glycine max (L.) Merrill] embryogenic suspension culture tissue. Plant Cell Rep. 1998, 17, 482–488. [CrossRef]
- Saravanan, K.; Vidya, N.; Appunu, C.; Gurusaravanan, P.; Arun, M. A simple and efficient genetic transformation system for soybean (Glycine max (L.) Merrill) targeting apical meristem of modified half-seed explant. 3 Biotech. 2023, 13, 293. [CrossRef]
- Da Silva, M.L.; Paim Pinto, D.L.; Passos, A.B.; Marcelino-Guimaräes, F.C.; Bandini Rossi, A.; Krause, W.; de Carvalho, I.; Batista, D.; Rocha, D.; Otoni, W. Novel and efficient transformation of wild passion fruit (Passiflora cincinnata Mast.) using sonication-assisted Agrobacterium-mediated transformation. In Vitro Cell. Dev. Biol. Plant. 2021, 57, 380–386. [CrossRef]
- Mohanan, S.; Satyanarayana, K.V.; Sridevi, V.; Gowda, K.; Giridhar, P.; Chandrashekar, A., Ravishankar, G. Evaluating the effect and effectiveness of different constructs with a conserved sequence for silencing of Coffea canephora N-methyltransferases. J. Plant Biochem. Biotechnol. 2013, 23, 399–409. [CrossRef]
- Wu, J.; Zhang, J.; Hao, X.; Lv, K.; Xie, Y.; Xu, W. Establishment of an efficient callus transient transformation system for Vitis vinifera cv. 'Chardonnay'. Protoplasma. 2023, 261, 1–16. [CrossRef]
- Chandrasekaran, A.; Pilavadi, T.; Venkatachalam, V.; Umapathy, D.; Arockiam, A.J.V.; Singarayar, M.S.; Lee, G-J.; Markandan, M. Bio-engineered As-Ag-TiO2 nanoparticles enhance the genetic transformation of Pisum sativum L. via proton-coupled electron transfer-dependent alternative protonation. Ind. Crops Prod. 2024, 215, 118604. [CrossRef]
- Vargas-Guevara, C.; Vargas-Segura, C.; Villalta-Villalobos, J.; Pereira, L.F.P.; Gatica-Arias, A. A. simple and efficient agroinfiltration method in coffee leaves (Coffea arabica L.): Assessment of factors affecting transgene expression. 3 Biotech. 2018, 8, 471. [CrossRef]
- Mirzaee, M.H.; Emadpour, M. Advances in the transformation of Cyclamen persicum Mill. through direct regeneration based on an optimized kanamycin selection scheme. Mol. Biotechnol. 2024, 66, 311–320. [CrossRef]
- Nanasato, Y.; Konagaya, K.I.; Okuzaki, A.; Tsuda, M.; Tabei, Y. Improvement of Agrobacterium-mediated transformation of cucumber (Cucumis sativus L.) by combination of vacuum infiltration and co-cultivation on filter paper wicks. Plant Biotechnol. Rep. 2013, 7, 267–276. [CrossRef]
- Duan, X.; Zheng, L.; Sun, J.; Liu, W.; Wang, W. An, H. Co-culturing on dry filter paper significantly increased the efficiency of Agrobacterium-mediated transformations of maize immature embryos. Physiol. Mol. Biol. Plants. 2019, 25, 549–560. [CrossRef]
- Pavlichenko, V.V.; Protopopova, M.V. Simplified method for Agrobacterium-mediated genetic transformation of Populus x berolinensis K. Koch. Methods Protoc. 2024, 7, 12. [CrossRef]
- Casarin, T.; Freitas, N.C.; Pinto, R.T.; Breitler J-C.; Zebral Rodrigues, L.A.; Marraccini, P.; Etienne, H.; Cardamone, L.E.; Carvalho, A.; Vilela, L. Multiplex CRISPR/Cas9-mediated knockout of the phytoene desaturase gene in Coffea canephora. Sci. Rep. 2022, 12, 17270. [CrossRef]
- Anwaar, S.; Jabeen, N.; Ahmad, K.S.; Shafique, S.; Irum, S.; Ismail, H.; Ullah, S.; Tahir, A.; Mehmood, N.; Gleason, M.L. Cloning of maize chitinase 1 gene and its expression in genetically transformed rice to confer resistance against rice blast caused by Pyricularia oryzae. PLoS ONE. 2024, 19, e0291939. [CrossRef]
- Molina, D.; Patino, L.; Quintero, M.; Cortes, J.; Bastos, S. Effects of the aspartic protease inhibitor from Lupinus bogotensis seeds on the growth and development of Hypothenemus hampei: an inhibitor showing high homologywith storage proteins. Phytochemistry. 2014, 98, 69–77. [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays tobacco tissues cultures. Physiol. Plant. 1962, 15, 472–497. [CrossRef]
- Jefferson R.A. Assaying chimeric genes in plants: the GUS gene fusion system. Plant Mol. Biol. Rep. 1987, 5, 387–405. [CrossRef]
- Doyle, J.J.; Doyle, J.L.A. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15.
- Noir, S.; Patheyron, S.; Combes, M.C.; Lashermes, P.; Chalhoub, B. Construction and characterisation of a BAC library for genome analysis of the allotetraploid coffee species (Coffea arabica L.). Theor. Appl. Genet. 2004, 109, 225–230. [CrossRef]
Figure 1.
Coffee plants transformed with the uidA gene. (a), X-gluc staining of early ET, composed of small embryogenic clusters; (b), X-Gluc staining of differentiated ET composed of pro-embryos and somatic embryos; (c), X-Gluc staining of selected embryogenic cells treated with 50 mg l-1 hygromycin transformed with vector pC1301; (d), X-Gluc staining of selected embryogenic cells treated with 20 mg l-1 geneticin transformed with vector pC2301 ; (e), somatic coffee embryos transformed with A. tumefaciens pC1301; (f), X-Gluc staining of plant leaves transformed with vector pC1301; (g), X-Gluc staining of leaves and roots of plants transformed with vector pC2301; (h), X-Gluc staining of untransformed plant leaves and roots; (i), transgenic coffee plants co-cultured with A. tumefaciens pC1301; (j) transgenic coffee plants co-cultured with A. tumefaciens pC2301.
Figure 1.
Coffee plants transformed with the uidA gene. (a), X-gluc staining of early ET, composed of small embryogenic clusters; (b), X-Gluc staining of differentiated ET composed of pro-embryos and somatic embryos; (c), X-Gluc staining of selected embryogenic cells treated with 50 mg l-1 hygromycin transformed with vector pC1301; (d), X-Gluc staining of selected embryogenic cells treated with 20 mg l-1 geneticin transformed with vector pC2301 ; (e), somatic coffee embryos transformed with A. tumefaciens pC1301; (f), X-Gluc staining of plant leaves transformed with vector pC1301; (g), X-Gluc staining of leaves and roots of plants transformed with vector pC2301; (h), X-Gluc staining of untransformed plant leaves and roots; (i), transgenic coffee plants co-cultured with A. tumefaciens pC1301; (j) transgenic coffee plants co-cultured with A. tumefaciens pC2301.

Figure 2.
Regression of the transient expression of the uidA gene according to pre-culturing time.
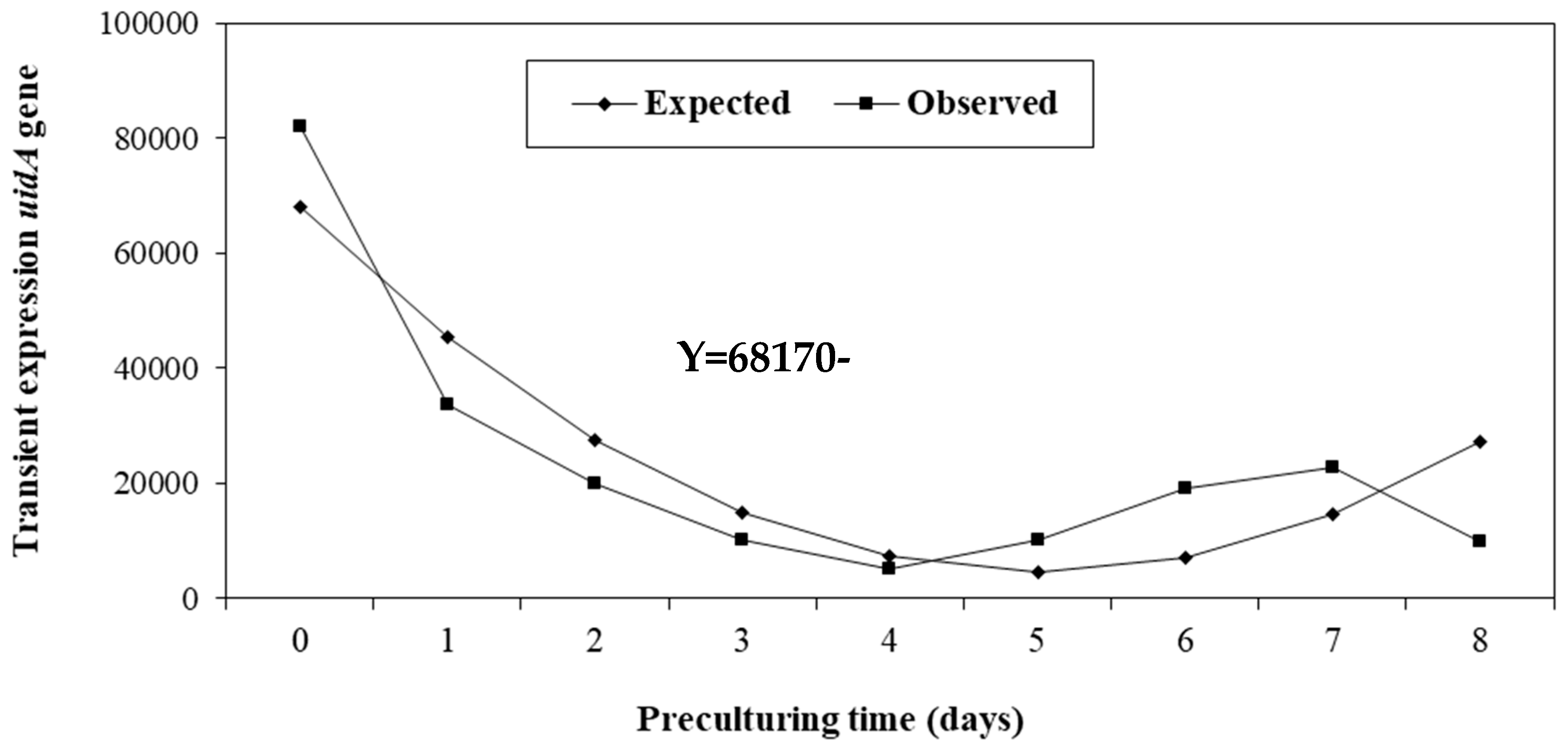
Figure 3.
Transient expression of the uidA gene according to sonication time in liquid and solid co-culturing media.
Figure 3.
Transient expression of the uidA gene according to sonication time in liquid and solid co-culturing media.
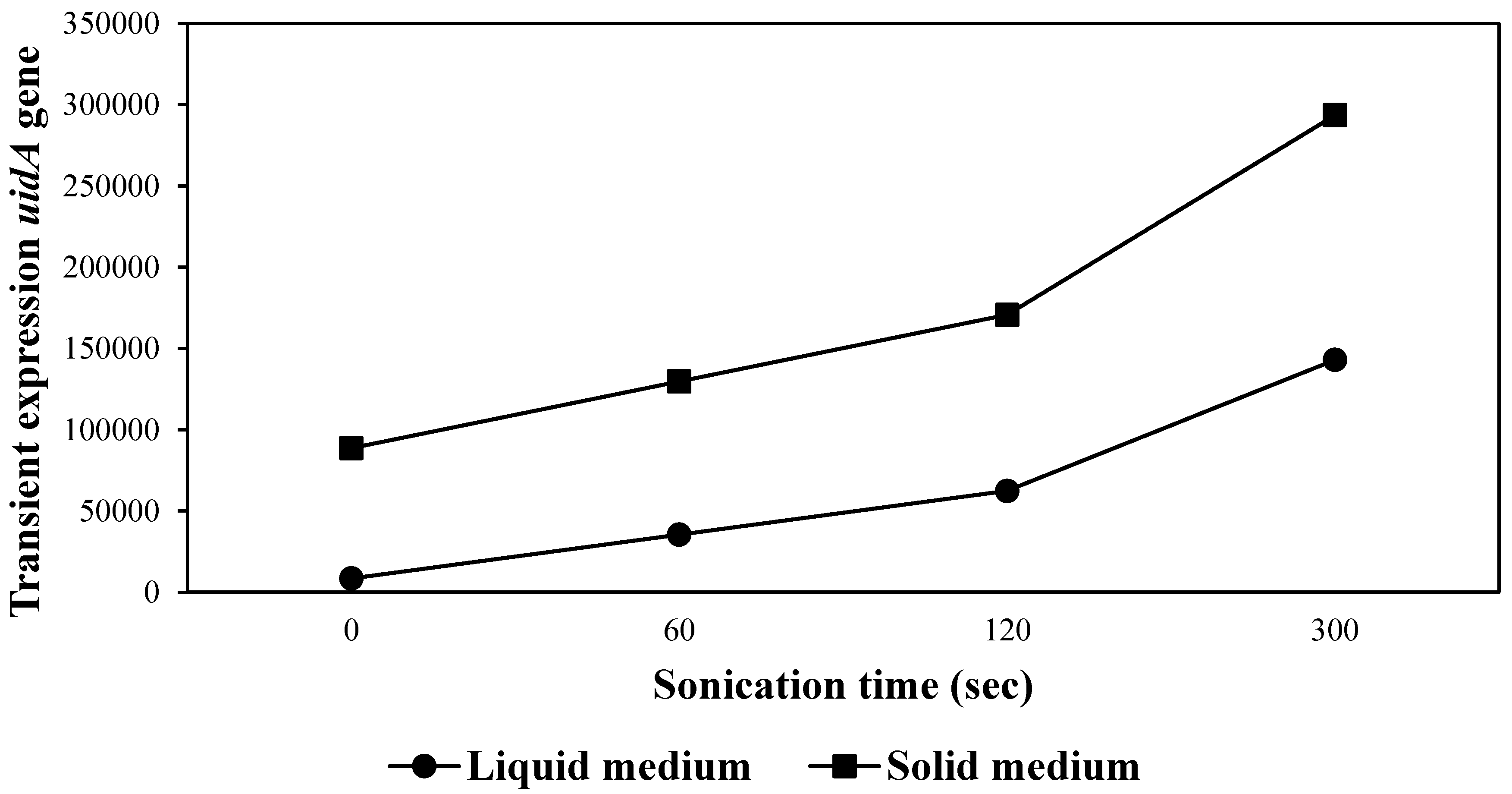
Figure 4.
Regression of the transient expression of the uidA gene according to the co-culturing time.
Figure 4.
Regression of the transient expression of the uidA gene according to the co-culturing time.
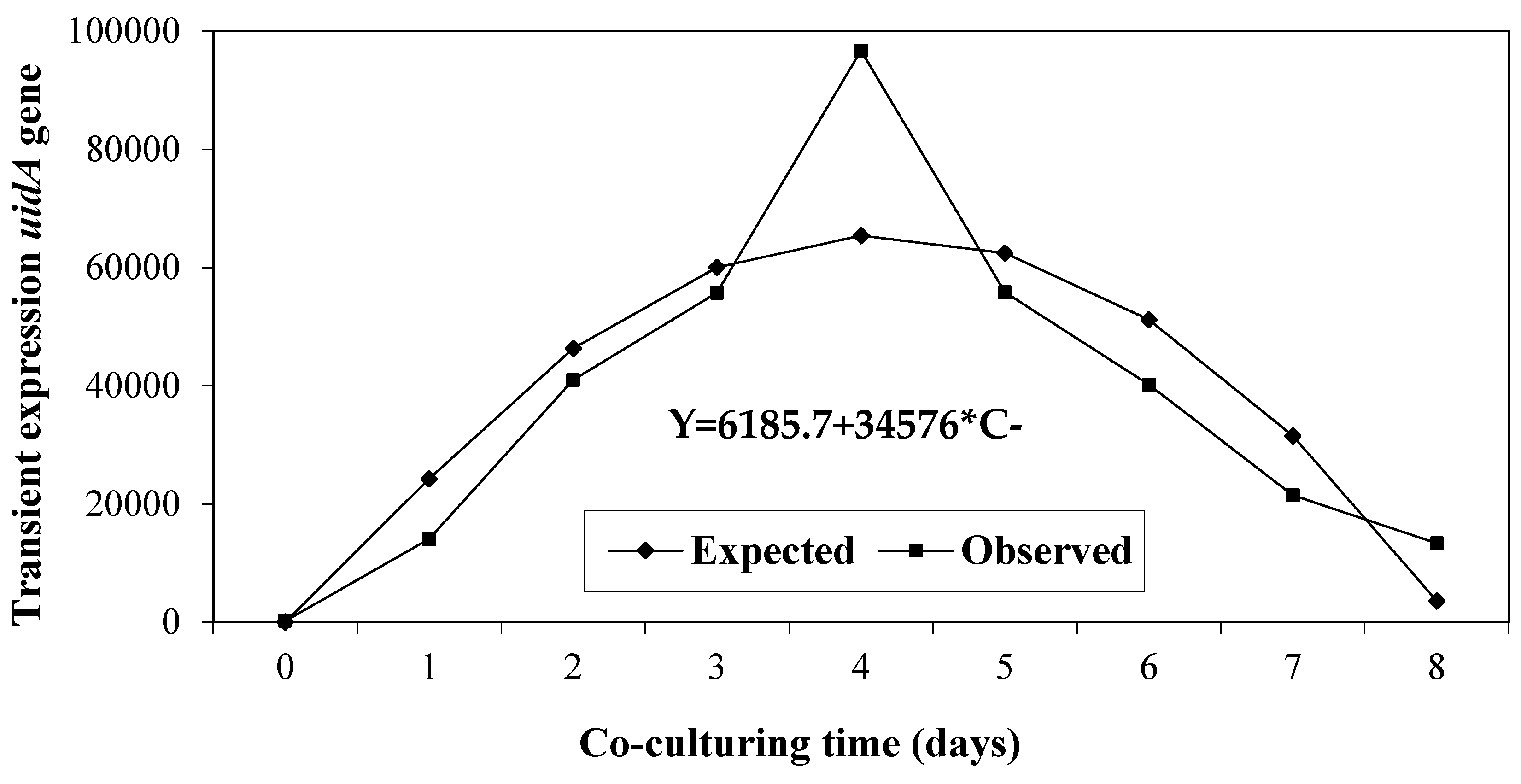
Figure 5.
Detection of the uidA gene in transgenic coffee plants. Lane 1, Molecular weight marker Low DNA Mass Ladder; lanes 2, 3, leaves of plants transformed with vector pC2301; lanes 4 and 5, leaves of plants transformed with vector pC1301; lane 6, untransformed plant leaves.
Figure 5.
Detection of the uidA gene in transgenic coffee plants. Lane 1, Molecular weight marker Low DNA Mass Ladder; lanes 2, 3, leaves of plants transformed with vector pC2301; lanes 4 and 5, leaves of plants transformed with vector pC1301; lane 6, untransformed plant leaves.
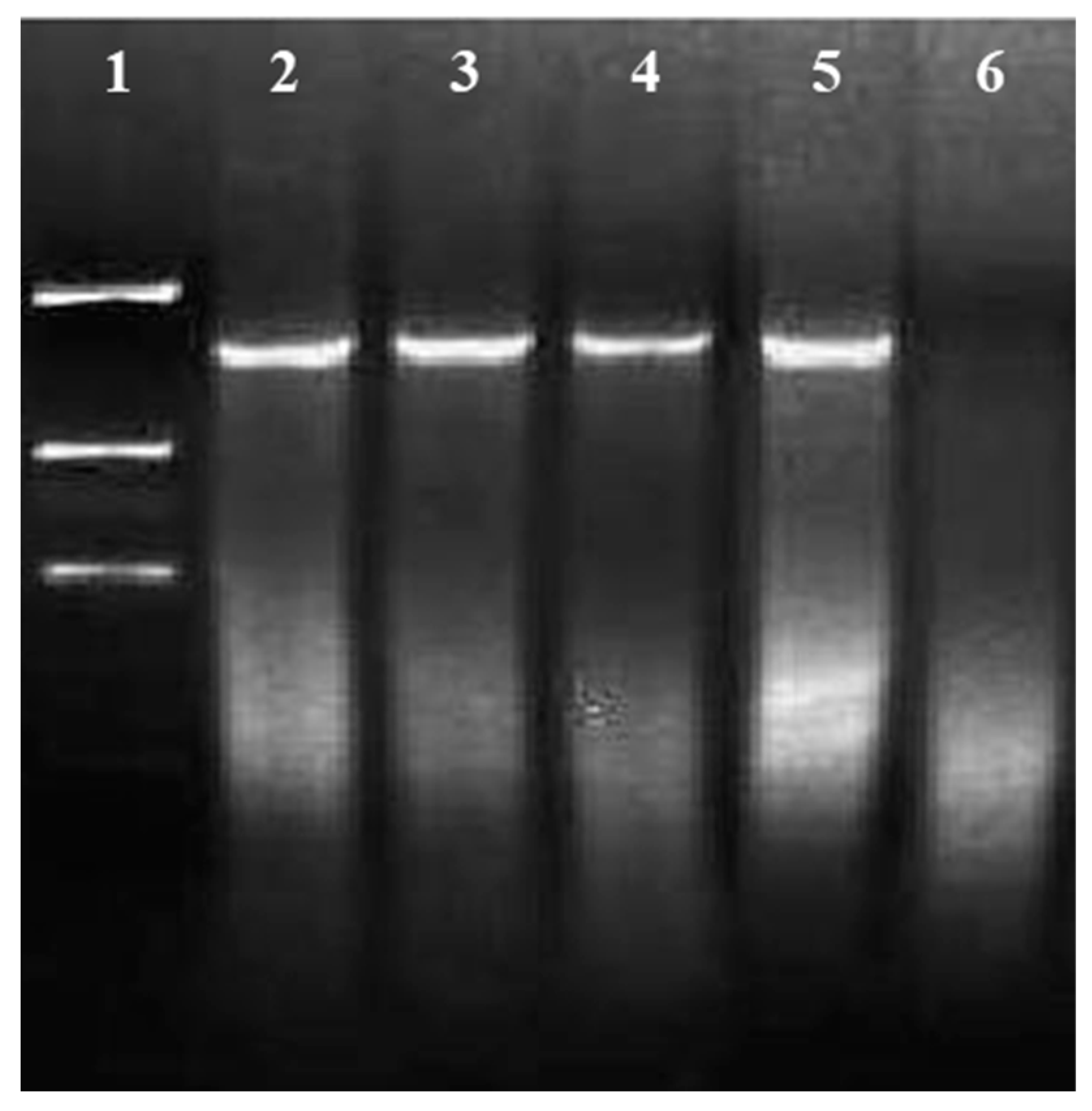
Figure 6.
Genomic DNA of coffee plants transformed with vector pC2301 and digested with enzymes BsteII and BglII. Lane 1, molecular weight marker λ Hind III; lanes 2, 3, 16 and 17, vector pC2301; lanes 4–14, 11 coffee plants transformed with pC2301; lane 15, negative control corresponding to untransformed coffee plant.
Figure 6.
Genomic DNA of coffee plants transformed with vector pC2301 and digested with enzymes BsteII and BglII. Lane 1, molecular weight marker λ Hind III; lanes 2, 3, 16 and 17, vector pC2301; lanes 4–14, 11 coffee plants transformed with pC2301; lane 15, negative control corresponding to untransformed coffee plant.

Figure 7.
Southern blot analysis of coffee plant DNA transformed with vector pC2301 and digested with enzymes Bste II and Bgl II. Lane 1, molecular weight marker λ Hind III; lanes 2, 3, 16, and 17, vector pC2301; lanes 4–14, 11 coffee plants transformed with pC2301; lane 15, negative control corresponding to untransformed coffee plant.
Figure 7.
Southern blot analysis of coffee plant DNA transformed with vector pC2301 and digested with enzymes Bste II and Bgl II. Lane 1, molecular weight marker λ Hind III; lanes 2, 3, 16, and 17, vector pC2301; lanes 4–14, 11 coffee plants transformed with pC2301; lane 15, negative control corresponding to untransformed coffee plant.
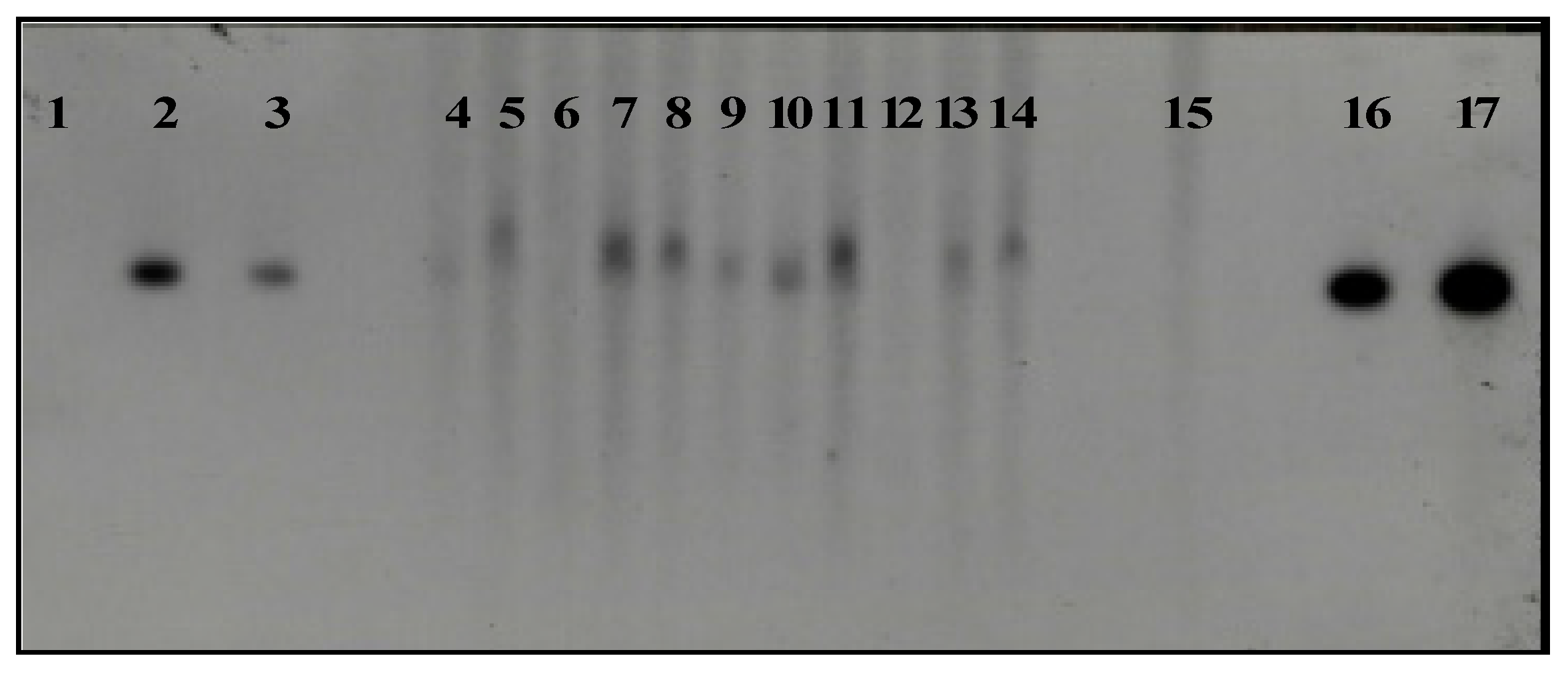
Figure 8.
Genomic DNA of coffee plants transformed with vector pC1301 and digested with enzyme Xho I. Lane 1, molecular weight marker λ Hind III; lanes 2, 3, 16, and 17, vector pC1301; lanes 4–14, 11 coffee plants transformed with vector pC1301; lane 15, negative control corresponding to untransformed coffee plant.
Figure 8.
Genomic DNA of coffee plants transformed with vector pC1301 and digested with enzyme Xho I. Lane 1, molecular weight marker λ Hind III; lanes 2, 3, 16, and 17, vector pC1301; lanes 4–14, 11 coffee plants transformed with vector pC1301; lane 15, negative control corresponding to untransformed coffee plant.

Figure 9.
Southern blot analysis of coffee plants genomic DNA transformed with vector pC1301 and digested with enzyme Xho I. Lane 1, molecular weight marker λ Hind III; lanes 2, 3, 16, and 17, vector pC1301; lanes 4–14, 11 coffee plants transformed with vector pC1301; lane 15, negative control corresponding to untransformed coffee plant.
Figure 9.
Southern blot analysis of coffee plants genomic DNA transformed with vector pC1301 and digested with enzyme Xho I. Lane 1, molecular weight marker λ Hind III; lanes 2, 3, 16, and 17, vector pC1301; lanes 4–14, 11 coffee plants transformed with vector pC1301; lane 15, negative control corresponding to untransformed coffee plant.
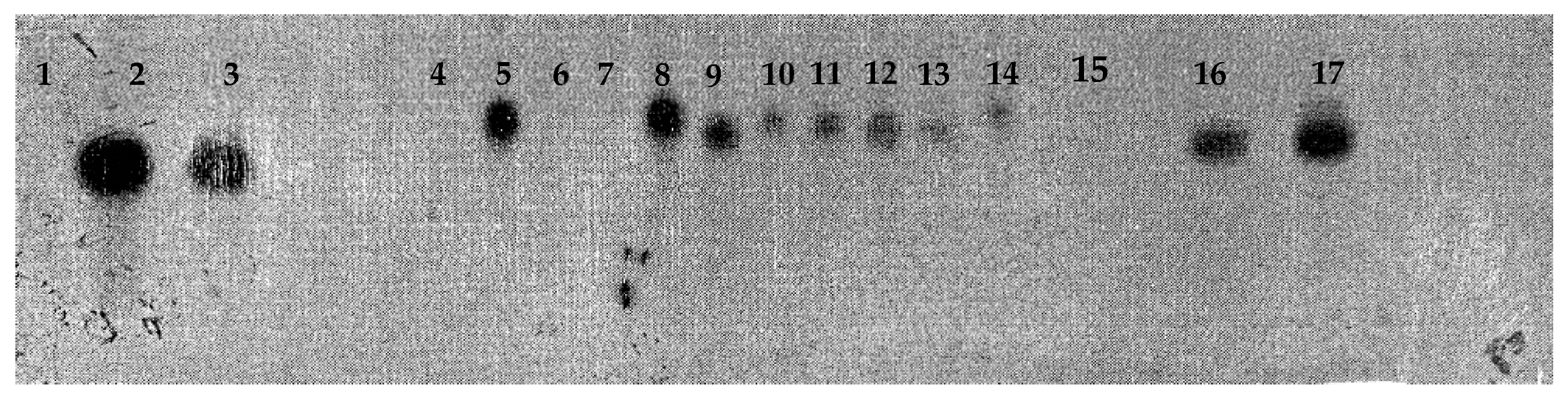

Table 1.
Average transient expression of the uidA gene according to the age of ET, Agrobacterium tumefaciens strain, co-culturing medium, transformation vector and genotype.
Table 1.
Average transient expression of the uidA gene according to the age of ET, Agrobacterium tumefaciens strain, co-culturing medium, transformation vector and genotype.
| Factor | Average transient expression of the uidA gene |
|---|---|
| Age of embryogenic tissue | |
| - Early - Differentiated |
229.535 ± 9.626 a 65.769 ± 5.340 b |
| A. tumefaciens strain | |
| - LBA4405 | 264.877 ± 12.176 a |
| - EHA105 | 99.759 ± 4.270 b |
| Co-culturing medium | |
| - Solid | 167.526 ± 80.923 a |
| - Liquid | 62.434 ± 54.765 b |
| Transformation vector | |
| - pC1301 | 164.199 ± 8.894 a |
| - pC2301 | 146.344 ± 9.150 b |
| Genotype | |
| - BK.620 | 172.437 ± 87.017 a |
| - BI.625 | 182.955 ± 87.810 a |
* Means with the same letter are not statistically different (Duncan = 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



