Submitted:
23 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
We recently identified that cerebral mRNA expression for inducible costimulator (ICOS) and its ligans, ICOSL, both significantly increases during elimination of Toxoplasma gondii cysts from the brains of infected mice by perforin-mediated cytotoxic activity of CD8+ T cells. In the present study, we examined the role of ICOS in activating the effector activity of CD8+ T cells in response to the cysts in infected mice. Following adoptive transfer of splenic CD8+ T cells from chronically infected ICOS-deficient (ICOS-/-) and wild-type (WT) mice to infected SCID mice, fewer CD8+ T cells were detected in the brains of the recipients of ICOS-/- than WT CD8+ T cells. However, such lower numbers of ICOS-/- CD8+ T cells, which migrated into the brains of the recipients, eliminated T. gondii cysts more efficiently than WT CD8+ T cells did. Consistently, the ICOS-/- CD8+ T cells secreted greater amounts of granzyme B in response to T. gondii antigens in vitro than did WT CD8+T cells. We identified that ICOS-/- CD8+ T cells express significantly greater levels of CD28 on their surface than WT CD8+ T cells, and relative expression levels of CD28 mRNA in ratios to CD8b mRNA levels in the brains of the recipients of those CD8+ T cells strongly correlated with their relative expression levels of mRNA for T-bet transcription factor and perforin. Furthermore, blocking of CD28 signaling by a combination of anti-CD80 and anti-CD86 antibodies abolished the increased cytotoxic activity of the ICOS-/- CD8+ T cells in vitro. The present study uncovered notable compensatory interactions between ICOS and CD28 to secure the cytotoxic effector activity of CD8+ T cells against a microbial infection in a murine model of chronic infection with T. gondii.
Keywords:
ICOS
; CD28
; costimulatory molecule
; CD8+ T cell
; cytotoxic activity
; host defense
; infection
; Toxoplasma gondii
1. Introduction
Toxoplasma gondii is an obligate intracellular protozoan parasite that can infect humans and all other mammals [1, 2}. This parasite forms tissue cysts in various organs especially in the brain and establishes a long-lasting chronic infection in these hosts [1,2]. Since the tissue cysts can persist in immunocompetent hosts for long periods of time during the chronic stage of infection, it was considered that the immune system is unable to detect or attack the tissue cysts of this parasite. However, our recent studies uncovered that CD8+ T cells have the capability to detect the host cells harboring T. gondii cysts and destroy them through their perforin-mediated effector activity [3,4]. Furthermore, we identified that CD8+ T cells penetrate into the cysts using their perforin-mediated activity and induce morphological deterioration and destruction of the cysts, which is followed by an accumulation of large numbers of phagocytes [4]. When mRNA levels for the immunity-related 734 molecules were compared in the brains of chronically infected SCID mice that had received CD8+ T cells from infected wild-type (WT) and perforin-deficient (Prf1-/-) mice, mRNA levels for only 6 molecules were identified to be significantly greater in the brains of the recipients of the WT CD8+ T cells than those of Prf1-/- CD8+ T cells [5]. These six molecules were two T cell costimulatory molecules (inducible costimulator [ICOS] and its ligand [ICOSL]); two chemokine receptors (C-X-C motif chemokine receptor 3 [CXCR3] and CXCR6); and two molecules related to an activation of microglia and macrophages (interleukin 18 receptor 1 [IL-18R1] and chitinase-like 3) [5].
The ICOS is a prominent costimulatory molecule that belongs to CD28 receptor family for supporting the activities of both CD4+ and CD8+ T cells. However, whereas CD28 is expressed on most T cells including naïve T cells, ICOS expression on CD8+ T cells is induced only after their activation through their T cell receptor engagement with target antigens presented by the MHC class I molecules [6,7]. In several bacterial and viral infections, blocking or deficiency of ICOS resulted in reduced numbers of the pathogen-specific [8,9] or IFN-γ+ [10,11] CD8+ T cells. Reduced IFN-γ production and cytotoxic activity of CD8+ T cells were also observed in ICOS-/- mice infected with Salmonella enterica serovar Typhimurium [12]. In contrast, in persistent infection with Plasmodium chabaude chabaude AS, an intracellular protozoan parasite that proliferates within red blood cells, ICOS-/- mice displayed increased numbers of IFN-γ+ CD8+ T cells [13]. Therefore, It is important to determine the roles of ICOS specifically on the cytotoxic effector activity of CD8+ T cells against T. gondii cysts during chronic infection with this intracellular protozoan parasite.
In the present study, we examined the roles of ICOS on the effector activity of CD8+ T cells against T. gondii cysts by transferring CD8+ immune T cells from infected wild-type (WT) and ICOS-deficient (ICOS-/-) mice into infected SCID mice lacking T cells to determine the effects of ICOS deficiency on the effector function of those CD8+ T cells to remove the tissue cysts from the brains of the recipient SCID mice. Unexpectedly, we found that ICOS-/- CD8+ T cells eliminated T. gondii cysts from the brains of the recipients more efficiently than WT CD8+ T cells did, whereas fewer ICOS-/- T cells migrated into the brains of the recipients than the WT T cells. We identified that the ICOS-/- CD8+ T cells expressed greater levels of CD28 on their surface than WT CD8+ T cells did, and a blocking of CD28 signaling pathway with a combination of anti-CD80 and anti-CD86 antibodies abolished increased secretion of GzmB by ICOS-/- CD8+ T cells in response to T. gondii antigens in vitro, indicating that the upregulated expression of CD28 compensated the absence of ICOS and maintained the cytotoxic effector activity of CD8+ T cells against T. gondii during chronic infection.
2. Materials and Methods
2.1. Mice
Female WT BALB/c, BALB/c-background ICOS-/-, and BALB/c-background SCID mice were from the Jackson Laboratory (Bar Harbor, ME). Outbred Swiss Webster mice were from Taconic (Germantown, NY). The studies were performed in accordance with approved protocols from the Institutional Animal Care and Use Committee of the University of Kentucky. There were 3 or 4 mice in each experimental group in each experiment.
2.2. Infection with T. gondii
Cysts of T. gondii were obtained from the brains of chronically infected Swiss Webster mice [14,15]. WT and ICOS-/- mice were infected orally with 10 cysts by gavage and treated with sulfadiazine in drinking water (400 mg/L) beginning at 7 days after infection for 10 days to assist controlling tachyzoite proliferation during the acute stage of infection and establish a chronic infection [3,16]. SCID mice were infected orally with 10 cysts by gavage and treated with sulfadiazine the same manner beginning at 9 days after infection for the entire period of the experiments to maintain a chronic infection in their brains [14,15].
2.3. Purification of CD8+ T Cells from Infected WT and ICOS-/- Mice and Adoptive Transfer of those T Cells into Infected SCID Mice
Spleen cells were obtained from ICOS-/- and WT mice infected with T. gondii for at least 2 months and suspended in Hank’s balanced salt solution (HBSS) (HyClone [Cytiva], Mariborough, MA) with 2 % heat-inactivated fetal bovine serum (FBS) (Millipore-Sigma, Burlington, MA). The spleen cells from four mice were pooled within the same experimental group, and CD8+ T cells were purified from the pooled spleen cell suspensions using magnetic beads-conjugated anti-mouse CD8α (clone 53-6.7) monoclonal antibodies (mAbs) (Miltenyi Biotech, Auburn, CA) and MACS column (Miltenyi) [14,17]. The purified CD8+ T cells were suspended in the 2% FBS-HBSS and injected intravenously from a tail vein into infected, sulfadiazine-treated SCID mice (2 x 106 cells/mouse) at 3 weeks after infection. Seven days later, the brain of each of the recipient mice was cut into halves, and one half was immediately frozen with dry ice for RNA purification and another half was fixed in a solution containing 10% formalin, 5% acetic acid, and 70% ethanol for immunohistochemical analyses. Two independent experiments were performed in the CD8+ T cell transfer study, which provided a total 7-8 mice in each experimental group.
2.4. RNA Purification and RT-PCR
RNA was purified from a half of the brain of each mouse using RNA STAT-60 (Tel-test, Friendswood, TX) and treated with DNase I (Invitrogen, Waltham, MA) to remove genomic DNA contamination as described previously [3,18]. cDNA was synthesized from 1 or 4 μg of the DNase I-treated RNA from each brain sample. Quantitative PCR reactions were performed with the cDNA using StepOnePlus real-time PCR system with Taqman reagents (Applied Biosystems, Branchburg, NJ) [14,17]. Primers and probes for mouse β-actin (a house-keeping control molecule), CD8β, perforin, GzmB, CD28, 4-1BB (TNFRSF9), and T-bet were ready-made products from Applied Biosystems. Primers and probe for bradyzoite (cyst)-specific BAG1 are as follows: 5’-TCACGTGGAGACCCAGAGT-3’ (forward), 5’-CTGGCAAGTCAGCCAAAATAATCAT-3’ (reverse), and 5’-TTTGCTGTCGAACTCC-3’ (probe) [18]. Amounts of mRNA levels for the targets of interest were normalized to amounts of mRNA for β-actin.
2.5. Immunohistochemistry
The fixed brains were embedded in paraffin, and sagittal sections (4 μm thickness) of the paraffin-embedded brains were stained for T. gondii or a combination of T. gondii and CD8+ cells. For the T. gondii staining, the sections were stained with rabbit polyclonal anti-T. gondii antibodies as previously described [18,19]. Dual staining for T. gondii and CD8α were performed using Ventana Discovery Ultra instrument (Roche Diagnostics, Indianapolis, IN). After deparaffinization and antigen retrieval with Ventana CC1 (Roche), the slides were incubated with rabbit polyclonal anti-T. gondii antibody at 1:1000 dilution, followed by alkaline phosphatase linked anti-rabbit IgG secondary antibody (Roche) and visualization with Discovery Red Chromogen (Roche). Residual antibody was denatured by heating with CC2 antigen retrieval buffer, prior to incubation with anti-CD8α antibody at 1:250 dilution (Cell Signaling Technology, Danvers, MA) at 37°C for 1 hour, followed by incubation with Ventana anti-rabbit-HQ (Roche) for 20 minutes, and Ventana anti-HQ-HRP (Roche). The staining was then amplified using Ventana’s Discovery TSA Amplification Kit (Roche) for 16 minutes, followed by linking with Discovery Amplification Multimer-HRP (Roche) for 20 minutes and DAB detection. Slides were counterstained with Meyer’s hematoxylin, blued, and permanently mounted. Cyst numbers in the entire field of each of the sagittal sections of the brains were microscopically counted. Three sections with 16 or 20 μm distance between the sections were counted for each brain and the mean value from the counts from the three sections was used for each mouse. CD8+ T cells in a total of 10 randomly selected fields at X200 magnification in a sagittal section of each brain were microscopically counted.
2.6. Culture of CD8+ T Cells Purified from Infected WT and ICOS-/- Mice with T. gondii Antigens
After purifying CD8+ T cells from the spleens of chronically infected ICOS-/- and WT mice using the anti-CD8-mAb-coated microbeads, the remaining CD8- T cell-depleted spleen cells were suspended in RPMI1640 medium (Gibco/Millipore Sigma, St. Louis, MO) containing 10% FBS (HyClone [Cytiva]) and 100 U/ml penicillin and 100 μg/ml streptomycin (Invitrogen/ThermoFisher, Carlsbad, CA) and cultured (5 × 105 cells/well) in a flat-bottom 96-well tissue culture plate (Costar, Corning, Lowell, MA) for 1.5 - 2 hrs. After the incubation, plastic non-adherent cells were removed by washing to prepare antigen-presenting cells (plastic adherent cells). Thereafter, CD8+ T cells purified from the spleens of those infected WT and ICOS-/- mice were placed to those wells (3 × 105 cells/well) containing the plastic adherent antigen-presenting cells from the corresponding strain of mice and cultured in the presence or absence of T. gondii tachyzoite lysate antigens (10 μg/ml) for 72 hrs [14,20]. In one experiment, blocking mAb against ICOSL (clone HK5.3, BioLegend, San Diego, CA), mAbs against CD80 and CD86 (clones 16-10A1 and PO3.1, respectively, Invitrogen/Thermo-Fisher), or a combination of both of these mAbs were added at 10 μg/ml to a part of these culture wells to block the ICOS-ICOSL or CD28-CD80/CD86, or both of these costimulatory pathways. As a control, isotype control mAbs were added in the same manner. There were 5 wells in each experimental group. The concentrations of GzmB in the culture supernatants were measured by ELISA using a commercial kit from R&D Biosystems (Minneapolis, MN) [21] by following their commercial inserts.
2.7. Flow Cytometry
CD8+ T cells purified from infected ICOS-/- and WT mice were incubated with anti-Fcγ receptor monoclonal antibody (mAb) to block antigen-nonspecific binding of mAbs to these cells, followed by incubation with FITC-labeled anti-CD8α and/or PE-labeled anti-CD28 for 30 min. As a control, these CD8+ T cells were incubated with FITC- or PE-labeled isotype control mAbs in the same manner. In a separate experiment, the purified CD8+ T cells were stained with FITC-labeled anti-CD8α, PE-labeled CD28, APC-labeled CD44, and APC-Cy7-labeled anti-CD62L mAbs. All of those antibodies were from BD Biosciences. There were duplicated or triplicated tubes for each staining. The cells were analyzed with BD Symphony A3 using DIVA software (BD Biosciences, San Jose, CA).
2.8. Statistical Analysis
Levels of significance in differences between experimental groups were determined by Student’s t test or Mann-Whitney U test using GraphPad Prism software 9.0. Levels of significance in correlations between two elements were determined by Pearson or Spearman test using the same software. Differences that had p values <0.05 were considered significant.
3. Results
3.1. CD8+ Immune T Cells from ICOS-/- Mice Chronically Infected with T. gondii have Increased Efficiency in Eliminating Tissue Cysts of the Parasite
To examine whether CD8+ T cells maintain their anti-cyst effector capability in the absence of ICOS during chronic infection with T. gondii, CD8+ T cells purified from the spleens of the infected WT and ICOS-/- mice were transferred to chronically infected (infected and treated with sulfadiazine) SCID mice. As a control, a part of the infected SCID mice did not receive any T cells. High levels of bradyzoite (cyst)-specific BAG1 mRNA levels were detected in the brains of the control SCID mice with no T cell transfer at both the day of the transfer of CD8+ T cells (Day 0) and seven days after the T cell transfer (Day 7), and their mRNA levels did not differ between these two time points (Figure 1A), indicating that the cyst burdens in the brains of these control mice were stable between these two time points. In contrast, seven days after the T cell transfer, BAG1 mRNA levels in the brains of the recipients of CD8+ immune T cells from the WT and ICOS-/- mice were both more than 20 times less than those of the control mice with no T cell transfer (P<0.001, Figure 1A). In addition, the cerebral BAG1 mRNA levels in the recipients of ICOS-/- CD8+ T cells tended to be slightly lower than those in the recipients of WT CD8+ T cells (1.10 ± 0.49 vs. 1.47 ± 0.40 [x 10-4] in BAG1 mRNA to β-actin ratios, Figure 1A), but this difference did not reach statistical significance.
We also examined the reduction of the cyst burdens in the recipients of the WT and ICOS-/- CD8+ T cells using the immunohistochemical staining for T. gondii on their brains. Consistent with the BAG1 mRNA levels in these mice, cyst numbers detected in the sagittal sections (three sections with 16-20 μm distance between sections) of the brains of these two groups of the CD8+ T cell recipients were markedly fewer than those in the brains of the control mice with no T cell transfer (P<0.05, Figure 1B). Notably, the cyst numbers in the recipients of the ICOS-/- CD8+ T cells were significantly lower than those of the WT CD8+ T cell recipients (P<0.01, Figure 1B).
To further address the capability of ICOS-/- CD8+ T cells to eliminate T. gondii cysts, we performed immunohistochemical analysis to compare numbers of CD8+ T cells detected in the brains of the recipients of WT and ICOS-/- CD8+ T cells. We counted numbers of CD8+ T cells detected in randomly selected 10 microscopic fields at x200 magnification in a sagittal section of the brain of each of these recipient mice. Numbers of CD8+ T cells in the recipients of ICOS-/- CD8+ T cells were markedly less than those in the recipients of WT CD8+ T cells (P<0.001, Figure 1C). A representative image of the CD8+ T cells detected in the immunohistologically stained sections of the brains of these two groups of mice are shown in Figure 1D,E.
An accumulation of CD8+ T cells to a T. gondii cyst detected in the brain of a recipient of WT CD8+ T cells is also shown in Figure 1F. These results indicate that whereas fewer ICOS-/- CD8+ T cells were present in the brains of the recipient SCID mice than the recipients of WT CD8+ T cells at 7 days after their transfer to the recipients, the former reduced the cerebral cyst burdens in the recipients as efficiently as the latter or even more efficiently than the latter during 7 days after their transfer to the recipient mice.
Consistent with the detection of fewer CD8+ T cells in the recipients of ICOS-/- CD8+ T cells than those of WT CD8+ T cells, amounts of CD8β mRNA in the brains of the former were a little less than a half of the latter (P<0.05, Figure 1G). To further depict the efficiency of WT and ICOS-/- CD8+ T cells that migrated into the brains of the recipient SCID mice in eliminating T. gondii cysts from their brains, we calculated the ratios of BAG1 mRNA level reduction (differences between the mean value of BAG1 mRNA levels in the control mice with no T cell transfer and BAG1 mRNA levels in each of the recipients of WT or ICOS-/- CD8+ T cells) to amounts of CD8β mRNA in the brain of each recipient mouse. The cyst removal efficiency ratios in the brains of the ICOS-/- CD8+ T cell recipients were 3.7 times greater than those of the WT CD8+ T cell recipients (P<0.01, Figure 1H).
In regard to the presence of lower numbers of ICOS-/- CD8+ T cells than WT CD8+ T cells in the recipient SCID mice, previous studies by others (22) and us (23) identified that CD8+ T cells infiltrated into the brains of mice chronically infected with T. gondii are CD44highCD62Llow effector memory phenotype. Therefore, we examined whether the frequencies of the effector memory population in the spleens of CD8+ T cells differ between infected ICOS-/- and WT mice. Notably, the frequencies of the CD44highCD62Llow effector memory population in the total splenic CD8+ T cell populations in infected ICOS-/- mice were approximately 40% less than those on infected WT mice (13.1 ± 2.04 % vs. 20.7 ± 0.53 %, P<0.01, Figure 1I). This difference is consistent with the difference in the levels of CD8β mRNA in the brains of infected SCID mice that received splenic CD8+ T cells from the ICOS-/- and WT mice shown in Figure 1G. Therefore, the low frequency of CD44highCD62Llow effector memory population in CD8+ T cells in the spleens of infected ICOS-/- mice would most likely a key factor that contributed to the presence of lower numbers of CD8+ T cells in the brains of the SCID mice that had received the CD8+ T cells from the ICOS-/- mice than in the recipients of WT CD8+ T cells.
3.2. ICOS-/- CD8+ T Cells Express Greater Levels of mRNA for Perforin and T-bet than WT CD8+ T Cells during Elimination of T. gondii Cysts in the Brain
Since perforin is required for anti-cyst effector activity of CD8+ T cells [3,4], we compared expression levels of mRNA for perforin by the ICOS-/- and WT CD8+ T cells in the brains of infected SCID mice that had received those T cells. For this purpose, we calculated the ratios of mRNA levels for perforin to mRNA levels for CD8β in the brains of those SCID mice at Day 7 after the T cell transfer. The ratios of perforin mRNA levels/CD8β mRNA levels were twice greater in the recipients of ICOS-/- CD8+ T cells than the recipients of WT CD8+ T cells (P<0.05, Figure 2A). Since GzmB is another key effector molecule in the cytotoxic activity of CD8+ T cells, we also compared the ratios of mRNA levels for GzmB to mRNA levels for CD8β in the brains of the recipients of the ICOS-/- and WT CD8+ T cells. The ratios of GzmB mRNA levels to CD8β mRNA levels tended to be slightly greater in the recipients of ICOS-/- CD8+ T cells than the recipients of WT CD8+ T cells, but the difference did not reach statistical significance (Figure 2B). Since the transcription factor T-bet plays critical roles for the cytotoxic activities of CD8+ T cells [24,25,26], we also compared the ratios of T-bet mRNA levels to CD8β mRNA levels in the recipients of the ICOS-/- and WT CD8+ T cells. These ratios were markedly greater in the brains of the recipients of ICOS-/- CD8+ T cells than those of the recipients of WT CD8+ T cells (P<0.05, Figure 2C).
3.3. Increased CD28 mRNA Levels in ICOS-/- CD8+ T Cells Correlate with Increased Levels of mRNA for T-bet Transcription Factor for their Cytotoxic Activity during Elimination of T. gondii Cysts in the Brain
In addition to ICOS, CD28 and 4-1BB expressed on the surface of CD8+ T cells play important roles as costimulatory molecules for their activation with their target antigens [27]. Since CD8+ T cells of infected ICOS-/- mice were found to have a significantly increased anti-cyst effector activity and perforin mRNA expression, there could be a possibility that CD8+ T cells of the infected ICOS-/- mice have increased expression levels of either 4-1BB or CD28, or both, to compensate the absence of ICOS. To address this possibility, the ratios of mRNA levels for 4-1BB (TNFRSF9) and CD28 to CD8β mRNA levels were compared between the brains of the recipients of the ICOS-/- and WT CD8+ T cells. Whereas the ratios of 4-1BB (TNFRSF9) mRNA levels to CD8β mRNA levels tended to be greater in the brains of the recipients of ICOS-/- CD8+ T cells than the recipients of WT CD8+ T cells, the difference did not reach statistical significance (Figure 3A). In contrast, the ratios of CD28 mRNA levels to CD8β mRNA levels were 3.7 times greater in the brains of the recipients of the ICOS-/- CD8+ T cells than the recipients of the WT CD8+ T cells (P<0.05, Figure 3B).
The transcription factor T-bet plays critical roles for the cytotoxic activities of CD8+ T cells [24,25,26] as mentioned earlier. To address the possibility that costimulatory signal mediated by the upregulated expression of CD28 in ICOS-/- CD8+ T cells contributes to the enhanced their cytotoxic effector activity to remove T. gondii cysts, we examined whether the increases in the ratios of CD28 mRNA levels to CD8β mRNA levels directly correlate with the increases in the ratios of T-bet mRNA levels to CD8β mRNA levels in the brains of the recipients of ICOS-/- and WT CD8+ T cells. Notably, the degrees of increases in the CD28 mRNA/CD8β mRNA ratios strongly correlated with the degrees of increases in the T-bet/CD8β mRNA ratios in the brains of these recipient mice (P<0.0001, Figure 3C).
To support the possibility that the increased expression of transcription factor T-bet contributed to upregulation of cytotoxic activity of ICOS-/- CD8+ T cells, we examined whether the increases in the ratios of T-bet mRNA levels to CD8β mRNA levels directly correlate with increases in the ratios of perforin mRNA levels to CD8β mRNA levels in the brains of recipients of ICOS-/- and WT CD8+ T cells. The degrees of increases in the ratios of T-bet mRNA levels/CD8β mRNA levels strongly correlated with the degrees of increases in the ratios of perforin mRNA levels/ CD8β mRNA levels in the brains of those T cell recipients (P=0.0009, Figure 3D). There results together suggest that the costimulatory signal provided by upregulated expression of CD28 in ICOS-/- CD8+ T cells contributes to their enhanced expression of transcription factor T-bet, and thereby induces upregulation of their perforin expression, which facilitates elimination of T. gondii cysts by these T cells in the brains of infected SCID mice that had received these T cells.
3.4. Surface Expression of CD28 Costimulatory Factor is Increased in ICOS-/- CD8+ T Cells during Chronic Infection with T. gondii
CD28 provides the costimulatory signal when this molecule expressed on the surface of CD8+ T cells interacts with their ligands, CD80 and CD86, expressed on the surface of antigen-presenting cells that present target antigens for the T cells. Based on the greater mRNA expression for CD28 in the ICOS-/- CD8+ T cells than WT CD8+ T cells in the brains of recipients of these T cells shown in Figure 3B, we performed flow cytometric analyses to examine whether expression levels of CD28 are increased on the surface of ICOS-/-CD8+ T cells when compared to WT CD8+ T cells during chronic T. gondii infection. Whereas WT CD8+ T cells displayed a single population based on their CD8 and CD28 expression levels (Figure 4A), the ICOS-/- CD8+ T cells showed two distinct populations based on the expression levels of these two molecules (Figure 4B). In addition, the larger population (population 2, composing 80% of ICOS CD8+ T cells) from these two populations in ICOS-/- CD8+ T cells had a significantly higher median fluorescent intensity (MFI) in CD28 expression than did the WT CD8+ T cells (P<0.01, Figs. 4D and 4E), although the MFI in CD28 expression in the smaller population (population 1, composing 20%) of ICOS-/- CD8+ T cells was equivalent to that of WT CD8+ T cells (Figure 4C). These results indicate that a majority of CD8+ T cells in chronically infected ICOS-/- mice have increased expression levels of CD28 costmulatory factor on their surface when compared to those T cells in infected WT mice.
3.5. Blockage of CD28−CD80/CD86 Costimulatory Pathway Abolishes the Increased Cytotoxic Activity of CD8+ T Cells of ICOS-/- Mice Chronically Infected with T. gondii
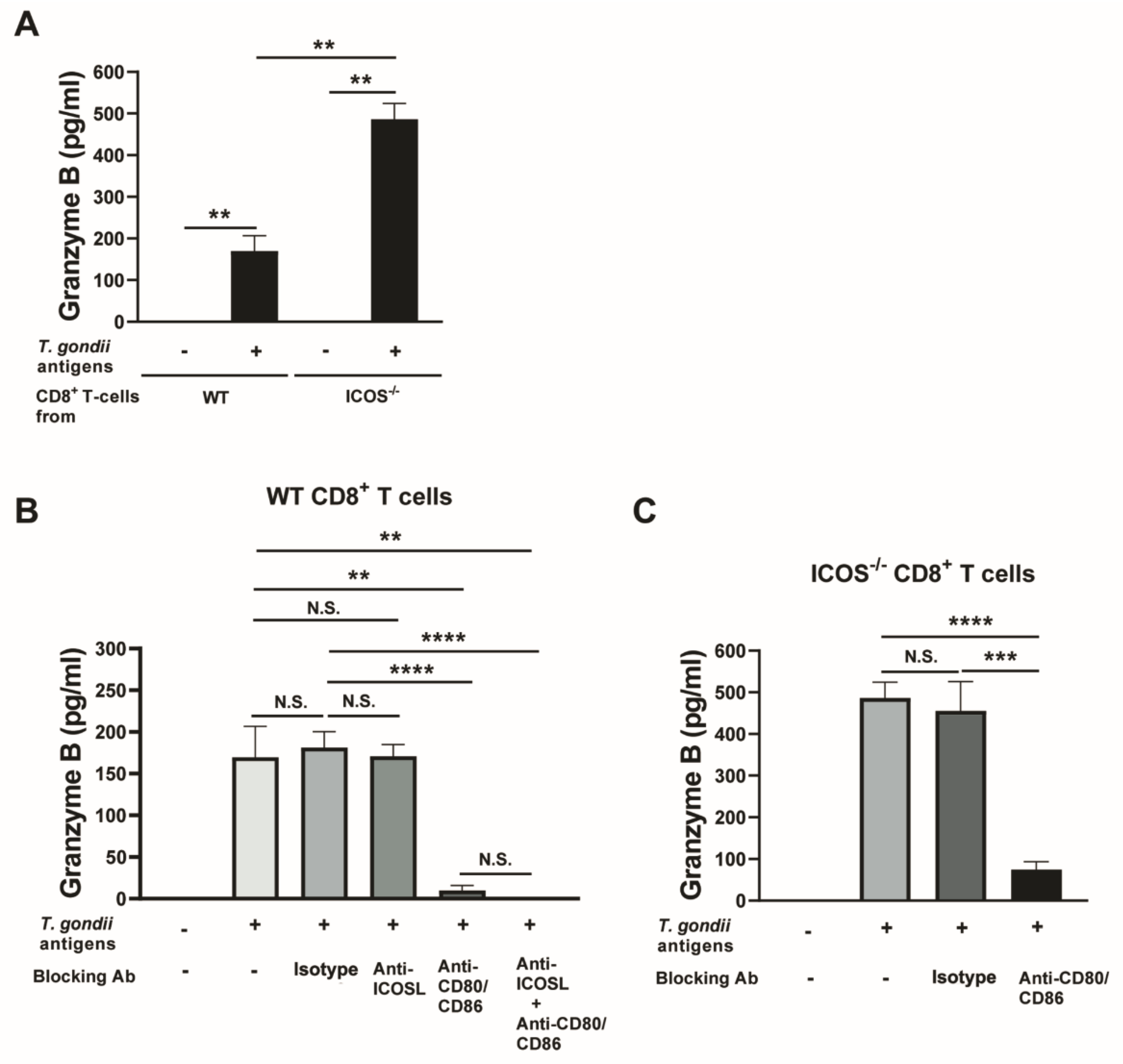
To confirm increased cytotoxic activity of CD8+ T cells from infected ICOS-/- mice against T. gondii when compared to CD8+ T cells from infected WT mice, we compared secretion of GzmB in response to T. gondii antigens between these T cells using in vitro cultures of these T cells, since we were unable to find a reliable ELISA kit to measure perforin levels. After culturing CD8+ T cells purified from the spleens of these mice with antigen-presenting cells (plastic adherent cells) from the corresponding mouse strain in the presence and absence of soluble T. gondii antigens for 72 hrs, the amounts of GzmB in the culture supernatants of the ICOS-/- CD8+ T cells stimulated with the T. gondii antigens were 5 times greater than those detected in the cultures of the WT CD8+ T cells stimulated with those antigens (P<0.01, Figure 5A). In the absence of the T. gondii antigens in the cultures, amounts of GzmB in the culture supernatants of both WT and ICOS-/- CD8+ T cells remained very low and close to the detection limit of the ELISA assay (Figure 5A).
Binding of CD28 expressed on the surface of CD8+ T cells to CD80 and CD86 expressed on the antigen-presenting cells that present their target antigens provides the costimulatory signal required for activating those T cells as mentioned earlier. To obtain a direct evidence on the contribution of the upregulated CD28 expression in the CD8+ T cells of chronically infected ICOS-/- mice to their upregulated cytotoxic activity against the parasite, we cultured CD8+ T cells from infected ICOS-/- and WT mice with antigen-presenting cells and T. gondii antigens in the presence and absence of blocking mAbs against CD80 and CD86. We also added anti-ICOSL mAb in a part of the cultures of WT CD8+ T cells to block ICOS-ICOSL costimulation signal.
In the cultures of WT CD8+ T cells, blocking of CD28-CD80/CD86 costimulatory signaling by anti-CD80 and anti-CD86 mAbs markedly reduced the levels of GzmB in their culture supernatants (P<0.0001, Figure 5B), whereas an addition of anti-ICOSL mAb to their cultures did not affect the GzmB levels in their culture supernatants (Figure 5B). When both the ICOS and CD28 costimulatory signaling pathways were blocked by anti-ICOSL mAb in combination with anti-CD80 and anti-CD86 mAbs, levels of GzmB on the culture supernatants of WT CD8+ T cells stimulated with T. gondii antigens were as low as those of cultures of those T cells without the stimulation with the parasite antigens (P<0.0001, Figure 5B). When comparing between the cultures with blocking of only CD28 costimulatory pathway and the cultures with blocking of both CD28 and ICOS costimulatory pathways, GzmB levels in the former tended to be higher than the latter, but this difference did not reach statistical significance (Figure 5B). These results indicate that the presence of CD28 costimulatory signaling without ICOS costimulatory signaling can provide the sufficient costimulatory signaling required for activation of the cytotoxic activity of WT CD8+ immune T cells in response to their target T. gondii antigens, although the absence of both CD28 and ICOS costimulatory signaling is required for completely blocking the cytotoxic activity of CD8+ T cells from chronically infected WT mice in response to T. gondii antigens.
In the cultures of ICOS-/- CD8+ T cells, blocking of CD28 costimulatory signaling by anti-CD80 and anti-CD86 mAbs markedly (6.4 times) reduced the levels of GzmB in their culture supernatants in response to T. gondii antigens (P<0.001, Figure 5C). This is consistent with the observations from WT CD8+ T cells, in which the presence of CD28 costimulation signaling without ICOS costimulation pathway can provide sufficient costimulatory signaling to efficiently activate the cytotoxic activity of the CD8+ T cells in response to T. gondii antigens, whereas the absence of the both costimulatory pathways ablated their cytotoxic activity in response to their target antignes. Thus, these results indicate that the increased cytotoxic activity of ICOS-/- CD8+ T cells against T. gondii antigens is mediated by their increased expression of CD28.
4. Discussion
The present study with adoptive transfer of CD8+ T cells from ICOS-/- and WT mice chronically infected with T. gondii to infected SCID mice revealed that a deficiency of ICOS during chronic infection with this parasite results in increased efficiency of the CD8+ T cells to eliminate tissue cysts of this parasite from the brains of the recipients. Our previous studies identified that anti-cyst activity of CD8+ T cells is operated by their perforin-dependent cytotoxic activity [3,4]. Consistently, the present study also revealed that relative expression levels of perforin mRNA in ratios to CD8β mRNA in the brains of the recipients of ICOS-/- CD8+ T cells are significantly greater than those of the recipients of WT CD8+ T cells, suggesting that ICOS-/- CD8+ T cells express greater levels of perforin mRNA in the brains of the recipient SCID mice than did WT CD8+ T cells. Furthermore, the present study identified that CD8+ T cells from infected ICOS-/- mice secrete much greater amounts of Gzm B in response to T. gondii antigens in vitro than did CD8+ T cells from infected WT mice. GzmB is a key effector molecule, in addition to perforin, in the cytotoxic activity of CD8+ T cells. In relation to our findings, a recent study with persistent infection with Plasmodium chabaude chabaude AS, an intracellular protozoan parasite closely related to T. gondii, demonstrated that infected ICOS-/- mice displayed increased numbers of IFN-γ+ CD8+ T cells when compared to infected WT mice [13]. In contrast, previous studies using infections with viruses [10,28], bacteria [8,11,12] showed that the absence of ICOS costimulatory signaling pathway, either by genetic deletion of ICOS or blocking of its functions by anti-ICOS mAb or ICOS-Ig (a fusion protein of ICOS and the Fc region of human IgG1), downregulates [8,10,11,12] or does not affect [28] the cytotoxic activity and/or IFN-γ production of CD8+ T cells during those microbial infections. The present study provides a new insight that ICOS deficiency induces an upregulation of the cytotoxic activity and the effector function of CD8+ T cells against T. gondii during chronic infection with this protozoan parasite. Therefore, the effects of the absence of ICOS on the functions of CD8+ T cells during microbial infections most likely differ depending of the types of pathogens.
The present study was performed during the chronic stage of T. gondii infection, in which WT and ICOS-/- mice were infected for at least 2 months. T. gondii resides and proliferate within the parasitophorous vacuole (PV) in infected host cells. The PV prevents its fusion with lysosomes and protects the parasite from their elimination [29,30]. Similarly, Mycobacterium tuberculosis, an intracellular bacterium, also resides within phagosomes in infected cells, and prevents their fusion with lysosomes [31,32,33]. In infection with Mycobacterium tuberculosis, the bacterial loads in the spleen did not differ between ICOS-/- and WT mice during the first 40 days of the infection, but the pathogen loads become significantly less in the former than the latter at 60 and 120 days after infection (11). Of interest, the significantly reduced bacterial loads in the ICOS-/- mice during the later time points of the infection are associated with increased numbers of IFN-γ+ CD4+ T cells in the spleens of these mice [11]. Therefore, the effects of ICOS deficiency on CD4+ and CD8+ T cells could differ depending on the time periods that the hosts have been infected with certain pathogens.
There are notable differences in resistance and susceptibility to chronic infection with T. gondii among inbred strains of mice [34,35,36]. Mice with the H-2b (e.g. C57BL/6) and H-2k haplotypes (e.g. C3H/He) are susceptible and develop progressive and ultimately fatal toxoplasmic encephalitis during the later stage of infection, whereas mice with the H-2d haplotype (e.g. BALB/c) are resistant and maintain a latency of the chronic infection in their brains [34,35,36]. The present study was performed in the genetically resistant BALB/c-background mice. A previous study by others [37] using the BALB/c-background ICOS-/- mice showed that whereas percentages of IFN-γ+ cells in CD4+ T cells in the spleens were reduced in ICOS-/- mice when compared to WT mice during the acute stage (day 7) of infection, percentages of IFN-γ+ cells in CD8+ T cells did not differ between ICOS-/- and WT mice in their spleens during the early stage of infection and in their brains during a later stage (weeks 4-6) of infection. In contrast, the present study revealed increased cytotoxic effector activity of CD8+ T cells against T. gondii cysts in BALB/c-background ICOS-/- than WT mice during the chronic stage of infection. Therefore, it may be possible that IFN-γ production and cytotoxic activity of CD8+ T cells are controlled in a different manner through ICOS-mediated pathways.
In contrast to the genetically resistant BALB/c mice, mice in genetically susceptible C57BL/6-background displayed that blocking of ICOS signaling by anti-ICOSL mAb or a genetic deletion of ICOS increases numbers of CD4+ and CD8+ T cells and IFN-γ+ CD8+ T cells in the spleens and brains during 5-6 weeks after infection, but significantly greater numbers of T. gondii cysts were detected in the brains of the infected ICOS-/- than WT mice [38,39]. In the present study with genetically resistant BALB/c-background mice, we identified that numbers of ICOS-/- CD8+ T cells in the brains of the SCID mice that had received those T cells were fewer than those in the brains of the recipients of WT CD8+ T cells, but the former eliminated T. gondii cysts from the brains of the recipients more efficiently than the latter did. It is most likely that the roles of ICOS on the protective activities of CD8+ T cells against T. gondii differ depending of the genetic resistance/susceptibility of the hosts to the infection.
Previous studies using infections with vaccinia virus [40], influenza virus [9], and Listeria monocytogenes [41] demonstrated a requirement of CD28 for optimal recall responses of CD8+ T cells. Notably, the present study using flow cytomotry identified that the absence of ICOS becomes compensated by the upregulation of CD28 expression levels on splenic CD8+ T cells during the chronic stage of T. gondii infection. Consistently, the present study also identified that the ratios of CD28 mRNA levels to CD8β mRNA levels in the brains of infected SCID mice that had received ICOS-/- splenic CD8+ T cells were significantly greater than those in those of the recipients of WT splenic CD8+ T cells. Furthermore, in vitro stimulation of CD8+ T cells purified from the spleens of infected ICOS-/- and WT mice with T. gondii antigens revealed that upregulated CD28 expression mediates the increased cytotoxic effector activity of the ICOS-/- CD8+ T cells in their recall responses to the pathogen.
The transcription factor T-bet plays critical roles for the cytotoxic activities of CD8+ T cells [24,25,26]. The present study identified that the degrees of increases in relative expression levels of mRNA for CD28 in ratios to CD8β mRNA levels strongly correlate with the degrees of increases in ratios of T-bet mRNA levels to CD8β mRNA levels in the brains of the recipients of ICOS-/- and WT CD8+ T cells. In addition, the degrees of increases in the ratios of T-bet mRNA levels/CD8β mRNA levels in the brains of the recipients of those CD8+ T cells strongly correlated with the degrees of increases in the ratios of perforin mRNA levels/ CD8β mRNA levels in those mice. Therefore, enhanced costimulatory signal through the increased expression of CD28 in ICOS-/- CD8+ T cells most likely induced upregulation of perforin mRNA levels through their increased expression of T-bet transcription factor and enhanced the efficiency of elimination of T. gondii cysts through their cytotoxic activity. To our knowledge, an upregulation of CD28 expression on CD8+ T cells in compensation of a deficiency of ICOS and an enhancement of their cytotoxic effector activity through the upregulated CD28 expression have not been reported before.
In relation to our finding on the compensation of the absence of ICOS by an upregulation of CD28 in the recall responses of the cytotoxic activity of CD8+ T cells against T. gondii, a previous study with infections with Lymphocytic choriomeningitis virus and vesicular stomatitis virus demonstrated that blocking of ICOS signaling by ICOS-Ig markedly impaired IFN-γ production of CD4+ T cells against the viruses in CD28-/- mice, whereas ICOS-Ig treatment in WT mice had only a limited downregulatory effect on their IFN-γ production [28]. Thus, it would be possible that not only a compensation of the absence of ICOS signaling by upregulation of CD28 expression but also a compensation of the absence of CD28 by upregulation of ICOS could be operated in maintaining the effector functions of not only CD8+ T cells but also in CD4+ T cells during microbial infections, although the interactions between ICOS and CD28 on regulating the functions of these T cell populations could differ depending on the pathogens as discussed earlier.
The present study provided a novel insight on a notable capability of the immune system to secure the protective activities of CD8+ T cells by utilizing compensatory interactions between two important costimulatory molecules, ICOS and CD28, for host resistance during chronic infection with T. gondii. The results of the present study may also suggest that under the presence of CD28 expression, increased expression of ICOS in WT CD8+ T cells as detected in our recent study [5] could enhances their anti-cyst effector activity to eliminate T. gondii cysts.
5. Conclusions
This work demonstrated that the absence of ICOS during chronic infection with T. gondii induces a significant upregulation of another costimulatory molecule, CD28, but not 4-1BB, on CD8+ T cells, and that the upregulated CD28 expression mediate increased cytotoxic effector activity of the CD8+ T cells to eliminate tissue cysts of the parasite from the brains of chronically infected mice. To our knowledge, an upregulation of CD28 expression on CD8+ T cells in compensation of deficiency in ICOS and an enhancement of the cytotoxic effector activity of the T cells through the upregulated CD28 expression have not been reported before. Thus, the present study revealed notable compensatory interactions between ICOS and CD28 to secure the cytotoxic effector activity of CD8+ T cells against a microbial infection in a murine model of chronic infection with T. gondii.
Author Contributions
Conceptualization, Y.S.; formal analysis, R.M. and, Y.S.; Investigation, R.M., K.E.B, and, Y.S.; data curation, R.M. and, Y.S.; writing, original draft preparation, Y.S.; writing-review, and editing, R.M., K.E.B., and, Y.S.; visualization, R.M. and, Y.S.; supervision, Y.S.; funding acquisition, Y.S.; All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by NIH grant (AI095032) and in part by a bridge funding from the University of Kentucky College of Medicine. The research was also supported by the Biospecimen Procurement and Translational Pathology Shared Resource Facility of the University of Kentucky Markey Cancer Center (P30CA177558).
Institutional Review Board Statement
The studies were performed in accordance with approved protocols from the Institutional Animal Care and Use Committee of the University of Kentucky.
Informed Consent Statement
“Not applicable.”
Data Availability Statement
The data generated from this study are presented in this article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors appreciate assistance in immunohistochemical analysis provided by the Biospecimen Procurement and Translational Pathology Shared Resource Facility of the University of Kentucky Markey Cancer Center.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Toxoplasma gondii . In Medical Microbiology; Baron, S., Ed.; The University of Texas Medical Branch at Galveston: Galveston, TX., 1996. [Google Scholar]
- Suzuki, Y.; Wang, X.; Jortner, B. , Payne, L.; Ni, Y.; Michie, S.A.; Xu, B.; Kudo T.; Perkins, S. Removal of Toxoplasma gondii cysts from the brain by perforin-mediated activity of CD8+ T cells. Am. J. Pathol. 2010, 176, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, A.; Hannah, R.; Lutshumba, J.; Ochiai, E.; Weiss, L.M.; Suzuki, Y. Penetration of CD8+ cytotoxic T cells into large target, tissue cysts of Toxoplasma gondii, leads to its elimination. Am. J. Pathol. 2019, 189, 1584–1607. [Google Scholar] [CrossRef] [PubMed]
- Lutshumba, J.; Ochiai, E.; Sa, Q.; Anand, N.; Suzuki, Y. Selective upregulation of transcripts for six molecules related to T cell costimulation and phagocyte recruitment and activation among 734 immunity-related genes in the brain during perforin-dependent, CD8+ T cell-mediated elimination of Toxoplasma gondii cysts. mSystems 2020, 5, e00189–20. [Google Scholar] [CrossRef] [PubMed]
- Hutloff, A.; Dittrich, A.M.; Beier, K.C.; Eljaschewitsch, B.; Kraft, R.; Anagnostopoulos, I.; Kroczek, R.A. ICOS is an inducible T-cell co-stimulator structurally and functionally related to CD28. Nature 1999, 397, 263–266. [Google Scholar] [CrossRef]
- Yoshinaga, S.K.; Whoriskey, J.S.; Khare, S.D.; Sarmiento, U.; Guo, J.; Horan, T.; Shih, G.; Zhang, M.; Coccia, M.A.; Kohno, T.; Tafuri-Bladt, A.; Brankow, D.; Campbell, P.; Chang, D.; Chiu, L.; Dai, T.; Duncan, G.; Elliott, G.S.; Hui, A.; McCabe, S.M.; Scully, S.; Shahinian, A.; Shaklee, C.L.; Van, G.; Mak, T.W.; Senaldi, G. T-cell co-stimulation through B7RP-1 and ICOS. Nature 1999, 402, 827–832. [Google Scholar] [CrossRef]
- Mittrucker, H.W.; Kursar, M.; Kohler, A.; Yanagihara, D.; Yoshinaga, S.K.; Kaufmann, S.H. Inducible costimulator protein controls the protective T cell response against Listeria monocytogenes. J. Immunol. 2002, 169, 5813–5817. [Google Scholar] [CrossRef]
- Bertram, E.M.; Tafuri, A.; Shahinian, A.; Chan, V.S.; Hunziker, L.; Recher, M.; Ohashi, P.S.; Mak, T.W.; Watts, T.H. Role of ICOS versus CD28 in antiviral immunity. Eur. J. Immunol. 2002, 32, 3376–3385. [Google Scholar] [CrossRef]
- Humphreys, I.R.; Edwards, L.; Snelgrove, R.J.; Rae, A.J.; Coyle, A.J.; Hussell, T. A critical role for ICOS co-stimulation in immune containment of pulmonary influenza virus infection. Eur. J. Immunol. 2006, 36, 2928–2938. [Google Scholar] [CrossRef]
- Nouailles, G.; Day, T.A.; Kuhlmann, S.; Loewe, D.; Dorhoi, A.; Gamradt, P.; Hurwitz, R.; Jorg, S.; Pradl, L.; Hutloff, A.; Koch, M.; Kursar, M.; Kaufmann, S.H. Impact of inducible co-stimulatory molecule (ICOS) on T-cell responses and protection against Mycobacterium tuberculosis infection. Eur. J. Immunol. 2011, 41, 981–991. [Google Scholar] [CrossRef]
- Vidric, M.; Bladt, A.T.; Dianzani, U.; Watts, T.H. Role for inducible costimulator in control of Salmonella enterica serovar Typhimurium infection in mice. Infect. Immun. 2006, 74, 1050–1061. [Google Scholar] [CrossRef] [PubMed]
- Wikenheiser, D.J.; Ghosh, D.; Kennedy, B.; Stumhofer, J.S. The Costimulatory molecule ICOS regulates host Th1 and follicular Th cell differentiation in response to Plasmodium chabaudi chabaudi AS Infection. J. Immunol. 2016, 196, 778–791. [Google Scholar] [CrossRef]
- Sa, Q. Ochiai, E.; Tiwari, A.; Mullins, J.; Shastri, N.; Mercier, C.; Cesbron-Delauw, M.F.; Suzuki, Y. Determination of a key antigen for immunological intervention to target the latent stage of Toxoplasma gondii. J. Immunol. 2017, 198, 4425–4434. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, E.; Sa, Q.; Brogli, M.; Kudo, T.; Wang, X.; Dubey, J.P.; Suzuki, Y. CXCL9 is important for recruiting immune T cells into the brain and inducing an accumulation of the T cells to the areas of tachyzoite proliferation to prevent reactivation of chronic cerebral infection with Toxoplasma gondii. Am. J. Pathol. 2015, 185, 314–324. [Google Scholar] [CrossRef]
- Kang, H.; Suzuki, Y. Requirement of non-T cells that produce gamma interferon for prevention of reactivation of Toxoplasma gondii infection in the brain. Infect. Immun. 2001, 69, 2920–2927. [Google Scholar] [CrossRef]
- Ochiai, E.; Sa, Q.; Perkins, S.; Grigg, M.E.; Suzuk, Y. CD8+ T cells remove cysts of Toxoplasma gondii from the brain mostly by recognizing epitopes commonly expressed by or cross-reactive between type II and type III strains of the parasite. Microbes. Infect. 2016, 18, 517–522. [Google Scholar] [CrossRef]
- Sa, Q.; Ochiai, E.; Sengoku, T.; Wilson, M.E.; Brogli, M.; Crutcher, S.; Michie, S.A.; Xu, B.; Payne, L.; Wang, X.; Suzuki, Y. VCAM-1/alpha4beta1 integrin interaction is crucial for prompt recruitment of immune T cells into the brain during the early stage of reactivation of chronic infection with Toxoplasma gondii to prevent toxoplasmic encephalitis. Infect. Immun. 2014, 82, 2826–2839. [Google Scholar] [CrossRef]
- Wang, X.; Kang, H.; Kikuchi, T.; Suzuki, Y. Gamma interferon production, but not perforin-mediated cytolytic activity, of T cells is required for prevention of toxoplasmic encephalitis in BALB/c mice genetically resistant to the disease. Infect. Immu. 2004, 72, 4432–4438. [Google Scholar] [CrossRef]
- Miller, R.; Wen, X.; Dunford, B.; Wang, X.; Suzuki, Y. Cytokine Production of CD8+ immune T cells but not of CD4+ T cells from Toxoplasma gondii-infected mice is polarized to a type 1 response following stimulation with tachyzoite-infected macrophages. J. Interferon Cytokine Res. 2006, 26, 787–792. [Google Scholar] [CrossRef]
- Mani, R.; Mercier, C.; Delauw, M.F.; Suzuki, Y. 2023, Immunization with the amino-terminus region of dense granule protein 6 (GRA6) of Toxoplasma gondii activates CD8+ cytotoxic T cells capable of removing tissue cysts of the parasite through antigen presentation by human HLA-A2.1. Microbes Infect 2023, 25, 105182. [Google Scholar] [CrossRef]
- Schluter, D.; Meyer, T.; Kwok, L.Y.; Montesinos-Rongen, M.; Lutjen, S.; Strack, A.; Schmitz, M.L.; Deckert, M. Phenotype and regulation of persistent intracerebral T cells in murine Toxoplasma encephalitis. J. Immunol. 2002, 169, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Michie, S.A.; Xu, B.; Suzuki, Y. Importance of IFN-gamma-mediated expression of endothelial VCAM-1 on recruitment of CD8+ T cells into the brain during chronic infection with Toxoplasma gondii. J. Interferon Cytokine Res. 2007, 27, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Moriya, O.; Yoshimoto, T.; Akatsuka, T. T-bet is required for protection against vaccinia virus infection. J. Virol. 2005, 79, 12798–12806. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, B.M.; Juedes, A.; Szabo, S.J.; von Herrath, M.; Glimcher, L.H. Antigen-driven effector CD8 T cell function regulated by T-bet. Proc. Natl. Acad. Sci. U S A 2003, 100, 15818–15823. [Google Scholar] [CrossRef]
- Intlekofer, A.M.; Takemoto, N.; Wherry, E.J.; Longworth, S.A.; Northrup, J.T.; Palanivel, V.R.; Mullen, A.C.; Gasink, C.R.; Kaech, S.M.; Miller, J.D.; Gapin, L.; Ryan, K.; Russ, A.P.; Lindsten, T.; Orange, J.S.; Goldrath, A.W.; Ahmed, R.; Reiner, S.L. Effector and memory CD8+ T cell fate coupled by T-bet and eomesodermin. Nat. Immunol. 2005, 6, 1236–1244. [Google Scholar] [CrossRef]
- Duttagupta, P.A.; Boesteanu, A.C.; Katsikis, P.D. Costimulation signals for memory CD8+ T cells during viral infections. Crit. Rev. Immunol. 2009, 29, 469–486. [Google Scholar] [CrossRef]
- Kopf, M.; Coyle, A.J.; Schmitz, N.; Barner, M.; Oxenius, A.; Gallimore, A.; Gutierrez-Ramos, J.C.; Bachmann, M.F. Inducible costimulator protein (ICOS) controls T helper cell subset polarization after virus and parasite infection. J Exp Med 2000, 192, 53–61. [Google Scholar] [CrossRef]
- Mordue, D.G.; Sibley, L.D. Intracellular fate of vacuoles containing Toxoplasma gondii is determined at the time of formation and depends on the mechanism of entry. J. Immunol. 1997, 159, 4452–4459. [Google Scholar] [CrossRef]
- Mordue, D.G.; Hakansson, S.; Niesman, I.; Sibley, L.D. Toxoplasma gondii resides in a vacuole that avoids fusion with host cell endocytic and exocytic vesicular trafficking pathways. Exp. Parasitol. 1999, 92, 87–99. [Google Scholar] [CrossRef]
- Armstrong, J.A.; Hart, P.D. Phagosome-lysosome interactions in cultured macrophages infected with virulent tubercle bacilli. Reversal of the usual nonfusion pattern and observations on bacterial survival. J. Exp. Med. 1975, 142, 1–16. [Google Scholar] [CrossRef]
- Kaufmann, S.H. How can immunology contribute to the control of tuberculosis? Nat. Rev. Immunol. 2001, 1, 20–30. [Google Scholar] [CrossRef]
- Russell, D.G. 2001, Mycobacterium tuberculosis: Here today, and here tomorrow. Nat. Rev. Mol. Cell. Biol. 2001, 2, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Joh, K.; Orellana, M.A.; Conley, F.K.; Remington, J.S. A gene(s) within the H-2D region determines the development of toxoplasmic encephalitis in mice. Immunology 1991, 74, 732–739. [Google Scholar] [PubMed]
- Suzuki, Y.; Joh, K.; Kwon, O.C.; Yang, Q.; Conley, F.K.; Remington, J.S. MHC class I gene(s) in the D/L region but not the TNF-alpha gene determines development of toxoplasmic encephalitis in mice. J. Immunol. 1994, 153, 4649–4654. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.R.; Hunter, C.A.; Estes, R.G.; Beckmann, E.; Forman, J.; David, C.; Remington, J.S.; McLeod, R. Definitive identification of a gene that confers resistance against Toxoplasma cyst burden and encephalitis. Immunology 1995, 85, 419–428. [Google Scholar]
- Wilson, E.H.; Zaph, C.; Mohrs, M.; Welcher, A.; Siu, J.; Artis, D.; Hunter, C.A. B7RP-1-ICOS interactions are required for optimal infection-induced expansion of CD4+ Th1 and Th2 responses. J Immunol 2006, 177, 2365–2372. [Google Scholar] [CrossRef]
- O'Brien, C.A.; Batista, S.J.; Still, K.M.; Harris, T.H. IL-10 and ICOS differentially regulate T cell responses in the brain during chronic Toxoplasma gondii infection. J. Immunol. 2019, 202, 1755–1766. [Google Scholar] [CrossRef]
- O'Brien, C.A.; Harris, T.H. ICOS-deficient and ICOS YF mutant mice fail to control Toxoplasma gondii infection of the brain. PLoS ONE.
- Fuse, S.; Zhang, W.; Usherwood, E.J. 2008, Control of memory CD8+ T cell differentiation by CD80/CD86-CD28 costimulation and restoration by IL-2 during the recall response. J. Immunol. 2008, 180, 1148–1157. [Google Scholar] [CrossRef]
- Frohlich, M.; Gogishvili, T.; Langenhorst, D.; Luhder, F.; Hunig. Interrupting CD28 costimulation before antigen rechallenge affects CD8+ T-cell expansion and effector functions during secondary response in mice. Eur. J. Immunol. 2016, 46, 1644–1655. [Google Scholar] [CrossRef]
Figure 1.
CD8+ immune T cells from ICOS-/- mice chronically infected with T. gondii possess an increased capability to eliminate T. gondii cysts. CD8+ T cells purified from the spleens from chronically infected WT and ICOS-/- mice were injected (2 x 106 cells/mouse) intravenously into chronically infected (infected and treated with sulfadiazine) SCID mice. As a control, two groups of the SCID mice did not receive any T cells. Seven days later (Day 7), the brains of the T cell recipients and one group of the control mice with no T cell transfer were obtained for measuring mRNA levels for (A) bradyzoite (cyst)-specific BAG1 by RT-PCR. Brain samples from another group of the control mice with no the T cell transfer were obtained on the day of the T cell transfer (Day 0) for the RT-PCR. (B) Numbers of T. gondii cysts per a sagittal section of the brain on Day 7 after their immunohistological staining. Three sections with 16 or 20 μm distance were counted for each mouse, and the mean value from the counts from the three sections was used for each mouse. (C) CD8+ T cells detected in a total of 10 randomly selected fields at X200 magnification of a sagittal section the brain of each of the recipients of CD8+ T cells from ICOS-/- or WT mice were counted microscopically after their immunohistological staining. A representative image (X200 magnification) of CD8+ T cells (stained in brown, some are arrowed) detected in a sagittal section of the brains of (D) WT CD8+ T cell recipients and (E) ICOS-/- CD8+ T cell recipients. (F) A representative image (X200 magnification) of a T. gondii cyst (stained in red, arrowed) attacked by WT CD8+ T cells (stained in brown). (G) CD8β mRNA levels in the brains of the recipients of WT and ICOS-/- CD8+ T cells, (H) the efficiency of cyst removal by CD8+ T cells that migrated into the brains of the recipients (the ratios of BAG1 mRNA level reduction by the CD8+ T cell transfer [differences between the mean value of BAG1 mRNA levels in the control mice with no T cell transfer and BAG1 mRNA levels in each of the recipients of WT or ICOS-/- CD8+ T cells] to amounts of CD8β mRNA in the brain of each recipient mouse). There were four SCID mice in each of the groups that received WT or ICOS-/- CD8+ T cells. (I) Frequencies of CD44highCD62Llow effector memory population in the splenic CD8+ T cells of chronically infected ICOS-/- and WT mice. There were three or four SCID mice in the control group without any T cell transfer at each of Day 0 and Day 7. In regard to the donors of the CD8+ T cells, there were three or four mice in each of infected WT and ICOS-/- mice, and their spleen cells were pooled within the same experimental group for purifying CD8+ T cells. Two independent experiments were performed. Panels A, G, and H show the RT-PCR results combined from the two independent experiments, which provided 7-8 mice in each experimental group. *P<0.05, **P<0.01, ***P<0.001, N.S., Not significant.
Figure 1.
CD8+ immune T cells from ICOS-/- mice chronically infected with T. gondii possess an increased capability to eliminate T. gondii cysts. CD8+ T cells purified from the spleens from chronically infected WT and ICOS-/- mice were injected (2 x 106 cells/mouse) intravenously into chronically infected (infected and treated with sulfadiazine) SCID mice. As a control, two groups of the SCID mice did not receive any T cells. Seven days later (Day 7), the brains of the T cell recipients and one group of the control mice with no T cell transfer were obtained for measuring mRNA levels for (A) bradyzoite (cyst)-specific BAG1 by RT-PCR. Brain samples from another group of the control mice with no the T cell transfer were obtained on the day of the T cell transfer (Day 0) for the RT-PCR. (B) Numbers of T. gondii cysts per a sagittal section of the brain on Day 7 after their immunohistological staining. Three sections with 16 or 20 μm distance were counted for each mouse, and the mean value from the counts from the three sections was used for each mouse. (C) CD8+ T cells detected in a total of 10 randomly selected fields at X200 magnification of a sagittal section the brain of each of the recipients of CD8+ T cells from ICOS-/- or WT mice were counted microscopically after their immunohistological staining. A representative image (X200 magnification) of CD8+ T cells (stained in brown, some are arrowed) detected in a sagittal section of the brains of (D) WT CD8+ T cell recipients and (E) ICOS-/- CD8+ T cell recipients. (F) A representative image (X200 magnification) of a T. gondii cyst (stained in red, arrowed) attacked by WT CD8+ T cells (stained in brown). (G) CD8β mRNA levels in the brains of the recipients of WT and ICOS-/- CD8+ T cells, (H) the efficiency of cyst removal by CD8+ T cells that migrated into the brains of the recipients (the ratios of BAG1 mRNA level reduction by the CD8+ T cell transfer [differences between the mean value of BAG1 mRNA levels in the control mice with no T cell transfer and BAG1 mRNA levels in each of the recipients of WT or ICOS-/- CD8+ T cells] to amounts of CD8β mRNA in the brain of each recipient mouse). There were four SCID mice in each of the groups that received WT or ICOS-/- CD8+ T cells. (I) Frequencies of CD44highCD62Llow effector memory population in the splenic CD8+ T cells of chronically infected ICOS-/- and WT mice. There were three or four SCID mice in the control group without any T cell transfer at each of Day 0 and Day 7. In regard to the donors of the CD8+ T cells, there were three or four mice in each of infected WT and ICOS-/- mice, and their spleen cells were pooled within the same experimental group for purifying CD8+ T cells. Two independent experiments were performed. Panels A, G, and H show the RT-PCR results combined from the two independent experiments, which provided 7-8 mice in each experimental group. *P<0.05, **P<0.01, ***P<0.001, N.S., Not significant.
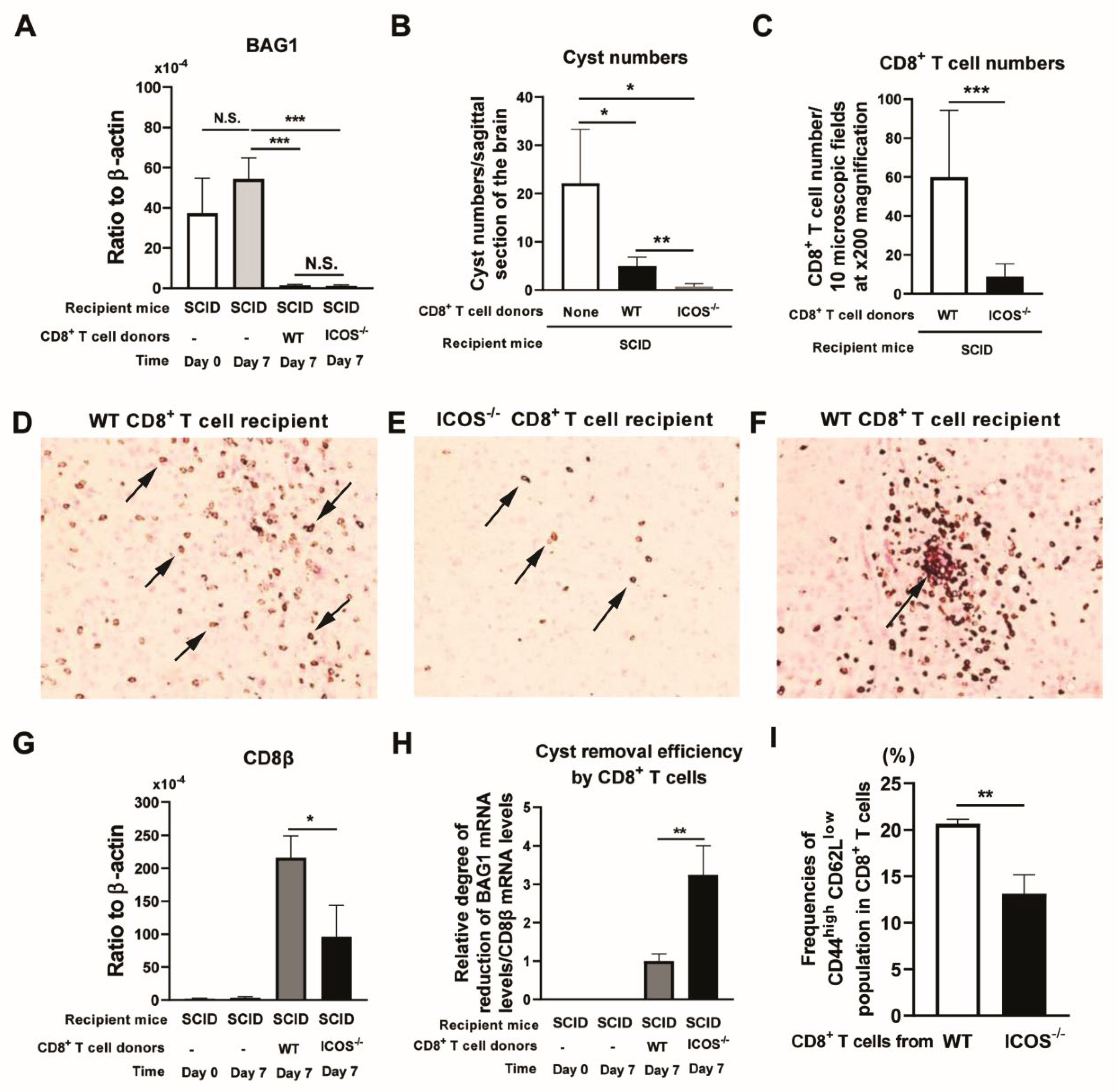
Figure 2.
Relative mRNA expression levels for perforin and T-bet in ratios to CD8β mRNA levels are greater in the brains of SCID mice that received ICOS-/- CD8+ T cells than those that received WT CD8+ T cells. CD8+ T cells purified from the spleens from chronically infected WT and ICOS-/- mice were injected (2 x 106 cells/mouse) intravenously into chronically infected (infected and treated with sulfadiazine) SCID mice. Seven days later (Day 7), the ratios of mRNA levels for (A) perforin, (B) GzmB, and (C) T-bet to mRNA levels to mRNA levels for CD8β were measured in the brains of those SCID mice by RT-PCR. There were four SCID mice in each of the groups. Two independent experiments were performed, and results combined from the two independent experiments (a total of 7-8 mice in each experimental group). *P<0.05, N.S., Not significant.
Figure 2.
Relative mRNA expression levels for perforin and T-bet in ratios to CD8β mRNA levels are greater in the brains of SCID mice that received ICOS-/- CD8+ T cells than those that received WT CD8+ T cells. CD8+ T cells purified from the spleens from chronically infected WT and ICOS-/- mice were injected (2 x 106 cells/mouse) intravenously into chronically infected (infected and treated with sulfadiazine) SCID mice. Seven days later (Day 7), the ratios of mRNA levels for (A) perforin, (B) GzmB, and (C) T-bet to mRNA levels to mRNA levels for CD8β were measured in the brains of those SCID mice by RT-PCR. There were four SCID mice in each of the groups. Two independent experiments were performed, and results combined from the two independent experiments (a total of 7-8 mice in each experimental group). *P<0.05, N.S., Not significant.
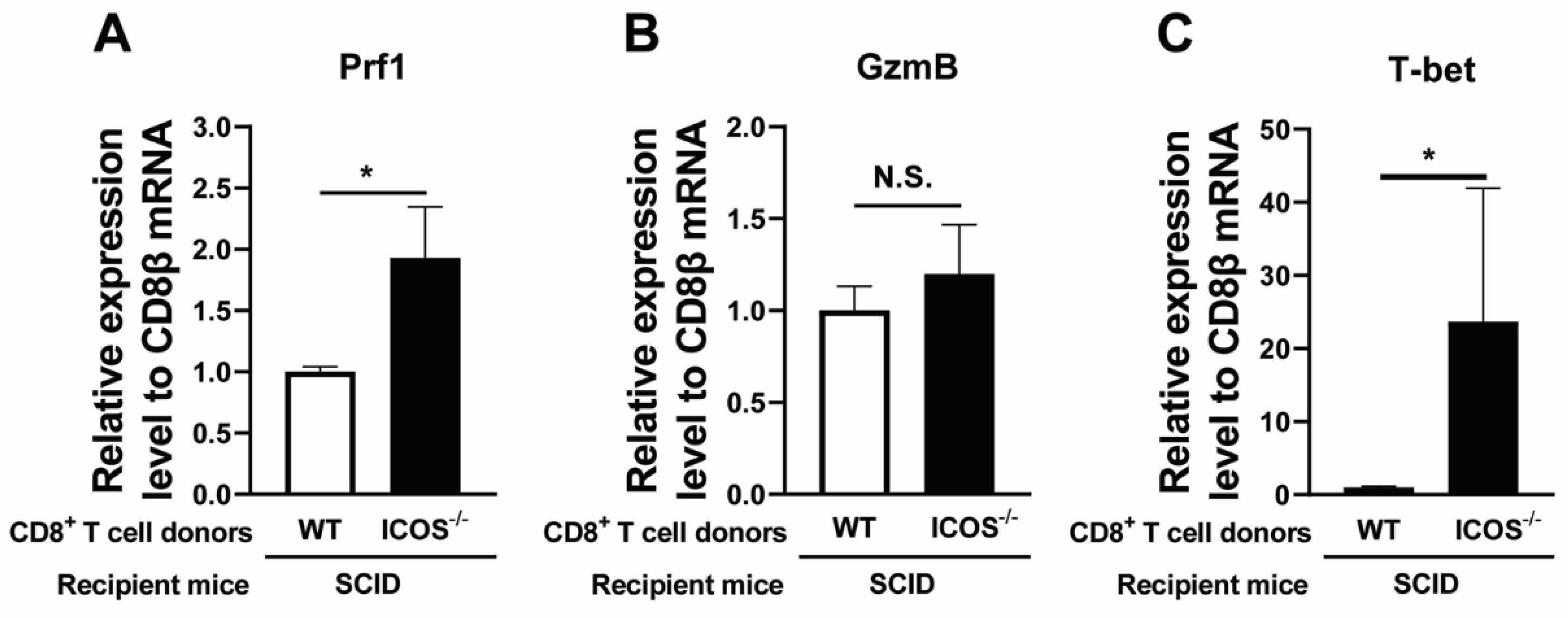
Figure 3.
Relative mRNA levels for CD28 in ICOS-/- CD8+ T cells that migrated into the brains of the recipient SCID mice are greater than those of the WT CD8+ T cells that migrated into the brains of recipient SCID mice (A and B), and strong correlations are present between the increased relative mRNA expression levels for CD28 and those for T-bet (C) and between relative mRNA expression levels for T-bet and those for perforin (D) in the CD8+ T cells that migrated into the brains of the recipients during their elimination of T. gondii cysts. CD8+ T cells purified from the spleens from chronically infected WT and ICOS-/- mice were injected (2 x 106 cells/mouse) intravenously into chronically infected (infected and treated with sulfadiazine) SCID mice. Seven days later, their brains were obtained for measuring (A) the ratios of 4-1BB (TNFRSF9) mRNA levels to CD8β mRNA levels, and (B) the ratios of CD28 mRNA levels to CD8β mRNA by RT-PCR. Correlations of (C) the ratios of CD28 mRNA/ CD8β mRNA levels with T-bet mRNA/CD8β mRNA levels and (D) the ratios of T-bet mRNA/ CD8β mRNA levels with perforin mRNA/CD8β mRNA levels with were examined in the brains of the recipients of the ICOS-/- and WT CD8+ T cells. In these correlation analyses, the data from both the recipients of ICOS-/- CD8+ T cells and those of WT CD8+ T cells were included. Two independent experiments were performed, and the results from these two experiments were combined (a total of 7-8 mice in each experimental group). *P<0.05. .
Figure 3.
Relative mRNA levels for CD28 in ICOS-/- CD8+ T cells that migrated into the brains of the recipient SCID mice are greater than those of the WT CD8+ T cells that migrated into the brains of recipient SCID mice (A and B), and strong correlations are present between the increased relative mRNA expression levels for CD28 and those for T-bet (C) and between relative mRNA expression levels for T-bet and those for perforin (D) in the CD8+ T cells that migrated into the brains of the recipients during their elimination of T. gondii cysts. CD8+ T cells purified from the spleens from chronically infected WT and ICOS-/- mice were injected (2 x 106 cells/mouse) intravenously into chronically infected (infected and treated with sulfadiazine) SCID mice. Seven days later, their brains were obtained for measuring (A) the ratios of 4-1BB (TNFRSF9) mRNA levels to CD8β mRNA levels, and (B) the ratios of CD28 mRNA levels to CD8β mRNA by RT-PCR. Correlations of (C) the ratios of CD28 mRNA/ CD8β mRNA levels with T-bet mRNA/CD8β mRNA levels and (D) the ratios of T-bet mRNA/ CD8β mRNA levels with perforin mRNA/CD8β mRNA levels with were examined in the brains of the recipients of the ICOS-/- and WT CD8+ T cells. In these correlation analyses, the data from both the recipients of ICOS-/- CD8+ T cells and those of WT CD8+ T cells were included. Two independent experiments were performed, and the results from these two experiments were combined (a total of 7-8 mice in each experimental group). *P<0.05. .

Figure 4.
Splenic CD8+ T cells of ICOS-/- mice chronically infected with T. gondii express greater levels of CD28 on their surfaces than CD8+ T cells of the infected WT mice. CD8+ T cells purified from the spleens of chronically infected WT and ICOS-/- mice were stained with FITC-labeled anti-mouse CD8α and PE-labeled anti-mouse CD28 mAbs and applied for flow cytometric analysis. For control, the cells were stained with FITC- and PE-labeled isotype control mAbs. A representative image of the FACS plots for expressions of CD8 and CD28 on CD8+ T cells from chronically infected (A) WT and (B) ICOS-/- mice. Comparisons of CD28 expression levels between (C) the population 1 of ICOS-/- CD8+ T cells and WT CD8+ T cells, and (D) between the population 2 of ICOS-/- CD8+ T cells and WT CD8+ T cells. (E) The median fluorescence intensity (MFI) of CD28 expressions on the population 2 of ICOS-/- CD8+ T cells and WT CD8+ T cells. *P<0.05, **P<0.01. N.S., Not significant.
Figure 4.
Splenic CD8+ T cells of ICOS-/- mice chronically infected with T. gondii express greater levels of CD28 on their surfaces than CD8+ T cells of the infected WT mice. CD8+ T cells purified from the spleens of chronically infected WT and ICOS-/- mice were stained with FITC-labeled anti-mouse CD8α and PE-labeled anti-mouse CD28 mAbs and applied for flow cytometric analysis. For control, the cells were stained with FITC- and PE-labeled isotype control mAbs. A representative image of the FACS plots for expressions of CD8 and CD28 on CD8+ T cells from chronically infected (A) WT and (B) ICOS-/- mice. Comparisons of CD28 expression levels between (C) the population 1 of ICOS-/- CD8+ T cells and WT CD8+ T cells, and (D) between the population 2 of ICOS-/- CD8+ T cells and WT CD8+ T cells. (E) The median fluorescence intensity (MFI) of CD28 expressions on the population 2 of ICOS-/- CD8+ T cells and WT CD8+ T cells. *P<0.05, **P<0.01. N.S., Not significant.
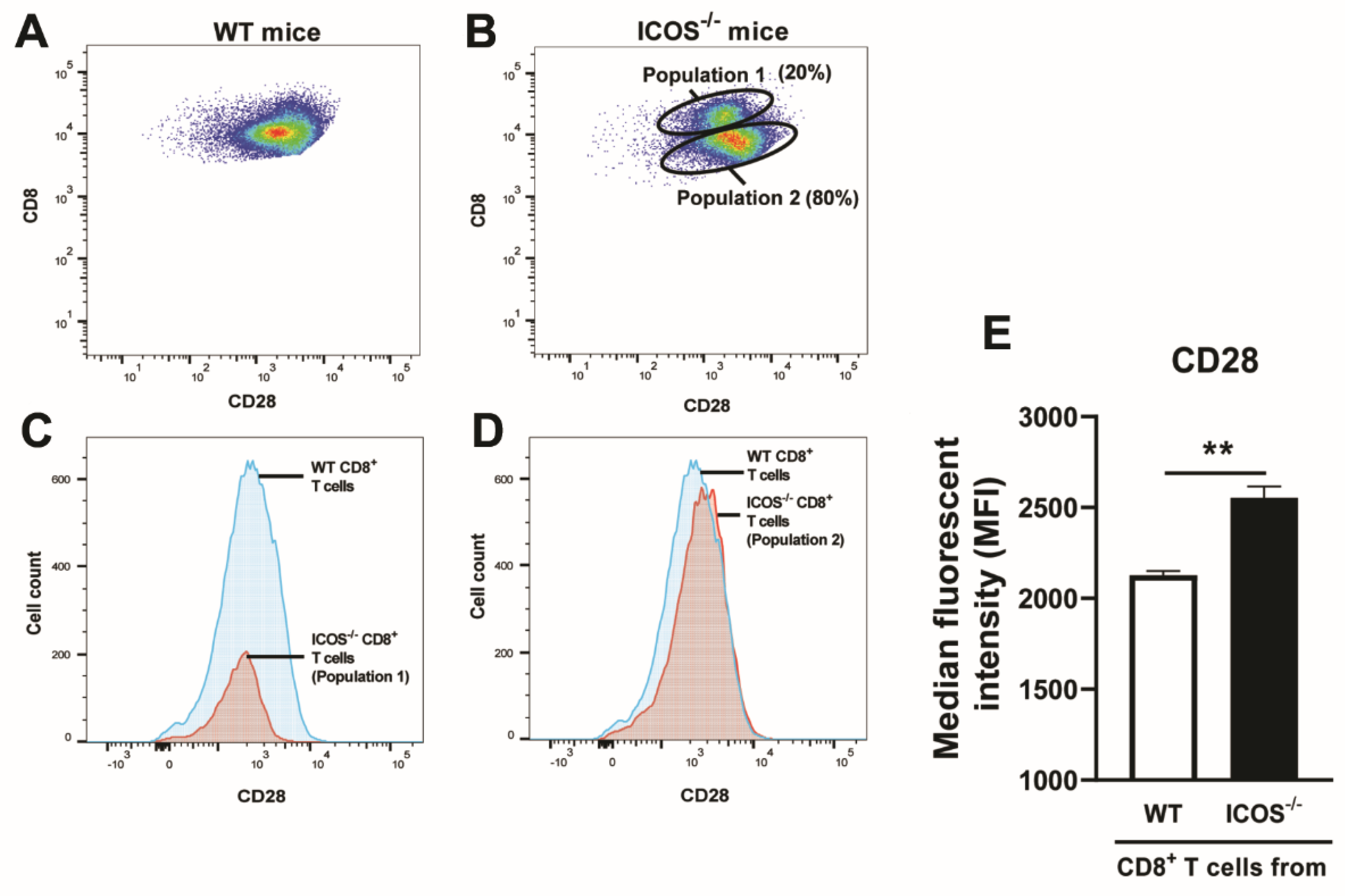
Figure 5.
Blockage of CD28−CD80/CD86 costimulatory pathway abolishes the cytotoxic functions of CD8+ T cells of ICOS-/- mice chronically infected with T. gondii. CD8+ T cells were purified from the spleens of chronically WT and ICOS-/- mice and cultured (3 x 105 cells/well) in 96 well culture plates with antigen-presenting cells (plastic-adherent cells) from the corresponding strain of mice in the presence or absence of T. gondii antigens (10 μg/ml) for 72 hrs. Blocking mAbs against ICOSL, both CD80 and CD86, or a combination of ICOSL, CD80, and CD86 were added at 10 μg/ml to a part of these cultures to block the ICOS-ICOSL, CD28-CD80/CD86, or both of these costimulatory pathways. As a control, isotype control mAbs were added in the same manner. Concentrations of GzmB in the culture supernatants in the cultures were measured by ELISA. (A) A comparison of GzmB levels in the culture supernatants of WT and ICOS-/- CD8+ T cells in the presence or absence of T. gondii antigens without any blocking mAbs. Comparisons of GzmB levels in the culture supernatants of (B) WT CD8+ T cells and (C) ICOS-/- CD8+ T cells in the presence and absence of the blocking mAbs against the ICOS-ICOSL or CD28-CD80/CD86 costimulatory pathways. There were 2 mice in each of infected WT and ICOS-/- mice, and their spleen cells were pooled within the same experimental group for purifying CD8+ T cells. There were 5 wells in each experimental group. **P<0.01, ***P<0.001, ****P<0.0001, N.S., Not significant.
Figure 5.
Blockage of CD28−CD80/CD86 costimulatory pathway abolishes the cytotoxic functions of CD8+ T cells of ICOS-/- mice chronically infected with T. gondii. CD8+ T cells were purified from the spleens of chronically WT and ICOS-/- mice and cultured (3 x 105 cells/well) in 96 well culture plates with antigen-presenting cells (plastic-adherent cells) from the corresponding strain of mice in the presence or absence of T. gondii antigens (10 μg/ml) for 72 hrs. Blocking mAbs against ICOSL, both CD80 and CD86, or a combination of ICOSL, CD80, and CD86 were added at 10 μg/ml to a part of these cultures to block the ICOS-ICOSL, CD28-CD80/CD86, or both of these costimulatory pathways. As a control, isotype control mAbs were added in the same manner. Concentrations of GzmB in the culture supernatants in the cultures were measured by ELISA. (A) A comparison of GzmB levels in the culture supernatants of WT and ICOS-/- CD8+ T cells in the presence or absence of T. gondii antigens without any blocking mAbs. Comparisons of GzmB levels in the culture supernatants of (B) WT CD8+ T cells and (C) ICOS-/- CD8+ T cells in the presence and absence of the blocking mAbs against the ICOS-ICOSL or CD28-CD80/CD86 costimulatory pathways. There were 2 mice in each of infected WT and ICOS-/- mice, and their spleen cells were pooled within the same experimental group for purifying CD8+ T cells. There were 5 wells in each experimental group. **P<0.01, ***P<0.001, ****P<0.0001, N.S., Not significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



