
Submitted:
24 September 2024
Posted:
25 September 2024
You are already at the latest version
Abstract
Genetic engineering has revolutionized our ability to modify microorganisms for various applications in agriculture, medicine, and industry. This review examines recent advances in genetic engineering techniques for bacteria, fungi, and oomycetes, with a focus on CRISPR-Cas systems. In bacteria, CRISPR-Cas9 has enabled precise genome editing, enhancing applications in antibiotic production and metabolic engineering. For fungi, despite challenges associated with their complex cell structures, CRISPR/Cas9 has advanced the production of enzymes and secondary metabolites. In oomycetes, significant plant pathogens, modified Agrobacterium-mediated transformation and CRISPR/Cas12a have contributed to developing disease-resistant crops. The review provides a comparative analysis of genetic engineering efficiencies across these microorganisms and addresses ethical and regulatory considerations. Future research directions include refining genetic tools to improve efficiency and expand applicability in non-model organisms. This comprehensive overview highlights the transformative potential of genetic engineering in microbiology and its implications for addressing global challenges in agriculture, medicine, and biotechnology.
Keywords:
Genetic engineering
; CRISPR
; Bacteria
; Fungi
; Oomycetes
; GEMs
1. Introduction
Genetic engineering is a transformative field that involves the manipulation of an organism's genetic material to alter its characteristics. In microorganisms, genetic engineering has become a crucial tool due to its potential to enhance various applications in medicine, agriculture, and industry. The advent of advanced genetic tools, such as CRISPR-Cas9, has revolutionized the ability to precisely edit microbial genomes, allowing for targeted modifications that can lead to significant improvements in microbial functions and applications [1].
Microbial genetic engineering involves the integration of target genes into chassis cells, enabling the production of recombinant genes that endow microorganisms with new phenotypes or enhance their ability to produce desired products [2]. This process has been facilitated by advancements in sequencing technologies and bioinformatics, which have uncovered a vast array of functional genes and gene clusters from even non-culturable microorganisms. These discoveries have opened new avenues for the construction of genetically engineered microorganisms (GEMs) that can be used in diverse applications, from bioremediation to the production of biofuels and pharmaceuticals [3].
The significance of genetic engineering in microorganisms is underscored by its ability to address critical challenges in various fields. For instance, in agriculture, GEMs can be used to enhance plant disease resistance, improve crop yields, and reduce reliance on chemical pesticides [1,4]. In medicine, they hold promise for the development of new therapeutic strategies, such as bacteriotherapy, where genetically engineered bacteria are used to produce therapeutic proteins and compounds directly within the human body [2]. The continued development and application of genetic engineering technologies in microorganisms have the potential to drive significant advancements across multiple sectors, contributing to improved health, sustainability, and economic growth [3].
Bacteria are indispensable in biotechnology due to their versatility and ease of genetic manipulation. They are extensively used in the production of antibiotics, enzymes, and biofuels. In agriculture, bacteria serve as biofertilizers and biopesticides, enhancing soil fertility and protecting crops from pests and diseases. Nitrogen-fixing bacteria, for instance, are crucial for sustainable agriculture as they naturally enrich soil with nitrogen, reducing the need for chemical fertilizers [5]. In medicine, bacteria are harnessed to produce therapeutic proteins and vaccines, and they are central to the development of new antibiotics to combat drug-resistant pathogens [6].
Fungi have long been utilized in biotechnology for their ability to produce a wide range of metabolites, including antibiotics, enzymes, and organic acids. They are essential in the food industry for the production of bread, beer, and cheese. They also improve agricultural crop yield and quality by enhancing plant physiology and stress tolerance. Fungi also play a significant role in environmental sustainability by decomposing organic matter, thus recycling nutrients in ecosystems [7]. In medicine, fungi are sources of important pharmaceuticals, such as penicillin, and are being explored for their potential in developing new drugs [8].
Oomycetes, although often less studied, are critical in agriculture as they include some of the most destructive plant pathogens, such as Phytophthora infestans, which causes late blight in potatoes. Managing oomycete diseases is vital for food security, and recent biotechnological advances have focused on understanding their pathogenic mechanisms to develop resistant crop varieties and effective control strategies [9]. Oomycetes also play roles in soil ecosystems, influencing nutrient cycling and plant health [10].
Together, these microorganisms are integral to advancing biotechnology, improving agricultural practices, and developing new medical therapies (Figure 1). This highlights their importance in addressing global challenges such as food security, environmental sustainability, and health care.
2. Genetic Engineering in Bacteria
2.1. Techniques and Tools
CRISPR-Cas9 has emerged as a groundbreaking tool in genetic engineering, particularly for bacteria, due to its precision and efficiency in editing genomes (Table 1). This technology harnesses the natural defense mechanism of bacteria against phages, allowing researchers to target and modify specific DNA sequences with unprecedented accuracy [11]. The CRISPR-Cas9 system utilizes a guide RNA (gRNA) to direct the Cas9 nuclease to a specific location in the genome, where it introduces a double-strand break. This break can then be repaired by the cell's natural repair mechanisms, facilitating the insertion, deletion, or modification of genetic material (Figure 1) [12].
In bacteria, CRISPR-Cas9 has been applied to a wide range of fields, including biotechnology and medicine. For instance, it has been used to engineer bacteria for the production of biofuels, pharmaceuticals, and other valuable compounds by enhancing metabolic pathways and optimizing production processes [11]. Moreover, CRISPR-Cas9 is being explored as a tool to combat antibiotic resistance, a growing global health concern. By precisely targeting and disrupting resistance genes, CRISPR-Cas9 can potentially restore the efficacy of existing antibiotics and help manage multidrug-resistant bacterial infections [13].
Recombineering is a technique that leverages homologous recombination facilitated by bacteriophage-derived recombination proteins, allowing for precise genetic modifications in bacterial genomes. This method is particularly advantageous because it requires only short homology arms (approximately 40–50 base pairs) to introduce genetic changes such as deletions, insertions, and point mutations [14]. Initially developed for Escherichia coli, recombineering has since been adapted for use in other bacteria, making it a versatile tool for genetic engineering across various species [15]. The integration of CRISPR-associated endonucleases has further enhanced recombineering by enabling efficient selection and screening of edited cells, thereby facilitating the creation of markerless and scarless mutants [14]. This approach is widely used for functional genomic studies, metabolic engineering, and the development of bacterial strains with improved traits for industrial applications [15].
Synthetic biology, on the other hand, involves the design and construction of new biological parts, devices, and systems, or the redesign of existing biological systems for useful purposes. This field has grown exponentially, driven by advances in DNA synthesis and assembly technologies, which allow for the creation of complex genetic circuits and metabolic pathways [16]. In bacteria, synthetic biology enables the rational engineering of microbial chassis to produce therapeutic compounds, biofuels, and other valuable products. By integrating computational models with genetic engineering, synthetic biology facilitates the design of organisms with tailored functionalities, enhancing their robustness and efficiency in various applications [17]. This approach not only improves the scalability of microbial production processes but also opens new avenues for addressing challenges in agriculture, medicine, and environmental sustainability [16].
2.2. Applications
Genetic engineering in bacteria, particularly through the use of Escherichia coli as a host system, has significantly advanced the production of therapeutic proteins. E. coli is favored for its simplicity, cost-effectiveness, and rapid growth, making it an ideal platform for large-scale production of protein therapeutics [18]. The ability to manipulate bacterial genomes using recombinant DNA technology allows for the production of proteins with modified features, enhancing their stability and functionality. Despite the lack of post-translational modifications in bacteria, which are often essential for the activity of human proteins, engineering strategies such as protein fusion and the expression of proteins in the periplasmic space have been developed to overcome these limitations (Table 2) [18].
Recent advancements have focused on optimizing E. coli systems for the expression of complex proteins that were traditionally difficult to produce in prokaryotic hosts. For instance, continuous manufacturing and cell-free production techniques have been explored to enhance the yield and quality of therapeutic proteins [19]. These advancements have expanded the range of proteins that can be produced in bacterial systems, including those that are typically extracted from mammalian tissues, such as alpha-1 antitrypsin and antithrombin III [19]. The development of these systems not only reduces production costs but also mitigates the risk of viral contamination associated with mammalian cell lines.
Moreover, genetic engineering has enabled the development of engineered probiotics that can deliver therapeutic proteins directly within the human body. These engineered probiotics interact with the host's immune system and can be used to deliver enzymes, vaccines, and other therapeutic agents, offering a more efficient and targeted drug delivery system compared to traditional methods [20]. However, challenges remain in ensuring the stability and efficacy of these engineered probiotics in the human gastrointestinal tract, necessitating further research and development to optimize their therapeutic potential [20]. The genetic engineering of bacteria for the production of therapeutic proteins continues to be a dynamic and promising field, with significant implications for medicine and biotechnology.
Genetic engineering in bacteria has been pivotal in advancing industrial applications, particularly in the fields of bioremediation and biofuel production. These applications leverage the unique capabilities of bacteria to metabolize a wide range of substrates, including pollutants and biomass, into valuable products. For instance, bioremediation is a process where genetically engineered bacteria are used to detoxify and remove pollutants from the environment. Bacteria such as Pseudomonas and Bacillus have been engineered to degrade complex organic compounds like hydrocarbons, pesticides, and heavy metals, which are major environmental contaminants [21]. Genetic modifications enhance the metabolic pathways of these bacteria, enabling them to break down pollutants more efficiently and even under extreme environmental conditions [22]. The use of genetically engineered bacteria in bioremediation not only aids in environmental cleanup but also offers a sustainable alternative to conventional chemical methods, reducing the ecological footprint of remediation efforts [23].
In the realm of biofuel production, genetic engineering has been instrumental in optimizing bacterial strains to convert biomass into biofuels such as ethanol, butanol, and biodiesel. The manipulation of metabolic pathways in bacteria like Clostridium thermocellum has improved the efficiency of converting lignocellulosic biomass into bioethanol, addressing one of the key challenges in biofuel production [23]. Genetic engineering techniques, including CRISPR-Cas9, have been employed to enhance the tolerance of bacteria to inhibitors and end-products, thereby increasing biofuel yield and reducing production costs [24]. These advancements in bacterial engineering are crucial for developing economically viable and sustainable biofuel industries, contributing to the transition towards renewable energy sources [25].
2.3. Challenges and Future Directions
Genetic engineering in bacteria faces several limitations that hinder its full potential, particularly when applied to non-model organisms. One major challenge is the diversity of microbial species, which poses difficulties in developing universal genetic tools that can be effectively applied across different bacterial species [26]. Many genetic engineering techniques, such as recombineering and CRISPR-Cas9, have been optimized for a few model organisms like E. coli, but their efficiency and applicability often decrease significantly in other bacteria. This limitation is compounded by the lack of detailed knowledge about the genetic and metabolic pathways in non-model bacteria, which complicates efforts to engineer these organisms for specific applications [26].
Recent advancements, however, are addressing these challenges. For instance, the development of serine recombinase-assisted genome engineering (SAGE) offers a promising solution by providing a high-efficiency genome integration tool that can be applied to a wide range of bacterial species [27]. This method allows for the stable insertion of multiple genetic elements into bacterial chromosomes, thereby expanding the toolkit available for engineering diverse bacterial hosts. Moreover, advances in omics technologies and computational tools are enabling a more systematic approach to understanding and manipulating the genetic makeup of non-model bacteria, facilitating the design of strains with enhanced capabilities for industrial applications [26].
Despite its potential, the application of CRISPR-Cas9 in bacteria also faces several challenges, including off-target effects and delivery issues. Researchers are actively working on improving the specificity of the gRNA and developing efficient delivery systems to introduce the CRISPR components into bacterial cells [12]. Advances in these areas could significantly enhance the utility of CRISPR-Cas9 in bacterial genetic engineering, paving the way for innovative solutions in biotechnology and medicine [11]. CRISPR-Cas9 represents a powerful tool with the potential to revolutionize bacterial genetic engineering, offering new opportunities for scientific and industrial applications.
Also, the genetic engineering of bacteria for industrial applications holds significant promise for addressing environmental and energy challenges. By enhancing the natural capabilities of bacteria, these technologies offer innovative solutions for sustainable industrial processes.
Looking to the future, the integration of synthetic biology with genetic engineering holds significant promise for overcoming current limitations. By designing modular genetic parts and systems, synthetic biology can provide more flexible and robust solutions for bacterial engineering [27]. Additionally, ongoing research into improving the specificity and efficiency of CRISPR-Cas systems, as well as developing novel gene-editing tools, is expected to enhance the precision of bacterial genetic modifications. These developments will likely lead to more efficient and versatile bacterial strains [28,29,30,31] that can be used in a variety of biotechnological applications, from environmental remediation to the production of biofuels and pharmaceuticals [25].
3. Genetic Engineering in Fungi
3.1. Transformation Techniques
Polyethylene glycol (PEG)-mediated transformation is a widely used technique in the genetic engineering of fungi, particularly for introducing foreign DNA into fungal protoplasts. This method capitalizes on the ability of PEG to facilitate the uptake of DNA by protoplasts, which are cells that have had their cell walls removed. The transformation process typically involves mixing the protoplasts with a solution containing PEG, calcium ions, and the DNA to be introduced. The PEG promotes cell agglomeration, which aids in DNA uptake, while calcium ions facilitate the DNA's entry into the cells [32]
Recent advancements have optimized PEG-mediated transformation systems for various fungal species, improving transformation efficiencies and expanding their applicability. For instance, in Pleurotus eryngii var. ferulae, comprehensive optimization of the transformation protocol, including the use of multiple endogenous promoters, has led to the establishment of an efficient PEG-mediated transformation system. This system is characterized by high conversion efficiency and mild reaction conditions, making it a cost-effective and convenient option for genetic manipulation [33].
The molecular weight of PEG is a critical factor influencing transformation efficiency. Studies have shown that lower molecular weight PEGs, such as PEG 3350, yield higher transformation efficiencies compared to higher molecular weight variants like PEG 8000, which can drastically reduce transformation success rates [32]. Additionally, the preparation of high-quality protoplasts is crucial, as the integrity and viability of protoplasts significantly affect the transformation outcome. The use of cell wall-degrading enzymes and appropriate osmotic stabilizers is essential to maintain protoplast stability during the transformation process [34].
PEG-mediated transformation has been successfully applied to a range of fungi, including economically important species like Pleurotus ostreatus, Ganoderma lucidum, and Lentinus edodes. In these species, the transformation efficiencies have been significantly improved, making PEG-mediated transformation a valuable tool for genetic engineering aimed at strain improvement and the study of gene function [33,35]. Despite its advantages, challenges such as low integration efficiency and the temporary expression of introduced genes remain, necessitating further refinement of the technique to enhance its reliability and effectiveness in different fungal systems.
Agrobacterium-mediated transformation (AMT) has become a pivotal technique in the genetic engineering of fungi, offering a robust method for introducing foreign DNA into fungal genomes. This method leverages the natural ability of Agrobacterium tumefaciens to transfer DNA to host cells, a process initially exploited in plants and now adapted for fungi. AMT is particularly valuable for its ability to achieve single-copy integration of transferred DNA, which is often more stable and predictable compared to other transformation methods. This technique has been successfully applied to a wide range of fungal species, including both filamentous fungi and yeasts, making it a versatile tool in fungal biotechnology [36,37].
Recent studies have also highlighted the effectiveness of AMT in various fungal species, such as Aspergillus vadensis and Simplicillium subtropicum. In A. vadensis, the optimization of co-cultivation conditions, particularly temperature, was found to be crucial for successful transformation, demonstrating the importance of fine-tuning environmental parameters to enhance transformation efficiency [37]. Similarly, in S. subtropicum, the choice of Agrobacterium strain and spore concentration during co-cultivation were key factors influencing the number of successful transformants, underscoring the need for species-specific protocol adjustments [36].
The application of AMT extends beyond simple gene insertion. It is a powerful tool for functional genomics, allowing for both forward and reverse genetic approaches. In forward genetics, AMT facilitates the creation of mutant libraries through random T-DNA insertional mutagenesis, which can be screened for phenotypic changes to identify gene functions. In reverse genetics, targeted gene disruptions or deletions can be achieved, enabling detailed studies of gene roles in fungal biology [38]. This versatility makes AMT a cornerstone technique for exploring complex genetic traits and improving industrially relevant fungal strains [36,37].
Despite its advantages, AMT is not without challenges. The efficiency of transformation can vary significantly between species and even among different strains of the same species. Factors such as the presence of phenolic inducers like acetosyringone, the pH of the medium, and the duration of co-cultivation are critical parameters that need careful optimization for each application. Nonetheless, the ongoing refinement of AMT protocols continues to expand its applicability and effectiveness, making it an indispensable tool in the genetic engineering of fungi [36,37].
3.2. CRISPR-Cas9
The CRISPR-Cas9 system has emerged as a transformative tool in the genetic engineering of fungi, enabling precise genome editing that has significantly advanced both basic research and industrial applications (Table 1). This technology is particularly effective in filamentous fungi, where it has been used to manipulate genes involved in the synthesis of secondary metabolites. For instance, CRISPR-Cas9 has facilitated the activation of silent gene clusters, leading to the discovery of new bioactive compounds. This capability is crucial for industries that rely on fungal metabolites, such as pharmaceuticals and agriculture, where the production of specific compounds can be enhanced or new ones can be discovered [39,40].
In the realm of edible fungi, CRISPR-Cas9 has been instrumental in the molecular breeding of various species, including Agaricus bisporus and Ganoderma lucidum. The technology allows for high-efficiency and high-precision genome modifications, which are essential for developing superior strains with enhanced nutritional and medicinal properties. This application is particularly relevant in the context of the rapidly growing edible fungi industry, where traditional breeding methods are often slow and labor-intensive. CRISPR-Cas9 offers a more streamlined and effective approach to strain improvement, addressing both consumer demand and industrial needs [41].
Despite its advantages, the application of CRISPR-Cas9 in fungi is not without challenges. One significant issue is the preference of many fungal species for non-homologous end joining (NHEJ) over homology-directed repair (HDR), which can limit the precision of genetic modifications. To overcome this, researchers have developed mutant strains with impaired NHEJ pathways, thereby enhancing HDR efficiency. Additionally, the design of single guide RNAs (sgRNAs) and the potential for off-target effects remain areas of ongoing research and development. Advances in computational tools for sgRNA design and the engineering of Cas9 variants are helping to address these challenges, making CRISPR-Cas9 an increasingly reliable tool for fungal genome editing [42,43].
3.3. Applications
Genetic engineering in fungi has significantly advanced the production of enzymes and secondary metabolites, which are crucial for various industrial applications. Fungi are prolific producers of enzymes, such as cellulases and amylases, which are widely used in industries ranging from biofuel production to food processing. Recent research has focused on optimizing fungal strains for enhanced enzyme production through genetic modifications. For instance, the use of CRISPR/Cas9 technology has facilitated targeted gene editing in fungi, allowing for the precise modification of genes involved in enzyme biosynthesis pathways. This has led to increased enzyme yields and improved enzyme characteristics, such as stability and activity under industrial conditions (Table 2) [44].
In addition to enzyme production, fungi are renowned for their ability to synthesize a diverse array of secondary metabolites, which have significant pharmaceutical and agricultural applications. These metabolites include antibiotics, immunosuppressants, and anticancer agents. The genetic engineering of fungi to enhance the production of these secondary metabolites involves manipulating biosynthetic gene clusters (BGCs) that encode the necessary enzymes and regulatory proteins. Advances in genomic and transcriptomic technologies have enabled the identification and activation of cryptic BGCs, leading to the discovery of new bioactive compounds. Techniques such as CRISPR/Cas9 have been instrumental in this process, allowing for the precise activation or silencing of specific genes within these clusters [43].
The application of systematic experimental design and chemometrics analysis has further improved the production of targeted secondary metabolites. By optimizing culture conditions and nutrient concentrations, researchers have been able to significantly increase the yield and diversity of secondary metabolites produced by fungi. This approach not only enhances the production of known metabolites but also facilitates the discovery of novel compounds with potential therapeutic applications. The integration of genetic engineering with advanced analytical techniques thus holds great promise for the sustainable production of valuable fungal metabolites [44].
Genetic engineering offers a powerful alternative to conventional breeding methods for increasing disease resistance in plants. Fungal diseases pose a significant threat to crops, and traditional methods, such as fungicide application, often have limitations, including non-specificity and environmental hazards. Genetic engineering allows for the introduction of genes that confer resistance to multiple pathogens simultaneously, even if these genes do not exist in the natural gene pool of the plant. Techniques like CRISPR/Cas9 have been utilized to enhance disease resistance in crops by enabling precise genome editing. This approach not only targets plant genomes but also the genomes of fungal pathogens, providing new strategies for plant disease management [1,45,46].
Fungi have also been used in food production for their nutritional benefits and sustainability. Genetic engineering can enhance these benefits by overcoming the limitations of non-engineered fungal strains. For example, the industrial fungus Trichoderma reesei has been genetically modified to produce significant quantities of proteins such as egg white and milk protein. Extending such synthetic biology tools to food-safe fungi could enhance the nutritional and sensory properties of fungal foods, such as mycoprotein, making them more suitable for human diets. This approach also offers a sustainable alternative to traditional animal agriculture, reducing environmental impact [47].
Filamentous fungi, known for their efficient protein secretion, serve as excellent cell factories for producing valuable agricultural products. Genetic engineering facilitates the development of fungi capable of producing secondary metabolites and proteins that can be used as bio-inputs in agriculture. These bio-inputs, which include biofertilizers and biopesticides, contribute to sustainable agricultural practices by enhancing soil health and crop productivity while minimizing the use of chemical fertilizers and pesticides [3,48].
3.4. Challenges and Innovations
One of the primary challenges in the genetic engineering of fungi is the low transformation efficiency, especially in filamentous fungi. This inefficiency is often due to the complex cell wall structure of fungi and their preference for non-homologous end joining (NHEJ) over homologous recombination (HR) for DNA repair. Traditional methods such as Agrobacterium-mediated transformation and protoplast transformation have been used, but they often result in low success rates, particularly in species with complex genomes like medicinal mushrooms [48]. Additionally, the lack of sufficient selection markers and the difficulty in achieving precise gene targeting further complicate genetic modifications in these organisms [43].
Recent advancements have focused on overcoming these challenges through the application of CRISPR/Cas technology. This system has been adapted to improve gene editing efficiency in various fungal species. For example, optimizing the CRISPR/Cas9 protocol in Penicillium digitatum has significantly increased editing efficiency from 10% to much higher levels by targeting genes associated with easily observable phenotypes [49]. Moreover, the development of modular synthetic biology toolkits has expanded the possibilities for genetic engineering in fungi, allowing for the integration and regulation of multiple genes and pathways simultaneously [47].
While genetic engineering in fungi faces significant challenges, particularly in transformation efficiency, recent innovations offer promising solutions. These advancements not only improve the feasibility of genetic modifications but also open new avenues for the application of fungi in various biotechnological fields. More specifically, genetic engineering in fungi holds great potential for advancing agricultural biotechnology. By enhancing disease resistance, improving nutritional value, and developing sustainable agricultural practices, genetically engineered fungi can play a crucial role in meeting the growing demands for food security and environmental sustainability.
The future of fungal genetic engineering looks promising with the continuous refinement of CRISPR/Cas systems and the development of new computational tools for sgRNA design. These innovations are expected to enhance the precision and efficiency of gene editing by considering factors such as chromatin architecture and sgRNA secondary structures [43]. Furthermore, the identification of neutral loci for gene integration in fungi like Aspergillus oryzae could lead to more stable and predictable expression of engineered traits, thereby expanding the applications of fungi in biotechnology and agriculture [47].
4. Genetic Engineering in Oomycetes
4.1. Challenges in Oomycete Genetics
Genetic engineering in oomycetes, a group of eukaryotic microorganisms including many plant pathogens, presents significant challenges due to their complex life cycles and genetic structures. These challenges have historically hindered the progress of functional genomics research in oomycetes compared to other microorganisms.
One of the primary difficulties in genetic manipulation of oomycetes is their lower transformation efficiency and homologous recombination frequency compared to fungi. This makes it more challenging to introduce genetic changes effectively [50,51]. Additionally, the manipulation of the sexual stage in oomycetes is notoriously difficult, particularly in species such as Phytophthora [52]. This complexity is compounded by the unique features of oomycete genomes, such as their large size, high ploidy levels, and unique cell wall compositions, which differ significantly from those of true fungi [51].
Recent advances in CRISPR-Cas technologies have begun to address some of these challenges. CRISPR-Cas systems, including Cas9 and Cas12a, have been adapted for use in oomycetes, offering new tools for genome editing (Table 1). However, the application of these technologies in oomycetes faces specific challenges related to the delivery, expression, and design of CRISPR components, leading to variable editing rates across different studies [53]. Furthermore, the lack of effective selection markers for oomycetes further complicates genetic engineering efforts. Although some progress has been made with the development of fungicide-resistance genes as selection markers, these are not yet widely applicable across different oomycete species [1,54].
Despite these challenges, the development of genome editing techniques is crucial for advancing our understanding of oomycete biology and improving disease management strategies. The ability to manipulate oomycete genomes could lead to significant breakthroughs in controlling plant diseases caused by these pathogens, which are of considerable economic importance due to their impact on global agriculture [1,9]. As research continues to overcome these genetic engineering challenges, it is expected that more efficient and effective methods will be developed, facilitating deeper insights into oomycete genetics and their interactions with host plants.
4.2. Recent Advances
Recent advances in genetic engineering of oomycetes have seen significant progress with the development of a modified Agrobacterium-mediated transformation (AMT) method (Figure 2). This method has been particularly effective for oomycete pathogens such as Phytophthora infestans and Plasmopara viticola, which are known for their destructive impact on agriculture [2]. The modified AMT technique involves the prokaryotic expression of the Arabidopsis bZIP gene AtVIP1 in Agrobacterium tumefaciens. This gene is crucial for AMT but is absent in oomycete genomes, and its introduction has led to increased transformation efficiency in P. infestans strains. The method also enabled the successful transformation of P. viticola, a species notoriously difficult to transform due to its inability to grow in axenic culture [2].
Recent advances in genetic engineering of oomycetes have also been significantly bolstered by the application of CRISPR-Cas12a for targeted mutagenesis. This technology has opened new avenues for genetic manipulation in oomycetes, a group of filamentous microbes that include many plant pathogens. The Cas12a system, derived from the Lachnospiraceae bacterium, has been particularly effective in species such as Phytophthora palmivora and Phytophthora infestans [55]. Unlike Cas9, which can be toxic to P. infestans, Cas12a offers a non-toxic alternative, making it a preferred choice for genome editing in these organisms [56].
By integrating this modified AMT method with CRISPR/Cas12a genome editing, researchers have achieved targeted mutagenesis in P. infestans. This approach has allowed for the creation of loss-of-function mutations in key genes, such as a MADS-box transcription factor gene, leading to reduced sporulation and virulence. Additionally, targeting the avirulence effector gene Avr8 demonstrated that its loss could enable the pathogen to evade the host's immune response, as seen in potato plants carrying the resistance gene R8. These findings underscore the potential of the modified AMT method as a powerful tool for molecular genetic studies in oomycetes and other microorganisms, facilitating advances in understanding pathogen biology and developing disease management strategies [2].
Another notable advancement in this area is the development of a multiplex CRISPR-Cas12a editing protocol for P. palmivora, which allows for the simultaneous knockout of multiple genes. This protocol employs a time-efficient transformation method using zoospore electroporation and a D156R point mutation in the Cas12a enzyme to enhance its activity at 25°C. This approach has been successfully used to knock out genes encoding adenine phosphoribosyltransferase (APT), rendering the mutants resistant to 2-fluoroadenine (2-FA) while retaining full virulence on Nicotiana benthamiana [57]. The use of 2-FA as a counter-selection marker provides a practical alternative to antibiotic selection, offering more flexibility for subsequent genetic manipulations.
These advances in CRISPR-Cas12a technology not only enhance the genetic toolkit available for oomycetes but also facilitate the study of gene function and pathogenicity. By enabling precise and efficient genome editing, researchers can better understand the molecular mechanisms underlying oomycete virulence and develop strategies for disease management. The ability to perform multiplex gene editing is particularly valuable for studying complex traits controlled by multiple genes, thereby accelerating the pace of research in this field [53].
4.3. Applications
Genetic engineering in oomycetes, a group of filamentous microorganisms that include significant plant pathogens, has seen substantial advancements in recent years. These advancements have provided deeper insights into pathogenesis and facilitated the development of disease-resistant crops.
Oomycetes, such as Phytophthora infestans and Plasmopara viticola, are notorious for causing severe agricultural damage. Recent studies have focused on enhancing genetic transformation methods to better understand these pathogens. For instance, Wang et al. [2] developed a modified Agrobacterium-mediated transformation method, which improved the efficiency of genetic manipulation in oomycetes. This approach, combined with CRISPR/Cas12a genome editing, enabled targeted mutagenesis in P. infestans, revealing the role of specific genes in pathogenicity and virulence [2]. Such genetic tools have been crucial in dissecting the molecular mechanisms of oomycete pathogenesis, which were previously challenging due to limited genetic manipulation techniques (Table 2).
The application of CRISPR technology has revolutionized the development of disease-resistant crops. This system allows precise genome editing, which has been applied to enhance resistance against oomycete pathogens in various crops, including rice, wheat, and grape [1]. By targeting specific genes in both the host plants and the pathogens, researchers have been able to enhance plant immunity and reduce susceptibility to infections. For example, the CRISPR/Cas9 system has been used to edit genes in oomycetes that are crucial for their pathogenicity, thereby reducing their ability to infect host plants [53].
4.4. Future Prospects
Recent advances in genetic engineering of oomycetes have significantly contributed to understanding pathogenesis and developing disease-resistant crops. The application of CRISPR-Cas technologies, particularly CRISPR-Cas9 and CRISPR-Cas12a, has facilitated precise genome editing in oomycetes, which are notorious for causing devastating plant diseases. These technologies have been instrumental in dissecting the molecular mechanisms of oomycete pathogenesis by enabling targeted mutagenesis of key pathogenicity genes. For instance, CRISPR-Cas9 has been employed to knock out genes in Phytophthora infestans, leading to insights into its virulence strategies and host interactions [1,53].
The integration of CRISPR-Cas systems into oomycete research has also opened new possibilities for developing disease-resistant crops. By editing the genomes of both the pathogens and host plants, researchers can enhance plant resistance to oomycete infections. This dual approach allows for the identification and modification of susceptibility genes in crops, thereby reducing their vulnerability to oomycete attacks. Recent studies have demonstrated the potential of this strategy by creating resistant phenotypes in crops such as rice, wheat, and tomatoes through CRISPR-mediated editing of specific resistance genes [58,59].
Moreover, the development of modified transformation methods, such as Agrobacterium-mediated transformation combined with CRISPR-Cas12a, has further advanced the genetic manipulation of oomycetes. This approach has been successfully applied to Phytophthora infestans and Plasmopara viticola, resulting in enhanced transformation efficiencies and effective targeted mutagenesis. Such innovations not only facilitate the study of oomycete biology but also aid in the development of crops with improved resistance to these pathogens, thereby contributing to sustainable agricultural practices and food security [2].
The future of genetic engineering in oomycetes looks promising, with ongoing research focusing on expanding the genetic toolbox available for these pathogens. The development of novel transformation systems and gene editing technologies continues to open new avenues for studying oomycete biology and improving crop resistance. As more genome sequences of oomycetes become available, researchers can perform comprehensive genomic comparisons, which will enhance our understanding of the evolution and mechanisms of pathogenicity in these organisms [60]. These insights will be instrumental in devising innovative strategies for sustainable agriculture and food security.
In conclusion, genetic engineering in oomycetes has significantly advanced our understanding of their pathogenesis and facilitated the development of disease-resistant crops. Continued research and technological innovations will likely yield further breakthroughs, contributing to more resilient agricultural systems.
5. Comparative Analysis
5.1. Comparison of Techniques
In bacteria, genetic engineering has been revolutionized by tools such as CRISPR/Cas9, which allows for precise genome editing. This technology has been widely adopted due to its simplicity, cost-effectiveness, and ability to target multiple genes simultaneously [61]. Bacteria's relatively simple and well-understood genetic systems make them ideal candidates for genetic manipulation, enabling advancements in areas like antibiotic resistance research and metabolic engineering [62]. The high efficiency of CRISPR/Cas9 in bacteria is largely due to their single-celled nature and the absence of complex cellular structures that can impede genetic modifications (Table 1).
In fungi, genetic engineering has faced more challenges compared to bacteria. The presence of a rigid cell wall and complex mycelial structures makes the transformation process more difficult [63]. However, advancements in CRISPR/Cas9 technology have significantly improved the precision and efficiency of genetic modifications in fungi [43]. Despite these advancements, the integration of exogenous DNA is often less efficient in fungi due to their intrinsic ploidy and propagation characteristics [64]. Nonetheless, CRISPR/Cas9 has facilitated targeted mutagenesis in fungi, allowing for the study of gene function and the development of disease-resistant strains [65].
Oomycetes, a group of filamentous microorganisms, present unique challenges for genetic engineering due to their distinct evolutionary lineage and complex life cycles. Traditional transformation methods, such as homologous recombination, have been less effective in oomycetes. However, recent developments in transformation systems and gene editing technologies, including CRISPR/Cas12a, have enhanced genetic manipulation in these organisms [66]. The modified Agrobacterium-mediated transformation method has improved transformation efficiency in oomycetes like Phytophthora infestans and Plasmopara viticola [2]. These advancements have opened new avenues for studying pathogenicity and developing disease-resistant crops, although the complexity of oomycete genomes still poses significant challenges [60,67].
While genetic engineering tools have been effectively applied across bacteria, fungi, and oomycetes, the success and applicability vary significantly among these groups due to their distinct biological characteristics. Continued advancements in genetic technologies are essential for overcoming the existing challenges and enhancing the applicability of these tools across different organisms.
5.2. Case Studies
In bacteria, genetic engineering has been extensively used to enhance industrial and clinical applications. For instance, CRISPR-Cas9 technology has been employed to modify bacterial genomes for the production of antibiotics and other valuable compounds [68]. This technology allows for precise editing of bacterial DNA, enabling the creation of strains with desirable traits such as increased production efficiency or resistance to certain environmental conditions [69]. Additionally, the engineering of gut microbiota has shown promise in medical applications, where genetically modified bacteria can produce therapeutic compounds directly in the human gut, offering potential treatments for metabolic diseases and cancer [70]).
In fungi, CRISPR/Cas9 has been used to improve strains of edible fungi, enhancing their nutritional and medicinal value. This technology has enabled targeted gene editing in species such as Agaricus bisporus and Ganoderma lucidum, leading to improved growth rates and resistance to diseases [71]. The application of CRISPR/Cas9 in fungi has also facilitated the discovery of new bioactive compounds by enabling the manipulation of biosynthetic gene clusters responsible for secondary metabolite production [63]. Despite challenges such as off-target effects and low editing efficiency, these advancements highlight the potential of genetic engineering to revolutionize fungal biotechnology [65].
Oomycetes, known for their role as plant pathogens, have benefited from recent advancements in genetic engineering techniques. A notable example is the use of a modified Agrobacterium-mediated transformation method combined with CRISPR/Cas12a to edit the genomes of Phytophthora infestans and Plasmopara viticola [2]. This approach has enabled the creation of loss-of-function mutations in genes critical for pathogenicity, providing insights into disease mechanisms and potential strategies for developing disease-resistant crops. These advancements represent significant progress in overcoming the historical challenges associated with genetic manipulation in oomycetes, offering new tools for plant disease management [53].
Genetic engineering has been effectively applied across bacteria, fungi, and oomycetes, each with specific examples demonstrating the potential of these technologies to address diverse biological and industrial challenges. Continued research and innovation in genetic tools are expected to further enhance their applicability and effectiveness across these groups.
6. Ethical and Regulatory Considerations
6.1. Safety Concerns
Genetic engineering in microorganisms, including bacteria, fungi, and oomycetes, presents several potential risks that need careful consideration. These risks stem from the inherent characteristics of microorganisms and the nature of genetic modifications.
In bacteria, one of the primary concerns is the potential for horizontal gene transfer (HGT), which can lead to the spread of antibiotic resistance genes and other undesirable traits across microbial communities [72]. This process can occur naturally among bacteria and is exacerbated by genetic engineering, potentially leading to unforeseen ecological impacts and public health challenges. Moreover, the release of genetically engineered bacteria into the environment, such as those used in agriculture or bioremediation, raises concerns about their persistence and impact on native microbial communities and ecosystem functions [73].
For fungi, genetic engineering poses risks related to ecological balance and biodiversity. Engineered fungi released into natural environments could outcompete native species, leading to reduced biodiversity and altered ecosystem dynamics [74]. Additionally, there is a risk of unintended effects due to off-target mutations, which can affect fungal physiology and interactions with other organisms [63]. These unintended changes could have cascading effects on ecosystems, particularly if engineered fungi are used in agricultural settings where they interact with a wide range of organisms.
Oomycetes, being plant pathogens, present unique risks when genetically engineered. The potential for engineered oomycetes to develop new virulence factors or overcome existing plant resistance mechanisms is a significant concern [66]. This could lead to more aggressive plant diseases, impacting agricultural productivity and food security. Additionally, the release of genetically modified oomycetes into the environment could disrupt natural plant-pathogen interactions, leading to unintended ecological consequences [60].
While genetic engineering offers significant benefits, such as improved agricultural productivity and bioremediation capabilities, the potential risks associated with genetically engineered microorganisms must be carefully managed. This includes robust risk assessments, stringent regulatory frameworks, and ongoing monitoring to mitigate negative impacts on human health and the environment [72,75].
6.2. Regulatory Frameworks
The regulation of genetically engineered microorganisms, including bacteria, fungi, and oomycetes, is governed by a complex and evolving set of guidelines that vary significantly across different regions. These regulations are designed to address the potential risks associated with the release and use of genetically modified organisms (GMOs) in various environments.
Globally, there is a lack of uniformity in the regulation of genetically engineered microorganisms. The European Union (EU) has one of the most stringent regulatory frameworks, where GMOs, including microorganisms, must undergo a rigorous approval process before being released into the environment. This process involves proving that the GMO is as safe as its conventional counterpart, both for human health and the environment [76]. The EU's regulations also require labeling of products containing GMOs, which reflects a high level of consumer protection and transparency [76].
In contrast, the regulatory landscape in the United States is more fragmented. The U.S. Department of Agriculture (USDA), Environmental Protection Agency (EPA), and Food and Drug Administration (FDA) share responsibilities for regulating genetically engineered microorganisms under various acts, including the Plant Protection Act (PPA), Federal Insecticide, Fungicide, and Rodenticide Act (FIFRA), and the Federal Food, Drug, and Cosmetic Act (FFDCA) [77]. The USDA's Animal and Plant Health Inspection Service (APHIS) regulates microorganisms that are considered plant pests, while the EPA oversees those used for pesticidal purposes [77]. This division of regulatory responsibilities can lead to complexities in the approval process but allows for specialized oversight based on the intended use of the genetically engineered organism.
The advent of genome editing technologies, such as CRISPR, has introduced new challenges for existing regulatory frameworks. Many countries are in the process of updating their regulations to address these technologies, which can create organisms with minimal genetic changes that might not fit traditional definitions of GMOs [78]. For instance, some countries differentiate between genome-edited organisms based on whether foreign DNA is present in the final product, which influences their regulatory classification [78].
The regulation of genetically engineered microorganisms is characterized by a diverse and dynamic landscape. While regions like the EU maintain strict regulatory standards, others like the U.S. adopt a more segmented approach. The rapid development of genome editing technologies continues to challenge existing regulations, necessitating ongoing revisions to ensure safety and address public concerns effectively.
7. Conclusions and Future Directions
This review has delved into the advancements and applied methodologies in genetic engineering for bacteria, fungi and oomycetes. A key focus has been on the utilization of the CRISPR system, highlighting its precision and efficacy in editing genomes to confer resistance against various pathogens. Notable successes include the transformation protocols for oomycete pathogens, which present promising avenues for future agricultural applications. Additionally, the exploration of bacterial, fungal and oomycete genomes has uncovered significant potential for creating more resilient species, contributing to sustainable agricultural practices and food security.
One promising area for future research is the refinement of CRISPR-based techniques to increase editing efficiency and specificity in microbial genomes. This involves developing novel Cas variants, optimizing delivery methods, such as nanoparticle-mediated delivery systems, to overcome existing barriers to transformation.
Another critical direction is the integration of multi-omics approaches, including genomics, transcriptomics, proteomics, and metabolomics, to gain a comprehensive understanding of host-pathogen interactions. This holistic view can reveal novel targets for genetic manipulation, potentially leading to the development of plants with enhanced disease resistance.
Moreover, exploring the potential of synthetic biology in genetic engineering offers exciting opportunities. By designing and constructing synthetic gene networks, researchers can create new metabolic pathways or regulatory circuits that can enhance desired traits. This interdisciplinary approach can also facilitate the development of custom-built organisms tailored to specific microbial cell factories.
Lastly, the ethical, environmental, and regulatory aspects of deploying genetically engineered organisms in the field warrant attention. Conducting thorough risk assessments and engaging with stakeholders, including farmers, consumers, and policymakers, will be crucial to ensure the responsible and sustainable application of these technologies. By addressing these areas, the field of genetic engineering can continue to advance, contributing significantly to sustainable agriculture and industrial biotechnology.
Author Contributions
Conceptualization, Supervision, Writing—original draft preparation, Writing—review & editing, Visualization, P.Y.; Writing—review & editing, A.C., S.S., C.H., L.L., M.S., M.A.A. All authors have read and approved the last version of the manuscript.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Paul, N.C.; Park, S.-W.; Liu, H.; Choi, S.; Ma, J.; MacCready, J.S.; Chilvers, M.I.; Sang, H. Plant and Fungal Genome Editing to Enhance Plant Disease Resistance Using the CRISPR/Cas9 System. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, F.; Liu, H.; Chen, H.; Zhang, F.; Li, S.; Sun, T.; Nekrasov, V.; Huang, S.; Dong, S. A Modified Agrobacterium-Mediated Transformation for Two Oomycete Pathogens. PLOS Pathogens 2023, 19, e1011346. [Google Scholar] [CrossRef]
- Oliveira, L.R.; Gonçalves, A.R.; Quintela, E.D.; Colognese, L.; Cortes, M.V. de C.B.; de Filippi, M.C.C. Genetic Engineering of Filamentous Fungi: Prospects for Obtaining Fourth-Generation Biological Products. Applied Microbiology 2024, 4, 794–810. [Google Scholar] [CrossRef]
- Yang, P.; Bokros, N.; Debolt, S.; Zhao, Z.; Xia, Y. Genome Sequence Source of Bacillus Amyloliquefaciens Strain GD4a, a Bacterial Endophyte Associated with Switchgrass Plants. Phytobiomes Journal 2022, 6, 354–357. [Google Scholar] [CrossRef]
- Malkawi, H.I.; Kapiel, T.Y.S. Microbial Biotechnology: A Key Tool for Addressing Climate Change and Food Insecurity. European Journal of Biology and Biotechnology 2024, 5, 1–15. [Google Scholar] [CrossRef]
- Santos-Beneit, F. What Is the Role of Microbial Biotechnology and Genetic Engineering in Medicine? Microbiologyopen 2024, 13, e1406. [Google Scholar] [CrossRef]
- Roth, M.G.; Westrick, N.M.; Baldwin, T.T. Fungal Biotechnology: From Yesterday to Tomorrow. Front. Fungal Biol. 2023, 4. [Google Scholar] [CrossRef]
- Corbu, V.M.; Gheorghe-Barbu, I.; Dumbravă, A. Ștefania; Vrâncianu, C.O.; Șesan, T.E. Current Insights in Fungal Importance—A Comprehensive Review. Microorganisms 2023, 11, 1384. [Google Scholar] [CrossRef]
- Sharma, S.; Sundaresha, S.; Bhardwaj, V. Biotechnological Approaches in Management of Oomycetes Diseases. 3 Biotech 2021, 11, 274. [Google Scholar] [CrossRef]
- Blakney, A.J.C.; Bainard, L.D.; St-Arnaud, M.; Hijri, M. Soil Chemistry and Soil History Significantly Structure Oomycete Communities in Brassicaceae Crop Rotations. Appl Environ Microbiol 89, e01314-22. [CrossRef]
- Li, T.; Yang, Y.; Qi, H.; Cui, W.; Zhang, L.; Fu, X.; He, X.; Liu, M.; Li, P.; Yu, T. CRISPR/Cas9 Therapeutics: Progress and Prospects. Sig Transduct Target Ther 2023, 8, 1–23. [Google Scholar] [CrossRef]
- Rabaan, A.A.; Al Fares, M.A.; Almaghaslah, M.; Alpakistany, T.; Al Kaabi, N.A.; Alshamrani, S.A.; Alshehri, A.A.; Almazni, I.A.; Saif, A.; Hakami, A.R.; et al. Application of CRISPR-Cas System to Mitigate Superbug Infections. Microorganisms 2023, 11, 2404. [Google Scholar] [CrossRef]
- Vialetto, E.; Miele, S.; Goren, M.G.; Yu, J.; Yu, Y.; Collias, D.; Beamud, B.; Osbelt, L.; Lourenço, M.; Strowig, T.; et al. Systematic Interrogation of CRISPR Antimicrobials in Klebsiella Pneumoniae Reveals Nuclease-, Guide- and Strain-Dependent Features Influencing Antimicrobial Activity. Nucleic Acids Research 2024, 52, 6079–6091. [Google Scholar] [CrossRef]
- Fels, U.; Gevaert, K.; Van Damme, P. Bacterial Genetic Engineering by Means of Recombineering for Reverse Genetics. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Li, R.; Li, A.; Zhang, Y.; Fu, J. The Emerging Role of Recombineering in Microbiology. Engineering Microbiology 2023, 3, 100097. [Google Scholar] [CrossRef]
- Matuszyńska, A.; Ebenhöh, O.; Zurbriggen, M.D.; Ducat, D.C.; Axmann, I.M. A New Era of Synthetic Biology—Microbial Community Design. Synthetic Biology 2024, 9, ysae011. [Google Scholar] [CrossRef]
- Sudheer, S.; Bai, R.G.; Usmani, Z.; Sharma, M. Insights on Engineered Microbes in Sustainable Agriculture: Biotechnological Developments and Future Prospects. Curr Genomics 2020, 21, 321–333. [Google Scholar] [CrossRef]
- Kamionka, M. Engineering of Therapeutic Proteins Production in Escherichia Coli. Curr Pharm Biotechnol 2011, 12, 268–274. [Google Scholar] [CrossRef]
- Niazi, S.K.; Magoola, M. Advances in Escherichia Coli-Based Therapeutic Protein Expression: Mammalian Conversion, Continuous Manufacturing, and Cell-Free Production. Biologics 2023, 3, 380–401. [Google Scholar] [CrossRef]
- Mugwanda, K.; Hamese, S.; Van Zyl, W.F.; Prinsloo, E.; Du Plessis, M.; Dicks, L.M.T.; Thimiri Govinda Raj, D.B. Recent Advances in Genetic Tools for Engineering Probiotic Lactic Acid Bacteria. Biosci Rep 2023, 43, BSR20211299. [Google Scholar] [CrossRef]
- Dell’ Anno, F.; Rastelli, E.; Sansone, C.; Brunet, C.; Ianora, A.; Dell’ Anno, A. Bacteria, Fungi and Microalgae for the Bioremediation of Marine Sediments Contaminated by Petroleum Hydrocarbons in the Omics Era. Microorganisms 2021, 9, 1695. [Google Scholar] [CrossRef]
- Bala, S.; Garg, D.; Thirumalesh, B.V.; Sharma, M.; Sridhar, K.; Inbaraj, B.S.; Tripathi, M. Recent Strategies for Bioremediation of Emerging Pollutants: A Review for a Green and Sustainable Environment. Toxics 2022, 10, 484. [Google Scholar] [CrossRef] [PubMed]
- Garg, D.; Samota, M.K.; Kontis, N.; Patel, N.; Bala, S.; Rosado, A.S. Revolutionizing Biofuel Generation: Unleashing the Power of CRISPR-Cas Mediated Gene Editing of Extremophiles. Microbiological Research 2023, 274, 127443. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Liu, D.; Jia, X.; Zheng, Y.; Liu, W.; Xiao, Y. CRISPR-Cas9/Cas12a Biotechnology and Application in Bacteria. Synthetic and Systems Biotechnology 2018, 3, 135–149. [Google Scholar] [CrossRef]
- Adegboye, M.F.; Ojuederie, O.B.; Talia, P.M.; Babalola, O.O. Bioprospecting of Microbial Strains for Biofuel Production: Metabolic Engineering, Applications, and Challenges. Biotechnology for Biofuels 2021, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Fong, S.S. Challenges and Advances for Genetic Engineering of Non-Model Bacteria and Uses in Consolidated Bioprocessing. Front Microbiol 2017, 8, 2060. [Google Scholar] [CrossRef]
- Egbert, R.; Elmore, J.; Guss, A. Taming Undomesticated Bacteria with a High-Efficiency Genome Engineering Tool Available online: https://www.energy.gov/science/ber/articles/taming-undomesticated-bacteria-high-efficiency-genome-engineering-tool (accessed on 26 August 2024). (accessed on null).
- Zhao, Z.; Bokros, N.; DeBolt, S.; Yang, P.; Xia, Y. Genome Sequence Resource of Bacillus Sp. RRD69, a Beneficial Bacterial Endophyte Isolated from Switchgrass Plants. MPMI 2021, 34, 1320–1323. [Google Scholar] [CrossRef]
- Yang, P.; Yuan, P.; Liu, W.; Zhao, Z.; Bernier, M.C.; Zhang, C.; Adhikari, A.; Opiyo, S.O.; Zhao, L.; Banks, F.; et al. Plant Growth Promotion and Plant Disease Suppression Induced by Bacillus Amyloliquefaciens Strain GD4a. Plants 2024, 13, 672. [Google Scholar] [CrossRef] [PubMed]
- Yang, P. Exploring Plant-Microbe Interactions through the Lens of Beneficial Bacteria, The Ohio State University, 2023.
- Sahib, M.R.; Yang, P.; Bokros, N.; Shapiro, N.; Woyke, T.; Kyrpides, N.C.; Xia, Y.; DeBolt, S. Improved Draft Genome Sequence of Microbacterium Sp. Strain LKL04, a Bacterial Endophyte Associated with Switchgrass Plants. Microbiol Resour Announc 2019, 8. [Google Scholar] [CrossRef]
- Rodriguez-Iglesias, A.; Schmoll, M. Protoplast Transformation for Genome Manipulation in Fungi. In Genetic Transformation Systems in Fungi, Volume 1; van den Berg, M.A., Maruthachalam, K., Eds.; Springer International Publishing: Cham, 2015; pp. 21–40. ISBN 978-3-319-10142-2. [Google Scholar]
- Zhang, Q.; Zhao, L.; Shen, M.; Liu, J.; Li, Y.; Xu, S.; Chen, L.; Shi, G.; Ding, Z. Establishment of an Efficient Polyethylene Glycol (PEG)-Mediated Transformation System in Pleurotus Eryngii Var. Ferulae Using Comprehensive Optimization and Multiple Endogenous Promoters. J Fungi (Basel) 2022, 8, 186. [Google Scholar] [CrossRef]
- Su, Z.-Z.; Dai, M.-D.; Zhu, J.-N.; Zeng, Y.-L.; Lu, X.-J.; Liu, X.-H.; Lin, F.-C. An Efficient Genetic Manipulation Protocol for Dark Septate Endophyte Falciphora Oryzae. Biotechnol Lett 2021, 43, 2045–2052. [Google Scholar] [CrossRef]
- Li, G.; Li, R.; Liu, Q.; Wang, Q.; Chen, M.; Li, B. A Highly Efficient Polyethylene Glycol-Mediated Transformation Method for Mushrooms. FEMS Microbiology Letters 2006, 256, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Sbaraini, N.; Tomazett, M.V.; Penteriche, A.B.; Gonçales, R.A.; Camargo, M. da S.; Bailão, A.M.; Borges, C.L.; Schrank, A.; Soares, C.M. de A.; Staats, C.C. An Efficient Agrobacterium Tumefaciens-Mediated Transformation Method for Simplicillium Subtropicum (Hypocreales: Cordycipitaceae). Genet Mol Biol 2021, 44, e20210073. [Google Scholar] [CrossRef] [PubMed]
- Ropero-Pérez, C.; Manzanares, P.; Marcos, J.F.; Garrigues, S. Agrobacterium Tumefaciens-Mediated Transformation for the Genetic Modification of the Biotechnologically Relevant Fungus Aspergillus Vadensis through Synthetic Biology. Current Research in Biotechnology 2024, 7, 100178. [Google Scholar] [CrossRef]
- Combier, J.-P.; Melayah, D.; Raffier, C.; Gay, G.; Marmeisse, R. Agrobacterium Tumefaciens-Mediated Transformation as a Tool for Insertional Mutagenesis in the Symbiotic Ectomycorrhizal Fungus Hebeloma Cylindrosporum. FEMS Microbiology Letters 2003, 220, 141–148. [Google Scholar] [CrossRef]
- Wei, T.-Y.; Wu, Y.-J.; Xie, Q.-P.; Tang, J.-W.; Yu, Z.-T.; Yang, S.-B.; Chen, S.-X. CRISPR/Cas9-Based Genome Editing in the Filamentous Fungus Glarea Lozoyensis and Its Application in Manipulating gloF. ACS Synth. Biol. 2020, 9, 1968–1977. [Google Scholar] [CrossRef] [PubMed]
- Wang, P. Two Distinct Approaches for CRISPR-Cas9-Mediated Gene Editing in Cryptococcus Neoformans and Related Species. mSphere 2018, 3. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, S.; Yang, L.; Zhang, Q. Application Progress of CRISPR/Cas9 Genome-Editing Technology in Edible Fungi. Front Microbiol 2023, 14, 1169884. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.; Valiante, V.; Nødvig, C.S.; Mattern, D.J.; Slotkowski, R.A.; Mortensen, U.H.; Brakhage, A.A. Functional Reconstitution of a Fungal Natural Product Gene Cluster by Advanced Genome Editing. ACS Synth. Biol. 2017, 6, 62–68. [Google Scholar] [CrossRef]
- Wang, D.; Jin, S.; Lu, Q.; Chen, Y. Advances and Challenges in CRISPR/Cas-Based Fungal Genome Engineering for Secondary Metabolite Production: A Review. J Fungi (Basel) 2023, 9, 362. [Google Scholar] [CrossRef]
- Michaliski, L.F.; Ióca, L.P.; Oliveira, L.S.; Crnkovic, C.M.; Takaki, M.; Freire, V.F.; Berlinck, R.G.S. Improvement of Targeted Fungi Secondary Metabolite Production Using a Systematic Experimental Design and Chemometrics Analysis. Methods and Protocols 2023, 6, 77. [Google Scholar] [CrossRef]
- van Esse, H.P.; Reuber, T.L.; van der Does, D. Genetic Modification to Improve Disease Resistance in Crops. New Phytologist 2020, 225, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Mekapogu, M.; Jung, J.-A.; Kwon, O.-K.; Ahn, M.-S.; Song, H.-Y.; Jang, S. Recent Progress in Enhancing Fungal Disease Resistance in Ornamental Plants. Int J Mol Sci 2021, 22, 7956. [Google Scholar] [CrossRef] [PubMed]
- Maini Rekdal, V.; van der Luijt, C.R.B.; Chen, Y.; Kakumanu, R.; Baidoo, E.E.K.; Petzold, C.J.; Cruz-Morales, P.; Keasling, J.D. Edible Mycelium Bioengineered for Enhanced Nutritional Value and Sensory Appeal Using a Modular Synthetic Biology Toolkit. Nat Commun 2024, 15, 2099. [Google Scholar] [CrossRef]
- Wang, Q.; Zhong, C.; Xiao, H. Genetic Engineering of Filamentous Fungi for Efficient Protein Expression and Secretion. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef]
- Ropero-Pérez, C.; Marcos, J.F.; Manzanares, P.; Garrigues, S. Increasing the Efficiency of CRISPR/Cas9-Mediated Genome Editing in the Citrus Postharvest Pathogen Penicillium Digitatum. Fungal Biology and Biotechnology 2024, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Hulbert, S.H. Host-Induced Gene Silencing (HIGS) for Elucidating Puccinia Gene Function in Wheat. In Plant Pathogenic Fungi and Oomycetes: Methods and Protocols; Ma, W., Wolpert, T., Eds.; Springer: New York, NY, 2018; pp. 139–150. ISBN 978-1-4939-8724-5. [Google Scholar]
- Feng, H.; Liu, T.; Li, J.; Wan, C.; Ding, F.; Wang, Y.; Zheng, X.; Ye, W. Gene Editing with an Oxathiapiprolin Resistance Selection Marker Reveals That PuLLP, a Loricrin-like Protein, Is Required for Oospore Development in Pythium Ultimum. Phytopathology Research 2023, 5, 34. [Google Scholar] [CrossRef]
- Lamour, K.H.; Win, J.; Kamoun, S. Oomycete Genomics: New Insights and Future Directions. FEMS Microbiol Lett 2007, 274, 1–8. [Google Scholar] [CrossRef]
- Vink, J.N.A.; Hayhurst, M.; Gerth, M.L. Harnessing CRISPR-Cas for Oomycete Genome Editing. Trends Microbiol 2023, 31, 947–958. [Google Scholar] [CrossRef]
- Wang, W.; Xue, Z.; Miao, J.; Cai, M.; Zhang, C.; Li, T.; Zhang, B.; Tyler, B.M.; Liu, X. PcMuORP1, an Oxathiapiprolin-Resistance Gene, Functions as a Novel Selection Marker for Phytophthora Transformation and CRISPR/Cas9 Mediated Genome Editing. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Ah-Fong, A.M.V.; Boyd, A.M.; Matson, M.E.H.; Judelson, H.S. A Cas12a-Based Gene Editing System for Phytophthora Infestans Reveals Monoallelic Expression of an Elicitor. Molecular Plant Pathology 2021, 22, 737–752. [Google Scholar] [CrossRef]
- Mendoza, C.S.; Findlay, A.; Judelson, H.S. A Variant of LbCas12a and Elevated Incubation Temperatures Enhance the Rate of Gene Editing in the Oomycete Phytophthora Infestans. MPMI 2023, 36, 677–681. [Google Scholar] [CrossRef]
- Verhoeven, T.; Pluis, M.H.; Peippo, M.; Couillaud, G.; Berg, G.C. van den; Evangelisti, E. LbCas12-Mediated Multiplex Gene Editing and 2-Fluoroadenine Counter-Selection in Phytophthora Palmivora 2024, 2024. 02.13.58 0060.
- Langner, T.; Kamoun, S.; Belhaj, K. CRISPR Crops: Plant Genome Editing Toward Disease Resistance. Annual Review of Phytopathology 2018, 56, 479–512. [Google Scholar] [CrossRef]
- Borrelli, V.M.G.; Brambilla, V.; Rogowsky, P.; Marocco, A.; Lanubile, A. The Enhancement of Plant Disease Resistance Using CRISPR/Cas9 Technology. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Kamoun, S. Molecular Genetics of Pathogenic Oomycetes. Eukaryot Cell 2003, 2, 191–199. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Karlin, S.; Mrázek, J. Predicted Highly Expressed Genes of Diverse Prokaryotic Genomes. Journal of Bacteriology 2000, 182, 5238–5250. [Google Scholar] [CrossRef]
- Jin, F.-J.; Hu, S.; Wang, B.-T.; Jin, L. Advances in Genetic Engineering Technology and Its Application in the Industrial Fungus Aspergillus Oryzae. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Yılmaz, Ş.G. Genome Editing Technologies: CRISPR, LEAPER, RESTORE, ARCUT, SATI, and RESCUE. EXCLI Journal 2021, 20, 19–45. [Google Scholar] [CrossRef]
- Capriotti, L.; Baraldi, E.; Mezzetti, B.; Limera, C.; Sabbadini, S. Biotechnological Approaches: Gene Overexpression, Gene Silencing, and Genome Editing to Control Fungal and Oomycete Diseases in Grapevine. International Journal of Molecular Sciences 2020, 21, 5701. [Google Scholar] [CrossRef]
- Ghimire, B.; Saraiva, M.; Andersen, C.B.; Gogoi, A.; Saleh, M.; Zic, N.; van West, P.; Brurberg, M.B. Transformation Systems, Gene Silencing and Gene Editing Technologies in Oomycetes. Fungal Biology Reviews 2022, 40, 37–52. [Google Scholar] [CrossRef]
- Kamoun, S.; Huitema, E.; Vleeshouwers, V.G. Resistance to Oomycetes: A General Role for the Hypersensitive Response? Trends Plant Sci 1999, 4, 196–200. [Google Scholar] [CrossRef]
- Pedrolli, D.B.; Ribeiro, N.V.; Squizato, P.N.; de Jesus, V.N.; Cozetto, D.A.; Tuma, R.B.; Gracindo, A.; Cesar, M.B.; Freire, P.J.C.; da Costa, A.F.M.; et al. Engineering Microbial Living Therapeutics: The Synthetic Biology Toolbox. Trends in Biotechnology 2019, 37, 100–115. [Google Scholar] [CrossRef]
- Peters, J.M.; Koo, B.-M.; Patino, R.; Heussler, G.E.; Hearne, C.C.; Qu, J.; Inclan, Y.F.; Hawkins, J.S.; Lu, C.H.S.; Silvis, M.R.; et al. Enabling Genetic Analysis of Diverse Bacteria with Mobile-CRISPRi. Nature Microbiology 2019, 4, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Arnold, J.; Glazier, J.; Mimee, M. Genetic Engineering of Resident Bacteria in the Gut Microbiome. J Bacteriol 205, e00127-23. [CrossRef]
- Pérez, G.; Lopez-Moya, F.; Chuina, E.; Ibañez-Vea, M.; Garde, E.; López-Llorca, L.V.; Pisabarro, A.G.; Ramírez, L. Strain Degeneration in Pleurotus Ostreatus: A Genotype Dependent Oxidative Stress Process Which Triggers Oxidative Stress, Cellular Detoxifying and Cell Wall Reshaping Genes. Journal of Fungi 2021, 7, 862. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.S.; Melo, E.O. Animal Transgenesis and Cloning: Combined Development and Future Perspectives. Methods Mol Biol 2023, 2647, 121–149. [Google Scholar] [CrossRef]
- Yaashikaa, P.R.; Devi, M.K.; Kumar, P.S. Engineering Microbes for Enhancing the Degradation of Environmental Pollutants: A Detailed Review on Synthetic Biology. Environmental Research 2022, 214, 113868. [Google Scholar] [CrossRef]
- Pant, G.; Garlapati, D.; Agrawal, U.; Prasuna, R.G.; Mathimani, T.; Pugazhendhi, A. Biological Approaches Practised Using Genetically Engineered Microbes for a Sustainable Environment: A Review. J Hazard Mater 2021, 405, 124631. [Google Scholar] [CrossRef]
- Bauer-Panskus, A.; Miyazaki, J.; Kawall, K.; Then, C. Risk Assessment of Genetically Engineered Plants That Can Persist and Propagate in the Environment. Environmental Sciences Europe 2020, 32, 32. [Google Scholar] [CrossRef]
- Green Genetic Engineering Law Available online: https://www.leopoldina.org/en/topics/green-genetic-engineering/green-genetic-engineering-law/ (accessed on 27 August 2024). (accessed on null).
- Hanlon, P.; Sewalt, V. GEMs: Genetically Engineered Microorganisms and the Regulatory Oversight of Their Uses in Modern Food Production. Critical Reviews in Food Science and Nutrition 2021, 61, 959–970. [Google Scholar] [CrossRef]
- Sprink, T.; Wilhelm, R. Genome Editing in Biotech Regulations Worldwide. In A Roadmap for Plant Genome Editing; Ricroch, A., Eriksson, D., Miladinović, D., Sweet, J., Van Laere, K., Woźniak-Gientka, E., Eds.; Springer Nature Switzerland: Cham, 2024; pp. 425–435. ISBN 978-3-031-46150-7. [Google Scholar]
- Vos, P.D.; Rossetti, G.; Mantegna, J.L.; Siira, S.J.; Gandadireja, A.P.; Bruce, M.; Raven, S.A.; Khersonsky, O.; Fleishman, S.J.; Filipovska, A.; et al. Computationally Designed Hyperactive Cas9 Enzymes. Nat Commun 2022, 13, 3023. [Google Scholar] [CrossRef] [PubMed]
- Cloud-Based Platform for Biotech R&D | Benchling. Available online: https://www.benchling.com (accessed on 2 September 2024).
- Bae, S.; Park, J.; Kim, J.-S. Cas-OFFinder: A Fast and Versatile Algorithm That Searches for Potential off-Target Sites of Cas9 RNA-Guided Endonucleases. Bioinformatics 2014, 30, 1473–1475. [Google Scholar] [CrossRef] [PubMed]
- Labuhn, M.; Adams, F.F.; Ng, M.; Knoess, S.; Schambach, A.; Charpentier, E.M.; Schwarzer, A.; Mateo, J.L.; Klusmann, J.-H.; Heckl, D. Refined sgRNA Efficacy Prediction Improves Large- and Small-Scale CRISPR–Cas9 Applications. Nucleic Acids Research 2018, 46, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Stemmer, M.; Thumberger, T.; Keyer, M. del S.; Wittbrodt, J.; Mateo, J.L. CCTop: An Intuitive, Flexible and Reliable CRISPR/Cas9 Target Prediction Tool. PLOS ONE 2015, 10, e0124633. [Google Scholar] [CrossRef]
- Labun, K.; Montague, T.G.; Krause, M.; Torres Cleuren, Y.N.; Tjeldnes, H.; Valen, E. CHOPCHOP v3: Expanding the CRISPR Web Toolbox beyond Genome Editing. Nucleic Acids Research 2019, 47, W171–W174. [Google Scholar] [CrossRef] [PubMed]
- Concordet, J.-P.; Haeussler, M. CRISPOR: Intuitive Guide Selection for CRISPR/Cas9 Genome Editing Experiments and Screens. Nucleic Acids Research 2018, 46, W242–W245. [Google Scholar] [CrossRef]
- Naito, Y.; Hino, K.; Bono, H.; Ui-Tei, K. CRISPRdirect: Software for Designing CRISPR/Cas Guide RNA with Reduced off-Target Sites. Bioinformatics 2015, 31, 1120–1123. [Google Scholar] [CrossRef]
- Liu, H.; Wei, Z.; Dominguez, A.; Li, Y.; Wang, X.; Qi, L.S. CRISPR-ERA: A Comprehensive Design Tool for CRISPR-Mediated Gene Editing, Repression and Activation. Bioinformatics 2015, 31, 3676–3678. [Google Scholar] [CrossRef]
- CRISPick Available online: https://portals.broadinstitute.org/gppx/crispick/public (accessed on 2 September 2024).
- CRISPR-Cas9 Guide RNA Design Checker | IDT Available online: https://www.idtdna.com/site/order/designtool/index/CRISPR_SEQUENCE (accessed on 2 September 2024).
- Pliatsika, V.; Rigoutsos, I. “Off-Spotter”: Very Fast and Exhaustive Enumeration of Genomic Lookalikes for Designing CRISPR/Cas Guide RNAs. Biology Direct 2015, 10, 4. [Google Scholar] [CrossRef]
- Enzmann, B.L.; Wronski, A. Synthego’s Engineered Cells Allow Scientists to “Cut Out” CRISPR Optimization [SPONSORED]. The CRISPR Journal 2018, 1, 255–257. [Google Scholar] [CrossRef]
- Patel, A.; Iannello, G.; Diaz, A.G.; Sirabella, D.; Thaker, V.; Corneo, B. Efficient Cas9-Based Genome Editing Using CRISPR Analysis Webtools in Severe Early-Onset-Obesity Patient-Derived iPSCs. Current Protocols 2022, 2, e519. [Google Scholar] [CrossRef]
- Yamada, S.; Suzuki, Y.; Kouzuma, A.; Watanabe, K. Development of a CRISPR Interference System for Selective Gene Knockdown in Acidithiobacillus Ferrooxidans. Journal of Bioscience and Bioengineering 2022, 133, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Schultenkämper, K.; Brito, L.F.; López, M.G.; Brautaset, T.; Wendisch, V.F. Establishment and Application of CRISPR Interference to Affect Sporulation, Hydrogen Peroxide Detoxification, and Mannitol Catabolism in the Methylotrophic Thermophile Bacillus Methanolicus. Appl Microbiol Biotechnol 2019, 103, 5879–5889. [Google Scholar] [CrossRef] [PubMed]
- Mougiakos, I.; Mohanraju, P.; Bosma, E.F.; Vrouwe, V.; Finger Bou, M.; Naduthodi, M.I.S.; Gussak, A.; Brinkman, R.B.L.; van Kranenburg, R.; van der Oost, J. Characterizing a Thermostable Cas9 for Bacterial Genome Editing and Silencing. Nat Commun 2017, 8, 1647. [Google Scholar] [CrossRef] [PubMed]
- Mougiakos, I.; Bosma, E.F.; Weenink, K.; Vossen, E.; Goijvaerts, K.; van der Oost, J.; van Kranenburg, R. Efficient Genome Editing of a Facultative Thermophile Using Mesophilic spCas9. ACS Synth. Biol. 2017, 6, 849–861. [Google Scholar] [CrossRef]
- Altenbuchner, J. Editing of the Bacillus Subtilis Genome by the CRISPR-Cas9 System. Applied and Environmental Microbiology 2016, 82, 5421–5427. [Google Scholar] [CrossRef]
- Wasels, F.; Jean-Marie, J.; Collas, F.; López-Contreras, A.M.; Lopes Ferreira, N. A Two-Plasmid Inducible CRISPR/Cas9 Genome Editing Tool for Clostridium Acetobutylicum. Journal of Microbiological Methods 2017, 140, 5–11. [Google Scholar] [CrossRef]
- Nagaraju, S.; Davies, N.K.; Walker, D.J.F.; Köpke, M.; Simpson, S.D. Genome Editing of Clostridium Autoethanogenum Using CRISPR/Cas9. Biotechnol Biofuels 2016, 9, 219. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.-T.; Seo, S.-O.; Lynn, P.; Lu, T.; Jin, Y.-S.; Blaschek, H.P. Bacterial Genome Editing with CRISPR-Cas9: Deletion, Integration, Single Nucleotide Modification, and Desirable “Clean” Mutant Selection in Clostridium Beijerinckii as an Example. ACS Synth. Biol. 2016, 5, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Li, Y.; Shi, Z.; Hemme, C.L.; Li, Y.; Zhu, Y.; Van Nostrand, J.D.; He, Z.; Zhou, J. Efficient Genome Editing in Clostridium Cellulolyticum via CRISPR-Cas9 Nickase. Applied and Environmental Microbiology 2015, 81, 4423–4431. [Google Scholar] [CrossRef]
- Cleto, S.; Jensen, J.V.; Wendisch, V.F.; Lu, T.K. Corynebacterium Glutamicum Metabolic Engineering with CRISPR Interference (CRISPRi). ACS Synth. Biol. 2016, 5, 375–385. [Google Scholar] [CrossRef]
- Woolston, B.M.; Emerson, D.F.; Currie, D.H.; Stephanopoulos, G. Rediverting Carbon Flux in Clostridium Ljungdahlii Using CRISPR Interference (CRISPRi). Metabolic Engineering 2018, 48, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Pyne, M.E.; Bruder, M.R.; Moo-Young, M.; Chung, D.A.; Chou, C.P. Harnessing Heterologous and Endogenous CRISPR-Cas Machineries for Efficient Markerless Genome Editing in Clostridium. Sci Rep 2016, 6, 25666. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.E.; Lanahan, A.A.; Zheng, T.; Toruno, C.; Lynd, L.R.; Cameron, J.C.; Olson, D.G.; Eckert, C.A. Development of Both Type I–B and Type II CRISPR/Cas Genome Editing Systems in the Cellulolytic Bacterium Clostridium Thermocellum. Metabolic Engineering Communications 2020, 10, e00116. [Google Scholar] [CrossRef]
- Zhang, J.; Zong, W.; Hong, W.; Zhang, Z.-T.; Wang, Y. Exploiting Endogenous CRISPR-Cas System for Multiplex Genome Editing in Clostridium Tyrobutyricum and Engineer the Strain for High-Level Butanol Production. Metabolic Engineering 2018, 47, 49–59. [Google Scholar] [CrossRef]
- Turco, F.; Garavaglia, M.; Van Houdt, R.; Hill, P.; Rawson, F.J.; Kovacs, K. Synthetic Biology Toolbox, Including a Single-Plasmid CRISPR-Cas9 System to Biologically Engineer the Electrogenic, Metal-Resistant Bacterium Cupriavidus Metallidurans CH34. ACS Synthetic Biology 2022, 11, 3617–3628. [Google Scholar] [CrossRef] [PubMed]
- Xiong, B.; Li, Z.; Liu, L.; Zhao, D.; Zhang, X.; Bi, C. Genome Editing of Ralstonia Eutropha Using an Electroporation-Based CRISPR-Cas9 Technique. Biotechnol Biofuels 2018, 11, 172. [Google Scholar] [CrossRef]
- Jiang, W.; Bikard, D.; Cox, D.; Zhang, F.; Marraffini, L.A. RNA-Guided Editing of Bacterial Genomes Using CRISPR-Cas Systems. Nat Biotechnol 2013, 31, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Rath, D.; Amlinger, L.; Hoekzema, M.; Devulapally, P.R.; Lundgren, M. Efficient Programmable Gene Silencing by Cascade. Nucleic Acids Research 2015, 43, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.; Ling, C.; Zhao, Y.; Yang, T.; Yin, J.; Guo, Y.; Chen, G.Q. CRISPR/Cas9 Editing Genome of Extremophile Halomonas Spp. Metabolic Engineering 2018, 47, 219–229. [Google Scholar] [CrossRef]
- Ganguly, J.; Martin-Pascual, M.; van Kranenburg, R. CRISPR Interference (CRISPRi) as Transcriptional Repression Tool for Hungateiclostridium Thermocellum DSM 1313. Microbial Biotechnology 2020, 13, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Baik, J.; Lim, S.-H. A Model Independent S/W Framework for Search-Based Software Testing. The Scientific World Journal 2014, 2014, 126348. [Google Scholar] [CrossRef] [PubMed]
- Mo, X.-H.; Zhang, H.; Wang, T.-M.; Zhang, C.; Zhang, C.; Xing, X.-H.; Yang, S. Establishment of CRISPR Interference in Methylorubrum Extorquens and Application of Rapidly Mining a New Phytoene Desaturase Involved in Carotenoid Biosynthesis. Appl Microbiol Biotechnol 2020, 104, 4515–4532. [Google Scholar] [CrossRef]
- Batianis, C.; Kozaeva, E.; Damalas, S.G.; Martín-Pascual, M.; Volke, D.C.; Nikel, P.I.; Martins dos Santos, V.A.P. An Expanded CRISPRi Toolbox for Tunable Control of Gene Expression in Pseudomonas Putida. Microbial Biotechnology 2020, 13, 368–385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yuan, J. CRISPR/Cas12a-Mediated Genome Engineering in the Photosynthetic Bacterium Rhodobacter Capsulatus. Microbial Biotechnology 2021, 14, 2700–2710. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Jiao, S.; Wang, M.; Yu, H.; Shen, Z. A CRISPR/Cas9-Based Genome Editing System for Rhodococcus Ruber TH. Metabolic Engineering 2020, 57, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Cobb, R.E.; Wang, Y.; Zhao, H. High-Efficiency Multiplex Genome Editing of Streptomyces Species Using an Engineered CRISPR/Cas System. ACS Synth. Biol. 2015, 4, 723–728. [Google Scholar] [CrossRef]
- Tong, Y.; Charusanti, P.; Zhang, L.; Weber, T.; Lee, S.Y. CRISPR-Cas9 Based Engineering of Actinomycetal Genomes. ACS Synth. Biol. 2015, 4, 1020–1029. [Google Scholar] [CrossRef]
- Choi, S.Y.; Woo, H.M. CRISPRi-dCas12a: A dCas12a-Mediated CRISPR Interference for Repression of Multiple Genes and Metabolic Engineering in Cyanobacteria. ACS Synth. Biol. 2020, 9, 2351–2361. [Google Scholar] [CrossRef]
- Gordon, G.C.; Korosh, T.C.; Cameron, J.C.; Markley, A.L.; Begemann, M.B.; Pfleger, B.F. CRISPR Interference as a Titratable, Trans-Acting Regulatory Tool for Metabolic Engineering in the Cyanobacterium Synechococcus Sp. Strain PCC 7002. Metabolic Engineering 2016, 38, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, B.; Duan, C.; Sun, B.; Yang, J.; Yang, S. Multigene Editing in the Escherichia Coli Genome via the CRISPR-Cas9 System. Applied and Environmental Microbiology 2015, 81, 2506–2514. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Ostrov, N.; Wong, B.G.; Gold, M.A.; Khalil, A.S.; Church, G.M. Functional Genomics of the Rapidly Replicating Bacterium Vibrio Natriegens by CRISPRi. Nat Microbiol 2019, 4, 1105–1113. [Google Scholar] [CrossRef]
- Nødvig, C.S.; Nielsen, J.B.; Kogle, M.E.; Mortensen, U.H. A CRISPR-Cas9 System for Genetic Engineering of Filamentous Fungi. PLOS ONE 2015, 10, e0133085. [Google Scholar] [CrossRef] [PubMed]
- Waltz, E. Gene-Edited CRISPR Mushroom Escapes US Regulation. Nature News 2016, 532, 293. [Google Scholar] [CrossRef]
- Nargesi, S.; Kaboli, S.; Thekkiniath, J.; Heidari, S.; Keramati, F.; Seyedmousavi, S.; Hedayati, M.T. Recent Advances in Genome Editing Tools in Medical Mycology Research. J Fungi (Basel) 2021, 7, 257. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Y.; Liu, T.; Zhang, H.; Zhang, H.; Li, J. The GlaA Signal Peptide Substantially Increases the Expression and Secretion of α-Galactosidase in Aspergillus Niger. Biotechnol Lett 2018, 40, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, D.I.; Olsthoorn, M.M.A.; Maillet, I.; Akeroyd, M.; Breestraat, S.; Donkers, S.; van der Hoeven, R.A.M.; van den Hondel, C.A.M.J.J.; Kooistra, R.; Lapointe, T.; et al. Effective Lead Selection for Improved Protein Production in Aspergillus Niger Based on Integrated Genomics. Fungal Genetics and Biology 2009, 46, S141–S152. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, W.; Sims, A.H.; Zhao, C.; Wang, A.; Tang, G.; Qin, J.; Wang, H. Isolation of Four Pepsin-like Protease Genes from Aspergillus Niger and Analysis of the Effect of Disruptions on Heterologous Laccase Expression. Fungal Genetics and Biology 2008, 45, 17–27. [Google Scholar] [CrossRef]
- Fiedler, M.R.M.; Barthel, L.; Kubisch, C.; Nai, C.; Meyer, V. Construction of an Improved Aspergillus Niger Platform for Enhanced Glucoamylase Secretion. Microbial Cell Factories 2018, 17, 95. [Google Scholar] [CrossRef]
- Ohno, A.; Maruyama, J.; Nemoto, T.; Arioka, M.; Kitamoto, K. A Carrier Fusion Significantly Induces Unfolded Protein Response in Heterologous Protein Production by Aspergillus Oryzae. Appl Microbiol Biotechnol 2011, 92, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Hoang, H.-D.; Maruyama, J.; Kitamoto, K. Modulating Endoplasmic Reticulum-Golgi Cargo Receptors for Improving Secretion of Carrier-Fused Heterologous Proteins in the Filamentous Fungus Aspergillus Oryzae. Applied and Environmental Microbiology 2015, 81, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Vyas, V.K.; Barrasa, M.I.; Fink, G.R. A Candida Albicans CRISPR System Permits Genetic Engineering of Essential Genes and Gene Families. Science Advances 2015, 1, e1500248. [Google Scholar] [CrossRef]
- Tsai, S.-C.; Tsai, L.-D.; Li, Y.-K. An Isolated Candida Albicans TL3 Capable of Degrading Phenol at Large Concentration. Bioscience, Biotechnology, and Biochemistry 2005, 69, 2358–2367. [Google Scholar] [CrossRef]
- Min, K.; Ichikawa, Y.; Woolford, C.A.; Mitchell, A.P. Candida Albicans Gene Deletion with a Transient CRISPR-Cas9 System. mSphere 2016, 1. [Google Scholar] [CrossRef]
- Yang, P.; Chen, Y.; Wu, H.; Fang, W.; Liang, Q.; Zheng, Y.; Olsson, S.; Zhang, D.; Zhou, J.; Wang, Z.; et al. The 5-Oxoprolinase Is Required for Conidiation, Sexual Reproduction, Virulence and Deoxynivalenol Production of Fusarium Graminearum. Curr Genet 2018, 64, 285–301. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.-Q.; Gao, J.; Wang, W.-J.; Wang, K.-F.; Xu, G.-Q.; Huang, H.; Ji, X.-J. CRISPR/Cas9-Based Genome Editing in the Filamentous Fungus Fusarium Fujikuroi and Its Application in Strain Engineering for Gibberellic Acid Production. ACS Synth. Biol. 2019, 8, 445–454. [Google Scholar] [CrossRef]
- Horwitz, S.B.; Goldman, I.D. A Conversation with Susan Band Horwitz. Annual Review of Pharmacology and Toxicology 2015, 55, 1–9. [Google Scholar] [CrossRef]
- Löbs, A.-K.; Engel, R.; Schwartz, C.; Flores, A.; Wheeldon, I. CRISPR–Cas9-Enabled Genetic Disruptions for Understanding Ethanol and Ethyl Acetate Biosynthesis in Kluyveromyces Marxianus. Biotechnol Biofuels 2017, 10, 164. [Google Scholar] [CrossRef]
- Chen, X.; Selvaraj, P.; Lin, L.; Fang, W.; Wu, C.; Yang, P.; Zhang, J.; Abubakar, Y.S.; Yang, F.; Lu, G.; et al. Rab7/Retromer-Based Endolysosomal Trafficking Is Essential for Proper Host Invasion in Rice Blast. New Phytologist 2023, 239. [Google Scholar] [CrossRef]
- Liu, W.; An, C.; Shu, X.; Meng, X.; Yao, Y.; Zhang, J.; Chen, F.; Xiang, H.; Yang, S.; Gao, X.; et al. A Dual-Plasmid CRISPR/Cas System for Mycotoxin Elimination in Polykaryotic Industrial Fungi. ACS Synth. Biol. 2020, 9, 2087–2095. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Gao, R.; Li, J.; Lin, L.; Zhao, J.; Sun, W.; Tian, C. Development of a Genome-Editing CRISPR/Cas9 System in Thermophilic Fungal Myceliophthora Species and Its Application to Hyper-Cellulase Production Strain Engineering. Biotechnology for Biofuels 2017, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Reilly, M.C.; Qin, L.; Craig, J.P.; Starr, T.L.; Glass, N.L. Deletion of Homologs of the SREBP Pathway Results in Hyper-Production of Cellulases in Neurospora Crassa and Trichoderma Reesei. Biotechnology for Biofuels 2015, 8, 121. [Google Scholar] [CrossRef]
- Qin, L.; Wu, V.W.; Glass, N.L. Deciphering the Regulatory Network between the SREBP Pathway and Protein Secretion in Neurospora Crassa. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Pohl, C.; Kiel, J.A.K.W.; Driessen, A.J.M.; Bovenberg, R.A.L.; Nygård, Y. CRISPR/Cas9 Based Genome Editing of Penicillium Chrysogenum. ACS Synth. Biol. 2016, 5, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Weninger, A.; Hatzl, A.-M.; Schmid, C.; Vogl, T.; Glieder, A. Combinatorial Optimization of CRISPR/Cas9 Expression Enables Precision Genome Engineering in the Methylotrophic Yeast Pichia Pastoris. Journal of Biotechnology 2016, 235, 139–149. [Google Scholar] [CrossRef]
- Zhong, Y.; Liu, X.; Xiao, P.; Wei, S.; Wang, T. Expression and Secretion of the Human Erythropoietin Using an Optimized Cbh1 Promoter and the Native CBH I Signal Sequence in the Industrial Fungus Trichoderma Reesei. Appl Biochem Biotechnol 2011, 165, 1169–1177. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, X.; Xue, X.; Luo, H.; Yao, B.; Xie, X.; Su, X. Overexpressing Key Component Genes of the Secretion Pathway for Enhanced Secretion of an Aspergillus Niger Glucose Oxidase in Trichoderma Reesei. Enzyme and Microbial Technology 2017, 106, 83–87. [Google Scholar] [CrossRef]
- Ronda, C.; Maury, J.; Jakočiu̅nas, T.; Baallal Jacobsen, S.A.; Germann, S.M.; Harrison, S.J.; Borodina, I.; Keasling, J.D.; Jensen, M.K.; Nielsen, A.T. CrEdit: CRISPR Mediated Multi-Loci Gene Integration in Saccharomyces Cerevisiae. Microb Cell Fact 2015, 14, 97. [Google Scholar] [CrossRef]
- Mans, R.; van Rossum, H.M.; Wijsman, M.; Backx, A.; Kuijpers, N.G.A.; van den Broek, M.; Daran-Lapujade, P.; Pronk, J.T.; van Maris, A.J.A.; Daran, J.-M.G. CRISPR/Cas9: A Molecular Swiss Army Knife for Simultaneous Introduction of Multiple Genetic Modifications in Saccharomyces Cerevisiae. FEMS Yeast Research 2015, 15, fov004. [Google Scholar] [CrossRef]
- Gilbert, B.J.; Miller, C.; Corrick, F.; Watson, R.A. Should Trainee Doctors Use the Developing World to Gain Clinical Experience? The Annual Varsity Medical Debate - London, Friday 20th January, 2012. Philosophy, Ethics, and Humanities in Medicine 2013, 8. [CrossRef]
- Li, J.; Zhang, Y.; Zhang, Y.; Yu, P.-L.; Pan, H.; Rollins, J.A. Introduction of Large Sequence Inserts by CRISPR-Cas9 To Create Pathogenicity Mutants in the Multinucleate Filamentous Pathogen Sclerotinia Sclerotiorum. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, C.; Shabbir-Hussain, M.; Frogue, K.; Blenner, M.; Wheeldon, I. Standardized Markerless Gene Integration for Pathway Engineering in Yarrowia Lipolytica. ACS Synth. Biol. 2017, 6, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Chi, Y.; Lin, D.; Tyler, B.M.; Liu, X. Mutations in ORP1 Conferring Oxathiapiprolin Resistance Confirmed by Genome Editing Using CRISPR/Cas9 in Phytophthora Capsici and P. Sojae. Phytopathology® 2018, 108, 1412–1419. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Tyler, B.M. Efficient Disruption and Replacement of an Effector Gene in the Oomycete Phytophthora Sojae Using CRISPR/Cas9. Molecular Plant Pathology 2016, 17, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Geiser, E.; Wiebach, V.; Wierckx, N.; Blank, L.M. Prospecting the Biodiversity of the Fungal Family Ustilaginaceae for the Production of Value-Added Chemicals. Fungal Biol Biotechnol 2014, 1, 2. [Google Scholar] [CrossRef]
- Schuster, M.; Schweizer, G.; Reissmann, S.; Kahmann, R. Genome Editing in Ustilago Maydis Using the CRISPR–Cas System. Fungal Genetics and Biology 2016, 89, 3–9. [Google Scholar] [CrossRef]
- Gao, S.; Tong, Y.; Wen, Z.; Zhu, L.; Ge, M.; Chen, D.; Jiang, Y.; Yang, S. Multiplex Gene Editing of the Yarrowia Lipolytica Genome Using the CRISPR-Cas9 System. Journal of Industrial Microbiology and Biotechnology 2016, 43, 1085–1093. [Google Scholar] [CrossRef]
Figure 1.
Methods of gene manipulation in microbes. (A) Knock-in adds new genetic material. (B) Knock-out removes or inactivates genes. (C) Knock-down reduces gene expression. Created with BioRender.com.
Figure 1.
Methods of gene manipulation in microbes. (A) Knock-in adds new genetic material. (B) Knock-out removes or inactivates genes. (C) Knock-down reduces gene expression. Created with BioRender.com.
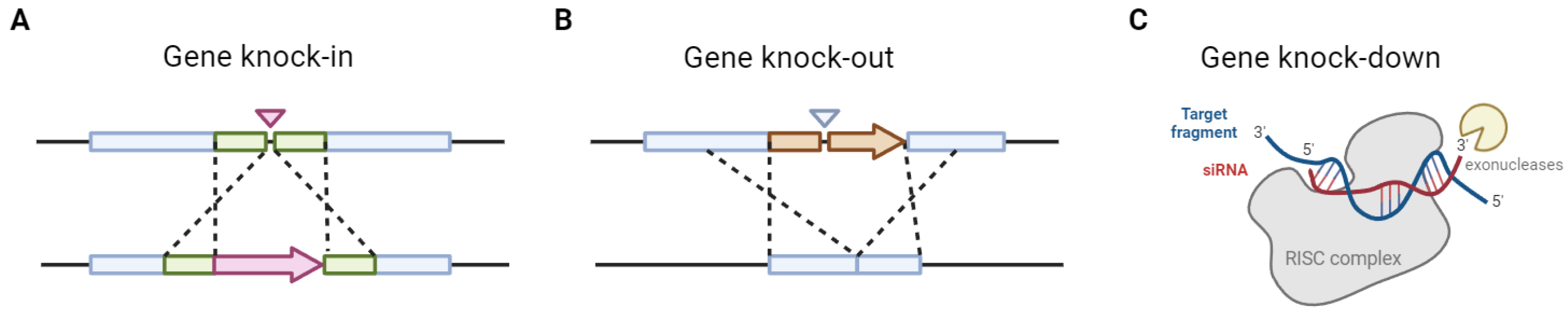
Figure 2.
Exemplary genetic engineering in oomycetes. Modified from [2]. (A) Zoospore induction. (B) Vector construction. (C) T-DNA integration. (D) AMT in zoospores. AMT, Agrobacterium-mediated transformation. (E) Isolation of positive colonies of engineered oomycete. Created with BioRender.com.
Figure 2.
Exemplary genetic engineering in oomycetes. Modified from [2]. (A) Zoospore induction. (B) Vector construction. (C) T-DNA integration. (D) AMT in zoospores. AMT, Agrobacterium-mediated transformation. (E) Isolation of positive colonies of engineered oomycete. Created with BioRender.com.
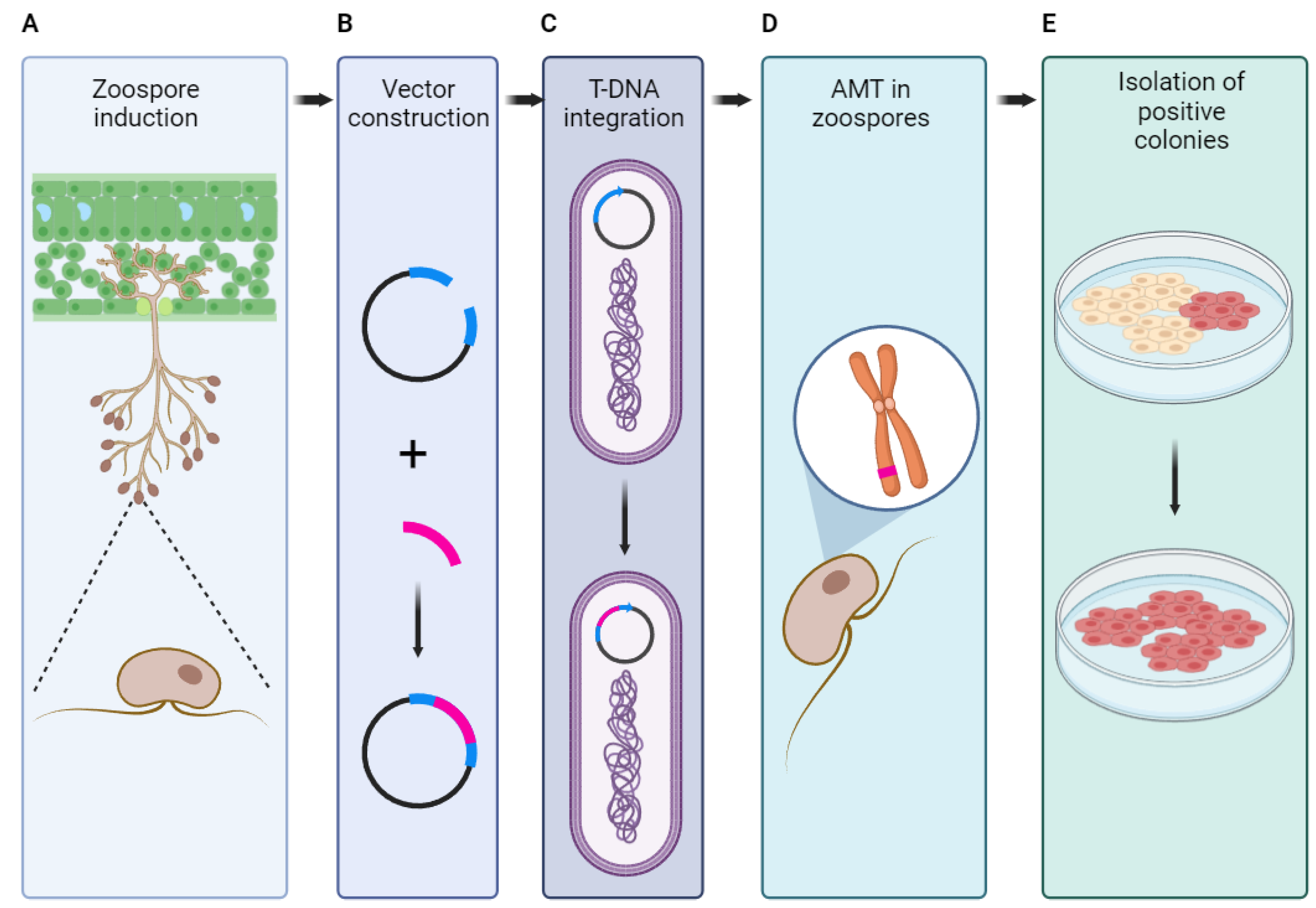

Table 1.
Popular tools for gRNA design of genetic engineering.
| Tool name | Access link | PAM sequence | Functions | Reference(s) |
|---|---|---|---|---|
| ATUM | https://www.atum.bio/eCommerce/cas9/input | NGG, NAG | Knock-in, knock-out | [79] |
| Benchling | https://www.benchling.com/crispr | Custom PAM | Knock-in, knock-out, knock-down | [80] |
| CasOFFinder | http://www.rgenome.net/cas-offinder/ | NGG, NRG, NNAGAAW, NNNNGMTT, NNGRRT, NNNVRYAC, NNNNRYAC, TTTN, TTTV, NNGTGA, TTN, NNNRRT, KYTV, NGCG, NGA, NGT, NG, TTN, ATTN, DTTN, NAAN, NNNNCC, TYCV, TATV, NNNNGNA, YTTV, NCC, NNNNCNR, NNNNCNAA, NNN, NRN, NGC, TTTR, NTTR, TTCN, NRTH, NNGG, NGGNG, NGGNG, NRTA, NNNA, TTNA, TTNA, NRNH, NGNG, NRCH | Knock-in, knock-out, knock-down | [81] |
| CCTop | https://cctop.cos.uni-heidelberg.de:8043/ | NGG, NRG, NG, NG-NRG, NGA, NGCG, TTTN, TTTV, YTN, TTN, YTTN, NNGRRT, NNNNGATT, NNAGAAW, NAAAAC, NNNNRYAC | Knock-in, knock-out | [82,83] |
| CHOPCHOP | https://chopchop.cbu.uib.no/ | NGG, NRG, NG, NG-NRG, NGA, NGCG, TTTN, TTTV, YTN, TTN, YTTN, NNGRRT, NNNNGATT, NNAGAAW, NAAAAC, NNNNRYAC | Knock-in, knock-out, activation, repression, nanopore enrichment | [84] |
| CRISPOR | http://crispor.gi.ucsc.edu/ | NGG, NNG, NGN, NNGT, NAA, NNG, NNG, NNGRRT, NGK, NNNRRT, NGA, NNNNCC, NGCG, NNAGAA, NGGNG, NNNNGMTT, NNNNACA, NNNNRYAC, TTTV, TTTN, ATTN, NTN, TYCV, TATV, TTTA, TCTA, TCCA, CCCA, GGTT, YTTV, TTYN, NNNNCNAA, NNN, NRN, NYN | Knock-in, knock-out | [85] |
| CRISPRdirect | https://crispr.dbcls.jp/ | NGG, NRG, NNGRRT, NG | Knock-in, knock-out | [86] |
| CRISPR-ERA | http://crispr-era.stanford.edu/ | NGG | Gene editing, gene repression, gene activation | [87] |
| CRISPick | https://portals.broadinstitute.org/gppx/crispick/public | NGG, NNGRR, TTTV | Knock-in, knock-out, knock-down | [88] |
| IDT | https://www.idtdna.com/site/order/designtool/index/CRISPR_SEQUENCE | Custom PAM | Knock-in, knock-out, knock-down | [89] |
| Off-Spotter | https://cm.jefferson.edu/Off-Spotter/ | NGG, NAG, NNNNACA, NNGRRT | Knock-in, knock-out | [90] |
| Synthego | https://design.synthego.com/#/ | Custom PAM | Knock-in, knock-out, knock-down | [91] |
| TrueDesign Genome Editor | https://www.thermofisher.com/us/en/home/life-science/genome-editing/invitrogen-truedesign-genome-editor.html | Custom PAM | Knockout a target gene, add a fluorescent or epitope tag, Insert, delete, or replace up to 30 bases, long insertion up to 10kb, Generate a SNP | [92] |
Table 2.
Important examples of genetic engineering in bacteria, fungi and oomycetes.
| Microbial species | Modifications | Importance | Reference(s) |
|---|---|---|---|
| Bacteria | |||
| Acidithiobacillus ferrooxidans | Knock-down | Utilized for the bioleaching of metals | [93] |
| Bacillus methanolicus | Knock-down | Methylotrophic bacteria | [94] |
| Bacillus smithii | Knock-in | Moderate thermophile capable of C5 and C6 sugar metabolism | [95,96] |
| Bacillus subtilis | Knock-in | Producer of industrial enzymes and valuable low-molecular-weight substances | [97] |
| Clostridium acetobutylicum | Knock-in | Isopropanol production | [98] |
| Clostridium autoethanogenum | Knock-in | Capable of fermenting CO, CO2, and H2 into biofuel ethanol and 2,3-butanediol | [99] |
| Clostridium beijerinckii | Knock-in | Production strain for biofuels and biochemical | [100] |
| Clostridium cellulolyticum | Knock-in | Capable of conversion of lignocellulosic biomass to valuable endproducts | [101] |
| Corynebacterium glutamicum | Knock-down | Producer of amino acids | [102] |
| Clostridium ljungdahlii | Knock-down | Capable of producing ethanol from synthesis gas | [103] |
| Clostridium pasteurianum | Knock-in | Capable of converting waste glycerol to butanol | [104] |
| Clostridium thermocellum | Knock-out | Thermophilic bacteria | [105] |
| Clostridium tyrobutyricum | Knock-out | Butanol production | [106] |
| Corynebacterium glutamicum | Knock-down | Producer of amino acids | [102] |
| Cupriavidus metallidurans | Knock-in | Facultative chemolithoautotroph | [107] |
| Cupriavidus necator | Knock-in | Facultative chemolithoautotroph | [108] |
| Escherichia coli | Knock-in, Knock-down, Knock-out | Programmed antimicrobial, recombination, multiplex recombination, CRISPRi, multiplexed CRISPRi, gene circuit, RNA targeting | [109,110] |
| Halomonas bluephagenesis | Knock-out | 3-Hydroxyvalerate production | [111] |
| Hungateiclostridium thermocellum | Knock-down | Thermophilic bacterium | [112] |
| Lactobacillus reuteri | Knock-in | Probiotic strain and producer of biotherapeutics | [113] |
| Methylobacterium extorquens | Knock-down | Methylotrophic bacterium | [114] |
| Pseudomonas putida | Knock-down | Exhibits solvent tolerance and highly versatile metabolism | [115] |
| Rhodobacter capsulatus | Knock-down | Facultative photo- and chemolithoautotroph species | [116] |
| Rhodococcus ruber TH | Knock-out | Acrylamide production | [117] |
| Streptococcus thermophilus | Knock-in | Probiotic and industrial fermentation strains | [118] |
| Streptomyces albus | Knock-in | Producer of heterologous secondary metabolites | [119] |
| Streptomyces coelicolor | Knock-in | Source of pharmacologically active and industrially relevant secondary metabolites | [119,120] |
| Synechococcus elongatus PCC 7942 | Knock-down | Squalene production | [121] |
| Synechococcus sp. PCC 7002 | Knock-down | Lactate production | [122] |
| Tatumella citrea | Knock-in | Producer of vitamin C precursor (2-keto-D-gluconic acid) | [123] |
| Vibrio natriegens | Knock-down | Fast-growing bacteria | [124] |
| Fungi | |||
| Aspergillus aculeatus | Knock-in | Source of and producer of enzymes | [125] |
| Agaricus bisporus | Knock-in | Anti-browning mushroom with extended shelf life and enhanced resistance to blemishes | [126] |
| Aspergillus fumigatus | Knock-in | For studying mechanisms of azole resistance | [127] |
| Aspergillus niger | Knock-in | Enhances the expression and secretion of α-galactosidase; improves protein production; facilitates heterologous laccase expression; and boosts glucoamylase secretion | [128,129,130,131] |
| Aspergillus oryzae | Knock-in | Enhances the expression and secretion of α-galactosidase; improves protein production; facilitates heterologous laccase expression; and boosts glucoamylase secretion | [132,133] |
| Candida albicans | Knock-in | Common production strain, capable of phenol and formaldehyde catabolism | [134,135,136] |
| Fusarium graminearum | Knock-out | Functional study of 5-oxoprolinase | [137] |
| Fusarium fujikuroi | Knock-out | GA4/GA7 mixtures production | [138] |
| Kluyveromyces lactis | Knock-in | Common production strain | [139] |
| Kluyveromyces marxianus | Knock-out | Ethyl acetate production | [140] |
| Magnaporthe oryzae | Knock-out | Study of endolysosomal trafficking mechanisms in rice blast | [141] |
| Monascus purpureus KL-001 | Knock-out | Monascus Red pigment production | [142] |
| Myceliophthora thermophile | Knock-in | Thermophilic strain and producer of cellulases | [143] |
| Neurospora crassa | Knock-in | hyper-production of cellulases | [144,145] |
| Penicillium chrysogenum | Knock-in | Producer of β-lactam antibiotics | [146] |
| Pichia pastoris | Knock-in | Common production strain | [147] |
| Trichoderma reesei | Knock-in | Expression and secretion of human erythropoietin; enhanced secretion of glucose oxidase; and hyper-cellulase production | [143,148,149] |
| Saccharomyces cerevisiae | Knock-in, Knock-down | Common production strain | [150,151,152] |
| Sclerotinia sclerotiorum | Knock-in | Introduction of large sequence inserts | [153] |
| Synechococcus elongatus PCC 7942 | Knock-in, Knock-out | Succinate production | |
| Yarrowia lipolytica | Knock-in | Lycopene production | [154] |
| Oomycetes | |||
| Phytophthora capsici | Knock-out | Conferring resistance to oxathiapiprolin | [155] |
| Phytophthora infestans | Knock-out | Increase in transformation efficiency; a modified genetic transformation and genome editing system | [2] |
| Phytophthora sojae | Knock-out | Efficient disruption and replacement of an effector gene. | [156] |
| Plasmopara viticola | Knock-out | Increase in transformation efficiency; a modified genetic transformation and genome editing system | [2] |
| Ustilago maydis | Knock-out | Natural producer of valuable biochemicals; causative agent of corn smut | [157,158] |
| Yarrowia lipolytica | Knock-in, Knock-out | Natural producer of valuable biochemical | [154,159] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



