
Submitted:
24 September 2024
Posted:
26 September 2024
You are already at the latest version
Abstract
We evaluated the individual and combined insecticidal and insectistatic activities of the acetonic extract of Jatropha curcas seeds (JcAE) and the enzymatic extract of Serratia marcescens (SmEE) against the fall armyworm Spodoptera frugiperda. An ingestion bioassay was performed using 0 ppm, JcAE (100, 250, 1000, and 2500 ppm), SmEE (40, 80, 150, and 300 ppm), or JcAE+SmEE (1000+150, 1000+300, 2500+150, and 2500+300 ppm). We determined the weight, development, and mortality of S. frugiperda larvae and pupae, identified the major components of JcAE, and evaluated the chitinolytic activity of SmEE. Mortality was compared via Fisher’s exact test, whereas continuous variables were compared via the Kruskal‒Wallis test followed by Dunn’s test. The percentage difference was also evaluated using 0 ppm as a baseline. The two highest concentrations of JcAE (1000 and 2500 ppm) and SmEE (150 and 300 ppm) resulted in significant insecticidal and insectistactic activity because they increased the cumulative larval and pupal mortality between 77.33% and 86.67%, decreased the weights of the larvae (range, -63.02% to -80.23%) and pupae (range, -8.45% to -22.72%), and retarded their development for up to 10 and 4 days, respectively. Additionally, 1000+300 and 2500+300 ppm JcAE+SmEE increased the cumulative mortality by 93.33% and 90.0%, reduced larval and pupal weights, and prolonged larval development. Seven major fatty acids were identified as the main components of JcAE, and SmEE showed exo- and endochitinase activity. JcAE and SmEE, either individually or in combination, are alternatives for sustainable biological control of S. frugiperda.
Keywords:
Physic nut
; chitinolytic
; insect pest
; biological control
; botanical extracts
; alternative insecticides
; sustainable crop production
1. Introduction
The fall armyworm, Spodoptera frugiperda J.E. Smith (Lepidoptera: Noctuidae), is a polyphagous pest that has detrimental effects on important crops in the Poaceae family, which includes corn (Zea mays) [1]. S. frugiperda represents a neotropical pest species from Central and South America that has spread to other continents, including Africa, India, and Asia [2]. The larvae of the fall armyworm represent the damaging stage of the pest, as they feed on corn buds and leaves. Furthermore, the moth of S. frugiperda has the capacity to travel distances up to 1600 km and lay up to 1500 eggs [3]. Therefore, in the absence of rapid and efficient control measures, this insect pest has the potential to cause significant yield losses in corn production, with estimated losses ranging from 8.3 to 20.6 million tons per year [4].
Synthetic insecticides, including emamectin benzoate, chlorpyrifos, and chlorantraniliprole, are frequently employed as means of combating the fall armyworm [5]. However, the excessive use of different synthetic chemical insecticides has contributed to various health and environmental problems, as well as the generation and dissemination of insecticide resistance in S. frugiperda [6]. The contamination of the environment by synthetic chemical insecticides has been extensively documented. These contaminants have been detected not only in soil and water but also in food for human consumption. The ingestion of these chemicals can cause a range of serious health problems, including vision loss, various types of cancer, liver disease [7], and neurodegenerative disorders such as Alzheimer’s disease and Parkinson’s disease [8]. Furthermore, the utilization of chemical insecticides has had a detrimental effect on a variety of beneficial insects that serve as parasitoids and predators [9]. In light of these concerns, recent research has concentrated on the pursuit of alternative sustainable methods for the control of S. frugiperda that are more environmentally friendly and do not result in toxicity to human health or adverse effects on other beneficial insects.
A number of studies have indicated the potential utility of botanical extracts derived from members of the Euphorbiaceae family as alternative agents for the control of fall armyworms. This is exemplified by Jatropha curcas L. (Euphorbiaceae), a plant also known as the physic nut, which belongs to this family and is widely distributed in tropical and subtropical regions of Asia, America, and Africa [10]. J. curcas is characterized by its durability, resilience to drought conditions, and low susceptibility to pest infestation [11]. Furthermore, the seeds of this species contain up to 60% oil, which is primarily composed of triglycerides, making it suitable for use as a biopesticide [12]. Recently, a systematic review indicated that botanical extracts obtained from different parts of J. curcas have insecticidal and insecticidal effects on different taxonomic orders of insect pests of important crops [13]. Furthermore, the hexanic and acetonic extracts, as well as the fatty acids present in the seeds of J. curcas, possess insecticidal activity, resulting in the death of the larvae and pupae of S. frugiperda [14]. Additionally, fractions enriched with phorbol esters have been shown to exert antifeedant effects [15]. In addition to botanical extracts, a variety of lytic enzymes derived from entomopathogenic bacteria, such as Serratia marcescens (Yersiniaceae), have been employed as a means of controlling insect pests [16,17]. In a previous investigation, the recombinant chitinase BL21 from S. marcescens was reported to increase the larval mortality of S. frugiperda and cause alterations in their development [18]. Moreover, it has been reported that the enzymatic extract of the bacterium, when administered via the parental route in larvae of S. frugiperda, caused toxicity [19].
The available evidence suggests that the use of extracts from J. curcas and S. marcescens may offer a promising alternative for the biological control of the fall armyworm. However, there is no evidence to suggest that the combination of these two compounds results in synergistic activity. Accordingly, the present study aimed to assess the insecticidal and insectist effects of J. curcas acetonic seed extract and S. marcescens enzymatic extract, both individually and in combination, on the larvae and pupae of S. frugiperda. The results obtained in the present study could be useful for providing evidence on the use, individually or in combination, of extracts of J. curcas and S. marcescens as alternatives to reduce or replace the use of synthetic chemical insecticides used for the control of the fall armyworm.
2. Materials and Methods
The specific objectives of the present study were as follows:
- To obtain the acetonic extract of J. curcas seeds (JcAE) and identify its major chemical compounds.
- To obtain an enzymatic extract of S. marcescens strain 81 (SmEE) and evaluate the endo- and exokitinase activity of SmEE.
- To test the individual and combined effects of JcAE and SmEE on S. frugiperda
- Determining the lethal concentrations (LC50s and LC90s) of JcAE and SmEE for larval mortality
2.1. Acetonic Extract of Jatropha curcas Seeds
2.1.1. Preparation of the Acetonic Extract of Jatropha curcas Seeds
The acetonic extract used in the study was obtained from J. curcas seeds, which were collected in October 2023 in the experimental field of Ingenio Atencingo, in the state of Puebla, Mexico (18°30′51.49” N, 98°36′26.59” W). The seeds were stored under refrigeration at 4 °C in the Entomology Laboratory of the Centro de Desarrollo de Productos Bióticos, Instituto Politécnico Nacional (CeProBi-IPN), for subsequent analysis. The seeds were dried for 24 hours in the Entomology Laboratory in the shade at room temperature (26 °C). The seeds were then disinfected with 1% sodium hypochlorite and washed with distilled water. A total of 400 g of seeds in good condition, uniform size, complete structure, adequate coloration and no obvious signs of damage were selected. The seeds were ground in a laboratory mill (KMF10 basic IKA WERKE; Wilmigton, USA) to obtain a fine powder with a final weight of 357 g, from which the plant extract was obtained. A total of 43 g of seed powder was lost due to its entrapment in the mill, which was difficult to access for recovery.
For the maceration method, technical grade acetone (J.T. Bayer®) was used as the solvent at a 1:5 ratio (357 g of J. cucas seed powder and 1785 mL of acetone). The mixture was then allowed to stand for 72 hours in an extraction hood and then filtered with Whatman No. 1 paper. The mixture was subjected to rotary evaporation (Büchi® R-205; Hamton, USA) for a period of 20 min, during which the solvent was removed under a reduced pressure of 540 mbar at 44 °C. This process yielded 250 mL of extract, which was transferred to a 300 mL Erlenmeyer flask. Finally, the mixture was dried in an extraction hood for 12 hours to ensure complete removal of the solvent.
2.1.2. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis of Jatropha curcas Seeds
The oils derived from J. curcas seeds were analyzed via an Agilent model 6850 gas chromatograph coupled to a 5973 N mass spectrometer with a single quadrupole detector (Agilent Technologies, Palo Alto, CA, USA). Chromatographic separation was performed on an Agilent HP-88 capillary column (100 m × 0.250 mm × 0.20 μm). The carrier gas was helium, and the flow rate was 1 mL/min. The injected sample volume was 1 μL, and the split ratio was 1:50. The oven temperature was started at 130 °C for 1 min, then increased to 170 °C at 6.5 °C/min, and finally increased to 230 °C at 6.5 °C/min. The injector temperature was maintained at 250 °C, and the transfer line temperature was 250 °C. The mass spectrometer was operated at 70 eV ionization energy, and the spectra were recorded in scan mode in the range of 30–500. The identification of the compounds was based on GC retention indices with reference to a homologous series of C10–C18 n-alkanes calculated via the van Dool and Kratz equation and via computer comparison with the system’s mass spectral library of GC–MS data (NIST 08) and other published mass spectra. Area percentages were obtained electronically from the GC–FID response without the use of an internal standard or correction factors.
2.2. Isolation of an Enzymatic Extract of Serratia marcescens (SmEE)
2.2.1. Serratia marcescens Culture
The primary culture of Serratia marcescens strain 81 was provided by the Laboratory of Molecular Pathogenicity, Centro de Investigación en Dinámica Celular, Universidad Autónoma del Estado de Morelos, Mexico. S. marcescens was grown at 30 °C on Lauria-Bertani (LB) solid media (10 g/L peptone, 5 g/L yeast extract and 10 g/L NaCl, 15 g/L agar, pH 7).
Five cryopreservation aliquots were made in liquid cultures in LB media at the Molecular Ecology Laboratory of the Universidad Autónoma Metropolitana Unidad Xochimilco. They were inoculated with a roast of the solid culture S. marcescens 81 and incubated for 18 hours at 37 °C with constant orbital shaking at 200 rpm. Subsequently, 1 mL aliquots of these cultures were taken and deposited in cryotubes, to which 1 mL of 50% glycerol was also added as a cryoprotective agent; the tubes were stored at -70 °C for later use.
2.2.2. Obtaining Colloidal Chitin for Chitinase-Producing Media
The collection of colloidal chitin is important for the subsequent preparation of the chitinase-producing medium. This procedure was carried out in three preparation steps where the commercial chitin powder underwent demineralization and deproteinization following the following steps: 1) demineralization, in which the commercial chitin powder was treated with a solution of 0.6 N hydrochloric acid in an 11:1 solid–liquid ratio (100 g of chitin powder in 1,100 mL HCl) for 3 h at room temperature; and 2) deproteinization, in which the demineralized chitin was treated with 1% sodium hydroxide for 24 h at room temperature. The product obtained was washed with distilled water and allowed to dry at 32 °C for 10 min (Young et al., 1985). 3) Colloidal chitin was prepared via a modified version of the method of Hsu and Lockwood (1975), which consisted of dissolving 80 g of demineralized and deproteinized chitin in 400 mL of concentrated HCl under constant stirring for 50 min. Subsequently, 2 L of cold distilled water was added. The mixture was left under continuous agitation for 18 h at room temperature and then filtered through Whatman No. 1 paper under vacuum and subsequently washed with distilled water to eliminate the acid. The paste obtained was sterilized for 30 min at 15 psi and stored at 4 °C (Castro et al., 2011). Once colloidal chitin was obtained, the preparation of the chitinase-producing medium was carried out.
2.2.3. Preparation of Serratia marcescens Inoculum
The primary inoculum of the cryopreserved strain of S. marcescens 81 was activated in liquid Luria–Bertani (LB) medium (tryptone 10 g/L, yeast extract 5 g/L, NaCl 10 g/L) and incubated at 30 °C under shaking for 24 h at pH 7. After the incubation, the inoculum was poured into enriched medium (yeast extract 0.025 g/L, magnesium sulfate 0.05 g/L, monobasic phosphate 0.068 g/L and colloidal chitin 1 g/L at pH 7) at 30 °C under constant agitation at 250 rpm for 24 h.
2.2.4. Serratia marcescens Enzymatic Extract-Producing Medium 81
The inoculum obtained from the enriched medium was poured into chitinase production medium (potassium dibasic phosphate 0.1 g/L, magnesium sulfate 0.05 g/L and colloidal chitin 2 g/L at pH 7), which was incubated at 30 °C with shaking at 250 rpm for five days. To obtain the enzymatic extract of S. marcescesns, the culture obtained was centrifuged at 2500 rpm for 10 min to obtain the cell-free supernatant. The extracellular protein concentration was determined via the Lowry method.
2.2.5. Endo and Exo Chitinase Activity Assay
A chitinase assay kit (Sigma–Aldrich®, St. Louis, MO, USA; CS0980-1KT) was used following the manufacturer’s suggested protocol to evaluate the enzymatic extract activity of S. marcescens. The kit includes three soluble substrates, 4-nitrophenyl N-acetyl-N-acetyl-β-D-glucosamine and 4-nitrophenyl N,N’-diacetyl-β-D-chitobiose, to detect exochitinase activity and 4-nitrophenyl β-D-N,N’,N’‘-triacetylchitobiose to detect endochitinase activity. Chitinase from Trichoderma viride was used as a positive control (included in the kit), while blank samples were prepared by mixing each of the soluble substrates with the stop solution (sodium carbonate). The S. marcescens enzyme extract test samples were made in duplicate. One unit of chitinase activity was defined as the release of one μmol of p-nitrophenol from each substrate per minute. Activity assays were performed at pH 4.8 and 37 °C for 30 min, and the absorbance of each sample was measured at 405 nm in a BioTek Epoch/2 ELISA microplate reader. The units per milliliter were calculated via the following formula:
U/mL: unit of enzyme activity
A405 sample: absorbance of the sample at 405 nm
A405white: absorbance of the blank at 405 nm
0.05: µmol/mL P-nitrophenol in the standard solution
0.3: final volume of the reaction in a 96-well plate after adding the stop solution (mL)
FD: dilution factor of commercial chitinase enzyme or biological solution used to prepare test samples
A405 standard: absorbance of the standard solution at 405 nm
Time: incubation time (min)
Vm: volume of the sample (mL)
2.3. Spodoptera frugiperda
2.3.1. Insect Breeding
Healthy S. frugiperda larvae were collected from corn crops in the municipality of Yautepec, in the state of Morelos, Mexico (18°53′09″N 99°03′38″W). The larvae showed no signs of disease or parasites and were fed an artificial diet throughout the developmental stages of their larval cycle until they emerged as adults. During the final stage of their cycle, they were fed only water and honey. The diet formula consisted of 800 mL of distilled water, 60 g of diet (Product No. F0635; S.W. Corn Borer, Bio-Serv, Frenchtown, NJ, USA), 20 g of sterile corn ear, 100 g of ground corn, 40 g of brewer’s yeast, 10 g of vitamins (Lepidopteran fortification mix, Bio-Serv), and 10 g of agar. Additionally, the diet was supplemented with 1.7 g of sorbic acid (dissolved in 17 mL of ethanol), 2.5 mL of formaldehyde, 1.7 g of methyl p-hydroxybenzoate, and 0.6 g of neomycin sulfate. Adults were then mated, and the females laid eggs in waxed paper bags with plastic lids containing absorbent cotton moistened with water and honey, which served as a food source for the moths. The bags were examined on a daily basis, and the eggs were then transferred to 4-oz plastic cups with adjustable lids and maintained at a temperature of 24 °C and 60% relative humidity for four days until hatching. The newly hatched larvae were maintained in a growth chamber at a temperature of 24 ± 2 °C and a relative humidity of 60 ± 5%, with a photoperiod of 12:12 h, and were fed the same artificial diet for three generations until the F3 generation was obtained and used in the experiment. Any larvae showing signs of disease due to parasitic, bacterial, or viral infestation were excluded from further analysis.
2.3.2. Spodoptera frugiperda Bioassay
Table 1 summarizes the defined treatment groups with the corresponding concentrations of J. curcas acetonic seed extract (JcAE) and S. marcescens enzymatic extract (SmEE) used in this study. These parameters were determined from preliminary bioassays and previous publications [20,21]. An artificial diet without JcAE or SmEE supplementation was used as a control group (0 ppm). Each of the treatments was mixed with the artificial diet, and 5 mL was poured into 4 oz plastic containers with adjustable lids and allowed to solidify for 4 h. One larva was placed in each container. One larva per cup was placed and incubated at 25 ± 2 °C with 70% RH and a 12:12 h photoperiod.
An ingestion bioassay was performed via a completely randomized design, with each larva considered an experimental unit and 30 neonate larvae per treatment. Intervention was assigned to the experimental units via a random number generator contained in an Excel spreadsheet.
The dependent variables evaluated in the study included 1) insectistatic activity, defined as larval weight at 7 and 14 days, pupal weight, and days of larval and pupal development, and 2) insecticidal activity, defined as larval and pupal mortality and cumulative mortality for both stages. Except for mortality and cumulative mortality, within the treatment groups (JcAE, SmEE, or JcAE+SmEE), each concentration (i.e., 100, 250, 1000, or 2500 ppm for JcAE) was compared with the control group and with the other concentrations. Mortality and cumulative mortality were expressed as the percentage of larvae and pupae that died due to the intervention divided by the total number of organisms examined.
2.4. Statistical Analysis
A probit analysis was performed to calculate the LC50 and LC90 for larval mortality with 95% confidence intervals (CIs) of the treatments (JcAE and SmEE) via BioStat (version 5.8.1).
Except for mortality and cumulative mortality, all variables were tested for normality (Shapiro–Wilk W test) and homoscedasticity (Bartlett’s test). However, since these assumptions were not met, the experimental groups were compared via the Kruskal–Wallis nonparametric test followed by Dunn’s multiple comparison test. To detect significant differences between the experimental groups, Dunn’s test was corrected for multiple comparisons via statistical hypothesis testing, with p values adjusted to account for multiple comparisons via a familywise alpha threshold and confidence level of 0.05.
Additionally, for a simplified comparative analysis, the percentage difference in larval and pupal weight and development was calculated by removing the baseline reference value (0 ppm, control group) for each concentration tested (value in ppm) via the following equation: 100*(value-baseline)/baseline. This analysis allowed a direct comparison of the effects of the concentrations, since if the treatment caused a decrease in the variable with respect to the baseline (control group), the result was a negative percentage difference, whereas if the intervention increased the response, a positive percentage difference was observed. Mortality and cumulative mortality recorded in each experimental group were compared with those recorded in the reference control group via the independent two-tailed Fisher’s exact test.
Continuous variables are presented as the means ± SDs, and a value of p < 0.05 was considered significant. Unless otherwise noted, all the statistical tests and analyses were performed in Prism 10 (GraphPad, Inc., USA).
3. Results
3.1. Chemical Compounds in J. curcas Extract
As illustrated in Figure 1, the GC–MS analysis demonstrated the presence of eight principal compounds in the oils derived from J. curcas seeds. These fatty acids included the following seven major fatty acids: linoleic acid (15.5%), oleic acid (10.4%), palmitic acid (12.9%), capric acid (13.1%), lauric acid (9.6%), stearic acid (5.2%), and myristic acid (2.6%) (Table 2).
3.2. Endo- and Exochitinase Activity of the Enzymatic Extract of Serratia marcescens
A total of 190 mL of cell-free enzyme extract with a protein concentration of 307.96 μg/mL was obtained from the chitinase-producing medium. This extract exhibited endo- and exochitinase activity when different substrates were employed, thereby promoting chitin hydrolysis. The enzymatic extract of chitinases produced by S. marcescens displayed total exochitinase activity on 4-nitrophenyl N-acetyl-β-D-glucosamine substrates of 0.039 U/mL, whereas the activity on the 4-nitrophenyl N,N′-diacetyl-β-D-chitobiose substrate reached 0.102 U/mL. Moreover, the enzyme demonstrated endo-chitinase activity for the substrate 4-nitrophenyl β-D-N,N′,N″-triacetylchitobiose, with an activity of 0.0007 U/mL.
3.3. Insectory and Insecticidal Effects of the Extracts
3.3.1. Larval and Pupal Weights
As illustrated in Figure 2A,B, exposing the larvae and pupae to the extracts resulted in a differential insectistatic effect, with only the higher concentrations of the individual extracts significantly reducing the larval and pupal weights of S. frugiperda in comparison with those of the control group. In contrast, when the extracts were combined, all the experimental groups presented a reduction in larval weight at 14 days. With the exception of the 2500+150 concentration, the remaining concentrations resulted in a significant decrease in pupal weight.
The baseline larval weight in the control group at 7 and 14 days (5.16 ± 2.23 and 76.60 ± 2.71 mg, respectively; Table 3) was used to establish the percentage reduction in larval weight. Following exposure to 1000 and 2500 ppm JcAE and 300 ppm SmEE, at 7 days, the reduction ranged from -32.01% to -42.92%, and at 14 days, it ranged from -65.44% to -80.23% (Figure 2A). However, following the exposure of S. frugiperda larvae to the combined extracts, no significant reduction was observed at 7 days. The percentage difference with respect to the baseline control group showed a variable reduction, ranging from -29.43% to -69.42%, across the four experimental groups. As shown in Table 3, the larval weights observed at lower concentrations of JcAE (100 and 250 ppm) and SmEE (40 and 80 ppm) were significantly greater than those at the highest concentration of each extract. These findings suggest that the lowest concentrations of these extracts have a limited insectistactic effect. Notably, the combined extracts demonstrated a synergistic effect that depended on the concentration tested. This is evidenced by the significant difference in the larval weight of the fall armyworm between the lowest combination (1000+150 ppm) and the highest combination (2500+300 ppm) at both 7 and 14 days.
As illustrated in Figure 2B, following exposure of the fall armyworm pupae to the two highest concentrations of the individual extracts, a notable reduction in pupal weight was observed, ranging from -8.45% to -22.72% when the control group was taken as the baseline (153.6 ± 2.6 mg, Table 4). In contrast, three of the four concentrations of the mixed extracts resulted in a significant reduction in pupal weight, with values ranging from -15.13% to -18.39%. As summarized in Table 4, the pupal weight was significantly reduced at concentrations of 1000 and 2500 ppm JcAE, with values of 140.6 ± 2.6 and 129.4 ± 2.3 mg, respectively. In comparison, the two lowest concentrations of this extract presented significantly lower values. Similarly, the pupal weight recorded with 150 and 300 ppm SmEE, which ranged between 118.7 and 135.3 mg, was lower than the weight of the pupae exposed to 40 and 80 ppm extract (ranging from 155.4 to 151.5 mg).
3.3.2. Larval and Pupal Development
The application of the extracts resulted in differential retardation of larval and pupal development in S. frugiperda, regardless of whether the extracts were used alone or in combination. Among the individual extracts, only those at relatively high concentrations demonstrated notable insectistatic effects. As shown in Figure 2C, the application of 1000 and 2500 ppm JcAE resulted in a significant increase in the number of days required to complete larval development when the control group was taken as the baseline (25.73 ± 0.77 days, Table 3). Similarly, the two higher concentrations of SmEE increased the time needed for larval development of the fall armyworm (150 and 300 ppm, 14.50 ± 0.87% and 36.02 ± 1.32%, respectively). Finally, a more heterogeneous effect was observed with all concentrations of the mixed extracts tested, which significantly increased the time needed for larval development by between 12.70% and 33.30%.
As summarized in Table 3, the larvae exposed to 100 and 250 ppm JcAE and 40 and 80 ppm JcAE needed fewer days to complete their development (range, 23.88--27.00 days) than did those exposed to the other concentrations. Therefore, the lower concentrations of the extracts had no insectistatic effect on the larval development of S. frugiperda. Conversely, 1000 and 2500 ppm JcAE and 300 ppm SmEE resulted in considerable insectistatic activity, as evidenced by prolonged larval development by approximately 10 days in comparison with that of the control group. With respect to the mixed extracts, exposure of fall armyworm larvae to 2500+150 ppm resulted in an approximately 8.57-day increase in development time compared with that of the control group. This last experimental group exhibited the greatest insectistatic activity, with larval development completed at 34.30 ± 0.82 days, which was significantly greater than that of the other concentrations tested for the combined extracts.
The pupal development of S. frugiperda in the control group required an average of 14.0 ± 1.2 days to complete (Table 4). As illustrated in Figure 2C, the pupal development time was markedly prolonged by 32.14% and 26.19% when the samples were exposed to 1000 and 2500 ppm JcAE, respectively, and between 28.57% and 30.35% when they were treated with 150 or 300 ppm SmEE. All of the concentrations tested for the mixed extracts had significant, albeit variable, insectistatic effects, with larval development being retarded by between 21.42% and 40.47% when the control group was used as the baseline. As summarized in Table 4, the number of days required to complete the pupal development stage was significantly prolonged in the experimental groups, with the longest period observed in the 2500 ppm JcAE group (17.6 ± 1.2 days) and the shortest in the 1000+300 JcAE+SmEE group (19.6 ± 0.5 days).
3.3.3. Larval and Pupal Mortality
The two highest concentrations of both JcAE and SmEE had notable insecticidal effects on S. frugiperda larvae (Table 3). Compared with the control treatment (13.3% mortality), the application of 1000 and 2000 ppm JcAE resulted in increases in larval mortality of up to 50% and 60%, respectively. A comparable effect was observed with the use of 150 and 300 ppm SmEE, which resulted in mortality rates of 56.67% and 60.0%, respectively. In contrast, the mixed extracts demonstrated more consistent insecticidal activity against the fall armyworm, as evidenced by a significant increase in larval mortality across all the evaluated concentrations compared with that of the control group (range, 50.0% to 66.6%; Table 3).
Table 4 shows that comparable insecticidal activity against S. frugiperda pupae was observed in response to 1000 and 2500 ppm JcAE, with a mortality rate of 66.67% each, whereas compared with the control, only 150 ppm SmEE significantly increased the mortality of fall armyworm pupae (69.23% vs. 23.08%, respectively). In stark contrast, the four concentrations of the mixed extracts tested presented pronounced insecticidal effects, resulting in pupal mortality rates between 72.73% and 80.0%.
The application of 1,000 and 2,500 ppm JcAE resulted in significant increases in cumulative mortality (larvae and pupae) of 83.33% and 86.67%, respectively, in comparison with those of the control group (33.33%). Similarly, the use of 150 and 300 ppm SmEE had notable effects on cumulative mortality, reaching 86.67% and 73.33%, respectively. All concentrations of the combined extracts demonstrated the highest insecticidal efficacy, with cumulative mortality rates ranging from 86.67% to 93.33% (Table 4).
The EAcJ treatment had the highest insecticidal activity, with the lowest lethal concentrations of CL50 = 1380 ppm and CL90 = 3085 ppm (gl= 2; X2 = 32.94; p < 0.0001) for S. frugiperda larval mortality, followed by SmEE, with a CL50 = 158.4 ppm and CL90 = 1,071 ppm (gl= 2; X2 = 26.24; p < 0.0001).
4. Discussion
The two higher concentrations of JcAE showed insectistatic activity, as the larval weight of S. frugiperda decreased markedly after 14 days. A similar effect was observed when this extract was used against the larvae of Copitarsia decolora (G.) (Noctuidae), whose larval weight was reduced by half [20]. This effect of feeding may be due to the metabolites present in the extract of J. curcas seeds. A study reported that the methanolic extract of J. curcas seed oil enriched with phorbol esters demonstrated antifeeding activity by affecting the relative growth rate of S. frugiperda larvae [15]. The results demonstrated that the insectistatic activity of JcAE at concentrations of 1000 and 2500 ppm had an antifeeding effect, resulting in a loss of up to 80% of the larval weight of the fall armyworm after 14 days of treatment. Furthermore, extracts derived from other parts of the J. curcas plant also demonstrated insecticidal activity. For example, the methanolic extract of J. curcas leaves resulted in a greater than forty percent reduction in larval weight in Helicoverpa armigera (H.) (Noctuidae) [22].
The findings of the present study indicate that SmEE exhibits insectistatic activity with chitinase activity at concentrations of 150 and 300 ppm. This resulted in a reduction in the larval weight of S. frugiperda by between 63% and 73% after 14 days. This activity was highly similar to the results obtained with purified chitinases expressed by the Chi60 and Chi70 genes of Xenorhabdus nematophila, which demonstrated the ability to reduce the second-instar larval growth of H. armigera by more than 80% [23]. Similarly, an insectistatic effect was demonstrated for purified chitinase produced by Bacillus subtilis, with a gradual reduction of up to 30% in larval weight in S. litura during the first, second, and third instars [24].
Furthermore, the combination of JcAE and SmEE at concentrations of 2500+300 ppm had a synergistic effect on both extracts, as evidenced by their insectistatic activity, which resulted in a 69% reduction in larval weight in S. frugiperda at 14 days. Similarly, the individual extracts JcAE at 2500 ppm and SmEE at 300 ppm, as well as their mixture JcAE+SmEE at 2500+300 ppm, significantly decreased pupal weight and prolonged larval and pupal development by ten and four days, respectively. These findings reaffirm the antifeedant effects of these extracts on the insect pest S. frugiperda. To date, no research has been conducted on the potential synergistic effects of mixtures of botanical and enzymatic extracts of entomopathogenic microorganisms. Consequently, further investigations into these natural formulations are imperative.
The results demonstrated that, compared with the control, JcAE treatment at 2500 ppm resulted in 60% mortality in larvae and 33% mortality in pupae of S. frugiperda. This effect has been previously reported in J. curcas seed oil, with 50% mortality of S. frugiperda larvae [25]. The insecticidal activity of this treatment has been evaluated in other species of agricultural pest insects of the Noctuidae family, such as C. decolora larvae. In these instances, the acetone extract of J. curcas produced mortality rates comparable to those reported in this study [20].
Diabaté, et al. [26] conducted field trials in which they sprayed an aqueous extract of J. curcas seeds on tomato plots to control Helicoverpa armigera (H.) (Noctuidae). They reported a 57% reduction in the number of larvae, indicating that the aqueous extract of J. curcas can be used in an integrated management program for H. armigera in tomato crops. It is necessary to determine the insecticidal effects of the fatty acids present in the acetonic seed extract of J. curcas on S. frugiperda. Some studies have demonstrated that oleic, linoleic, palmitic, stearic, arachidonic, and myristic fatty acids increase the mortality and inhibition rates of Helicoverpa zea (B.) in neuronal cells (AW1 cells), which suggests that lauric acid induces apoptosis in AW1 cells [27]. Furthermore, linoleic acid extracted from Ricinus communis (another member of the Euphorbiaceae family) was reported to result in 75% larval mortality in S. frugiperda [22].
The present study demonstrated the efficacy of JcAE in combating S. frugiperda. Other studies have reported the use of other vegetative parts with insecticidal activity, such as the methanolic extract of fresh and dried leaves of J. curcas, which has been shown to cause larval mortality in S. frugiperda at rates of 60–56% [28]. Similarly, the use of extracts from different parts of J. curcas has demonstrated comparable insecticidal effects on other insect pest species of the Noctuidae family, including several botanical extracts of J. curcas. For example, the methanolic extract of J. curcas leaves has been demonstrated to cause an average mortality percentage of sixty percent in larvae of Achaea janata (L.) (Noctuidae) [29], whereas curcas leaves have been shown to cause the same mortality percentage in larvae of Spodoptera litura (F.) (Noctuidae) [30].
The results demonstrated that the insecticidal activity of the chitinases present in the SmEE treatment at 300 ppm resulted in 60% mortality of S. frugiperda larvae with respect to the control group. Similarly, purified chitinases from S. marcescens have been demonstrated to exhibit insecticidal activity, resulting in 80% mortality in H. armigera larvae [31]. In a different study, a protein extract with chitinase activity derived from Paenibacillus sp. strain D1 bacteria resulted in 63% larval mortality in H. armigera [32]. Notably, the percentage of insecticidal activity may vary according to the microorganism producing the chitinases and the pest insect it controls. The results demonstrated that the chitinases derived from S. marcescens at 150 ppm presented insecticidal activity, resulting in a substantial mortality rate in S. frugiperda larvae. However, the chitinase purified from Pseudomonas ssp. Strain TXG6-1 exhibited minimal insecticidal activity, with a mortality rate of only 17% in S. litura larvae [33]. However, the exochitinase activity of the enzyme extract of B. thuringiensis strain aizawa was the most pronounced, resulting in complete larval mortality in Spodoptera littoralis (B.) (Noctuidae) [34]. Conversely, our findings demonstrated the insecticidal activity of the JcAE+SmEE mixture at concentrations of 1000+300 and 2500+300 ppm, which resulted in 62% and 63% mortality in S. frugiperda larvae, respectively. The current findings demonstrate that the combination of JcAE and SmEE has a synergistic effect, which could prove beneficial in the management of S. frugiperda, a significant agricultural pest.
The results of the chemical compound identification analysis confirmed the presence of fatty acids, including capric, lauric, and myristic acids, as well as the major linoleic, palmitic, and oleic fatty acids present in the JcAE extract (Table 2). These findings are consistent with those of other studies, which identified the major compounds oleic, linoleic, and palmitic acid [35,36]. Similarly, these same three fatty acids and vaccenic acid have been previously identified as the primary compounds present in J. curcas seeds [20], with capric acid also being identified in our study as one of the major compounds present in JcAE.
SmEE treatment resulted in the production of chitinolytic endo- and exochitinase activity for the substrates glucosamine, chitobiose, and triacetylchitobiose. These same activities have been previously reported for the SEN strain of S. marcescens [16]. Furthermore, S. marcescens both exhibited chitinolytic activities, in contrast to the crude extract of rChit37 chitinases derived from the entomopathogenic fungus Beauveria bassiana, which demonstrated solely exochitinase activity on the substrate chitobiose [21]. Similarly, single exochitinase activity was observed in the bacterium Bacillus thuringiensis strain Aizawa [34].
5. Conclusions
The results of the present study indicate that the SmEE and JcAE treatments, when administered individually or in combination, promoted insecticidal and insecticidal activity in S. frugiperda. The combination of JcAE and SmEE had a synergistic effect, exhibiting efficacy against the larvae and pupae of S. frugiperda. In light of these findings, the use of these mixtures of JcAE+SmEE may be considered a potential sustainable alternative for the agroecological management of the fall armyworm. The utilization of these mixtures would result in a reduction in the application of synthetic chemical insecticides, thereby affording protection to the environment and preventing the development of insect resistance to these extracts when applied in mixtures. Nevertheless, future research should concentrate on conducting bioassays of these mixtures on corn plants under greenhouse conditions and in experimental plots.
Author Contributions
Conceptualization, Armando Valdez-Ramirez, Maria de la Torre-Hernandez, Rodolfo Figueroa-Brito and Antonio Flores-Macias; Data curation, Miguel Ramos-Lopez; Formal analysis, Armando Valdez-Ramirez, Pavel Hernandez-Carreño and Daniel Diaz; Funding acquisition, Rodolfo Figueroa-Brito; Investigation, Antonio Flores-Macias, Maria Nuñez-Valdez and Miguel Ramos-Lopez; Methodology, Armando Valdez-Ramirez and Eloy Rodriguez del Leon; Project administration, Rodolfo Figueroa-Brito; Supervision, Maria de la Torre-Hernandez; Validation, Maria Nuñez-Valdez; Visualization, Eloy Rodriguez del Leon; Writing—original draft, Armando Valdez-Ramirez and Daniel Diaz; Writing—review & editing, Erick Gutierrez-Grijalva, Erik Lizarraga-Verdugo, Perla Gutierrez-Arzapalo and Daniel Diaz.
Funding
This work was funded by the National Science and Technology of México (CONAHCYT) and the National Polytechnic Institute (SIP20221518).
Data Availability Statement
All data generated and used to produce the study is available upon reasonable request to the corresponding author.
Acknowledgments
The principal author acknowledges the scholarship with number CVU 632879 and researchers for Mexico from the National Council of Humanities, Sciences and Technologies of Mexico (CONAHCYT) and the Center for the Development of Biotic Products of the National Polytechnic Institute for this scientific research.
Conflicts of Interest
The authors declare that they have no conflicts of interest, financial or otherwise.
References
- Overton, K.; Maino, J.L.; Day, R.; Umina, P.A.; Bett, B.; Carnovale, D.; Ekesi, S.; Meagher, R.; Reynolds, O.L. Global crop impacts, yield losses and action thresholds for fall armyworm (Spodoptera frugiperda): A review. Crop Protection 2021, 145, 105641. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Goergen, G.; Koffi, D.; Agboka, K.; Adjevi, A.K.M.; Du Plessis, H.; Van den Berg, J.; Tepa-Yotto, G.T.; Winsou, J.K.; Meagher, R.L.; et al. Genetic studies of fall armyworm indicate a new introduction into Africa and identify limits to its migratory behavior. Sci Rep 2022, 12, 1941. [Google Scholar] [CrossRef] [PubMed]
- Cock, M.J.W.; Beseh, P.K.; Buddie, A.G.; Cafa, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci Rep 2017, 7, 4103. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.M.; Gadratagi, B.-G.; Paramesh, V.; Kumar, P.; Madivalar, Y.; Narayanappa, N.; Ullah, F. Sustainable Management of Invasive Fall Armyworm, Spodoptera frugiperda. Agronomy 2022, 12, 2150. [Google Scholar] [CrossRef]
- Ahmed, K.S.; Idrees, A.; Majeed, M.Z.; Majeed, M.I.; Shehzad, M.Z.; Ullah, M.I.; Afzal, A.; Li, J. Synergized Toxicity of Promising Plant Extracts and Synthetic Chemicals against Fall Armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) in Pakistan. Agronomy 2022, 12, 1289. [Google Scholar] [CrossRef]
- Gutierrez-Moreno, R.; Mota-Sanchez, D.; Blanco, C.A.; Whalon, M.E.; Teran-Santofimio, H.; Rodriguez-Maciel, J.C.; DiFonzo, C. Field-Evolved Resistance of the Fall Armyworm (Lepidoptera: Noctuidae) to Synthetic Insecticides in Puerto Rico and Mexico. J Econ Entomol 2019, 112, 792–802. [Google Scholar] [CrossRef]
- Gupta, A. Pesticide residues in food commodities; Agrobios (India): 2006.
- Asghar, U.; Malik, M. Pesticide Exposure and Human Health: A Review. Journal of Ecosystem & Ecography 2016, 01. [Google Scholar] [CrossRef]
- Tavares, W.S.; Costa, M.A.; Cruz, I.; Silveira, R.D.; Serrao, J.E.; Zanuncio, J.C. Selective effects of natural and synthetic insecticides on mortality of Spodoptera frugiperda (Lepidoptera: Noctuidae) and its predator Eriopis connexa (Coleoptera: Coccinellidae). J Environ Sci Health B 2010, 45, 557–561. [Google Scholar] [CrossRef]
- Nunes, C.F.; Santos, D.N.d.; Pasqual, M.; Valente, T.C.T. External morphology of fruits, seeds and seedlings of physic nut. Pesquisa Agropecuária Brasileira 2009, 44, 207–210. [Google Scholar] [CrossRef]
- Pinto, T.L.F.; Marcos Filho, J.; Forti, V.A.; Carvalho, C.d.; Gomes Junior, F.G. Evaluation of the Jatropha curcas L. seed viability by tetrazolium and X-ray tests. Revista Brasileira de Sementes 2009, 31, 195–201. [Google Scholar] [CrossRef]
- Thi, H.T.; Le, B.A.; Le, H.N.T.; Okitsu, K.; Imamura, K.; Takenaka, N.; Luu, B.V.; Maeda, Y. Screening of fatty acids, saccharides, and phytochemicals in Jatropha curcas seed kernel as their trimethylsilyl derivatives using gas chromatography/mass spectrometry. J Chromatogr B Analyt Technol Biomed Life Sci 2018, 1102-1103, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Ramirez, A.; Flores-Macias, A.; Figueroa-Brito, R.; Torre-Hernandez, M.E.d.l.; Ramos-Lopez, M.A.; Beltran-Ontiveros, S.A.; Becerril-Camacho, D.M.; Diaz, D. A Systematic Review of the Bioactivity of Jatropha curcas L. (Euphorbiaceae) Extracts in the Control of Insect Pests. Sustainability 2023, 15, 11637. [Google Scholar] [CrossRef]
- Valdez-Ramírez, A.; Flores-Macías, A.; Ramos-López, M.Á.; Castañeda-Espinoza, J.D.; Rodríguez-González, F.; Herrera-Figueroa, L.E.; Figueroa-Brito, R. Effect of Extracts and Compounds of Jatropha curcas L. Seeds Against the Fall Armyworm Spodoptera frugiperda1. Southwestern Entomologist 2024, 49, 120–132. [Google Scholar] [CrossRef]
- Devappa, R.K.; Angulo-Escalante, M.A.; Makkar, H.P.S.; Becker, K. Potential of using phorbol esters as an insecticide against Spodoptera frugiperda. Industrial Crops and Products 2012, 38, 50–53. [Google Scholar] [CrossRef]
- Aggarwal, C.; Paul, S.; Tripathi, V.; Paul, B.; Khan, M. Chitinolytic activity in Serratia marcescens (strain SEN) and potency against different larval instars of Spodoptera litura with effect of sublethal doses on insect development. BioControl 2015, 60, 631–640. [Google Scholar] [CrossRef]
- Aggarwal, C.; Paul, S.; Tripathi, V.; Paul, B.; Khan, M. Characterization of putative virulence factors of Serratia marcescens strain SEN for pathogenesis in Spodoptera litura. J Invertebr Pathol 2017, 143, 115–123. [Google Scholar] [CrossRef]
- El-Sayed, G.M.; Emam, M.T.H.; Hammad, M.A.; Mahmoud, S.H. Gene Cloning, Heterologous Expression, and In Silico Analysis of Chitinase B from Serratia marcescens for Biocontrol of Spodoptera frugiperda Larvae Infesting Maize Crops. Molecules 2024, 29, 1466. [Google Scholar] [CrossRef]
- Pineda-Castellanos, M.L.; Rodriguez-Segura, Z.; Villalobos, F.J.; Hernandez, L.; Lina, L.; Nunez-Valdez, M.E. Pathogenicity of Isolates of Serratia Marcescens toward Larvae of the Scarab Phyllophaga Blanchardi (Coleoptera). Pathogens 2015, 4, 210–228. [Google Scholar] [CrossRef]
- Figueroa-Brito, R.; Tabarez-Parra, A.S.; Avilés-Montes, D.; Rivas-González, J.M.; Ramos-López, M.Á.; Sotelo-Leyva, C.; Salinas-Sánchez, D.O. Chemical Composition of Jatropha curcas Seed Extracts and Its Bioactivity Against Copitarsia decolora under Laboratory and Greenhouse Conditions. Southwestern Entomologist 2021, 46, 103–114. [Google Scholar] [CrossRef]
- Lovera, A.; Belaich, M.; Villamizar, L.; Patarroyo, M.A.; Barrera, G. Enhanced virulence of Beauveria bassiana against Diatraea saccharalis using a soluble recombinant enzyme with endo- and exochitinase activity. Biological Control 2020, 144, 104211. [Google Scholar] [CrossRef]
- Ingle, K.P.; Deshmukh, A.G.; Padole, D.A.; Dudhare, M.S. Screening of insecticidal activity of Jatropha curcas (L.) against diamond back moth and Helicoverpa armigera. Seed 2017, 5, 20. [Google Scholar]
- Liu, J.; Bai, H.; Song, P.; Nangong, Z.; Dong, Z.; Li, Z.; Wang, Q. Insecticidal Activity of Chitinases from Xenorhabdus nematophila HB310 and Its Relationship with the Toxin Complex. Toxins (Basel) 2022, 14, 646. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, R.; Revathi, K.; Thanigaivel, A.; Kirubakaran, S.A.; Senthil-Nathan, S. Bacillus subtilis chitinase identified by matrix-assisted laser desorption/ionization time-of flight/time of flight mass spectrometry has insecticidal activity against Spodoptera litura Fab. Pestic Biochem Physiol 2014, 116, 1–12. [Google Scholar] [CrossRef] [PubMed]
- López, J.J.; Chirinos, D.T.; Ponce, W.H.; Solórzano, R.F.; Alarcón, J.P. Insecticide activity of botanical formulates on the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Revista Colombiana De Entomología 2022, 48. [Google Scholar] [CrossRef]
- Diabaté, D.; Gnago, J.A.; Koffi, K.; Tano, Y. The effect of pesticides and aqueous extracts of Azadirachta indica (A. Juss) and Jatropha carcus L. on Bemisia tabaci (Gennadius) (Homoptera: Aleyrididae) and Helicoverpa armigera (Hübner)(Lepidoptera: Noctuidae) found on tomato plants in Côte d’Ivoire. Journal of Applied Biosciences 2014, 80, 7132–7143. [Google Scholar] [CrossRef]
- Ren, Y.; Shi, J.; Mu, Y.; Tao, K.; Jin, H.; Hou, T. AW1 Neuronal Cell Cytotoxicity: The Mode of Action of Insecticidal Fatty Acids. J Agric Food Chem 2019, 67, 12129–12136. [Google Scholar] [CrossRef]
- Ribeiro, S.S.; Silva, T.B.d.; Moraes, V.R.d.S.; Nogueira, P.C.d.L.; Costa, E.V.; Bernardo, A.R.; Matos, A.P.; Fernandes, B.; Silva, M.F.d.G.F.d.; Pessoa, Â.M.d.S. Chemical constituents of methanolic extracts of Jatropha curcas L and effects on spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae). Química Nova 2012, 35, 2218–2221. [Google Scholar] [CrossRef]
- Devanand, P.; Rani, P.U. Biological potency of certain plant extracts in management of two lepidopteran pests of Ricinus communis L. Journal of Biopesticides 2008, 1, 170–176. [Google Scholar]
- Ingle, K.P.; Deshmukh, A.G.; Padole, D.A.; Dudhare, M.S. Bioefficacy of crude extracts from Jatropha curcas against Spodoptera litura. Seed 2017, 5, 0. [Google Scholar]
- Danişmazoğlu, M.; DemİR, İ.; Sezen, K.; MuratoĞLu, H.; NalÇAcioĞLu, R. Cloning and expression of chitinase A, B, and C (chiA, chiB, chiC) genes from Serratia marcescens originating from Helicoverpa armigera and determining their activities. Turkish Journal of Biology 2015, 39, 78–87. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, A.; Joshi, P. Combined application of chitinolytic bacteriumPaenibacillussp. D1 with low doses of chemical pesticides for better control ofHelicoverpa armigera. International Journal of Pest Management 2016, 62, 222–227. [Google Scholar] [CrossRef]
- Zhong, W.; Ding, S.; Guo, H. The chitinase C gene PsChiC from Pseudomonas sp. and its synergistic effects on larvicidal activity. Genet Mol Biol 2015, 38, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Sampson, M.N.; Gooday, G.W. Involvement of chitinases of Bacillus thuringiensis during pathogenesis in insects. Microbiology (Reading) 1998, 144 Pt 8, 2189–2194. [Google Scholar] [CrossRef]
- Martinez-Diaz, Y.; Gonzalez-Rodriguez, A.; Rico-Ponce, H.R.; Rocha-Ramirez, V.; Ovando-Medina, I.; Espinosa-Garcia, F.J. Fatty Acid Diversity is Not Associated with Neutral Genetic Diversity in Native Populations of the Biodiesel Plant Jatropha curcas L. Chem Biodivers 2017, 14, e1600188. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, J.P.F. Teor e composição do óleo de sementes de Jatropha spp. Bragantia 1987, 46, 151–157. [Google Scholar] [CrossRef]
Figure 1.
Major chemical compounds in the acetonic extract of Jatropha curcas seeds.
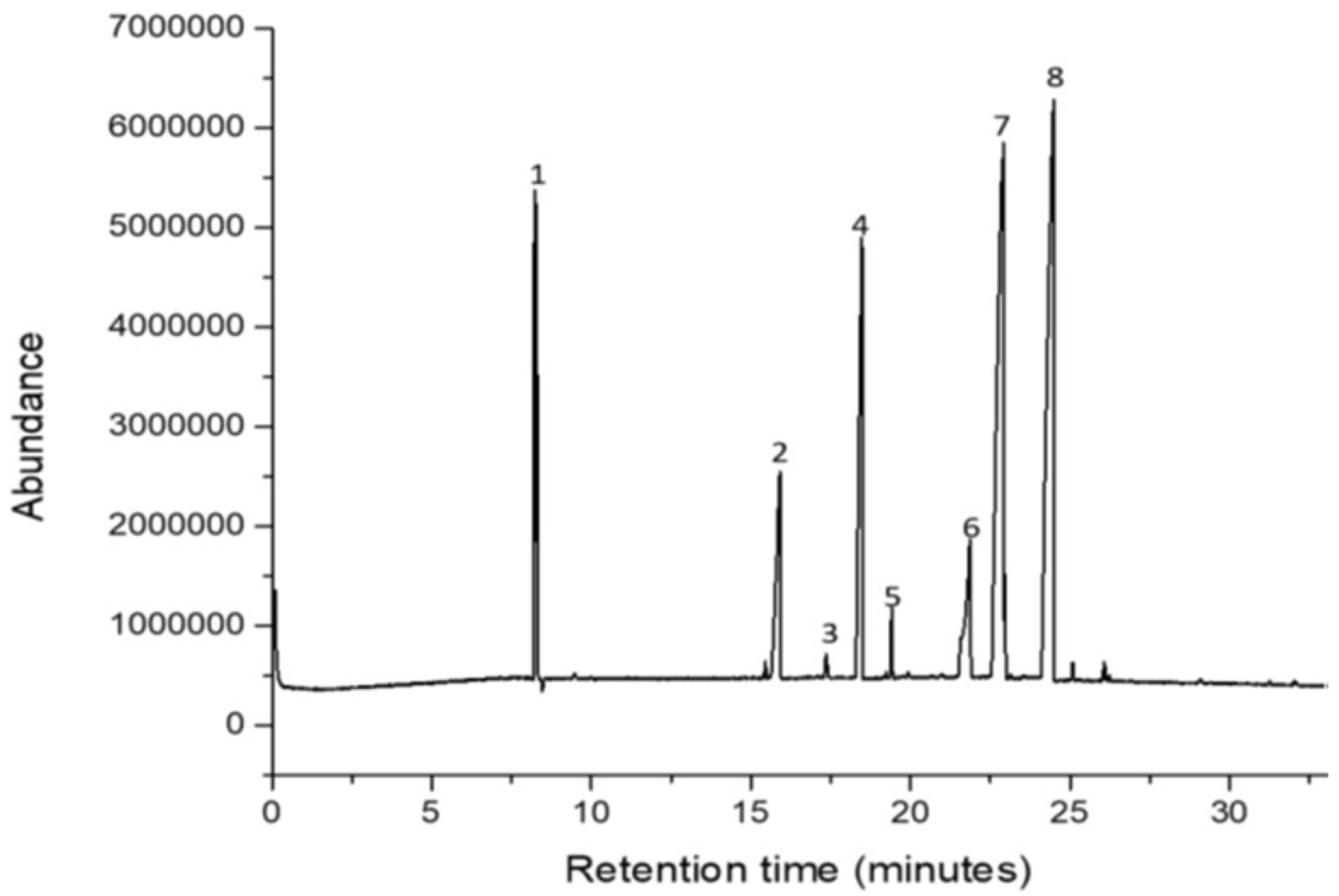
Figure 2.
Insectory effects of the extracts on the larvae and pupae of Spodoptera frugiperda.
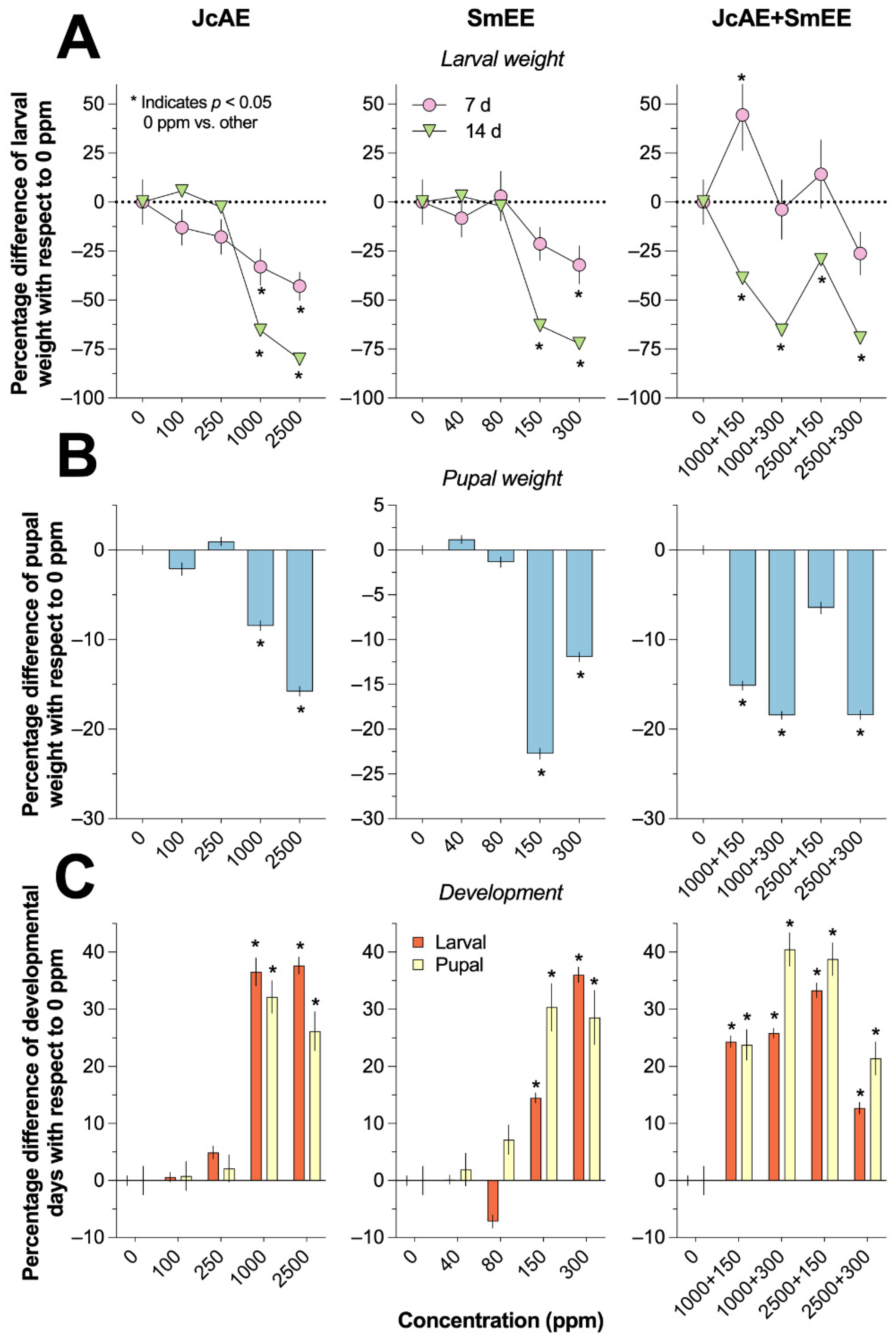

Table 1.
Experimental groups used to evaluate the insecticidal and insectistatic activities of the acetonic extract of Jatropha curcas seeds and the enzymatic extract of Serratia marcescens against Spodoptera frugiperda.
Table 1.
Experimental groups used to evaluate the insecticidal and insectistatic activities of the acetonic extract of Jatropha curcas seeds and the enzymatic extract of Serratia marcescens against Spodoptera frugiperda.
| Treatment | Acetonic extract of J. curcas seeds (JcAE) ppm | Enzymatic extract of S. marcescens (SmEE) ppm |
|---|---|---|
| Control | 0 | 0 |
| JcAE | 100 | 0 |
| 250 | 0 | |
| 1000 | 0 | |
| 2500 | 0 | |
| SmEE | 0 | 40 |
| 0 | 80 | |
| 0 | 150 | |
| 0 | 300 | |
| JcAE+SmEE | 1000 | 150 |
| 1000 | 300 | |
| 2500 | 150 | |
| 2500 | 300 |
JcAE = acetonic extract of J. curcas seeds; SmEE = enzymatic extract of S. marcescens; JcAE+SmEE = mixture of both treatments.
Table 2.
Chemical composition of the acetonic extract of Jatropha curcas seeds.
| Elution order | Retention time (minutes) | Compound | KIL | KI | % Area* |
|---|---|---|---|---|---|
| 1 | 8.17 | Capric acid (C10:0) | 1279 | 1231 | 13.1 |
| 2 | 15.91 | Lauric acid (C12:0) | 1562 | 1543 | 9.6 |
| 3 | 17.34 | Myristic acid (C14:0) | 1780 | 1771 | 2.6 |
| 4 | 18.47 | Palmitic acid (C16:0) | 1942 | 1902 | 12.9 |
| 5 | 19.41 | Stearic acid (C18:0) | 2063 | 2053 | 5.2 |
| 6 | 21.85 | Oleic acid (C18:1 (cis-9)) | 2095 | 2087 | 10.4 |
| 7 | 22.91 | Linoleic acid (C18:2(cis-9,12)) | 2175 | 2170 | 15.5 |
| 8 | 24.48 | Unknown | ------ | ------- | 17.1 |
KIL: Kovats index reported in the literature [22]. KI: Kovats index, relative to (C10--C18) n-alkanes in the column (Agilent 100 m × 0.250 mm × 0.20 μm); *Values reported as the percentage of the total area of all compounds in the extract.
Table 3.
Insecticidal and insectistatic activity of acetonic extract of Jatropha curcas and enzymatic extract of Serratia marcescens on larvae of Spodoptera frugiperda.
Table 3.
Insecticidal and insectistatic activity of acetonic extract of Jatropha curcas and enzymatic extract of Serratia marcescens on larvae of Spodoptera frugiperda.
| Treatment | ppm | Larval weight (mg) | Larval development (d) | Larval mortality (%) | |
|---|---|---|---|---|---|
| 7 d | 14 d | ||||
| Control | 0 | 5.16 ± 2.23 | 76.60 ± 2.71 | 25.73 ± 0.77 | 4/30 (13.33) |
| *JcAE | 100 | 4.49 ± 1.51 a | 80.89 ± 4.37 a | 25.89 ± 0.75 b | 3/30 (10.00) |
| 250 | 4.25 ± 1.61 a | 74.62 ± 2.26 b | 27.00 ± 1.27 b | 3/30 (10.00) | |
| 1000 | 3.46 ± 1.93 b | 26.47 ± 2.49 c | 35.13 ± 2.26 a | 15/30 (50.00)* | |
| 2500 | 2.95 ± 1.27 b | 15.14 ± 2.29 c | 35.42 ± 1.08 a | 18/30 (60.00)* | |
| SmEE | 40 | 4.74 ± 1.60 a | 78.85 ± 3.09 a | 25.77 ± 0.66 b | 5/30 (16.67) |
| 80 | 5.32 ± 2.51 a | 74.99 ± 2.95 a | 23.88 ± 1.33 c | 5/30 (16.67) | |
| 150 | 4.06 ± 1.01 a,b | 28.32 ± 2.34 b | 29.46 ± 0.51 a | 17/30 (56.67)* | |
| 300 | 3.51 ± 1.89 b | 21.31 ± 2.76 b | 35.00 ± 0.89 a | 18/30 (60.00)* | |
| JcAE+SmEE | 1000+150 | 7.46 ± 2.96 a | 46.85 ± 2.45 a | 32.00 ± 0.70 b | 15/30 (50.00)* |
| 1000+300 | 4.97 ± 2.83 b | 26.53 ± 2.65 b | 32.39 ± 0.50 b | 20/30 (66.67)* | |
| 2500+150 | 5.90 ± 3.33 a,b | 54.06 ± 2.94 a | 34.30 ± 0.82 a | 15/30 (50.00)* | |
| 2500+300 | 3.81 ± 1.84 b | 23.42 ± 2.81 b | 29.00 ± 0.75 c | 19/30 (63.33)* | |
* Indicates control vs. other p < 0.05; a, b, c: Distinct literals between groups within a treatment indicate p < 0.05.
Table 4.
Insecticidal and insectistatic activity of acetonic extract of Jatropha curcas and enzymatic extract of Serratia marcescens on pupae of Spodoptera frugiperda.
Table 4.
Insecticidal and insectistatic activity of acetonic extract of Jatropha curcas and enzymatic extract of Serratia marcescens on pupae of Spodoptera frugiperda.
| Treatment | ppm | Pupal weight (mg) | Pupal development (d) | Pupal mortality (%)+ | Larvae and pupae cumulative mortality (%) |
|---|---|---|---|---|---|
| Control | 0 | 153.6 ± 2.6 | 14.0 ± 1.2 | 6/26 (23.08) | 10/30 (33.33) |
| *JcAE | 100 | 150.3 ± 4.8 a | 14.1 ± 1.3 b | 5/27 (18.52) | 8/30 (26.67) |
| 250 | 155.1 ± 2.4 a | 14.3 ± 1.1 b | 6/27 (22.22) | 9/30 (30.00) | |
| 1000 | 140.6 ± 2.6 b | 18.5 ± 0.7 a | 10/15 (66.67)* | 25/30 (83.33)* | |
| 2500 | 129.4 ± 2.3 b | 17.6 ± 1.2 a | 8/12 (66.67)* | 26/30 (86.67)* | |
| SmEE | 40 | 155.4 ± 2.3 a | 14.2 ± 1.5 c | 4/25 (16.00) | 9/30 (30.00) |
| 80 | 151.5 ± 3.8 a | 15.0 ± 1.2 b,c | 5/25 (20.00) | 10/30 (33.33) | |
| 150 | 118.7 ± 3.0 b | 18.2 ± 0.9 a | 9/13 (69.23)* | 26/30 (86.67)* | |
| 300 | 135.3 ± 2.1 b | 18.0 ± 1.0 a,b | 4/12 (33.33) | 22/30 (73.33)* | |
| JcAE+SmEE | 1000+150 | 130.3 ± 2.5 a | 17.3 ± 0.5 a | 11/15 (73.33)* | 26/30 (86.67)* |
| 1000+300 | 125.3 ± 2.0 b | 19.6 ± 0.5 b | 8/10 (80.00)* | 28/30 (93.33)* | |
| 2500+150 | 143.7 ± 2.7 a | 19.4 ± 0.5 b | 11/15 (73.33)* | 26/30 (86.67)* | |
| 2500+300 | 125.3 ± 2.4 b | 17.0 ± 0.7 a | 8/11 (72.73)* | 27/30 (90.00)* |
* Indicates control vs. other p < 0.05; a, b, c; Distinct literals between groups within a treatment indicate p < 0.05; + The number of pupae assessed represented the remaining individuals not killed during the larval evaluation shown in Table 3.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



