
Submitted:
24 September 2024
Posted:
25 September 2024
You are already at the latest version
Abstract
The associations among increased pain sensitivity, obesity, and systemic inflammation have not been described as a direct link between BBB dysfunctions. To analyze the metabolic, behavioral, and inflammatory effects of a HFD and ultrastructural modifications in brain regions, we used an experimental model. Adult male Wistar rats were randomly assigned to one of two conditions, an ad libitum control group, or a high-fat-diet (60%) group (HFD-fed), for eight weeks. At the end of the protocol, glucose, and insulin tolerance tests were performed. Additionally, we analyzed the response to a normally innocuous mechanical stimulus and changes in motor coordination. At the end of the protocol, HFD-fed individuals presented increased HOMA-IR and metabolic syndrome (MetS) prevalence. HFD-fed rats also developed an increased nociceptive response to mechanical stimuli and neurological injury, resulting in impaired motor function. Hypothalamus and cerebellum neurons from HFD-fed presented with nuclear swelling, an absence of nucleoli, and karyolysis. These results reveal that HFD consumption affecting vital brain structures such as the cerebellum, hippocampus, and hypothalamus. This, in turn, causes neuronal damage, impairs cellular communication, and alters motricity and pain sensitivity. Understanding the link between BBB impairment and sensory and motor changes could lead to targeted therapeutic strategies.
Keywords:
neuroimmunology
; nociception
; neuronal integrity
; motor impairment
1. Introduction
The consequences of obesity are diverse and include the development of metabolic syndrome (MetS) with abdominal obesity, elevated triglycerides, low HDL cholesterol, high blood pressure, glucose homeostasis disruption, and insulin resistance [1]. The pathophysiology of MetS involves a positive energy balance, characterized by hyperplasia and hypertrophy of adipose tissue, with changes in leptin release [2]. One of the main methods for inducing obesity is to feed animals a high-fat diet (HFD) [3,4,5,6]. Obesity and consumption of a HFD may contribute to the development of cognitive disorders [7], neuroinflammation [8], and dysfunction of the blood‒brain barrier (BBB) [9,10,11], causing alterations in gap (connexin-43) [12], and tight (occludin) junctions [13], which can cause damage to certain areas and produce behavioral changes. Cell modifications have been reported in hypothalamic nuclei with decreased tight junction protein expression [14], and Evans blue diffusion is altered in arcuate nuclei [15]. Some obesity models have shown that immunoreactivity to glial fibrillary acidic protein (GFAP) increases in the hypothalamus [16], including the arcuate nucleus [17], leading to gliosis. In some models, modifications in the cellular ultrastructure include vacuolar degeneration in the hippocampus [18], and damage to Purkinje and granular cells in the cerebellum [19].
This emerging understanding highlights a crucial link between metabolic conditions such as obesity and altered pain perception, as obesity-induced systemic inflammation and blood‒brain barrier dysfunction may influence these brain regions, thereby affecting how pain is processed and experienced. Recent research has revealed the involvement of nonconventional brain areas, with a special emphasis on the hippocampus [20]. Surprisingly, in obese animals fed a Western diet, an increase in sensitivity to pain was observed starting at week 8 [21,22], with persistence at 12 weeks. Furthermore, a Western diet was shown to impair motor coordination at 4, 12, and 22 weeks [23,24,25], and investigations in a short model of changes in brain structure and function are essential. The aim of this study was to investigate the relationship between ultrastructural changes in the BBB in neuroanatomical regions associated with pain and motricity, such as the cerebellum and hippocampus, in an integral model of functional alterations in behavior caused by an HFD. Additionally, we included the hypothalamus as a structure known for the presence of MetS.
2. Results
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
2.1. Establishing a Model of Obesity
2.1.1. Body Weight, Food Consumption, and Adiposity
To evaluate the effects of a high-fat diet (HFD) in rats, weight gain and caloric intake were recorded over eight weeks. The pattern of obesity induced by a HFD was compared with the RM ANOVA results, which revealed significant differences in the interaction effect of week × diet (F(8,272) = 8.36, P < 0.0001), and we observed that HFD-fed rats gained more weight (126.3 ± 3.36 SEM) than did those fed a standard chow diet (98.02 ± 4.19 SEM) at week 4, and significant differences (P < 0.0001) were maintained until the end of the protocol (Figure 1A). However, the HFD-fed animals consumed more calories in the first two weeks than did the CTRL-fed animals did (Figure 1B), and the RM ANOVA revealed significant differences in the interaction of week × diet (F(8,272) = 4.23, P = 0.0001).
The proportion of adipose tissue in both groups was measured to determine whether the weight gain was due to fat mass expansion. Compared with those in the CTRL group, HFD-fed animals accumulated more epididymal (t=8.84, df=34, P < 0.0001) and subcutaneous (t=5.62, df=34, P = 0.0001) fat, as measured by weighing dissected fat pads after necropsy (Figure 1C). The size of the adipocytes increased (hypertrophy) in both the epididymal (t=2.94, df=10, P < 0.0147) and subcutaneous (t=2.39, df=10, P = 0.0379) tissues of the HFD-fed animals (Figure 1D–F). These findings demonstrate that the increased weight gain observed in HFD-fed rats is attributed to increased accumulation of epididymal and subcutaneous fat, alongside increased adipocyte hypertrophy.
2.1.2. Effects of an 8-Week HFD on Somatometric and Metabolic Modifications
Somatometric and metabolic parameters in both groups were summarized. As previously mentioned, HFD consumption induced increased body mass due to increased fat depots; in addition, HFD-fed rats presented an increased mass: body length ratio (determined as body mass index and the Lee index). Moreover, HFD-fed animals presented an increased waist circumference, which is noteworthy because central obesity is a crucial feature of MetS in humans. In addition to increased FPG, plasma analysis revealed that HFD-fed rats presented increased plasma leptin and insulin concentrations (Table 1). Moreover, HFD-fed animals presented an increased HOMA index. Eight weeks of HFD consumption led to somatometric and metabolic changes, marked by increased body mass, central obesity, and elevated plasma leptin and insulin levels and a high HOMA index, reflecting key features associated with metabolic syndrome.
FPG was measured to identify the presence of metabolic alterations, and an IPGTT and ITT were performed. The HFD-fed animals exhibited impaired glucose tolerance via the IPGTT (Figure 2 A and B), and the RM ANOVA revealed significant differences over time (F(5,70)= 38.11, P < 0.0001) and between groups (F(1,14)= 5.81, P = 0.03) but not in the interaction; the area under the curve for impaired glucose tolerance was greater for the HFD-fed group than for the CTRL group (t= 2.593, df =7, P = 0.0358). In the ITT (Figure 2C and D), the RM ANOVA revealed significant differences over time (F(2.797,34.68) = 15.03, P < 0.001) and between groups (F(1,14) = 16.60, P = 0.0011) but not in the interaction; the area under the curve was greater for the HFD-fed group than for the CTRL group (t= 2.706, df= 14, P = 0.0170). These results suggest that HFD treatment induced impaired glucose metabolism and was associated with typical signs of insulin resistance and MetS.
2.2. Behavioral Tests and Damage Induced by an HFD
Behavioral response assessment was performed via tests of pain and motor coordination. When mechanical allodynia via von Frey filaments was evaluated, rats fed a HFD for 8 weeks (obese rats) presented a significant (P < 0.05) decrease in the 50% PWT on both hind paws compared with those in the CTRL group (Figure 3A and B), indicating that rats fed a HFD for 8 weeks developed an increased nociceptive response to mechanical stimuli. When motor coordination was assessed via the rotarod with accelerating speed, obese rats also presented a significant (P < 0.05) decrease in latency to fall compared with that of CTRL (Figure 3C), indicating that the motor capacity of obese rats was impaired.
2.3. Evaluating BBB Integrity
Initially, to study brain tissue damage and assess morphological changes in cells in each neuroanatomical region, processed tissues were analyzed via hematoxylin‒eosin (H&E) staining. In HFD-fed mice, the cerebellar cortex (Figure 4B, H and N) presented an increased number of cells in the molecular layer. In the hippocampus (Figure 4 3D, J and P), the nuclei of neurons from the superior and inferior blades and the hinge of the dentate gyrus (DG) seemed to have more compacted chromatin and smaller neuropils. In the hypothalamus (Figure 4F, L and R), dilated vessels and many cell nuclei were more hyperchromatic in the HFD-fed group than in the CTRL group. HFD consumption led to notable morphological changes across multiple neuroanatomical regions, supporting the study's aim to investigate brain tissue damage induced by dietary factors.
Additionally, the penetration of Evans dye into the 3 tissues was significant for the cerebellum (t=4.543, df=4, P = 0.0105) of CTRL, with an average of 5.68 ± 0.64 SEM, and HFD-fed, with an average of 21.17 ± 3.35 SEM; the hippocampus (t=4.295, df=4, P = 0.0127) for CTRL, with an average of 5.94 ± 1.02 SEM, and HFD-fed, with an average of 11.96 ± 0.96 SEM; and the hypothalamus (t=4.212, df=4, P = 0.0136) for CTRL, with an average of 7.67± 2.06 SEM, and HFD-fed, with an average of 26.51 ± 3.98 SEM. We observed differences in the intensity of GFAP (Figure 5) across regions; greater intensity was observed in the cerebellum (Figure 5A vs. D; t=10.65, df=16, P < 0.0001), hippocampus (Figure 5B vs. E; t=5.867, df=14, P < 0.0001), and hypothalamus (Figure 5C vs. F; t=7.964, df=16, P < 0.0004) in the HFD-fed group than in the CTRL group. The increase in Evans dye penetration and GFAP intensity across the cerebellum, hippocampus, and hypothalamus in HFD-fed animals underscores the impact of a high-fat diet on blood‒brain barrier integrity and astrocyte activation.
The intensity also differed across regions for connexin-43 (Figure 5), with greater intensities for the HFD-fed group than for the CTRL group in the cerebellum (Figure 5H vs. K; t=72.04, df=16, P< 0.0001), hippocampus (Figure 5I vs. L; t=20.66, df=16, P < 0.0001), and hypothalamus (Figure 5J vs. M; t=32.92, df=16, P < 0.0001).
When we evaluated the ultrastructural level in those areas, we observed that in the cerebellum (Supplementary Figure S1), the endothelial cells of the blood vessel next to the Purkinje cell bodies had a similar thickness around the contour of the vessel, and the capillaries presented a continuous morphology. In HFD-fed mice, the endothelium of the blood vessels of the cerebellar cortex showed remarkable homogeneous thinning and capillaries, with thinning of endothelial cells and a loss of continuous and resembling fenestrated capillaries.
Hippocampal capillaries from normal rats presented characteristics of continuous capillaries with homogeneous endothelial thickness (Figure 6). The main ultrastructural alteration observed in HFD-treated rats was thinning of the capillary endothelium, many of which had numerous electron-lucent spaces, most likely occupied by interstitial fluid present in the neuropil. No ultrastructural differences in neuronal nuclei were observed between the CTRL-fed and HFD-fed groups.
Finally, regarding the capillaries in the hypothalamus (Figure 7), we observed the morphology of continuous capillaries and nuclei with abundant euchromatin, and heterochromatin associated with the nuclear envelope (Figure 7A). In HFD-fed mice, the thickness of the cytoplasm of endothelial cells decreased (Figure 7C), and alterations in the nuclear chromatin condensation pattern were detected (Figure 7C). Some capillaries in these animals were partially surrounded by clear spaces, which were probably occupied by tissue fluid (Figure 7C). Numerous neurons presented nuclei with prominent nucleoli in normal rats (Figure 7B), whereas the neurons from HFD-treated rats presented swollen nuclei and no nucleoli or karyolysis (Figure 7D). Some neurons were highly dense in electrons (Figure 7D). HFD consumption led to alterations in capillary and neuronal morphology in the cerebellum, hippocampus, and hypothalamus, including decreased endothelial cell cytoplasm thickness, changes in nuclear chromatin condensation, and signs of neuronal damage.
3. Discussion
In this study, we aimed to investigate the structure‒function relationship between allodynia and motor impairment in rats fed a high-fat diet (HFD) under conditions of obesity and metabolic syndrome (MetS). We demonstrated that, within a short period (8 weeks), a HFD (60% of daily intake) leads to significant changes in metabolism and triggers the development of MetS, with impacts on allodynia and motor activity, as well as changes in the BBB, GFAP, and connexin-43 intensity and ultrastructural modifications in key brain regions, including the hippocampus, cerebellum, and hypothalamus. This highlights the profound impact of a HFD on both the peripheral and central nervous systems.
Our results align with observations from other research groups who reported that a high-fat commercial diet [26], induces obesity [27,28,29]. An interesting precedent to this work is the findings of Buettner and colleagues, who reported that a HFD using lard and olive oil significantly increased the weight of rats compared with that of CTRL [28]. This finding is reasonable because of the caloric content of fats (1 g of fat equals 9 kcal). In our study, the weight of the animals increased despite decreased food intake (Figure 1). Additionally, the effects of HFD on the development of obesity depend on the duration of treatment, ranging from 4 to 40 weeks, and the percentage of calories from fat, ranging from 37% to 60% [30]. Hu et al [31] revealed that after four weeks of consuming a HFD (with 60% of calories from lipids), the body weights of the animals did not significantly differ between groups. However, in our results, which were obtained via an 8-week HFD (60%), the change in weight gain began at four weeks. This change is consistent with previous findings and the study by Hu et al., where rats fed a HFD accumulated more adipose tissue in the gonadal and visceral regions. Our data confirm that the duration and fat content of a HFD are critical factors in the development of obesity, with significant weight gain observed as early as four weeks. These findings align with those of previous studies and underscore the role of high-fat diets in adipose tissue accumulation and obesity onset.
Saturated fats induce insulin resistance and other metabolic defects, whereas monounsaturated fats are considered protective agents against cardiovascular and metabolic alterations.40 However, previous studies have shown that olive oil or lard have nearly similar metabolic effects when they are used in high percentages [32]. Indeed, chronic, long-term (11 weeks) high-fat intake may subsequently promote the development of MetS [33,34]. Despite anticipating increased liver lipid accumulation, our acute 8-week exposure model did not produce significant changes in this parameter (data not shown). However, metabolic alterations were evident, as indicated by the significant increase in HOMA-IR, reflecting impaired insulin sensitivity associated with the high-fat diet.
Previous studies have suggested that mechanical stress affects skeletal tissues, leading to chronic and painful disorders [35], and on the basis of the results obtained in our study, we propose that overweight is a significant contributor to pain. Specifically, hypersensitivity to mechanical stimulation is a typical sign of neuropathic pain [36]; a study showed that obese animals fed a Western diet (high fat, high sucrose, high cholesterol) for eight weeks experienced this type of pain [21]. Our findings revealed that an 8-week HFD resulted in mechanical allodynia, with a sensitivity to von Frey filaments approximately 40% greater than that in CTRL rats. Compared with the CTRL group, the HFD-fed group of Sprague‒Dawley rats presented a lower mechanical threshold after eight weeks (significantly lower after ten weeks), which persisted after 12 weeks [22]. Numerous studies have examined the nociceptive response to mechanical allodynia in mice of different C57BL/6 strains and at various feeding durations (6, 7, 8, and 16 weeks), and a significant reduction in the mechanical threshold was observed [37,38,39]. According to Guillford and colleagues, mechanical sensitivity increased in nondiabetic C57BL/6 mice after eight weeks on a HFD compared with that in CTRL-fed C57BL/6 mice [40]; in contrast, our model exhibited decreased mechanical sensitivity.
In addition, the accelerating or constant-speed rotarod test is commonly used to diagnose neurological injuries through motor coordination impairment [41]. As performing a task requires both kinematic (spatial direction) and kinetic (force) bodily movements, motor coordination is needed [23]. In our research, rats fed a HFD for eight weeks presented greater neurological and motor coordination impairments than did rats fed a healthy diet. This decline in motor coordination has been observed in Wistar rats as early as four weeks [23], 5 weeks in C57BL/6J mice [42], 12 weeks in Wistar rats [43], and 12 weeks [44] to 22 weeks in C57BL/6J mice [25]. Our research aligns with previous studies demonstrating that a HFD leads to motor coordination impairments, as evidenced by the rotarod test, reinforcing the utility of this test in diagnosing neurological deficits associated with metabolic disturbances.
In this study, we demonstrated that physiological changes, such as allodynia and motor alterations, are closely associated with morphological changes in BBB permeability and vary according to extrahypothalamic structure (see Figure 5). In this context, we detected differences when we analysed the cerebellum, hippocampus, and hypothalamus. These differences are so complex that the close relationships between the major cellular components of the BBB, such as endothelial cells and astrocytic cells, must be understood. Our GFAP data align with those of Valdez and colleagues [45], who reported the absence of reactive microglia in the hypothalamus, suggesting that there was no neuroinflammatory response in animals exposed to an olive oil-rich diet. However, according to Severi and colleagues [46], our ultrastructural data revealed thinning of the capillary endothelium in HFD-treated rats, many of which had numerous electron-lucent spaces in the hypothalamus. In our study, we observed similar changes in the hippocampus, and previous studies in animal models have shown that HFDs induce neuroinflammation in the hippocampus, which is associated with pain-related behaviours [47,48]. Our model highlights the relationship between physiological changes, such as allodynia and motor alterations, and morphological changes in BBB permeability across different brain structures. These results underscore the importance of understanding the interactions between BBB components, such as endothelial and astrocytic cells, in the context of diet-induced neuroinflammation and its impact on neurological functions.
Recent research on the hippocampus of adult offspring revealed a positive correlation between GFAP and the gap junction protein connexin-43 in response to a HFD [12]. In our study, the observed decrease in GFAP intensity was associated with a decrease in CX43, including in the hippocampus, cerebellum, and hypothalamus. However, ultrastructural analysis reveals an irregular profile and is partially surrounded by clear spaces such as those described previously in the median eminence [49], hippocampus [50], and cerebellum [19]; these morphological changes could explain the alterations in functional tests.
While our ultrastructural analysis provided valuable insights, it was limited to specific brain regions, leaving the potential effects on other regions unexplored. The partial decrease in GFAP and connexin-43 (CX43) expression observed only in the hippocampus, cerebellum, and hypothalamus may reflect region-specific responses. Despite these limitations, the findings of this study are robust, aligned with the literature, and provide consistent data on HFD-induced alterations in the structural and functional integrity of the brain. Expanding the ultrastructural analysis to include other brain regions and incorporating a broader array of neuroinflammatory markers could provide a comprehensive understanding of the brain's response to an HFD.
4. Materials and Methods
Animals and general housing conditions
Adult male Wistar rats with weighing 250–300 g was obtained from the vivarium of the School of Medicine at Universidad Nacional Autónoma de México. For acclimatization, the animals were housed in pairs and maintained in a soundproof monitoring room with a light/dark (LD) cycle (lights on at 07:00 h). The space was maintained at 22 ± 1°C, 50% humidity, and continuous airflow. The rats had free access to food with standard chow (Laboratory Rodent Diet 5001, LabDiet®; Minneapolis, MN, USA), and water. The testing procedures used in this study were in strict accordance with the Mexican norms for animal handling Norma Official Mexicana NOM-062-ZOO-1999, which conforms to international guidelines for animal handling and is approved by the Ethics Committee of the Medicine School UNAM FMED/DI/067/2019 06082019. Furthermore, all efforts were made to minimize the number of animals and their suffering. Human endpoints were established for this study (Supplementary Table S1).
4.1. Establishing a Model of Obesity
4.1.1. High Fat Diet
The HFD was prepared with the control diet as a base, adding fats of animal and vegetable origins. The preparation of the HFD [51] (100 g) consisted of pulverizing Laboratory Rodent Diet 5001 pellets (50 g), to which lard (17.5 g), olive oil (17.5 g) and lyophilized egg albumin (15 g) were added. Fatty acid analysis of the paste obtained was performed (Supplementary Table S2).
4.1.2. Experimental Design
A total of 36 rats (the experimental unit) were used for this study. By simple randomization method, the rats were randomly assigned to one of two conditions: the CTRL group (n = 18) or the HFD group (HFD-fed, n = 18). After two weeks of acclimatization with free access to food (Laboratory Rodent Diet 5001) and water, the rats in the experimental group were fed a HFD for eight weeks. Food consumption and weight were quantified with a digital weighing scale (Kent Scientific Corporation) each week throughout the testing period. One animal in each group were excluded in the analysis (Table 1) because died during the insulin test.
4.1.3. Glucose Tolerance Test (IPGTT) and Insulin Tolerance Test (ITT)
At the end of the eight weeks of treatment, eight animals in each group were fasted for 12 hours to determine their fasting plasma glucose (FPG) levels, and IPGTTs and ITTs were performed. For the IPGTT, 2 grams of glucose/kg body weight was administered intraperitoneally; for the ITT, 0.2 IU of human recombinant insulin (Humulin, Lilly; Indianapolis, IN, USA) was administered intraperitoneally. Blood glucose was measured at 0, 30, 60, 90, and 120 minutes after the intervention via a handheld blood glucose monitor (OneTouch® UltraMini®, Pennsylvania, USA).
4.1.4. Insulin Resistance Surrogate Measures
Insulin resistance was estimated according to the Homeostatic Model Assessment, which was previously adapted for use in Wistar rats [52]: HOMAir = (FPG (mg/dL) * FPI (µUI/ml))/2,430.
Insulin concentrations (pmol/L) were converted for use in the above formula: 1 µUI/ml = 6 pmol/L [53].
4.1.5. Plasma, Adipose Tissue, and Brain Samples
At the end of the eight weeks of treatment, 18 animals in each group, under anesthesia with intraperitoneal (i.p.) sodium pentobarbital (Pisabental®, Aranda; 100 mg/kg; Mexico City, Mexico), epidydimal and subcutaneous fat pads were obtained. The weight of the fat pads was reported as a proportion of the total body weight. Additionally (n =11 by group), 5 ml of blood was drawn from the inferior vena cava into tubes with EDTA. To obtain plasma and measure glucose, leptin, and insulin concentrations, the blood was immediately centrifuged at 1800 rpm for 10 minutes at room temperature and subsequently frozen in 0.2 ml aliquots at -80°C until analysis. The brain, liver, and adipose tissue were removed. Leptin and insulin levels were determined via a Milliplex® MAP Rat Adipokine Magnetic Bead Panel (RADPKMAG-80K; Merck Millipore; Hesse, Germany) according to the manufacturer’s instructions.
4.1.6. Histological Preparation
The brain and adipose tissue were washed in ice-cold phosphate-buffered saline (PBS), followed by the addition of 4% paraformaldehyde in PBS at 4°C. After fixation, the tissues were dehydrated and embedded in paraffin for sectioning.
These tissues were analyzed via hematoxylin‒eosin-stained paraffin sections (cut into four-μm-thick sections) from both groups.
4.2. Behavioral Tests and Damage Induced by an HFD
4.2.1. Mechanical Allodynia
Behavioral tests were performed in all the rats at the basal stage (day zero) and after 8 weeks in the CTRL (n =6) and HFD-fed (n =6) rats, according to a method published by Chaplan [54]. Von Frey filaments were used to stimulate the plantar aspect of the foot, and the 50% paw withdrawal threshold (PWT) was determined via the up-and-down method [55].
4.2.2. Motor Coordination
All the rats were trained with a 5-minute run at a steady speed of 4 rpm for 3 days before testing. The next day, the animals were placed in the rotating cylinder at a rotational speed that was increased from 4 rpm to 40 rpm for 5 minutes. The latency to fall was recorded up to a limit of 300 seconds [56].
4.3. Evaluating BBB Integrity
4.3.1. Evans Blue Administration
After the experiment, three rats in each group were anaesthetized via an intraperitoneal injection of 60 mg/kg sodium pentobarbital. A 2 cm thoracic incision was made to expose the thoracic cavity. The rats were then perfused intracardially with 250 ml of 0.9% saline solution, followed by 4% paraformaldehyde (PFA) prepared in a buffered 0.1 M sodium phosphate pH 7.2 (PB) solution plus Evans blue [57].
4.3.2. Immunofluorescence
To conduct immunofluorescence, the extracted brains were postfixed in 4% PFA at 4°C for 24 hours. Afterwards, they were cryoprotected in a 30% sucrose solution. The tissue was embedded in optimal cutting temperature (O.C.T.) compound (FreezeMount, USA) and processed under freezing conditions. A cryostat (Leica CM1100) was used to obtain four series of 20-micrometer-thick coronal sections. Sections were collected in two series: one series was processed for GFAP, and the other series was processed for Connnexin-43. Free floating sections were blocked with 2% albumin, and the primary antibodies mouse anti-GFAP (1:100; PM065AA, BioCare Medical, CA, USA) or anti-connexin 43 (F-7) (1:50; SC-271837, Santa Cruz, TX, USA) were added and incubated overnight at 4°C. Each well was then washed four times for ten minutes with 1X PBS while agitated. The secondary antibody, goat anti-mouse IgG (1:100; 62-6511, Invitrogen, MA, USA), was incubated at room temperature for two hours in the dark. DAPI-containing mounting medium was added (VECTASHIELD®, CA, USA). The cover slip, which was previously cleaned with gauze and alcohol, was placed on the slide 5 minutes later. The edge was protected by a transparent varnish coating. After microscopic examination, the samples were stored at 4°C.
4.3.3. Image Digitalization
The slides were examined via a Zeiss (LSM 880 Zeiss; Oberkochen, Germany) confocal laser scanning microscope, and the laser intensity and digital gain were held constant between groups. Using the ImageJ software package, the marker density, corrected fluorescence, morphological parameters of the cells, and cell type were determined from the images.
4.3.4. Transmission Electron Microscopy
Three rats (by group) were anaesthetized i.p. with sodium pentobarbital (Pisabental®, Aranda; 100 mg/kg; Mexico City, Mexico), and their whole bodies were fixed via a gravity-fed perfusion apparatus with two bottles interconnected with three-way keys, one containing heparinized physiological saline solution (PSS) and the other containing 2.5% glutaraldehyde fixative solution in 0.1 M sodium cacodylate buffer (pH 7.4). This apparatus was placed approximately 130 cm above the animal [58]. A thoracotomy was carried out to insert a needle into the left ventricle of the heart. Afterwards, a cut was made in the right atrium allowing the exit of blood, whereas the blood was removed from the circulatory system with the perfusion PSS for 20 min. When only PSS exited from the right atrium, the fixative solution was perfused for 20 min. Next, the rats were decapitated to remove the brains from the skull. Manual brain samples (1 mm3 each) containing the cerebellum cortex, hippocampus, and hypothalamus (near the III ventricle) were obtained and immersed in the same fixative for 3 h at 4°C. The samples were then washed in the same buffer (0.2 M, pH 7.4) and postfixed in 1% osmium tetroxide diluted in the same buffer for 1 h at 4°C. The samples were dehydrated in gradually increasing ethanol concentrations, transferred to propylene oxide, and embedded in EPON 812 embedding medium. Thin 0.9-μm-thick sections obtained with a glass knife were stained with toluidine blue in borax to select the microscopic field from which ultrathin sections were cut with a diamond knife. Ultrathin sections were mounted on copper grids, contrasted with 2% aqueous uranyl acetate and 2% plumber citrate, and observed with a Zeiss STEM electron microscope (Crossbeam 550). Six ultrathin sections were analyzed for each CNS region.
4.3.5. Data Analysis
GraphPad Prism software (GraphPad Software Inc., San Diego, CA, United States) was used for the statistical analyses. Descriptive statistics are presented as the mean ± standard error of the mean (S.E.M.) for the quantitative variables BMI, abdominal circumference, body length, Lee index, FPG, leptin, insulin, triacylglycerols, HOMA-IR, epididymal fat, subcutaneous fat, optical density of Evans blue, quantification of GFAP, and CX43 staining intensity. To quantify the global response of glucose and insulin in the blood after the test, area under the curve (AUC) analysis was performed. The data were tested for normality with the Shapiro‒Wilk test. When suitable, differences between groups were assessed through unpaired t tests or with Mann‒Whitney U tests when the data did not meet the normality assumption.
Statistical differences between groups in terms of body weight gain, diet consumption, and IPGTT and ITT results were determined via repeated measures (RM) ANOVA followed by Sidak’s multiple comparisons test or one- or two-way ANOVA followed by Tukey’s test or the Kruskal‒Wallis test followed by the Dunn test for variables that were not normally distributed.
A sample size calculation was performed to detect a 75% difference in altered behavior proportions between the control group and the high-fat diet group, with 95% confidence and 80% power, yielding a sample size of 6. Mechanical allodynia and motor coordination data are presented as the means ± S.E.M. of 6 independent animals. Between-group differences were assessed via one-way analysis of variance (ANOVA) or the Kruskal‒Wallis test followed by the Mann‒Whitney U test. P values < 0.05 were considered to indicate statistical significance.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Transmission electron micrograph of cerebellum cortex; Table S1: Guidelines Human Endpoints; Table S2: Fatty acid composition of the High Fat Diet.
Author Contributions
Conceptualization, L.M.U.-R., and E. E.-B.; Data curation, L.M.U.-R.; R. M.-T., A. B.M., R.E., J. C.-H., V. R.-M. ,and A.P.-T.; Formal analysis, L.M.U.-R., A. B.M., R. N.-N.; J. C.-H., and L. N.; Funding acquisition, L.M.U.-R., and L. N.; Investigation, L.M.U.-R., R. N.R., and R.E.; Validation, L.M.U.-R., E. E-B., and R.E.; Investigation, L.M.U.-R., A. B.M., R. N.-N., R.E., and A. P.-T.; Methodology, L.M.U.-R.; Resources, L.M.U.-R., and L. N.; Supervision, L.M.U.-R., Validation, L.M.U.-R., Writing—original draft preparation, L.M.U.-R.; Writing—Review and Editing, L.M.U.-R., E. E-B., R. M.-T.,A. B.M., and R.E.; visualization, L.M.U.-R., J. C.-H., and V. R.-M.; project administration, L.M.U.-R.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT), grant numbers IA207321. Additionally, we thank the support for the project FMED/DI/067/2019 from División de Investigación Facultad de Medicina, UNAM.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of the Medicine School UNAM (protocol code 063-CIC-2019; date of approval 08 October 2019).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We would like to extend our gratitude to Pamela Pedraza Flores, for help and animal care. Dana León Aguilar for help during immunofluorescence.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Levesque:, R.J.R. Obesity and Overweight. In Encyclopedia of Adolescence; Springer International Publishing, 2018; pp. 2561–2565.
- Cornier, M.A.; Dabelea, D.; Hernandez, T.L.; Lindstrom, R.C.; Steig, A.J.; Stob, N.R.; Van Pelt, R.E.; Wang, H.; Eckel, R.H. The Metabolic Syndrome. Endocr. Rev. 2008, 29, 777–822. [Google Scholar] [CrossRef] [PubMed]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; De Angelis, M.H.; Schürmann, A.; et al. Animal Models of Obesity and Diabetes Mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef]
- Small, L.; Brandon, A.E.; Turner, N.; Cooney, G.J. Modeling Insulin Resistance in Rodents by Alterations in Diet: What Have High-Fat and High-Calorie Diets Revealed? Am. J. Physiol. - Endocrinol. Metab. 2018, 314, E251–E265. [Google Scholar] [CrossRef] [PubMed]
- Keita, H.; Ramírez-San Juan, E.; Paniagua-Castro, N.; Garduño-Siciliano, L.; Quevedo, L. The Long-Term Ingestion of a Diet High in Extra Virgin Olive Oil Produces Obesity and Insulin Resistance but Protects Endothelial Function in Rats: A Preliminary Study. Diabetol. Metab. Syndr. 2013, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Larqué, C.; Lugo-Martínez, H.; Mendoza, X.; Nochebuena, M.; Novo, L.; Vilchis, R.; Sánchez-Bringas, G.; Ubaldo, L.; Velasco, M.; Escalona, R. Paternal Obesity Induced by High-Fat Diet Impairs the Metabolic and Reproductive Health of Progeny in Rats. Reprod. Heal. Progeny Rats. Metab. 2023, 13, 1098. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.L.; Norhaizan, M.E. Effect of High-Fat Diets on Oxidative Stress, Cellular Inflammatory Response and Cognitive Function. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- Lama, A.; Pirozzi, C.; Severi, I.; Morgese, M.G.; Senzacqua, M.; Annunziata, C.; Comella, F.; Del Piano, F.; Schiavone, S.; Petrosino, S.; et al. Palmitoylethanolamide Dampens Neuroinflammation and Anxiety-like Behavior in Obese Mice. Brain. Behav. Immun. 2022, 102, 110–123. [Google Scholar] [CrossRef]
- Hargrave, S.L.; Davidson, T.L.; Zheng, W.; Kinzig, K.P. Western Diets Induce Blood-Brain Barrier Leakage and Alter Spatial Strategies in Rats. Behav. Neurosci. 2016, 130, 123–135. [Google Scholar] [CrossRef]
- Machida, T.; Takata, F.; Matsumoto, J.; Miyamura, T.; Hirata, R.; Kimura, I.; Kataoka, Y.; Dohgu, S.; Yamauchi, A. Contribution of Thrombin-Reactive Brain Pericytes to Blood-Brain Barrier Dysfunction in an in Vivo Mouse Model of Obesity-Associated Diabetes and an in Vitro Rat Model. PLoS One 2017, 12. [Google Scholar] [CrossRef]
- Ogata, S.; Ito, S.; Masuda, T.; Ohtsuki, S. Changes of Blood-Brain Barrier and Brain Parenchymal Protein Expression Levels of Mice under Different Insulin-Resistance Conditions Induced by High-Fat Diet. Pharm. Res. 2019, 36, 141. [Google Scholar] [CrossRef]
- Shiadeh, S.M.J.; Goretta, F.; Svedin, P.; Jansson, T.; Mallard, C.; Ardalan, M. Long-Term Impact of Maternal Obesity on the Gliovascular Unit and Ephrin Signaling in the Hippocampus of Adult Offspring. J. Neuroinflammation 2024, 21. [Google Scholar] [CrossRef] [PubMed]
- Tucsek, Z.; Toth, P.; Tarantini, S.; Sosnowska, D.; Gautam, T.; Warrington, J.P.; Giles, C.B.; Wren, J.D.; Koller, A.; Ballabh, P.; et al. Aging Exacerbates Obesity-Induced Cerebromicrovascular Rarefaction, Neurovascular Uncoupling, and Cognitive Decline in Mice. J. Gerontol. A. Biol. Sci. Med. Sci. 2014, 69, 1339–1352. [Google Scholar] [CrossRef]
- Salameh, T.S.; Mortell, W.G.; Logsdon, A.F.; Butterfield, D.A.; Banks, W.A. Disruption of the Hippocampal and Hypothalamic Blood-Brain Barrier in a Diet-Induced Obese Model of Type II Diabetes: Prevention and Treatment by the Mitochondrial Carbonic Anhydrase Inhibitor, Topiramate. Fluids Barriers CNS 2019, 16, 1. [Google Scholar] [CrossRef]
- Kim, D.W.; Glendining, K.A.; Grattan, D.R.; Jasoni, C.L. Maternal Obesity in the Mouse Compromises the Blood-Brain Barrier in the Arcuate Nucleus of Offspring. Endocrinology 2016, 157, 2229–2242. [Google Scholar] [CrossRef]
- Higuchi, S.; Irie, K.; Mishima, S.; Araki, M.; Ohji, M.; Shirakawa, A.; Akitake, Y.; Matsuyama, K.; Mishima, K.; Mishima, K.; et al. The Cannabinoid 1-Receptor Silent Antagonist O-2050 Attenuates Preference for High-Fat Diet and Activated Astrocytes in Mice. J. Pharmacol. Sci. 2010, 112, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.; Joaquim, A.; Bonamin, L.V.; Martins, M. de F.M.; Kirsten, T.B.; Cardoso, C.V.; Bernardi, M.M.; Bondan, E.F. Reduced Astrocytic Expression of GFAP in the Offspring of Female Rats That Received Hypercaloric Diet. Nutr. Neurosci. 2020, 23, 411–421. [Google Scholar] [CrossRef]
- Severi, I.; Fosca, M.; Colleluori, G.; Marini, F.; Imperatori, L.; Senzacqua, M.; Di Vincenzo, A.; Barbatelli, G.; Fiori, F.; Rau, J. V.; et al. High-Fat Diet Impairs Mouse Median Eminence: A Study by Transmission and Scanning Electron Microscopy Coupled with Raman Spectroscopy. Int. J. Mol. Sci. 2021, 22, 8049. [Google Scholar] [CrossRef] [PubMed]
- El-Sayyad, H.I.H.; El-Gallil, H.A.; El-Ghaweet, H.A. Synergistic Effects of Pomegranate Juice and Atorvastatin for Improving Cerebellar Structure and Function of Breast-Feeding Rats Maternally Fed on a High Cholesterol Diet. J. Chem. Neuroanat. 2020, 107. [Google Scholar] [CrossRef]
- Woo, C.W.; Schmidt, L.; Krishnan, A.; Jepma, M.; Roy, M.; Lindquist, M.A.; Atlas, L.Y.; Wager, T.D. Quantifying Cerebral Contributions to Pain beyond Nociception. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Gavini, C.K.; Bookout, A.L.; Bonomo, R.; Gautron, L.; Lee, S.; Mansuy-Aubert, V. Liver X Receptors Protect Dorsal Root Ganglia from Obesity-Induced Endoplasmic Reticulum Stress and Mechanical Allodynia. Cell Rep. 2018, 25, 271–277.e4. [Google Scholar] [CrossRef]
- Guo, X.; Tao, X.; Tong, Q.; Li, T.; Dong, D.; Zhang, B.; Zhao, M.; Song, T. Impaired AMPK-CGRP Signaling in the Central Nervous System Contributes to Enhanced Neuropathic Pain in High-fat Diet-induced Obese Rats, with or without Nerve Injury. Mol. Med. Rep. 2019, 20, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Uppin, V.; Acharya, P.; Kempaiah, B.B.; Talahalli, R.R. Zerumbone Augments Cognitive Enhancement Potentials of EPA+DHA: Insight from a Hyperlipidaemic Rat Model. Br. J. Nutr. 2020, 124, 1353–1360. [Google Scholar] [CrossRef]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.J.; et al. Time-Restricted Feeding without Reducing Caloric Intake Prevents Metabolic Diseases in Mice Fed a High-Fat Diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef]
- Fajardo, L.; Sanchez, P.; Salles, J.; Rigaudière, J.P.; Patrac, V.; Caspar-Bauguil, S.; Bergoglgio, C.; Moro, C.; Walrand, S.; Le Bacquer, O. Inhibition of the Endocannabinoid System Reverses Obese Phenotype in Aged Mice and Partly Restores Skeletal Muscle Function. Am. J. Physiol. Endocrinol. Metab. 2023, 324, E176–E184. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.R. Use of High-Fat Diets to Study Rodent Obesity as a Model of Human Obesity. Int. J. Obes. 2019, 43, 1491–1492. [Google Scholar] [CrossRef] [PubMed]
- Boustany, C.M.; Bharadwaj, K.; Daugherty, A.; Brown, D.R.; Randall, D.C.; Cassis, L.A. Activation of the Systemic and Adipose Renin-Angiotensin System in Rats with Diet-Induced Obesity and Hypertension. Am. J. Physiol. - Regul. Integr. Comp. Physiol. 2004, 287. [Google Scholar] [CrossRef]
- Buettner, R.; Schölmerich, J.; Bollheimer, L.C. High-Fat Diets: Modeling the Metabolic Disorders of Human Obesity in Rodents. Obesity (Silver Spring). 2007, 15, 798–808. [Google Scholar] [CrossRef]
- Suman, R.K.; Ray Mohanty, I.; Borde, M.K.; Maheshwari, U.; Deshmukh, Y.A. Development of an Experimental Model of Diabetes Co-Existing with Metabolic Syndrome in Rats. Adv. Pharmacol. Sci. 2016, 2016. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Fairus, A.; Ima-Nirwana, S. Animal Models of Metabolic Syndrome: A Review. Nutr. Metab. (Lond). 2016, 13, 65. [Google Scholar] [CrossRef]
- Hu, S.; Wang, L.; Yang, D.; Li, L.; Togo, J.; Wu, Y.; Liu, Q.; Li, B.; Li, M.; Wang, G.; et al. Dietary Fat, but Not Protein or Carbohydrate, Regulates Energy Intake and Causes Adiposity in Mice. Cell Metab. 2018, 28, 415–431.e4. [Google Scholar] [CrossRef]
- Buettner, R.; Parhofer, K.G.; Woenckhaus, M.; Wrede, C.E.; Kunz-Schughart, L.A.; Schölmerich, J.; Bollheimer, L.C. Defining High-Fat-Diet Rat Models: Metabolic and Molecular Effects of Different Fat Types. J. Mol. Endocrinol. 2006, 36, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Kettner, N.M.; Mayo, S.A.; Hua, J.; Lee, C.; Moore, D.D.; Fu, L. Circadian Dysfunction Induces Leptin Resistance in Mice. Cell Metab. 2015, 22, 448–459. [Google Scholar] [CrossRef]
- Mwangi, S.M.; Nezami, B.G.; Obukwelu, B.; Anitha, M.; Marri, S.; Fu, P.; Epperson, M.F.; Le, N.A.; Shanmugam, M.; Sitaraman, S. V.; et al. Glial Cell Line-Derived Neurotrophic Factor Protects against High-Fat Diet-Induced Obesity. Am. J. Physiol. - Gastrointest. Liver Physiol. 2014, 306. [Google Scholar] [CrossRef]
- Marques Miranda, C.; de Lima Campos, M.; Leite-Almeida, H. Diet, Body Weight and Pain Susceptibility – A Systematic Review of Preclinical Studies. Neurobiol. Pain 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.; Binder, A.; Wasner, G. Neuropathic Pain: Diagnosis, Pathophysiological Mechanisms, and Treatment. Lancet Neurol. 2010, 9, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; O’Meara, B.; Jack, M.M.; Elliot, D.; Lamb, B.; Khan, Z.W.; Menta, B.W.; Ryals, J.M.; Winter, M.K.; Wright, D.E. Intrinsic Activity of C57BL/6 Substrains Associates with High-Fat Diet-Induced Mechanical Sensitivity in Mice. J. Pain 2018, 19, 1285–1295. [Google Scholar] [CrossRef]
- Liang, Y.J.; Feng, S.Y.; Qi, Y.P.; Li, K.; Jin, Z.R.; Jing, H.B.; Liu, L.Y.; Cai, J.; Xing, G.G.; Fu, K.Y. Contribution of Microglial Reaction to Increased Nociceptive Responses in High-Fat-Diet (HFD)-Induced Obesity in Male Mice. Brain. Behav. Immun. 2019, 80, 777–792. [Google Scholar] [CrossRef] [PubMed]
- Lupachyk, S.; Watcho, P.; Obrosov, A.A.; Stavniichuk, R.; Obrosova, I.G. Endoplasmic Reticulum Stress Contributes to Prediabetic Peripheral Neuropathy. Exp. Neurol. 2013, 247, 342–348. [Google Scholar] [CrossRef]
- Guilford, B.L.; Ryals, J.M.; Wright, D.E. Phenotypic Changes in Diabetic Neuropathy Induced by a High-Fat Diet in Diabetic C57Bl/6 Mice. Exp. Diabetes Res. 2011, 2011, 18–21. [Google Scholar] [CrossRef]
- Brooks, S.P.; Dunnett, S.B. Tests to Assess Motor Phenotype in Mice: A User’s Guide. Nat. Rev. Neurosci. 2009, 10, 519–529. [Google Scholar] [CrossRef]
- Grover, L.; Sklioutovskaya-Lopez, K.; Parkman, J.K.; Wang, K.; Hendricks, E.; Adams-Duffield, J.; Kim, J.H. Diet, Sex, and Genetic Predisposition to Obesity and Type 2 Diabetes Modulate Motor and Anxiety-Related Behaviors in Mice, and Alter Cerebellar Gene Expression. Behav. Brain Res. 2023, 445. [Google Scholar] [CrossRef]
- Bhandari, A.; Kalotra, S.; Bajaj, P.; Sunkaria, A.; Kaur, G. Dietary Intervention with Tinospora Cordifolia Improved Aging-Related Decline in Locomotor Coordination and Cerebellar Cell Survival and Plasticity in Female Rats. Biogerontology 2022, 23, 809–824. [Google Scholar] [CrossRef] [PubMed]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E. a.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J. a J.; et al. Time-Restricted Feeding without Reducing Caloric Intake Prevents Metabolic Diseases in Mice Fed a High-Fat Diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [PubMed]
- Valdez, M.C.; Freeborn, D.; Valdez, J.M.; Johnstone, A.F.M.; Snow, S.J.; Tennant, A.H.; Kodavanti, U.P.; Kodavanti, P.R.S. Mitochondrial Bioenergetics in Brain Following Ozone Exposure in Rats Maintained on Coconut, Fish and Olive Oil-Rich Diets. Int. J. Mol. Sci. 2019, Vol. 20, Page 6303 2019, 20, 6303. [Google Scholar] [CrossRef] [PubMed]
- Severi, I.; Fosca, M.; Colleluori, G.; Marini, F.; Imperatori, L.; Senzacqua, M.; Di Vincenzo, A.; Barbatelli, G.; Fiori, F.; Rau, J. V.; et al. High-Fat Diet Impairs Mouse Median Eminence: A Study by Transmission and Scanning Electron Microscopy Coupled with Raman Spectroscopy. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Boitard, C.; Cavaroc, A.; Sauvant, J.; Aubert, A.; Castanon, N.; Layé, S.; Ferreira, G. Impairment of Hippocampal-Dependent Memory Induced by Juvenile High-Fat Diet Intake Is Associated with Enhanced Hippocampal Inflammation in Rats. Brain. Behav. Immun. 2014, 40, 9–17. [Google Scholar] [CrossRef]
- Guillemot-Legris, O.; Muccioli, G.G. Obesity-Induced Neuroinflammation: Beyond the Hypothalamus. Trends Neurosci. 2017, 40, 237–253. [Google Scholar] [CrossRef]
- Severi, I.; Fosca, M.; Colleluori, G.; Marini, F.; Imperatori, L.; Senzacqua, M.; Di Vincenzo, A.; Barbatelli, G.; Fiori, F.; Rau, J. V.; et al. High-Fat Diet Impairs Mouse Median Eminence: A Study by Transmission and Scanning Electron Microscopy Coupled with Raman Spectroscopy. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Ding, H.; Liu, C.; Zhang, S.; Li, B.; Xu, Q.; Shi, B.; Li, S.; Dong, S.; Ma, X.; Zhang, Y.; et al. Sleeve Gastrectomy Attenuated Diabetes-Related Cognitive Decline in Diabetic Rats. Front. Endocrinol. (Lausanne). 2022, 13. [Google Scholar] [CrossRef]
- Escalona, R.; Larqué, C.; Cortes, D.; Vilchis, R.; Granados-Delgado, E.; Sánchez, A.; Sánchez-Bringas, G.; Lugo-Martínez, H. High-Fat Diet Impairs Glucose Homeostasis by Increased P16 Beta-Cell Expression and Alters Glucose Homeostasis of the Progeny in a Parental-Sex Dependent Manner. Front. Endocrinol. (Lausanne). 2023, 14, 1246194. [Google Scholar] [CrossRef]
- Cacho, J.; Sevillano, J.; de Castro, J.; Herrera, E.; Ramos, M.P. Validation of Simple Indexes to Assess Insulin Sensitivity during Pregnancy in Wistar and Sprague-Dawley Rats. Am. J. Physiol. Metab. 2008, 295, E1269–E1276. [Google Scholar] [CrossRef] [PubMed]
- Knopp, J.L.; Holder-Pearson, L.; Chase, J.G. Insulin Units and Conversion Factors: A Story of Truth, Boots, and Faster Half-Truths. J. Diabetes Sci. Technol. 2019, 13, 597–600. [Google Scholar] [CrossRef] [PubMed]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative Assessment of Tactile Allodynia in the Rat Paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Dixon, W.J. Efficient Analysis of Experimental Observations. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 441–462. [Google Scholar] [CrossRef]
- Harauma, A.; Tomita, M.; Muto, D.; Moriguchi, T. Effect of Long-Term Administration of Arachidonic Acid on n-3 Fatty Acid Deficient Mice. Prostaglandins Leukot. Essent. Fat. Acids 2015, 95, 41–45. [Google Scholar] [CrossRef]
- del Valle, J.; Camins, A.; Pallàs, M.; Vilaplana, J.; Pelegrí, C. A New Method for Determining Blood-Brain Barrier Integrity Based on Intracardiac Perfusion of an Evans Blue-Hoechst Cocktail. J. Neurosci. Methods 2008, 174, 42–49. [Google Scholar] [CrossRef]
- Bozzola, J.J.; Russell, L.D. Specimen Preparation for Transmission Electron Microscopy. In Principles and Techniques for Biologists; McKean, B.L., Ed.; Jones and Bartlett Publishers Inc.: Massachusetts, 1998; pp. 20–22. ISBN 0763701920. [Google Scholar]
Figure 1.
Model of obesity induced by a high-fat diet during eight weeks. (A) Weekly body weight gain; (B) Diet consumed; (C) Fat accumulation. Data are expressed as mean ± SEM, n =18 rats. (D) and (E) adipocyte area of the epididymal and subcutaneous tissue, respectively. Data are expressed as mean ± SEM, n = 6 rats. The control group is represented with black symbols and HFD-fed group is represented with gray symbols. Asterisks indicate significant differences compared with the control group (*P < 0.05, *** P < 0.0005, **** P < 0.0001. (F) Representative microphotograph (C) of adipose tissue (4 µm) by hematoxylin-eosin-stained paraffin sections.
Figure 1.
Model of obesity induced by a high-fat diet during eight weeks. (A) Weekly body weight gain; (B) Diet consumed; (C) Fat accumulation. Data are expressed as mean ± SEM, n =18 rats. (D) and (E) adipocyte area of the epididymal and subcutaneous tissue, respectively. Data are expressed as mean ± SEM, n = 6 rats. The control group is represented with black symbols and HFD-fed group is represented with gray symbols. Asterisks indicate significant differences compared with the control group (*P < 0.05, *** P < 0.0005, **** P < 0.0001. (F) Representative microphotograph (C) of adipose tissue (4 µm) by hematoxylin-eosin-stained paraffin sections.
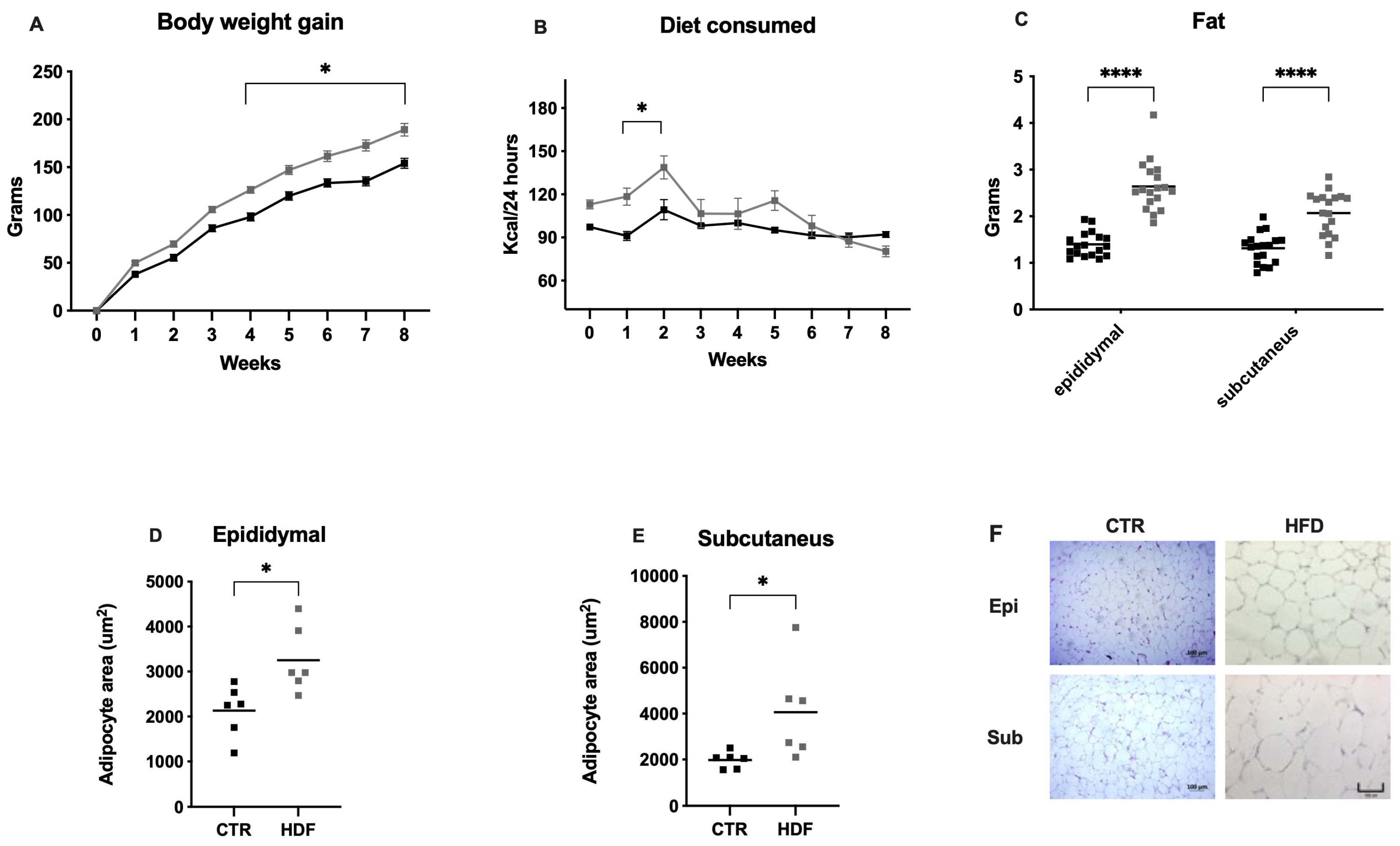
Figure 2.
Tolerance Tests Glucose and Insulin. (A) Intraperitoneal glucose tolerance test; (B) Area under the curve for glucose levels. (C) insulin tolerance test; (D) Area under the curve for insulin levels. The control group (CTR) is represented with black symbols and HFD-fed group is represented with gray symbols. Data are expressed as mean ± SEM, n = 8 rats. Asterisks indicate significant differences compared with the control group (P < 0.05). The horizontal gray line over the X-axis indicates baseline 0.
Figure 2.
Tolerance Tests Glucose and Insulin. (A) Intraperitoneal glucose tolerance test; (B) Area under the curve for glucose levels. (C) insulin tolerance test; (D) Area under the curve for insulin levels. The control group (CTR) is represented with black symbols and HFD-fed group is represented with gray symbols. Data are expressed as mean ± SEM, n = 8 rats. Asterisks indicate significant differences compared with the control group (P < 0.05). The horizontal gray line over the X-axis indicates baseline 0.
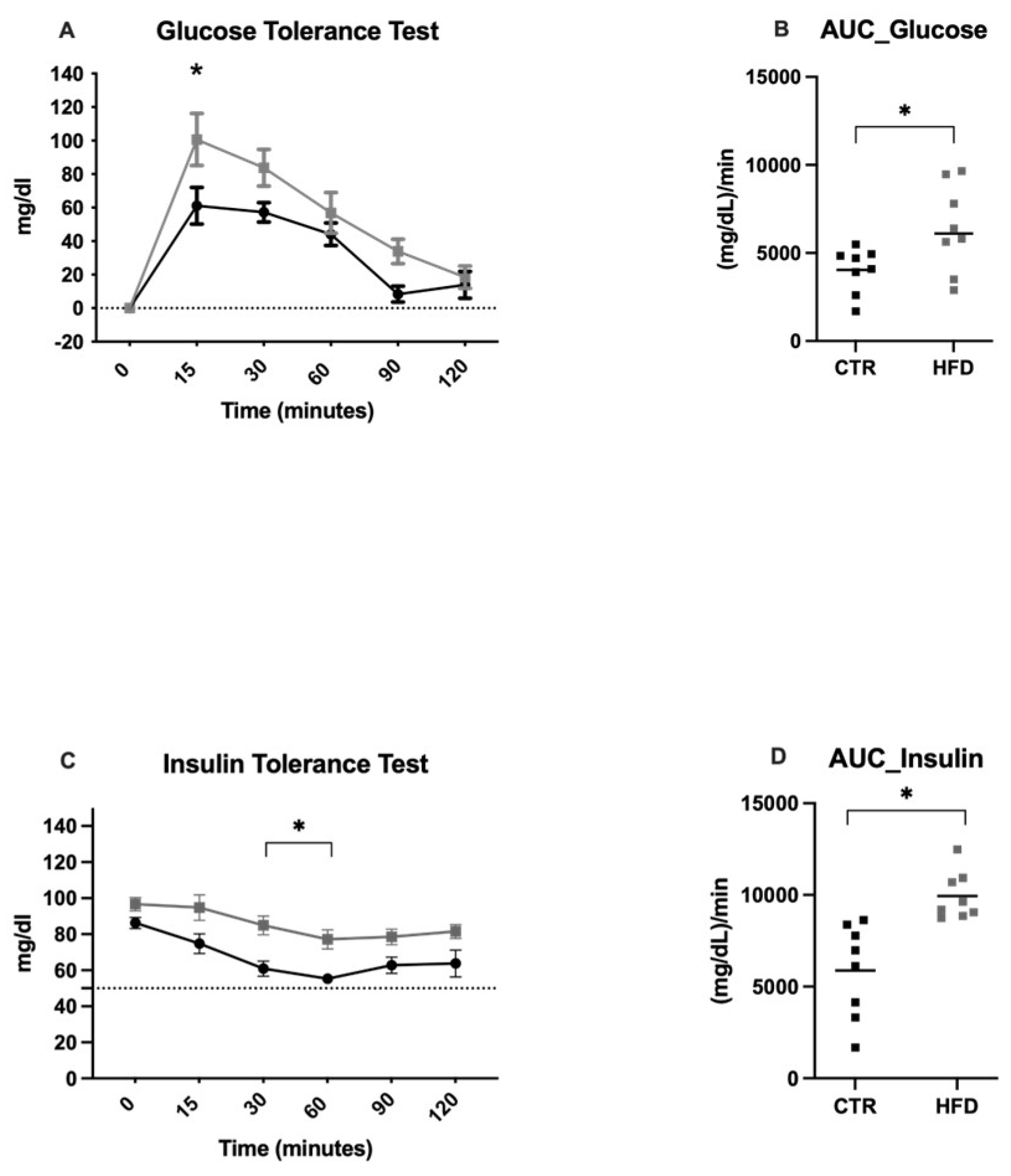
Figure 3.
Nociceptive behavior and Motor coordination induced by a high-fat diet after eight weeks. Mechanical allodynia was determined with von Frey filaments using the up-down method in the right paw (A) and (B) the left paw. (C) Latency of fall during Rota-Rod testing. The control group is represented with black symbols and HFD-fed group is represented with gray symbols. Data are expressed as mean ± SEM (n = 6). Basal*, Control & after 8 weeks with chow diet P < 0.05 vs. groups Control and HFD-fed groups after 8 weeks. (PWT) 50% paw withdrawal threshold.
Figure 3.
Nociceptive behavior and Motor coordination induced by a high-fat diet after eight weeks. Mechanical allodynia was determined with von Frey filaments using the up-down method in the right paw (A) and (B) the left paw. (C) Latency of fall during Rota-Rod testing. The control group is represented with black symbols and HFD-fed group is represented with gray symbols. Data are expressed as mean ± SEM (n = 6). Basal*, Control & after 8 weeks with chow diet P < 0.05 vs. groups Control and HFD-fed groups after 8 weeks. (PWT) 50% paw withdrawal threshold.
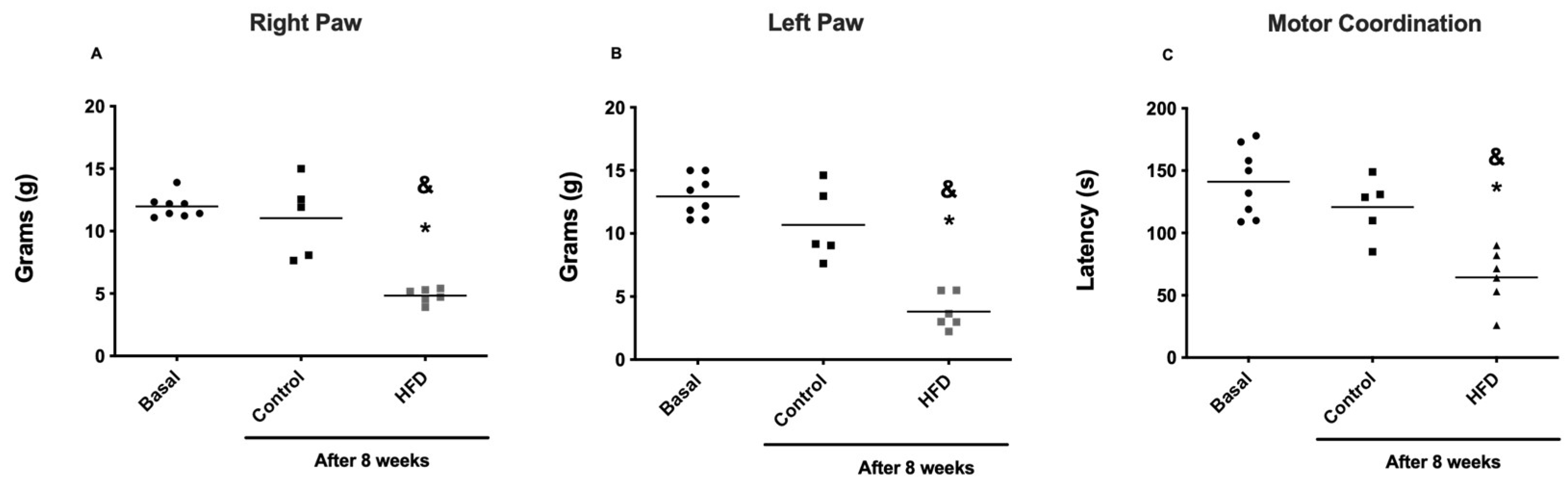
Figure 4.
Photomicrographs of different regions of the CNS from control and HFD-treated rats. The cerebellar cortex of HFD-treated rats showed an increased number of cells at the molecular layer, probably corresponding to stellate cells and basket cells, and a more prominent blood capillary network [B, H, N (arrows)] compared to animal controls (A, G, M). In the hippocampus, nuclei of neurons from the superior and inferior blades and hinge of the dentate gyrus (DG) HFD-treated rats (D, J, P) seem to have more compacted chromatin, and neuropil was observed to be smaller; note the increased cellularity of GD and normal eosinophilia of the neuropil in controls (C, I, O). In the hypothalamus anterior to the tuberal region, more dilated vessels were observed in the HFD-treated rats (F, L, R, arrows) and many cell nuclei were more hyperchromatic than in control animals (E, K, Q). H&E stain.
Figure 4.
Photomicrographs of different regions of the CNS from control and HFD-treated rats. The cerebellar cortex of HFD-treated rats showed an increased number of cells at the molecular layer, probably corresponding to stellate cells and basket cells, and a more prominent blood capillary network [B, H, N (arrows)] compared to animal controls (A, G, M). In the hippocampus, nuclei of neurons from the superior and inferior blades and hinge of the dentate gyrus (DG) HFD-treated rats (D, J, P) seem to have more compacted chromatin, and neuropil was observed to be smaller; note the increased cellularity of GD and normal eosinophilia of the neuropil in controls (C, I, O). In the hypothalamus anterior to the tuberal region, more dilated vessels were observed in the HFD-treated rats (F, L, R, arrows) and many cell nuclei were more hyperchromatic than in control animals (E, K, Q). H&E stain.
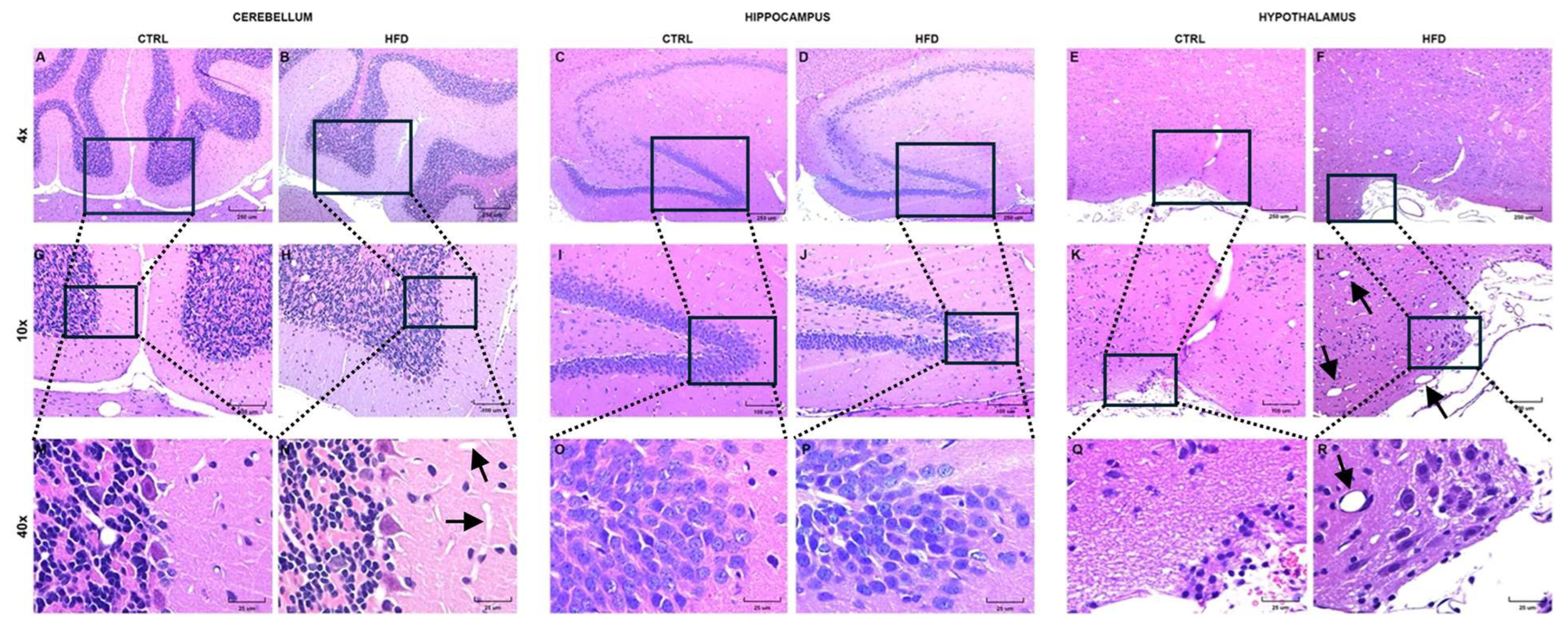
Figure 5.
GFAP (A-G) and Connexin-43 (H-N) expression at different regions on the CNS. The figure shows different CNS regions: Cerebellum GFAP (A, D), and Cx-43 (H, K), Hippocampus GFAP (B, E), and Cx-43 (I, L), Hypothalamus GFAP (C, F), and Cx-43 (J, M) in the control group (A-C, H-J), and HFD group (D-F, K-M). GFAP (G) and Cx-43 (N) intensity analysis. The control group is represented with black symbols and HFD-fed group is represented with gray symbols. Scale bar, 20 μm. Data are expressed as the mean ± SEM. Green color (GFAP or Cx-43), blue color (DAPI), red color (Evans Blue). A statistically significant difference was established as *P < 0.05.
Figure 5.
GFAP (A-G) and Connexin-43 (H-N) expression at different regions on the CNS. The figure shows different CNS regions: Cerebellum GFAP (A, D), and Cx-43 (H, K), Hippocampus GFAP (B, E), and Cx-43 (I, L), Hypothalamus GFAP (C, F), and Cx-43 (J, M) in the control group (A-C, H-J), and HFD group (D-F, K-M). GFAP (G) and Cx-43 (N) intensity analysis. The control group is represented with black symbols and HFD-fed group is represented with gray symbols. Scale bar, 20 μm. Data are expressed as the mean ± SEM. Green color (GFAP or Cx-43), blue color (DAPI), red color (Evans Blue). A statistically significant difference was established as *P < 0.05.
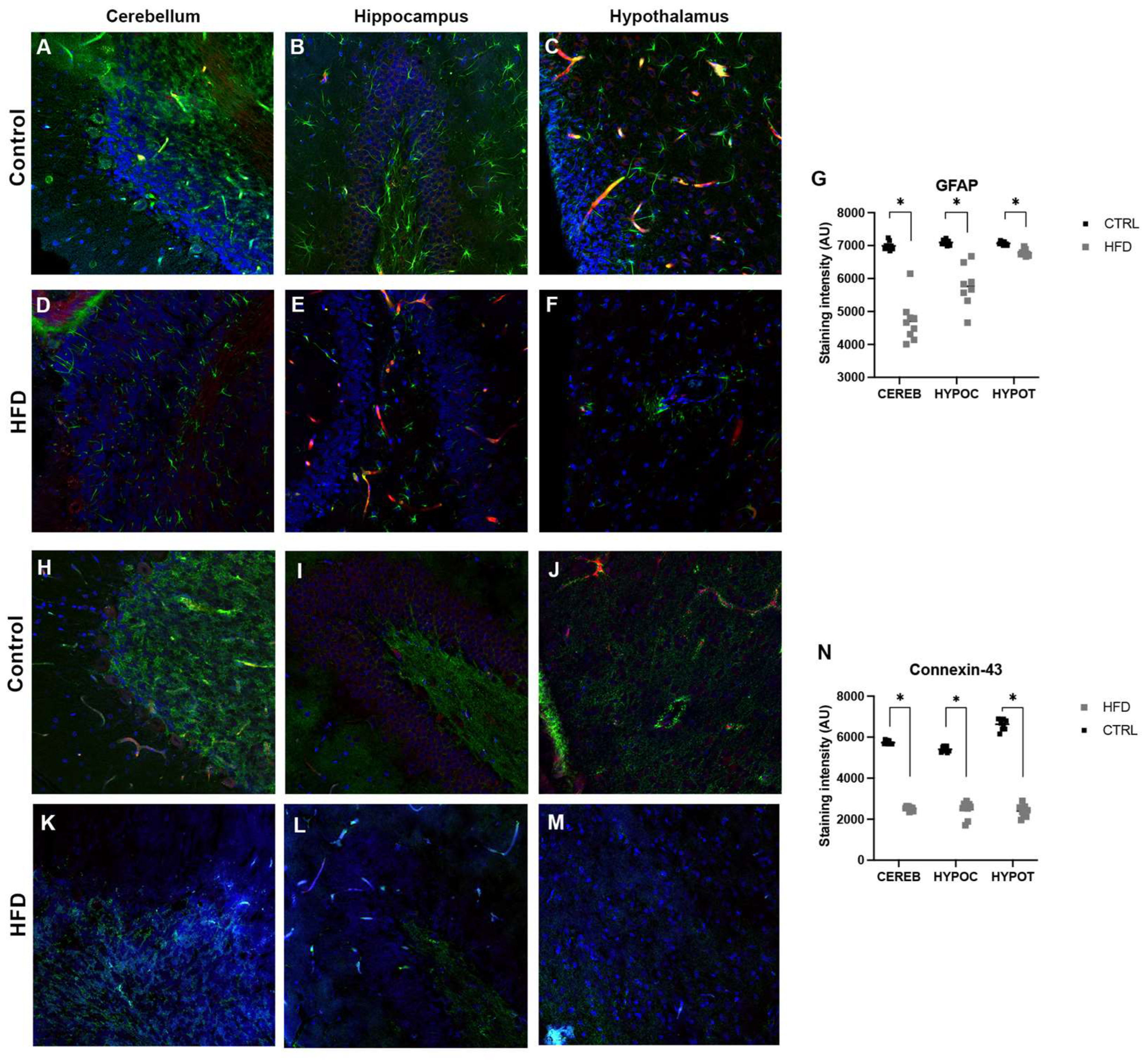
Figure 6.
Transmission electron micrograph regions of hippocampus from normal rats. (A, B) and HFD-treated rats (C-F). Hippocampal capillaries with homogeneous endothelial thickness (A, black arrowheads), neuropil (B), and myelin sheath (A, White arrowheads). Some small electro lucent spaces were occasionally observed near the blood capillaries (A, Asterisk). HFD-treated rats (C-F). Capillaries (Cap) were observed with an irregular profile and partially surrounded by clear spaces (asterisks). Spaces and apparent ruptures of axonal integrity (arrows). Nuclei of neurons (N), microglia (m) and oligodendrocytes (o). Spaces at neuropil were also observed (B, C). .
Figure 6.
Transmission electron micrograph regions of hippocampus from normal rats. (A, B) and HFD-treated rats (C-F). Hippocampal capillaries with homogeneous endothelial thickness (A, black arrowheads), neuropil (B), and myelin sheath (A, White arrowheads). Some small electro lucent spaces were occasionally observed near the blood capillaries (A, Asterisk). HFD-treated rats (C-F). Capillaries (Cap) were observed with an irregular profile and partially surrounded by clear spaces (asterisks). Spaces and apparent ruptures of axonal integrity (arrows). Nuclei of neurons (N), microglia (m) and oligodendrocytes (o). Spaces at neuropil were also observed (B, C). .
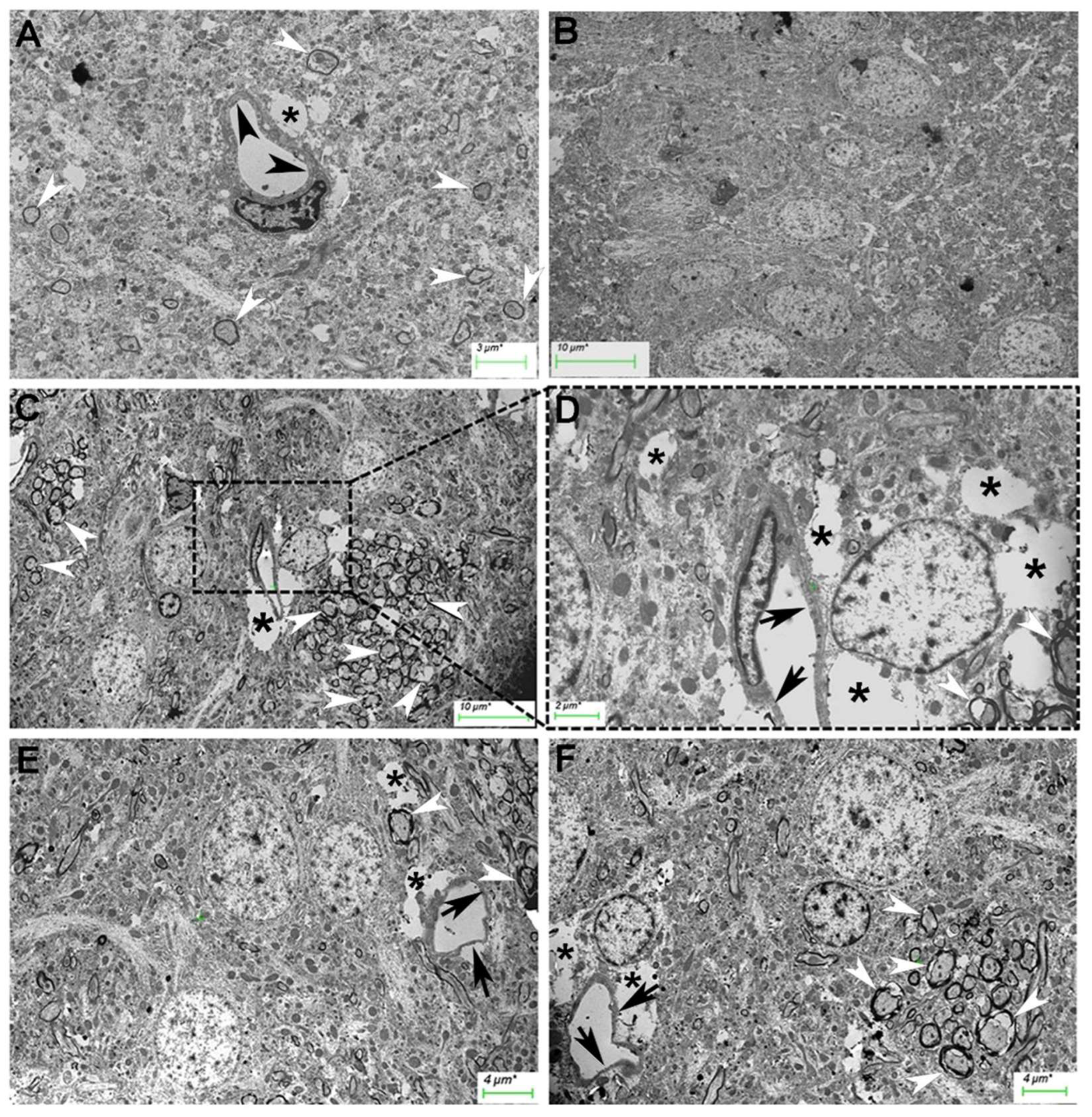
Figure 7.
Transmission electron micrograph of hypothalamus from normal rats. (A, B) and HFD-treated rats (C, D). Capillaries (A, arrowheads; nu: nucleus). Endothelial cells (C, arrowheads). Nuclear chromatin condensation pattern (C, nu). Tissue fluid (C, asterisk). Nuclei with prominent nucleoli in normal rats (B, arrows). Swelling nuclei, absent nucleoli, and karyolysis in HFD (D, arrows). Some neurons were highly dense in electrons (D, arrowheads).
Figure 7.
Transmission electron micrograph of hypothalamus from normal rats. (A, B) and HFD-treated rats (C, D). Capillaries (A, arrowheads; nu: nucleus). Endothelial cells (C, arrowheads). Nuclear chromatin condensation pattern (C, nu). Tissue fluid (C, asterisk). Nuclei with prominent nucleoli in normal rats (B, arrows). Swelling nuclei, absent nucleoli, and karyolysis in HFD (D, arrows). Some neurons were highly dense in electrons (D, arrowheads).
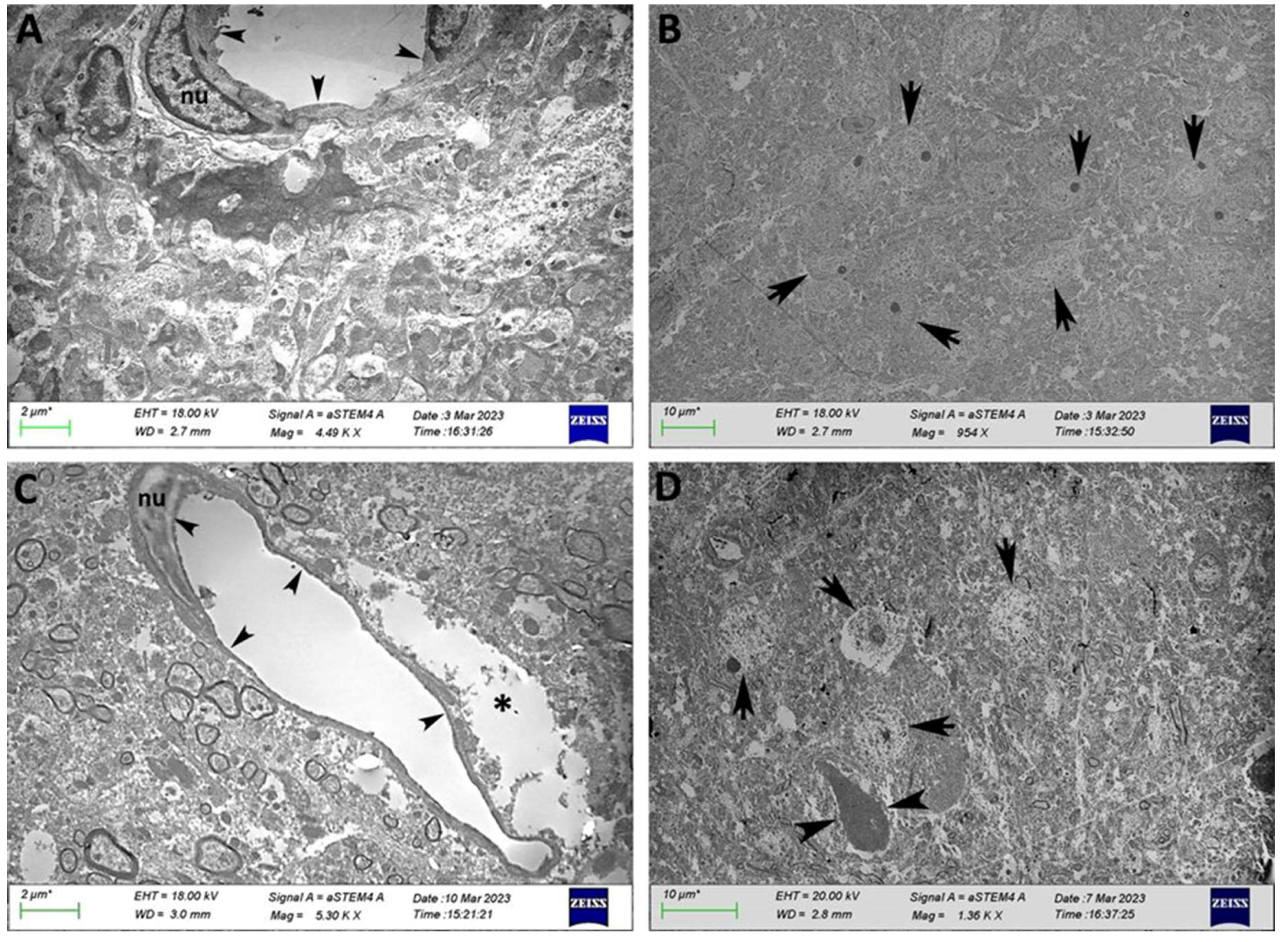

Table 1.
Effect of diets on somatometric and physiological parameters after 8 weeks.
| Parameters | Groups | P | |
|---|---|---|---|
| CTR | HFD-fed | ||
| BMI (kg/m2) | 0.67 ± 0.02 | 0.79 ± 0.03 | 0.002* |
| Abdominal circumference (cm) | 19.09 ± 0.13 | 21.23 ± 0.28 | 0.001* |
| Body length (cm) | 25.86 ± 0.35 | 24.50 ± 0.44 | 0.026* |
| Lee index (g/cm) | 295.34 ± 4.42 | 318.70 ± 5.68 | 0.004* |
| Fasting plasma glucose (mg/dL) | 81 ± 1.58 | 91 ± 4.54 | 0.022* |
| Leptin (pg/ml) | 26547.19 ± 7785 | 58447.19 ± 9227 | 0.007* |
| Insulin (pg/ml) | 3180.77 ± 366.2 | 4621.67 ± 691.1 | 0.042* |
| Triacylglycerols (mg/dL) | 99.11 ± 4.47 | 118.16 ± 18.39 | 0.177 |
| HOMA-IR (%) | 18.38 ± 1.86 | 31.76 ± 4.59 | 0.012* |
Bold numbers indicate significant differences. Data are expressed as mean ± SEM, n = 11 rats. A statistically significant difference was established as *P < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



