
Submitted:
24 September 2024
Posted:
25 September 2024
You are already at the latest version
Abstract
Bacterial structures formed from the outer membrane and the periplasm components carry biomolecules to expel cellular material and interact with other cells. These outer membrane vesicles (OMVs) can encapsulate bioactive content, which gives OMVs a high potential as alternative drug delivery vehicles or as a platform for novel vaccine development. Single-gene mutants of the hypervesiculating strain Escherichia coli JC8031 were engineered to further enhance OMV production based on metabolic network modelling and in silico gene knockout design (ΔpoxB, ΔsgbE, ΔgmhA, and ΔallD). Mutants were experimentally obtained by genome editing using CRISPR-Cas9 and tested for OMVs recovery observing an enhanced OMV production in all of them. Lipidomic analysis through LC-ESI-QTOF-MS was performed for OMVs obtained from each engineered strain and compared to the wild type E. coli JC8031 strain. The lipid profile of OMVs from the wild type E. coli JC8031 did not change significantly when compared to the mutant strains. The obtained results suggest that the vesicle production can be further improved while the obtained vesicles are not altered in their composition, allowing further study for stability and integrity for use in therapeutic settings.
Keywords:
Outer Membrane Vesicles
; Lipidomics
; CRISPR
; Metabolic Network Models
; E. coli JC8031
1. Introduction
By shipping away bioactive compounds, Gram-negative bacteria are able to communicate, relieve stress, and attack foreign entities [1,2]. Proteins, nucleic acids, and compounds found in the periplasm are packaged into spheres made of their own outer membrane (OM) material. Their cargo, protected during travel by the lipid bilayer, can reach host organisms more reliably than compounds secreted directly to the environment. Proteins displayed on the OM can become a part of outer membrane vesicles (OMVs) and provoke adherence with other organisms [3]. Manipulation of membrane proteins and OMV cargo can be leveraged to target selected organisms and inject a desired bioactive material [4,5].
While the effectiveness of widely used antibiotics is declining, naturally occurring phenomena continue to inspire antibiotic therapies, alternative antibiotic sources, and drug delivery [1]. Pseudomonas aeruginosa produces OMVs that have bacterial toxicity. They attack other bacteria using a periplasmic autolysin encapsulated in OMVs [6]. Lisobacter spp. have also shown similar activity with bioactive compounds present in its OMVs cargo [7]. Other gram-negative bacteria, such as Escherichia coli have a high OMVs production potential [8]. This presents an opportunity to use E. coli as a platform for the production of OMVs for drug delivery, the development of novel vaccines [9], or to emulate an OMVs-based antibacterial system.
Several factors affect the production of OMVs. Mechanisms affecting their biogenesis are related to deformations in the OM including: 1.) disrupted links between the inner membrane, the peptidoglycan layer, and the outer membrane; 2.) accumulation of periplasmic proteins; and 3.) curvatures induced by OM proteins [2,3,9,10]. Kulp et al. suggest that vesicle production is affected by structural characteristics of the periplasmic membrane while metabolic pathways for the production of membrane proteins, lipopolysaccharides (LPS), enterobacterial common antigen (ECA), and stress response pathways also affect their formation [11]. However, the metabolic pathways involved in the production of OMVs have yet to be elucidated. Metabolic network models connect enzymatic reactions predicted from the genome to describe the metabolic states and biological capabilities of an organism. Finding what compounds are needed by the cells to achieve a certain state, growth rate, or compound production can help accomplish a greater in vivo productivity. Further exploring the effect of gene editing in the metabolic space can increase our understanding of OMV formation and improve its yield.
Being able to produce OMVs in large quantities is an important target. Additionally, the activity and stability of OMVs are important when considering their therapeutic use [12]. Proteins in the cargo and displayed on the membrane affect their bioactivity [3]. Toxic LPS in the outer membrane can also affect their medical use [13].The OMVs proteome analyses of various Gram-negative bacteria, including E. coli, have been reported and lipidomic profiles for Klebsiella pneumoniae OMVs [14,15,16,17] have given an insight into their composition. However, there are currently no lipidomic profiles reported for E. coli OMVs.
In this study, we adapted a genome-scale metabolic network model for E. coli K12 strain and employed constraint-based optimization methods to develop strategies that enhance OMVs production in E. coli JC8031. Description of the vesicle production was achieved through the model constraints and the objective functions driving the optimization problem. We found four E. coli JC8031 mutant strains with enhanced vesicle production through in silico gene knockouts. To our knowledge, this is the first use of metabolic network models to study OMVs production and the first report on the lipid composition of isolated E. coli OMVs.
2. Results
2.1. A refined Metabolic Network Model Was Reconstructed for E. coli JC8031
We decided to use the E. coli K12 metabolic network model iML1515 [18] as base and adapt it to obtain a computational model that adequately describes vesicle production in E. coli JC8031 strain. We initially accounted for the JC8031 strain-specific deletion ΔtolRA [19]. We implemented GrowMatch [19] to verify the consistency of in silico results of cell growth and vesicle production with reported experimental data of 150 samples from the Keio collection [11]. Implementation of GrowMatch resolved modeled growth inconsistencies in 16 instances and helped calibrate the in silico predictions to cell growth data by including additional reactions in the carbon and lipid metabolism or additional constraints. Further additions were made to model protein secretion of proteins to the periplasm through transport or exchange reactions. Model data is available in Additional file 1. The employed model consisted of 2160 reactions, 724 transport and exchange reactions, and 2057 metabolites divided into two separate compartments (cytosol and periplasm) along with the extracellular space.
The objective function employed in the constraint-based methods resulted from the metabolites involved in the formation of the OM in the bacterium, which is further incorporated in the vesicle [1,3,20]. This included phosphatidylethanolamines (PEs) and phosphatidylglycerols (PGs) as well as the core oligosaccharide lipid A (COLIPA) as a precursor to LPS. The reactions that compose the objective function were key steps in the production and translocation of membrane material to the periplasm.
2.2. E. coli JC8031 Mutant Strains Were Designed in Silico and Constructed In Vitro Using CRISPR-Cas9
We performed a strain design analysis with the objective of identifying mutations that result in increased vesicle production as described by the objective function. We used OptKnock [21] to find gene knockouts that could be conducive to a greater vesicle production. The in silico strain design was performed by implementing OptKnock using the refined model of E. coli JC8031as input, the developed objective function for vesicle production, and the exchange reactions that accounted for the LB culture media. OptKnock implementation resulted in sets of up to four gene knock-outs that increase the value of the vesicle production objective function while maintaining cell growth levels. A set of four gene knock-outs was obtained after filtering results (Table 1). A combination of the four gene deletions was found to improve vesicle production in silico. Additionally, single-gene knock-outs were also found to produce an enhancing effect in silico.
Given the enhanced vesicle production found in silico, we constructed four E. coli JC8031 mutants with each single-gene deletion to experimentally assess the increased vesicle production. Four single deletion mutants (ΔpoxB, ΔsgbE, ΔgmhA, and ΔallD) were obtained by genome editing using CRISPR-Cas9. PCR results confirmed the complete deletion of the selected gene in each of the four mutants.
2.2.1. Constructed Mutants Presented an Enhanced Vesicle Production
We recovered OMVs produced by the four constructed mutants and JC8031 strain in cell cultures to determine the impact of the gene deletions on the total vesicle yield. Vesicle recovery was quantified through UV-absorbance in a wavelength spectrum ranging from 200 to 320nm [22]. The area under the absorbance spectrum curve (A.U.C) was used as a measure of vesicle quantity and concentration in each sample. Vesicle production varied within sample groups and showed a distinction in the vesicle production between the wild type E. coli JC8031 and the four designed mutant strains (Figure 1-A). All four single deletion mutants surpassed the E. coli JC8031 vesicle production. Three single deletion mutants displayed a seven-fold increase in recovered vesicle quantity, while deletion of gene ΔsgbE produced 10 times as much as JC8031 strain. Protein concentration in the samples was evaluated using the Warburg-Christian equation [23]. Protein concentration was significantly higher in three of the single deletion mutants, ΔpoxB, ΔsgbE and ΔallD (Figure 1-B).
2.3. Lipidomic Analysis Shows Knock-Out Strains Keep a Stable Composition
Given that significant changes were found in vesicle production when knocking out selected genes, a lipidomic analysis was performed to assess the possible impacts of these knock-outs on the molecular composition of the OMVs (See Additional file 2). Figure 2 shows a summary of all the detected changes in lipid composition as measured by LC-ESI-QTOF-MS. Importantly, depending on the specific analysis and metabolite annotation strategy, the lipids that can be tracked will change [24], so in this case special attention should be paid to changes in the PEs content, taking into account that these are one of the prevalent lipid compounds expected to be present in OMVs obtained from E. coli [25].
OMV lipidomics was characterized by LC-ESI-QTOF-MS for WT strain JC8031 and four single deletion mutants. Average lipid composition across all OMV samples consisted mainly of glycerolipids (~52%), glycerophospholipids (~12%), polyketides (~6%), fatty acyls (~5%), and prenol lipids (~4%). Other lipid components in minor quantities consisted of and sphingolipids (<1%), saccharolipids (<1%), and sterol lipids (<1%). Additionally, other OMV compounds remained unidentified (~20%).
The lipid profile of OMVs from single deletion mutants was compared to OMVs from the WT E. coli JC8031. Figure 2 shows the qualitative change in terms of lipid groups content. OMVs from strain ΔgmhA showed a decrease in glycerolipids as compared to WT JC8031. OMVs from strain ΔpoxB exhibited an increase in fatty acyl, polyketides, and glycerophospholipid participation in the lipid profile. The lipid profile of OMVs from the other single deletion mutant strains ΔsgbE and ΔallD, did not show evident differences with OMVs from the JC8031 strain.
It is worth mentioning that the computational algorithms used for the strain design focused on maximizing a linear combination of the fluxes of the metabolic reactions involving the production of PGs, PEs, and lipid A. A preliminary analysis of the qualitative changes showed that PEs had increased production in three of the single deletion mutants. PGs did not have a fold change greater than 3 in any of the single deletion mutant strains.
On the other hand, to obtain quantitative data, an unsupervised principal component analysis (PCA) was implemented to analyze the stability of the lipidomic analytical system and the unsupervised distribution of analyzed samples. The quality control samples appear grouped, evidencing the stability of the analytical system during the analysis of all samples (Figure 3a). Then, a discriminant PLS-DA analysis was built to maximize and inspect the differences between the study groups, obtaining no model (negative values of Q2 and CV-ANOVA with p-value=1) between the OMVs sample groups (Figure 3b). No lipids were found to significantly differentiate groups in either multivariate or univariate analysis, so it can be assumed that there are no significant changes in OMVs lipids composition obtained from the knock-out strains when compared to OMVs from E. coli JC8031.
3. Discussion
Deletion analysis performed on the refined model showed that knock-outs enhance vesicles production as modeled by the objective function. Interestingly, the gene deletions suggested by the OptKnock algorithm are not directly related to phospholipid metabolism. However, two genes have a direct relation to the outer membrane (poxB, gmhA). poxB encodes for a membrane protein that acts as a pyruvate dehydrogenase. This protein is related to membrane binding and presents interactions with lipids [26]. gmhA encodes for a phosphoheptose isomerase involved in the lipopolysaccharide biosynthesis and ΔgmhA have shown lipopolysaccharides lacking heptose [27]. The involvement of these genes with the formation of OM components supports previously proposed mechanisms of vesicle biogenesis through perturbation of membrane proteins, induced curvatures, and membrane integrity [3]. However, the results show a lack of significance in a ΔgmhA alteration over the protein concentration. The other two deleted genes (allD, sgbE) showed an indirect relation to vesicle production. sgbE encodes for an enzyme from an L-ascorbate utilization operon and is involved in the metabolism of nucleotide and aromatic amino acid precursors [28]. allD gene encodes for an enzyme involved in the assimilation of allantoin and allantoin use as a nitrogen source [29]. These single deletion targets are also indirectly related to vesicle production through the alteration of biomass growth. The use of the Warburg-Christian equation through spectrophotometric methods shows limitation to quantify vesicle production. This method can be complemented with LPS measurements through the Purpald test [23].
Results from in silico deletion experiments showed that there is a high biomass growth rate, multiple deletions did not reduce growth rate to an extent where the designed strain is no longer valuable. In silico vs. in vitro differences could be observed experimentally through deletion suggested genes. This implies a challenge in generating multiple subsequent deletions and achieving the expected biomass growth. Here, we found that deletions associated to a reduction on cellular growth also showed a reduced yield for vesicle recovery.
On the other hand, from the industrial point of view biomass growth was not significantly affected for all mutants while OMVs generation was enhanced up to 10 times in the single deletion mutant strain ΔsgbE, therefore more attractive when scaling up the process from an economical perspective.
The composition of the produced OMVs is also important when considering their use in therapeutic applications. Previous studies have raised concern on the effect of the recovery method on the size distribution, morphology and composition of OMVs. However, a comparison of isolation methods has shown that different methods produce a similar effect on vesicle morphology [5]. The lipidomic analysis showed no statistically significant change in composition that separated the single deletion mutants from the JC8031 strain. This gives an insight into the response of general OMVs composition with respect to genome alteration. The lack of change in the lipid profile could be attractive for biomedical applications as it facilitates the study and analysis of material stability of OMVs as envelopes for the desired cargo and the effect of external factors on their structural integrity. Compared to detergent
To the best of our knowledge, the studies reporting OMVs composition are relatively limited and they are mostly focused on proteomics analysis rather than lipidomics. Besides the information that can be obtained from searching in academic databases, it is also possible to consult specialized compendiums available online, such as vesiclepedia [30]. A search in this compendium can also show how information of bacterial OMVs lipidomics is very limited and suggests that the data obtained during the present study may be one of few attempts to characterize E. coli OMVs from the lipidomics point of view (See Additional file 3).
On the other hand, regardless of the relative lack of information, it is possible to make further analysis of the data if assuming that OMVs lipid composition is similar to the E. coli outer membrane composition, as suggested by previous studies [31]. In that case, special attention should be paid to compounds like PGs and PEs, as these are reported to be the prevalent lipid components of the E. coli outer membrane [20,25], particularly for the K12 strain, which is the strain from which the JC8031 strain was derived [19]. Indeed, PGs and PEs were detected in our OMVs samples although no significant changes in lipid composition can be inferred from the results when comparing OMVs from the designed single deletion mutants against OMVs from the JC8031 strain. This result is consistent with a previous study where all possible single gene deletions, in E. coli, were analyzed with respect to their effect on the membranes lipid composition [32]; none of the genes that were deleted during the present study (poxB, sgbE, gmhA, and allD) were reported previously as responsible of causing a significant change in the lipid composition of the bacterium and it might be assumed that they should not cause significant changes in OMVs lipid composition either.
Another important aspect to take into account is about the prevalent fatty acyl chains forming part of the OMVs. In our case, the prevalent species are vaccenic acid (18:1) and palmitic acid (16:0) (Additional file 3), both of them previously reported as prevalent in E. coli membranes [25]. In this last report, it was also stressed that palmitic acid was slightly more prevalent in the outer membrane. Our results show that palmitic acid is indeed slightly more prevalent when analyzed in the PEs context, but a more quantitative analysis is necessary to obtain conclusive data. This kind of analysis will be particularly important if considering that OMVs are expected to be relatively rigid, thanks to a higher content of saturated fatty acyl chains [31].
The lipid profile of E. coli OMVs presented an unexpected major component in glycerolipids, while expected major components glycerophospholipids were found in second place. While glycerophospholipids are usually present in the outer membrane in large proportions, it is not uncommon for membranes composed of glycerolipids [33]. The lack of phosphorus in membrane lipids is associated with the depletion of phosphorus in the growth media [34]. In this case, LB culture media presented a rich nutritious environment for cell growth which provided phosphorous. This implies small participation of glycerophospholipids in the E. coli OMV lipid profile is not due to phosphorus availability. It was not possible to determine if OMV lipid profile matches the outer membrane profile in whole E. coli cells, or if vesicle formation was preferential in regions low in phosphorus or with higher amounts of glycerolipids. Additionally, proteomic studies have found an enhanced carryover of inner membrane to the generated vesicles [5]. This could have implications for the engineering of membrane proteins for interaction with host cells and could partly account for the unexpected high glycerolipids content in the isolated OMVs. Further work is needed to analyze differences in OMVs from the originating outer membrane and the effect of phosphorus availability on the lipid profile.
Genomic advances have helped to determine the set of genes in an organism. Annotation of the genome of E. coli has produced a map of enzymes expressed by these bacteria. A network of these reactions comprises the metabolic states the cells are capable of undertaking. A key limitation in metabolic modeling arises from genome annotation. Missing pieces in annotation results in disconnections between reactions which alter the distribution of metabolic fluxes and could result in blocked reactions, or no in silico growth. In the case of E. coli, several network models that tackle these limitations have been developed [18]. The E. coli model has increased in the number of metabolites and reactions and expanded from the core metabolism to secondary metabolism and other functions. Disconnection of reactions still exists. Algorithms such as GrowMatch allow closing the gap between the computational model and the observed biological results. Results from GrowMatch suggest that this gap was somewhat but not fully closed in the proposed model of a high vesicle yield E. coli strain. Even though this step is not completely successful in restoring growth-no growth consistency in the model, it brings it closer to the biological growth data available.
Vesicle generation is mediated by structural mechanisms. Some metabolic pathways have been related to the production of OMVs [11]. However, these pathways are related to changes in the structure of phospholipids that cause instability of the cell envelope and higher vesicle production. In this study, we focused on exploring the metabolic pathways involved in the generation of OMV precursors: cell envelope precursors, lipidA and glycerophospholipids. This results in modeling a system of multiple metabolites. Connectivity in metabolic networks made the optimization of the selected objective function harder. In comparison, enhancement of the production of a single metabolite is easier to study as the effects of changes in the metabolic network can be traced back easier. To improve the predictive power of the metabolic model, additional information is required as input to further constrain the model and contextualize it to experimental data. Future work should include the improvement of the model through the integration of lipidomic and transcriptomic data. Integration of lipidomic data is challenging as lipids are usually expressed in generic metabolites in metabolic models and experimental data is not easily associated with the network.
4. Materials and Methods
4.1. Vesicle Recovery and Quantification
High vesicle yield E. coli JC8031 strain was selected as the base strain (also referred to as WT strain). E. coli JC8031 is a tol/pal mutant [19]. This strain was modeled in silico and used for experimental procedures, vesicle recovery, and gene editing. E. coli JC8031 was transformed with a GFP production plasmid with an ampicillin resistance cassette for selection.
E. coli was cultured in 50mL supplemented LB medium, with ampicillin for GFP plasmid selection (100 ug/mL ampicillin final concentration), and incubated for 24 hours at 37ºC in a shaker incubator at 250 rpm to obtain OMVs at a late log phase and avoid cell debris collection [11]. The culture was transferred to 50mL falcon tubes and centrifuged at 4500 rpm and 20ºC for 30 minutes to remove cells. After this, 30 mL of supernatant was filtered through a 45um polyamide membrane in a filter housing using a 50 mL syringe to remove any debris present in the sample. The filtrate was ultracentrifuged at 50,000G for 3 hours at 4ºC [5]. Presence of vesicles was verified as a translucent pellet with green fluorescence under blue light. Vesicles were resuspended in 1000uL of PBS with 10uL of spectinomycin antibiotic and stored at -80ºC until further analysis. Vesicle solution in PBS was inoculated in LB medium agar with ampicillin to check that no viable cells were still present.
Vesicle production was quantified indirectly through UV absorption spectra. Absorption was obtained in a UV spectrophotometer in a spectrum ranging from 200nm to 320nm at intervals of 2nm. Vesicle production values were calculated as the area under the UV absorption curve in the specified spectrum range. Additionally, the protein concentration in each sample was semi-quantitatively estimated using the Warburg-Christian equation using absorbance values at 280nm for protein content and 260nm to account for the influence of nucleic acid content [23]. Vesicle production and protein concentration was normalized to the mass of sample and OD600 to account for varying growth rates between sample groups.
4.2. Metabolic Network Model
A metabolic network model for E. coli K12 was used as initial input to assess the OMVs production and to find optimal culture conditions in silico. This model was initially developed by Monk et al. [18] as iML1515 and modified to ensure the presence of biomass precursors and a functional biomass growth reaction. The model was adapted to strain JC8031 to account for different mutations.
4.3. Model Refinement
Vesicle production values and growth rates of 3908 E. coli mutants in the Keio collection were reported by Kulp et al. [11]. Flux balance analysis (FBA) [35] was used to simulate biomass growth of 150 Keio mutants reported by Kulp to exhibit high vesicle production phenotype. The results of the FBA were used to compare simulated and experimental cell growth rates and check the consistency of in silico results and reported in vitro data. Subsets of this dataset were used to perform improvements through constraint based optimization methods [36]. Matching results from in silico predictions and reported experimental values are present as either growth-growth (GG), or no growth-no growth (NGNG). When inconsistencies appear, in silico predictions display growth while experimental results do not, or vice versa. These are cataloged as growth-no growth (GNG) or no growth-growth (NGG) inconsistencies. GrowMatch [37] was used to calibrate in silico predictions to cell growth data of the Keio collection through the exclusion of model reactions, while NGG inconsistencies were resolved by the inclusion of reactions from an external database.
4.4. Model Objective Function Definition
The objective function for vesicle production is key to properly model biological behavior. As the objective function is maximized or minimized, the distribution of metabolic fluxes changes. This has the effect of guiding the model towards a biologically relevant solution. The objective functions found by using available algorithms [38] were linear combinations of reactions found in the model but do not appear to be related to known mechanisms of vesicle production, important metabolic steps in vesicle production, or tied to the composition of vesicles and were over-fitted to the data. The objective function was described according to the expected composition of the E. coli OM, glycerophospholipids and lipopolysaccharides [34] as well as the expectation for an increased phospholipid production to increase OMV formation [39]. This approach is similar to the constitution of biomass objective functions which are also key to the model [40].
4.5. Strain Design
Targeted deletions to improve vesicle production were studied using the calibrated model with the defined objective function for vesiculation. The constraint-based method OptKnock [21] was used to simulate gene deletions by inactivating reaction fluxes while maximizing vesiculation and biomass production. This approach ensures lethal deletions were not evaluated. OptKnock was implemented to simulate gene deletions on the refined model taking into account that LB medium metabolites are available as active exchange reactions. OptKnock algorithm was used limited to 5 deletions.
4.6. Genome Editing
Target genes were knocked-out by complete gene deletion by using CRISPR-Cas9. Guide RNAs were designed for the genes obtained through strain design using the tools from Benchling [41] and a reference E. coli genome (NCBI Accession: NC_000913.3). Synthetic cassettes were designed including the N20 from gRNA design (Table 2) and two segments of 500bp corresponding to sequence flanking the target gene to provide template availability for homology directed repair of the double stranded DNA lesion thereby completely eliminating the targeted gene from the bacterial genome. A system of two plasmids was employed: a plasmid including Cas9 genes and a plasmid for gRNA expression [42].
4.7. Lipidomics
Lipidomic analysis was carried out at MetCore Uniandes (Bogota, Colombia). 40 uL of type I water and 160 uL of HPLC-grade methanol were added to the lyophilized samples. Samples were resuspended in a vortex for two minutes. 400 uL of MTBE were added and samples were vortexed for 60 minutes. 250 uL of type I water were added and samples were vortexed for two minutes. Samples were centrifuged at 6190 rpm and 25°C for 10 minutes. 20 uL of organic phase were transferred to HPLC fixed-insert vials and diluted with 80 uL of MTBE.
Quality control samples were prepared by mixing equal volumes of metabolite extract from each sample. Quality control runs were performed to stabilize the analytic platform. Subsequent quality control runs were employed every five randomized samples.
Lipidomic analysis was implemented in an Agilent Technologies 1260 Liquid Chromatography system coupled to a quadrupole time of flight mass analyzer and ionization by electrospray (LC-ESI-QTOF-MS). 1 uL of each sample was injected in a C8 column (InfinityLab Poroshell 120 EC-C8 (150 x 3.0 mm, 2.7 µm)) at 60°C. A gradient elution was employed composed of 5 mM ammonium formiate in Milli-Q water (Phase A) and 5 mM of ammonium formiate in isopropanol-methanol 15:85 (Phase B) with a constant flux of 0,4 mL/min. Mass spectrometry detection was performed in ESI positive mode in full scan from 100 to 1100 m/z. Mass correction was employed throughout the analysis with two reference masses: m/z 121,0509 (C5H4N4) and m/z 922,0098 (C18H18O6N3P3F24).
Lipidomic profiles were obtained by using Agilent Mass Hunter Profinder 10 software employing the Recursive Feature Extraction (RFE) with extraction conditions: 0-31 min and 10000 counts, positive ion species: (-H, +Cl, +NH4), and no additional mass or species filters. Molecular characteristics found in the solvent blank control were eliminated. Data from alignment, deconvolution and integration were filtered by calculation of a variation coefficient (VC) of the area in QC samples. Characteristics with CV > 20% were filtered out. Data was normalized based on vesicle sample information: quantity and sample volume.
Selection of statistically significant molecular characteristics was performed through multivariate statistical analysis (MVA) and univariate (UVA). MVA was performed using SIMCA-P + 16.0 software (Umetrics) and UVA was performed by implementing the MetaboAnalyst 4.0 tool (#). Annotation of molecular characteristics was performed using the CEU MASS MEDIATOR tools (#) by batch analysis with the following parameters: tolerance=10 ppm; databases=LipidMaps; Metabolites=OnlyLipids; input masses mode= m/z masses; ionization mode=positive; adducts= (M+H, M+Cl, M+NH4). Additionally, molecular formulas were generated for statistically significant molecular characteristics by using the Agilent MassHunter Qualitative 10 software with positive ions= (-H, +Cl, +NH4) and elements= (H, C, O, N, P) as parameters.
4.8. Statistical Analysis
Data was analysed using GraphPad Prism version 9.3.1 (GraphPad Software). An ordinary one-way ANOVA was used to estimate the statistical significance of differences within groups. Sidak’s multiple comparison test was used to evaluate statistical difference with wild type group JC8031. Differences were considered significant at P values of <0.05.
5. Conclusions
In this study, we generated four single deletion mutants through in silico metabolic network model strain design and CRISPR-Cas9 genome editing (ΔpoxB, ΔsgbE, ΔgmhA, ΔallD). The four designed strains showed an enhanced vesicle production of up to 10 times that of the wild-type JC8031 strain while maintaining cell growth, while only three displayed a greater protein concentration (ΔpoxB, ΔsgbE, ΔallD). This is a step ahead to obtain OMVs in a large-scale setting and evidences it is plausible to use metabolic network modelling the complex vesicle formation process while targeting single genes.
Lipidomic analysis though LC-ESI-QTOF-MS showed that there were no significant changes in lipid profile that separated the four mutants and WT in different groups. OMV lipid profile remained stable across strains. Recovered E. coli OMVs displayed a lack of phosphorus in their composition as glycerolipids were the majority component while glycerophospholipids were present in a lesser quantity. Knowledge of E. coli OMV lipid profile and lack of alterations in different single deletion strains is useful to employ OMVs as therapeutic alternatives and drug transport and delivery.
Strain design using metabolic network models encounters difficulties when the target for production is a complex structure such as OMVs instead of a single metabolite. Integration of multiple source omic data should be used to improve the model.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Additional file 1.xlsx Title Model data Description: Metabolic network model information. List of metabolites and extracellular, transport, exchange, cytosolic, and periplasmic reactions. Additional file 2.xlsx Title: Lipidomic data Description: Data obtained by LC_MS of the lipid profile of outer-membrane vesicles obtained from Escherichia coli JC8031 and four single-deletion mutants. Additional file 3.xlsx Title: Lipidomic analysis. Fold change Description: Detected changes in lipid profile between outer-membrane vesicles from Escherichia coli JC8031 and four single-deletion mutants.
Author Contributions
Conceptualization: AFGB, JDV, HAR, Data curation: HAR, MPC, JDV, Formal analysis: HAR, JDV, MPC, Funding acquisition: JDV, AFGB, Investigation: HAR, MPC, MF, JDV, Methodology: HAR, JDV, MPC, MF, AFGB, Project administration: JDV, AFGB,Supervision & Validation: AFGB, JDV, Visualization: HAR, MPC, Writing; original draft: HAR, JDV, MPC, Writing; review & editing: HAR, JDV, MPC, MF, AFGB.
Funding
This project was funded by COLCIENCIAS through a grant from the National Program on Science, Technology, and Innovation in Health; contract 822-2017, Bogotá D.C. - Colombia.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest
References
- Jan, A.T. Outer Membrane Vesicles (OMVs) of Gram-negative Bacteria: A Perspective Update. Front. Microbiol. 2017, 8, 1053–1053. [Google Scholar] [CrossRef] [PubMed]
- Nagakubo, T.; Nomura, N.; Toyofuku, M. Cracking Open Bacterial Membrane Vesicles. Front. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kulp, A.; Kuehn, M.J. Biological Functions and Biogenesis of Secreted Bacterial Outer Membrane Vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Pillai, J. Bacterial membrane vesicles as novel nanosystems for drug delivery. Int. J. Nanomed. 2017, 12, 6329–6341. [Google Scholar] [CrossRef]
- Reimer, S.L.; Beniac, D.R.; Hiebert, S.L.; Booth, T.F.; Chong, P.M.; Westmacott, G.R.; Zhanel, G.G.; Bay, D.C. Comparative Analysis of Outer Membrane Vesicle Isolation Methods With an Escherichia coli tolA Mutant Reveals a Hypervesiculating Phenotype With Outer-Inner Membrane Vesicle Content. Front. Microbiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Kadurugamuwa, J.L.; Beveridge, T.J. Bacteriolytic effect of membrane vesicles from Pseudomonas aeruginosa on other bacteria including pathogens: conceptually new antibiotics. J. Bacteriol. 1996, 178, 2767–2774. [Google Scholar] [CrossRef]
- Vasilyeva, N.V.; Tsfasman, I.M.; Kudryakova, I.V.; Suzina, N.E.; Shishkova, N.A.; Kulaev, I.S.; Stepnaya, O.A. The role of membrane vesicles in secretion of lysobacter sp. bacteriolytic enzymes. Microb. Physiol. 2013, 23, 142–151. [Google Scholar] [CrossRef]
- Li, Z.; Clarke, A.J.; Beveridge, T.J. Gram-negative bacteria produce membrane vesicles which are capable of killing other bacteria. J. Bacteriol. 1998, 180, 5478–5483. [Google Scholar] [CrossRef]
- Furuyama, N.; Sircili, M.P. Outer membrane vesicles (OMVs) produced by gram-negative bacteria: Structure, functions, biogenesis, and vaccine application. BioMed Res. Int. 2021. [Google Scholar] [CrossRef]
- Avila-Calderón, E.D.; Ruiz-Palma, M.d.S.; Aguilera-Arreola, M.G.; Velázquez-Guadarrama, N.; Ruiz, E.A.; Gomez-Lunar, Z.; Witonsky, S.; Contreras-Rodríguez, A. Outer Membrane Vesicles of Gram-Negative Bacteria: An Outlook on Biogenesis. Front. Microbiol. 2021. [Google Scholar] [CrossRef]
- Kulp, A.J.; Sun, B.; Ai, T.; Manning, A.J.; Orench-Rivera, N.; Schmid, A.K.; Kuehn, M.J. Genome-wide assessment of outer membrane vesicle production in Escherichia coli. PLOS ONE 2015, 10, e0139200. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liu, Q. Engineered Bacterial Outer Membrane Vesicles as Multifunctional Delivery Platforms. Front. Mater. 2020, 7. [Google Scholar] [CrossRef]
- Jasim, R.; Han, M.-L.; Zhu, Y.; Hu, X.; Hussein, M.H.; Lin, Y.-W.; Zhou, Q.; Da Dong, C.Y.; Li, J.; Velkov, T. Lipidomic Analysis of the Outer Membrane Vesicles from Paired Polymyxin-Susceptible and -Resistant Klebsiella pneumoniae Clinical Isolates. Int. J. Mol. Sci. 2018, 19, 2356. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.; Choi, D.S.; Kim, K.P.; Gho, Y.S. Proteomics in gram-negative bacterial outer membrane vesicles. Mass Spectrom. Rev. 2008, 27, 535–555. [Google Scholar] [CrossRef]
- Lee, J.; Kim, O.Y.; Gho, Y.S. Proteomic profiling of Gram-negative bacterial outer membrane vesicles: Current perspectives. Proteom. – Clin. Appl. 2016, 10, 897–909. [Google Scholar] [CrossRef]
- Tiwari, V.; Solanki, V.; Roy, R.; Biswas, D.; Tiwari, M. Significances of OMV and Extracellular Vesicle Proteomics. Journal of Data Mining in Genomics & Proteomics 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Monk, J.M.; Lloyd, C.J.; Brunk, E.; Mih, N.; Sastry, A.; King, Z.; Takeuchi, R.; Nomura, W.; Zhang, Z.; Mori, H.; et al. iML1515, a knowledgebase that computes Escherichia coli traits. Nat. Biotechnol. 2017, 35, 904–908. [Google Scholar] [CrossRef]
- Bernadac, A.; Gavioli, M.; Lazzaroni, J.-C.; Raina, S.; Lloubès, R. Escherichia coli tol-pal mutants form outer membrane vesicles. J. Bacteriol. 1998, 180, 4872–4878. [Google Scholar] [CrossRef]
- Oursel, D.; Loutelier-Bourhis, C.; Orange, N.; Chevalier, S.; Norris, V.; Lange, C.M. Lipid composition of membranes of Escherichia coli by liquid chromatography/tandem mass spectrometry using negative electrospray ionization. Rapid Commun. Mass Spectrom. 2007, 21, 1721–1728. [Google Scholar] [CrossRef]
- Burgard, A.P.; Pharkya, P.; Maranas, C.D. OptKnock: A Bilevel Programming Framework for Identifying Gene Knockout Strategies for Microbial Strain Optimization. Biotechnol. Bioeng. 2003, 84, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Ayed, Z.; Cuvillier, L.; Dobhal, G.; Goreham, R.V. Electroporation of outer membrane vesicles derived from Pseudomonas aeruginosa with gold nanoparticles. SN Appl. Sci. 2019, 1, 1600. [Google Scholar] [CrossRef]
- Roier, S.; Zingl, F.G.; Cakar, F.; Durakovic, S.; Kohl, P.; Eichmann, T.O.; Klug, L.; Gadermaier, B.; Weinzerl, K.; Prassl, R.; et al. A novel mechanism for the biogenesis of outer membrane vesicles in Gram-negative bacteria. Nat. Commun. 2016, 7, 10515. [Google Scholar] [CrossRef]
- Markowitz, V.M.; Chen, I.-M.A.; Palaniappan, K.; Chu, K.; Szeto, E.; Grechkin, Y.; Ratner, A.; Jacob, B.; Huang, J.; Williams, P.; et al. IMG: the integrated microbial genomes database and comparative analysis system. Nucleic Acids Res. 2011, 40, D115–D122. [Google Scholar] [CrossRef]
- Lugtenberg, E.; Peters, R. Distribution of lipids in cytoplasmic and outer membranes of Escherichia coli K12. Biochim. et Biophys. Acta (BBA) - Lipids Lipid Metab. 1976, 441, 38–47. [Google Scholar] [CrossRef]
- Grabau, C.; Chang, Y.Y.; E Cronan, J. Lipid binding by Escherichia coli pyruvate oxidase is disrupted by small alterations of the carboxyl-terminal region. J. Biol. Chem. 1989, 264, 12510–12519. [Google Scholar] [CrossRef] [PubMed]
- Brooke, J.S.; Valvano, M.A. Biosynthesis of inner core lipopolysaccharide in enteric bacteria identification and characterization of a conserved phosphoheptose isomerase. Journal of Biological Chemistry 1996, 271, 3608–3614. [Google Scholar] [CrossRef]
- Yew, W.S.; Gerlt, J.A. Utilization of L-ascorbate by Escherichia coli K-12: Assignments of functions to products of the yjf-sga and yia-sgb operons. J. Bacteriol. 2002, 184, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-I.; Shin, I.; Cho, S.; Lee, J.; Rhee, S. Structural and Functional Insights into (S)-Ureidoglycolate Dehydrogenase, a Metabolic Branch Point Enzyme in Nitrogen Utilization. PLOS ONE 2012, 7, e52066. [Google Scholar] [CrossRef]
- Kalra, H.; Simpson, R.J.; Ji, H.; Aikawa, E.; Altevogt, P.; Askenase, P.; Bond, V.C.; Borràs, F.E.; Breakefield, X.; Budnik, V.; et al. Vesiclepedia: A Compendium for Extracellular Vesicles with Continuous Community Annotation. PLoS Biol. 2012, 10, e1001450. [Google Scholar] [CrossRef]
- Kulkarni, H.M.; Jagannadham, M.V. Biogenesis and multifaceted roles of outer membrane vesicles from Gram-negative bacteria. Microbiology 2014, 160, 2109–2121. [Google Scholar] [CrossRef] [PubMed]
- Jeucken, A.; Molenaar, M.R.; van de Lest, C.H.; Jansen, J.W.; Helms, J.B.; Brouwers, J.F. A Comprehensive Functional Characterization of Escherichia coli Lipid Genes. Cell Rep. 2019, 27, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Geiger, O.; Sohlenkamp, C.; López-Lara, I. M. Formation of Bacterial Membrane Lipids: Pathways, Enzymes, Reactions. In Handbook of Hydrocarbon and Lipid Microbiology; 2010. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: diversity in structures and pathways. 2015. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.D.; Thiele, I.; Palsson, B.Ø. What is flux balance analysis? 2010. [Google Scholar] [CrossRef]
- Maranas, C.D.; Zomorrodi, A.R. Optimization Methods in Metabolic Networks; 2016. [Google Scholar]
- Kumar, V.S.; Maranas, C.D. GrowMatch: An automated method for reconciling in silico/in vivo growth predictions. PLOS Comput. Biol. 2009, 5, e1000308. [Google Scholar] [CrossRef]
- Burgard, A.P.; Maranas, C.D. Optimization-based framework for inferring and testing hypothesized metabolic objective functions. Biotechnol. Bioeng. 2003, 82, 670–677. [Google Scholar] [CrossRef]
- Balhuizen, M.D.; Veldhuizen, E.J.A.; Haagsman, H.P. Outer Membrane Vesicle Induction and Isolation for Vaccine Development. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Feist, A.M.; O Palsson, B. The biomass objective function. Curr. Opin. Microbiol. 2010, 13, 344–349. [Google Scholar] [CrossRef]
- Inc. Benchling, “Benchling [Biology Software],” Retrieved from https://benchling.com.
- Jiang, Y.; Chen, B.; Duan, C.; Sun, B.; Yang, J.; Yang, S. Multigene editing in the Escherichia coli genome via the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2015, 81, 2506–2514. [Google Scholar] [CrossRef]
Figure 1.
A) Spectrophotometric determination of mean vesicle production for E. coli JC8031 and the four constructed mutants, ΔpoxB, ΔsgbE, ΔgmhA, ΔallD, normalized to mass of sample and OD600. B) Mean protein concentration according to the Warburg-Christian equation. (One Way Anova presented p<0.0001; Asterisks show Sidak’s multiple comparison test results, ns: p>0.05; *: p<0.05; **: p<0.01; ****: p<0.0001, all groups have n=3).
Figure 1.
A) Spectrophotometric determination of mean vesicle production for E. coli JC8031 and the four constructed mutants, ΔpoxB, ΔsgbE, ΔgmhA, ΔallD, normalized to mass of sample and OD600. B) Mean protein concentration according to the Warburg-Christian equation. (One Way Anova presented p<0.0001; Asterisks show Sidak’s multiple comparison test results, ns: p>0.05; *: p<0.05; **: p<0.01; ****: p<0.0001, all groups have n=3).
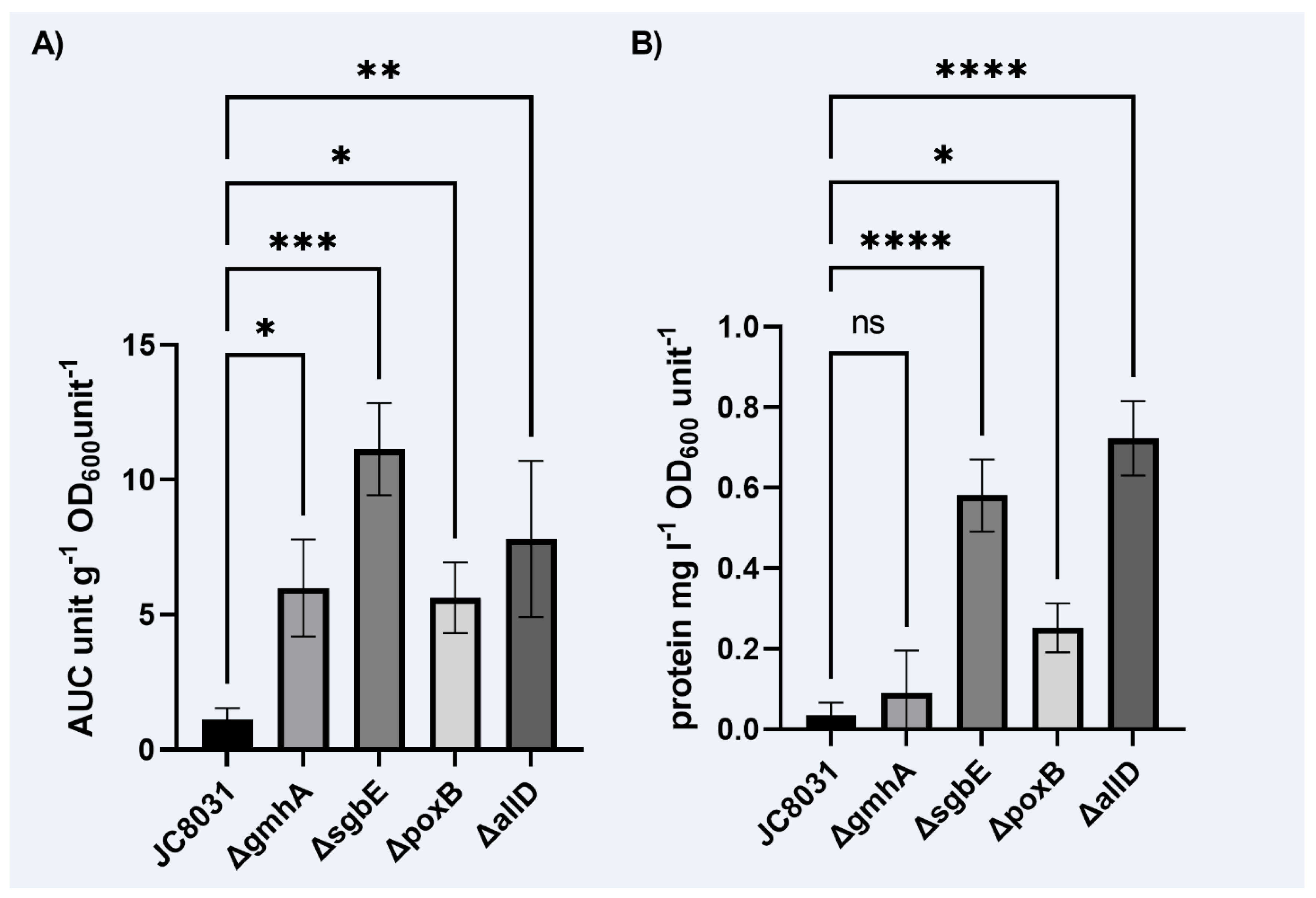
Figure 2.
Lipid groups percentage change of four constructed E. coli mutants, ΔpoxB, ΔsgbE, ΔgmhA, ΔallD compared to WT E. coli JC8031 strain. Darker shades of blue indicatee a higher lipid group content for the WT strain (negative change in relative abundance), darker shades of red indicate higher lipid group content in the respective mutant (positive change in relative abundance).
Figure 2.
Lipid groups percentage change of four constructed E. coli mutants, ΔpoxB, ΔsgbE, ΔgmhA, ΔallD compared to WT E. coli JC8031 strain. Darker shades of blue indicatee a higher lipid group content for the WT strain (negative change in relative abundance), darker shades of red indicate higher lipid group content in the respective mutant (positive change in relative abundance).
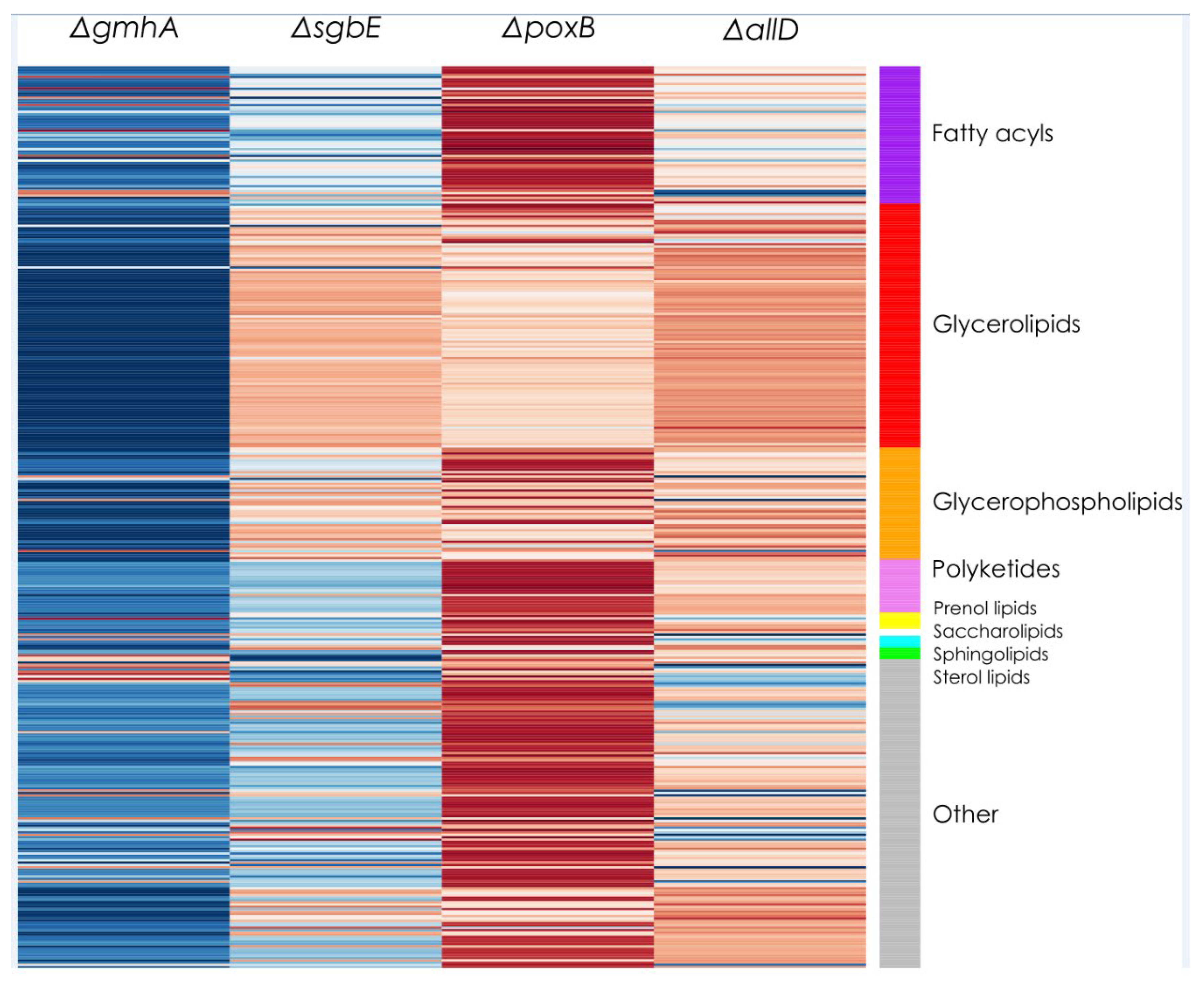
Figure 3.
PCA and PLS-DA models for lipidomic analysis by LC-ESI-QTOF-MS of OMVs from E.coli JC8031 and ΔpoxB, ΔsgbE, ΔgmhA, ΔallD mutants. a) PCA score plot and Quality control samples (QC). R2(cum): 0.954, Q2 (cum): 0.906; b) PLS-DA model, R2(cum): 0.885, Q2 (cum): -0.159, CV-ANOVA=1.
Figure 3.
PCA and PLS-DA models for lipidomic analysis by LC-ESI-QTOF-MS of OMVs from E.coli JC8031 and ΔpoxB, ΔsgbE, ΔgmhA, ΔallD mutants. a) PCA score plot and Quality control samples (QC). R2(cum): 0.954, Q2 (cum): 0.906; b) PLS-DA model, R2(cum): 0.885, Q2 (cum): -0.159, CV-ANOVA=1.
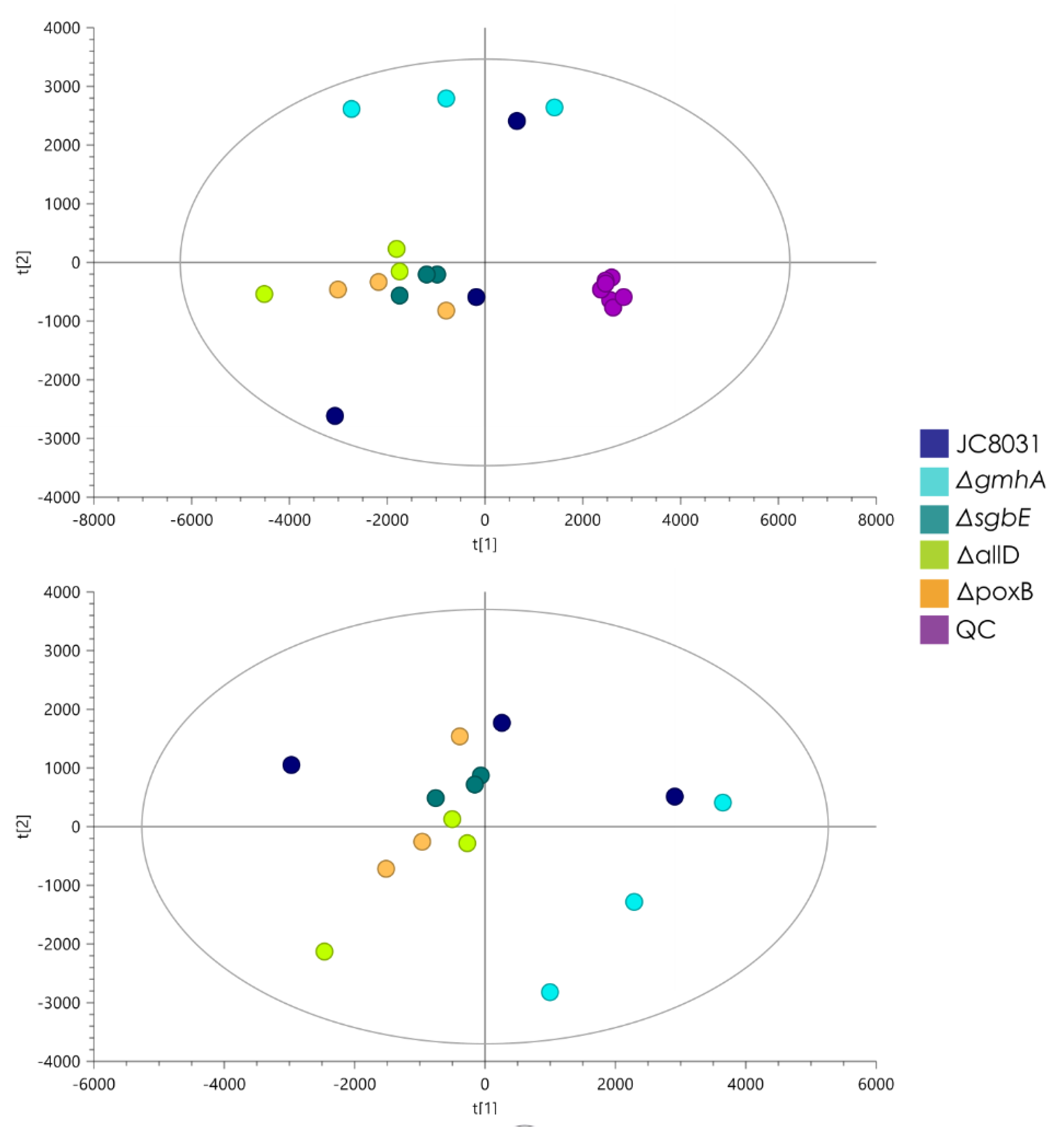

Table 1.
Gene deletions obtained through strain design using OptKnock.
| Gene | Keio Code | Description |
|---|---|---|
| poxB | B0871 | Peripheral membrane protein |
| sgbE | B3583 | L-ribulose-5-phosphate-4-epimerase |
| gmhA | B0222 | Phosphoheptose isomerase |
| allD | B0517 | Ureodiglycerol dehydrogenation |
Table 2.
CRISPR-Cas9 gRNA design.
| Target gene | N20 | Platform | Cassette size | Plasmid cut sites |
|---|---|---|---|---|
| gmhA | 5’ CGTGATCAAAGCGATCGCAG 3´ | pTarget (Jiang et al., 2015) | 1133 bp | XbaI, AvrII |
| sgbE | 5´ CATCGGCGCTCACAGCAAGG 3´ | pTarget (Jiang et al., 2015) | 1133 bp | XbaI, AvrII |
| allD | 5´ GCGACTACCGTACAGGCATG 3´ | pTarget (Jiang et al., 2015) | 1133 bp | XbaI, AvrII |
| poxB | 5´ GCGACTACCGTACAGGCATG 3´ | pTarget (Jiang et al., 2015) | 1133 bp | XbaI, AvrII |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



