
Submitted:
24 September 2024
Posted:
25 September 2024
You are already at the latest version
Abstract
Modification of canopy architecture is fundamental to productivity in grapevine. This research focused on evaluating the impact of canopy openness on photosynthetically active radiation, photosynthetic activity, and the physicochemical properties of fruit in Cabernet Sauvignon grapevines. A completely randomised design was used to compare open and closed canopies, with ten replicates per treatment (20 trees in total), during the vegetative growth period and after harvest. Key measurements included photosynthetically active radiation (PAR), daily light inte-gral (DLI), photosynthetic rate (PR), stomatal conductance (SC), intercellular CO2 concentration (IC), leaf area (LA), transpiration, ambient CO2 concentration, and temperature. Additionally, we assessed fruit quality parameters such as total soluble solids (TSS), glucose-fructose, total titratable acidity (TTA), pH, TSS/TTA, and total phenols (TP). significantly (p≤0.05) increased PAR, DLI, PR, IC, and LA, while after harvest, only PR and IC showed variation. Closed canopies increased CO2/H2O by 62.5%, while H2O/CO2 and temperature were higher in open canopies. Canopy openness increased contacts, gaps and visible sky and reduced leaf area index. Open canopy fruits showed higher TSS, glucose-fructose, pH, TSS/TTA and TP contents. Canopy openness is essential for improving light interception, photosynthetic efficiency and fruit quality in Cabernet Sauvi-gnon grapevine cultivated in northern Mexico.
Keywords:
climate change
; gas exchange
; photosynthetic rate
; solar radiation
; training system
; Vitis vinífera L
1. Introduction
It is estimated that by 2050, about 70% of the world's population, i.e. 6.79 billion people, will live in cities and urbanised areas. This will pose a challenge to agro-ecosystems and the supply of fresh horticultural products [1,2]. In addition, depletion and pollution of natural resources including water, soil and air [3] and climate change is likely to affect agricultural production and requiring adjustments to be made in agronomic management in most locations [4]. Among the effects of climate change as predicted by various models, are more extreme fluctuations in air temperature [5], in CO2 concentrations and in precipitation, including atmospheric humidity and cloud cover. Many of these atmospheric factors are interrelated and, together, are likely to bring about a net reduction in direct solar radiation and a corresponding increase in diffuse radiation. Such changes in the radiation environment will affect gas exchange and thus the photosynthetic performance of horticultural plants and so not only their biomass accumulation [6] but also both the yield and quality of their harvested products. Hence it becomes important that canopy design and management should be re-optimised so as to maximise the crop’s interception and utilisation of these increases in scattered light [7].
But light quality, intensity and direction influence not only the photosynthetic process but also plant development through photomorphogenesis. Light interception by the canopy is fluctuating and complex, hence good canopy design and management is critical to optimising light distribution among plant organs [8]. Previous studies have demonstrated the importance of diffuse radiation to productivity in perennial horticulture [9]. Photosynthesis and biomass increase are maximised at optimal light levels [10] but excessive light levels can cause photorespiration and irreversible cell damage [8]. Thus, maintaining an optimal light distribution within the canopy is key to maximising crop yield and quality under varying climate change scenarios [11].
Grapevine (Vitis vinifera L.) (Vitacea:Viteae) is among the crops that enjoy some of the highest cultural, nutritional and commercial importances around the world. However, the commercial production of this fruit requires good canopy management and efficient use of water, light and CO2 [12]. This requires the design of structures and training systems (vertical, lyra, pergola, among others) by pruning the canopy in winter or summer [13]. Among temperate perennial fruit plants, pruning of grapevines plays a key role in the spatial distribution and control of canopy components (leaves, shoots and branches) [14]. In addition, pruning brings about significant improvements in physiological response through carbohydrate synthesis derived from the efficient use of photosynthetically active radiation (PAR) with stomatal opening, gas exchange, photosynthetic activity and canopy architecture [15].
The productivity of any fruit species, including grapevine, is linked to the optimal use of PAR [1]. Previous studies on this crop have shown that canopy openness is a determining factor in fruit growth and development [16]. As a vine is a liana, the distribution of its branches, shoots and leaves depends on mechanical supports [12] and the implementation of open canopy involves hiring trained personnel and the removal of about 50% of the shoots, which significantly increases production costs.
Modern vine production systems include the optimisation of the shape and management of vegetative growth, aspects that have favoured the development and implementation of multiple training systems. In addition, these systems facilitate the mechanisation process and cost reduction in terms of manual pruning, shoot positioning, foliar spraying of pesticides, nutrients, as well as fruit harvesting [17]. Therefore, the establishment of new vineyards should consider annual pruning and the optimal selection of the training system according to the geographical location of the vineyard, soil type, climatic conditions and the destination of the fruit, always seeking to increase the exposure of the leaf surfaces to solar radiation [1,18].
Mexico is a nation with one of the greatest climatic diversities. This represents an opportunity for the establishment of vineyards for the commercial production of fruit for fresh consumption, dehydrated and processed in the form juices, jams and wines of high quality [19]. In addition, global climate change is causing increased incidences of extreme events in terms of air temperatures and precipitation rates and amounts. These events already affect the development of traditional fruit trees such as apple, peach and pecan nut. In this context, grapevines have been demonstrated to offer improved agronomic performances, including optimal development of fruit oenological attributes (soluble solids, acidity, pH, phenols, anthocyanins, tannins, etc.) and consequently a processed product of high quality [20]. Finally, understanding the relationship between biological and environmental factors modulating shoot growth and their interaction with reproductive organs is essential to optimise the regulation of carbon allocation between vegetative and reproductive organs, as well as to understand the canopy microclimate. Therefore, the objective of this research was to evaluate changes in photosynthetically active radiation, photosynthetic activity parameters, canopy architecture and some aspects of physicochemical fruit quality in response to open versus closed canopy managements in Cabernet Sauvignon grapevines.
2. Materials and Methods
2.1. Site, Plant Material and Experimental Design
This study was conducted in 2022 (June and September) and 2023 (June-July) in Meoqui, Chihuahua, Mexico (28°23′23″N,105°37′25″W) at an altitude of 1200 m, where mean annual temperature was 18.6 °C and precipitation was 369.8 mm. Grapevines of cv. Cabernet Sauvignon grafted on P-111 were used, these were 13 years old and planted with north-south rows at 4000 vines ha-1 (1 x 2.5 m). The horizontal training system (VSP) was defined as closed canopy. A V-shaped training system, defined as open canopy, was also implemented by pruning 50% of shoots at the cardinal points east and west.
The plants selected as the experimental units had average heights of 2.5 ± 0.5 m and trunk diameters of 35 ± 2 cm (measured at 20 cm above ground level). The vineyard was established on a loam/sandy textured soil with 55% sand, 24% silt, 20% clay in the top (0 to 35 cm) layer. Here the pH was 7.6, the organic matter 0.71% and the electrical conductivity 0.4 dS m-1. Soil nutrient levels were: 148 N and 15 P (kg ha-1); and 175 K+, 3200 Ca2+, 738 Mg2+, 0.32 Fe2+, 2.86 Mn2+, 1.62 Zn2+ and 3.46 Cu2+ (mg kg-1). Nutrients were applied to the soil surface at the end of March using a fertiliser of general formula 80 N: 80 P2O5: 100 K2O. Standard commercial practices for weed control and irrigation scheduling were used throughout the experiment.
Plants were selected randomly in the vineyard but uniformly for trunk perimeter. Two treatments were applied: ‘open canopy’ and ‘closed canopy’ each with ten replicates, where each vine was defined as an experimental unit. The intensity values of photosynthetically active radiation (PAR) were recorded and integrated daily light (DLI), with measurements of net photosynthesis, stomatal conductance, intracellular CO2 level, leaf area, transpiration, air CO2, air and leaf temperatures, the numbers and sizes of stomata, and the canopy architecture as the number of leaf layers, the leaf area index, the fraction (%) of canopy gaps and the fraction (%) of sky visible from below the canopy. These measurements were made at 08:00, 11:00, 14:00 and 17:00 h. Fruit quality was analysed with reference to canopy type and time based on the sampling dates of June 27 and 29, and July 4, 6, 10, 13, 18, 20, 23 and 27 2023.
2.2. Photosynthetically Active Radiation and Daily Light Integral
Photosynthetically active radiation was determined on mature, undamaged leaves in the middle part of the plant canopy using a 1 m long linear radiometer LI-191 (LI-COR, NE, USA) [21]. The radiometer was placed on a horizontal support, oriented parallel to the north: south planting line. The PAR readings were taken at three orientations (east, west and centre) of the plant canopy. In each orientation, three measurements were averaged for each radiometer angle (45°E, 45°W, incident light and ground reflected light). Data were recorded at 08:00, 11:00, 14:00 and 17:00 h. Results are expressed as µmol m-2 s-1.
The DLI was estimated according to the equation ∑PAR (µmol m-2 s-1) x 0.0144; where the factor 0.0144 corresponds to the total number of seconds of measurement per day, divided by 1,000,000 [22]. The determinations of PAR and DLI were carried out during berry ripening (on 9 June) and after harvest (on 22 September) 2022.
2.3. Leaf Area
Leaf area (LA) was determined according to the procedure described by [8] with slight modifications. Briefly, during the vegetative growth stage, 60 leaves were selected per canopy type (open and closed) undamaged leaves were selected from average canopies, from the cardinal points. Average leaf area was determined using a scanner (Cano Scan LIDE 100, Japan) and Photoshop CS software (Adobe, USA). The LA results are expressed in cm2.
2.4. Number and Dimensions of Stomata, Stomatal Pore and Guard Cell
Stomatal measurements were made on 9 June 2022 according to the method of [23] with slight modifications. Casts were made using transparent nail polish (i.e., nitrocellulose). A thin layer of nail polish was applied to the leaf surface (abaxial and adaxial) and allowed to dry for approximately 120 s. The film was then removed using transparent adhesive tape and placed on a slide to obtain an epidermal cast of the leaf. The number of stomata per mm2 was quantified using an optical microscope (Olympus BX51, NY, USA) with 20X objective and 0.15 mm2 field of view. The numbers and dimensions (length and width) of stomata, stomatal pore and guard cells were also determined.
2.5. Parameters of Photosynthetic Activity and Canopy Architecture
Determination of photosynthetic rate, stomatal conductance, intercellular CO2, transpiration, air CO2, temperature (air and leaf) and stomatal conductance were made on previously selected plants with mature leaves, recording canopy type (open or closed) and with four readings per experimental unit (two on the east and two on the west). These parameters were recorded during veraison (9 June) and after harvest (22 September 2022) at the times 08:00, 11:00, 14:00 and 17:00 h [18]. A non-dispersive infrared CO2 analyser LI-6200 (Li-Cor., Lincon, NE, USA) was used, equipped with a 1 L capacity chamber.
Prior to harvest on 9 June 2022, the number of leaf contacts (leaf layers), the fraction (%) of canopy gaps (GAPs), the fraction (%) of sky visible from below the canopy and the leaf area index were assessed. These data were recorded using a LI-2000 canopy analyser (Li-Cor, Inc., Lincoln, NE, USA), equipped with a fisheye hemispherical lens and an optical system with five concentric detectors useful for determining the distribution of canopy foliage at five angles (7, 23, 38, 53 and 68°). The device was placed on a structure composed of a vertical bar at the planting line and a vertically-mobile horizontal bar which facilitated measurements at the same point, but at different heights within the canopy (≈ 80 cm).
2.6. Fruit Quality
In this study, some physicochemical parameters of the fruit were evaluated. For this purpose, 200 fruits were harvested per canopy type and per sampling date (27, 29 June; 4, 6, 10, 13, 18, 20, 23 and 27 July 2023). Fruit selection was carried out considering their position in the cluster (shoulder, middle and lower) with three replicates per canopy type. The samples were transported in polystyrene boxes for analysis in the wine laboratory of the Faculty of Agrotechnological Sciences of the Autonomous University of Chihuahua.
Must extraction was performed manually by mashing the samples without breaking the seeds and then stored at 4°C. The must was centrifuged at 10,000 rpm for 15 min at 4°C and allowed to stand for 15 min, after which the supernatant was taken. Total soluble solids (TSS) was determined with an Atago ATC-1 hand-held refractometer (Atago, Japan) at 20 °C. The results are expressed in °Brix. D-glucose/D-fructose concentration was determined using the enzymatic method described by [24] with modifications. A 2 mL sample of juice was taken, centrifuged at 3,000 rpm for 15 min at 4°C and absorbance was then measured at 340 nm with a UV visible spectrophotometer Y15 (Biosystems, Spain). The results were expressed in g L-1. Total titratable acidity (TTA) in g L-1 of tartaric acid was determined by titration with 0.1 N NaOH and pH was determined using a HI-2002 potentiometer (Hanna Instruments, USA). Total phenols (TP) were determined by the Folin-Ciocalteu method [25]. The results were expressed as mg GAE L-1.
2.5. Statistical Analyses
The statistical analyses consisted of determining differences between open and closed canopy with respect to PAR, DLI, leaf area and photosynthetic parameters (photosynthetic activity, stomatal conductance, intercellular CO2, transpiration, air CO2, temperature (air and leaf) and stomatal conductance by means of paired samples, for which the means of the observations were grouped according to the angle of measurement of the radiometer, the orientation of the plant and the time of recording. Data on the number and dimensions of stomata, ostioles and guard cells and on canopy architecture, the number of leaf contacts or layers, fraction of GAPs (%), fraction of visible sky below the canopy (%) and leaf area index, were analysed with Student's t-test for independent samples. The values of GAPs and the fraction of sky visible from below the canopy were subjected to linear regression analysis. Finally, fruit quality data were analysed with a curvilinear regression model for which sampling date was considered the independent variable. All analyses were performed with the statistical analysis software SPSS (Statistical Package for the Social Sciences) 19.0.
3. Results
3.1. Photosynthetically Active Radiation and Photosynthetic Activity Parameters
Efficient canopy management combining severe winter pruning and summer pruning, including thinning of clusters, shoots and leaf removal in the fruiting zone prior to veraison, contribute to improved yield and fruit quality in grapevine [26]. In this regard, at the vegetative growth stage (9 June), the opening of the canopy in Cabernet Sauvignon plants significantly increased (p≤0.05) the interception of PAR, daily integrated light (DLI), photosynthetic rate (PR), stomatal conductance (SC), intracellular CO2 (CI) and leaf area (LA) with values of 415 µmol CO2 m-2 s-1, 5.9 mol d-1 m-2, 8.9 mmol CO2 m-2 s-1, 291 mmol CO2 mol-1 and 98 cm2, respectively (Table 1). In the evaluation of 22 September 2022 (post-harvest), both canopies showed statistically equal values (p>0.05) for PAR, DLI, SC and LA, where the only significant difference was for PR (4.2 mmol CO2 m-2 s-1) and CI (mmol CO2 mol-1) in the open canopy.
Compared with the closed canopy vines, the leaves expanding prior to fruit ripening in the open canopy vines had a significantly greater (p≤0.05) ratio between transpiration and CO2 uptake (H2O/CO2). The less efficient with a difference of about 33% (1180 vs 1760 g L-1) for this sampling date or phenological stage, therefore, monitoring the moisture level and minimising the occurrence of water stress linked to evapotranspiration should be considered. However, the change from closed to open canopy had no effect on transpiration, CO2/H2O, LT (leaf temperature) or AT (air temperature) parameters (Table 2).
After harvest, leaves of open and closed canopy plants did not differ in transpiration values (2.9 vs. 2.8 mmol H2O m-2 s-1). However, the value of the ration H2O/CO2 in the closed canopy was 62.5 % higher, while the H2O/CO2 ratio and leaf and ambient temperatures were higher in the open canopy plants with mean values of 359.5 g L-1, 32.9°C and 33.2°C, respectively (Table 2). No significant differences were found for the number of stomata per mm2 or for the dimensions (length and width) of stomata, stomatal pores and guard cells. However, the dimension values evaluated in plants with open canopy are slightly higher than those observed with closed canopy.

Table 3.
Number and dimensions of stomata, stomatal pore and guard cells of leaves of the grapevine cv Cabernet Sauvignon with canopy structure open or closed.
Table 3.
Number and dimensions of stomata, stomatal pore and guard cells of leaves of the grapevine cv Cabernet Sauvignon with canopy structure open or closed.
| Treatment | NS mm2 | LS | WS | LSP | SPO | WGC |
|---|---|---|---|---|---|---|
| Open canopy | 47.5 a | 31.62 a | 24.10 a | 13.62 a | 1.88 a | 8.42 a |
| Closed canopy | 47.5 a | 31.35 a | 23.15 a | 15.2 a | 1.55 a | 7.40 a |
1 NS: number of stomata; LS: length of stomata; WS: width of stomata; LSP: length of stomatal pore; SPO: stomatal pore opening; WGC: width of guard cell. Means with the same letter within a column are not significantly different (Student’s t test, p≤0.05). The data are expressed in µm.
Canopy architecture is described by the number of leaf contacts or layers, gaps between the canopy, visible sky below the canopy and leaf area index (Figure 1). The numbers of leaf layers and gaps were higher for open canopies with values of r2 = 0.87 leaf layers and 0.71 gaps. For canopy layers, a highly significant interaction (p≤0.001) was found between angle and canopy with a coefficient of determination for the model of 0.967. This means that canopy openness has a strong effect on plant development. The open canopy showed a higher percentage of visible sky with a lower leaf area index and no significant interaction (p≤0.05) in both cases with a coefficient of determination of 0.884 and 0.970, respectively.
3.2. Fruit Quality
Fruit quality is linked to leaf, branch and fruit pruning. In this study, the effect of canopy management, open vs closed, on several fruit quality parameters important for determining harvest and for processing including TSS, d-glucose/fructose ratio, TTA, pH, TSS/TTA and total phenols, was evaluated (Figure 2). Significant interactions between phenological stage and canopy management were found for all fruit quality parameters: TTA (p≤0.023), pH (p≤0.020), TSS/TTA (p≤0.000), total phenols (p≤0.041), 0.000, 0.007, respectively), where canopy types (open vs. closed canopy) were analysed individually the results was also significant (p ≤ 0.000, in all cases). Except for pH and TSS/TTA, phenological stage significantly affected TTA content, d-glucose/fructose ratio, TTA and total phenols (p ≤ 0.026, 0.026, 0.027, 0.037, respectively).
In general, fruits harvested from open canopies showed higher TSS values (between 14.09 and 21.8°Brix, r2= 0.637) and d-glucose/fructose ratios (between 129.97 and 214.93 g L-1, r2 = 0.636) than those from closed canopies (Figure 2). Likewise, pH 2.82 was higher in open canopies at the first sampling (27/06/2023, 178 Julian days) with an increasing trend, but at the end of the period of evaluations (27/07/2023, Julian day 207) open canopy pH 3.77 was less than closed canopy pH 4.03. The TTA results were the inverse of those for pH, i.e. the open canopy values lay between 14.91 and 5.73 g tartaric acid L-1 and the closed canopy values between 15.37 and 5.63 g tartaric acid L-1. A parameter quantifying flavour is linked to the dimensionless TSS/TTA ratio, where there is an inverse relationship between TSS content and tartaric acid content as a consequence of respiration and ripening (Jesús et al., 2021).
High correlations were observed for TSS and TTA (r2 = 0.853 and 0.794), where throughout the sampling period TSS/TTA values were higher in open canopy fruits (0.95/5.50) than closed canopy fruits (0.69/3.97). Of the secondary metabolites, phenols are in greatest abundance in the fruit, where they play a key role in the sensory properties (colour, astringency and bitterness) and are highly correlated with antioxidant activity [27]. In open canopy fruit (between 177 and 202 Julian days), the total phenols content was higher than in the closed canopy fruit, i.e. between 2080 and 2650 mg GAE L-1. In the final sampling at 207 Julian days, a reverse behaviour was observed for closed canopy fruits (3020 mg GAE L-1) (r2= 0.89 and 0.81, respectively).
4. Discussion
4.1. Photosynthetically Active Radiation and Photosynthetic Activity Parameters
Solar radiation plays an essential role in biomass production, but excess can cause photorespiration, organ and tissue damage and, consequently, irreversible damage to the photosynthetic apparatus [7]. Therefore, the determination of the total amount of PAR (400-700 nm) received by at plant surface is paramount, as it represents a useful tool for predicting the yield of any horticultural crop, including deciduous perennials such as the grapevine [3]. In this study, an open canopy in the grapevine cv. Cabernet Sauvignon increased the interception of photosynthetically active radiation and the daily light integral compared with a closed canopy, which is likely to bear a direct relationship with biomass accumulation, growth, irrigation needs and fruit composition and quality [21,28]. Climate change has increased the intensity of diffuse radiation, and the rates of gas exchange and consequently of canopy photosynthesis [10]. This is confirmed by our results in this study for photosynthetic rate, levels of intercellular CO2 and canopy architecture.
Photosynthesis is a complex biochemical process and is the basis of productivity in all autotrophic organisms, where the assimilation of atmospheric CO2 is central to the functioning of ecosystems and agroecosystems [1]. The efficiency of the photosynthetic systems of higher plants is very sensitive to variations in atmospheric (temperature, CO2 concentration etc) and edaphic conditions (water and salinity etc) [8]. With these abiotic factors affecting photosynthetic electron transfer and the photophosphorylation, key processes in the preservation of thylakoid membranes and the ultrastructure of chloroplast organelles [5] Drought has been shown to be the main abiotic stress factor limiting photosynthetic processes and thus crop plant productivity [6,28]. Recent advances in genomics, spatial modelling tools and canopy conduction systems have facilitated improvements in photosynthesis and yield in horticultural crops under climate change scenarios [3,29].
In deciduous perennial crops, such as in Cabernet Sauvignon grapevines, optimal flower and fruit development is generally related to current photosynthetic rates and to photo-assimilate reserves held in the secondary tissues of stem, branches and roots [8]. Leaves have been shown to sense and to respond not only to their own microenvironment but also to that of the other leaves on the same plant [21]. This behaviour is related some signal of systemic irradiance, where the shading of mature leaves causes significant changes in the anatomy of developing young leaves, including reductions in stomatal index and in leaf area [17,18]. One aspects of agronomic management in grape growing relates to management of the canopy. Here, cordon training systems, the number, spacing and vigour of shoots, and the removal of leaves in the fruiting zone [30] are all cultural practices that set out to reduce excessive shading within the canopy.
Canopy structure also affects soil microbial stability and microclimate [27], but the central objective is to maintain optimal levels of light interception and uniform light distribution within the canopy in order to increase the biological productivity of the plant as well as the quality of the harvested product [5]. The different grapevines genotypes exhibit a wide range of physiological responses with respect to drought tolerance, across the isohydric-anohydric spectrum, showing variations in stomatal control responses over evaporative demand and soil moisture with large fluctuations in leaf water potential [31]. When evaluating the effects of irrigation management on the relationship between stomatal conductance and stem water potential in the grapevine cv. Cabernet Sauvignon, [32], report an unclear behaviour with respect to the level of stomatal control, i.e. behaviour changed in this isohydric or anisohydric cultivar depending on the irrigation regime, leaf orientation and age, as well as the plant training system.
In field studies of radiation interception, photosynthesis, transpiration and growth analysis, leaf area estimation is a useful tool for identifying fast-growing and early-flowering genotypes [33]. Grapevines of cv. Cabernet Sauvignon have been shown to respond strongly to variations in agronomic management, including to irrigation and mineral nutrition, by varying their leaf area but this response has the potential for causing excessive shading which has direct effects on the physicochemical quality of the fruit and consequently of the wine [34].
The regulation of CO2 assimilation and transpiration are two physiological processes central to determining the rate of biomass accumulation and to temperature control in plant tissues, for which stomatal density and distribution vary with leaf surface structure [35]. The grapevine is hypostomatic, with both stomatal density (the number of stomata per unit leaf area) and stomata dimensions (width-length) varying among the various species and cultivars [19]. Our results show that stomatal density and stomatal dimensions (length and width) are similar in both open and closed canopy types. Similar results were reported by [36] when evaluating stomata density and size over two growing seasons and among a range of grapevine cultivars, including ‘Cabernet Sauvignon’ with density values between 36 and 41 stomata mm2. The factor(s) that determine variation in stomatal density are not clear. Some authors suggest it is genetically determined while others conclude it is associated with light intensity, or humidity, or air temperature etc [10,23]. These are all variables that, in our study with Cabernet Sauvignon were not modified by our opening vs. closed canopy structure.
Previous studies report an unclear relationship between stomatal dimensions (length and width) and stomatal density, because these may vary between growing seasons, species, cultivars and even clones [19]. However, it is known that cultivars with high stomatal densities tend to have higher stomatal conductance and water use efficiencies. Stomatal density is regulated by hydraulic and chemical signals as a response to water deficit and signalling linked to abscisic acid and leaf water potential [20]. When evaluating different irrigation levels and atmospheric vapour pressure deficits over two growing seasons in ‘Cabernet Sauvignon’ grapevines, [37] reported values of stomatal conductance that varied between 84.6 ± 23.5 and 456.6 ± 41.2 mmol m-2 s-1, where the reductions in stomatal conductance were linked to the lower water availabilities in the roots and to the increases in abscisic acid and symplastic fluid levels.
Canopy management (leaves and shoots) can be defined managements that adjust the canopy dimensions at the boundaries in space (i.e. canopy volume = width x height-x lenght), and the numbers×areas of the leaves within this volume. Canopy management still represents an active area of research in viticulture that over the years has generated multiple options for training systems, pruning (winter and summer), shoot positioning, leaf removal and vigour control [33]. This set of techniques seeks to improve the efficiency solar radiation interception, to minimise shading, to improve crop production and/or quality, to reduce the incidence of diseases and to facilitate mechanisation of pruning, pathogen control, weed control, fertilisation, harvest etc [32]. The leaves of most land plants (including of grapevines) absorb solar radiation between 400-700 nm. Only a small proportion (~6%) of this radiation is transmitted by the leaves, so light levels in the centre of a dense canopy is low, often only ~1% of that at the surface of the canopy [29] (Reshef et al., 2017). To minimise shading, one of the most common agronomic practices is to increase the proportion of gaps in the canopy [15], especially in the cluster/renewal zones, where uniformity in the microclimate is sought [27].
Light and air temperature modify the biology and physiology of the grapevine [33]. Under a scenario of climate change, with increasingly extreme fluctuations in air temperature and solar radiation, the photosynthetic apparatus can be subjected to conditions of light saturation and drastic reductions in photochemical efficiency are induced, leading to the phenomenon of chronic photoinhibition, acceleration of phenological phases and early ripening. These conditions do not favour obtaining quality fruit and, consequently, quality wine [3,17]. Interception of photosynthetically active radiation is related leaf area and so to the level of leaf shading [38]. In our study, data for number of contacts, percentage of visible sky and GAPs were higher for the open canopy with a leaf area index (LAI) <2 showing better light capture according to the Jefferies and Heilbronn model [39]. This model is used to describe the relation between LAI and percent ground cover to estimate intercepted radiation in multiple crops, including grapevine cv Cabernet Sauvignon [40].
The open canopy modification allows grapevines to acclimatise (i.e. alter their growth) in response to shade conditions by sensing different portions of the (PAR) spectrum (ultraviolet, blue, red and far-red) through a complex phototropic sensory system made up of different components including UVR8, cryptochromes, phototropins, ZTL-type receptors and phytochromes [41]. Hence, the grapevine cv. Cabernet Sauvignon’ under open canopy conditions is sensitive to variations in light levels and light quality. However, under closed canopy conditions, they cope with dim light and optimise light capture by increasing leaf area, leaf thinning (reductions in leaf mass per unit area) and accumulation of photosynthetic pigments, maximising efficiency in light interception and transformation of photons to photoassimilates [42]. All these morphological and physiological responses are linked to changes in water relations and atmospheric CO2 management.
Available evidence indicates that shading reduces transpiration, stomatal conductance, photosynthesis, stomatal density, hydraulic conductivity and water use efficiency [36]. Under the conditions of our study, differences were detected only for gas exchange and leaf temperature. However, during the postharvest fruiting stage, a non-significant reduction was observed for all parameters assessed in both canopy types. The leaf area index, which was higher for the closed canopy, may indicate lower light interception, affecting critical parameters such as transpiration, root development and photosynthetic capacity. Such physiological behaviours modify the optimal frequency of irrigation application, nutrient supply and may be yield limiting [11].
4.2. Quality of Fruit
The ripening of the grape berry for the production of high-quality wine represents a complex process that involves accumulations of some compounds (glucose, fructose, organic acids, polyphenols and aromatic substances) and reductions in others (tyramine, phenylethylamine, putresin and cadaverine) which can impart undesirable aromas to the wines [27]. Such fruit can be consumed as a fresh product, or dehydrated, or processed to juice or wine. Where harvesting aims to promote maximum quality, expressed in target sugar and organic acid contents, it is necessary to subject the plant to a certain level of water stress [9]. However, this does not correspond to oenological maturity, therefore, to establish the optimal harvest time it is necessary to consider soluble solids, acidity, aromas and polyphenols, i.e. phenolic maturity [4]. In red grape cultivars, including Cabernet Sauvignon, phenolic maturity is related to the evolution in the concentrations of anthocyanins and tannins in the epidermis ‘pomace’ and seeds [15]. However, training systems, vine age, water regime, soil type, environmental temperature and solar radiation are a set of biotic and abiotic factors that intervene in the accumulation and synthesis of phenolic compounds, responsible for colour, aroma and texture - the main sensory characteristics for any high-quality red wine [38].
The Cabernet Sauvignon cultivar has outstanding characteristics for red wine production with excellent quality but has shown varied behaviour with respect to its genetic characteristics, productivity and phenotypic quality, so a gradual selection of clones has been made to improve bunch size and fruit with greater homogeneity in aroma, colour and flavour [40]. Therefore, specific assessments of the effects of climate change on the wine industry are important to prioritise adaptation strategies [26], as most wine regions worldwide are exposed to increased occurrence of extreme events - temperature, hail, floods, drought etc [8]. These weather phenomena affect the mesoclimate and often cause a lag in vegetative growth and alterations in ripening and fruit composition. However, the intensity of the physiological response between vineyards can vary according to slope, altitude, surrounding vegetation and the primary and secondary branch [27].
The ‘pomace’ (epidermis and seeds) of the grape berry are characterised by a high content of mineral nutrients, vitamins and phytochemicals with high biological and antioxidant activity [13]. Most of these phytochemicals are polyphenols, including flavonoids (anthocyanins, flavonols, flavan-3-ols, flavones and chalcones) and non-flavonoids (phenolic acids, stilbenes, tannins, coumarins and neolignans) [40]. The concentrations of these phytochemicals vary with cultivar, with the vagaries of climate (temperature, radiation, water availability etc), with soil characteristics and with cultural practices, including training systems [26], where a dominant factor is the harvest time, determined by analysis of the juice (technological maturity), pulp and seeds (phenolic maturity) [16]. Therefore, grape berry quality is connected with the composition of the pulp, pomace and seeds and indirectly with the winemaking process [9]. In these climatic conditions, a VSP training system is widely used, due to its greater compatibility with mechanisation of agronomic activities (pruning, pathogen control, harvest etc) and commercial production at the regional level [16]. However, with more extreme seasonal variations in air temperature and solar radiation, VSP has favoured overexposure of the bunch and consequently the presence of sunburned fruit, lower yields, degradation of colour pigments (anthocyanins, flavonols and proanthocyanidins) and variation in fruit acidity [43].
The modification of leaf area prior to flowering by removing the basal leaves around the cluster (east or north in the Southern Hemisphere) during fruit development aims to improve the exposure of the cluster to solar radiation (to the west and south) and increase the content of soluble solids, anthocyanins and flavonols [44]. Traditionally in wine production systems, canopy management through shoot removal has made a significant contribution to secondary metabolism, affecting the numbers and concentrations of secondary metabolite molecules (phenols, flavonoids, anthocyanins etc) with high biological and antioxidant activity [45]. In addition, canopy management helps maintenance of the source-sink balance and improves cluster microclimate [5]. In this regard, [21] evaluates the effects of leaf and shoot removal on Merlot grapevines, reporting significant increases in the proportion of quercetin and kaempferol to the detriment of myricetin derivatives. However, the phytochemical and antioxidant composition of the fruit depends on a complex balance between compounds of primary and secondary metabolism, which are highly sensitive to environmental factors, including solar radiation. On the other hand, post-bloom leaf removal in Merlot vines [46] report a significant improvement in the reduction of Botrytis incidence and herbaceous aromas, but without affecting yield and bunch weight. In general, berry quality in grapevine is the net result of a multiplicity of agronomic practices, including pruning, irrigation, mineral nutrition, rootstock and, above all, training system [14].
5. Conclusions
This study highlights the significant benefits of modifying canopy architecture for enhancing productivity in Cabernet Sauvignon grapevines. Open canopies, compared to closed canopies, notably increased photosynthetically active radiation, photosynthetic rates, and key fruit quality parameters such as total soluble solids and total phenols. This underscores the importance of canopy openness in optimizing light interception and photosynthetic efficiency. However, to fully understand the impacts on grapevine productivity, further research should incorporate additional variables, including soil evaporation, rootstock, different cultivars, and various growing locations.
Author Contributions
original draft, E.H.-O.; conceptualisation, O.C.-A.; data curation, J.A.O.-A.; investigation, O.A.H.-R.; visualisation, supervision, R.A.-V.; software and methodology, J.L.J.-C.; writing – review & editing and resources, A.A.G.-B. and D.L.O.-B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Dr. Jorge Jiménez Castro for his valuable assistance in the statistical analysis of the data.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
AT: air temperature; Ci: intracellular CO2; DLI: daily light integral; LA: leaf area; LSP: length of stomatal pore; LS: length of stomata; LT: leaf temperature; NS: number of stomata; PAR: photosynthetically active radiation; PR: photosynthetic rate; SC: stomatal conductance; SPO: stomatal pore opening; Tr: transpiration; WGC: width of guard cell; WS: width of stomata.
References
- Yu R, Torres N, Tanner JD, Kacur SM, Marigliano LE, et al. Adapting wine grape production to climate change through canopy architecture manipulation and irrigation in warm climates. Frontiers in Plant Science. 2022;13:1015574. [CrossRef]
- Ontiveros-Capurata RE, Juárez-López P, Mendoza-Tafolla RO, Alia-Tejacal I, Villegas-Torres OG, et al. Relationship between chlorophyll and nitrogen concentration, and fresh matter production in basil ‘Nufar’ (Ocimum basilicum) with three handheld chlorophyll meter readings: SPAD, atLEAF and MC-100. Revista Chapingo. Serie Horticultura. 2022;28(3):189-202. [CrossRef]
- Gavhane KP, Hasan M, Singh DK, Kumar SN, Sahoo RN, et al. Determination of optimal daily light integral (DLI) for indoor cultivation of iceberg lettuce in an indigenous vertical hydroponic system. Scientific Reports. 2023;13(1):10923. [CrossRef]
- Furini G, de Oliveira Anese R, Reinehr J, Silva FN, Casa RT, et al. Ethephon effect on defoliation of cluster zone, Botrytis bunch rot, and viticultural performance of ‘Cabernet Sauvignon’ grapevine in highland region of southern Brazil. Ciencia Rural. 2024;54(6):e20230207. [CrossRef]
- Durand M, Murchie EH, Lindfors AV, Urban O, Aphalo PJ, et al. Diffuse solar radiation and canopy photosynthesis in a changing environment. Agricultural and Forest Meteorology. 2021;311:108684. [CrossRef]
- Dechant B, Ryu Y, Badgley G, Zeng Y, Berry JA, et al. Canopy structure explains the relationship between photosynthesis and sun-induced chlorophyll fluorescence in crops. Remote Sensing of Environment. 2020;241:111733. [CrossRef]
- Li Z, Zhao T, Liu J, Li H, Liu B. Shade-induced leaf senescence in plants. Plants. 2023;12(7):1550. [CrossRef]
- Liu X, Xu Y, Wang Y, Yang Q, Li Q. Rerouting artificial light for efficient crops production: A review of lighting strategy in PFALs. Agronomy. 2022;12(5):1021. [CrossRef]
- Jediyi H, Naamani K, Elkoch AA, Dihazi A, El Fels AE, et al. First study on technological maturity and phenols composition during the ripeness of five Vitis vinifera L grape varieties in Morocco. Scientia Horticulturae. 2019;246:390-7. [CrossRef]
- Emmel C, D'Odorico P, Revill A, Hörtnagl L, Ammann C, et al. Canopy photosynthesis of six major arable crops is enhanced under diffuse light due to canopy architecture. Global Change Biology. 2020;26(9):5164-77. [CrossRef]
- Neo DCJ, Ong MMX, Lee YY, Teo EJ, Ong Q, et al. Shaping and tuning lighting conditions in controlled environment agriculture: A review. ACS Agricultural Science & Technology. 2022;2(1):3-16. [CrossRef]
- Torregrosa L, Carbonneau A, Kelner JJ. The shoot system architecture of Vitis vinifera ssp. sativa. Scientia Horticulturae. 2021;288:110404. [CrossRef]
- Jogaiah S, Striegler KR, Bergmeier E, Harris J. Influence of canopy management practices on canopy characteristics, yield, and fruit composition of ‘Norton’ grapes (Vitis aestivalis Michx). International Journal of Fruit Science. 2013;13(4):441-58. [CrossRef]
- Prieto JA, Louarn G, Pérez-Peña J, Ojeda H, Simonneau T, et al. A functional–structural plant model that simulates whole-canopy gas exchange of grapevine plants (Vitis vinifera L.) under different training systems. Anales de Botánica. 2020;126(4):647-60. [CrossRef]
- Gao XT, Sun D, Wu MH, Li HQ, Liu FQ, et al. Influence of cluster positions in the canopy and row orientation on the flavonoid and volatile compound profiles in Vitis vinifera L. Cabernet franc and Chardonnay berries. Food Research International. 2021;143:110306. [CrossRef]
- Nistor E, Dobrei A, Dobrei A, Ciorica G. Climate variability and canopy management influence on grape berries quality in Merlot and Pinot Noir varieties. Scientific Papers. Series B. Horticulture. 2020;64(1):299-306.
- Greer DH, Ghannoum O. Changes in photosynthesis and chlorophyll a fluorescence in relation to leaf temperature from just before to after harvest of Vitis vinifera cv. Shiraz vines grown in outdoor conditions. Functional Plant Biology. 2021;49(2):170-85. [CrossRef]
- Wu YS, Gong WZ, Wang YM, Yang WY. Shading of mature leaves systemically regulates photosynthesis and leaf area of new developing leaves via hormones. Photosynthetica. 2019;57(1):303-10. [CrossRef]
- Espinoza S, Ortega-Farías S, Ahumada-Orellana L. Characterization of stomatal density and size of different Vitis vinifera L. cultivars growing in Mediterranean climate conditions. Ciência e Técnica Vitivinícola. 2024;39(1):196-208. [CrossRef]
- Pereira GE, Padhi EM, Girardello RC, Medina-Plaza C, Tseng D, et al. Trunk girdling increased stomatal conductance in Cabernet Sauvignon grapevines, reduced glutamine, and increased malvidin-3-glucoside and quercetin-3-glucoside concentrations in skins and pulp at harvest. Frontiers in Plant Science. 2020;11:707. [CrossRef]
- Wang X, De Bei R, Fuentes S, Collins C. Influence of canopy management practices on canopy architecture and reproductive performance of Semillon and Shiraz grapevines in a hot climate. American Journal of Enology and Viticulture. 2019;70(4):360-72. [CrossRef]
- Korczynski PC, Logan J, Faust JE. Mapping monthly distribution of daily light integrals across the contiguous United States. HortTechnology. 2002;12(1):12-6. [CrossRef]
- Sánchez C, Fischer G, Sanjuanelo DW. Stomatal behavior in fruits and leaves of the purple passion fruit (Passiflora edulis Sims) and fruits and cladodes of the yellow pitaya [Hylocereus megalanthus (K. Schum. ex Vaupel) Ralf Bauer]. Agronomía Colombiana. 2013;31(1):38-47.
- Olmedo L, Henning MF, Pappalardo B, García SM, Pellon-Maison M. Validation of an enzymatic colorimetric assay for fructose content determination in soft drinks. Revista Española de Nutrición Humana y Dietética. 2021;25(1):69-77. [CrossRef]
- Waterman PG, Mole S. Analysis of Phenolic Plant Metabolites. Oxford, UK: Blackwell Scientific Publications; 1994.
- Rescic J, Mikulic-Petkovsek M, Rusjan D. The impact of canopy managements on grape and wine composition of cv. ‘Istrian Malvasia’ (Vitis vinifera L.). Journal of the Science of Food and Agriculture. 2016;96(14):4724-35. [CrossRef]
- Bubola M, Sivilotti P, Janjanin D, Poni S. Early leaf removal has a larger effect than cluster thinning on grape phenolic composition in cv. Teran. American Journal of Enology and Viticulture. 2017;68(2):234-42. [CrossRef]
- Sanchez-Rodriguez LA, Spósito MB. Influence of the trellis/training system on the physiology and production of Vitis labrusca cv. Niagara Rosada in Brazil. Scientia Horticulturae. 2020;261:109043. [CrossRef]
- Reshef N, Walbaum N, Agam N, Fait A. Sunlight modulates fruit metabolic profile and shapes the spatial pattern of compound accumulation within the grape cluster. Frontiers in Plant Science. 2017;8:70. [CrossRef]
- Sebastian V, Nicolas O, Alvaro G, Samuel OF. Effect of irrigation management on the relationship between stomatal conductance and stem water potential on cv. Cabernet Sauvignon. BIO Web of Conferences. 2023;56:01012. [CrossRef]
- Anić M, Osrečak M, Andabaka Ž, Tomaz I, Večenaj Ž, et al. The effect of leaf removal on canopy microclimate, vine performance and grape phenolic composition of Merlot (Vitis vinifera L.) grapes in the continental part of Croatia. Scientia Horticulturae. 2021;285:110161. [CrossRef]
- Zhang J, Serra S, Leisso RS, Musacchi S. Effect of light microclimate on the quality of ‘d’Anjou’ pears in mature open-centre tree architecture. Biosystems Engineering. 2016;141:1-11. [CrossRef]
- Munitz S, Schwartz A, Netzer Y. Effect of timing of irrigation initiation on vegetative growth, physiology and yield parameters in Cabernet Sauvignon grapevines. Aust J Grape Wine Res. 2020;26(3):220-32. [CrossRef]
- Munitz S, Schwartz A, Netzer Y. Water consumption, crop coefficient and leaf area relations of a Vitis vinifera cv. 'Cabernet Sauvignon' vineyard. Agric Water Manag. 2019;219:86-94. [CrossRef]
- Hunter JJ, Volschenk CG, Zorer R. Vineyard row orientation of Vitis vinifera L. cv. Shiraz/101-14 Mgt: climatic profiles and vine physiological status. Agricultural and Forest Meteorology. 2016;228:104-19. [CrossRef]
- Boso S, Gago P, Alonso-Villaverde V, Santiago JL, Martinez MC. Density and size of stomata in the leaves of different hybrids (Vitis sp.) and Vitis vinifera varieties. Vitis. 2016;55(1):17-22. [CrossRef]
- Speirs J, Binney A, Collins M, Edwards E, Loveys B. Expression of ABA synthesis and metabolism genes under different irrigation strategies and atmospheric VPDs is associated with stomatal conductance in grapevine (Vitis vinifera L. cv Cabernet Sauvignon). Journal of Experimental Botany. 2013;64(7):1907-16. [CrossRef]
- Hunter JK, Tarricone L, Volschenk C, Giacalone C, Melo MS, et al. Grapevine physiological response to row orientation-induced spatial radiation and microclimate changes. Oeno One. 2020;54(2):411-33. [CrossRef]
- Römermann C, Bucher SF, Hahn M, Bernhardt-Römermann M. Plant functional traits–fixed facts or variable depending on the season? Folia Geobotanica. 2016;51:143-59. [CrossRef]
- Niimi J, Tomic O, Næs T, Bastian SE, Jeffery DW, et al. Objective measures of grape quality: From Cabernet Sauvignon grape composition to wine sensory characteristics. LWT - Food Science and Technology. 2020;123:109105. [CrossRef]
- Sams B, Bramley RG, Sanchez L, Dokoozlian N, Ford C, et al. Remote sensing, yield, physical characteristics, and fruit composition variability in Cabernet Sauvignon vineyards. American Journal of Enology and Viticulture. 2022;73(2):93-105. [CrossRef]
- González CV, Prieto JA, Mazza C, Jeréz DN, Biruk LN, et al. Grapevine morphological shade acclimation is mediated by light quality whereas hydraulic shade acclimation is mediated by light intensity. Plant Science. 2021;307:110893. [CrossRef]
- Torres N, Martínez-Lüscher J, Porte E, Kurtural SK. Optimal ranges and thresholds of grape berry solar radiation for flavonoid biosynthesis in warm climates. Frontiers in Plant Science. 2020;11:931. [CrossRef]
- Güler S, Kunter B, Şehit A. Stomatal density, type and their relationships with leaf morphological traits in Vitis vinifera L. varieties. International Journal of Agriculture Environment and Food Sciences. 2023;8(1):78-87. [CrossRef]
- Sivilotti P, Herrera JC, Lisjak K, Česnik HB, Sabbatini P, et al. Impact of leaf removal applied before and after flowering on anthocyanin, tannin, and methoxypyrazine concentrations in ‘Merlot' (Vitis vinifera L.) grapes and wines. Journal of Agricultural and Food Chemistry. 2016;64:4487-96. [CrossRef]
- Alonso R, Muñoz F, Bottini R, Piccoli P, Berli FJ. Effects of wind exposure and deficit irrigation on vegetative growth, yield components and berry composition of Malbec and Cabernet Sauvignon. Plants. 2024;13(10):1292. [CrossRef]
Figure 1.
Number of contacts or layers of leaves, spaces or GAPs between canopy, spaces or gaps between canopy, sky visible under canopy and leaf area index. Bars represent the standard error ± 1.
Figure 1.
Number of contacts or layers of leaves, spaces or GAPs between canopy, spaces or gaps between canopy, sky visible under canopy and leaf area index. Bars represent the standard error ± 1.
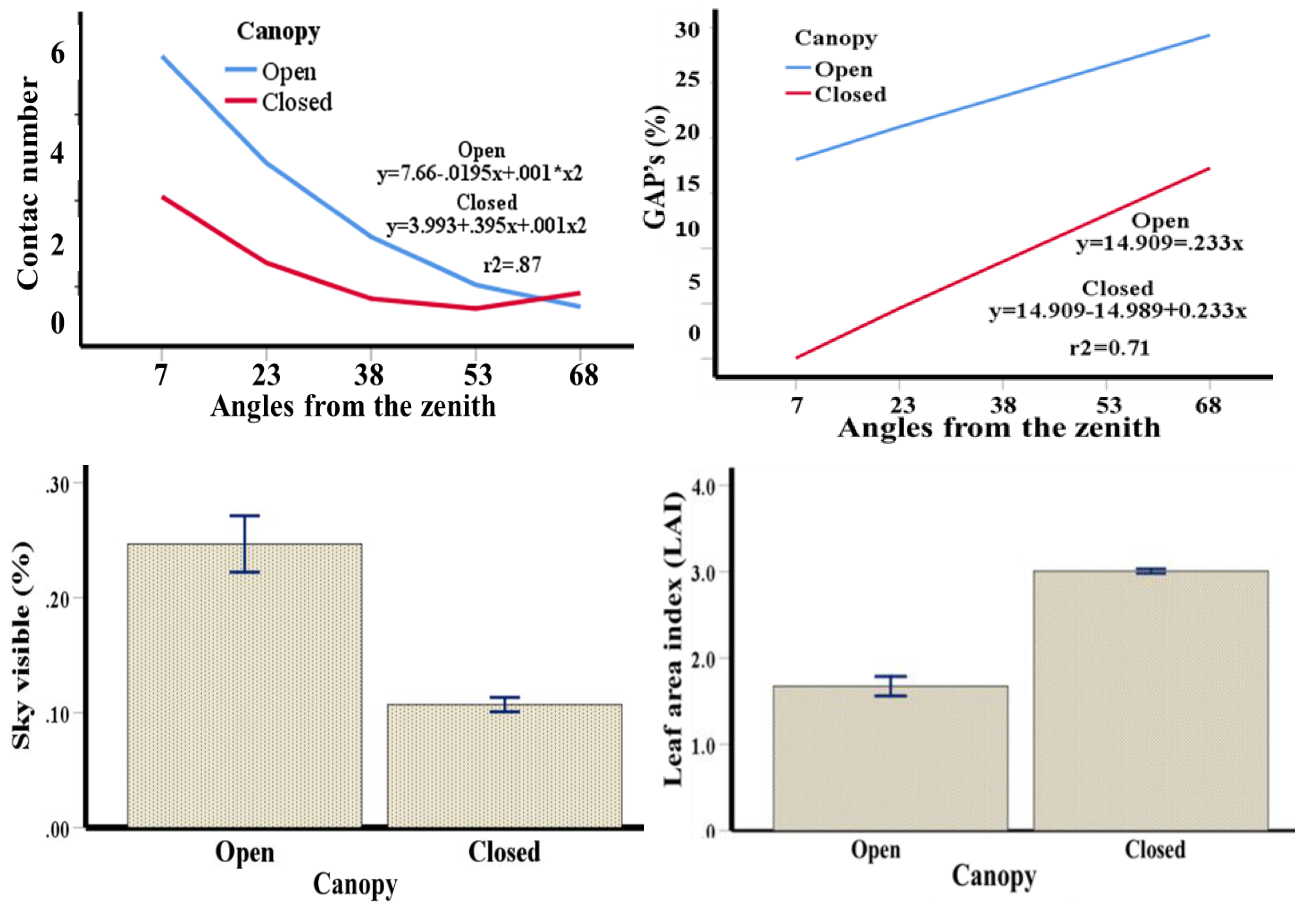
Figure 2.
Changes in fruit quality with time in the grapevine cv Cabernet Sauvignon managed with closed and open canopies.
Figure 2.
Changes in fruit quality with time in the grapevine cv Cabernet Sauvignon managed with closed and open canopies.
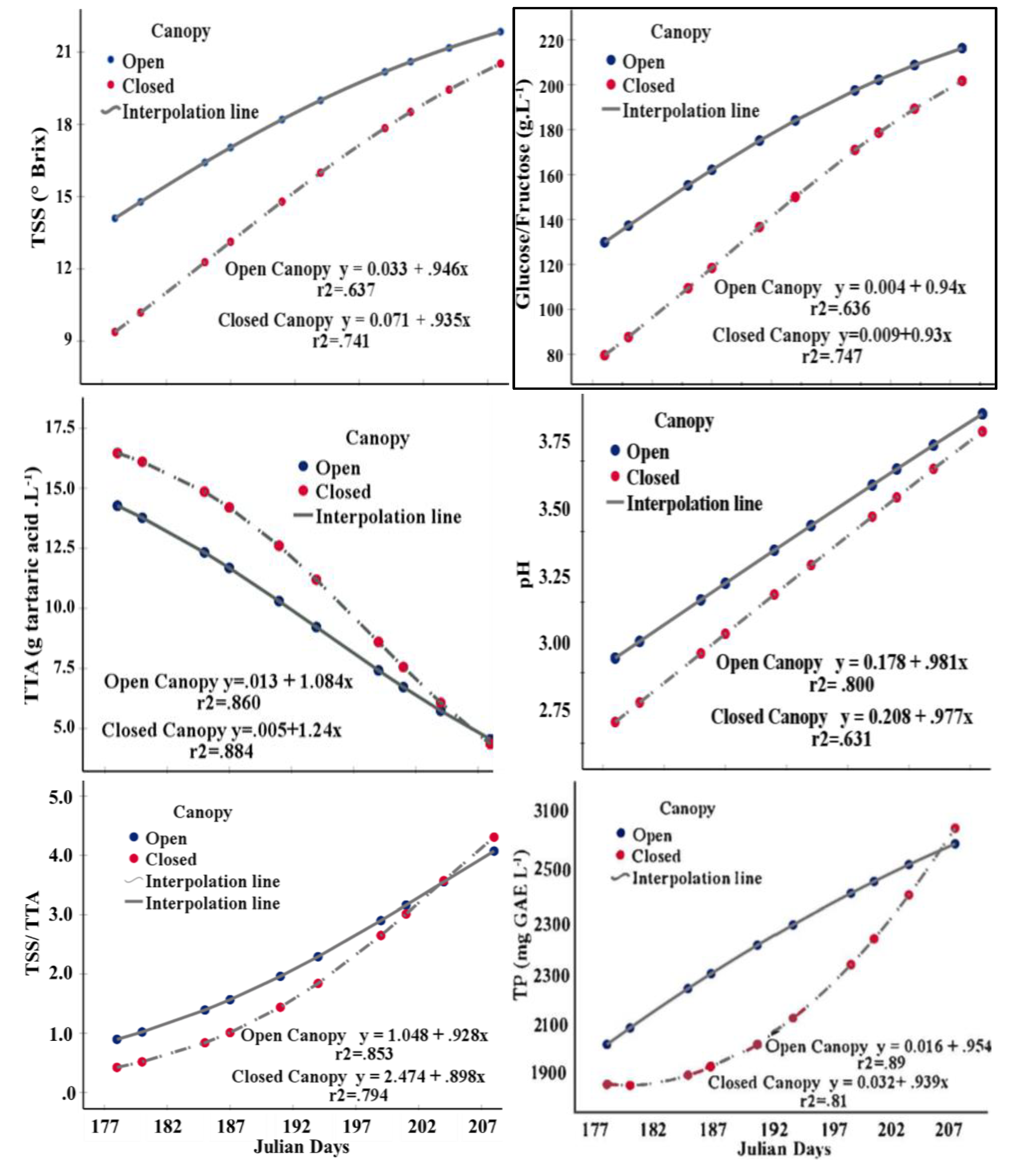
Table 1.
Changes in photosynthetically active radiation, daily light integral, photosynthetic rate, stomatal conductance, intracellular CO2 and leaf area in the grapevine cv Cabernet Sauvignon with canopy structure, open or closed.
Table 1.
Changes in photosynthetically active radiation, daily light integral, photosynthetic rate, stomatal conductance, intracellular CO2 and leaf area in the grapevine cv Cabernet Sauvignon with canopy structure, open or closed.
| Treatment | 9 June 2022 (vegetative growth) | |||||
|---|---|---|---|---|---|---|
| PAR | DLI | PR | SC | CI | LA | |
| Open canopy | 415.9 a | 5.9 a | 8.9 a | 150.1 a | 291.4 a | 97.92 a |
| Closed canopy | 266.0 b | 3.8 b | 7.2 b | 140.2 a | 272.1 b | 95.20 b |
| 22 September 2022 (after harvest ) | ||||||
| Open canopy | 320.7 a | 4.6 a | 4.2 a | 90.34 a | 304.5 a | 91.39 a |
| Closed canopy | 298.6 a | 4.3 a | 3.0 b | 90.82 a | 289.6 b | 90.67 a |
1 PAR: photosynthetically active radiation (µmol m-2 s-1); DLI: daily light integral (mol d-1 m-2); PR: photosynthetic rate (mmol CO2 m-2 s-1); SC: stomatal conductance (mol H2O m-2 s-1); Ci: intracellular CO2 (mmol CO2 mol-1); LA: leaf area (cm2). Means with the same letter within a column are not different (Student’s t test, p≤0.05).
Table 2.
Changes in transpiration, gas exchange and leaf and air temperature in grapevine cv Cabernet Sauvignon with an open or a closed canopy structure.
Table 2.
Changes in transpiration, gas exchange and leaf and air temperature in grapevine cv Cabernet Sauvignon with an open or a closed canopy structure.
| Treatment | 9 June 2022 (vegetative growth) | ||||
|---|---|---|---|---|---|
| Transpiration | CO2 /H2O | H2O/CO2 | LT | AT | |
| Open canopy | 6.4 a | 3.5 a | 1180.4 b | 35.9 a | 36.9 a |
| Closed canopy | 6.0 a | 3.4 a | 1760.4 a | 36.2 a | 37.3 a |
| 22 September 2022 (after harvest) | |||||
| Open canopy | 2.9 a | 4.0 a | 359.5 a | 32.9 a | 33.2 a |
| Closed canopy | 2.8 a | 2.5 b | 355.2 b | 32.17 b | 32.4 b |
1 Tr: transpiration (mmol H2O m-2 s-1); CO2/H2O: g L-1; H2O/CO2: g L-1; LT: leaf temperature (°C) and AT: air temperature (°C). Means with the same letter within a column are not different (Student’s t test, p≤0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



