
Submitted:
24 September 2024
Posted:
26 September 2024
You are already at the latest version
Abstract
A number of standard molecules are used for the molecular and histological characterization of lymphatic endothelial cells (LECs) such as Lyve1, Podoplanin (D2-40), VEGFR3, Prox1 and CD31. The number of molecules whose mutations cause lymphatic malformations or primary congenital lymphedema is considerable, but the majority of these diseases have not yet been characterized at the molecular level. Therefore, there is still considerable scope for molecular and functional studies of the lymphatic vasculature. Using RNASeq, we have previously characterized LECs under normoxic and hypoxic conditions. We used this information to compare it with immunohistochemical data. We carried out some of the immunohistology ourselves, and systematically studied the Human Protein Atlas (https://www.proteinatlas.org/). Here we describe molecules that are expressed at RNA and protein levels in LECs, hoping to stimulate future functional studies of these molecules.
Keywords:
Lymphatic endothelial cell
; ANKRD37
; CAV1
; CAV2
; CD59
; CNN3
; DYSF
; KANK3
; MARCKSL1
; MMRN1
; NXN
; SPTAN1
; SPTBN1
1. Introduction
The lymphatic vascular system was already known in ancient times, although its functions were misinterpreted. Venae albae or Ductus lactei were already known to Hippocrates of Kos (460-370 BC), Aristoteles (384 – 322 BC), and the doctors of the Alexandrian school (approx. 300 BC – 600 AC) (cited from [1]). And when Gaspare Aselli (re)discovered the lymphatics [2], he knew very well what he had to look for. He realized that the Venae albae were easy to find in the mesenteries e.g. of dogs, but he was probably the first to make a connection of these milky vessels with food intake. The anatomical representation of the lymphatic vascular system was especially advanced at Italian universities [3], although the function of the system was still very mysterious. One of the first to recognize the importance of directed fluid transport in the lymphatics was [4]. Contrary to the prevailing doctrine, which postulated a lymph flow into the liver, he described the connection to the central venous system. He also recognized a valve system in lymphatics and the coagulability of the lymph.
We should be very humble in evaluating ancient knowledge, because one thing is certain: we are still far from fully understanding the functions of the lymphatic vascular system, although more and more functions are being discussed [5,6,7,8]. The complexity is immediately apparent when we visualize the heterogeneity of lymphatic endothelial cells (LECs) [9], and the long lists of molecules expressed in LECs [10,11], lymph collectors [12] or lymph nodes [13,14]. Thereby, development and behavior of LECs is critically regulated by the transcription factor PROX1 [15,16,17]; and the histological characterization of human LECs is typically performed with antibodies against PROX1 and CD31 [18], the Vascular endothelial growth factor receptor-3 (VEGFR3, FLT4) [19], the Lymphatic vessel endothelial hyaluronic acid receptor 1 (LYVE1) [20], and the type-I integral membrane glycoprotein Podoplanin (PDPN) [21].
Congenital malformations of the blood vascular system are very common, those of the lymphatic system are much rarer, mostly located in the head-neck region and can be life threatening. Most of the lymphatic malformations are caused by somatic mutations in genes involved in the VEGFR3 signaling pathway [22,23,24,25]. However, most genetic causes of lymphatic malformations and primary lymphedema have not yet been discovered. This illustrates that there are still a very large number of molecules whose significance for the development and function of the lymphatic vessels is not yet known. We have previously used RNASeq to study human foreskin-derived LECs under normoxic and hypoxic conditions, and defined 162 genes that are significantly regulated by hypoxia [10], as well as highly expressed genes that influence the composition of the extracellular matrix and may be involved in lymphedema-induced fibrosis [26]. Here, we went through the RNASeq list and studied the large number of highly expressed LEC genes. We compared RNA expression with protein expression by systematically studying the Human Protein Atlas [27] (https://www.proteinatlas.org/). In this way, we sought to define additional molecules that can be used for deeper characterization of lymphatic vessels and to define further functions of LECs.
2. Results and Discussion
We recently performed expression analyses of three well characterized human foreskin-derived LEC lines under normoxic and hypoxic conditions [10,26], and received a list of approx. 16,000 LEC-expressed genes. Only a very small number of these molecules have been studied in LECs so far. We have tried to systematically match RNA expression with protein expression using the Human Protein Atlas [27]. We have concentrated on molecules whose function has not yet been investigated in LECs. However, the number of matches between RNA and protein expression was not very high, but there can be many technical reasons (fixation time of tissue, paraffine permeability of the antibodies, etc.) for this.
One of the main functions of the lymphatics is regulation of fluid homeostasis. The fluid is mainly absorbed via the flexible microvalves of the initial lymphatic vessels. However, part of the fluid uptake also occurs by means of active transcytosis, which has also been observed for the uptake of chylomicrons by lacteals [28,29,30]. Very recently the uptake of myofibroblast-derived microvesicles (MVs) was studied in human dermal BECs and LECs [31]. The authors describe that MVs cross an LEC layer but not a BEC layer in vitro. Internalization into BECs is mainly via the caveolin-dependent pathway, while LECs mainly use the macropinocytosis pathway. High expression of caveolin 1 (CAV1) and CAV2 (Table 1), for endo- and transcytosis, seems to be in line with the free passage of MVs through LECs; and antibodies against CAV1 and CAV2 clearly stain lymphatics, and selected blood vessels (Figure 1 and Figure 2).
Very active transcytosis by LECs is also reflected by the RNA expression the ferlin family members: myoferlin (MYOF; very high), dysferlin (DYSF; high), and otoferlin (OTOF; moderate) (Table 1). Ferlins regulate membrane fusion, and fusion of vesicles to cell membranes e.g. for exocytosis or membrane regeneration [32]. Of the three ferlins, we observed clear immunostaining against DYSF (dystrophy-associated fer-1-like protein) in various organs (Figure 3) and performed immunodouble staining with CD31 in human foreskin (Figure 4). We observed DYSF in dermal lymphatics. Blood vessels were mostly negative, except for the subepithelial capillary plexus. We tested various OTOF antibodies, but did not receive a positive result; and, in contrast to the high RNA expression, we found no immunopositivity for MYOF in the Human Protein Atlas. However, DYSF is clearly present in LECs, which is in line with high transcytotic activity.
Both the initial lymphatic vessels and the lymph collectors exhibit strong functional fluctuations in their diameter, and they have valves, whose elasticity is of great importance for reliable valve closure. Spectrins have originally been identified as the major elastic component of erythrocytes, linking the actin cytoskeleton to the cell membrane. However, nonerythroid spectrins have also been identified [33,34,35]. Spectrins form tetrameric proteins of alpha and beta subunits. SPTAN1 (spectrin, alpha, nonerythrocytic 1) and SPTBN1 (spectrin, beta, nonerythrocytic 1) are highly expressed in LECs (Table 1), and they are well detectable at protein level in lymphatics of various organs (Figure 5 and Figure 6). Neuropathies related to malfunction of the two molecules have been observed, however, lymphedema has not been described yet.
Among the genes most highly expressed in LECs is Multimerin 1 (MMRN1) (Table 1). Lymph contains fibrinogen and can coagulate [36], and we have previously pointed out that LECs are an important source for factor VIII (F8, antihemophilic globulin A) and its carrier protein von-Willebrand factor (VWF) [8]. MMRN1 is a specific coagulation factor V binding platelet protein with a role in hemostasis and coagulation, and accordingly it is highly expressed in megakaryocytes [37]. Besides that, it is found with extremely high specificity in endothelial cells, however, a clear distinction between BECs and LECs has not been made [38]. However, it appears that LECs are clearly stronger positive than BECs (https://www.proteinatlas.org/ENSG00000138722-MMRN1/single+cell+type). We found immunopositivity for MMRN1 in lymphatics of various organs (Figure 7). During lymphostasis, the accumulation of pro-coagulatory factors (F8, VWF, MMRN1) and the decrease in the anti-coagulatory and anti-inflammatory 5'-nucleotidase (CD73) of the LECs [39] can be important reasons for the increased tendency to thrombosis and inflammation.
The MARCKS-like protein-1 (MARCKSL1 = MLP) has great similarity with the myristoylated, alanine-rich protein MARCKS [40,41], a substrate for protein kinase C. MARCKSL1 has mainly been studied in neural development, and in cancer cell migration [42]. High MARCKSL1 expression has a strong prognostic value in lymph node-negative breast cancer patients [43]. Upon phosphorylation, MARCKSL1 induces actin bundling and inhibits cell migration [42]. We observed high RNA expression of MARCKSL1 in LECs (Table 1), and immune-positive lymphatics in various organs (Figure 8). The influence of MARCKSL1 on LEC stability and function has not been studied yet.
Another molecule associated with the actin cytoskeletal system is calponin 3 (CNN3). It consists of an acidic C terminus and a basic N terminus [44,45]. CNN3 regulates contractility of actomyosin-containing stress fibers of non-muscle cells [46]. Thereby, control of stress fiber contractility by CNN3 was found to be associated with mechanosensitive Yap/Taz (Yes-associated protein/transcriptional coactivator with PDZ binding motif) transcriptional activation [47]. We observed high RNA expression of CNN3 in LECs (Table 1) and immune-positive lymphatics in various organs (Figure 9), making LECs an attractive model for further studies on the function of CNN3. The importance of the Yap/Taz signaling in the Hippo pathway for lymphangiogenesis has been well recognized [48,49].
The cytoskeleton of LECs appears to be very stable, and accordingly, the invasiveness of LECs is usually extremely low; the only exception being the vanishing bone disease (Gorham-Stout disease, GHS) with approx. 350 cases being reported [50,51,52]. The ankyrin repeat domain protein 37 (ANKRD37) is not only associated with preeclampsia during pregnancy. Its knock-down enhances trophoblast invasiveness, migration, and regulation of key invasion proteins [53]. ANKRD37 is expressed at very low levels in LECs and significantly upregulated by hypoxia (Table 1) [26]. It is a hypoxia-inducible factor-1 (HIF1) target gene [54]. Its expression in GHS has, to the best of our knowledge, not been studied yet. We observed immune-positivity for ANKRD37 in LECs (Figure 10), however, due to the large number of ANKRD family members expressed in LECs (data not shown), specificity of antibody staining might be problematic.
Another molecule involved in actin stress fiber formation, and containing an ankyrin repeat domain, is KANK3 (KN motif- and ankyrin repeat domain-containing protein 3) [55]. It is highly expressed in LECs (Table 1), and immune-positivity is seen in lymphatics of various organs (Figure 11). The four members of the KANK family regulate integrin-mediated adhesion, actomyosin contractility, and link focal adhesions to the cortical microtubule stabilization complex [56]. Specific expression of KANK3 in endothelial cells has been noted [56], but functional studies are lacking.
Lymph contains high amounts of immunoglobulin G (IgG) heavy chain [36]. Antigen-IgG complexes represent a starting point for the classical complement system, which can stimulate phagocytes, inflammation and the cell-killing membrane attack complex (MAC) [57]. LECs express high amounts of CD59, aka protectin (Table 1), a glycoprotein functioning as a membrane-bound inhibitor of MAC [58]. Malfunctioning of CD59 causes hemolytic anemia with immune-mediated polyneuropathy [59]. The specific function in LECs has not been investigated yet. Immune-positivity is seen in lymphatics and blood vessels (Figure 12), suggesting an important immune-suppressive function in vessels. Strong expression of CD59 in colorectal cancer is associated with higher incidence of lymph node metastasis [55]. In ulcerative colitis, a chronic inflammatory disease of the colon, CD59 belongs to the complement components that have shown to safeguard the intestinal barrier and reduce intestinal inflammation [60].
It was previously shown that the non-canonical WNT (Wingless-type MMTV integration site) signaling pathway is of great importance for the elongation of lymphatics during embryonic lymphangiogenesis [61,62]. Nucleoredoxin (NXN) is a 48kDa protein and a redox-dependent negative regulator of the Wnt signaling pathway [63]. NXN mRNA is highly expressed in LECs (Table 1), and by immunohistology it can be detected in lymphatics of various organs (Figure 13). Similar to mutations in WNT5A [64] and other WNT signaling members, mutations in NXN cause the Robinow syndrome [65], which is mainly characterized by dysmorphic facial features and short-limbed dwarfism. To our knowledge, morphology and function of the lymphatics have not yet been investigated in Robinow patients.
3. Conclusion
The exome of lymphatic endothelial cells is very extensive, as in other cell types. The correspondence with the protein expression that we have studied in the Human Protein Atlas is comparatively low. However, this can probably be attributed to simple technical reasons when studying paraffin-embedded human tissues. We found good agreement for molecules associated with the high transcytosis activity of LECs (caveolin, dysferlin). There was very good agreement for molecules that have important functions in the structure and regulation of the cytoskeleton. This may be due to the fact that the LECs of the initial lymphatic vessels have a unique morphology and function. This is expressed in the oak leaf-like morphology of the cells with the formation of specialized button-like junctions and microvalves, which are of essential importance for the directed lymph flow [66,67,68]. Accordingly, we found expression of molecules that mediate cellular elasticity (spectrins) and interact with actin microfilaments (ANKRD37, CNN3, KANK3, MARCKSL1). In terms of immune regulation, lymphatic vessels are a double-edged sword [8]. They are the main route for the emigration of leukocytes towards the lymph nodes. However, they can also have an immunosuppressive effect, which is reflected in the expression of CD59. The influence of LECs on coagulation, which has already been described several times, was confirmed in our investigations (multimerin1). The importance of the WNT signaling pathway for development and function of lymphatics still needs to be investigated in greater detail (NXN). In sum, we hope our studies can stimulate further studies on the complex morphology and functions of lymphatics.
4. Materials and Methods
4.1. Cell Culture
4.2. RNA Sequencing
4.3. Immunofluorescence (IF)
IF was performed on human foreskin derived from operations performed at the University Medical Center Goettingen (UMG). Specimens were fixed in 4% paraformaldehyde for 1h, embedded in tissue freeze medium and sectioned at 12µm. Studies were performed with the informed consent of the patients or their parents, and were approved by the ethics committee of the UMG (application no. 18/1/18). Primary antibodies were: mouse-anti-human CD31(BD Pharmingen, dilution 1:50), rabbit-anti-human dysferlin (Sigma, dilution 1:200). Secondary antibodies were: Alexa 488-conjugated goat-anti-mouse IgG (H+L) (Invitrogen, dilution 1:200), and Alexa 594-conjugated goat-anti-rabbit IgG (H+L) (Invitrogen, dilution 1:200). Nuclei were counterstained with Dapi.
4.4. Immunohistochemistry
We compared our RNASeq expression data with protein expression by systematically studying the Human Protein Atlas [27] (https://www.proteinatlas.org/). All figures shown here can be found and further studied at variable magnification in this repository.
Author Contributions
J.B.: investigation, data curation; J.W.: conceptualization, data curation, writing - review and editing.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are included in the manuscript.
Acknowledgments
We are grateful to Ch. Zelent and B. Manshausen for their expert technical assistance. The studies were supported by the Verein zur Förderung der Lymphologie e.V., 79856 Hinterzarten, Germany. The grant supplier did not have any influence on the performance of the studies.
Conflicts of Interest Statement
The authors declare no competing interests.
Abbreviations
ANKRD37 - ankyrin repeat domain protein 37; BEC - Blood vascular endothelial cell; CAV1/2 – Caveolin 1/2; CD59 - CD59 glycoprotein aka Protectin; CNN3 - Calponin 3; DYSF – Dysferlin; GHS - Gorham-Stout disease/syndrome; KANK3 - KN motif- and ankyrin repeat domain-containing protein 3; LEC - Lymphatic endothelial cell; MAC - Membrane attack complex; MARCKSL1 - MARCKS-like protein-1; MMRN1 - Multimerin 1; MYOF – Myoferlin; NXN – Nucleoredoxin; OTOF – Otoferlin; SPTAN1 - Spectrin, alpha, nonerythrocytic 1; SPTBN1 - Spectrin, beta, nonerythrocytic 1; WNT - Wingless-type MMTV integration site.
References
- Rusznyák, I.; Földi, M.; Szabó, G. Lymphologie. Physiologie und Pathologie der Lymphgefäße und des Lymphkreislaufes; Akadémiai Kiadó, 1969;
- Asellius, G. De lactibus, sive lacteis venis, quarto vasorum mesaraicorum genere, novo invento Gasparis Asellii Cremo. Dissertatio; ex officina Iohannis Maire 1640, Milan 1628;
- Mascagni, P.; Sanctius, C. Vasorum Lymphaticorum Corporis Humani Historia et Ichnographia; Ex typographia Pazzini Carli, 1787;
- Rudbeck, O. Nova exercitatio anatomica, exhibens ductos hepaticos aquosos, et vasa glandularum serosa (1653); Almquist and Wiksells: Uppsala, 1930;
- Witte, M.H.; Jones, K.; Wilting, J.; Dictor, M.; Selg, M.; McHale, N.; Gershenwald, J.E.; Jackson, D.G. Structure Function Relationships in the Lymphatic System and Implications for Cancer Biology. Cancer Metastasis Rev. 2006, 25, 159–184. [CrossRef]
- Oliver, G.; Kipnis, J.; Randolph, G.J.; Harvey, N.L. The Lymphatic Vasculature in the 21st Century: Novel Functional Roles in Homeostasis and Disease. Cell 2020, 182, 270–296. [CrossRef]
- Martin-Almedina, S.; Mortimer, P.S.; Ostergaard, P. Development and Physiological Functions of the Lymphatic System: Insights from Human Genetic Studies of Primary Lymphedema. Physiol. Rev. 2021, 101, 1809–1871. [CrossRef]
- Wilting, J.; Becker, J. The Lymphatic Vascular System: Much More than Just a Sewer. Cell Biosci. 2022, 12, 157. [CrossRef]
- Petrova, T.V.; Koh, G.Y. Organ-Specific Lymphatic Vasculature: From Development to Pathophysiology. J. Exp. Med. 2018, 215, 35–49. [CrossRef]
- Becker, J.; Schwoch, S.; Zelent, C.; Sitte, M.; Salinas, G.; Wilting, J. Transcriptome Analysis of Hypoxic Lymphatic Endothelial Cells Indicates Their Potential to Contribute to Extracellular Matrix Rearrangement. Cells 2021, 10, 1008. [CrossRef]
- Podgrabinska, S.; Braun, P.; Velasco, P.; Kloos, B.; Pepper, M.S.; Skobe, M. Molecular Characterization of Lymphatic Endothelial Cells. Proc. Natl. Acad. Sci. U. S. A. 2002, 99, 16069–16074. [CrossRef]
- Hasselhof, V.; Sperling, A.; Buttler, K.; Ströbel, P.; Becker, J.; Aung, T.; Felmerer, G.; Wilting, J. Morphological and Molecular Characterization of Human Dermal Lymphatic Collectors. PloS One 2016, 11, e0164964. [CrossRef]
- Fujimoto, N.; He, Y.; D’Addio, M.; Tacconi, C.; Detmar, M.; Dieterich, L.C. Single-Cell Mapping Reveals New Markers and Functions of Lymphatic Endothelial Cells in Lymph Nodes. PLOS Biol. 2020, 18, e3000704. [CrossRef]
- Park, S.M.; Angel, C.E.; McIntosh, J.D.; Mansell, C.J.; Chen, C.-J.J.; Cebon, J.; Dunbar, P.R. Mapping the Distinctive Populations of Lymphatic Endothelial Cells in Different Zones of Human Lymph Nodes. PloS One 2014, 9, e94781. [CrossRef]
- Wigle, J.T.; Oliver, G. Prox1 Function Is Required for the Development of the Murine Lymphatic System. Cell 1999, 98, 769–778. [CrossRef]
- Harvey, N.L.; Srinivasan, R.S.; Dillard, M.E.; Johnson, N.C.; Witte, M.H.; Boyd, K.; Sleeman, M.W.; Oliver, G. Lymphatic Vascular Defects Promoted by Prox1 Haploinsufficiency Cause Adult-Onset Obesity. Nat. Genet. 2005, 37, 1072–1081. [CrossRef]
- Wong, B.W.; Wang, X.; Zecchin, A.; Thienpont, B.; Cornelissen, I.; Kalucka, J.; García-Caballero, M.; Missiaen, R.; Huang, H.; Brüning, U.; et al. The Role of Fatty Acid β-Oxidation in Lymphangiogenesis. Nature 2017, 542, 49–54. [CrossRef]
- Wilting, J.; Papoutsi, M.; Christ, B.; Nicolaides, K.H.; von Kaisenberg, C.S.; Borges, J.; Stark, G.B.; Alitalo, K.; Tomarev, S.I.; Niemeyer, C.; et al. The Transcription Factor Prox1 Is a Marker for Lymphatic Endothelial Cells in Normal and Diseased Human Tissues. FASEB J. 2002, 16, 1271–1273. [CrossRef]
- Kaipainen, A.; Korhonen, J.; Mustonen, T.; van Hinsbergh, V.W.; Fang, G.H.; Dumont, D.; Breitman, M.; Alitalo, K. Expression of the Fms-like Tyrosine Kinase 4 Gene Becomes Restricted to Lymphatic Endothelium during Development. Proc. Natl. Acad. Sci. U. S. A. 1995, 92, 3566–3570. [CrossRef]
- Banerji, S.; Ni, J.; Wang, S.X.; Clasper, S.; Su, J.; Tammi, R.; Jones, M.; Jackson, D.G. LYVE-1, a New Homologue of the CD44 Glycoprotein, Is a Lymph-Specific Receptor for Hyaluronan. J. Cell Biol. 1999, 144, 789–801. [CrossRef]
- Breiteneder-Geleff, S.; Soleiman, A.; Kowalski, H.; Horvat, R.; Amann, G.; Kriehuber, E.; Diem, K.; Weninger, W.; Tschachler, E.; Alitalo, K.; et al. Angiosarcomas Express Mixed Endothelial Phenotypes of Blood and Lymphatic Capillaries: Podoplanin as a Specific Marker for Lymphatic Endothelium. Am. J. Pathol. 1999, 154, 385–394. [CrossRef]
- Mattonet, K.; Wilting, J.; Jeltsch, M. Genetic Causes of Primary Lymphedema [German: Die Genetischen Ursachen Des Primären Lymphödems]. In; 2015; pp. 210–229.
- Modaghegh, M.H.S.; Tanzadehpanah, H.; Kamyar, M.M.; Manoochehri, H.; Sheykhhasan, M.; Forouzanfar, F.; Mahmoudian, R.A.; Lotfian, E.; Mahaki, H. The Role of Key Biomarkers in Lymphatic Malformation: An Updated Review. J. Gene Med. 2024, 26, e3665. [CrossRef]
- Mäkinen, T.; Boon, L.M.; Vikkula, M.; Alitalo, K. Lymphatic Malformations: Genetics, Mechanisms and Therapeutic Strategies. Circ. Res. 2021, 129, 136–154. [CrossRef]
- Petkova, M.; Ferby, I.; Mäkinen, T. Lymphatic Malformations: Mechanistic Insights and Evolving Therapeutic Frontiers. J. Clin. Invest. 2024, 134, e172844. [CrossRef]
- Wilting, J.; Felmerer, G.; Becker, J. Control of the Extracellular Matrix by Hypoxic Lymphatic Endothelial Cells: Impact on the Progression of Lymphedema? Dev. Dyn. 2023, 252, 227–238. [CrossRef]
- Thul, P.J.; Lindskog, C. The Human Protein Atlas: A Spatial Map of the Human Proteome. Protein Sci. Publ. Protein Soc. 2018, 27, 233–244. [CrossRef]
- Casley-Smith, J.R.; Florey, H.W. The Structure of Normal Small Lymphatics. Q. J. Exp. Physiol. Cogn. Med. Sci. 1961, 46, 101–106. [CrossRef]
- Sabesin, S.M.; Frase, S. Electron Microscopic Studies of the Assembly, Intracellular Transport, and Secretion of Chylomicrons by Rat Intestine. J. Lipid Res. 1977, 18, 496–511.
- Zhang, F.; Zarkada, G.; Han, J.; Li, J.; Dubrac, A.; Ola, R.; Genet, G.; Boyé, K.; Michon, P.; Künzel, S.E.; et al. Lacteal Junction Zippering Protects against Diet-Induced Obesity. Science 2018, 361, 599–603. [CrossRef]
- Arif, S.; Richer, M.; Larochelle, S.; Moulin, V.J. Microvesicles Derived from Dermal Myofibroblasts Modify the Integrity of the Blood and Lymphatic Barriers Using Distinct Endocytosis Pathways. J. Extracell. Biol. 2024, 3, e151. [CrossRef]
- Bansal, D.; Miyake, K.; Vogel, S.S.; Groh, S.; Chen, C.-C.; Williamson, R.; McNeil, P.L.; Campbell, K.P. Defective Membrane Repair in Dysferlin-Deficient Muscular Dystrophy. Nature 2003, 423, 168–172. [CrossRef]
- McMahon, A.P.; Giebelhaus, D.H.; Champion, J.E.; Bailes, J.A.; Lacey, S.; Carritt, B.; Henchman, S.K.; Moon, R.T. cDNA Cloning, Sequencing and Chromosome Mapping of a Non-Erythroid Spectrin, Human Alpha-Fodrin. Differ. Res. Biol. Divers. 1987, 34, 68–78. [CrossRef]
- Chang, J.G.; Scarpa, A.; Eddy, R.L.; Byers, M.G.; Harris, A.S.; Morrow, J.S.; Watkins, P.; Shows, T.B.; Forget, B.G. Cloning of a Portion of the Chromosomal Gene and cDNA for Human Beta-Fodrin, the Nonerythroid Form of Beta-Spectrin. Genomics 1993, 17, 287–293. [CrossRef]
- Liem, R.K.H. Cytoskeletal Integrators: The Spectrin Superfamily. Cold Spring Harb. Perspect. Biol. 2016, 8, a018259. [CrossRef]
- Leak, L.V.; Liotta, L.A.; Krutzsch, H.; Jones, M.; Fusaro, V.A.; Ross, S.J.; Zhao, Y.; Petricoin, E.F. Proteomic Analysis of Lymph. Proteomics 2004, 4, 753–765. [CrossRef]
- Posner, M.G. Multimerin-1 and Cancer: A Review. Biosci. Rep. 2022, 42, BSR20211248. [CrossRef]
- Hayward, C.P.; Hassell, J.A.; Denomme, G.A.; Rachubinski, R.A.; Brown, C.; Kelton, J.G. The cDNA Sequence of Human Endothelial Cell Multimerin. A Unique Protein with RGDS, Coiled-Coil, and Epidermal Growth Factor-like Domains and a Carboxyl Terminus Similar to the Globular Domain of Complement C1q and Collagens Type VIII and X. J. Biol. Chem. 1995, 270, 18246–18251. [CrossRef]
- Ji, R.C.; Kato, S. Histochemical Analysis of Lymphatic Endothelial Cells in Lymphostasis. Microsc. Res. Tech. 2001, 55, 70–80. [CrossRef]
- Umekage, T.; Kato, K. A Mouse Brain cDNA Encodes a Novel Protein with the Protein Kinase C Phosphorylation Site Domain Common to MARCKS. FEBS Lett. 1991, 286, 147–151. [CrossRef]
- Stumpo, D.J.; Eddy, R.L.; Haley, L.L.; Sait, S.; Shows, T.B.; Lai, W.S.; Young, W.S.; Speer, M.C.; Dehejia, A.; Polymeropoulos, M.; et al. Promoter Sequence, Expression, and Fine Chromosomal Mapping of the Human Gene (MLP) Encoding the MARCKS-like Protein: Identification of Neighboring and Linked Polymorphic Loci for MLP and MACS and Use in the Evaluation of Human Neural Tube Defects. Genomics 1998, 49, 253–264. [CrossRef]
- Björkblom, B.; Padzik, A.; Mohammad, H.; Westerlund, N.; Komulainen, E.; Hollos, P.; Parviainen, L.; Papageorgiou, A.C.; Iljin, K.; Kallioniemi, O.; et al. C-Jun N-Terminal Kinase Phosphorylation of MARCKSL1 Determines Actin Stability and Migration in Neurons and in Cancer Cells. Mol. Cell. Biol. 2012, 32, 3513–3526. [CrossRef]
- Jonsdottir, K.; Zhang, H.; Jhagroe, D.; Skaland, I.; Slewa, A.; Björkblom, B.; Coffey, E.T.; Gudlaugsson, E.; Smaaland, R.; Janssen, E.A.M.; et al. The Prognostic Value of MARCKS-like 1 in Lymph Node-Negative Breast Cancer. Breast Cancer Res. Treat. 2012, 135, 381–390. [CrossRef]
- Maguchi, M.; Nishida, W.; Kohara, K.; Kuwano, A.; Kondo, I.; Hiwada, K. Molecular Cloning and Gene Mapping of Human Basic and Acidic Calponins. Biochem. Biophys. Res. Commun. 1995, 217, 238–244. [CrossRef]
- Liu, R.; Jin, J.-P. Calponin Isoforms CNN1, CNN2 and CNN3: Regulators for Actin Cytoskeleton Functions in Smooth Muscle and Non-Muscle Cells. Gene 2016, 585, 143–153. [CrossRef]
- Ciuba, K.; Hawkes, W.; Tojkander, S.; Kogan, K.; Engel, U.; Iskratsch, T.; Lappalainen, P. Calponin-3 Is Critical for Coordinated Contractility of Actin Stress Fibers. Sci. Rep. 2018, 8, 17670. [CrossRef]
- Maddala, R.; Mongan, M.; Xia, Y.; Rao, P.V. Calponin-3 Deficiency Augments Contractile Activity, Plasticity, Fibrogenic Response and Yap/Taz Transcriptional Activation in Lens Epithelial Cells and Explants. Sci. Rep. 2020, 10, 1295. [CrossRef]
- Cho, H.; Kim, J.; Ahn, J.H.; Hong, Y.-K.; Mäkinen, T.; Lim, D.-S.; Koh, G.Y. YAP and TAZ Negatively Regulate Prox1 During Developmental and Pathologic Lymphangiogenesis. Circ. Res. 2019, 124, 225–242. [CrossRef]
- Zhong, W.; Jiang, H.; Zou, Y.; Ren, J.; Li, Z.; He, K.; Zhao, J.; Zhou, X.; Mou, D.; Cai, Y. The YAP Signaling Pathway Promotes the Progression of Lymphatic Malformations through the Activation of Lymphatic Endothelial Cells. Pediatr. Res. 2021, 89, 110–117.
- Lee, S.; Finn, L.; Sze, R.W.; Perkins, J.A.; Sie, K.C. Gorham Stout Syndrome (Disappearing Bone Disease): Two Additional Case Reports and a Review of the Literature. Arch. Otolaryngol. Head Neck Surg. 2003, 129, 1340–1343. [CrossRef]
- Leite, I.; Hernández-Martín, A.; Colmenero, I.; López-Gutiérrez, J.C.; Torrelo, A. Invasive Lymphatic Malformation (Gorham-Stout) of the Pelvis with Prominent Skin Involvement. Pediatr. Dermatol. 2013, 30, 374–378. [CrossRef]
- Ghaffarpour, N.; Baselga, E.; Boon, L.M.; Diociaiuti, A.; Dompmartin, A.; Dvorakova, V.; El Hachem, M.; Gasparella, P.; Haxhija, E.; Kyrklund, K.; et al. The VASCERN-VASCA Working Group Diagnostic and Management Pathways for Lymphatic Malformations. Eur. J. Med. Genet. 2022, 65, 104637. [CrossRef]
- Tan, W.; Fu, H.; Zhou, X.; Duan, Y.; Yin, N.; Huang, J.; Liu, X. ANKRD37 Inhibits Trophoblast Migration and Invasion by Regulating the NF-κB Pathway in Preeclampsia. J. Gene Med. 2022, 24, e3416. [CrossRef]
- Benita, Y.; Kikuchi, H.; Smith, A.D.; Zhang, M.Q.; Chung, D.C.; Xavier, R.J. An Integrative Genomics Approach Identifies Hypoxia Inducible Factor-1 (HIF-1)-Target Genes That Form the Core Response to Hypoxia. Nucleic Acids Res. 2009, 37, 4587–4602. [CrossRef]
- Zhu, Y.; Kakinuma, N.; Wang, Y.; Kiyama, R. Kank Proteins: A New Family of Ankyrin-Repeat Domain-Containing Proteins. Biochim. Biophys. Acta 2008, 1780, 128–133. [CrossRef]
- Guo, S.S.; Seiwert, A.; Szeto, I.Y.Y.; Fässler, R. Tissue Distribution and Subcellular Localization of the Family of Kidney Ankyrin Repeat Domain (KANK) Proteins. Exp. Cell Res. 2021, 398, 112391. [CrossRef]
- Janeway, C.; Travers, P.; Walport, M.; Shlomchik, M. Immunobiology: The Immune System in Health and Disease; Garland Pub. New York, 2001; Vol. 2;.
- Okada, N.; Harada, R.; Fujita, T.; Okada, H. A Novel Membrane Glycoprotein Capable of Inhibiting Membrane Attack by Homologous Complement. Int. Immunol. 1989, 1, 205–208. [CrossRef]
- Nevo, Y.; Ben-Zeev, B.; Tabib, A.; Straussberg, R.; Anikster, Y.; Shorer, Z.; Fattal-Valevski, A.; Ta-Shma, A.; Aharoni, S.; Rabie, M.; et al. CD59 Deficiency Is Associated with Chronic Hemolysis and Childhood Relapsing Immune-Mediated Polyneuropathy. Blood 2013, 121, 129–135. [CrossRef]
- Ma, Y.; Zhang, K.; Wu, Y.; Fu, X.; Liang, S.; Peng, M.; Guo, J.; Liu, M. Revisiting the Relationship between Complement and Ulcerative Colitis. Scand. J. Immunol. 2023, 98, e13329. [CrossRef]
- Dejana, E. The Role of Wnt Signaling in Physiological and Pathological Angiogenesis. Circ. Res. 2010, 107, 943–952. [CrossRef]
- Lutze, G.; Haarmann, A.; Demanou Toukam, J.A.; Buttler, K.; Wilting, J.; Becker, J. Non-Canonical WNT-Signaling Controls Differentiation of Lymphatics and Extension Lymphangiogenesis via RAC and JNK Signaling. Sci. Rep. 2019, 9, 4739. [CrossRef]
- Kurooka, H.; Kato, K.; Minoguchi, S.; Takahashi, Y.; Ikeda, J.; Habu, S.; Osawa, N.; Buchberg, A.M.; Moriwaki, K.; Shisa, H.; et al. Cloning and Characterization of the Nucleoredoxin Gene That Encodes a Novel Nuclear Protein Related to Thioredoxin. Genomics 1997, 39, 331–339. [CrossRef]
- Person, A.D.; Beiraghi, S.; Sieben, C.M.; Hermanson, S.; Neumann, A.N.; Robu, M.E.; Schleiffarth, J.R.; Billington, C.J.; van Bokhoven, H.; Hoogeboom, J.M.; et al. WNT5A Mutations in Patients with Autosomal Dominant Robinow Syndrome. Dev. Dyn. 2010, 239, 327–337. [CrossRef]
- White, J.J.; Mazzeu, J.F.; Coban-Akdemir, Z.; Bayram, Y.; Bahrambeigi, V.; Hoischen, A.; van Bon, B.W.M.; Gezdirici, A.; Gulec, E.Y.; Ramond, F.; et al. WNT Signaling Perturbations Underlie the Genetic Heterogeneity of Robinow Syndrome. Am. J. Hum. Genet. 2018, 102, 27–43. [CrossRef]
- von Recklinghausen, F. Die Lymphgefässe und ihre Beziehung zum Bindegewebe; A. Hirschwald, 1862;
- Trzewik, J.; Mallipattu, S.K.; Artmann, G.M.; Delano, F.A.; Schmid-Schonbein, G.W. Evidence for a Second Valve System in Lymphatics: Endothelial Microvalves. FASEB J. 2001, 15, 1711–1717. [CrossRef]
- Baluk, P.; Fuxe, J.; Hashizume, H.; Romano, T.; Lashnits, E.; Butz, S.; Vestweber, D.; Corada, M.; Molendini, C.; Dejana, E. Functionally Specialized Junctions between Endothelial Cells of Lymphatic Vessels. J. Exp. Med. 2007, 204, 2349–2362.
Figure 1.
Immunostaining of CAV1 in lymphatics (arrows) of human A) oral mucosa, Antibody CAB003791 B) rectum, Antibody CAB003791 C) Breast, Antibody CAB003791 and D) parietal layer of lymph node marginal sinus, Antibody HPA049326. From: The Human Protein Atlas.
Figure 1.
Immunostaining of CAV1 in lymphatics (arrows) of human A) oral mucosa, Antibody CAB003791 B) rectum, Antibody CAB003791 C) Breast, Antibody CAB003791 and D) parietal layer of lymph node marginal sinus, Antibody HPA049326. From: The Human Protein Atlas.
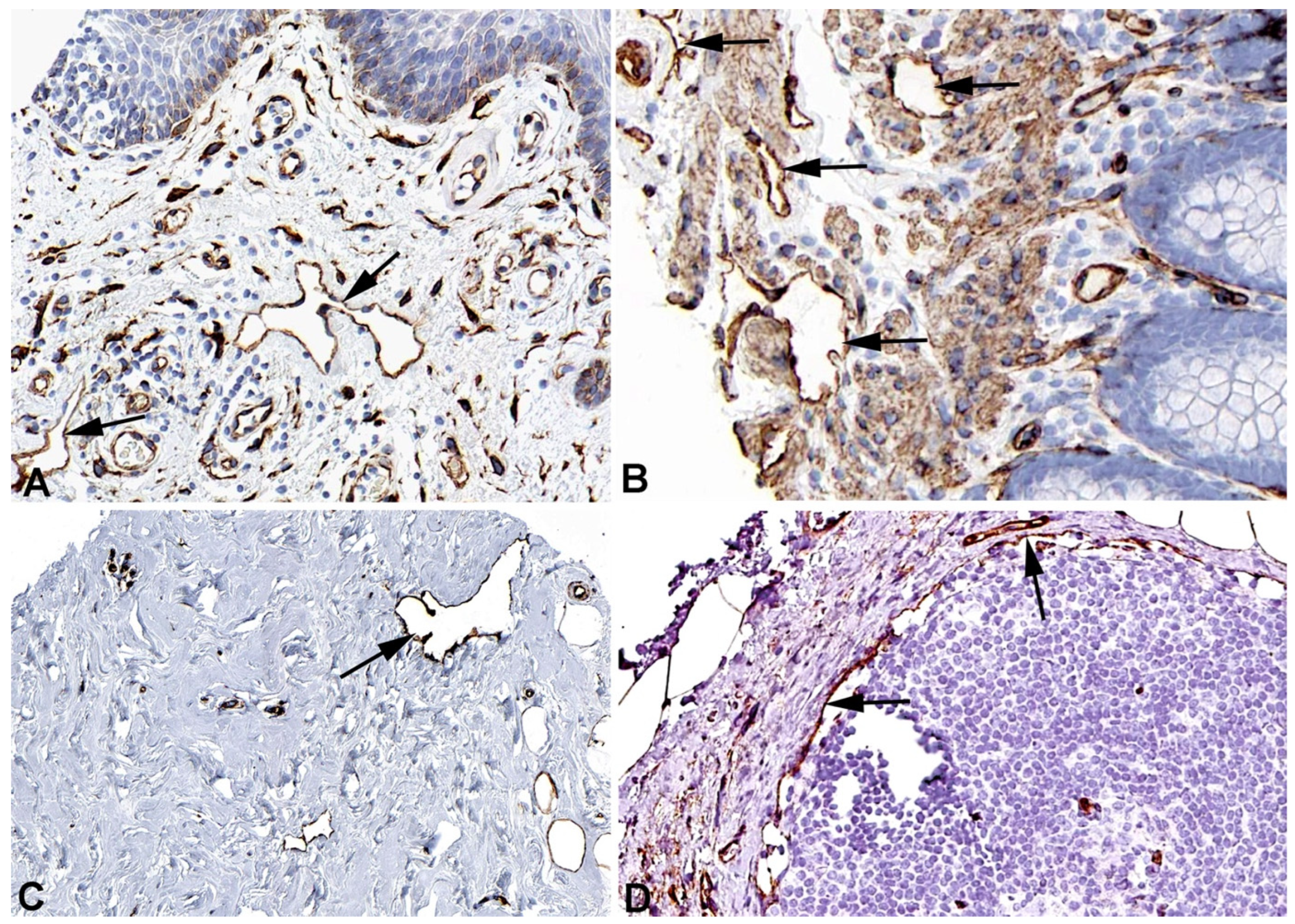
Figure 2.
Immunostaining of CAV2 in lymphatics (arrows) of human A) Esophagus, Antibody HPA044810 B) Colon, Antibody CAB013488, C) Oral mucosa, Antibody HPA044810 and D) Skin, Antibody HPA044810. From: The Human Protein Atlas.
Figure 2.
Immunostaining of CAV2 in lymphatics (arrows) of human A) Esophagus, Antibody HPA044810 B) Colon, Antibody CAB013488, C) Oral mucosa, Antibody HPA044810 and D) Skin, Antibody HPA044810. From: The Human Protein Atlas.
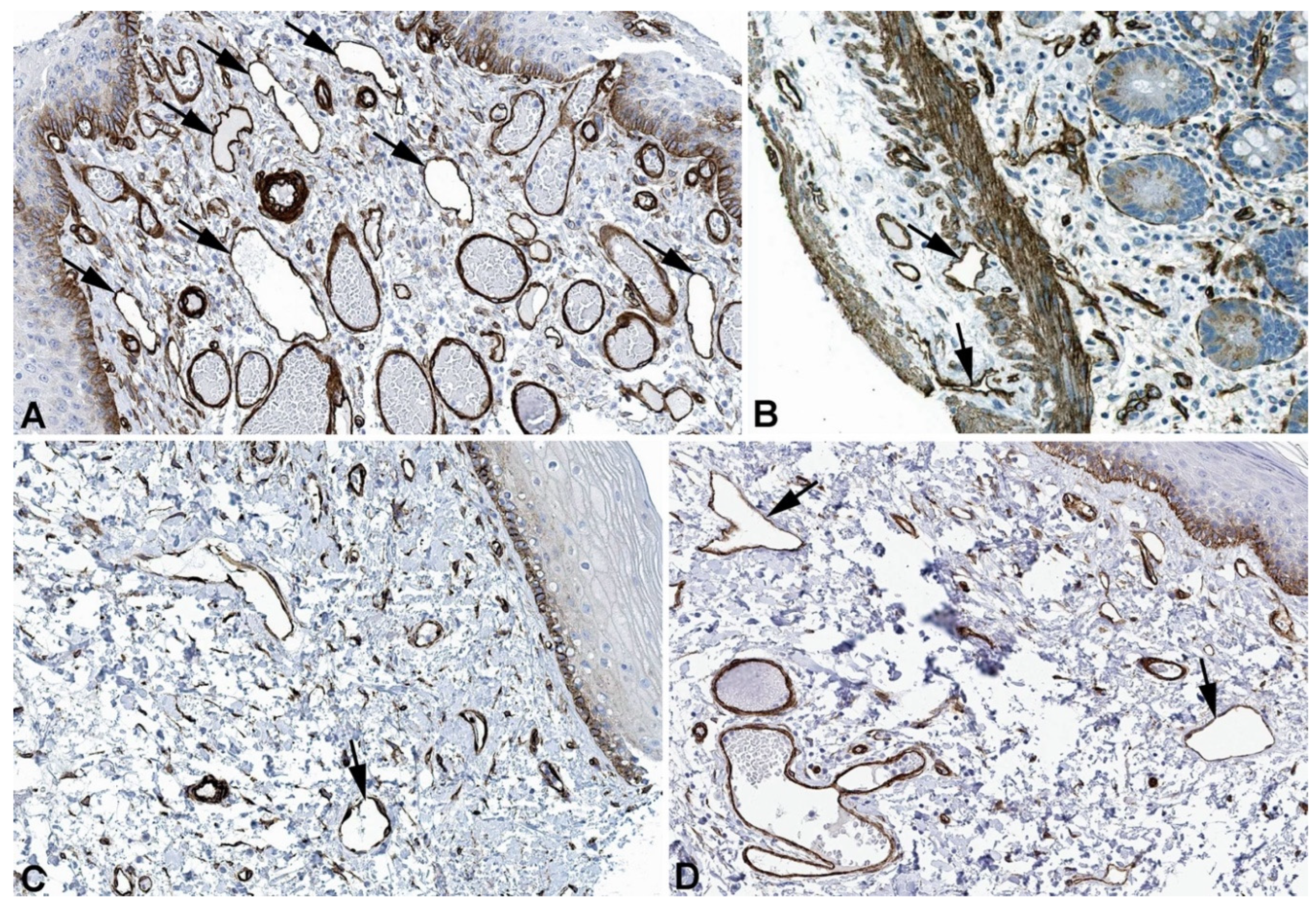
Figure 3.
Immunostaining of DYSF in lymphatics (arrows) of human A) Breast, Antibody CAB002510, B) Duodenum, Antibody CAB002510, C) Colon, Antibody HPA017071, D) Skin anal, Antibody CAB002510. From: The Human Protein Atlas.
Figure 3.
Immunostaining of DYSF in lymphatics (arrows) of human A) Breast, Antibody CAB002510, B) Duodenum, Antibody CAB002510, C) Colon, Antibody HPA017071, D) Skin anal, Antibody CAB002510. From: The Human Protein Atlas.
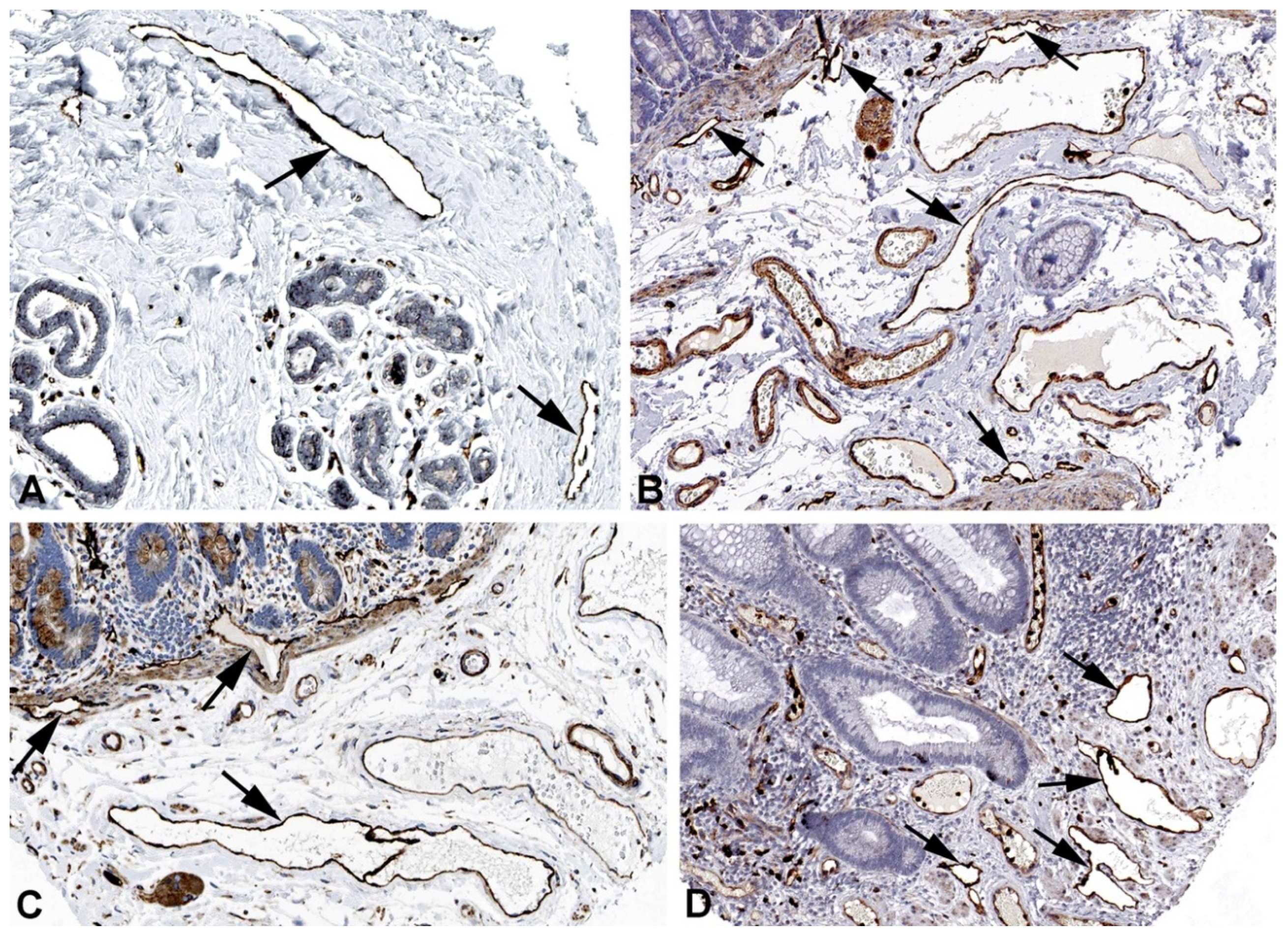
Figure 4.
Immunostaining of DYSF (red) and CD31 (green) in lymphatics of human foreskin. Blood vessels strongly express CD31; lymphatics have a punctate weak staining. Nuclei are stained blue with Dapi. In lymphatics the red DYSF staining is dominant, but subepithelial capillaries also express DYSF. A-C) 10x objective; Bar = 100 µm, D) 40x objective; Bar = 25µm.
Figure 4.
Immunostaining of DYSF (red) and CD31 (green) in lymphatics of human foreskin. Blood vessels strongly express CD31; lymphatics have a punctate weak staining. Nuclei are stained blue with Dapi. In lymphatics the red DYSF staining is dominant, but subepithelial capillaries also express DYSF. A-C) 10x objective; Bar = 100 µm, D) 40x objective; Bar = 25µm.
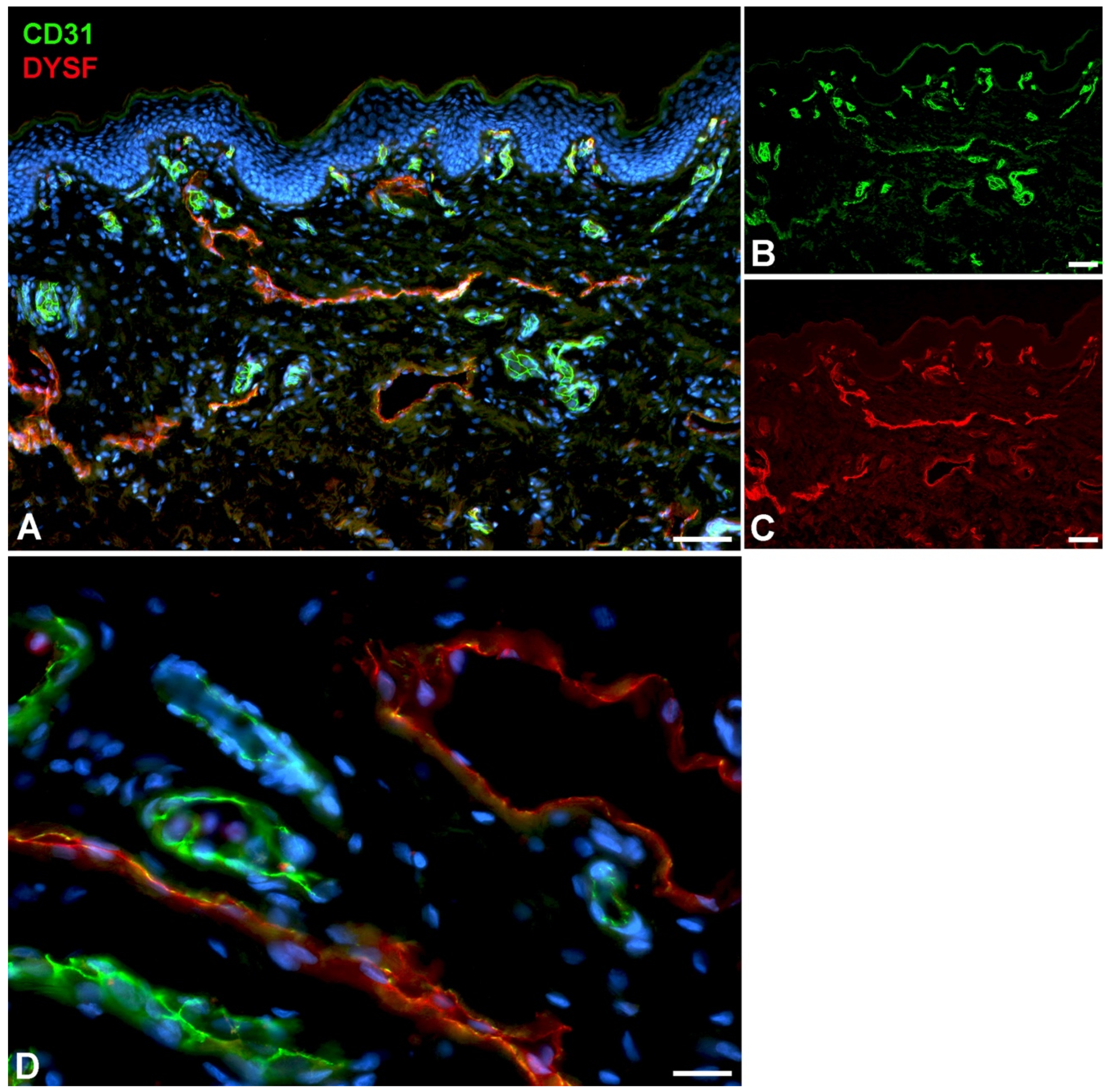
Figure 5.
Immunostaining of SPTAN1 in lymphatics (arrows) of human A) Oral mucosa, Antibody HPA007927, B) Small intestine. Antibody CAB004581, C) Breast Antibody HPA007927, D) Epididymis, Antibody HPA007927. From: The Human Protein Atlas.
Figure 5.
Immunostaining of SPTAN1 in lymphatics (arrows) of human A) Oral mucosa, Antibody HPA007927, B) Small intestine. Antibody CAB004581, C) Breast Antibody HPA007927, D) Epididymis, Antibody HPA007927. From: The Human Protein Atlas.
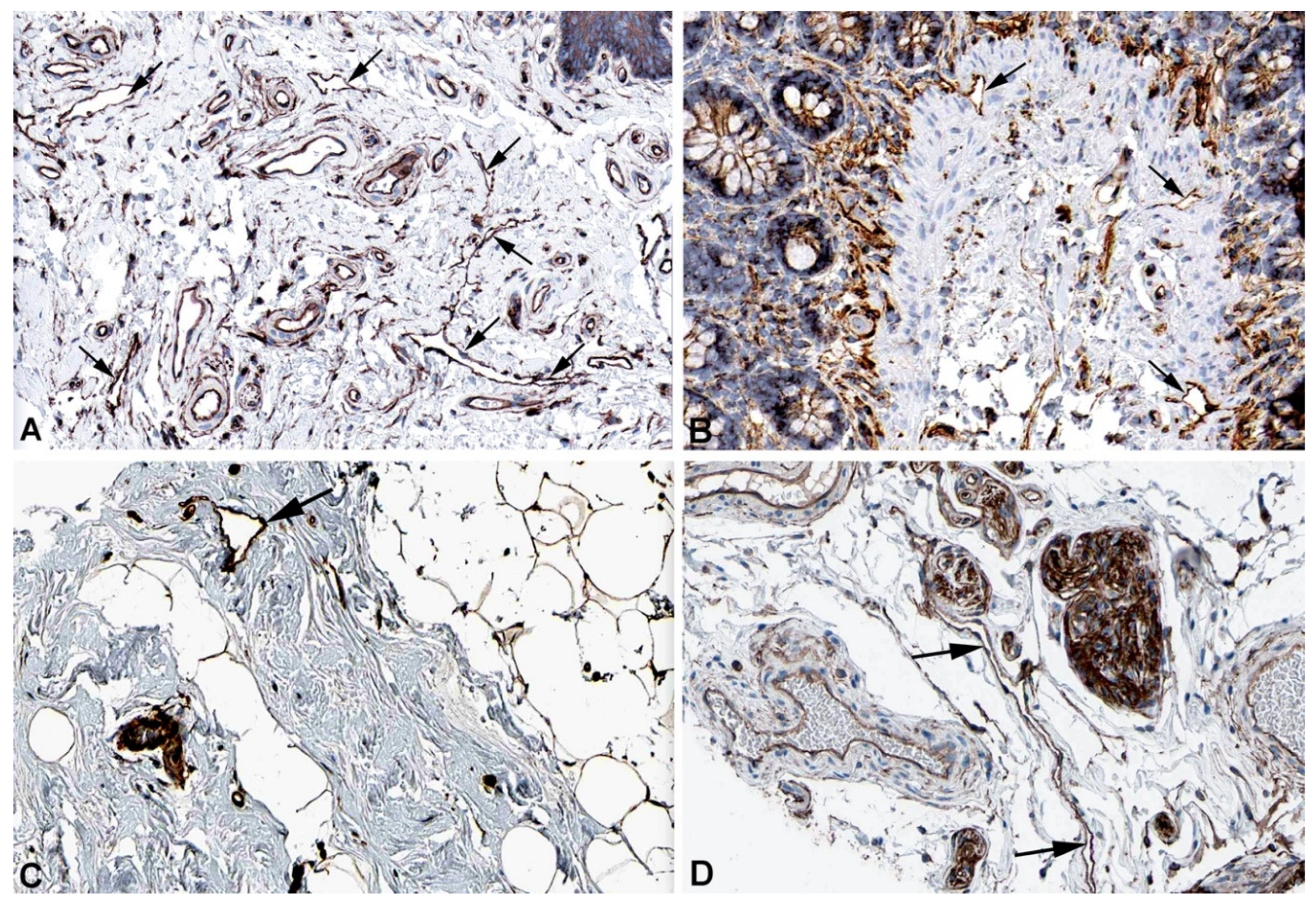
Figure 6.
Immunostaining of SPTBN1 in lymphatics (arrows) of human A) Esophagus, Antibody HPA013149, B) Colon, Antibody HPA013149, C) Breast, Antibody HPA013149. From: The Human Protein Atlas.
Figure 6.
Immunostaining of SPTBN1 in lymphatics (arrows) of human A) Esophagus, Antibody HPA013149, B) Colon, Antibody HPA013149, C) Breast, Antibody HPA013149. From: The Human Protein Atlas.
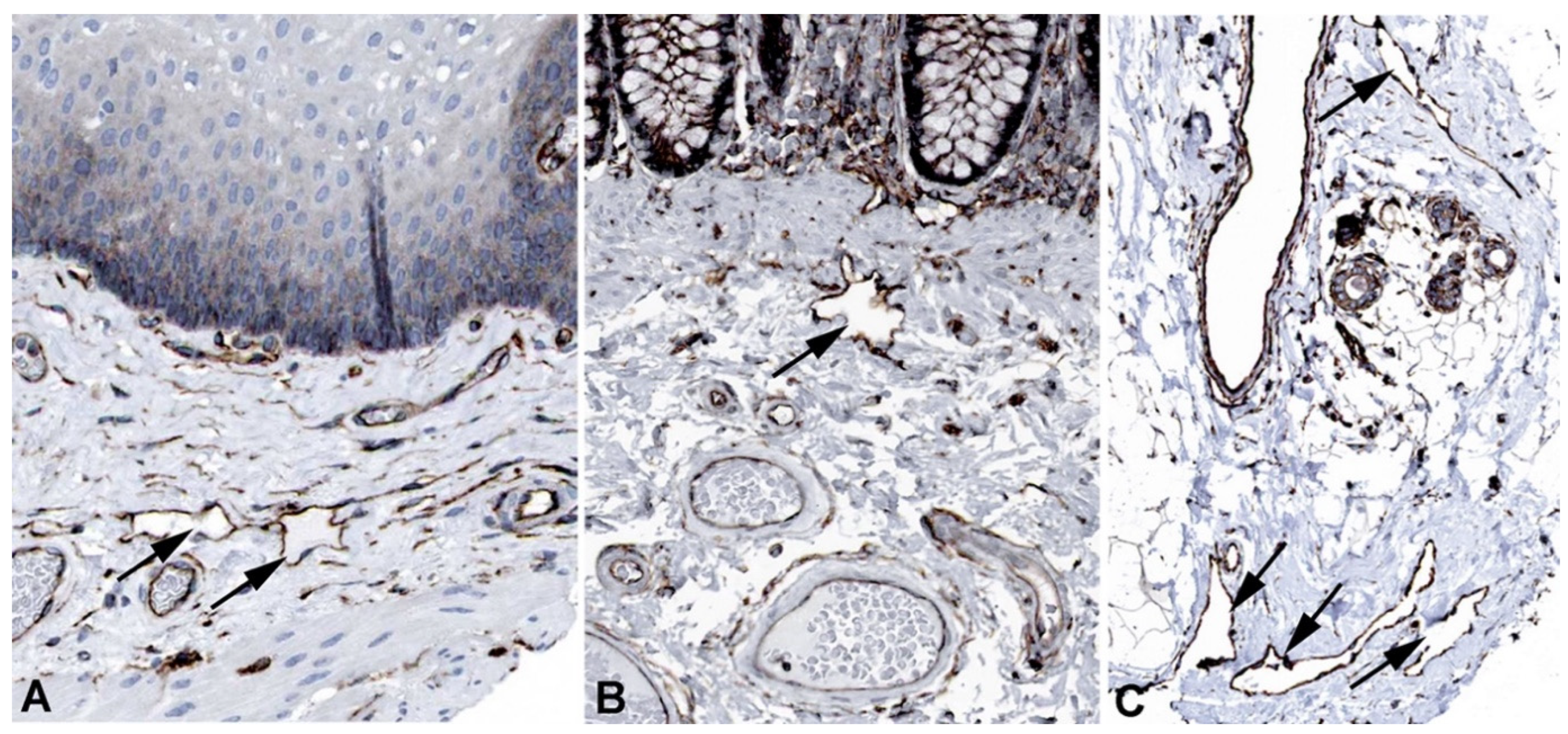
Figure 7.
Immunostaining of MMRN1 in lymphatics (arrows) of human A) Skin, Antibody HPA035769, B) Oral mucosa, Antibody HPA035769, C) Urinary bladder, Antibody HPA035769. From: The Human Protein Atlas.
Figure 7.
Immunostaining of MMRN1 in lymphatics (arrows) of human A) Skin, Antibody HPA035769, B) Oral mucosa, Antibody HPA035769, C) Urinary bladder, Antibody HPA035769. From: The Human Protein Atlas.

Figure 8.
Immunostaining of MARCKSL1 in lymphatics (arrows) of human A) Oral mucosa, Antibody HPA030528, B) Esophagus, Antibody HPA030528, C) Duodenum, Antibody HPA030528, D) Cervix uteri, Antibody HPA030528. From: The Human Protein Atlas.
Figure 8.
Immunostaining of MARCKSL1 in lymphatics (arrows) of human A) Oral mucosa, Antibody HPA030528, B) Esophagus, Antibody HPA030528, C) Duodenum, Antibody HPA030528, D) Cervix uteri, Antibody HPA030528. From: The Human Protein Atlas.
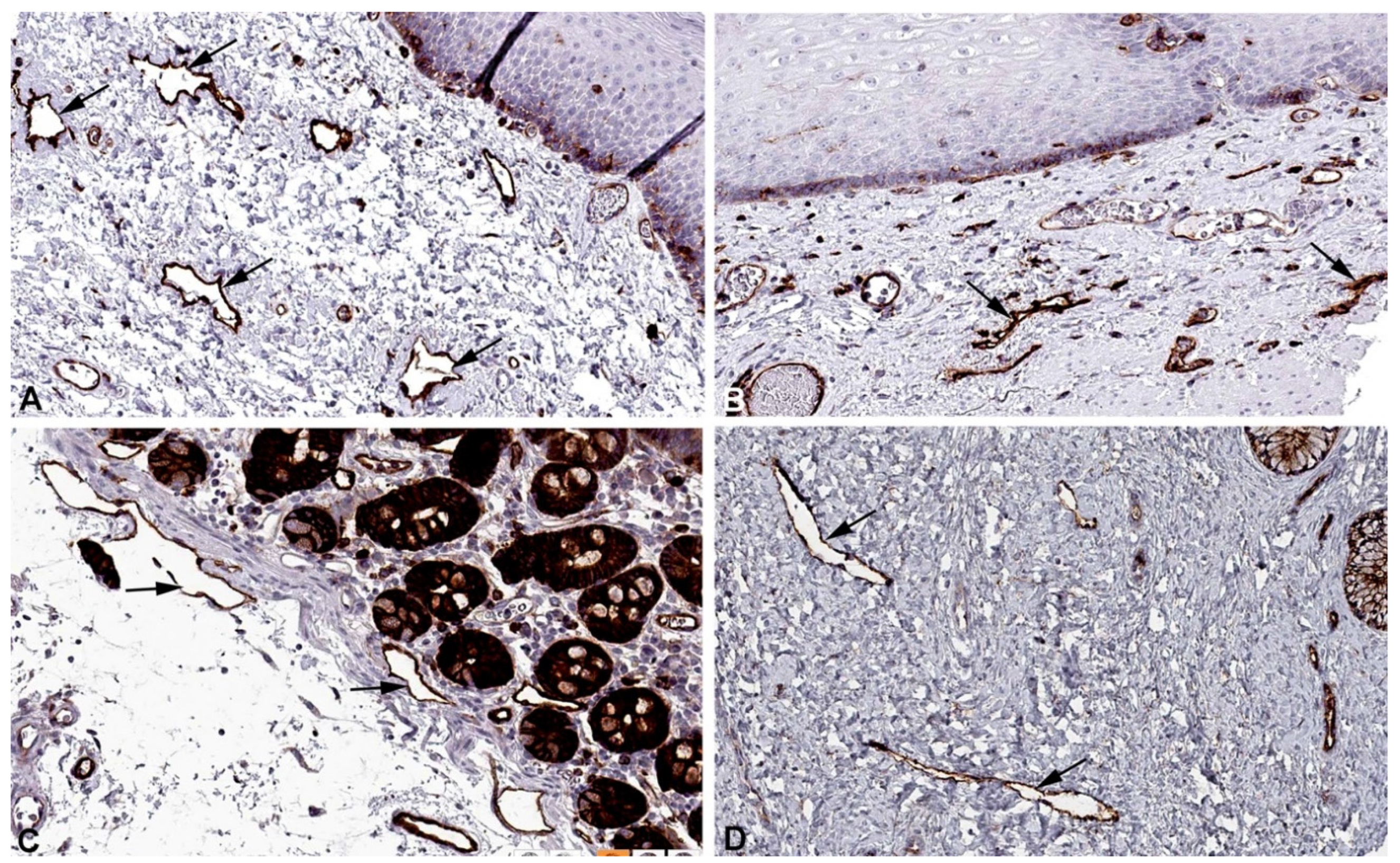
Figure 9.
Immunostaining of CNN3 in lymphatics (arrows) of human A) Oral mucosa, Antibody HPA051237, B) Esophagus, Antibody CAB009849, C) Skin, Antibody CAB009849, D) Colon, Antibody CAB009849. From: The Human Protein Atlas.
Figure 9.
Immunostaining of CNN3 in lymphatics (arrows) of human A) Oral mucosa, Antibody HPA051237, B) Esophagus, Antibody CAB009849, C) Skin, Antibody CAB009849, D) Colon, Antibody CAB009849. From: The Human Protein Atlas.
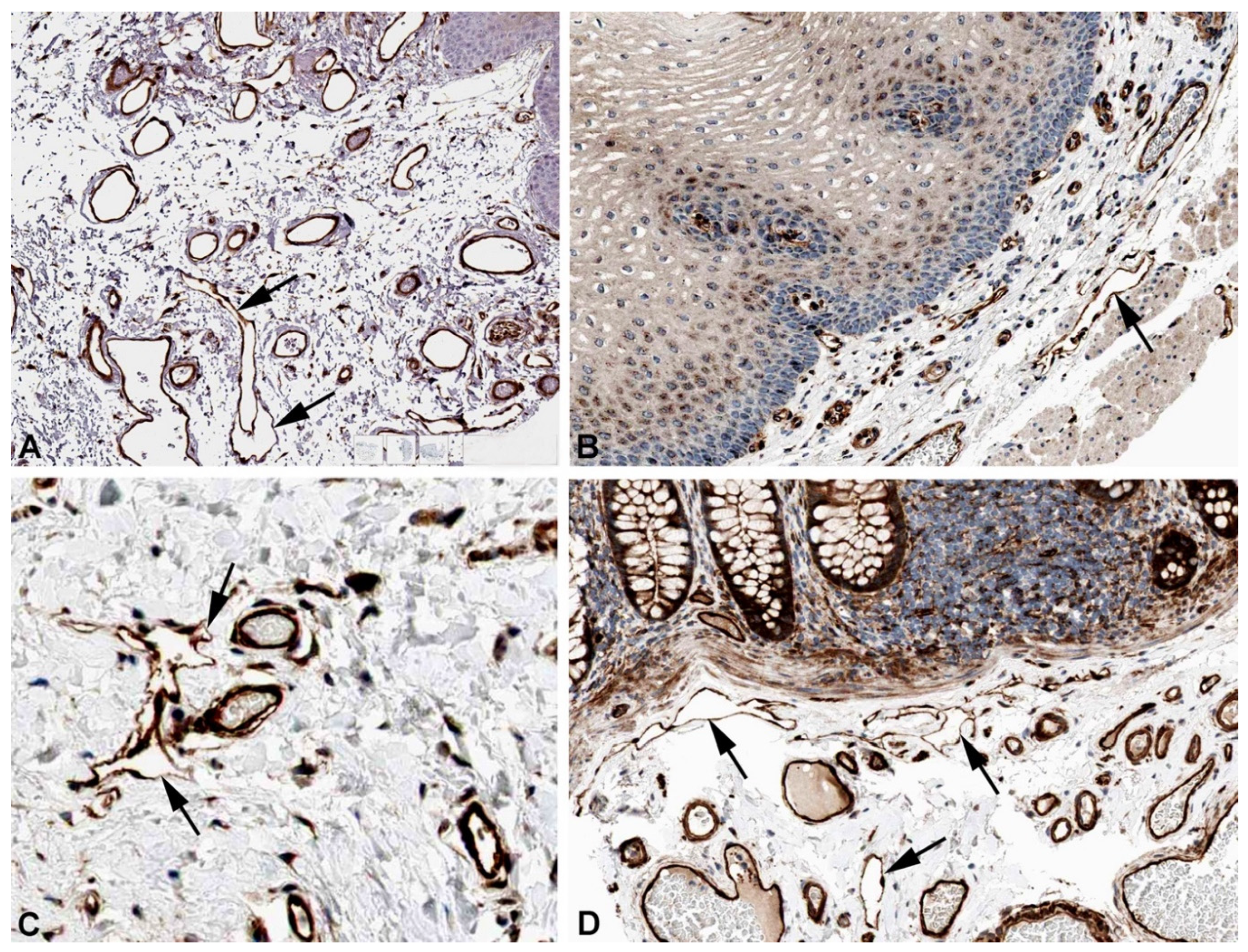
Figure 10.
Immunostaining of ANKRD37 in lymphatics (arrows) of human A) Breast, Antibody HPA036626, B) Esophagus, Antibody HPA036626, C) Colon, Antibody HPA036626, D) Lymph node, Antibody HPA036626. From: The Human Protein Atlas.
Figure 10.
Immunostaining of ANKRD37 in lymphatics (arrows) of human A) Breast, Antibody HPA036626, B) Esophagus, Antibody HPA036626, C) Colon, Antibody HPA036626, D) Lymph node, Antibody HPA036626. From: The Human Protein Atlas.
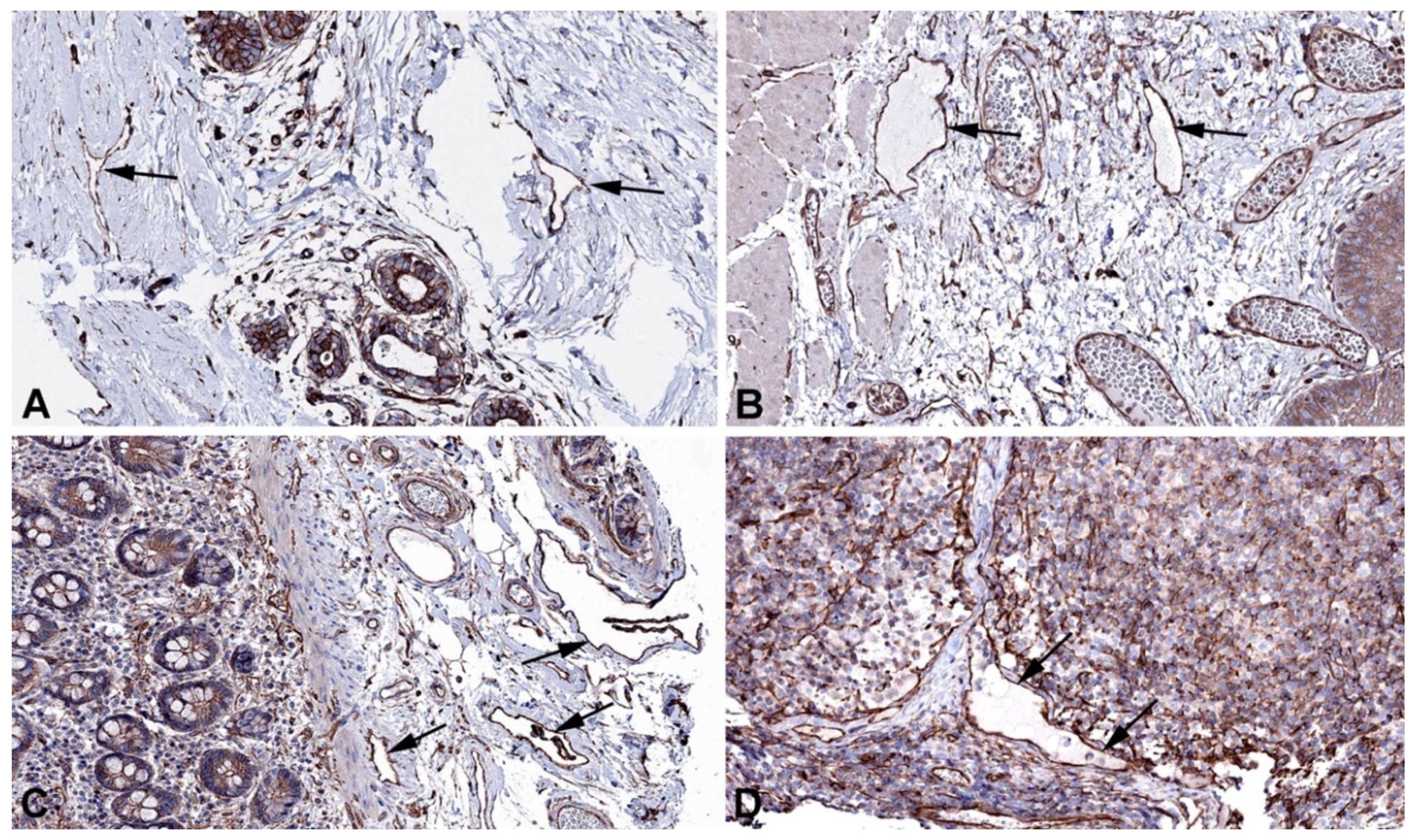
Figure 11.
Immunostaining of KANK3 in lymphatics (arrows) of human A) Skin, (obviously edematous), Antibody HPA051153, B) Colon, Antibody HPA051153, C) Rectum, Antibody HPA051153, D) Lymph node, Antibody HPA051153. From: The Human Protein Atlas.
Figure 11.
Immunostaining of KANK3 in lymphatics (arrows) of human A) Skin, (obviously edematous), Antibody HPA051153, B) Colon, Antibody HPA051153, C) Rectum, Antibody HPA051153, D) Lymph node, Antibody HPA051153. From: The Human Protein Atlas.
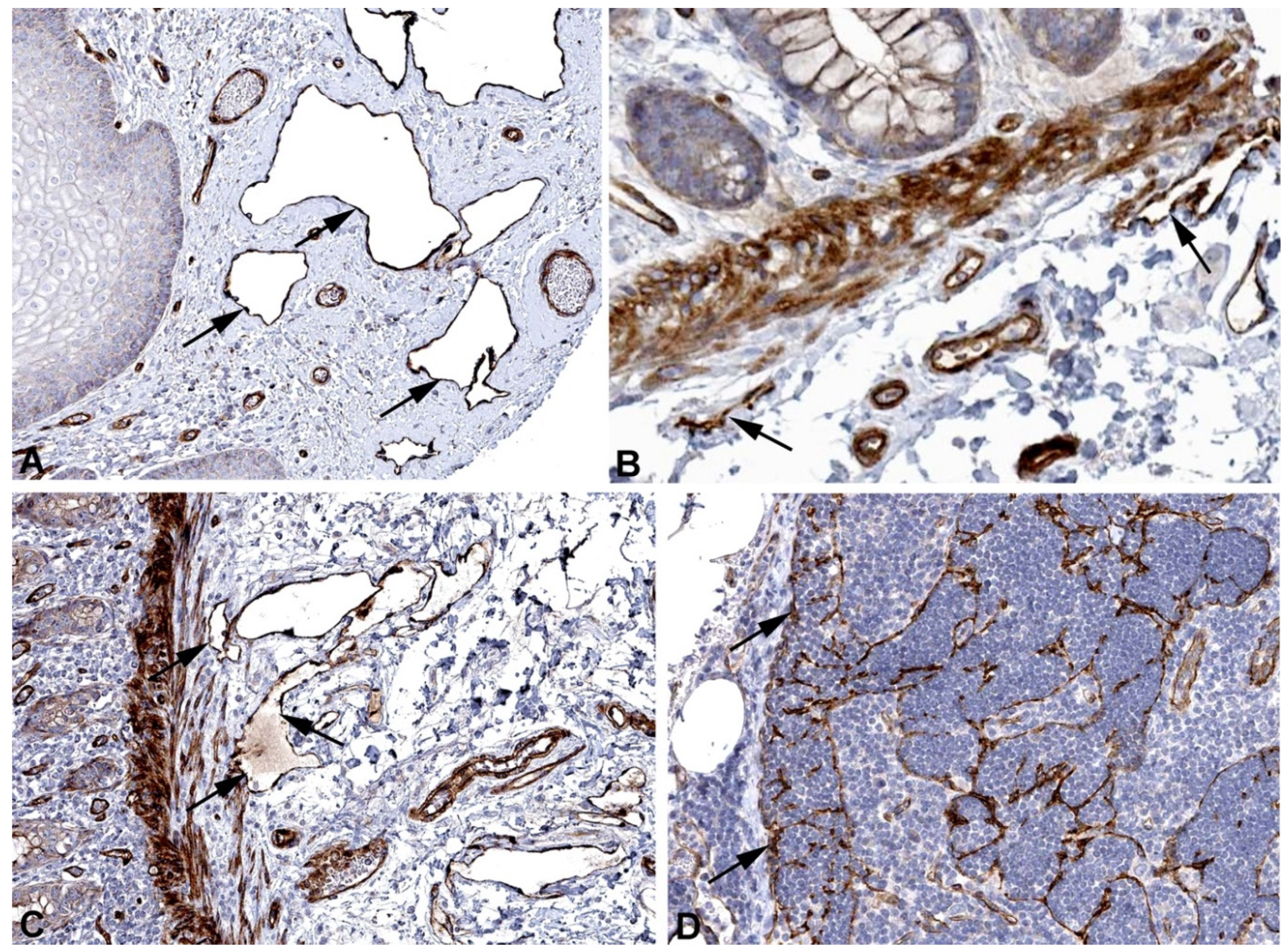
Figure 12.
Immunostaining of CD59 in lymphatics (arrows) of human A) Skin, Antibody HPA026494, B) Breast, Antibody HPA0264949, C) Oral mucosa, Antibody HPA026494, D) Colon, Antibody HPA026494. From: The Human Protein Atlas.
Figure 12.
Immunostaining of CD59 in lymphatics (arrows) of human A) Skin, Antibody HPA026494, B) Breast, Antibody HPA0264949, C) Oral mucosa, Antibody HPA026494, D) Colon, Antibody HPA026494. From: The Human Protein Atlas.
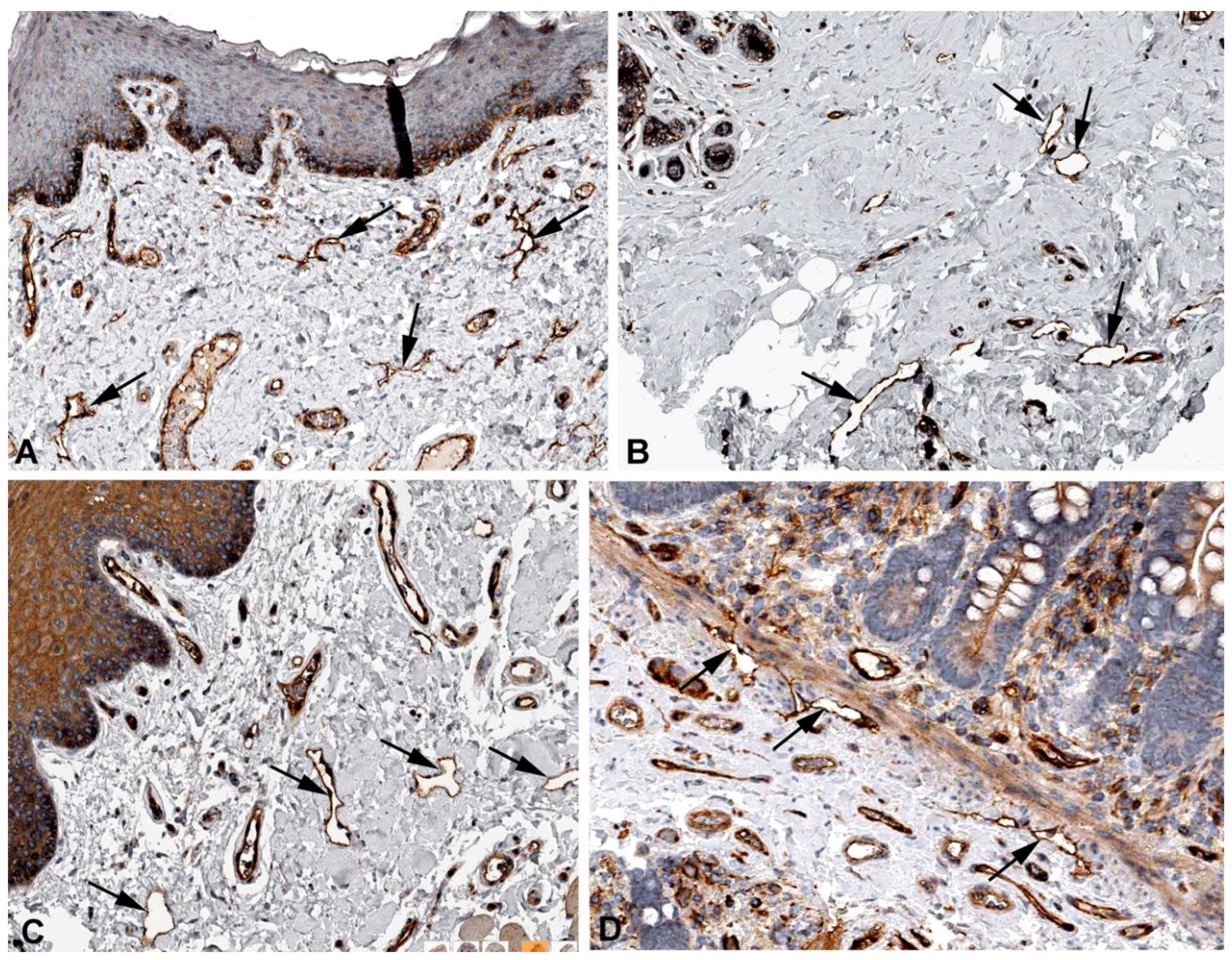
Figure 13.
Immunostaining of NXN in lymphatics (arrows) of human A) Skin, Antibody HPA023566, B) Breast, Antibody HPA023566, C) Duodenum, Antibody HPA023566, D) Colon, Antibody HPA023566. From: The Human Protein Atlas.
Figure 13.
Immunostaining of NXN in lymphatics (arrows) of human A) Skin, Antibody HPA023566, B) Breast, Antibody HPA023566, C) Duodenum, Antibody HPA023566, D) Colon, Antibody HPA023566. From: The Human Protein Atlas.

Table 1.
RNASeq analysis of 3 defined human dermal LEC lines under normoxia and hypoxia.
| gene_id | gene_name | chromos. | start | end | width | strand | gene_biotype | HDLEC-5 | HDLEC-6 | HDLEC-7 | Hypox-5 | Hypox-6 | Hypox-7 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ENSG00000186352 | ANKRD37 | 4 | 1.85E+08 | 1.85E+08 | 4608 | + | protein_cod. | 92.68 | 117.12 | 124.35 | 312.56 | 549.57 | 650.6 |
| ENSG00000105974 | CAV1 | 7 | 1.17E+08 | 1.17E+08 | 36184 | + | protein_cod. | 28254.02 | 22816.55 | 40041.39 | 41886.36 | 29070.25 | 44229.96 |
| ENSG00000105971 | CAV2 | 7 | 1.16E+08 | 1.17E+08 | 221162 | + | protein_cod. | 5923.02 | 6362.42 | 8338.77 | 8258.02 | 7425.45 | 10627.94 |
| ENSG00000085063 | CD59 | 11 | 33703010 | 33736491 | 33482 | - | protein_cod. | 25996.8 | 30542.27 | 33419.64 | 25877.99 | 36425.42 | 34222.24 |
| ENSG00000117519 | CNN3 | 4 | 94896949 | 94927223 | 30275 | - | protein_cod. | 16690.62 | 24868.18 | 11453.55 | 14356.97 | 20710.53 | 8937.79 |
| ENSG00000135636 | DYSF | 2 | 71453722 | 71686768 | 233047 | + | protein_cod. | 2655.52 | 1698.22 | 4384.61 | 3353.58 | 2204.57 | 5193.08 |
| ENSG00000186994 | KANK3 | 19 | 8322584 | 8343262 | 20679 | - | protein_cod. | 2257.23 | 4007.72 | 2948.58 | 3945.85 | 4201.93 | 5177.89 |
| ENSG00000175130 | MARCKSL1 | 1 | 32333839 | 32336233 | 2395 | - | protein_cod. | 8786.11 | 9698.24 | 8003.97 | 8408.33 | 7819.15 | 4811.13 |
| ENSG00000138722 | MMRN1 | 4 | 89879532 | 89954629 | 75098 | + | protein_cod. | 19949464 | 204322.7 | 16868.85 | 263839.6 | 303443.4 | 38908.39 |
| ENSG00000138119 | MYOF | 10 | 93306429 | 93482334 | 175906 | - | protein_cod. | 12506.28 | 4986.79 | 5959.34 | 11865.43 | 2877.56 | 11681.51 |
| ENSG00000167693 | NXN | 17 | 799310 | 979776 | 180467 | - | protein_cod. | 5646.52 | 7277.79 | 2263.45 | 4812.87 | 5743.42 | 2401.48 |
| ENSG00000115155 | OTOF | 2 | 26457203 | 26558698 | 101496 | - | protein_cod. | 1187.21 | 1087.97 | 463.93 | 498.71 | 109.01 | 146 |
| ENSG00000197694 | SPTAN1 | 9 | 1.29E+08 | 1.29E+08 | 81105 | + | protein_cod. | 10798.23 | 8180.84 | 6609.8 | 8411.32 | 9523.71 | 8408.68 |
| ENSG00000115306 | SPTBN1 | 2 | 54456317 | 54671446 | 215130 | + | protein_cod. | 23119.16 | 28282.09 | 11418.88 | 25384.26 | 39159.74 | 14476.62 |
Three human dermal lymphatic endothelial cell lines (HDLEC-5,6, and 7) were investigated under 21% pO2 as well as under 1% pO2 (Hypox-5,6, and7). RNASeq was performed by the NGS-Integrative Genomics Core Unit, UMG, Göttingen (details see: [10]. The number of reads is presented. The molecules shown here are not regulated by hypoxia, with two exceptions: Otoferlin (OTOF), which is a moderately high expressed gene, is downregulated by hypoxia. OTOF belongs to the ferlin family of proteins, additionally including dysferlin (DYSF) and myoferlin (MYOF). ANKRD37 is a lowly expressed gene, which is upregulated by hypoxia.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



