
Submitted:
25 September 2024
Posted:
26 September 2024
You are already at the latest version
Abstract
Five new species of the spider genus Toxopsoides Forster & Wilton, 1973 are described from Tasmanian surveys by DPIPWE and Bush Blitz (T. driesseni, sp. nov, T. tarkayna, sp. nov., T. wybalena, sp. nov., T. deux, sp. nov.) and one from an intensive IBISCA survey (T. ibisca, sp. nov.) in southern Queensland. It is suggested that Toxopsoides is transferred to Cycloctenidae with which it shares most characters.
Keywords:
Bush Blitz
; IBISCA
; new species
Introduction
Toxopsoides Forster & Wilton, 1979 are small tree dwelling spiders strongly resembling the larger Cycloctenus L. Koch, 1878: they differ principally in that Cycloctenus has the ctenid-like eye arrangement with the ALE set up high in a line with the PME; whereas in Toxopsoides, the eye ALE are low, as in the Zorinae. At the inception of this project, Toxopsoides were placed in the family Desidae, remote from a very similar genus, Cycloctenus, Cycloctenidae. Toxopsoides are small spiders usually found on trees and both genera have featured commonly in Tasmanian Bush Blitz’, e.g., Tarkine, Gowen Brae, Flinders Island. Of Cycloctenus, four species from Tasmania were described by Hickman: more undescribed are known. The contentiously placed Toxopsoides include one species putatively shared with New Zealand and three from New South Wales and Queensland recently described by Smith (2013) who followed conventional wisdom and placed them in the Desidae. They are now placed in the Toxopidae (Wheeler et al., 2017). Several new species were taken in the above Bush Blitz’ as well as the IBISCA project (Staunton et al., 2011) in Queensland.
The Tasmanian species taken on two Bush Blitz surveys, plus one taken on a Button Grass survey by the Department of Primary Industry, Parks, Water and the Environment, Tasmania (DPIPWE), and one from Queensland are described.
This project was to establish the presence of genus in other locations, further supporting the statements of Smith (2013).
Material and Methods
Abbreviations. Morphology: Abbreviations are standard for the Araneae; in addition, RTA, retrolateral tibial apophysis; CO, copulatory orifice; FD, fertilisation duct. Leg measurements are given in the descriptions as leg 1: femur, patella, tibia, metatarsus (absent in palp), tarsus, total. Digital photographs were taken with a Leica microscope L16 with a Leica DFC 500 camera and the image stacking software program Zerene Stacker (http://www.zerenesystems.com/). The epigyne was drawn with a camera lucida on a Zeiss Stemi SV6. Scanning electron micrographs were done on a Hitachi TM1000; material was sputter coated with gold.
Taxonomy
Toxopsoides Forster & Wilton, 1973
Toxopsoides Forster & Wilton, 1973, p. 309. Type species, Toxopsoides huttoni Forster and Wilton, 1973, by original designation.
Diagnosis. Differs from the often sympatric Cycloctenus by the eyes being in the standard zorine (Miturgidae) configuration: AER viewed from above slightly recurved with ALE clearly closer to the AME (Figure 1b) than to the PME. Toxopsoides also have 5-7 pairs of strong ventral spines on tibiae, I, II and 3-4 pairs of strong ventral spines in both males and females on metatarsi I and II (Figure 1a) and a much less elaborate palpal bulb and RTA than Cycloctenus (e.g., Figure 6a).
Relationships. The primary diagnostic character mentioned by Smith (2013), the two rows of trichobothia on the metatarsi, is a lapsus of Forster & Wilton (1973) and has not been been seen on any material herein (e.g., Figure 1e,f). Equally, the line of setae on the anal tubercle was used by Silva (2002) as a diagnostic character of the ctenid subfamily Calocteninae but such a line is also evident in Toxopsoides (Figure 1c).
As noted, although Toxopsoides is presently placed in the Toxopidae (Wheeler et al., 2017, Gorneau et al., 2023), it differs from Cycloctenus (Cycloctenidae) somatically only in the ALE being set low in a miturgid position, rather than near the PME, as in the ctenid like condition in Cycloctenus. It has not been sequenced or included in a cladogram but appears in the Toxopidae by what I call “guilty by association”. Thus, it is suggested that Toxopsoides should be transferred to the Cycloctenidae.
The male palp of Toxopsoides species is minimally divergent with a large tegulum and the small interspecific differences confined to the apical fifth of the bulb; similarly, the female epigyne is very similar across species which maximum difference shown in T. driesseni, sp. nov.
Toxopsoides
Included species: Toxopsoides huttoni Forster & Wilton, 1973; T. erici Smith, 2013, T. kathleenae Smith, 2013, T. macleayi, Smith, 2013, T. driesseni, sp. nov., T. wybalena, sp. nov., T. takayna, sp. nov., T. deux, sp. nov., T. ibisca, sp. nov.
Toxopsoides driesseni, sp. nov.
Diagnosis. Males and females have a similar abdominal pattern to those of T. erici and T. huttoni but males differ in that the long grooved median apophysis is basal on the bulb and lacks strong rugosity in the much less extensive or protruding processes on the male palpal bulb.
Diagnostic description. Male palpal tibia at most half length of cymbium; with RTA with two lobes; short, broad, smooth conductor, tegular envelope large, dominant with large (embolic) process (Figure 2a); tegular base =0.65 of bulb. Female with spermathecae ca. 1.35 times longer than wide (Figure 2d); epigyne with long median scape with widely curving lateral ridge (Figure 2c).
Material Examined. Tasmania. Holotype male, Lake St Clair, Airstrip Rd (site 5M) [42.06S, 146.17E], 30 Oct 2001, M. Driessen, QM S65850. Paratype female, QM S86836, same data but 10 Apr 2001, QM S86036; 1 male, McPartlan Pass, 12 March 1998, M. Driessen, QM S51336;1 male, Lake St Clair (A6-2), 6 Apr 2000, M. Driessen, QM S62054.
Description. Holotype male QM S65850
Carapace 2.04 long, 1.86 wide. Abdomen 2.25 long, 1.11 wide. Total 4.30.
Leg measurements. I: 2.00; 0.57; 2.01; 2.18; 0.84; 7.59. II: 1.74; 0.51; 1.50; 0.98; 0.32; 5.04. III: 1.55; 0.65; 1.28; 1.22; 0.69; 5.37. IV: 2.00; 0.57; 2.01; 2.18; 0.84; 7.59. Palp: 0.90; 0.47; 0.95; —; 1.22; 3.53.
Colour. Carapace pallid with narrow dark edges and broad subcentral dark bands extending from PLE; abdomen pallid with dark irregular mottling laterally and a pair of long dark bands anteriorly and smaller dark spots posteriorly.
Legs pallid with a dark spot anteriorly on trochanter I—IV; femora with two dark bands; medially and indistinct dark zone apically; pedal patellae with dark zone anteriorly; two annulations on mid-tibiae I-IV; one in distal half and one apically on metatarsi I—IV. Palp entirely pallid. Sternum, maxillae and labium pallid; abdomen ventrally with large dark median arrow-shaped area, flanked with pallid zone and the incursion of the dark flanks.
Palp. Tibia clearly wider than long; RTA is two similarly long lobes, overlaid in ventral view (Figure 2a). Cymbium with narrow lateral margins, apical cone symmetrical short conical with 2 spines on the ventral and lateral faces. Bulb ovoid, with dominant tegulum distally traversed partially by sinuous ridge; sperm ducts form tight basal fold and extend in one wide fold to embolus; large curved median apophysis and below it a long rugose conductor tip.
Female QM S86836. Similar to male except: Trichobothria, long distal on tarsi; none evident on dorsal metatarsi I. Spines: I; tibia I, II, 7 long pairs of spines; metatarsi I, I with 4 long pairs. STC with 7 teeth; ITC, small, bare.
Epigyne. Wide with large opening and narrow median plate.
Distribution. Known only buttongrass plains in the Lake St Clair area, central Tasmania.
Toxopsoides wybalena, sp. nov.
Figure 4a, b
Diagnosis. Males differ from those of T. takayna, sp. nov. in the absence of two large spines on the distal cymbium and the more slender cymbial cone (Figure 4b).
Diagnostic description. Male palpal tibia slightly more than half length of cymbium; RTA with two lobes; short triangular conductor; tegular envelope moderately large, not dominant with small (embolic) process (Figure 4b; tegular base =0.74 of bulb. Female unknown.
Material Examined. Tasmania. Holotype male, Flinders Island, Walkers Lookout, 40.054°S, 148.08°E, 20-30 Mar 2014, R. Raven, TMAG.
Description Holotype Male TMAG
Carapace 2.30 long, 2.15 wide. Abdomen 2.41 long, 3.07 wide. Total length, 4.7.
Leg measurements. I: 1.66; 0.62; 2.19; 1.56; 0.66; 6.69. II: 1.66; 0.63; 1.56; 1.51; 0.73; 6.09. III: 1.59; 0.33; 1.32; 1.12; 0.71; 5.06. IV: 2.07; 0.38; 1.63; 2.04; 0.89; 7.01. Palp: 0.90; 0.18; 0.38; --; 0.60; 2.07.
Colour. Carapace like T. takayna but with dark divergent lines along caput margins; abdomen like T. takayna. Legs orange brown, femora with 2-3 dark annulations and one distally on tibiae. Spines like T. takayna.
Palp. Tibia clearly longer than wide; RTA is two unequal lobes inner lobe slender spinelike, outer dorsal lobe short conical (Figure 4b). Cymbium with narrow lateral margins, apical cone symmetrical trianguloid with 4 spines apically and 4 more basal and lateral. Bulb long ovoid, with dominant tegulum with small lateral median apophysis and small hooked embolus, sperm ducts a sinuous lobe of two folds extending to broad distal loop (Figure 4b).
Distribution. Known only from Flinders Island, Tasmania. According to indigenous history (consistent with geology), Flinders Island is the focal point of radical drying about 4800 years ago, as much of the world’s water was bound up in polar ice and glaciations. At that time, Flinders Island connected Tasmania to Victoria as the Bass Strait closed. More significantly, it is recorded as a major catastrophe for the indigenous people. Concomitantly, in 1847, up to 130 First Nations people had died at Wybalena on Flinders Island and all were moved to Tasmania where many more died.
Toxopsoides takayna, sp. nov.
Diagnosis. Similar abdominal pattern to T. erici and T. huttoni but differs in the much less extensive or protruding processes on the male palpal bulb.
Diagnostic description. Male palpal tibia at most half length of cymbium; with RTA with two lobes; short, broad, smooth conductor, tegular envelope large, dominant with large (embolic) process (Figure 5a); tegular base =0.79 of bulb. Females unknown.
Material examined. Tasmania; Meredith Range Regional Reserve; E of Arthur River, 41.48467°S, 145.4737°E, Nothofagus with little understorey, open ground, on bark, 27 Jan-6 Feb 2015, R. Raven (RR20).
Description Holotype Male TMAG
Carapace 2.50 long, 2.00 wide. Abdomen 3.03 long, 1.75 wide. Colour in alcohol. Carapace pallid with irregular dark bands from caput margins submarginally, bands narrow posteriorly. Abdomen dorsally with black flanks for anterior half, a bright white zone with laterally dark ostiate region with posterior flares and dark irregular transverse band just anterior to spinnerets. Legs pallid: femora with two irregular dark bands medially, patellae distally dark, two brown annuli on tibiae; chelicerae fawn anteriorly darkly smudged.
Eyes. Eight in two rows, anterior row clearly narrower (0.72 of PER), eyes smaller and upcurved; AME their length from clypeal margin and two-thirds their diameter apart; all oval; AME: ALE: PME: PLE, 1.38. 1.00, 1.62. 1.90, respectively. Lower edge of ALE bisects AME. PME one AME from each other and PLE. PER so recurved as to be almost 2 rows.
Chelicerae. With a downwardly directed spine on the upper third (Fig. xx).
Spines. Tibia I, II with 7 pairs of long spines, lateroventrally; metatarsi I, II with 5 such pairs. Palp: fe: p1d1.1.1; pa d1; ti d1; cymbial cone with 10 spines. Palp: 1.18, 0.47, 0.53, --. 1.22, 3.40.
Palp. Tibia not much longer than wide; RTA is two unequal lobes overlaid in ventral view (Figure 5a, b), ventral lobe a low broad ridge (Figure 5b, outer dorsal lobe slender pointed. Cymbium with narrow lateral margins, apical cone symmetrical digitiform with 6 spines apically and 4 more basal and lateral 7 spines over the ventral and lateral faces. Bulb long ovoid, with dominant tegulum with small lateral median apophysis and small hooked embolus, sperm ducts a basal lobe extending to broad distal loop (Figure 5a).
Distribution. Known only from the Meredith Range, in the Tarkine region, south-western Tasmania.
Toxopsoides deux, sp. nov.
Figure 7a,b
Diagnosis. Similar abdominal pattern to T. erici and T. huttoni but differs in the much less extensive or protruding processes on the male palpal bulb.
Diagnostic description. Male palpal tibia at most half length of cymbium; with RTA with two lobes; tegulum deeply incursed distally for one-third tegulum; very small, indistinct conductor, tegular envelope almost absent with small (embolic) (Figure 7b) process; tegular base =0.66 of bulb. Females unknown.
Material Examined. Tasmania. Holotype male, Meredith Range Regional Reserve; 41.5328°S, 145.1075°E, 27 Jan-6 Feb 2015, R. Raven, TMAG.
Description. Holotype Male TMAG
Similar to T. deux, sp. nov. but: colour in ethanol. Carapace fawn with irregular submarginal dark bands; legs fawn with dark annuli; abdomen dorsally mottled brown with two small dark chevrons posteriorly.
Abdomen. A line of long distinct setae on the anal tubercle (Fig. xx).
Palp. Tibia clearly longer than wide; RTA is two unequal lobes, juxtaposed in ventral view (Fig. xx), inner dorsal lobe slender pointer, outer lobe widely triangular. Cymbium with narrow lateral margins, apical cone symmetrical triangular with 7 spines over the ventral and lateral faces. Bulb subcircular, with dominant tegulum traversed by deeply procurved embolic ridge, sperm ducts confined to basal and prolateral; small hooked median apophysis beside large triangular lobe.
Distribution. Known only from the Tarkine region in western Tasmania.
Toxopsoides ibisca, sp. nov.
Diagnosis. Males share with T. huttoni (sensu Smith, 2013) the rugose conductor tip (Figure 8a) but differ in the longer, more slender conductor, the shorter pointed tegular envelope (TEP, Figure 8a). They differ from all other species in the long male palpal tibia (Figure 8a).
Diagnostic description. Male palpal tibia longer than cymbium; with RTA with two lobes; narrow, fluted conductor, tegular envelope large, dominant with large (embolic) process (Figure 8b); tegular base =0.82 of bulb. Female with spermathecae set diagonally (Figure 2d); epigyne with broad median scape with paired lateral lobes (Figure 8d).
Material Examined Queensland: holotype male, Lamington National Park, IBISCA survey, site 900B, 28°238”S, 153°145”E, 950m, pitfall Trap, 12-21/02/2007, K. Staunton, QM S76179; paratype female, same data but site 900C, QM S76193.
Other Material Examined. LAMINGTON NATIONAL PARK, IBSICA SURVEY: 1 fe., IQ-1100-B, SC20035, 28°259”S, 153°162”E, 1142m, Malaise Trap, 7-17/10/2006, C. Lambkin, N. Starick, QM S76454; 1 fe., IQ-500-A, SC20173, 28°216”S, 153°142”E, 560m, pitfall Trap, 8-12/10/2006, K. Staunton, QM S81032; 1 fe., IQ-300-A, SC20359, 28°148”S, 153°137”E, 267m, pitfall Trap, 9-11/10/2006, K. Staunton, QM S81047; 2 fe., IQ-900-C, SC20436, 28°24”S, 153°149”E, 944m, pitfall Trap, 11-20/10/2006, A. Cameron, QM S81079; 1 ma., 1 fe., IQ-300-B, SC20441, 28°155”S, 153°139”E, 282m, pitfall Trap, 11-21/10/2006, D. Putland, QM S81087; 1 fe., IQ-700-D, SC20443, 28°204”S, 153°129”E, 748m, pitfall Trap, 11-20/10/2006, S. Maunsell, QM S81120; 1 fe., IQ-1100-C, SC20507, 28°26”S, 153°167”E, 1106m, flight intercept, 17-27/10/2006, G. Monteith, QM S76230; 1 fe., IQ-1100-B, SC30742, 28°259”S, 153°162”E, 1142m, dung baited pitfall, 22-27/10/2006, R. Menendez, G. Monteith, QM S76425; 1 ma., IQ-1100-D, SC30714, 28°262”S, 153°17”E. 1140m, dung baited pitfall, 22-27/10/2006, R. Menendez, G. Monteith, QM S76434; 1 ma., IQ-300-A, SC30564, 28°148”S, 153°137”E, 267m, mushroom baited pitfall, 16-21/10/2006, R. Menendez, G. Monteith, QM S76361; 1 fe., IQ-300-B, SC30556, 28°155”S, 153°139”E, 282m, mushroom baited pitfall, 16-21/10/2006, R. Menendez, G. Monteith, QM S76426; 1 fe., IQ-300-D, SC30704, 28°142”S, 153°133”E, 248m, mushroom baited pitfall, 21-26/10/2006, R. Menendez, G. Monteith, QM S76372; 1 fe., IQ-500-B, SC30668, 28°212”S, 153°141”E, 514m, mushroom baited pitfall, 18-23/10/2006, R. Menendez, G. Monteith, QM S76410; 1 fe., IQ-700-A, SC30566, 28°188”S, 153°121”E, 746m, mushroom baited pitfall, 16-21/10/2006, R. Menendez, G. Monteith, QM S76365; 1 ma., IQ-700-A, SC30705, 28°188”S, 153°121”E, 746m, dung baited pitfall, 21-26/10/2006, R. Menendez, G. Monteith, QM S76337; 1 ma., IQ-700-C, SC30757, 28°193”S, 153°128”E, 748m, dung baited pitfall, 24-29/10/2006, R. Menendez, G. Monteith, QM S76335; 1 ma., IQ-700-C, SC30759, 28°193”S, 153°128”E, 748m, dung baited pitfall, 24-29/10/2006, R. Menendez, G. Monteith, QM S76382; 1 fe., IQ-900-C, SC30750, 28°24”S, 153°149”E, 944m, mushroom baited pitfall, 22-27/10/2006, R. Menendez, G. Monteith, QM S76394; 1 ma., IQ-500-B, SC22168, 28°212”S, 153°141”E, 514m, flight intercept, 15-25/01/2007, G. Monteith, QM S76302; 1 ma., IQ-1100-C, SC22177, 28°26”S, 153°167”E, 1106m, flight intercept, 16-26/01/2007, G. Monteith, QM S76279; 1 fe., IQ-1100-D, SC22178, 28°262”S, 153°17”E, 1140m, flight intercept, 16-26/01/2007, G. Monteith, QM S76291; 7 ma., fe., IQ-900-A, SC22254, 28°234”S, 153°141”E, 904m, pitfall Trap, 12-21/02/2007, K. Staunton, QM S76187; 6 ma., 6 fe., IQ-900-C, SC22256, 28°24”S, 153°149”E, 944m, pitfall Trap, 12-21/02/2007, K. Staunton, QM S76193; 7 ma., 3 fe., IQ-900-D, SC22257, 28°227”S, 153°131”E, 920m, pitfall Trap, 12-21/02/2007, K. Staunton, QM S76183; 7 ma., 3 fe., IQ-1100-A, SC22258, 28°258”S, 153°159”E, 141m, pitfall Trap, 12-21/02/2007, K. Staunton, QM S76215; 11 ma., 6 fe., IQ-1100-B, SC22259, 28°259“S, 153°162”E, 1142m, pitfall Trap, 12-21/02/2007, K. Staunton, QM S76203; 6 ma., 6 fe., IQ-1100-C, SC22260, 28°26”S, 153°167”E, 1106m, pitfall Trap, 12-21/02/2007, K. Staunton, QM S76200; 8 ma., 5 fe., IQ-1100-D, SC22261; 28°262”S, 153°17”E, 1140m, pitfall Trap, 12-21/02/2007, K. Staunton, QM S76206; 1 fe., IQ-900-D, SC20756, 28°227”S, 153°131”E, 920m, flight intercept, 15-25/10/2006, G. Monteith, R. Menendez, QM S7624.
Holotype male QM S76179
Carapace 1.12 long, 0.97 wide. Abdomen 1.08 long, 0.59 wide. Total length 2.2.
Leg measurements. I: 1.11; 0.41; 1.35; 1.19; 0.41; 4.46. II: 1.03; 0.35; 1.03; 1.00; 0.35; 3.76. III: 1.08; 0.41; 0.95; 1.03; 0.38; 3.84. IV: 1.38; 0.41; 1.30; 1.35; 0.46; 4.89. Palp: 0.49; 0.22; 0.27; --; 0.41; 1.38.
Colour. Carapace, legs and chelicerae yellow brown, carapace altar with darker central region. Abdomen dorsally pallid (other material greenish brown) with darker median band.
Carapace. Pear-shaped, caput not raised, no “saddle” anterior to fovea. Cover of light brown hairs. Eyes. Configuration: 2-4-2 from above, 2-2-4 from in front. From above, ALE beside PME. PER in two rows. Chelicerae. Small, fangs short, transverse. Pair of inwardly directed spines on upper face; other male showed spine on only one chelicera (Figure 1b, arrow). Furrow with line of 6--8 teeth medially, dorsal of which is line of 6--8 long curved spines on ridge; 4 small spaced teeth on retromargin; boss small, distinct. Maxillae rectanguloid, lacking grooves. Labium subquadrate. Serrula long. Sternum cordate.
Legs. Femora appear dark through uniform cover of short black bristles. Trochanteral notches very shallow. Plumose hairs absent. Weak preening comb on metatarsi III, IV. Trichobothria. Metatarsi and tarsi with trichobothria in single narrow band. Erect trichobothria-like hairs on ventral tibiae and metatarsi I, II (Figure 1d, e).
Spines. Patellae with two dorsal spines one basally and one on apex; 6 pairs of strong spines on raised bases on tibiae I, II, 4 such pairs on metatarsi I, II; ventral spines on tibiae and metatarsi III, IV weak. I: fe p1d2; pa d1.1; ti d1w v2.2.2.2.2.2; me v2.2.2.2. II: like I but fe p1d2. III: fe p1d3r1; pa d1w.1; ti p2d2r2v2.2.2. IV: fe p1d3r1; pa d1.1w; ti p2d2r2v2.2.2; me p3r4v.2+3 distal. Palp: p1d3; pa d1 basal plus strong erect spine distally; ti v2w d6w.
Palp. RTA small, distolateral, bipartite: basal a conical point, distal is broad flange that appeared acute in ventral view. Tibia as long as patella, longer than cymbium, line of setae marks invagination of distal edge of tibia ventrally. Cymbium apically rounded with 3 upwardly curved spines apically on lower face and 2 on prolateral edge; dorsal scopula distinct; no groove retrolaterally. Tegulum dominant with all sclerites in distal quarter. Spermatic duct sinuous and visible. Embolus small, distal.
Abdomen. Spiracle distinct, transverse. Colulus distinct with 6 setae. PLS conical, apical segment domed. Spinnerets. PLS 0.5 size and diameter of ALS. PMS as long as PLS but conical and less sclerotised. Fringe on ventral face of anal tubercle.
Female QM S76193
Colour in Alcohol. Carapace and abdomen dark, carapace with broad submedian lateral pallid band and median pallid band. Abdomen dorsally dark with small pallid areas, unpaired; ventrally dark with pallid genital area and booklungs, triangular area points distal from it, plus a pair of irregular pallid lines laterally, fusing posteriorly to large diamond shaped area including tracheal spiracle. Legs light brown with slighlt darker annulations on femora.
Epigyne. Spermathecae set diagonally (Figure 8d); epigyne with broad median scape with paired lateral lobes (Figure 8d) and median pair of small crescentic shaped insemination apertures.
Otherwise, similar to male.
Distribution. Known only from Lamington National Park, southeastern Queensland.
Discussion
Two main points emerge from this work: reflections upon the presumed diversity of the Australian spider fauna and the unexplored diversity of this group in Australia,
The curious issue about the Tasmanian diversity is that V. V. Hickman (1894-1984) did not describe any Toxopsoides species and yet he was a specialist in small spiders and did describe new species of the slightly larger and related genus, Cycloctenus; both occur in similar habitats and indeed near Hickman’s Fern Tree home on Mt Wellington, kunanyi, In Bush Blitz surveys of Tasmania from 2010-2016 (https://bushblitz.org.au/tasmania-2010/; https://bushblitz.org.au/skullbone-plains-tas-2012/; https://bushblitz.org.au/flinders-island-tasmania-2014/; https://bushblitz.org.au/tarkine-tasmania-2015/; https://bushblitz.org.au/southwest-national-park-tasmania-2016/; https://bushblitz.org.au/bruny-island-tasmania-2016/), a number of Toxopsoides species were encountered (see above reports). Not all were described here as the scope would then grow considerably. The most likely possibility is that the relative abundance of Toxopsoides in Tasmania has changed since Hickman collected and they have become much more common.
How many spider species are there in Australia. Raven (1988) posited that the spider fauna of Australia was then about 25% known based on a rough approximation from recent works. At the time, it was thought that the more diverse araneomorph groups, like Salticidae and Thomisidae, would not show such a substantial increase because the fauna seemed better know and the groups were larger. Smaller lesser known groups, like the Oonopidae have exploded in diversity and clearly many more remain to be described. However, in 2008, before of the Maratus “explosion” (2012), only eight species Maratus were known, not withstanding genus synonymies and despite species synonymies, the time at writing (July 2024) is 111, an increase of 1387% in 16 years in a group that was thought to be well known. However, that meteoric rise in the number of species is against the total diversity of around 800 saliticid species, then known
Yeates et al. (2003) used the “revision multiplier approach” and concluded that the actual number of spider species in Australia is closer to 20,000; it is presently still less than 5000. Given that Raven et al. (2023) increased the known species diversity in one group of Miturgidae by 16 fold (3 known to 48), that figure seems more likely. However, to approach the the multiplier required, the increase needs to occur in groups presently well represented, like the Salticidae and in that context, the Maratus “explosion” seems to be an exception, rather than the rule. In the WSC (2024), we see that worldwide, the most diverse groups are Salticidae, Linyphiidae and Araneidae (6697, 4858, 3128 species, respectively). However, in Australia, only 36 species of Linyphiidae are named and most are introduced; without doubt, the linyphiid fauna is diverse but again, the starting value is small. Likewise, although the Australian Oonopidae have increased by 2300% (presently numbering 193 species growing from 8 in 1988.
Thus, although revisions may show very high relative increases in diversity, they are usually in groups that have low species numbers. Looking over a number of projects underway and indications from molecular studies, the most diverse and unnamed families (with expected new species) are likely to be Salticidae, Mygalomorphae and the litter based marronoid families (each 500), Gnaphosidae, Theridiidae, Miturgidae, Lycosidae (each 300), Clubionidae, Araneidae, Linyphiidae, the cribellate families, (each 200), Zodariidae (100), Oxyopidae, Orsolobidae, Cycloctenidae, Selenopidae, Hahniidae, Thomisidae, Ctenidae (each maximally 50); thus a predicted increase in about 3950 species. Given that Australia is still not at 5000 species, the initial predection of 10,000 species in Australia (Raven, 1988) still seems likely.
Acknowledgments
This project was initiated with the support of a Bush Blitz grant managed by Biodiversity Assessment and Management (BAAM), Queensland. It is based upon collections made in Tasmania with Tasmanian Museum and Art Gallery, Hobart while on Bush Blitz surveys to the Tarkine and Flinders Island. Collections were also made by Dr M. Driessen in Lake St Clair region as well as extensive collections made by Griffith University and the Queensland Museum at Lamington National Park, Queensland (IBISCA, see Staunton et al., 2011). All line drawings are by Wendy Hebron; Figure 1a is by Dr B Baehr.
References
- Forster, R. R. & Wilton, C. L. (1973). The spiders of New Zealand. Part IV. Otago Museum Bulletin 4: 1-309.
- Gorneau, J. A., Crews, S. C., Cala-Riquelme, F., Montana, K. O., Spagna, J. C., Ballarin, F., Almeida-Silva, L. M. & Esposito, L. A. (2023). Webs of intrigue: museum genomics elucidate relationships of the marronoid spider clade (Araneae). Insect Systematics and Diversity 7(5, 5): 1-18. [CrossRef]
- Hazzi, N. A. & Silva-Dávila, D. (2012). A new species of Caloctenus Araneae: Ctenidae) from Colombia. Zootaxa 3315: 65-68. [CrossRef]
- Raven, R.J. (1988). The current status of Australian spider systematics, pp. 37-47, figs. 1, 2, 1 table. In, Australian Arachnology, (eds.) Austin, A. D. and Heather, N.W., Australian Entomological Society, Miscellaneous Publ. No. 5, 137pp.
- Silva-Dávila, D. (2004). Revision of the spider genus Caloctenus Keyserling, 1877 (Araneae, Ctenidae). Revista Peruana de Biología 11: 5-26.
- Silva-Dávila, D. (2007) Mahafalytenus, a new spider genus from Madagascar (Araneae, Ctenidae). Proceedings of the California Academy of Sciences 58: 59-98.
- Silva-Dávila, D. (2003). Higher-level relationships of the spider family Ctenidae (Araneae: Ctenoidea). Bulletin of the American Museum of Natural History 274: 1-86. [CrossRef]
- Smith, H. M. (2013). The spider genus Toxopsoides (Araneae: Desidae: Toxopinae): new records and species from Australia. Proceedings of the Linnean Society of New South Wales 135: 19-43.
- Staunton, K.M., Fellows, C.S., Kitching, R.L., Nakamura, A., Burwell, C.J., Raven, R.J., 2011. Limited surrogacy between predatory arthropods along an altitudinal gradient in subtropical rainforest. Memoirs of the Queensland Museum Nature 55(2): 291-302.
- Wheeler, W. C., Coddington, J. A., Crowley, L. M., Dimitrov, D., Goloboff, P. A., Griswold, C. E., Hormiga, G., Prendini, L., Ramírez, M. J., Sierwald, P., Almeida-Silva, L. M., Álvarez-Padilla, F., Arnedo, M. A., Benavides, L. R., Benjamin, S. P., Bond, J. E., Grismado, C. J., Hasan, E., Hedin, M., Izquierdo, M. A., Labarque, F. M., Ledford, J., Lopardo, L., Maddison, W. P., Miller, J. A., Piacentini, L. N., Platnick, N. I., Polotow, D., Silva-Dávila, D., Scharff, N., Szűts, T., Ubick, D., Vink, C., Wood, H. M. & Zhang, J. X. (2017). The spider tree of life: phylogeny of Araneae based on target-gene analyses from an extensive taxon sampling. Cladistics 33(6): 576-616. [CrossRef]
- Yeates, D. K., Harvey, M. S. & Austin, A. D. 2003. New estimates for terrestrial arthropod species-richness in Australia. Records of the South Australian Museum Monograph Series No. 7: 231–241.
Figure 1.
Toxopsoides. a, T. erici, male, QM S31334, habitus showing strong paired spines; b--e, T. takayna, sp. nov., b, “face” showing spine on chelicerae and ALE set low; c, line of setae on anal tubercle; d, metatarsus showing two trichobothria (red arrows) and e, line of setae laterally.
Figure 1.
Toxopsoides. a, T. erici, male, QM S31334, habitus showing strong paired spines; b--e, T. takayna, sp. nov., b, “face” showing spine on chelicerae and ALE set low; c, line of setae on anal tubercle; d, metatarsus showing two trichobothria (red arrows) and e, line of setae laterally.
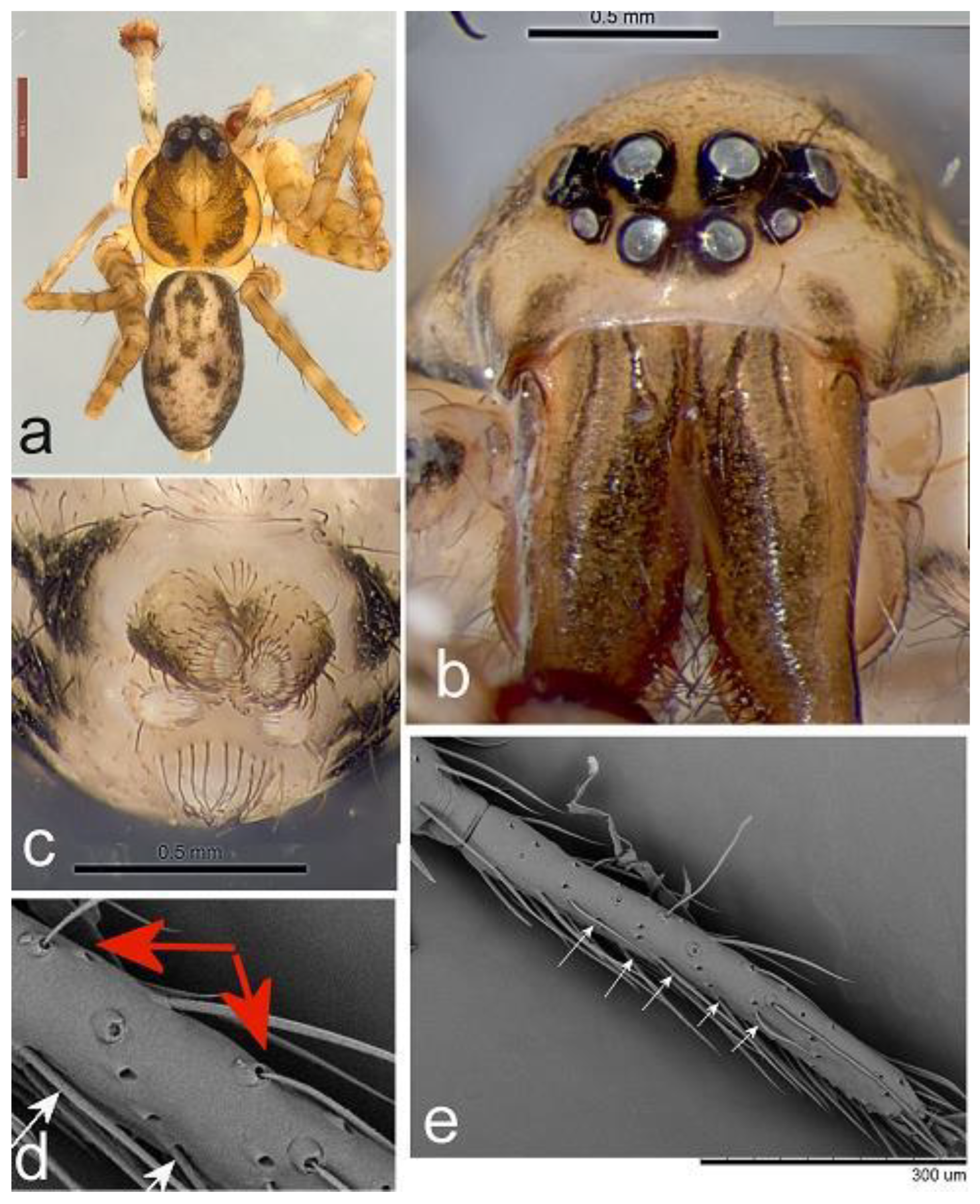
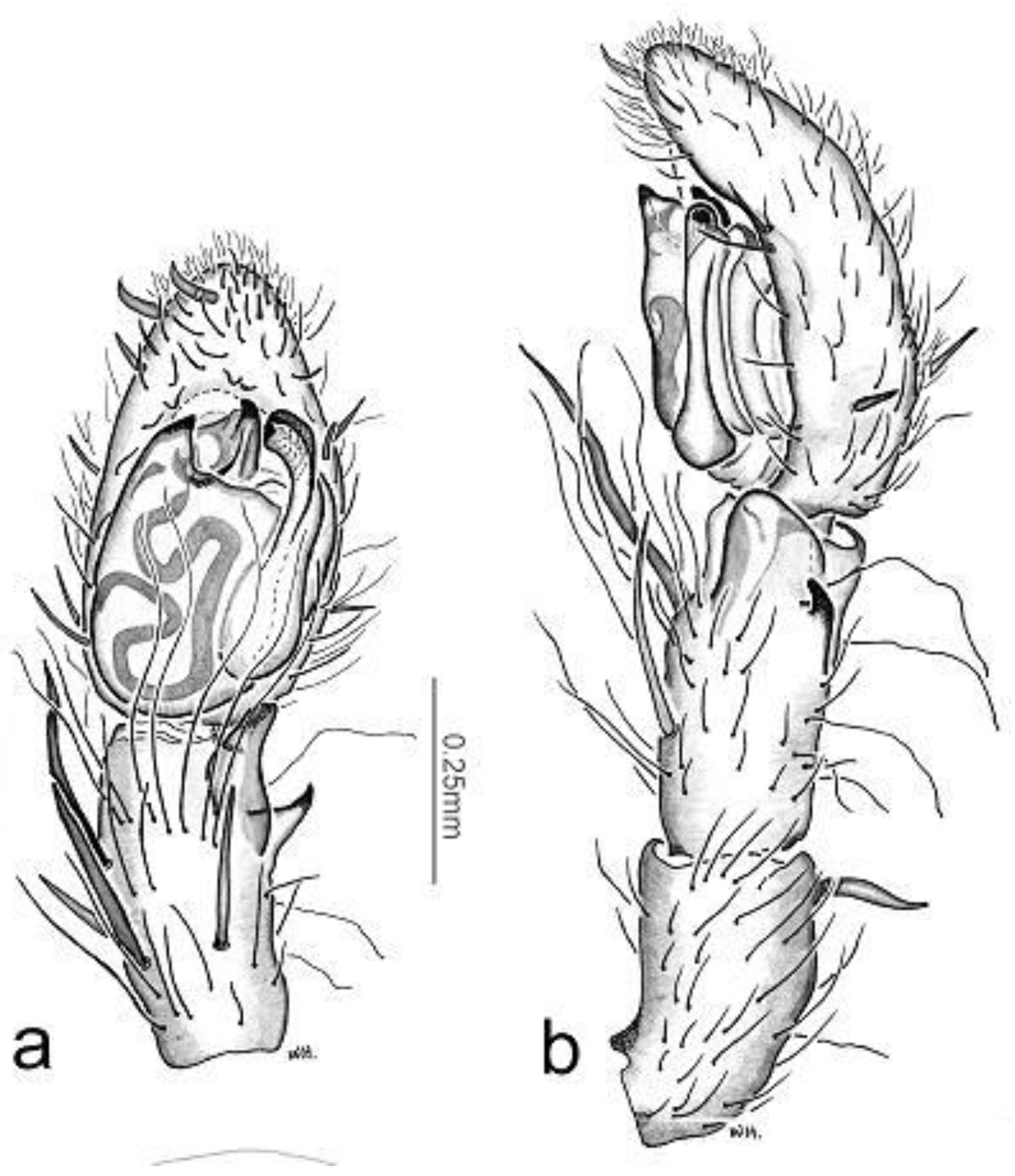
Figure 2.
Toxopsoides driesseni, sp. nov. a, b. Holotype male, QM S65850. a, b, palpal tibia, cymbium, and bulb, ventral (a) and retrolateral (b) views. c, d, epigyne, Paratype female, QM S86036: external (c) and internal (d) views.
Figure 2.
Toxopsoides driesseni, sp. nov. a, b. Holotype male, QM S65850. a, b, palpal tibia, cymbium, and bulb, ventral (a) and retrolateral (b) views. c, d, epigyne, Paratype female, QM S86036: external (c) and internal (d) views.
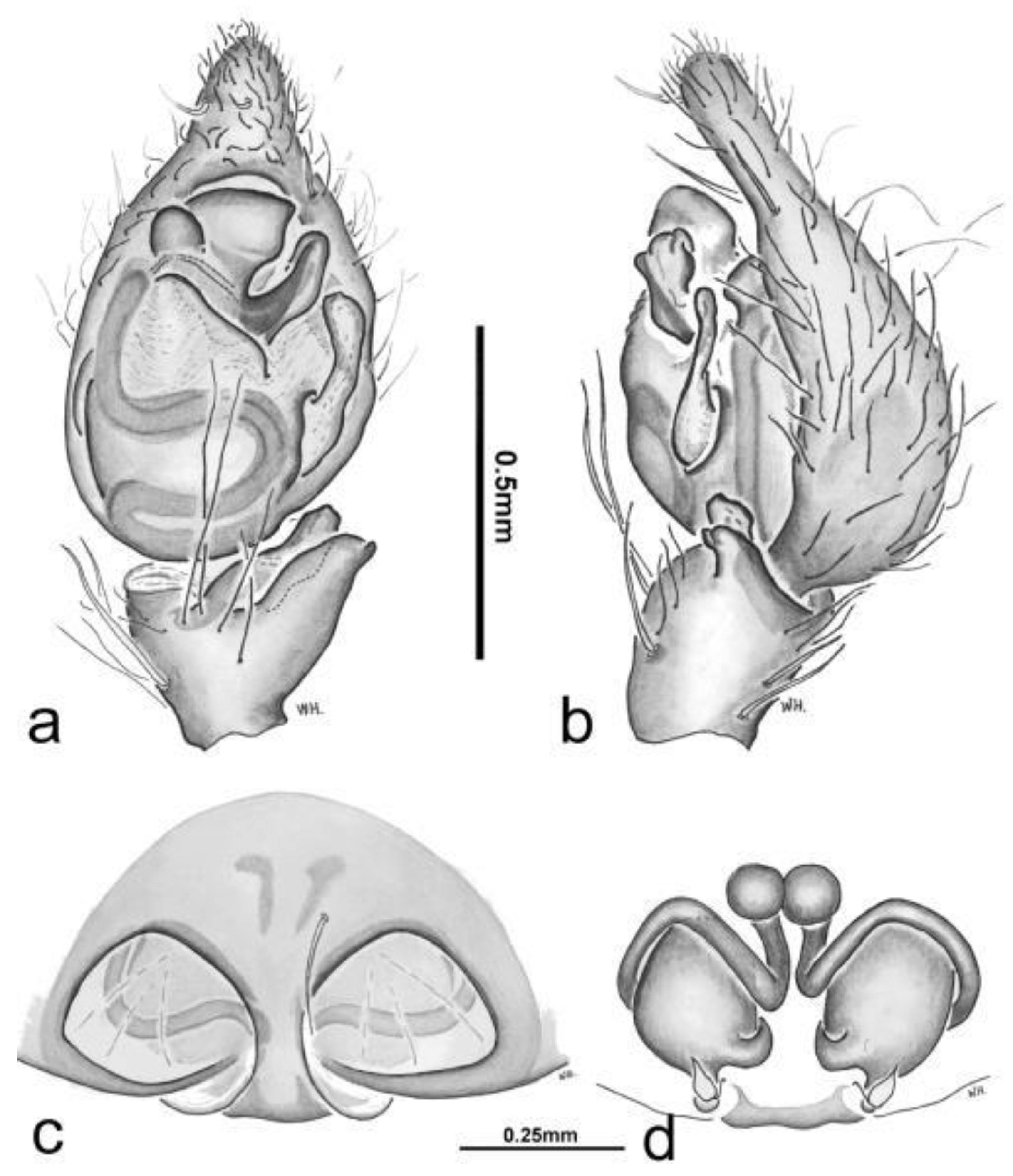
Figure 3.
Toxopsoides driesseni, sp. nov. Male, SEM: palpal tibia, cymbium, and bulb, ventral view; b, leg I, claws, third claw concealed; c, tarsal trichobothrium; d, leg I, prolateral view showing strong paired spines on tibia and metatarsus and long tarsal trichobothrium (inset).
Figure 3.
Toxopsoides driesseni, sp. nov. Male, SEM: palpal tibia, cymbium, and bulb, ventral view; b, leg I, claws, third claw concealed; c, tarsal trichobothrium; d, leg I, prolateral view showing strong paired spines on tibia and metatarsus and long tarsal trichobothrium (inset).

Figure 4.
Toxopsoides wybalena, sp. nov. Holotype male, TMAG: : a, habitus; b, palpal patella, tibia, cymbium and bulb, ventral view.
Figure 4.
Toxopsoides wybalena, sp. nov. Holotype male, TMAG: : a, habitus; b, palpal patella, tibia, cymbium and bulb, ventral view.

Figure 5.
Toxopsoides takayna, sp. nov. Holotype male, TMAG: palpal tibia, cymbium and bulb, ventral (a) and retrolateral (c) view; c, enlarged distal region of palpal bulb, ventral view.
Figure 5.
Toxopsoides takayna, sp. nov. Holotype male, TMAG: palpal tibia, cymbium and bulb, ventral (a) and retrolateral (c) view; c, enlarged distal region of palpal bulb, ventral view.
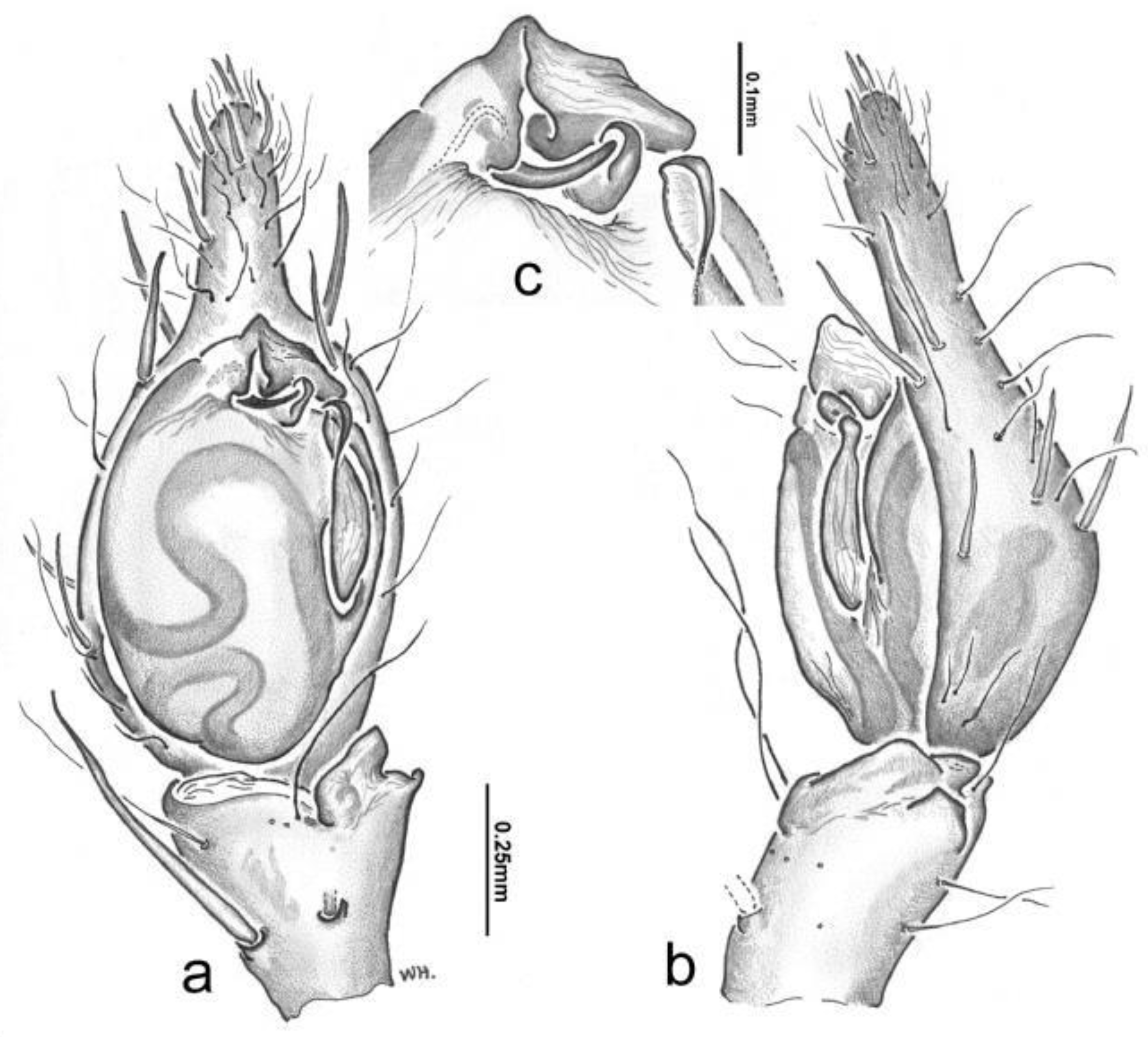
Figure 6.
Toxopsoides takayna, sp. nov. Male, TMAG. a, habitus; b, c, palpal tibia, cymbium and bulb, ventral (b) and retrolateral (c) views; d, SEM, metatarsus showing trichobothria.
Figure 6.
Toxopsoides takayna, sp. nov. Male, TMAG. a, habitus; b, c, palpal tibia, cymbium and bulb, ventral (b) and retrolateral (c) views; d, SEM, metatarsus showing trichobothria.

Figure 7.
Toxopsoides deux, sp. nov., males: a: habitus of two males; b, palpal patella, tibia, cymbium and bulb, ventral view.
Figure 7.
Toxopsoides deux, sp. nov., males: a: habitus of two males; b, palpal patella, tibia, cymbium and bulb, ventral view.
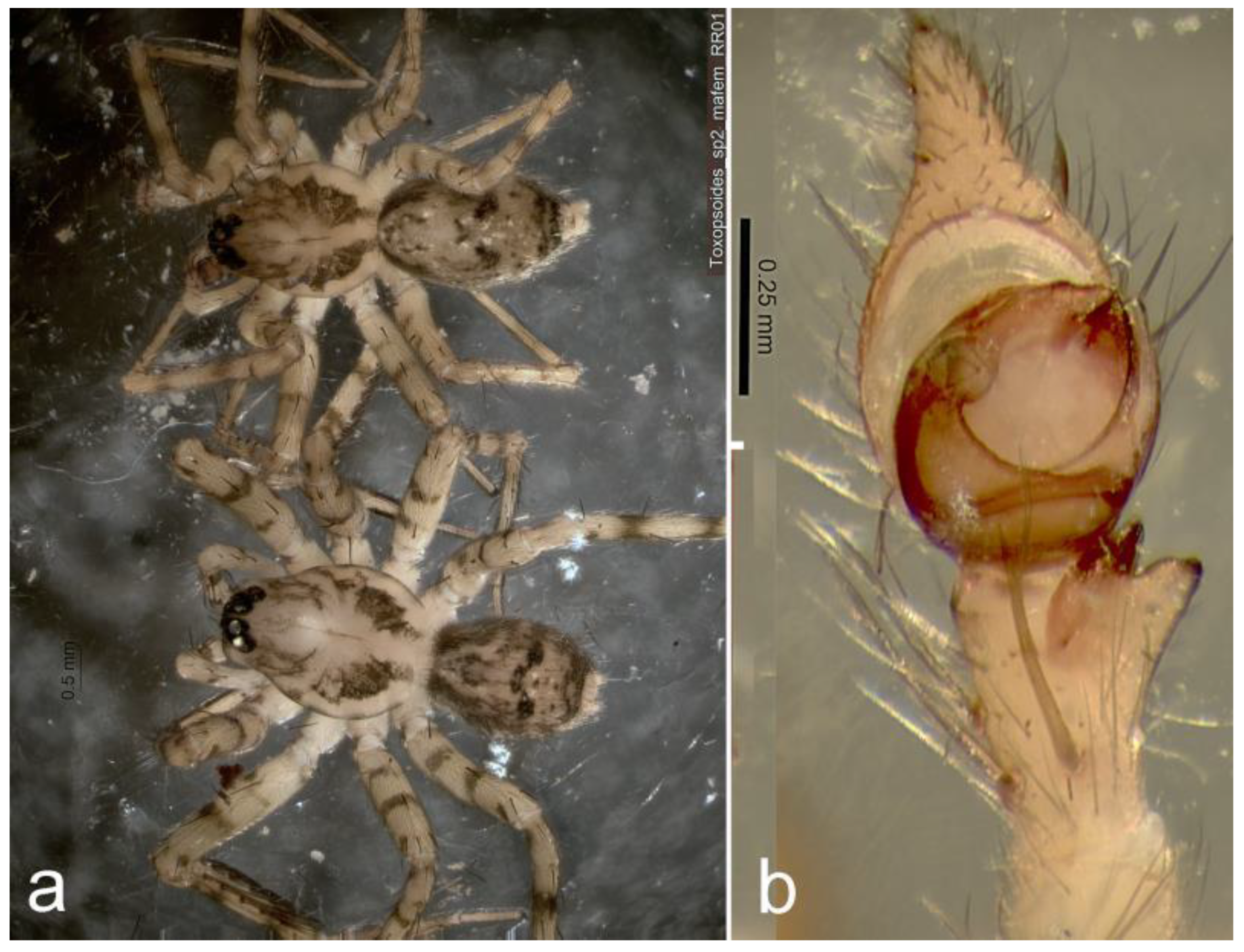
Figure 8.
Toxopsoides ibisca, sp. nov. a, b, Holotype male, QM S76179: palpal tibia, cymbium and bulb, ventral (a) and retrolateral (b) views; c, d, female epigyne, QM S76193, external (c) and internal (d) views.
Figure 8.
Toxopsoides ibisca, sp. nov. a, b, Holotype male, QM S76179: palpal tibia, cymbium and bulb, ventral (a) and retrolateral (b) views; c, d, female epigyne, QM S76193, external (c) and internal (d) views.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



