
Submitted:
27 September 2024
Posted:
29 September 2024
You are already at the latest version
Abstract
Tryptophan is an essential aromatic amino acid widely used in the pharmaceutical, agricultural, and food industries. Microbial fermentation, mainly using Escherichia coli, has become the preferred method for its production due to sustainability and lower costs. Optimizing tryptophan production requires careful control of various fermentation parameters, including nutrients, pH, temperature, and dissolved oxygen (DO) levels. Glucose, as the primary carbon source, must be fed at controlled rates to avoid metabolic overflow, which leads to byproduct accumulation and reduced production efficiency. Nitrogen sources, both organic (such as yeast extract) and inorganic (like ammonium), influence biomass growth and tryptophan yield, with ammonium levels requiring careful regulation to avoid toxic accumulation. Phosphate enhances growth but can lead to byproduct formation if used excessively. pH is another critical factor, with an optimal range between 6.5 and 7.2, where enzyme activity is maximized. Temperature control promotes growth and production, particularly between 33°C and 36°C. High DO levels increase tryptophan titers by boosting the pentose phosphate pathway and reducing byproducts like acetate. Furthermore, surfactants and supplements such as betaine monohydrate and citrate help alleviate osmotic stress and enhance precursor availability, improving production efficiency. Careful manipulation of these parameters allows for high-density cell cultures and significant tryptophan accumulation, making microbial fermentation competitive for large-scale production.
Keywords:
Tryptophan
; fermentation
; metabolic engineering
; biosynthesis
; Escherichia coli
1. Introduction
L-tryptophan is an aromatic amino acid discovered in casein extracts by Hopkins and Cole in 1901 [1,2]. This amino acid is used in the food and feed industries for its essential nutritional properties [3,4,5,6]. Humans and animals are unable to synthesize it internally, as its production incurs a substantial metabolic cost; thus, it is more advantageous for them to acquire it from their environment [6,7,8]. Tryptophan is widely used in the pharmaceutical industry for the treatment of schizophrenia and alcoholism and is a precursor to antidepressant and antitumor molecules such as serotonin and violacein [6,9,10,11,12]. In the agricultural industry, tryptophan is a precursor of the phytohormone indole-3-acetic acid [13].
Tryptophan can be obtained through chemical synthesis, enzymatic methods, and fermentations [1,14]. Chemical synthesis is unsuitable for complex compounds [15], as it produces mixtures of L- and D- isomers [12,16]. Like most amino acids, tryptophan can be produced by microorganisms that consume low-cost raw materials such as sugars and ammonium [12,17,18,19,20,21], promising an affordable and sustainable alternative [5,22].
High levels of production are desired at an industrial level [23]. The annual tryptophan production is estimated at 4.1x104 metric tons per year [24] and continues to increase [18,24,25]. Although other microorganisms have been used for tryptophan production [16,18,25,26,27,28,29], Corynebacterium glutamicum and Escherichia coli are the main study models and have been well characterized genetically [1,4,22,25,30,31,32]. E. coli has the advantage of being fast-growing, easy to culture, well-studied, has metabolic plasticity, and a broad range of genetic tools [24,30].
The production efficiency of a strain can be represented by the conversion of the carbon source to the desired product (Yp/s) [33]; this rate should be maximized by modifying its metabolism to reduce production costs and be able to use them industrially [6,20,33,34]. There are two theoretical maximum conversion rates of glucose to tryptophan [11]; the first is 0.227 g/g of glucose and is used for strains that import glucose through the PEP: glucose phosphotransferase system (PTS) [26,34,35] since this consumes 50 % of the PEP produced by the cell and cannot be directly channeled to tryptophan production. The second value of 0.46 g/g of glucose is for strains that incorporate glucose by a system independent of PEP consumption [20]. Tryptophan production is still low compared to lysine and glutamate production [16]. The maximum conversion rates achieved are around 0.238 g/g of glucose, representing 50% of the theoretical maximum conversion rate, making industrial production processes expensive and encouraging further research to improve tryptophan production [5,6,15].
Production titers are the most used parameter to represent a strain's efficiency. Paradoxically, strains with a higher conversion rate are not always those with the highest titers [11]. To define which parameter is most important in tryptophan production, it is necessary to consider the cost of the carbon source against the cost of purification in downstream processes.
Although much effort has been invested in developing new engineered strains and mathematical models to improve tryptophan production [16,23,36,37], their metabolic behavior is affected by specific environmental parameters and substrate availability [38]. The most notable example is the strain FB-04 harboring the plasmid pSV03, which produced 13.3 g/L of tryptophan [39], while a later publication managed to increase production to 38.1 g/L by modifying the culture conditions [40]. The optimization of tryptophan production must be considered from the culture medium formulation, carbon source supply, fermentation parameters, precursor supply, and the simultaneous regulation of biosynthetic pathways and gene expression [23,41,42,43]. The genetic modification strategies used for tryptophan production have been previously reviewed in depth [44,45]. However, fermentation strategies have not been analyzed in the same detail [46]. For this reason, in this review, we will focus mainly on describing how fermentation conditions affect tryptophan production.
2. Genetic Modifications
The tryptophan biosynthetic pathway is extensive and complex, so it has been arbitrarily divided by several authors into modules that include the tricarboxylic acid cycle (TCA), the glycolysis (EMP pathway), pentose phosphate pathway (PPP), shikimate pathway, chorismate pathway, as well as pathways involving glutamine, serine, and the terminal branch of tryptophan synthesis [5,11,30,33,47,48,49]. These pathways are tightly regulated because tryptophan formation consumes an appreciable amount of resources and energy [50]. Some mechanisms that have been described are attenuation, feedback inhibition, feedback repression, and feed-forward inhibition [5,15,30,47,48,51]. To improve tryptophan production, researchers have removed attenuation and repression of critical enzymes, overexpressed enzymes involved in rate-limiting steps, blocked competitive and degradative reactions, improved precursor supply, modified transporters, and boosted the tryptophan biosynthesis pathway (see Figure 1) [4,24,50,52,53].
More than 40 individual genetic modifications have been identified in E. coli to improve tryptophan production [54]. However, a small group of these modifications have been consistently used to achieve titers of more than 30 g/L, as shown in Figure 2. These modifications are closely related to the final pathway of aromatic compound production, particularly tryptophan, so their use is stable in various genetic backgrounds. One of the most representative strains is the TRTH strain, which has been tested under different fermentation conditions and has been used as parental of other strains, achieving production between 32.24 and 60.2 g/L [38,53,55,56], which is the highest titer achieved.
2.1. Strains Derived from Mutagenesis
One of the first strategies to generate overproducing strains was the introduction of random mutations into the genome by physical, chemical, or biological agents. The mutants generated by these strategies can have diverse behaviors, even adverse to the objective sought, so a selective environment is necessary [10,20]. To find strains that produce tryptophan enough to excrete it, analogs such as 5-methyl-tryptophan or 5-fluoro-tryptophan are used, which represses the enzymes involved in the formation of tryptophan. Strains that manage to grow in the presence of these analogs usually present mutations in their regulatory mechanisms and should be overproducers [25,57].
Another strategy for strain selection is the use of biosensors that indicate the level of tryptophan production in each cell, such as riboswitch-based high throughput screening (HTS) [51] or the tnaC regulatory element fused to a reporter gene [10]. The overproducing strains that use these systems can be isolated using fluorescence-activated cell sorting (FACS) [10,51]
Another strategy is CRISPR/Cas9-facilitated engineering with growth-coupled and sensor-guided in vivo screening (CGSS), in which growth depends on the efficiency with which an enzyme works, and production is estimated by a reporter gene. This strategy does not require equipment such as FACS to select cells that show improved production [58].
Obtaining randomly generated overproducing mutants can lead to new genetic modifications that can be identified utilizing new sequencing technologies, which can be used in the rational design of strains that improve tryptophan production [10].
2.2. Modification and Overexpression of the Tryptophan Operon
The enzymes that produce tryptophan from chorismate are expressed from the trpEDCBA operon [16]. This operon is regulated by the trpL attenuator in their mRNA 5’ UTR, whose activity depends on tryptophan. Their transcription is regulated by the TrpR, which acts as a repressor when bound to tryptophan. The overexpression of this operon enhances tryptophan production and can be achieved by removing the trpL attenuator, swapping the trp promoter, and cloning the operon in stable and multicopy vectors [3,4,11,24,26,27,34,35,38,41,43,47,51,52,53,55,56,59,60,61,62,63,64,65,66,67,68,69,70,71].
Overexpression of the trp operon may sound logical. However, the anthranilate synthase enzyme encoded by the trpE gene is, in fact, the main bottleneck in the operon. This enzyme catalyzes the first reaction of the pathway and is inhibited by tryptophan. The trpES40F mutant is insensitive to inhibition and has been extensively used to create overproducing strains. The trpES40 and trpD genes have been overexpressed in the FB-04/pSV03 strain and their derivatives without modifying the native trp operon, achieving yields of up to 45.6 g/L [33,39,40]. Other mutant alleles insensitive to feedback inhibition are trpEM293T/N168D/A478T/C237R [50]. The regulation of the final tryptophan pathway is very susceptible to the accumulation of intermediates, so tryptophan must be produced in a balanced manner with their precursors. The heterologous expression of the trpCR378F gene from Aspergillus niger improves tryptophan production, mainly when anthranilate accumulates because it inhibits the indole glycerol phosphate synthase (IGPS) of E. coli [11,35].
2.3. Eliminating the Degradation Pathway of Tryptophan
The tryptophan degradation pathway comprises the tryptophanase enzyme encoded by the tnaA gene. In this degradative pathway, tryptophan is converted to pyruvate, ammonium, and indole, which, as noted below, influence tryptophan synthesis. Under normal conditions, this enzyme has low activity, so it does not interfere with the metabolism of E. coli. However, high concentrations of tryptophan, such as those in overproducing strains, activate the transcription of tryptophanase, preventing reaching high production levels [72]. Silencing the tnaA gene is one of the most widely used modifications, along with overexpression of the trp operon to enhance tryptophan production [6,40,72].
2.4. Modifications to the Central Metabolic Pathways
Regarding tryptophan synthesis, the central metabolism can be divided into four main pathways: glycolysis, TCA cycle, pentose phosphate pathway (PPP), and glyoxylate shunt [30]. These pathways provide the metabolites and energy the cell needs to function. For the biosynthesis of aromatic amino acids, glycolysis is responsible for providing phosphoenolpyruvate (PEP), while PPP provides erythrose-4-phosphate (E4P), phosphoribosyl pyrophosphate (PRPP) and the reducing cofactor Nicotinamide Adenine Dinucleotide Phosphate Reduced (NADPH). Glutamine, which donates an amino group to tryptophan production, is derived from the TCA cycle [30].
PEP is a crucial precursor of aromatic compounds and the TCA cycle and is central to glucose uptake [30]. To transport a mol of glucose by PTS is necessary to consume a mol of PEP, generating pyruvate that can be channeled to the production of acetate [4,22,30,33,34,62,69]. In this condition, 50% of the cellular PEP is used to import sugar, while around 1.5-3% is redirected towards producing aromatic amino acids [6,22]. The PTS system can be inactivated by deleting the ptsI [11], ptsG [4,6,52,69], or ptsH genes [5,63]. Still, it reduces glucose consumption so significantly that it affects growth, impairing tryptophan production, although it favors the conversion rate of glucose to tryptophan [4,6,11,22,52,69]. The PTS has been attenuated with the ptsHN12S mutant instead of deleting it [33]. In strains with the PTS inactive, glucose transport has been complemented by overexpressing galactose permease (galP) [5,59,63] or expressing the glucose facilitator (glf) of Zymomonas mobilis [6]. When transporting glucose through both systems, overexpression of glucose kinase (glk) is required to efficiently incorporate glucose into glycolysis [6,59].
PEP is consumed by pyruvate kinase (pykF or pykA) to perform its primary function: to supply the TCA cycle [33,47,73]. Deleting these genes allows the accumulation of PEP; however, when made simultaneously attenuates the TCA cycle, affecting biomass production [6,30,34,48]. Deletion of pykF reduces the flux on glycolysis and the acetate production pathway, redirecting it towards the PPP [33]. Eliminating pyruvate kinase activity forces the redistribution of metabolic fluxes to generate pyruvate through other enzymes such as anthranilate synthase; by this means, tryptophan production can be growth-coupled, as it compels the organism to produce anthranilate to support its growth [73].
PEP can be regenerated from pyruvate or oxaloacetate by overexpression of PEP synthase (ppsA), which catalyzes the reverse reaction to pyruvate kinase, or by overexpressing PEP carboxykinase (pckA), which consumes oxaloacetate [24,41,51,71]. The ppc gene encodes phosphoenolpyruvate carboxylase, which consumes PEP to produce oxaloacetate conversely to pckA; deleting it reduces PEP consumption [47]. Pyruvate has also been channeled to oxaloacetate production by expression of C. glutamicum pyruvate carboxylase (pycP458S) to create a PEP-pyruvate-OAA node in which oxaloacetate is converted to PEP by pckA [6]. Similarly, PEP has been regenerated from acetyl-CoA by overexpressing the genes aceB, mdh, and pckA. The aceB gene encodes malate synthase in the glyoxylate shunt that consumes acetyl-CoA, while mdh encodes malate dehydrogenase, producing oxaloacetate (see Figure 1) [47].
When strategies to increase PEP production deplete the supply of the TCA cycle, this can be reactivated by adding citrate to the medium. To efficiently metabolize citrate in the TRP07 strain, it was necessary to overexpress the citrate transporter citT and the TCA cycle genes acnAB and icd [48]. Something that is striking in Figure 1 is that the reactions of the TCA cycle that include the conversion of α-ketoglutarate to malate have not been overexpressed even though the rest have been done in different studies, possibly because carbon atoms would be lost because these are part of the TCA cycle that produces CO2.
In E. coli, aromatic metabolites are generated from the condensation reaction between PEP and E4P, which gives rise to DAHP [33,48,50,51,71]. The shikimate pathway requires equal proportions of E4P and PEP, but E4P cellular levels are lower than PEP and can be a limiting substrate [6]. E4P can be increased by overexpression of the tktA gene, which encodes a transketolase, increasing tryptophan production [5,24,34,41,50,52,69,71]. An alternative that is more efficient in yeast to increase the supply of E4P is the introduction of the xfpk gene from Bifidobacterium adolescentis, which converts a molecule of fructose-6-phosphate and phosphate into E4P, with the production of acetyl phosphate. This strategy has also increased tryptophan production in E. coli and can bypass the carbon-losing enzyme 6-phosphogluconate dehydrogenase that produces CO2. However, it is necessary to reinforce the incorporation of acetyl phosphate into the metabolism to avoid acetate accumulation [6].
Another function of the PPP is to provide the NADPH required for tryptophan synthesis. The pntAB genes have been overexpressed, which encode a membrane-bound complex with transhydrogenase function that transfers a proton from NADH to NADPH. This allows for a more suitable NADPH:NADH ratio for tryptophan production [6,74]. The sthA gene encodes a transhydrogenase that catalyzes the reverse reaction to that of pntAB and helps control NADPH accumulation [74].
2.5. Modification of the Pathways Involved in Acetate Accumulation
Acetate accumulation is detrimental to biomass production and constitutes a waste of carbon that prevents achieving high conversion rates for tryptophan production. There are two pathways for acetate production; the main one is formed by the genes pta-ackA, while the secondary one is formed by the gene poxB [41,47,55,62,70]. Deleting the pta gene effectively decreases acetate production and favors tryptophan production [4,24,41,51,55,56,61,70]. However, it has also been reported that pta deletion impairs biomass and tryptophan production in some strains. As alternatives, the transcription of the pta gene has been attenuated using a small RNA, or the wild-type gene has been replaced by a less active allele called pta1 so that biomass production is not affected [33,40,63]. Another strategy to reduce acetate production is eliminating the acetate kinase activity catalyzed by two enzymes encoded by the genes ackA and tdcD [38]. Silencing of poxB has little impact on acetate accumulation [47]; in strain TRP07, it was used as an insertional site, and its elimination alone was not evaluated [48]. However, two or more genes involved in acetate production can be silenced to increase its effect, as in strains TRTHBPA, TRP12, or TRP30 [24,51,56,70].
Gecse and colleagues propose that eliminating poxB and pta is more effective in glucose-limited culture media. In contrast, overexpression of the gltA gene for a non-limited medium is better for preventing the metabolism overflow that generates acetate [5,75]. However, the harmful effect of pta deletion is also influenced by dissolved oxygen (DO) levels; at low DO levels, the impairment of growth is more pronounced [76].
In E. coli, acetate can be inefficiently consumed by its conversion to acetyl-CoA through two irreversible reactions catalyzed by the enzyme acetyl-CoA synthetase. The first step consumes one molecule of ATP to form acetyl-adenosine monophosphate (AMP). The second step converts acetyl-AMP to acetyl-CoA. Overexpression of the acs gene allowed strain T16 to consume the acetate produced [47].
By decreasing acetate production, the accumulation of pyruvate and acetyl-CoA may increase, which can be funneled into the synthesis of other byproducts [4,38,55,62]. After acetate, the most produced compounds are lactate, ethanol, and formate; to avoid their accumulation in the TRP12 strain, the genes ldhA, adhE, and pflB that code for the enzymes that branch towards the synthesis of these compounds have been deleted [24].
2.6. Modification to the Common aromatic Amino Acid Pathway
There are three isoforms of the enzyme 3-deoxy-7-phosphoheptulonate synthase, encoded by the genes aroG, aroH, and aroF, and are mainly regulated by phenylalanine, tryptophan, and tyrosine, respectively. The enzyme encoded by the gene aroG has the most significant activity in E. coli, followed by aroF and aroH [45]. To increase the activity of this enzyme, it has been overexpressed, and N-terminal coding sequence libraries have been used to improve their translation levels [51]. Overexpression of the feedback inhibition-resistant mutant aroGS180F is commonly used to enhance tryptophan production [4,5,6,24,26,34,35,41,48,51,52,59,63,71]; other mutants used for tryptophan production are aroF394 [21], aroFI11- [39,40] and the aroGD6G/D7A which seems to be a better alternative to the aroGS180F [11,58].
After overexpressing the genes aroB, aroD, aroE, aroK, aroA, and aroC, the enzymes shikimate dehydrogenase (aroE) and shikimate kinase I (aroK) have been identified as rate-limiting steps in shikimate production and have therefore been successfully overexpressed [5]. The insufficient activity of shikimate kinase has also been corrected by overexpressing the isoform II encoded by the gene aroL. This gene was identified after introducing perturbations in the supply of the carbon source and allowed to identify the gene aroB encoding a 3-dehydroquinate synthase as a control step in tryptophan production [50].
2.7. Supply of Tryptophan Precursors
2.7.1. Glutamine
The glutamine pathway comprises the enzyme glutamine synthase, which combines glutamate and ammonia. This enzyme is encoded by the gene glnA and may be a rate-limiting step in tryptophan production [5]. E. coli can supply glutamine to the enzyme anthranilate synthase during the first hours of fermentation but then become scarce as glutamate accumulates [30]. Wild-type glutamine synthase from E. coli can be inhibited by high concentrations of NH4+. This does not happen in Gram-positive bacteria such as Bacillus subtilis and Bacillus megaterium, although they are sensitive to glutamine [74]. Mutations have been introduced into the glnA gene of Bacillus subtilis (glnABs) to eliminate the inhibition by glutamine and increase its affinity for ammonia and glutamate [77]. Overexpression of the glnAL159I/E304A mutant of Bacillus subtilis has improved tryptophan production [5]. Another modification used to improve tryptophan production is glnAY405F from C. glutamicum [10].
2.7.2. Serine
Serine is another precursor that can be limiting in tryptophan production [5,26]. The serine synthesis pathway involves dehydrogenation, transamination, and finally, dephosphorylation of 3-phosphoglycerate from glycolysis [49,78]. Its production has been improved through the overexpression of the genes constituting its synthesis pathway: serA, serB, and serC [5,24]. The serA gene encodes a phosphoglycerate dehydrogenase, which is inhibited by serine accumulation, so a serAH344A/N364A mutant is commonly used [5,6,10]. Another mutant used is serAT372N, accompanied by deleting the sdaB gene, which codes for the serine degradation reaction to boost serine supply for tryptophan production [24].
2.8. Modification of the Tryptophan Transport Systems
The tryptophan transport system in E. coli consists of the permeases aroP, tnaB, and mtr and the aromatic amino acid exporter yddG [5,20,53]. Modifying transport systems to accumulate tryptophan in the medium reduces the inhibition of its synthesis when it accumulates intracellularly [16]. Eliminating the tnaB gene favors the extracellular accumulation of tryptophan by preventing its re-incorporation into the cell [5] and improving the production of tryptophan in the T13 strain [20]. In the T12 strain, a possible bottleneck in the tryptophan synthesis pathway prevented production from improving when overexpressing the yddG gene; instead, there was a greater secretion of phenylalanine and tyrosine that crystallized in the medium, decreasing the intracellular accumulation of anthranilate [5].
Deleting aroP and overexpressing yddG promotes extracellular accumulation of tryptophan but depletes the cell's reserves for protein synthesis, affecting growth [53]. For E. coli, this is 0.054 mmol of tryptophan/g of biomass [20]. Deleting aroP is the least used method to produce tryptophan but has shown promising results for the strain GTP1017 [52], while overexpressing yddG in the strain TRTH-Y improved tryptophan production [53].
The tnaB gene is the most deleted transporter that improves the accumulation of extracellular tryptophan. Its elimination is favored because most strains have their tnaA gene silenced in the same operon, and both genes can be eliminated simultaneously. This gene has been modified independently or in combination with the other genes that constitute the tryptophan transport system [4,5,6,11,26,35,51,52,59,61,71].
2.9. Silencing of Transcriptional Regulators
The TrpR repressor regulates the transcription of genes involved in the production, degradation, transport, and regulation of tryptophan; among these are aroL, aroH, trpEDCBA, mtr, and trpR. Eliminating the trpR gene is a minimum requirement for a marked production of tryptophan by releasing the expression of multiple genes responsible for its production [6,27,40].
Similarly, the tyrosine repressor of transcription (tyrR) controls the production of common precursors for tryptophan and tyrosine, so silencing it has favorable effects on tryptophan production [6,48]. TyrR represses the aroF gene, a limiting factor in DAHP production [21].
The fructose repressor, FruR, is a global regulator of the central metabolic pathway. Its deletion activates genes involved in glycolysis, alternative sugar catabolism, the Entner-Doudoroff pathway, and PPP. Furthermore, it represses gluconeogenesis, the TCA cycle, and the electron transport chain. Although the fruR mutant produces more pyruvate and acetate by activating glycolysis, its deletion improves the production of tryptophan in a stable manner using sugars in different mixtures. The accumulation of pyruvate can stimulate the conversion of 3-phosphoglycerate to serine-phosphate, which can be converted to serine, improving tryptophan production and reducing the accumulation of precursors of the common pathway of aromatic amino acids such as quinone [49].
Tang and collaborators identified transcriptional regulators whose expression changes in overproducing strains. To test their effect, they modified several genes but only found an improvement in tryptophan production when deleting the putative regulator yihL, whose function needs to be better characterized [51].
2.10. Modulating Pathways Competing for Precursors
Chorismate is a common precursor for the synthesis of aromatic amino acids [51]; when it is not efficiently directed towards tryptophan synthesis, it accumulates, causing feedback inhibition, and phenylalanine and tyrosine can be produced as byproducts [23,26]. Blocking phenylalanine and tyrosine production by silencing the pheA and tyrA genes can improve tryptophan production [21,40]. However, auxotrophic strains are generated, which increases the manufacturing cost, and supplementation of the medium with these amino acids inhibits the common pathway for synthesizing aromatic amino acids, reducing the supply of precursors for synthesizing tryptophan [5]. To reduce competition for precursors and avoid generating auxotrophic strains, the native promoters of the tyrA and pheA genes have been replaced by the self-regulated promoter PfliC, increasing tryptophan production without affecting growth [51].
3. Fermentation Parameters
Fermentations to produce tryptophan are performed starting from an isolated colony activated in a rich medium, commonly Lysogenic Broth, then transferred to a seed culture medium like the fermentation medium but differing mainly because it has a higher amount of yeast extract and nitrogen. This medium produces healthy biomass and adapts the strain to the final medium. Depending on the scale of the fermentation, it can be done in a flask controlling only temperature and agitation, or it can be done in a fermenter controlling all the relevant parameters. Fermentation in seed medium is used to inoculate the fermentation medium, which is more focused on tryptophan production, so parameters such as DO, growth rate, feed rate, pH, agitation, aeration, and temperature are controlled, as described in Figure 3. When the initial glucose in the fermentation medium is exhausted, a concentrated glucose solution, sometimes mixed with other substrates, is pumped into the fermenter. Fermentations are stopped when tryptophan accumulation no longer increases. Table 1 summarizes the main parameters analyzed in this review for tryptophan production.
3.1. Factors Affecting Biomass Accumulation
Biomass in fermentations contains all the enzymes, precursors, and energy required for tryptophan synthesis [15,20]. Increasing tryptophan production depletes cell resources and can produce a metabolic overload that limits growth [20,30,82]. During the exponential phase of fed-batch fermentations, tryptophan production is proportional to the available biomass [70]. In theory, if the production rate is maintained in high-density cell cultures, the titers of the desired metabolite will be improved, and the efficiency of the fermentation and reduction of production costs will be possible [43,48].
The investigation of metabolites that could be limiting the production of tryptophan in the E. coli 10303 strain identified vitamins B1 and B2, the amino acids methionine, cysteine, leucine and threonine, and the nucleotides guanidine monophosphate (GMP) and guanine as substrates that improve biomass production, with leucine having the most significant impact on tryptophan production allowing alone to produce 0.983 g/L, which represents an improvement of 34.8% [19].
Serine is essential as a precursor of tryptophan and in biomass production. If serine is insufficient, tryptophan production is affected, while if there is an excess in its accumulation, homoserine dehydrogenase (thrA) is inhibited, which participates in the isoleucine synthesis pathway [5]. Adding isoleucine or threonine can correct the growth inhibition caused by an excess of serine [5,89]. The addition of corn liquor boosts growth in the T13 strain. After analyzing its composition, it was identified that its high isoleucine content was responsible for such an effect. For this strain, adding 0.5 g/L of isoleucine increased glucose consumption by 41.2% [5]. The Addition of casamino acids also promotes biomass production when serine accumulates. It is known that phenylalanine in casamino acids is responsible for restoring growth. However, the mechanism of action has yet to be entirely understood [89].
Previous studies have shown that biomass can be increased by supplementing the culture medium with yeast extract [59]. However, the results may vary depending on the commercial supplier [19].
Increasing the inoculum also increases the final production of biomass and tryptophan [43,59]. Higher amounts of inoculum reduce the lag phase of fermentation and allow it to enter the exponential phase quickly because it contains many extracellular enzymes that favor using fresh medium and allow it to adapt faster. However, too much inoculum causes excessively accelerated growth. It favors autolysis that occurs in the last stages of fermentation, so 20% inoculum (V/V) containing 5-6 g/L of dry cells was the optimal value for the TRTH strain [43].
Optimizing the activity of glycolytic enzymes also improves tryptophan production [23]. Adding KH2PO4 to the medium activates glycolysis and boosts biomass production by increasing the growth rate in the initial stage of fermentation; nevertheless, excessive KH2PO4 values boost glucose consumption so much that it causes an overflow of the metabolism that ends in the accumulation of organic acids. Optimizing the amount of inoculum and KH2PO4 in the TRTH strain made it possible to achieve one of the highest tryptophan titers in E. coli (59.55 g/L) without the need to use additional equipment to the fermenter [43].
High glucose concentrations generate osmotic stress and lower the activity of the TCA cycle through the Crabtree Effect, affecting biomass production. For the TRTHΔpta strain, 10 g/L of initial glucose accumulates the highest biomass [55]; high glucose concentrations induced acetate accumulation from the initial fermentation phase. Acetate profoundly affects bacterial metabolism, reduces glucose consumption, inhibits cell growth, and causes autolysis to the detriment of desired metabolites [55,62]. Acetate accumulation above 0.5-2 g/L prevents cultures from achieving high cell density because it inhibits the growth of E. coli [4,40,47,60,68]. For the TRJH strain, adding 2 g/L of acetate reduces biomass and tryptophan production by 11.0% and 17.9%, respectively [68].
Acetate production by E. coli responds to at least three environmental conditions: low dissolved oxygen (DO), metabolic overflow when excess glucose is available, and high pH values [61,64]. Acetate inhibits growth by depleting the energy of bacteria. When protonated, it can diffuse through the membrane into the cytoplasm, lowering the pH by dissociating from its H+, and to compensate for pH, bacteria need to pump protons into the extracellular domain, expending ATP, becoming a futile cycle that causes an energy imbalance [30,55]. Acetate concentrations above 40 mM [55,80] also affect recombinant protein expression, reducing gene overexpression, especially in modified strains [38,68].
Acetate reduces the activity of isocitrate dehydrogenase by promoting its phosphorylation by the enzyme phosphatase/kinase (aceK), which triggers the activation of the glyoxylate shunt [81,82]. The greater flux in the glyoxylate cycle reduces oxidative respiration, affecting growth, a result also observed when overexpressing aceBA. In Addition, the aldehyde group of glyoxylate makes it highly reactive and toxic, so malate synthase (aceB) metabolizes it rapidly [82].
To increase cell density, acetate, NH4+, and other inhibitory metabolites could be removed using a disk stack centrifugal separator that recycles the cells to continue the fermentation [70]. An alternative process is using a vacuum scraper concentrator evaporation, which produces the highest reported titer of tryptophan in E. coli (60.2 g/L) [60]. Recycling cells reduces operating costs by reducing fermentation times. They are more efficient if only used in the final fermentation stage when inhibitory metabolites accumulate and tryptophan begins to crystallize [70].
Cations derived from alkaline solutions that control pH, such as NH4+, K+, and Na+, inhibit biomass production when accumulated in high concentrations. Strategies like combining two alkaline solutions to regulate pH, by reducing the inhibitory effect of each generated cation, have been assayed [61]. The following sections will describe this strategy in more detail.
For the T13 strain, biomass and tryptophan production increased as the temperature increased from 33°C to 36°C; 36°C was the highest temperature tested with the highest production obtained [5]. This high temperature, however, could affect serine production since it has been reported that a temperature of 30°C is better for serine production [90].
It is also necessary to consider the fermentation equipment's capacities, which can be a limitation in producing high-density cell cultures since they can be physically limited in the distribution of nutrients in the medium [43]. For example, due to the large scale of industrial reactors, zones with significantly high glucose concentrations are formed near the feed inlet, which triggers acetate formation [75]. Increasing the stirring speed cannot correct this phenomenon because the high shear forces cause mechanical damage to the bacteria [32].
3.2. Modulation of Growth Rate
The growth rate is a critical parameter in fermentations and can be controlled by manipulating the availability of a limiting substrate, commonly glucose [55,60,61]. By reducing the growth rate, bacteria require fewer amino acids for protein production, allowing resources to be redirected toward tryptophan synthesis and secretion [16,53,91].
An excess of glucose under aerobic conditions triggers metabolic overflow in E. coli, favoring ATP synthesis through acetogenesis and accelerating its growth [3,61,64,68,75]. In contrast, a low growth rate reduces acetate formation and favors the availability of dissolved oxygen. The specific growth rate should be controlled below 0.25 1/h to avoid the formation of byproducts and favor the accumulation of the desired metabolites [3,61,68]. A threshold growth rate of between 0.14-0.17 1/h has also been recommended, although this value may depend on the strain and the culture medium [3,65,68]. For the TRTHΔpta strain, a rate of 0.25 1/h produces the highest biomass and tryptophan. A low growth rate, especially during the early fermentation phase, reduces acetate accumulation, extending fermentation for longer and maintaining higher growth rates in the late phase, which favors biomass accumulation [38]. Lactic acid and alanine are other byproducts that can accumulate when the growth rate is too high; this leads to the accumulation of NH4+ due to the utilization of NH3∙H2O to neutralize them, which reduces biomass production [3].
Plasmids are commonly used to overexpress genes that enhance metabolite production. Bacteria that maintain a plasmid show impaired growth rates compared to those that do not, and bacteria with unstable plasmids tend to lose them or accumulate adaptive mutations that affect antibiotic resistance or gene expression [92]. Plasmid stability is reduced when the growth rate increases, preventing them from contributing to metabolite production [3,61]; this instability may be due to the accumulation of short-chain organic acids like acetate that interfere in the synthesis of nucleotides, lipids, and proteins and to the accumulation of NH4+ [3]. Nevertheless, highly stable plasmids could be used in industrial-scale fermentations even without adding antibiotics [71].
3.3. Ensuring Dissolved Oxygen (DO) Concentration
DO insufficiency affects cellular metabolism, especially the TCA cycle activity, and promotes acetate and glutamate accumulation. In fed-batch fermentations, dissolved oxygen can be maintained with aeration and agitation. However, when high cell density is reached, it is also necessary to control glucose availability to reduce metabolism and oxygen consumption [4]. Optimizing DO improves biomass and tryptophan production through a balanced metabolism [41,62].
3.3.1. Avoiding Acetate Accumulation
Acetate accumulation is a common problem when performing fermentations in E. coli [38,70]. Increasing the dissolved oxygen concentration boosts acetate consumption, decreasing their accumulation [4]. DO regulates the genes involved in acetate production at the transcriptional level. The main acetate synthesis pathway is expressed at low and high DO levels [41,47,62,70], so it has been considered a constitutive pathway [80]. However, this pathway is promoted at low DO levels with the transcriptional activation of the tdcD gene, whose product has acetate kinase activity [41]. This reversible pathway can produce small amounts of acetyl-CoA when acetate is too high [47]. The secondary pathway contributes less to acetate production [55]. However, it is activated in low oxygen levels [41,62] or in the presence of acetate itself [84]. The availability of oxygen during fermentation is a factor that influences the effect of mutations in the pta and ackA genes on biomass production; with low DO, biomass production is more affected [76,79].
The citrate synthase enzyme encoded by the gltA gene incorporates acetyl-CoA and oxaloacetate into the TCA cycle, oxidizing acetyl-CoA to CO2 [80]. This enzyme can carry a high metabolic flow; under aerobic conditions, around 70% of the total glucose is consumed by the glycolysis pathway and then by the TCA cycle [30] for the strain JB102/pBE7 up to 47.78% of the carbon atoms can be consumed to produce CO2 [26]. Low DO levels decrease the TCA cycle's activity by inactivating the citrate synthase, causing acetyl-CoA to accumulate and be redirected toward acetate production [55,80]. The accumulation of acetyl-CoA, in turn, inhibits the pyruvate dehydrogenase complex, increasing the accumulation of pyruvate [55]; in strains that have a decreased capacity to produce acetate, pyruvate is redirected to the synthesis of lactate and glutamate [4,38,55,62].
Under aerobic conditions, the enzyme isocitrate lyase (aceA) activity is lower than that of isocitrate dehydrogenase, channeling the flow of carbon toward the TCA cycle. When DO decreases, isocitrate dehydrogenase activity is reduced, redirecting the flow of carbon toward the glyoxylate shunt, which can reduce the accumulation of acetate [82].
High oxygen availability decreases the transcription of the fba gene, which is critical in glycolysis, preventing the accumulation of pyruvate and acetyl-CoA, which are acetate precursors. In Addition, it promotes the consumption of these precursors by increasing the transcription of the anaplerotic and gluconeogenesis genes, such as pckA, ppsA, ppc, and maeA [41]. Reducing the accumulation of these metabolites increases glucose conversion efficiency to tryptophan [41,62].
The rpoS gene transcribes the expression of several stress response genes that prepare the bacteria to enter the stationary phase; one of these genes is acs, which is repressed at low DO levels and is activated in the late fermentation phase when there is more DO [62,80].
Biomass and tryptophan production does not follow the acetate trend concerning DO levels. The highest biomass and tryptophan productions are 30% and 20%, respectively, requiring an intermediate DO concentration to achieve maximum production [62].
3.3.2. Redistribution of Flows between Metabolic Pathways
The DO availability profoundly affects gene transcription and metabolic flux distribution. Among the main pathways affected by DO levels are glycolysis, PPP, TCA cycle, and cytochrome activity [41,62]. A high level of DO enhances the supply of PEP, E4P, and NADPH, positively affecting tryptophan production [62].
By reducing glucose consumption by the PTS, PEP availability can be increased [22,62,69]. High DO levels reduce glucose uptake because the TCA cycle is activated, boosting ATP production that inhibits the activity of the phosphofructokinase encoded by the pfk gene [62]. The glycolysis and PPP divide at glucose-6-phosphate, which is a point that determines glycolysis and PPP fluxes efficiency [23]. Shifts in DO from 5% to 20% in the THTR strain can increase carbon flux into the PPP by up to 2.25-fold, supplying more E4P for aromatic amino acid production. Thus, favoring the oxidative branch of the PPP produces the cofactor NADPH, driving biomass and tryptophan synthesis [62].
Elevated DO levels stimulate gluconeogenesis by increasing the transcription of the genes pckA and ppsA, increasing the availability of PEP. The PPP is also activated by increasing the transcription levels of zwf, pgl, tktA, and araD, which supply E4P for tryptophan synthesis [41,62]. Finally, elevated DO levels stimulate the transcription of the genes aroG, aroK, and trpEDCBA involved in the final branch of tryptophan [41].
3.3.3. Oxidative Stress
Fermentations with a high DO level allow high-density growth in E. coli, favoring the production of tryptophan [41,62]. An adverse effect of high DO levels is that they cause oxidative stress by generating Reactive Oxygen Species (ROS), such as hydrogen peroxide, which promotes the formation of free radicals through Fenton reactions, damaging proteins, membranes, and DNA, resulting in cell death [38,41,81,83]. Oxidative stress causes a redistribution of the metabolic fluxes of E. coli; among the pathways that are attenuated are the TCA cycle and glycolysis, while the PPP, acetate pathway and glyoxylate shunt are activated, resulting in a high NADPH:NADH ratio and the production of α-ketoglutarate [82,83].
The glyoxylate shunt plays an essential role in the response to oxidative stress [81,82,83], participating in the metabolism of acetate and other substrates that can be converted to acetyl-CoA for redirection towards gluconeogenesis and cell growth. The overall effect of this cycle is to form one mol of malate from two mol of acetyl-CoA, thanks to the fact that it avoids the oxidative decarboxylation steps of the TCA cycle and conserves the carbon skeleton for the generation of biomass [81,82]. The glyoxylate cycle competes with the TCA cycle by reducing the production of α-ketoglutarate and glutamate [62,82]. Under conditions of oxidative stress, α-ketoglutarate and glutamate accumulate even though the TCA cycle is attenuated and the glyoxylate shunt is active because the enzyme α-ketoglutarate dehydrogenase is almost wholly inactivated, forming a bottleneck [62,83].
The accumulation of glutamate diverts resources that could be used to produce tryptophan, reducing the efficiency with which glucose is converted [62,70]. The production of glutamate from α-ketoglutarate by glutamate dehydrogenase (gdhA) consumes NADPH as a cofactor [93], causing an imbalance in the reducing power of the cell when produced in high quantities [41,62]. The reason why E. coli accumulates glutamate in strains seeking to enhance tryptophan production, which is not very clear; however, it could be due to an imbalance in the redox state that prevents the regeneration of NADP from NADPH. E. coli has a high demand for NADPH during its growth, supplied by the PPP and isocitrate dehydrogenase; however, when its growth is reduced, the excess NADPH is consumed, producing glutamate [26].
3.4. Effect of pH
The pH in fermentations can influence DO levels, growth rate, and metabolite production [4,41]. E. coli can adapt its metabolism to grow at a pH between 4.4 and 9.2, although it usually acidifies the medium by producing organic acids [61]. Low pH reduces its growth rate due to decreased respiratory activity that increases DO, improves plasmid stability, and reduces acetate production [25,32,61]. Low pH reduces the activity of enzymes such as phosphofructokinase, becoming a rate-limiting step in glycolysis, reducing glucose consumption, growth rate, and tryptophan production. However, attenuation of the glycolysis also reduces acetate accumulation because it reduces pyruvate and acetyl-CoA production [41].
Acetate production increases as pH increases; acetate accumulation has been observed to change from 6 to 12 g/L when pH changes from 6.0 to 7.5 in the Dpta/mtr-Y strain [41]. Acetate production could serve as a mechanism to neutralize high pH by activating the transcription of the pta gene [61]. In contrast, reducing pta activity can redirect carbon flow through the PPP and the common aromatic amino acid pathway [40].
Low pH induces sucB and sucC genes by activating the TCA cycle to exploit the high proton potential. The increased consumption of acetyl-CoA by the TCA cycle may reduce the metabolic overflow that produces acetate [4,41,61].
The tryptophan production pathway is also affected by pH. The enzymes encoded by the genes aroK, tnaA, and tnaB change their activity depending on pH. For example, shikimate kinase I (aroK) is induced at high pH [61], while at pH 8, the highest productions of tryptophan from indole and serine have been achieved [12].
Multiple factors determine the optimal pH for tryptophan production: lowering the pH to prevent acetate accumulation affects stable and optimally active enzymes at near-neutral pH [41]. In Aureobacterium flavescens, maintaining the pH to levels close to neutrality allows for the extension of the active growth period and improves tryptophan production [25]. Using a high pH to activate the enzymes involved in tryptophan production demands using alkaline solutions, causing the accumulation of cations such as NH4+, Na+, or K+ that make the medium toxic for bacteria. In contrast, acidic pH reduces the solubility of tryptophan, favoring its crystallization and avoiding feedback inhibition [41]. Although studies have determined that the optimal pH for tryptophan production is between 6.5-7.2 and maintaining the pH at 6.8 can produce 32.15 g/L of tryptophan in the TRJH/ΔPB strain, fermentations can be divided into several stages to first improve biomass production at low pH avoiding acetate production and then increase the pH for tryptophan production [61].
3.5. Culture Feeding Strategies
Substrates are added initially in batch fermentations, and their consumption continues until exhaustion. In fed-batch fermentations, the carbon source can be controlled to produce the metabolite of interest and maintain biomass production. In these last fermentations, there is an initial phase (trophophase) in which priority is given to the accumulation of biomass with rich media and an initial concentration of glucose [20,44]; after this initial glucose is exhausted, a more concentrated glucose solution is fed at a rate that varies depending on the feeding strategy, to start the formation of the desired product (idiophase) [20,55]. The feeding rate changes the bacteria's metabolic activity and the medium's volume, affecting the growth rate and the formation of products and byproducts [44]. If the feeding rate and the growth rate are not coordinated, low substrate concentrations result in low productivity, with long culture times; on the contrary, overfeeding induces an overflow of the metabolism that favors the generation of byproducts, such as small organic acids, which can result in the loss of the batch because the carbon flow is not being adequately incorporated into the tryptophan production pathway, causing a bottleneck [4,26,47,61,75,84].
E. coli tends to overload its metabolism, which is inconvenient because the high flux through the glycolysis generates pyruvate and acetyl-CoA that cannot be processed by the TCA cycle, redirecting the conversion of glucose toward acetate accumulation [4,68,80]. Despite the efforts that have been made to improve tryptophan production, the generation of byproducts is a problem, with acetate being the primary inhibitory metabolite [3,4,38,40]. Acetate accumulation enhances the formation of other byproducts and is toxic in the mid-logarithmic phase, making it challenging to achieve high cell density [38,63]. In fed-batch fermentations, longer growth phases are achieved than with batch fermentations, so the acetate accumulation significantly affects biomass production [68].
Acetate accumulation is related to glucose consumption and growth rate [55,68]. Many fed-batch fermentation strategies seek to reduce acetate production to achieve high-density cultures. One of the most used strategies is to reduce the feed rate to maintain glucose concentration below 2 g/L. This reduces growth rate and prevents metabolic overflow [4,38,55,62,63,64,68].
Although acetate is considered a waste product, it can be consumed with glucose and plays a vital role in the overall regulation of the E. coli metabolic network. However, acetate accumulation inhibits its recycling through the pta-ackA-acs pathway [62,84]. Consequently, glucose consumption decreases due to acetate accumulation [63] because acetate represses the transcription of genes involved in PTS glucose uptake and the TCA cycle [84].
Glutamate can be found at extremely high levels in overproducing strains such as S028 and JB102/pBE7 [26,59]. Glutamate accumulation can be decreased by reducing the glucose feed rate and enhancing tryptophan production [26,41,62]. Glutamate can be stocked in amounts that are problematic for tryptophan production [5,26]. Glutamate is not efficiently converted to glutamine, even though the accumulation of its precursor α-ketoglutarate has been reported to impulse glutamine synthesis [49], which acts as an amino group donor in the conversion of chorismate to anthranilate, making it an essential precursor for chorismate synthesis and potentially a limiting factor in tryptophan production [74].
3.5.1. Exponential Feeding
The exponential feeding strategy aims at adjusting the specific growth rate (µ) to a predetermined value by controlling the feed rate; this is calculated using an equation that considers how the glucose demand grows exponentially during the mid-logarithmic phase following the biomass production [3,65,94]. One of the objectives is to limit the specific growth rate below a threshold value (µcritical) to avoid acetate accumulation. For example, for the TRJH strain, it has been observed that a µ of 0.2 1/h gives the maximum production of tryptophan and biomass [68], while Liu, Chen, and Wu selected a growth rate of 0.15 1/h for several strains [33]. Although this approach seems simple in theory, slight variations in growth rates trigger glucose starvation and overfeeding intervals, which reduce biomass and tryptophan production, contributing to the formation of byproducts [65,75]. An attempt has been made to improve this strategy by eventually correcting the feed function with real-time measurements of glucose concentration, varying the growth rate at different stages of fermentation in a process known as pseudo-exponential feed rate [65], or combining the pseudo-exponential with glucose-stat feeding strategy [68].
The exponential feeding strategy is very effective during the exponential growth phase, where E. coli can take advantage of high feeding rates without acetate accumulation. However, when the growth rate is reduced at late stages, this strategy causes the accumulation of up to 80 g/L of glutamate, as was shown in the JB102/pBE7 strain due to metabolic overflow. For this reason, decreasing the feeding rate when E. coli reduces its growth rate while still producing tryptophan improves the glucose conversion rate [26].
3.5.2. Glucose-Stat Feeding Strategy
Lack of glucose in fermentation harms bacterial metabolism, affecting the production of metabolites [68]. Glucose feeding can be adjusted after measuring its concentration in the medium; glucose concentrations close to 1 g/L produce high concentrations of biomass and tryptophan [65], although concentrations lower than 0.2 g/L have also been used to reduce acetate production [26]. Performing measurements and controls manually is a tedious process if there are no online measuring instruments, which is why other strategies have been adapted to regulate glucose concentration through indirect measurements of its concentration, taking advantage of the most used sensors in fermenters.
Adjusting the glucose concentration is an effective control to regulate microbial metabolism, especially if it is done by computers that predict the glucose consumption behavior. The glucose concentration can be kept low to avoid the accumulation of acetate but sufficient to sustain growth; in the TRJH strain, it was possible to produce 51.4 g/L of biomass and 36.3 g/L of tryptophan by maintaining the glucose concentration at 0.15 g/L with a maximum specific growth rate of 0.275 1/h [68].
3.5.3. DO Feedback Control
DO levels affect tryptophan biosynthesis and glucose consumption [41]. Reactions in growth deplete available oxygen; high oxygen demand indicates an accelerated growth rate with high glucose consumption. As glucose is depleted, metabolism is reduced, allowing DO levels to increase; thus, glucose availability and growth rate can be indirectly controlled by monitoring DO levels [62]. In this strategy, when an upper limit of DO is reached, the system is fed with high glucose until DO reaches a lower limit, effectively avoiding glucose overfeeding and DO limitation. Limits are typically between 20-30%, as E. coli can suffer from glucose starvation when DO exceeds 40% saturation [64]. This control strategy can reduce acetate excretion and increase tryptophan and biomass production; however, the significant variation in glucose and DO throughout the fermentation can lead to glucose starvation and metabolic overflow with subsequent acetate production [64]. Despite this, one of the highest production titers in the TRTH strain, 59.55 g/L, was achieved following this strategy [43].
3.5.4. DO-Stat Control
DO-stat feeding can avoid substrate overfeeding and oxygen limitation [55,62,68]. When glucose is the limiting reagent, a DO level of 5% corresponds to 0.35 g/L residual glucose, while 50% corresponds to less than 0.10 g/L [62]. The DO-stat control strategy aims to maintain the DO level at a constant value by regulating the glucose feed so the glucose concentration and growth rate remain fixed; this strategy has been applied for phenylalanine production with almost zero acetate accumulation [62,68,85]. A commonly used value for DO in this type of fermentation is 20% [55,64].
Maintaining oxygen consumption at an optimal fixed value reduces the risks of glucose starvation and overfeeding. This strategy leads to low glucose concentrations by reducing acetate levels to less than 0.61 g/l. This strategy can achieve better results for biomass and tryptophan production than DO feedback control; regardless, it has a lower glucose-to-tryptophan conversion rate than E. coli mutants for the acetate production pathway [64].
3.5.5. DO Stage Control Strategy
DO-stat strategy can take advantage of the metabolic state of E. coli by using different DO levels for each fermentation stage [62]. The fermentation can be divided into two phases, focusing on biomass and tryptophan production. For the TRTH strain of E. coli, the highest tryptophan production is achieved with a DO level of 20% (0-20 h) and 30% (20-40 h); these conditions lead to reduce the accumulation of byproducts such as lactate, acetate and glutamate at the beginning of the fermentation allowing to produce more biomass and stimulate the transcription of genes necessary for the biosynthesis of tryptophan at the end of the fermentation when the biomass production is less active [41,62].
3.5.6. pH-stat Control
Glucose consumption and organic acid production by E. coli are linearly correlated. Likewise, the amount of alkali needed to neutralize these organic acids is proportional to the amount of glucose consumed. The pH-stat feed control strategy uses a feed solution where glucose and alkali are mixed to regulate the pH of the fermentation and maintain the glucose concentration in a stable state. The pump introducing this solution is activated when the pH is below a lower limit [61].
3.5.7 pH stage Control
As with the DO stage strategy, fermentation can be divided into phases to shift the range of optimal pH values, allowing for better tryptophan and biomass production [61]. Low pH in the initial fermentation phase reduces glycolysis activity, reducing acetate accumulation, glucose consumption, and growth rate. High pH increases the activity of enzymes involved in tryptophan production, but more ammonia is required to maintain the high pH. The supply of a more alkaline solution to raise the pH inhibits growth due to NH4+ accumulation, causing reduced energy efficiency [41].
The pH with the highest tryptophan and biomass production in the Dpta/pta-Y strain is pH 7.0 (0-20 h) and 6.5 (20-40 h), which leads to a high glucose conversion rate, producing 52.15 g/L of biomass and 52.57 g/L of tryptophan. The change in pH between the fermentation stages reduced the activity of the critical gene fbp, decreasing the flux through glycolysis and glucose consumption. In contrast, genes involved in the PPP (zwf, pgl, and araD) and tryptophan biosynthesis (pckA, ppsA, tktA, aroG, aroK, and trpEDBA) were more actively transcribed. Transcription of genes involved in acetate production (poxB, ackA, and tdcD) was decreased, and the acetate-consuming gene acs was more actively transcribed [41].
In the BCTRPG strain, fermentation was also divided into stages to adjust the pH, starting with low values and increasing them over time (6.5 from 0 to 12 h, 6.8 from 12 to 24 h, and 7.2 from 24 to 39 h). This strategy achieved higher tryptophan production than when a constant pH of 7 was used during fermentation [4].
3.6. Composition of the Culture Medium
The culture media used to produce tryptophan are very similar. Although the doses of the substrates vary, they are mainly composed of a source of PO4-, Mg2+, SO4-, NH4+, carbon, citrate, Fe, K+, vitamins, and trace elements.
3.6.1. Effect of calcium on fermentation
CaCO3 can be used as a buffer in fermentations for tryptophan production [40,67]. Ca2+ is a catalytic cofactor that activates specific metabolic pathways' enzymes, affecting the distribution of metabolic fluxes, cell growth, morphology, and accumulation of desired metabolites. CaCO3 has been observed to strengthen the TCA cycle and increase ATP production, which may benefit biomass production. The central genes CaCO3 activates are gltA, icd, and sucC, which belong to the TCA cycle [42]. Isocitrate dehydrogenase can complement the PPP in the production of NADPH by increasing its supply for amino acid production and biomass formation [23,42]. Activation of gltA has been reported to reduce acetate production in non-glucose-limited media [75].
3.6.2. Addition of Betaine Monohydrate
Betaine monohydrate is used as an osmoprotectant of bacterial cells, reverses the inhibitory effect of salts and urea on enzymes, protects proteins from chemical and thermal denaturation, is a donor of methyl groups, prevents caramelization of glucose, among other applications in fermentations [85,86]. An unexpected effect of betaine on E. coli fermentation is the activation of the transcription of the zwf gene, improving the activity of the glucose-6-phosphate dehydrogenase enzyme to produce NADPH, which is required as a cofactor to produce several metabolites, for example, serine production in the THRD strain increases when NADPH availability is high. This transcriptional activation is specific to the zwf gene and is not observed in the other central metabolic pathway genes gnd, icd, pfkA, or pykF [85].
High concentrations of tryptophan can be toxic to bacteria, accelerating their death phase. Intracellular concentrations of tryptophan in overproducing strains can exceed the concentration at which it crystallizes (11.4 g/L in water at 24°C) [31,59,67]. Adding betaine monohydrate to the culture medium could also protect E. coli from the effects of tryptophan [59].
Osmotically active solutes (osmolytes) such as betaine, proline, K+, glutamate, trehalose, or 3-(N-morpholino) propane sulfonate (MOPS) allow E. coli to incorporate free water to recover its cytoplasmic volume as an adaptation mechanism to osmotic stress conditions, avoiding dehydration. When betaine is present, E. coli uses it preferentially, replacing the other osmolytes, preventing K+ and MOPS from being taken from the medium and decreasing the amount of glutamate and trehalose synthesized. Betaine at 1 mM increases the growth rate of E. coli more efficiently than other osmoprotectants, such as proline [86]. Adding betaine could decrease the harmful effects of K+ accumulation under osmotic stress conditions [86], which can occur when KOH is used as an alkaline solution to regulate pH in fermentations.
3.6.3. Citrate supplementation
Adding sodium citrate enhances tryptophan production in E. coli, with an optimal concentration of 2 g/L improving tryptophan titers from 33.8 to 35.7 g/L in strain TRTH. Production of other metabolites in bacteria suggests sodium citrate increases Ca2+ uptake, inhibiting phosphofructokinase, and pyruvate kinase enzymes, resulting in less glucose being processed in glycolysis. Inhibition of pyruvate kinase increases the availability of the critical precursor phosphoenolpyruvate by reducing the production of pyruvate and acetyl-CoA; it also reduces the formation of acetate and lactate. The glycolysis is also weakened by increased activity of the competing PPP due to activation of the enzyme glucose-6-phosphate dehydrogenase; the oxidative branch of the PPP produces NADPH, which is required to produce tryptophan by balancing precursor availability [66].
Another pathway affected by citrate is the TCA cycle; it inhibits citrate synthase, reducing the incorporation of acetyl-CoA into the TCA cycle [66]. Decreased activity of the TCA cycle may prevent the accumulation of glutamate, which has been observed in some strains [59].
The Addition of citrate promotes the production of ATP, NADH, and NADPH, along with the reduction of byproducts, favoring the formation of biomass [66]. Adding 6 g/L of citric acid to the medium provides intermediates to reactivate the TCA cycle in the strain TRP04, which lacks the pykA and ppc genes [48].
3.6.4. Effect of Organic Nitrogen on Tryptophan Production
Yeast extract is a common ingredient of culture media, obtained from autolysis, hydrolysis, or plasmolysis, that serves as a source of organic nitrogen. Due to its complex composition, it provides amino acids, peptides, carbohydrates, nucleic acids, vitamins, and minerals for the growth of microorganisms [19]. Adding yeast extract to the culture medium can reduce acetate formation and improve protein production [66]. However, it has been reported that cultures on rich media based on acid-hydrolyzed casein stimulate tryptophanase activity [7].
Due to the complex composition of yeast extracts, levels of critical nutrients can vary widely between suppliers and even batches, affecting tryptophan production [19,56]. Impurities in yeast extract, such as pigments, proteins, and endotoxins, reduce tryptophan production [56].
Residual E. coli bacteria from fermentation can be chemically or enzymatically hydrolyzed to serve as an alternative organic nitrogen source to yeast extract. It provides amino acids, nucleotides, and other components at levels appropriate for E. coli growth while enhancing tryptophan production. However, rapid nitrogen uptake from the hydrolysate leads to acetate accumulation [56].
3.6.5. Inorganic Nitrogen Supply
The imbalance in the carbon-nitrogen ratio can reduce tryptophan production [63]. Ammonium assimilation in E. coli occurs through two assimilation pathways: the glutamine synthetase/glutamate synthase (GS/GOGAT) cycle and the glutamate dehydrogenase (GDH) pathway, which involve three key metabolites: glutamine, glutamate, and α-ketoglutarate [95].
For fermentations, a source of NH4+ is aqueous ammonia, the preferred alkaline solution to regulate pH. NH4+ has a high similarity to K+ in size and charge, allowing E. coli to exchange them in some cellular functions, reducing the importance of K+. This similarity allows NH4+ at high concentrations to be transported by the high-affinity potassium uptake system (Kpd); when NH4+ is incorporated into the metabolism, it dissociates from its proton, lowering the pH of the cytoplasm and the cell has to spend ATP to pump out protons in a futile cycle increasing the rate of oxygen consumption, decreasing the energetic efficiency of the bacteria and inhibiting their growth [3,55,96]. For optimal biomass and tryptophan production, the NH4+ concentration should be less than 120 mmol/L [3,41,60,61].
In modified strains that use plasmids to overexpress genes, the accumulation of NH4+ decreases tryptophan production because it decreases the stability of plasmids. The population of bacteria that have lost plasmids does not contribute to tryptophan production [60].
Although glutamine synthase (glnA) requires NH4+ to produce glutamine, its activity is regulated by several mechanisms, such as end-product feedback inhibition. At high ammonium concentrations, the adenylation of GlnA causes it to lose activity, turning it into a limiting reagent to produce tryptophan [74].
3.6.6. Increased Phosphate in the Medium
Phosphate is essential for cellular metabolism; it is part of ribonucleic acids and membrane phospholipids and is part of the intermediates of glycolysis and PPP [43]. Its importance makes phosphate a critical factor in the production of tryptophan and biomass; its participation in glycolysis causes its consumption to be proportional to that of glucose. In strain W3110-ZDrr, using 20 g/L of phosphate instead of 7.5 g/L increased tryptophan production from 15.8 to 26.8 g/L, but 25 g/L of phosphate resulted in growth inhibition [65].
For the TRTH strain, increasing KH2PO4 considerably boosted biomass production by increasing the initial growth rate. Tryptophan production also increased to 10 g/L of KH2PO4 and then decreased. Phosphate was consumed simultaneously as biomass production and was wholly depleted during fermentations. The increase in phosphate consumption caused an increase in glucose consumption and an improvement in glucose conversion rate to tryptophan. However, concentrations greater than 10 g/L facilitated the accumulation of acetic acid and lactic acid, exceeding 5 g/L and affecting growth, possibly due to a metabolic overload of the TCA cycle and the tryptophan pathway due to the high glycolysis activity. Due to its ubiquity, phosphate can affect the expression of plasmids in E. coli, affecting the expression of recombinant proteins [43]. The activity of tryptophan synthase is improved as the concentration of the phosphate buffer increases, and this buffer turns out to be more suitable than borate or Tris buffers [12].
3.6.7. The addition of Surfactants
Microorganisms use feedback inhibition as a mechanism to avoid producing more metabolites than required; intracellular concentrations of more than 4.5 g/L of tryptophan can affect its accumulation because it inhibits tryptophan synthase by up to 50% [67]; this enzyme can also be inhibited by indole [9]. Anthranilate synthase is another critical enzyme that is regulated by the negative feedback of tryptophan [39]; its inhibition when tryptophan reaches 30 g/L in strain AGX1757 favors the accumulation of indole [67], which inhibits the growth of E. coli at concentrations as low as 0.04% [12]. Blocking tryptophan and anthranilate synthases can redirect carbon flow toward phenylalanine and tyrosine synthesis [39].
The solubility of tryptophan in the culture medium is 22 g/L but increases to 32.1 g/L in the culture supernatant, possibly due to interaction with macromolecules secreted by E. coli [67]. Rationally engineered E. coli strains can accumulate over 40 g/L of tryptophan above the saturation concentration under typical culture conditions [35]. High intracellular concentrations of metabolites such as tryptophan can inhibit cell growth [17,24,30], which could be prevented by enhancing tryptophan export to reduce feedback inhibition [24,52].
Aromatic amino acids can diffuse through the cell membrane due to their hydrophobic properties [5,53]. Surfactants can affect membrane fluidity and permeability by facilitating the exchange of metabolites [9,65]. The surfactant Triton X-114 has been shown to enhance tryptophan production but is a poor candidate for use in fermentations because it generates large amounts of foam [65]. Triton X-100 served as an indole reservoir to reduce tryptophan synthase inhibition, boosting the production of the non-overproducing strain E. coli ATCC 11303 when cane molasses was used as a substrate [9]. Using Triton X-100 allowed production levels of 141.4 g/L to be reached in the E. coli B 10 strain from indole and serine. Another indole reservoir used to enhance tryptophan production is Amberlite XAD-2 because it does not encapsulate tryptophan [12].
Pluronic L-61 surfactant also enhances tryptophan production; its mechanism of action is through the precipitation of tryptophan in the medium when it reaches concentrations of 25 g/L by breaking its interaction with macromolecules that improve its solubility; this prevents the total inhibition of tryptophan synthase at 30 g/L, facilitating its secretion and recovery by centrifugation [65,67]. Tryptophan production in fed-batch fermentation improves as the Pluronic L-61 surfactant increases by up to 0.5% (V/V); higher concentrations inhibit growth. In discontinuous batch fermentations, the addition of this surfactant has no effect [65]. It is recommended to add Pluronic L-61 after the tryptophan reaches its solubility threshold to allow biomass accumulation [67].
Tween 60 also improves tryptophan production in fed-batch and batch fermentations. The mechanism of action seems related to the activation of transmembrane exporters that, by secreting tryptophan, reduce negative feedback. Tween 60 also promotes glutamate production. The improvement in tryptophan production with surfactants does not generalize to other amino acids; for example, it impairs the production of lysine, proline, leucine, valine, and the aromatic amino acids tyrosine and phenylalanine [65].
3.6.8. Accumulation of Sodium and Potassium Cations
K+ is a cofactor for many vital enzymes [3] and is the most critical cation for the growth of non-halophilic microbes; growth at low K+ concentrations requires high energy expenditure to maintain the ion gradient generated across the membrane when the optimal K+ concentration is reached inside the bacterium [96].
High concentrations of salt ions interact non-covalently with proteins and DNA, modifying the pH and ionic strength and adversely affecting enzyme activity [86,87]. Under conditions of osmotic stress, such as those present when tryptophan is produced in high concentrations, E. coli modifies its enzymatic activity and transport systems to accumulate ionic solutes such as K+, glutamate, and other anions that allow it to maintain the turgor of its membrane [88]. In minimal media, K+ plays the leading role in regulating osmolarity [86]; however, ionic solutes have adverse effects on growth, so they are only accumulated at low osmolarities, to be replaced at high osmolarities by compatible solutes that interfere little with most enzymes [88].
As osmolarity increases, K+ is neutralized by glutamate to form potassium glutamate, which serves as an activation signal for other osmotic stress response mechanisms [87]. Glutamate accumulation is probably due to the inhibition of glutamate-consuming pathways rather than an increase in glutamate synthesis [88]. As previously mentioned, glutamate accumulation hurts tryptophan production.
Alkaline solutions of NaOH and KOH that are commonly used to regulate the pH of fermentations release Na+ and K+ cations that affect nutrient uptake and growth when present above 40 mmol/L and inhibit biomass and tryptophan production when present above 60 and 78 mmol/L, respectively [61]. Purification and production costs of tryptophan can be impacted because the presence of K+ in the medium reduces the adsorption capacity for additional organic compounds in tryptophan synthesis [41]. To avoid high K+ concentrations, A mixture of KOH/NH3∙H2O in a 1:2 v/v ratio was used to control the pH, achieving high tryptophan titers [61].
3.6.9. Addition of Tryptophan Precursors in Media
The supply of precursors inside the cell is critical for amino acid productivity [71]. However, it is essential to control their feeding and production rate to avoid toxic effects due to the accumulation of metabolites. The supply of precursors must be balanced to achieve efficient production of tryptophan; the biosynthesis of 1 mol of tryptophan requires 1 mol of E4P and PEP as initial precursors and also consumes the equivalent of 1 mol of PEP in each glutamine, phosphoribosyl-5-pyrophosphate, and serine pathways, so favoring any of these pathways can unbalance the production of the other precursors [17]. For this reason, from 1 mol of glucose, 0.86 mol of DAHP can be produced, but only 0.45 mol of tryptophan can be produced [23].
The addition of precursors can reveal new bottlenecks; it has been described that the supply of anthranilate can cause an accumulation of indole when tryptophan accumulation is higher than 30 g/L because it inhibits the activity of the tryptophan synthase [41]. A higher production of tryptophan is associated with a lower accumulation of indole [78]. Adding serine to the medium did not improve tryptophan production because it was not a limiting reagent for tryptophan synthase [41].
Anthranilate is derived from chorismate and is a direct precursor of tryptophan; the supply of anthranilate improves the glucose conversion rate to tryptophan by preventing acetate production at high glucose concentrations up to 2.0 g/L. However, anthranilate can inhibit biomass production at concentrations higher than 0.2 g/L [64]. Anthranilate inhibits the growth of E. coli, possibly due to the blocking of tryptophan production by feed-forward inhibition of the enzyme IGPS since, in wild-type bacteria, adding tryptophan restores the growth [35].
3.6.10. Effect of Methionine on Tryptophan Production
The amino acid methionine is the precursor of S- adenosyl methionine (SAM), a methyl group donor involved in the biosynthesis of nucleic acids, phospholipids, proteins, and other molecules [42]. In tryptophan-producing strains, it can boost biomass synthesis [19]. Some publications use 1 g/L of methionine in the fermentation media [6,51]. For tryptophan production, the effect of methionine has yet to be studied in detail. However, adding methionine or glycine to the culture medium reduces acetate formation and improves protein production [66]. Adding methionine can alleviate the effects of acetate accumulation because its accumulation inhibits methionine synthesis, causing the accumulation of homocysteine, which is an inhibitor [68].
3.6.11. Effect of Carbon Sources
Most fermentations to produce tryptophan have been carried out efficiently using glucose, which is inexpensive but has the disadvantage of being obtained through agroindustrial processes, which compete with the food and feed industries with socioeconomic implications [15]. As an alternative, other carbon sources have been used, such as glycerol, which is obtained as a byproduct of biodiesel production, has a high reducing power, is inexpensive, and enters the cell through protein-assisted facilitated diffusion without consuming PEP [15,33,50].
Strategies implemented to utilize other carbon sources may vary due to changes in the distribution of metabolic fluxes; for example, glycerol is incorporated into cellular metabolism through gluconeogenesis [50]. When glycerol is used as a carbon source, and DAHP accumulates, or there is a phosphate deficiency, the highly toxic methylglyoxal is generated, an electrophile that reacts with proteins and DNA [15].
4. Models for Tryptophan Production
Constructing mathematical models from experimental observations and measurements facilitates studying biochemical processes. Simulating variations in these models can guide the selection of regulatory elements that allow the generation of stable tryptophan-producing strains [91,97]. The main mechanisms that regulate the genetic network for tryptophan production were discovered in the 1960s, and since then, they have become one of the most studied [8].
These models have been built using different approaches. Models based on Ordinary Differential Equations (ODE) are inherently quantitative and, therefore, require precise equations to describe biochemical reactions; the constants of these functions are difficult to obtain through experiments and can be inferred from mathematical techniques that adjust the equations to a defined behavior [8]. Models based on ODE for tryptophan production have been gradually developed by different research groups that incorporate various components and regulatory mechanisms, making them increasingly complex as the understanding of the functioning of the trp operon advances. [37], due to this complexity, the models must be solved numerically [98]. The representation of the tryptophan production models has focused mainly on the analysis of its synthesis through the trp operon, transport, and degradation [8]. The effect of central metabolism [23], the demand for tryptophan by the cell [16,97], or the impact of the availability of extracellular tryptophan [99] have also been added. Santillan and Mackey included the delay time and highlighted the importance of anthranilate synthase in the production of tryptophan [37]. The delay time between transcription and translation creates an oscillation in the transcription of the genes of the trp operon, which is not linear but exists in both on and off states; this oscillation is characteristic of repressed systems [100]. For the expression of the tna operon, it was also found that it has a bi-stable behavior that depends on the concentration of glucose and tryptophan; however, when the trp operon is included in the simulation, this behavior is no longer observed, possibly because the system enters in a state of homeostasis [8,101]. Xiu, Chang, and Zeng included attenuation in their model and the effect of the growth rate [100]. Sun and collaborators considered tryptophan secretion an essential factor in tryptophan accumulation and included the interaction between gene structure, regulatory elements and regulators [102]. Simulations in these models have found that feedback inhibition increases trp operon stability. However, several models omit it, while attenuation reduces the response time when the bacteria undergo a nutritional shift [98,102]. The ODE models have also been used to describe the behavior of strains that overproduce tryptophan [36].
Gene regulatory networks can be called Boolean networks, where nodes represent genes associated with Boolean values, 0 represents inactive genes and 1 represents active genes [8,101]. Changes in the values of Boolean networks represent their dynamics and are defined by Boolean functions. The order in which these values are updated is called the update scheme: they can be updated simultaneously, in sequential order, or asynchronously. It has been observed that each update scheme can give information about different properties of the network [101]. Boolean networks have the advantage of being wholly qualitative and do not require precise values to carry out their simulations [8].
A variant of Boolean networks is the threshold network, in which a weight is assigned to each of the Boolean functions of each node belonging to a directed graph [101].
Unlike Boolean representations, complex metabolic networks can be presented as qualitative models to take advantage of the large amount of scientific information. These models are known as Petri Nets (PNs), which facilitate the identification of attractors and the recognition of critical regulatory elements thanks to bioinformatics tools and techniques specific to PNs [102].
Machine Learning has gained momentum in recent years and permeated different branches of science. It can complement other tools, such as metabolic flux balance models, and incorporate experimental data, as is the case of the optimization of tryptophan production in yeast carried out by Zhang and collaborators [103]. Another model that uses Machine Learning is the QPAML classification model, which is fed by metabolic fluxes generated by introducing perturbations in a metabolic network [54].
5. Conclusions
Optimizing tryptophan production in Escherichia coli through fermentation requires a multifaceted approach that integrates genetic, metabolic, and environmental factors. This review highlights that precise control over nutrients, pH, temperature, and dissolved oxygen (DO) is crucial for maximizing production efficiency. The balance between growth rate and substrate consumption is essential, as excessive feed rates or poor oxygen control can lead to byproduct accumulation, reducing overall yields. Key strategies include adjusting glucose feed rates to avoid metabolic overflow, managing nitrogen and phosphate levels to support biomass and tryptophan production, and maintaining optimal pH and temperature conditions. Additionally, supplements like betaine and surfactants enhance production by reducing stress and increasing precursor availability. Combining these fermentation strategies with genetic modifications that enhance the biosynthetic pathway makes it possible to achieve high-density cultures capable of producing significant tryptophan titers. Continued research in fine-tuning these parameters will further improve microbial tryptophan production's economic viability and scalability for industrial applications.
Author Contributions
Conceptualization, M.A.R.-V. and A.M.-A.; methodology, M.A.R.-V.; software, M.A.R.-V.; validation, M.A.R.-V. and A.M.-A.; formal analysis, M.A.R.-V.; investigation, M.A.R.-V.; resources, A.M.-A.; data curation, M.A.R.-V.; writing—original draft preparation, M.A.R.-V.; writing—review and editing, A.M.-A.; visualization, M.A.R.-V.; supervision, A.M.-A.; project administration, A.M.-A.; funding acquisition, A.M.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This research and APC were funded by CONAHCYT “Paradigmas de la Ciencia” grant number 319732 and “Infrastructure” grant number 317147, given to A.M.-A.; M.A.R.-V. was a recipient of CONAHCYT Ph.D. fellowship no. 486528.
Acknowledgments
The authors thank Alejandro Coreño-Alonso and Diego A. Castro López, who contributed to a previous version of this review, and Mario Alberto Pantoja Alonso for discussing the theme.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mohamed, H.; El Nady, G.; Ali, A.; Abdel-Razik, A.; Ibrahim, S. Production of L-Tryptophan by Mutants of Corynebacterium Glutamicum. Arab Universities Journal of Agricultural Sciences 2018, 26, 1187–2018. [Google Scholar] [CrossRef]
- Richard, D.M.; Dawes, M.A.; Mathias, C.W.; Acheson, A.; Hill-Kapturczak, N.; Dougherty, D.M. L-Tryptophan: Basic Metabolic Functions, Behavioral Research and Therapeutic Indications. International Journal of Tryptophan Research 2009, 2, IJTR–S2129. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Xu, Q.; Liang, J.; Xie, X.; Zhang, C.; Chen, N. Control Strategy of Specific Growth Rate in L-Tryptophan Production by Escherichia Coli. In; 2014; pp. 241–249.
- Lu, N.; Zhang, B.; Cheng, L.; Wang, J.; Zhang, S.; Fu, S.; Xiao, Y.; Liu, H. Gene Modification of Escherichia Coli and Incorporation of Process Control to Decrease Acetate Accumulation and Increase ʟ-Tryptophan Production. Ann Microbiol 2017, 67, 567–576. [Google Scholar] [CrossRef]
- Liu, S.; Wang, B.-B.; Xu, J.-Z.; Zhang, W.-G. Engineering of Shikimate Pathway and Terminal Branch for Efficient Production of L-Tryptophan in Escherichia Coli. Int J Mol Sci 2023, 24, 11866. [Google Scholar] [CrossRef]
- Xiong, B.; Zhu, Y.; Tian, D.; Jiang, S.; Fan, X.; Ma, Q.; Wu, H.; Xie, X. Flux Redistribution of Central Carbon Metabolism for Efficient Production of L-tryptophan in Escherichia Coli. Biotechnol Bioeng 2021, 118, 1393–1404. [Google Scholar] [CrossRef]
- Yanofsky, C.; Horn, V.; Gollnick, P. Physiological Studies of Tryptophan Transport and Tryptophanase Operon Induction in Escherichia Coli. J Bacteriol 1991, 173, 6009–6017. [Google Scholar] [CrossRef]
- Deal, I.; Macauley, M.; Davies, R. Boolean Models of the Transport, Synthesis, and Metabolism of Tryptophan in Escherichia Coli. Bull Math Biol 2023, 85, 29. [Google Scholar] [CrossRef]
- Sadeghiyan-Rizi, T.; Fooladi, J.; Sadrai, S. Preliminary Study on Cost-Effective L-Tryptophan Production from Indole and L-Serine by E. Coli Cells. Avicenna J Med Biotechnol 2016, 8, 188–192. [Google Scholar]
- Ding, D.; Bai, D.; Li, J.; Mao, Z.; Zhu, Y.; Liu, P.; Lin, J.; Ma, H.; Zhang, D. Analyzing the Genetic Characteristics of a Tryptophan-Overproducing Escherichia Coli. Bioprocess Biosyst Eng 2021, 44, 1685–1697. [Google Scholar] [CrossRef]
- Minliang, C.; Chengwei, M.; Lin, C.; Zeng, A.-P. Integrated Laboratory Evolution and Rational Engineering of GalP/Glk-Dependent Escherichia Coli for Higher Yield and Productivity of L-Tryptophan Biosynthesis. Metab Eng Commun 2021, 12, e00167. [Google Scholar] [CrossRef]
- Bang, W.; Lang, S.; Sahm, H.; Wagner, F. Production L-tryptophan by Escherichia Coli Cells. Biotechnol Bioeng 1983, 25, 999–1011. [Google Scholar] [CrossRef] [PubMed]
- Gang, S.; Sharma, S.; Saraf, M.; Buck, M.; Schumacher, J. Analysis of Indole-3-Acetic Acid (IAA) Production in Klebsiella by LC-MS/MS and the Salkowski Method. Bio Protoc 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Sano, K.; Yokozeki, K.; Eguchi, C.; Kagawa, T.; Noda, I.; Mitsugi, K. Enzymatic Production of L-Tryptophan from l- and Dl-5-Indolyl-Methylhydantoin by Newly Isolated Bacterium. Agric Biol Chem 1977, 41, 819–825. [Google Scholar] [CrossRef]
- Schoppel, K.; Trachtmann, N.; Korzin, E.J.; Tzanavari, A.; Sprenger, G.A.; Weuster-Botz, D. Metabolic Control Analysis Enables Rational Improvement of E. Coli L-Tryptophan Producers but Methylglyoxal Formation Limits Glycerol-Based Production. Microb Cell Fact 2022, 21, 201. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Z.-L.; Zeng, A.-P.; Deckwer, W.-D. Model Analysis Concerning the Effects of Growth Rate and Intracellular Tryptophan Level on the Stability and Dynamics of Tryptophan Biosynthesis in Bacteria. J Biotechnol 1997, 58, 125–140. [Google Scholar] [CrossRef]
- Ikeda, M. Towards Bacterial Strains Overproducing L-Tryptophan and Other Aromatics by Metabolic Engineering. Appl Microbiol Biotechnol 2006, 69, 615–626. [Google Scholar] [CrossRef]
- Roy, A. Enzymatic Production of L-Tryptophan by a Thermophilic Strain of Bacillus Licheniformis Isolated from a Local Hot Spring of Paniphala, Asansol Area of West Bengal. J Pure Appl Microbiol 2023, 17, 2307–2315. [Google Scholar] [CrossRef]
- Chen, X.; Jin, Z.W.; Li, X.; Wang, Z. Effect of Yeast Extract on L-Tryptophan Production in Batch Culture of Escherichia Coli. Adv Mat Res 2013, 807–809, 2009–2013. [Google Scholar] [CrossRef]
- Varma, A.; Boesch, B.W.; Palsson, B.O. Biochemical Production Capabilities of Escherichia Coli. Biotechnol Bioeng 1993, 42, 59–73. [Google Scholar] [CrossRef]
- Tribe, D.E.; Pittard, J. Hyperproduction of Tryptophan by Escherichia Coli: Genetic Manipulation of the Pathways Leading to Tryptophan Formation. Appl Environ Microbiol 1979, 38, 181–190. [Google Scholar] [CrossRef]
- Niu, H.; Li, R.; Liang, Q.; Qi, Q.; Li, Q.; Gu, P. Metabolic Engineering for Improving <scp>l</Scp> -Tryptophan Production in Escherichia Coli. J Ind Microbiol Biotechnol 2019, 46, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.W.; Mauch, K.; Reuss, M.; Gilles, E.D.; Kremling, A. Metabolic Design Based on a Coupled Gene Expression—Metabolic Network Model of Tryptophan Production in Escherichia Coli. Metab Eng 2004, 6, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Ding, S.; Liu, Y.; Gao, C.; Hu, G.; Song, W.; Liu, J.; Chen, X.; Liu, L. Enhancing Tryptophan Production by Balancing Precursors in Escherichia Coli. Biotechnol Bioeng 2022, 119, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Mukhopadhyay, S.K. L-Tryptophan Production by Auxotrophic and Analogue Resistant Mutants of Aureobacterium Flavescens. International Journal of Tryptophan Research 2011, 4, IJTR–S5984. [Google Scholar] [CrossRef] [PubMed]
- Dodge, T.C.; Gerstner, J.M. Optimization of the Glucose Feed Rate Profile for the Production of Tryptophan from Recombinant E Coli. Journal of Chemical Technology & Biotechnology 2002, 77, 1238–1245. [Google Scholar] [CrossRef]
- Aiba, S.; Tsunekawa, H.; Imanaka, T. New Approach to Tryptophan Production by Escherichia Coli: Genetic Manipulation of Composite Plasmids in Vitro. Appl Environ Microbiol 1982, 43, 289–297. [Google Scholar] [CrossRef]
- Malin, B.; Westhead, J. Production of L-tryptophan in Submerged Culture. Journal of Biochemical and Microbiological Technology and Engineering 1959, 1, 49–57. [Google Scholar] [CrossRef]
- Deshpande, A.; Vue, J.; Morgan, J. Combining Random Mutagenesis and Metabolic Engineering for Enhanced Tryptophan Production in Synechocystis Sp. Strain PCC 6803. Appl Environ Microbiol 2020, 86. [Google Scholar] [CrossRef]
- Liu, S.; Xu, J.-Z.; Zhang, W.-G. Advances and Prospects in Metabolic Engineering of Escherichia Coli for L-Tryptophan Production. World J Microbiol Biotechnol 2022, 38, 22. [Google Scholar] [CrossRef]
- Ikeda, M.; Nakanishi, K.; Kino, K.; Katsumata, R. Fermentative Production of Tryptophan by a Stable Recombinant Strain of Coryne-Bacterium Glutamicum with a Modified Serine-Biosynthetic Pathway. Biosci Biotechnol Biochem 1994, 58, 674–678. [Google Scholar] [CrossRef]
- Plachý, J.; Ulbert, S. Production of L-tryptophan. Acta Biotechnol 1990, 10, 517–522. [Google Scholar] [CrossRef]
- Liu, L.; Chen, S.; Wu, J. Phosphoenolpyruvate:Glucose Phosphotransferase System Modification Increases the Conversion Rate during <scp>l</Scp> -Tryptophan Production in Escherichia Coli. J Ind Microbiol Biotechnol 2017, 44, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Berry, A. Improving Production of Aromatic Compounds in Escherichia Coli by Metabolic Engineering. Trends Biotechnol 1996, 14, 250–256. [Google Scholar] [CrossRef]
- Chen, L.; Chen, M.; Ma, C.; Zeng, A.-P. Discovery of Feed-Forward Regulation in L-Tryptophan Biosynthesis and Its Use in Metabolic Engineering of E. Coli for Efficient Tryptophan Bioproduction. Metab Eng 2018, 47, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Castro-López, D.A.; González de la Vara, L.E.; Santillán, M.; Martínez-Antonio, A. A Molecular Dynamic Model of Tryptophan Overproduction in Escherichia Coli. Fermentation 2022, 8, 560. [Google Scholar] [CrossRef]
- Santillán, M.; Mackey, M.C. Dynamic Regulation of the Tryptophan Operon: A Modeling Study and Comparison with Experimental Data. Proceedings of the National Academy of Sciences 2001, 98, 1364–1369. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Cheng, L.; Wang, J.; Shen, Z.; Chen, N. Impact of Deletion of the Genes Encoding Acetate Kinase on Production of L-Tryptophan by Escherichia Coli. Ann Microbiol 2016, 66, 261–269. [Google Scholar] [CrossRef]
- Zhao, Z.-J.; Zou, C.; Zhu, Y.-X.; Dai, J.; Chen, S.; Wu, D.; Wu, J.; Chen, J. Development of L-Tryptophan Production Strains by Defined Genetic Modification in Escherichia Coli. J Ind Microbiol Biotechnol 2011, 38, 1921–1929. [Google Scholar] [CrossRef]
- Liu, L.; Duan, X.; Wu, J. L-Tryptophan Production in Escherichia Coli Improved by Weakening the Pta-AckA Pathway. PLoS One 2016, 11, e0158200. [Google Scholar] [CrossRef]
- Zhao, C.; Fang, H.; Wang, J.; Zhang, S.; Zhao, X.; Li, Z.; Lin, C.; Shen, Z.; Cheng, L. Application of Fermentation Process Control to Increase <scp>l</Scp> -tryptophan Production in <scp> Escherichia Coli </Scp>. Biotechnol Prog 2020, 36. [Google Scholar] [CrossRef]
- Zhou, H.-Y.; Wu, W.-J.; Xu, Y.-Y.; Zhou, B.; Niu, K.; Liu, Z.-Q.; Zheng, Y.-G. Calcium Carbonate Addition Improves L-Methionine Biosynthesis by Metabolically Engineered Escherichia Coli W3110-BL. Front Bioeng Biotechnol 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiong, H.; Xu, Q. L-Tryptophan Fermentation by High Cell Density Culture of Escherichia Coli. Food and Fermentation Industries 2019, 45, 15–20. [Google Scholar]
- Mears, L.; Stocks, S.M.; Sin, G.; Gernaey, K. V. A Review of Control Strategies for Manipulating the Feed Rate in Fed-Batch Fermentation Processes. J Biotechnol 2017, 245, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Panichkin, V.B.; Livshits, V.A.; Biryukova, I. V.; Mashko, S. V. Metabolic Engineering of Escherichia Coli for L-Tryptophan Production. Appl Biochem Microbiol 2016, 52, 783–809. [Google Scholar] [CrossRef]
- Liu, L.; Bilal, M.; Luo, H.; Zhao, Y.; Iqbal, H.M.N. Metabolic Engineering and Fermentation Process Strategies for L-Tryptophan Production by Escherichia Coli. Processes 2019, 7. [Google Scholar] [CrossRef]
- Du, L.; Zhang, Z.; Xu, Q.; Chen, N. New Strategy for Removing Acetic Acid as a By-Product during L-Tryptophan Production. Biotechnology & Biotechnological Equipment 2019, 33, 1471–1480. [Google Scholar] [CrossRef]
- Du, L.; Zhang, Z.; Xu, Q.; Chen, N. Central Metabolic Pathway Modification to Improve L-Tryptophan Production in Escherichia Coli. Bioengineered 2019, 10, 59–70. [Google Scholar] [CrossRef]
- Liu, L.; Duan, X.; Wu, J. Modulating the Direction of Carbon Flow in Escherichia Coli to Improve l -Tryptophan Production by Inactivating the Global Regulator FruR. J Biotechnol 2016, 231, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Tröndle, J.; Schoppel, K.; Bleidt, A.; Trachtmann, N.; Sprenger, G.A.; Weuster-Botz, D. Metabolic Control Analysis of L-Tryptophan Production with Escherichia Coli Based on Data from Short-Term Perturbation Experiments. J Biotechnol 2020, 307, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Pan, X.; Yang, T.; You, J.; Zhu, R.; Yang, T.; Zhang, X.; Xu, M.; Rao, Z. Multidimensional Engineering of Escherichia Coli for Efficient Synthesis of L-Tryptophan. Bioresour Technol 2023, 386, 129475. [Google Scholar] [CrossRef]
- Gu, P.; Yang, F.; Li, F.; Liang, Q.; Qi, Q. Knocking out Analysis of Tryptophan Permeases in Escherichia Coli for Improving L-Tryptophan Production. Appl Microbiol Biotechnol 2013, 97, 6677–6683. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Cheng, Y.; Xie, X.; Xu, Q.; Chen, N. Modification of Tryptophan Transport System and Its Impact on Production of L-Tryptophan in Escherichia Coli. Bioresour Technol 2012, 114, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Valdovinos, M.A.; Salas-Navarrete, P.C.; Amores, G.R.; Hernández-Orihuela, A.L.; Martínez-Antonio, A. Qualitative Perturbation Analysis and Machine Learning: Elucidating Bacterial Optimization of Tryptophan Production. Algorithms 2024, 17, 282. [Google Scholar] [CrossRef]
- Wang, J.; Huang, J.; Shi, J.; Xu, Q.; Xie, X.; Chen, N. Fermentation Characterization of an L-Tryptophan Producing Escherichia Coli Strain with Inactivated Phosphotransacetylase. Ann Microbiol 2013, 63, 1219–1224. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Z.; Liu, Z.; Xu, Q. Using Enzymatic Hydrolyzate as New Nitrogen Source for L-Tryptophan Fermentation by E.Coli. Bioengineered 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Bandiera, M.; Morpurgo, G.; Ricci, R. Tryptophan Production by Mutant Strains of Escherichia Coli K 12. Experientia 1967, 23, 724–725. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chen, L.; Zeng, A.-P. CRISPR/Cas9-Facilitated Engineering with Growth-Coupled and Sensor-Guided in Vivo Screening of Enzyme Variants for a More Efficient Chorismate Pathway in E. Coli. Metab Eng Commun 2019, 9, e00094. [Google Scholar] [CrossRef]
- Chen, L.; Zeng, A.-P. Rational Design and Metabolic Analysis of Escherichia Coli for Effective Production of L-Tryptophan at High Concentration. Appl Microbiol Biotechnol 2017, 101, 559–568. [Google Scholar] [CrossRef]
- Xu, Q.; Bai, F.; Chen, N.; Bai, G. Removing the By-Products Acetic Acid and NH 4 + from the l -Tryptophan Broth by Vacuum Thin Film Evaporation during l -Tryptophan Production. Electronic Journal of Biotechnology 2018, 33, 46–51. [Google Scholar] [CrossRef]
- Cheng, L.-K.; Wang, J.; Xu, Q.-Y.; Zhao, C.-G.; Shen, Z.-Q.; Xie, X.-X.; Chen, N. Strategy for PH Control and PH Feedback-Controlled Substrate Feeding for High-Level Production of l-Tryptophan by Escherichia Coli. World J Microbiol Biotechnol 2013, 29, 883–890. [Google Scholar] [CrossRef]
- Zhao, C.; Cheng, L.; Xu, Q.; Wang, J.; Shen, Z.; Chen, N. Improvement of the Production of L-Tryptophan in Escherichia Coli by Application of a Dissolved Oxygen Stage Control Strategy. Ann Microbiol 2016, 66, 843–854. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y.; Ding, D.; Cong, L.; Zhang, D. Rational Design and Analysis of an Escherichia Coli Strain for High-Efficiency Tryptophan Production. J Ind Microbiol Biotechnol 2018, 45, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Jing, K.; Tang, Y.; Yao, C.; del Rio-Chanona, E.A.; Ling, X.; Zhang, D. Overproduction of L-tryptophan via Simultaneous Feed of Glucose and Anthranilic Acid from Recombinant Escherichia Coli W3110: Kinetic Modeling and Process Scale-up. Biotechnol Bioeng 2018, 115, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Huang, J.; Zhu, X.; Huang, L.; Cai, J.; Xu, Z. Enhanced Production of L-Tryptophan with Glucose Feeding and Surfactant Addition and Related Metabolic Flux Redistribution in the Recombinant Escherichia Coli. Food Sci Biotechnol 2013, 22, 207–214. [Google Scholar] [CrossRef]
- Xu, Q.; Cheng, L.; Xie, X.; Zhang, C.; Li, Y.; Ning, C. Effect of Sodium Citrate on L-Tryptophan Fermentation by Escherichia Coli. In; 2015; pp. 335–342.
- Azuma, S.; Tsunekawa, H.; Okabe, M.; Okamoto, R.; Aiba, S. Hyper-Production of l-Trytophan via Fermentation with Crystallization. Appl Microbiol Biotechnol 1993, 39, 471–476. [Google Scholar] [CrossRef]
- Cheng, L.-K.; Wang, J.; Xu, Q.-Y.; Xie, X.-X.; Zhang, Y.-J.; Zhao, C.-G.; Chen, N. Effect of Feeding Strategy on L-Tryptophan Production by Recombinant Escherichia Coli. Ann Microbiol 2012, 62, 1625–1634. [Google Scholar] [CrossRef]
- Gu, P.; Yang, F.; Kang, J.; Wang, Q.; Qi, Q. One-Step of Tryptophan Attenuator Inactivation and Promoter Swapping to Improve the Production of L-Tryptophan in Escherichia Coli. Microb Cell Fact 2012, 11, 30. [Google Scholar] [CrossRef]
- Xu, Q.; Bai, F.; Chen, N.; Bai, G. Gene Modification of the Acetate Biosynthesis Pathway in Escherichia Coli and Implementation of the Cell Recycling Technology to Increase L-Tryptophan Production. PLoS One 2017, 12, e0179240. [Google Scholar] [CrossRef]
- Shen, T.; Liu, Q.; Xie, X.; Xu, Q.; Chen, N. Improved Production of Tryptophan in Genetically Engineered Escherichia Coli with TktA and PpsA Overexpression. J Biomed Biotechnol 2012, 2012, 605219. [Google Scholar] [CrossRef]
- Li, G.; Young, K.D. Indole Production by the Tryptophanase TnaA in Escherichia Coli Is Determined by the Amount of Exogenous Tryptophan. Microbiology (N Y) 2013, 159, 402–410. [Google Scholar] [CrossRef]
- von Kamp, A.; Klamt, S. Growth-Coupled Overproduction Is Feasible for Almost All Metabolites in Five Major Production Organisms. Nat Commun 2017, 8, 15956. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ding, D.; Wang, H.; Liu, L.; Fang, H.; Chen, T.; Zhang, D. Engineering Escherichia Coli to Improve Tryptophan Production via Genetic Manipulation of Precursor and Cofactor Pathways. Synth Syst Biotechnol 2020, 5, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Gecse, G.; Labunskaite, R.; Pedersen, M.; Kilstrup, M.; Johanson, T. Minimizing Acetate Formation from Overflow Metabolism in Escherichia Coli: Comparison of Genetic Engineering Strategies to Improve Robustness toward Sugar Gradients in Large-Scale Fermentation Processes. Front Bioeng Biotechnol 2024, 12. [Google Scholar] [CrossRef]
- Chang, D.-E.; Jung, H.-C.; Rhee, J.-S.; Pan, J.-G. Homofermentative Production of D- or L-Lactate in Metabolically Engineered Escherichia Coli RR1. Appl Environ Microbiol 1999, 65, 1384–1389. [Google Scholar] [CrossRef]
- Wray, L. V.; Fisher, S.H. Functional Roles of the Conserved Glu304 Loop of Bacillus Subtilis Glutamine Synthetase. J Bacteriol 2010, 192, 5018–5025. [Google Scholar] [CrossRef]
- Ashworth, D.J.; Chen, C.S.; Mascarenhas, Desmond. Direct Observation of Tryptophan Biosynthesis in Escherichia Coli by Carbon-13 Nuclear Magnetic Resonance Spectroscopy. Anal Chem 1986, 58, 526–532. [Google Scholar] [CrossRef]
- Schütze, A.; Benndorf, D.; Püttker, S.; Kohrs, F.; Bettenbrock, K. The Impact of AckA, Pta, and AckA-Pta Mutations on Growth, Gene Expression and Protein Acetylation in Escherichia Coli K-12. Front Microbiol 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Phue, J.-N.; Shiloach, J. Impact of Dissolved Oxygen Concentration on Acetate Accumulation and Physiology of E. Coli BL21, Evaluating Transcription Levels of Key Genes at Different Dissolved Oxygen Conditions. Metab Eng 2005, 7, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Jung, J.; Jang, I.-A.; Madsen, E.L.; Park, W. Role of Glyoxylate Shunt in Oxidative Stress Response. Journal of Biological Chemistry 2016, 291, 11928–11938. [Google Scholar] [CrossRef]
- Yang, P.; Liu, W.; Chen, Y.; Gong, A.-D. Engineering the Glyoxylate Cycle for Chemical Bioproduction. Front Bioeng Biotechnol 2022, 10. [Google Scholar] [CrossRef]
- Rui, B.; Shen, T.; Zhou, H.; Liu, J.; Chen, J.; Pan, X.; Liu, H.; Wu, J.; Zheng, H.; Shi, Y. A Systematic Investigation of Escherichia Coli Central Carbon Metabolism in Response to Superoxide Stress. BMC Syst Biol 2010, 4, 122. [Google Scholar] [CrossRef] [PubMed]
- Millard, P.; Enjalbert, B.; Uttenweiler-Joseph, S.; Portais, J.-C.; Létisse, F. Control and Regulation of Acetate Overflow in Escherichia Coli. Elife 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, D.; Cai, N.; Han, C.; Mao, Q.; Wang, T.; Zhou, Q.; Chen, N.; Xie, X. Betaine Supplementation Improved L-Threonine Fermentation of Escherichia Coli THRD by Upregulating Zwf (Glucose-6-Phosphate Dehydrogenase) Expression. Electronic Journal of Biotechnology 2019, 39, 67–73. [Google Scholar] [CrossRef]
- Cayley, S.; Lewis, B.A.; Record, M.T. Origins of the Osmoprotective Properties of Betaine and Proline in Escherichia Coli K-12. J Bacteriol 1992, 174, 1586–1595. [Google Scholar] [CrossRef]
- Gauthier, M.J.; Flatau, G.N.; Le Rudulier, D.; Clément, R.L.; Combarro Combarro, M.P. Intracellular Accumulation of Potassium and Glutamate Specifically Enhances Survival of Escherichia Coli in Seawater. Appl Environ Microbiol 1991, 57, 272–276. [Google Scholar] [CrossRef]
- McLaggan, D.; Naprstek, J.; Buurman, E.T.; Epstein, W. Interdependence of K+ and Glutamate Accumulation during Osmotic Adaptation of Escherichia Coli. Journal of Biological Chemistry 1994, 269, 1911–1917. [Google Scholar] [CrossRef]
- Tazuya-Murayama, K.; Aramaki, H.; Mishima, M.; Saito, K.; Ishida, S.; Yamada, K. Effect of L-Serine on the Biosynthesis of Aromatic Amino Acids in Escherichia Coli. J Nutr Sci Vitaminol (Tokyo) 2006, 52, 256–260. [Google Scholar] [CrossRef]
- Tran, K.-N.T.; Eom, G.T.; Hong, S.H. Improving L-Serine Production in Escherichia Coli via Synthetic Protein Scaffold of SerB, SerC, and EamA. Biochem Eng J 2019, 148, 138–142. [Google Scholar] [CrossRef]
- Sinha, S. Theoretical Study of Tryptophan Operon: Application in Microbial Technology. Biotechnol Bioeng 1988, 31, 117–124. [Google Scholar] [CrossRef]
- Dwivedi, C.P.; Imanaka, T.; Aiba, S. Instability of Plasmid-harboring Strain of E. Coli in Continuous Culture. Biotechnol Bioeng 1982, 24, 1465–1468. [Google Scholar] [CrossRef]
- Kumar, R.; Shimizu, K. Metabolic Regulation of Escherichia Coli and Its GdhA, GlnL, GltB, D Mutants under Different Carbon and Nitrogen Limitations in the Continuous Culture. Microb Cell Fact 2010, 9, 8. [Google Scholar] [CrossRef]
- Li, X.; Kang, Y.; Yu, C.; Dai, J.; Wang, Z.; Li, Z.; Yao, J.; Li, P.; Zheng, G.; Chen, X. Exponential Feeding Strategy of High-Density Cultivation of a Salt-Tolerant Aroma-Producing Yeast Zygosaccharomyces Rouxii in Stirred Fermenter. Biochem Eng J 2016, 111, 18–23. [Google Scholar] [CrossRef]
- Yuan, J.; Doucette, C.D.; Fowler, W.U.; Feng, X.; Piazza, M.; Rabitz, H.A.; Wingreen, N.S.; Rabinowitz, J.D. Metabolomics-driven Quantitative Analysis of Ammonia Assimilation in E. Coli. Mol Syst Biol 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Buurman, EdT. ; Joost Teixeira de Mattos, M.; Neijssel, OenseM. Futile Cycling of Ammonium Ions via the High Affinity Potassium Uptake System (Kdp) of Escherichia Coli. Arch Microbiol 1991, 155. [Google Scholar] [CrossRef]
- Sen, A.K.; Liu, W. Dynamic Analysis of Genetic Control and Regulation of Amino Acid Synthesis: The Tryptophan Operon in Escherichia Coli. Biotechnol Bioeng 1990, 35, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Santillán, M.; Zeron, E.S. Dynamic Influence of Feedback Enzyme Inhibition and Transcription Attenuation on the Tryptophan Operon Response to Nutritional Shifts. J Theor Biol 2004, 231, 287–298. [Google Scholar] [CrossRef]
- Bhartiya, S.; Rawool, S.; Venkatesh, K. V. Dynamic Model of Escherichia Coli Tryptophan Operon Shows an Optimal Structural Design. Eur J Biochem 2003, 270, 2644–2651. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Z.; Chang, Z.; Zeng, A. Nonlinear Dynamics of Regulation of Bacterial Trp Operon: Model Analysis of Integrated Effects of Repression, Feedback Inhibition, and Attenuation. Biotechnol Prog 2002, 18, 686–693. [Google Scholar] [CrossRef]
- Orozco-Gómez, D.I.; Sosa-Hernández, J.E.; Gallardo-Navarro, Ó.A.; Santana-Solano, J.; Santillán, M. Bistable Behaviour and Medium-Dependent Post-Translational Regulation of the Tryptophanase Operon Regulatory Pathway in Echerichia Coli. Sci Rep 2019, 9, 5451. [Google Scholar] [CrossRef]
- Sun, Y.; Mu, X.; Li, Z.; Teng, H.; Xiu, Z. Robustness and Nonlinear Dynamic Analysis for Trp Operon and Optimization of Tryptophan Production: An Integrated Model Considering Gene Regulation, Genes Interaction and Product Excretion. In Proceedings of the 2010 4th International Conference on Bioinformatics and Biomedical Engineering; IEEE, June 2010; pp. 1–7. [Google Scholar]
- Zhang, J.; Petersen, S.D.; Radivojevic, T.; Ramirez, A.; Pérez-Manríquez, A.; Abeliuk, E.; Sánchez, B.J.; Costello, Z.; Chen, Y.; Fero, M.J.; et al. Combining Mechanistic and Machine Learning Models for Predictive Engineering and Optimization of Tryptophan Metabolism. Nat Commun 2020, 11, 4880. [Google Scholar] [CrossRef]
Figure 1.
Gene modifications used to increase tryptophan production in Escherichia coli. Genes deleted or attenuated are shown in orange, overexpressed genes in light green, and genes mutated to be insensitive to feedback inhibition are shown in dark green. Heterologous genes have a subscript that identifies the species from which they were obtained: Corynebacterium glutamicum (Cg), Aspergillus niger (An), Zymomonas mobilis (Zm), Bacillus subtilis (Bs), and Bifidobacterium adolescentis (Ba). Abbreviations for genes and reactions are: high-affinity ATP-driven potassium transport system (kdp), ATP-depending proton transporter (PT), citrate transporter (citT), tryptophan permease (tnaB), aromatic amino acid transport protein (aroP), tryptophan-specific transport protein (mtr), aromatic amino acid exporter (yddG), galactose permease (galP), glucose facilitator from Zymomonas mobilis (glfZm), glucokinase (glk), enzyme IIBC of PTS (ptsG), enzyme IIAGlc of PTS (crr), phosphocarrier protein HPr (ptsH), enzyme I of PTS (ptsI), glucose-6-phosphate isomerase (pgi), 6-phosphofructokinase (pfkA), fructose-1,6-bisphosphatase (fbp), fructose-bisphosphate aldolase (fba), triose-phosphate isomerase (tpiA), glyceraldehyde-3-phosphate dehydrogenase (gapA), phosphoglycerate kinase (pgk), phosphoglycerate mutase (gpmA), enolase (eno), pyruvate oxidase (poxB), pyruvate kinase 1 (pykF), pyruvate kinase 2 (pykA), phosphoenolpyruvate synthetase (ppsA), pyruvate dehydrogenase complex (PDH), phosphate acetyltransferase (pta), acetate kinase (ackA), propionate/acetate kinase (tdcD), acetyl-CoA synthetase (acs), phosphoenolpyruvate carboxylase (ppc), acetaldehyde-CoA dehydrogenase/alcohol dehydrogenase (adhE), citrate synthase (gltA), aconitate hydratase A (acnA), aconitate hydratase B (acnB), isocitrate dehydrogenase (icd), 2-ketoglutarate dehydrogenase complex (AKGDH), succinyl-CoA synthetase (sucCD), succinate:quinone oxidoreductase complex (sdhABCD), fumarase A (fumA), fumarase B (fumB), fumarase C (fumC), pyruvate carboxylase from Corynebaterium glutamicum (pycCg), phosphenolpyruvate carboxykinase (pckA), malate dehydrogenase (mdh), NAD-dependent malic enzyme (maeA), isocitrate lyase (aceA), malate synthase A (aceB), D-lactate dehydrogenase (ldhA), pyruvate formate-lyase (pflB), glutamate dehydrogenase (gdhA), glutamine synthetase (glnA), glutamine synthetase form B. subtilis or C. glutamicum (glnABs/Cg), glutamate synthase (gltB), phosphoglycerate dehydrogenase (serA), phosphoserine phosphatase (serB), phosphoserine aminotransferase (serC), membrane-bound proton-translocating pyridine nucleotide transhydrogenase (pntAB), soluble pyridine nucleotide transhydrogenase (sthA), L-serine deaminase II (sdaB), glucose-6-phosphate dehydrogenase (zwf), 6-phosphogluconolactonase (pgl), 6-phosphogluconate dehydrogenase (gnd), L-ribulose-5-phosphate 4-epimerase (araD), transketolase 1 (tktA), ribose-5-phosphate isomerase A (rpiA), ribose-phosphate diphosphokinase (prs), 3-deoxy-7-phosphoheptulonate synthase (aroG/H/F), 3-dehydroquinate synthase (aroB), 3-dehydroquinate dehydratase (aroD), shikimate dehydrogenase (aroE), shikimate kinase 1 (aroK), shikimate kinase 2 (aroL), 3-phosphoshikimate 1-carboxyvinyltransferase (aroA), chorismate synthase (aroC), chorismate mutase/ prephenate dehydrogenase (tyrA), chorismate mutase/prephenate dehydratase (pheA), anthranilate synthase complex (trpED), anthranilate synthase subunit TrpD (trpD), indole-3-glycerol phosphate synthase/phosphoribosylanthranilate isomerase (trpC), tryptophan synthase (trpBA), tryptophanase (tnaA), trp operon leader peptide (trpL), transcriptional regulator YihL (yihL), tyrosine repressor (tyrR), tryptophan repressor (trpR), catabolite repressor activator (fruR) and phosphoketolase from Bifidobacterium adolescentis (xfpkBa). Abbreviations for metabolites are: proton (H+), potassium ion (K+), sodium ion (Na+), ammonia ion (NH4+), phosphoenolpyruvate (PEP), acetyl-phosphate (acetyl-P), glucose 6-phosphate (G6P), fructose 6-phosphate (F6P), fructose 1,6-biphosphate (F1,6BP), glyceraldehyde 3-phosphate (G3P), dihydroxyacetone phosphate (DHAP), 3-phospho-D-glyceroyl phosphate (1,3DPG), 3-phospho-D-glycerate (3PG), D-glycerate 2-phosphate (2PG), oxaloacetate (OAA), succinyl-CoA (Suc-CoA), α-ketoglutarate (AKG), 3-phosphohydroxypyruvate (3PHP), O-phospho-L-serine (SOP), 6-phospho-D-glucono-1,5-lactone (6PGL), 6-phospho-D-gluconate (6PG), D-ribulose 5-phosphate (Ru5P), D-xylulose 5-phosphate (Xu5P), D-erythrose 4-phosphate (E4P), D-ribose 5-phosphate (R5P), sedoheptulose 7-phosphate (SH7P), 5-phospho-D-ribose 1-diphosphate (PRPP), 3-deoxy-D-arabinoheptulosonate 7-phosphate (DAHP), 3-dehydroquinate (DHQ), dehydroshikimate (DHS), shikimate-5-phosphate (S5P), 5-O-(1-carboxyvinyl)-3-phosphoshikimate (CVPS), N-(5-phospho-D-ribosyl)anthranilate (PRAN), 1-(2-carboxyphenylamino)-1-deoxy-D-ribulose 5-phosphate (CDRP) and indole-3-glycerol-phosphate (IGP).
Figure 1.
Gene modifications used to increase tryptophan production in Escherichia coli. Genes deleted or attenuated are shown in orange, overexpressed genes in light green, and genes mutated to be insensitive to feedback inhibition are shown in dark green. Heterologous genes have a subscript that identifies the species from which they were obtained: Corynebacterium glutamicum (Cg), Aspergillus niger (An), Zymomonas mobilis (Zm), Bacillus subtilis (Bs), and Bifidobacterium adolescentis (Ba). Abbreviations for genes and reactions are: high-affinity ATP-driven potassium transport system (kdp), ATP-depending proton transporter (PT), citrate transporter (citT), tryptophan permease (tnaB), aromatic amino acid transport protein (aroP), tryptophan-specific transport protein (mtr), aromatic amino acid exporter (yddG), galactose permease (galP), glucose facilitator from Zymomonas mobilis (glfZm), glucokinase (glk), enzyme IIBC of PTS (ptsG), enzyme IIAGlc of PTS (crr), phosphocarrier protein HPr (ptsH), enzyme I of PTS (ptsI), glucose-6-phosphate isomerase (pgi), 6-phosphofructokinase (pfkA), fructose-1,6-bisphosphatase (fbp), fructose-bisphosphate aldolase (fba), triose-phosphate isomerase (tpiA), glyceraldehyde-3-phosphate dehydrogenase (gapA), phosphoglycerate kinase (pgk), phosphoglycerate mutase (gpmA), enolase (eno), pyruvate oxidase (poxB), pyruvate kinase 1 (pykF), pyruvate kinase 2 (pykA), phosphoenolpyruvate synthetase (ppsA), pyruvate dehydrogenase complex (PDH), phosphate acetyltransferase (pta), acetate kinase (ackA), propionate/acetate kinase (tdcD), acetyl-CoA synthetase (acs), phosphoenolpyruvate carboxylase (ppc), acetaldehyde-CoA dehydrogenase/alcohol dehydrogenase (adhE), citrate synthase (gltA), aconitate hydratase A (acnA), aconitate hydratase B (acnB), isocitrate dehydrogenase (icd), 2-ketoglutarate dehydrogenase complex (AKGDH), succinyl-CoA synthetase (sucCD), succinate:quinone oxidoreductase complex (sdhABCD), fumarase A (fumA), fumarase B (fumB), fumarase C (fumC), pyruvate carboxylase from Corynebaterium glutamicum (pycCg), phosphenolpyruvate carboxykinase (pckA), malate dehydrogenase (mdh), NAD-dependent malic enzyme (maeA), isocitrate lyase (aceA), malate synthase A (aceB), D-lactate dehydrogenase (ldhA), pyruvate formate-lyase (pflB), glutamate dehydrogenase (gdhA), glutamine synthetase (glnA), glutamine synthetase form B. subtilis or C. glutamicum (glnABs/Cg), glutamate synthase (gltB), phosphoglycerate dehydrogenase (serA), phosphoserine phosphatase (serB), phosphoserine aminotransferase (serC), membrane-bound proton-translocating pyridine nucleotide transhydrogenase (pntAB), soluble pyridine nucleotide transhydrogenase (sthA), L-serine deaminase II (sdaB), glucose-6-phosphate dehydrogenase (zwf), 6-phosphogluconolactonase (pgl), 6-phosphogluconate dehydrogenase (gnd), L-ribulose-5-phosphate 4-epimerase (araD), transketolase 1 (tktA), ribose-5-phosphate isomerase A (rpiA), ribose-phosphate diphosphokinase (prs), 3-deoxy-7-phosphoheptulonate synthase (aroG/H/F), 3-dehydroquinate synthase (aroB), 3-dehydroquinate dehydratase (aroD), shikimate dehydrogenase (aroE), shikimate kinase 1 (aroK), shikimate kinase 2 (aroL), 3-phosphoshikimate 1-carboxyvinyltransferase (aroA), chorismate synthase (aroC), chorismate mutase/ prephenate dehydrogenase (tyrA), chorismate mutase/prephenate dehydratase (pheA), anthranilate synthase complex (trpED), anthranilate synthase subunit TrpD (trpD), indole-3-glycerol phosphate synthase/phosphoribosylanthranilate isomerase (trpC), tryptophan synthase (trpBA), tryptophanase (tnaA), trp operon leader peptide (trpL), transcriptional regulator YihL (yihL), tyrosine repressor (tyrR), tryptophan repressor (trpR), catabolite repressor activator (fruR) and phosphoketolase from Bifidobacterium adolescentis (xfpkBa). Abbreviations for metabolites are: proton (H+), potassium ion (K+), sodium ion (Na+), ammonia ion (NH4+), phosphoenolpyruvate (PEP), acetyl-phosphate (acetyl-P), glucose 6-phosphate (G6P), fructose 6-phosphate (F6P), fructose 1,6-biphosphate (F1,6BP), glyceraldehyde 3-phosphate (G3P), dihydroxyacetone phosphate (DHAP), 3-phospho-D-glyceroyl phosphate (1,3DPG), 3-phospho-D-glycerate (3PG), D-glycerate 2-phosphate (2PG), oxaloacetate (OAA), succinyl-CoA (Suc-CoA), α-ketoglutarate (AKG), 3-phosphohydroxypyruvate (3PHP), O-phospho-L-serine (SOP), 6-phospho-D-glucono-1,5-lactone (6PGL), 6-phospho-D-gluconate (6PG), D-ribulose 5-phosphate (Ru5P), D-xylulose 5-phosphate (Xu5P), D-erythrose 4-phosphate (E4P), D-ribose 5-phosphate (R5P), sedoheptulose 7-phosphate (SH7P), 5-phospho-D-ribose 1-diphosphate (PRPP), 3-deoxy-D-arabinoheptulosonate 7-phosphate (DAHP), 3-dehydroquinate (DHQ), dehydroshikimate (DHS), shikimate-5-phosphate (S5P), 5-O-(1-carboxyvinyl)-3-phosphoshikimate (CVPS), N-(5-phospho-D-ribosyl)anthranilate (PRAN), 1-(2-carboxyphenylamino)-1-deoxy-D-ribulose 5-phosphate (CDRP) and indole-3-glycerol-phosphate (IGP).
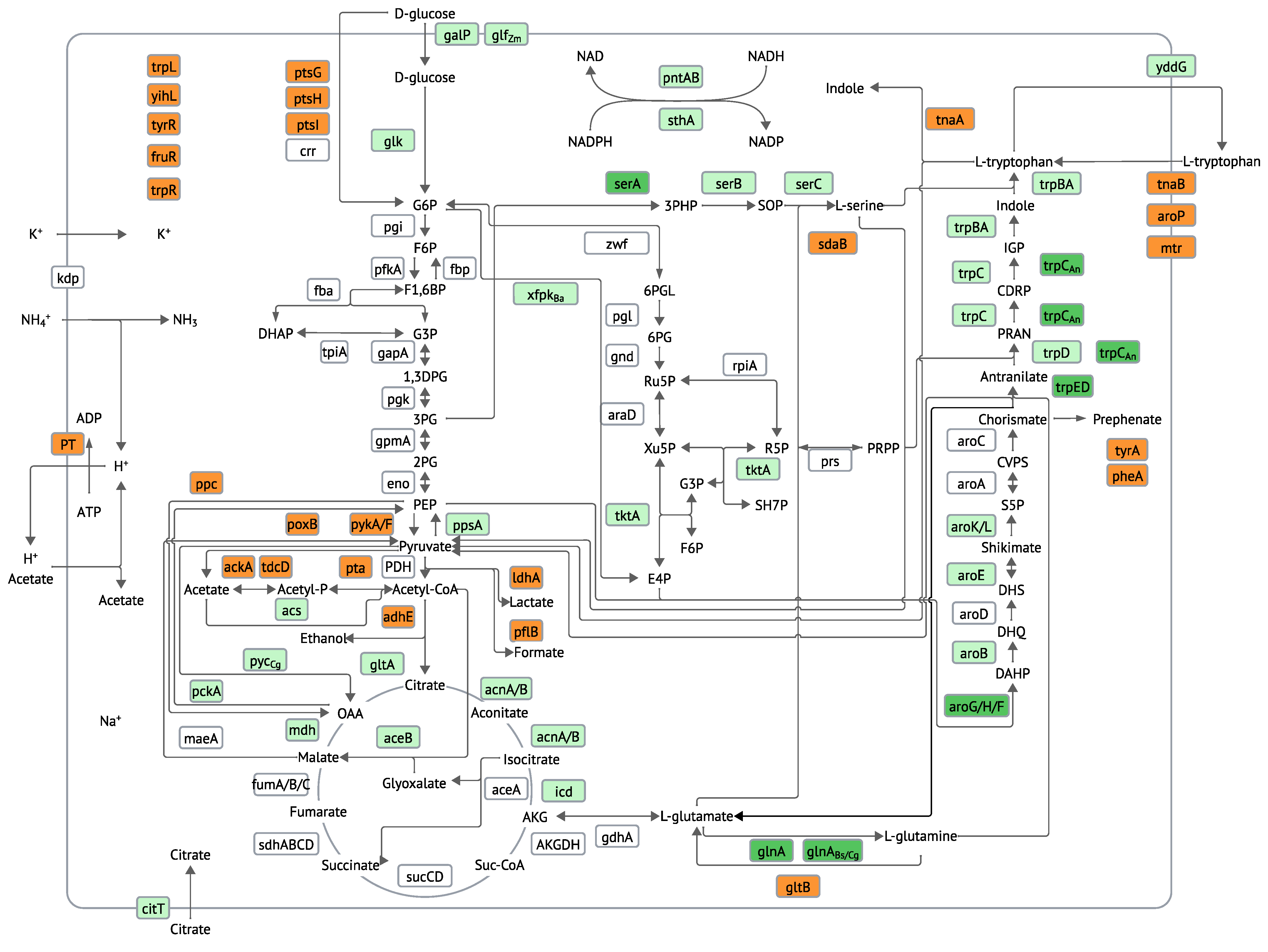
Figure 2.
E. coli strains modified to improve tryptophan production. Light green indicates overexpression, dark green represents mutations resistant to negative feedback, yellow indicates transcriptional or enzymatic attenuation, orange represents deletion, and gray indicates that the strain was obtained through random mutagenesis. Heterologous expression of genes from Corynebacterium glutamicum (Cg), Aspergillus niger (An), Zymomonas mobilis (Zm), and Bifidobacterium adolescentis (Ba) is represented by a subscript with its initials.
Figure 2.
E. coli strains modified to improve tryptophan production. Light green indicates overexpression, dark green represents mutations resistant to negative feedback, yellow indicates transcriptional or enzymatic attenuation, orange represents deletion, and gray indicates that the strain was obtained through random mutagenesis. Heterologous expression of genes from Corynebacterium glutamicum (Cg), Aspergillus niger (An), Zymomonas mobilis (Zm), and Bifidobacterium adolescentis (Ba) is represented by a subscript with its initials.
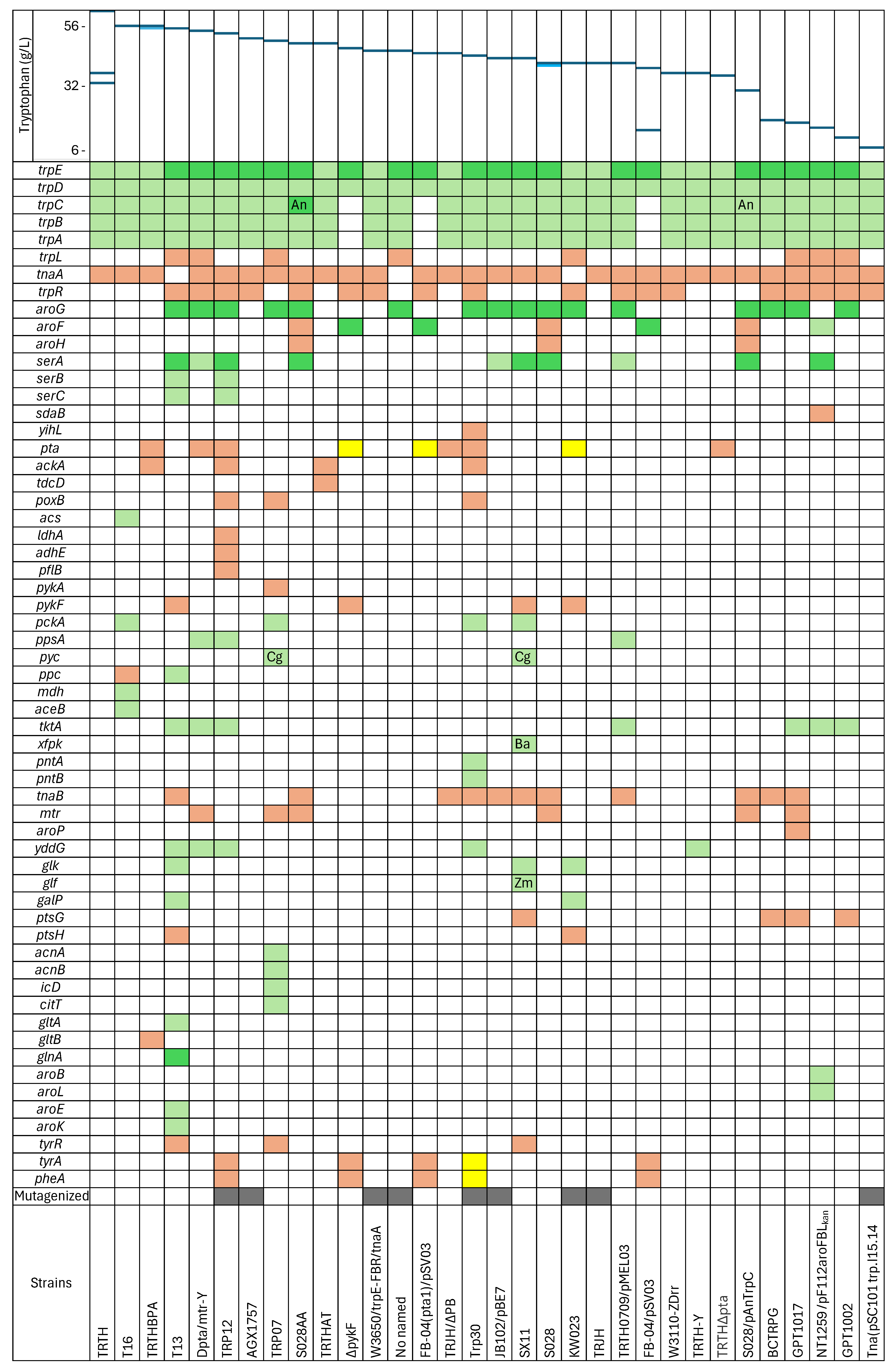
Figure 3.
Diagram with elements of a basic fermentation process.
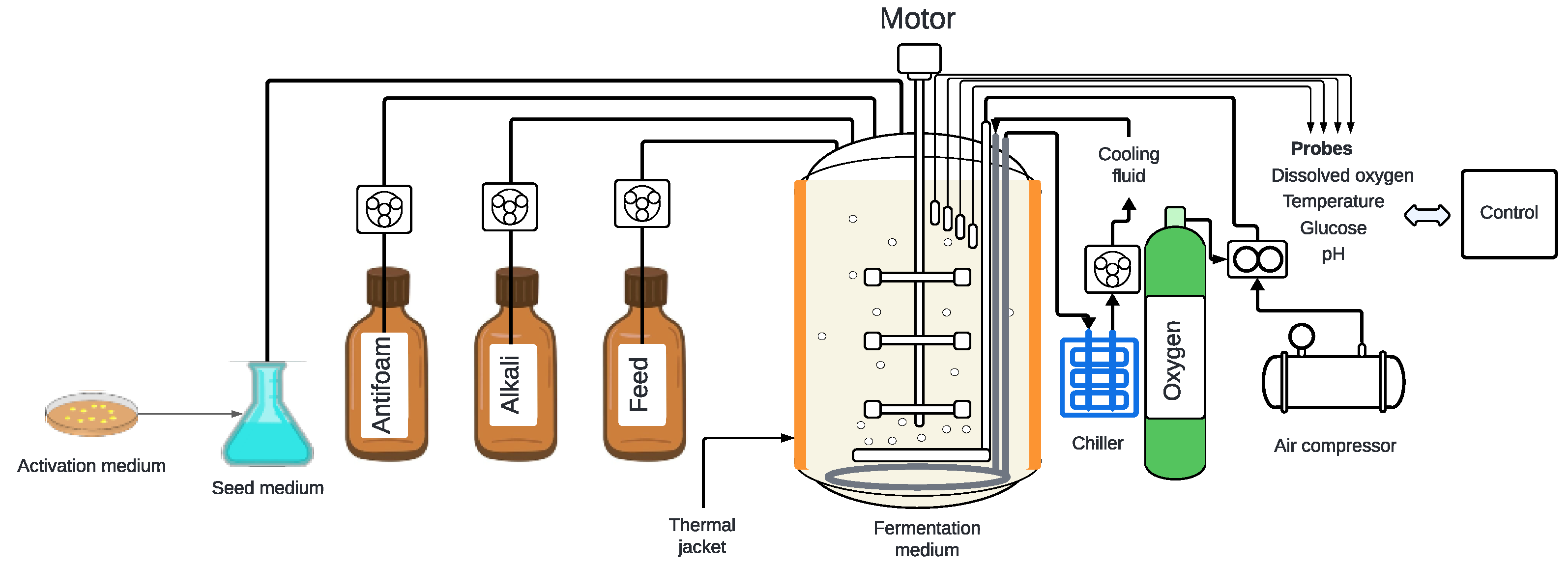

Table 1.
Parameters affecting tryptophan production.
| Parameter | Pathways | Genes* | Effect | Ref. |
|---|---|---|---|---|
| Low DO | Acetate, glyoxylate shunt, TCA cycle | pta, ackA, poxB, tdcD, gltA, icd | Acetyl-CoA, pyruvate, lactate and acetate accumulation. | [4,38,41,55,62,79,80] |
| High DO | Acetate degradation, glycolysis, gluconeogenesis, PTS, PPP, glutamate and tryptophan | fba, rpoS, acs, pckA, ppsA, ppc, maeA, pfk, zwf, pgl, tktA, araD, aroG, aroK, trpEDCBA, gdhA, pntAB | Acetate consumption, production of PEP, E4P, NADPH, tryptophan and glutamate, reduction of glucose consumption and oxidative stress. | [4,38,41,62,74,81,82,83] |
| Low pH | Glycolysis, acetate and TCA cycle | pfk, sucB, sucC | Low growth rate, stable plasmids, low acetate production and tryptophan crystallization. | [4,25,32,41,61] |
| High pH | Acetate, shikimate and tryptophan | pta, aroK, tnaA, tnaB | Accumulation of acetate, NH4+ and K+, and tryptophan production. | [12,41,61] |
| Low growth rate | Glycolysis, acetate | Plasmid stability, low acetate production, increase tryptophan production. | [3,16,53,61] | |
| High growth rate | Glycolysis, acetate | Biomass and acetate accumulation. | [3,61,64,68,75] | |
| Low feed rate | Glycolysis, acetate and tryptophan | Low production of acetate, biomass and tryptophan. | [4,26,38,41,55,62,63,64,68] | |
| High feed rate | Acetate and glutamate | gdhA | Acetate and glutamate accumulation. | [4,26,47,61,68,75,80,84] |
| Calcium | TCA cycle | gltA, icd, sucC | Increase ATP and biomass production | [40,42] |
| Betaine | PPP, stress response and K+ uptake | zwf | Activation of PPP, osmotic stress relief, reduce glutamate accumulation, reduce K+ uptake. | [85,86] |
| Citrate | Ca2+ uptake, glycolysis, PPP and TCA cycle | pykAF, zwf, gltA | Reduce production of acetate and glutamate while increase accumulation of PEP, NADH, NADPH and ATP. | [48,66] |
| High organic nitrogen | Incrase acetate and biomass production. | [56,66] | ||
| Inorganic nitrogen | Glutamate, tryptophan, glutamine | glnA | Increase energy waste, change pH, affect ionic strength, provide of NH4+ for tryptophan production and reduce plasmid stability. | [3] |
| Phosphate | Glycolysis | Increase growth rate, increase acetate accumulation. | [12,43,65] | |
| Surfactants | PPP, tryptophan transport, glutamate | zwf, yddG, trpBA | Enhance crystallization and secretion of tryptophan, reduce inhibition and is a sink for indole. | [9,12,65] |
| Cation’s accumulation | Change pH and ionic strength, difficult the purification of tryptophan and increase glutamate accumulation. | [41,61,86,87,88] | ||
| Precursors | Tryptophan, isoleucine, threonine | trpC, serA, thrA | Improve tryptophan production, feedback inhibition of precursor pathway. | [9,41,64] |
| Phenylalanine | Relieves serine inhibition, block common pathways of aromatic amino acids. | [5,89] | ||
| Leucine | Relieves serine inhibition. | [89] | ||
| Isoleucine | Isoleucine | Relieves serine inhibition. | [89] | |
| Methionine | Methionine | Increase production of proteins and reduce accumulation of acetate. | [6,19,51,66,68] | |
| High volume of inoculum | Increase rate growth and reduce lag-phase. | [43,59] |
* Genes symbols are the same as in Figure 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



